 Mário Barletta
Mário Barletta André R. A. Lima
André R. A. Lima- Laboratório de Ecologia e Gerenciamento de Ecossistemas Costeiros e Estuarinos, Departamento de Oceanografia, Universidade Federal de Pernambuco, Recife, Brazil
Research on estuarine ecology in South America (SA) increased quali-quantitatively since the early 1980 in search of consistent recommendations for estuarine conservation. The most important ecological theory achieved is that the seasonal fluctuation of the salinity gradient creates an ecocline influenced by gradual changes between river-dominated to marine-like waters. Estuarine fish fauna adapts to these changeable abiotic characteristics, including the spatial, and seasonal bioavailability of dissolved oxygen and numerous pollutants. However, studies on the influence of the estuarine ecocline are still missing for key estuarine systems. This study provides an overview of fish ecology and anthropogenic impacts within estuarine systems of SA and discusses priorities for environmental conservation. Research on fish reached important conclusions regarding essential habitats and fish interaction with other biological and abiotic compartments over spatio-temporal settings, including conditions of severe anthropogenic impacts. These impacts are related to unplanned urban settlements, industrial estates, ports, damming of major rivers, dredging activities, and deforestation for extensive farming. Changes in estuarine morphology alter natural flows and lead to habitat losses, disrupting the ecocline and impairing fishes from moving among formerly connected habitats, especially earlier ontogenetic phases. In addition, industrial, urban, and farming activities often result in high loads of metals and persistent organic pollutants, organic enrichment and oxygen depletion. Moreover, plastic debris, a ubiquitous contaminant with sources on every human activity, including fishing, when fragmented into microplastics, become preferably concentrated in semi-enclosed environments, as estuaries. Metals, POPs and microplastics are actually asserted to be persistent. When in high concentrations, they become bioavailable to the estuarine trophic web through bioaccumulation, being biomagnified or biotransfered toward higher trophic level organisms, such as top predator fishes. Therefore, research on environmental quality and fish ecology must be based on robust sampling designs along the whole ecocline using long-term approaches. In addition, basic sanitation, co-management, an improved licensing system and scientifically-based risk assessments/monitoring for all sorts of enterprise are also urgent. These conservation priorities need to be in place before human-driven changes surpass the ecosystem's capacity to produce resources and maintain services.
Introduction
Estuaries are characterized by gradual changes between freshwater inputs and marine waters intrusion (Barletta et al., 2017a). The encounter of these two water masses creates zones of mixing and vertical stratification of various strengths. Such stratification results in a variety of abiotic habitats that function as boundaries defining different faunal communities, which are influenced mainly by the seasonal fluctuation of the salinity gradient as induced by diel tidal variation and river runoff. Thus, an ecocline can be defined as a “gradation from one ecosystem to another when there is no sharp boundary between the two” (Attrill and Rundle, 2002). Usually, these boundaries are freshwater-oligohaline/upper, mesohaline-mixoeuhaline/middle, and euhaline-hyperhaline/lower reaches (Day et al., 2012). Each of these reaches can actually move horizontally, advancing seawards according to increased rainfall patterns or upstream under tidal influence (Barletta et al., 2005). Moreover, the seasonal balance of dissolved oxygen, suspended solids, pollutants, and microbiological contaminants are also ruled by the estuarine ecocline and temperature variability (Barletta et al., 2017a). Not surprisingly, this ecocline is the strongest forcing structuring physico-chemical conditions and fish communities within any estuarine ecosystems (Barletta et al., 2008).
Human interventions, such as damming of major rivers and dredging of channels along the estuarine course, might weakens and even disrupt the ecocline, reducing the original ecological functions, impairing the sustainable use of estuarine resources and services (Blaber and Barletta, 2016). These modifications are also responsible for disrupting sediment flows, change bottom composition and impair freshwater flow seawards, leading to changes in habitat availability, biodiversity damages, and shifts in biogeochemical cycles of several chemicals and pollutants (Barletta et al., 2016, 2019). Moreover, entire river basins, estuarine courses, and adjacent coastal waters are subjected to the influence of industrial, urban and farming activities, which are responsible for high loads of wastewaters and solid wastes (Huang et al., 2014; Pereira et al., 2015). Thereby, plastic debris, metals, nutrients, POPs and emerging pollutants present high concentrations closer to their sources, usually urban settlements, industrial estaes, and farms. Their chemical behavior and bioavailability for fishes along the way to the sea are also influenced by the major annual water quality shifts. The solubilisation, speciation, sorption, precipitation, diffusion, advection, sinking toward sediments, complexation, and absorption by the estuarine food web is the result of the seasonal fluctuation of the estuarine ecocline. Overall, pollutants are entraped in estuaries when saline intrusion is great and flushed out to the sea when river runoff increases (Barletta et al., 2012, 2019; Lima et al., 2014). Metals, POPs and microplastics are all persistent in the environment. The bioaccumulation and biomagnification of metals and POPs (Lanfranchi et al., 2006; Cappelletti et al., 2015), as well as the trophic transfer of microplastics (Galloway et al., 2017) are common in estuarine trophic webs. In addition to habitat loss by water quality/volume changes and damming of major rivers, fishes are also subjected to fishing pressure and to compete for resources with introduced species, often resulting in decreased native fish biomass within estuaries (Neuhaus et al., 2016; Barletta et al., 2017b). Currently, it is asserted that if human-driven changes remain uncontrolled, estuaries may become increasingly vulnerable to co-working global changes (Costa and Barletta, 2016; Ríos-Pulgarín et al., 2016). However, despite these impacts, changes in local climate and hydrodynamic variability might contribute to reduce pollution through biodilution and transport of contaminants out of the system, especially during the rainy season, when river flow increases, in a process known as environmental homeostasis (Elliott and Quintino, 2007). On the other hand, systems that suffer with severe droughts may not be able to recover and will face long and lasting antropogenic modifications.
This work is an overview of estuarine fish research and anthropogenic influences on estuarine fishes and estuarine ecosystems of SA located in tropical, subtropical and warm temperate provinces (Figure 1 and Table 1; study areas are described as Supplementary Material). Whereas, the current knowledge of estuarine fish fauna are poorly understood in this continent, this review focused in the description of the available information on fish ecology and how human-driven contamination and modifications have affected several estuarine systems and fishes of commercial and subsistence importance. To gather and interpret all the relevant literature aiming at generating managerial information was a daunting task, but a unique opportunity to promote well-informed decision-making, especially considering the transition among environments from equatorial to temperate regimes (Costa and Barletta, 2016). However, the ecocline theory is hardly ever attempeted in most studies. Therefore, the review also discusses on how to work in favor of the seasonal fluctuation of the ecocline is important to understand the distribution pattern of dominant fish species and major contaminants along estuarine systems and food webs.
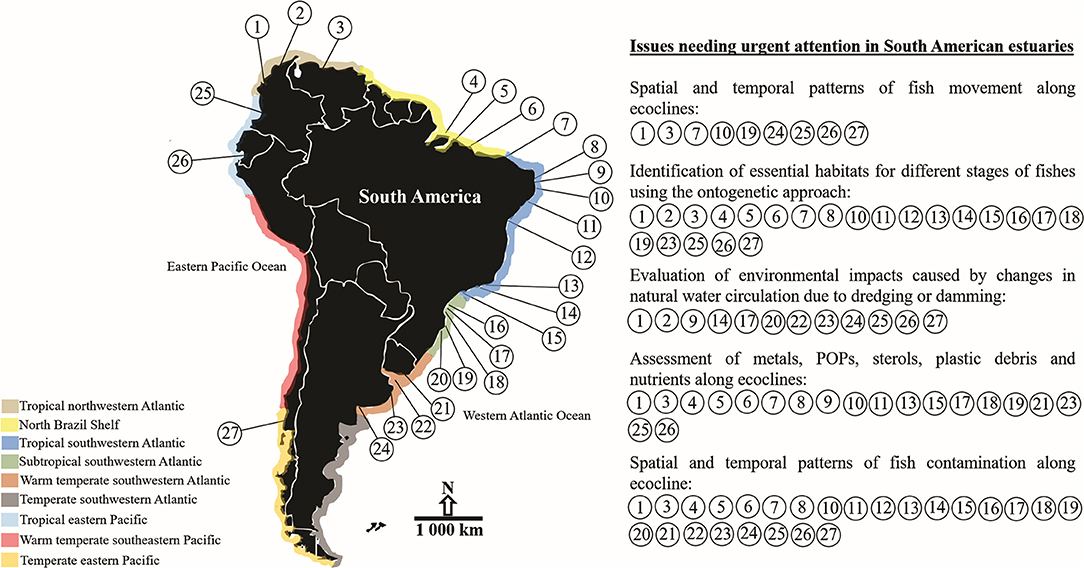
Figure 1. Location and main conservation issues identified in South American estuaries. Tropical northwestern Atlantic [(1) Atrato River Delta; (2) Ciénaga Grande de Santa Marta Lagoon; (3) Orinoco River Delta]. North Brazil Shelf [(4) Amazon Estuarine Complex; (5) Curuçá Estuary; (6) Caeté Estuary]. Tropical southwestern Atlantic [(7) Parnaiba River Delta; (8) Potengi Estuary; (9) Goiana Estuary; (10) Itamaracá Estuarine System; (11) São Francisco River Estuary; (12) Paraguaçu Estuary; (13) Guanabara Bay; (14) Sepetiba Bay; (15) Santos Estuary]. Subtropical southwestern Atlantic [(16) Paranaguá Estuarine Complex; (17) Guaratuba Bay; (18) Babitonga Bay; (19) Laguna Estuarine Complex; (20) Patos Lagoon Estuary]. Warm temperate southwestern Atlantic [(21) Pando Sub-Estuary; (22) Río de La Plata Estuary; (23) Mar Chiquita Coastal Lagoon; (24) Bahía Blanca Estuary]. Tropical eastern Pacific [(25) Bahía Málaga Estuary; (26) Rio Palmar and Rio Javita Estuaries]. Temperate eastern Pacific [(27) Valdivia Estuary]. Source: Google Earth.
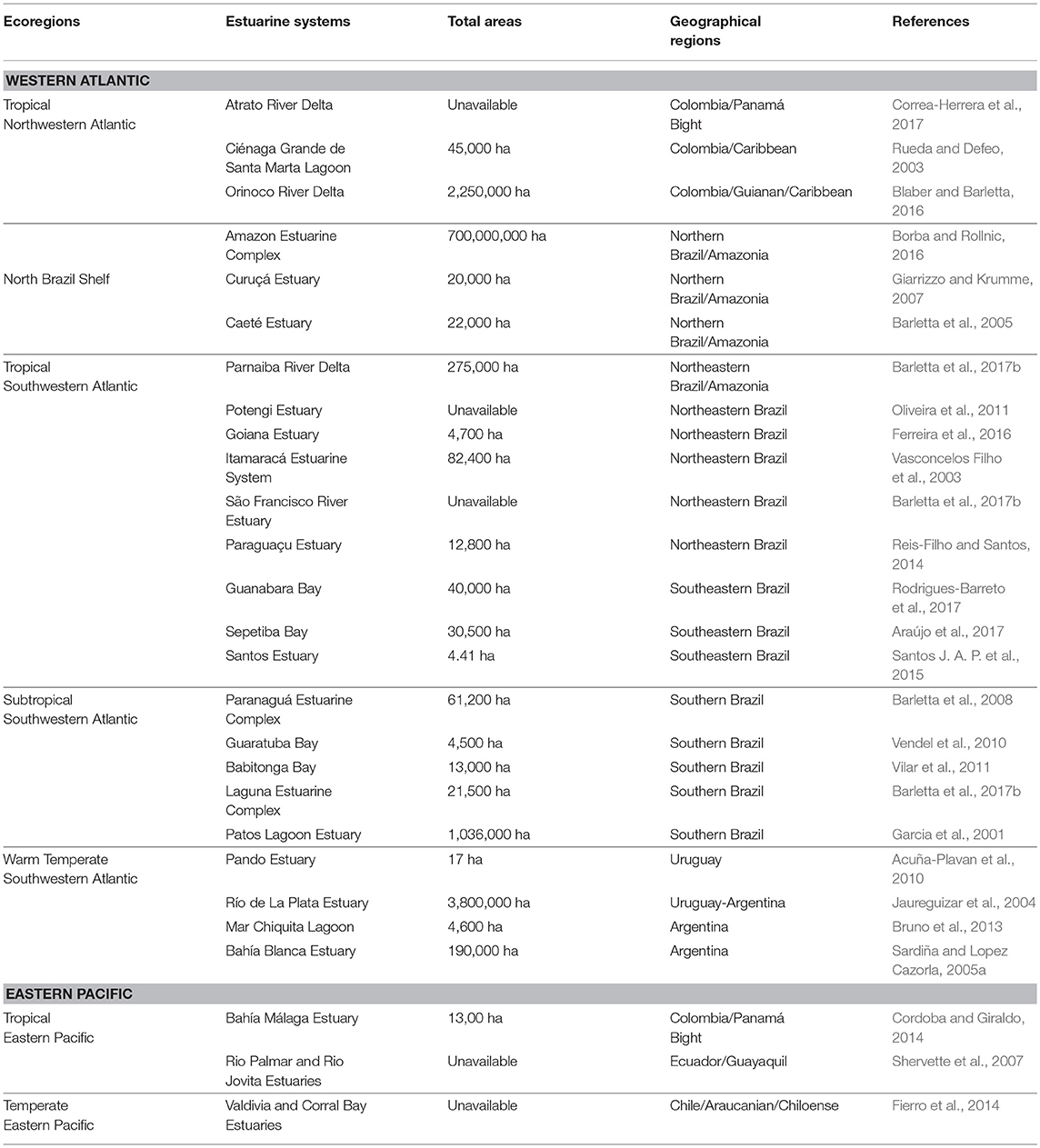
Table 1. Ecoregions, geographical regions, and total area of key estuarine systems in South America based on available published data.
Methods
To address fish ecology and environmental quality of SA estuaries into a single manuscript, we performed a vast scientific search aiming at collect the greatest amount of studies in estuarine systems of every coastal country sourrunding the continent. We compiled information of nearly 420 articles found in the databases Web of Science, Scopus, and Google Scholar.
Results
Western Atlantic Estuaries
Tropical Northwestern Atlantic
Ciénaga Grande de Santa Marta Lagoon: Colombian Caribbean Coast
A complete description on the influence of the seasonal fluctuation of the estuarine ecocline on fish assemblages is available encompassing two annual cycles of 1993–1994 and 1997 within the Ciénaga Grande de Santa Marta Lagoon (Rueda and Defeo, 2003; Figure S1). The most contrasting seasons (major rainy and major dry), were used as temporal factors to reveal a strong relation between salinity variability and fish density ruled by seasonal shifts. Three distinct assemblages were determined by the salinity gradient, changing from freshwater to marine-estuarine and marine assemblages (Rueda and Defeo, 2003). Eugerres plumieri, Diapterus rhombeus (Gerreidae), Micropogonias furnieri (Sciaenidae), Mugil incilis (Mugilidae), Cathorops spixii (Ariidae), Elops saurus (Elopidae), and Anchovia clupeoides (Engraulidae) were the most frequent species. Anchovia clupeoides typified the rainy season of 1997; Cathorops spixii and E. saurus typified the dry season; and M. incilis and M. furnieri the rainy season in both annual cycles. D. rhombeus and E. plumieri typified the dry and rainy seasons of 1993–1994 (Rueda and Santos-Martínez, 1999; Rueda and Defeo, 2003). The highest densities of E. plumieri and M. incilis; and a peak of C. spixii occurred during the rainy season, near the opening of the lagoon (Rueda, 2001).
Regarding biological studies, the feeding ecology of A. clupeoides (Duque and Acero, 2003) and Oligoplites spp. (Carangidae) (Duque-Nivia et al., 1996) were described using spatio-temporal patterns along the salinity ecocline. The reproduction of Bairdiella ronchus (Sciaenidae) was also described according to the relationship between monthly gonadosomatic index and salinity variability, however no relationship was identified (Castro et al., 1999).
North Brazil Shelf
Amazon Estuarine Complex: North Brazil/Amazonia
In the Amazon Estuarine Complex (Figure S2), ecological guilds and fish diversity were assessed according to spatial and sesonal structures within the main channel and tidal creeks of the Pará River estuary (Mourão et al., 2014, 2015). Fish composition, abundance, and the use of estuarine habitats as nursery and reproduction grounds were mainly structured by salinity. The highest diversity and abundance of fishes occurred during the dry season in the main channel, while tidal creeks were used for reproduction. Migrants and freshwater stragglers were dominant in the lower estuary (Guamá River and Guajará Bay), while estuarine, marine stragglers and migrants predominated along the main channel (Mourão et al., 2014). Piscivorous and zoobenthivorous were the dominant feeding groups in all studied areas. Most species were occasional, characterizing the estuary as a transition zone (Mourão et al., 2015).
Starting from this estuary and traveling all the way to the Andes foothills, Amazonian catfishes (Brachyplatystoma spp.) perform one of the longest fish migrations in the world. They are commercially important and utilize the entire Amazon Basin to complete their life cycles. Populations are declining due to overfishing and to the presence of dams in the upper reaches of the migration range, which impairs spawning (Barthem et al., 1991). Otolith microchemistry revealed that larvae migrate downstream from the Andean piedmont to the lower Amazon (Duponchelle et al., 2016), while juveniles exhibit diverse strategies, rearing upstream, or downstream (Hegg et al., 2015).
Studies on the ecology of Amazonian freshwater stingrays (Potamotrygonidae) were performed within the Amazon complex. Potamotrygon motoro is a predominant species in Marajó Island (between the Amazon and Pará estuaries) showing preference for intermediate salinities and being resistant to critical conditions of pH, dissolved oxygen and temperature (Almeida et al., 2009). Paratrygon aiereba was studied according to cytogenetic approaches. The results revealed that their populations are structured within each one of the five rivers of the Amazon Basin, with no gene flow. Thus, the species is acknowledged as three distinct biological species, emphasizing that the management of their exploitation needs to focus on each river, rather than the entire river basin (Frederico et al., 2012).
In the Pará River Estuary, length–weight relationships of fish species (Loureiro et al., 2017), as well as the reproductive biology of commercial sciaenids are available (Santos et al., 2010; Barbosa et al., 2012). Plagioscion squamosissimus (sciaenidae) reproduces primarily in February/March and August/September (Barbosa et al., 2012), while Plagioscion magdalenae (Sciaenidae) spawns mainly from August to February (Santos et al., 2010).
Curuçá Estuary: North Brazil/Eastern Amazon
In Curuçá Estuary (Figure S3), forty intertidal fish species had their weight-length relationship studied by Giarrizzo et al. (2006). Moreover, the ontogenetic and seasonal diet shifts of Sciades herzbergii (Ariidae) (Giarrizzo and Saint-Paul, 2008), Lutjanus jocu (Lutjanidae) (Monteiro et al., 2009), and Colomesus psittacus (Tetraodontidae) (Giarrizzo et al., 2010) were also assessed for tidal creeks. The seasonal availability of food was more important than ontogenetic diet shifts; and stable isotope approaches revealed different types of food webs within the same ecosystem (Giarrizzo et al., 2011; Schwamborn and Giarrizzo, 2015).
The seasonal changes in the composition of intertidal fish assemblages inhabiting four creeks of the same salinity zone in the upper and middle reaches were investigated (Giarrizzo and Krumme, 2007). Fish composition changed seasonally, while density and biomass differed spatially, suggesting that spatial composition within habitats of homogenous salinity are dependent on landscape features.
Caeté River Estuary: North Brazil/Eastern Amazon
In the Caeté Estuary (Figure S4), fluctuations of the salinity gradient were the main forcing structuring fish assemblages over the annual cycle of 1996–1997 (Barletta et al., 2005). Stellifer rastrifer, Stellifer microps (Sciaenidae), C. spixii, and Aspredo aspredo (Aspredinidae) were the most abundant species. The species S. rastrifer was found along the entire main channel, regardless seasons; S. microps and A. aspredo were abundant in the upper estuary over the year, while C. spixii peaked in the middle and lower reaches in the early-rainy season. Larval phases of these four frequent species were also found in the main channel (Barletta-Bergan et al., 2002a). In addition, larvae of Stellifer and Cathorops genera were also found in mangrove creeks, emphasizing the importance of the system for the life cycle of subsistence species (Barletta-Bergan et al., 2002b).
Larval A. Clupeoides and S. microps were abundant in the main channel, peaking in the upper estuary in the dry season (Barletta-Bergan et al., 2002a). Guavina guavina (Eleotridae) and A. clupeoides were also frequent in the creeks, peaking in August (Barletta-Bergan et al., 2002b). Larval Cynoscion acoupa (Sciaenidae) was the most abundant commercial species in terms of density in both main channel and creeks. However, later stages were not frequent in the main channel (Barletta et al., 2005) or even in creeks (Barletta et al., 2003), placing this species as marine-estuarine dependent. Moreover, in a mangrove lake at the middle estuary, Achirus achirus (Achiridae), and the commercial taxa Centropomus pectinatus, Centropomus undecimalis (Centropomidae) and M. curema, accounted for 82% of the total catch in weight along 24 h sampling-cycles in wet and dry months, reinforcing the importance of the connectivity among estuarine habitats for fishes (Goch et al., 2005).
Tropical Southwestern Atlantic
Goiana River Estuary: Northeast Brazil
The main channel of the Goiana Estuary (Figure S5) was year-round characterized along its ecocline. In a rare effort, the influence of the ecocline on the movement patterns of fishes have been assessed throughout their ontogeny, from larval to adult phases along the annual cycles of 2005–2006 and 2012–2013. Fluctuations of the salinity ecocline and levels of dissolved oxygen structured the ariids C. spixii and Cathorops agassizii (Dantas et al., 2012). Juveniles peaked in the middle estuary in the early-rainy season (March–May), characterizing their nursery ground. For C. spixii, the nursery shifted to the lower estuary during the late-rainy season (June–August), while for C. agassizii it remained in the middle estuary. Sub-adults and adults peaked in the upper estuary in the rainy season. High densities of mouth-brooder males, free embryos and young juveniles were observed in the upper estuary during the late-dry season (December–February), characterizing their reproductive grounds. Mangrove creeks are important grounds only for later phases of these species, since earliest phases receive parental care (Ramos et al., 2011; Lima et al., 2012, 2013).
Salinity and water temperature shifts structured the patterns of estuarine use by Stellifer brasiliensis and Stellifer stellifer (Sciaenidae) along the system (Dantas et al., 2015). The middle estuary was an important nursery ground for juveniles and a feeding ground for sub-adults and adults of both species in the late-rainy season (June–August), except for adult S. stellifer, which peaked in the early-dry season (September–November). Later phases and larval S. brasiliensis were also found in mangrove creeks (Ramos et al., 2011; Lima et al., 2016), but were absent in the main channel, while larval S. stellifer were only recorded in the main channel (Lima et al., 2015). The species C. acoupa uses the upper estuary as a nursery in the early-rainy season (Ferreira et al., 2016). Sub-adults peak in the upper estuary during the entire annual cycle and migrate to the middle estuary in the late-rainy season, while adults inhabit adjacent coastal waters. Larval C. acoupa peaks in the lower estuary over the entire year (Lima et al., 2015), suggesting that spawning occurs in coastal waters; and later, larval and sub-adult phases inhabit the estuary (Ramos et al., 2011; Ferreira et al., 2016; Lima et al., 2016).
The patterns of estuarine use by Gerreidae species were influenced by changes in salinity gradient and dissolved oxygen (Ramos et al., 2016). Juvenile Eugerres brasilianus uses the upper estuary and the middle estuary as nursery grounds during the late-dry and early-rainy seasons, respectively. Sub-adults peaked in the upper estuary in the early-dry season, while adults were abundant over the entire year. On the other hand, juvenile Eucinostomus melanopterus uses estuarine beaches as nursery in the early-rainy and dry seasons. Independent of seasons, the abundance of sub-adults and adults at beaches toward the north of the estuarine mouth points to a degree of competition for estuarine resources, since both phases rely on the same food items (Ramos et al., 2016). Both species were found in tidal creeks (Ramos et al., 2011), but the larval phase of E. brasilianus was recorded in the main channel (Lima et al., 2015).
São Francisco River Estuary: Northeast Brazil
Flood pulses and river flow dynamics structured fish movement in the São Francisco River basin (Nestler et al., 2012). However, the estuarine portion is severely affected by eight upstream hydroelectric power dams that control flood pulses and alternative fish habitats (Coimbra et al., 2017; Figure S6). Alterations in flow regime facilitated the establishment of the non-native Metynnis lippincottianus (Serrasalmidae), while some endemic fishes disappeared (Assis et al., 2017). Restocking programs for the endemic Prochilodus argenteus (Prochilodontidae) were successful, but are no further recommended because the low allelic richness of the hatchery stock can minimize genetic diversity and increase the divergence from ancestral generations (Coimbra et al., 2017). Restocking programs were also tried for Brycon orthotaenia (Bryconidae), and now, after 40 years, they are recorded in the system (Brito et al., 2016).
The estuary, however, still maintains important ecological functions regarding connectivity. Its mouth is an important ground for the gonadal maturation of E. brasilianus (Rodrigues et al., 2017). Moreover, coastal demersal fishes are also influence by changes in abiotic conditions. The freshwater flux during the rainy months increases the functional diversity of fishes at shallower depths along the coast and estuarine-adapted fishes invade shallow coastal regions in a process known as estuarization of the coast (Passos et al., 2016).
Paraguaçu River Estuary: Northeast Brazil
Fish assemblages and functional guilds in the Paraguaçu Estuary are distributed according to differences in the salinity gradient (Reis-Filho et al., 2010; Figure S7). The middle estuary was dominated by S. rastrifer and Cynoscion microlepdotus (Sciaenidae). Eucinostomus argenteus (Gerreidae), Pellona harroweri (Pristigasteridae), and D. rhombeus were frequent in the middle and lower estuaries, while Sphoeroides greeleyi (Tetraodontidae) was frequent over the main channel. Deep-water and estuarine-resident fishes were abundant in the middle estuary, while shallow-water fishes were common in more saline areas. Marine-migrants were found in all sectors, while marine-stragglers were common from the middle to the mouth of the estuary (Reis-Filho et al., 2010). Organic matter and salinity were the main factors influencing overall fish distribution. Dissolved oxygen and organic matter structured the distribution of Eleotris pisonis (Eleotridae), Trinectes paulistanus, Achirus declives (Achiridae), Sphoeroides testudineus (Tetraodontidae), D. rhombeus, Citharichthys spilopterus (Paralichthyidae), Ctenogobius stomatus and Ctenogobius stigmaticus (Gobiidae) in habitats unaffected by aggardation, and Dormitator maculatus (Eleotridae), E. argenteus and Atherinella brasiliensis (Atherinopsidae) in affected habitats of the upper estuary.
Guanabara Bay: Southeast Brazil
In Guanabara Bay, the influence of the seasonal variability of local environmental factors on spatial patterns of demersal fishes was assessed in a fortnightly survey among 2005–2007 (Silva et al., 2016; Figure S8). Salinity, dissolved oxygen, ammonium, and phosphorus were responsible for quali-quantitative changes in spatio-temporal estuarine use. Chilomycterus spinosus (Diodontidae), G. genidens, E. argenteus, and M. furnieri were abundant species. The species C. spinosus were concentrated in the middle bay exhibiting high tolerance to eutrophic conditions and variations in salinity and temperature (Santos S. R. et al., 2015). A rise in the abundance of small specimens were observed when rainfall increased. The species G. genidens had strong association with the upper bay, where ammonium, total phosphorus and temperatures were higher (Silva et al., 2016).
The species M. furnieri (Mulato et al., 2015) and E. Argenteus (Corrêa and Vianna, 2016) were also frequent throughout the year. Juveniles had higher frequency in the upper and middle bays, while larger fishes were frequent in the lower bay. Recruitment of M. furnieri is observed in late fall, winter and spring (Mulato et al., 2015), and E. argenteus recruits in the first semester of each year (Corrêa and Vianna, 2016).
At the seawards end of the bay, Haemulidae, Pomacentridae and Labrisomidae were the richest families, and Diplodus argenteus (Sparidae), Haemulon aurolineatum (Haemulidae), Stephanolepis hispidus (Monacanthidae), and Abudefduf saxatilis (Pomacentridae) were the abundant species inhabiting an exposed estuarine beach located in a marine-estuarine transitional zone (Vermelha beach) (Rodrigues-Barreto et al., 2017). For a sheltered estuarine beach (Flamengo beach), richness and abundance were higher during spring (September to November) and lower during winter (June to August), while the relative abundance of fishes varied according to seasonal shifts in day length (Vasconcellos et al., 2010). Harengula clupeola (Clupeidae) peaked in spring, A. brasiliensis in austral summer/autumn, and Trachinotus carolinus (Carangidae), Umbrina coroides (Sciaenidae), and M. liza in winter.
At the entrance of the Bay, fish larvae peaked with highest temperatures and lowest salinities (March) (Castro et al., 2005). Abundance was higher at night, and lower during the day. Larval Cetrengraulis edentulus, Anchoa lyolepis (Engraulidae), and Harengula jaguana (Cupeidae) were dominant during both ebb and flood tides.
The species C. edentulus is the main commercially exploited species in this environment, and spawns from late winter to spring with a peak in November (Franco et al., 2014). Genidens genidens (Ariidae) is a potential sentinel species due to its abundance and non-migratory habits; and southern and southeastern Brazil populations of this species exhibit similar reproductive season and length-weight relationship (Silva et al., 2013).
Sepetiba Bay Estuary: Southeast Brazil
Salinity, depth, and transparency are the main variables influencing fish assemblages along the Sepetiba Bay and sandy beaches in the inner and outer bay areas (Araújo et al., 2002; Pessanha and Araújo, 2003; Figure S9). The species D. rhombeus, C. spixii, and G. genidens dominated the system and preferred shallow, less saline and less transparent waters of the inner bay, where the lowest diversity and highest abundance were also observed. In the outer bay, it was observed the opposite situation (highest diversity and lowest abundance), and Cynoscion leiarchus (Sciaenidae), Diplectrum radiale (Serranidae), and G. aprion were frequent species. The middle bay is inhabited by fishes that prefer average salinities, or with no clear preference [e.g., G. genidens, G. aprion, and Eucinostomus gula (Gerreidae)] (Araújo et al., 2002).‘
In a long-term survey, the salinity gradient influenced spatial changes in fish communities (Araújo et al., 2017). The inner bay was characterized by Chloroscombrus chrysurus (Carangidae), G. genidens, and M. furnieri. In the middle bay, Prionotus punctatus (Triglidae), G. genidens, and M. furnieri were frequent species, while in the outer bay P. punctatus and D. radiale were typical species. Fish richness and abundance decreased over the last three decades (1990–2010s) and sharpest changes were observed in the inner and middle bays since they are the most impacted areas due to high loads of metal and to the enlargement of the Sepetiba Port (Araújo et al., 2017).
Santos Estuary: Southeast Brazil
In the Santos Estuary, D. rhombeus, A. brasiliensis, B. ronchus, G. genidens, and H. clupeola were the most abundant species in the upper estuary during a short-term survey of one rainy (March) and one dry month (August) (Santos J. A. P. et al., 2015; Figure S10). The dry period was dominated by H. clupeola and A. brasiliensis, while the rainy period was dominated by D. rhombeus, G. genidens and B. ronchus. Fish diversity peaked during the rainy month and it was associated to the appearance of a species with affinities for lower salinities (Santos J. A. P. et al., 2015).
Environmental Impacts in Tropical Western Atlantic Estuaries
Habitat Changes
In the Amazon Estuarine Complex, the replacement of culture of sugar cane to permanent agroforests, timber logging (including a shipping terminal), and an industrial estate at Pará River Estuary resulted in habitat loss and critical alterations of fish communities (Barros and Uhl, 1995; Viana and Lucena Frédou, 2014). For the Goiana Estuary, sugarcane plantations, milling, aquaculture, limestone mining, dredging, and urban development are among the landscape modifications (Barletta et al., 2017b).
Alterations in river flow patterns and volume due to damming and dredging, as well as water quality changes due to wastewater disposal, building, and operation of artisanal fishing harbors are severely degrading the São Francisco River Estuary (Barletta et al., 2017b). The decreased river flow reduced dissolved inorganic nitrogen and silicate seawards, and the estuary became oligotrophic (Martins et al., 2011; Medeiros et al., 2011; Genz and Luz, 2012).
Urbanization and pipeline routes for oil and gas industry have caused coastal erosion problems in Guanabara Bay (Araruna et al., 2014), and fish diversity became low in estuarine sandy beaches exposed to wave action with unlimited human access (Franco et al., 2016). In Sepetiba Bay, human modification include the enlargement of Sepetiba Port, dredging activities, construction of a steel factory and a terminal for building submarines (Araújo et al., 2016).
Environmental and Fish Contamination With Plastic Debris
Contamination by microplastics is a concern in the main channel and mangrove creeks of the Goiana Estuary and exibits complex patterns (Lima et al., 2014, 2016). Microplastics are found in every habitat throughout the entire year, sharing the same habitats as fishes and their prey (Lima et al., 2014, 2015, 2016). When precipitation increases, the freshwater flow seawards is a powerful exporter of this contaminant from the upper estuary to the marine environment. Total density of microplastics present the same order of magnitude as total fish larvae and fish eggs in the main channel. Simultaneous comparable amounts of microplastics and fish increase the chances of interactions of these two compartments (Lima et al., 2015). The ingestion of plastic debris by demersal fishes was recorded in ariid catfishes (Possatto et al., 2011), gerreids (Ramos et al., 2012), sciaenids (Dantas et al., 2012; Ferreira et al., 2016), and centropomids (Ferreira et al., 2019).
Sediments in beaches within Guanabara Bay are also polluted with plastic debris, especially microplastics (Carvalho and Baptista Neto, 2016). Microplastics peaked in the warm-rainy season (January-February), but also presented high concentration during the cold-dry season (June–July). Sources are associated to local rivers and streams, fishing and harbor activities, domestic effluents, maritime terminals, and mussel farming (Carvalho and Baptista Neto, 2016; Castro et al., 2016).
Sewage Contamination and Eutrophication
In Santos (Braga et al., 2000; Aguiar and Braga, 2007) and Curuça Estuarine Systems (Venekey and de Melo, 2016), fecal sterols, fecal coliforms, and high loads of nutrients are higher near industrial estates, farms, and cities, revealing a strong sewage contamination. High levels of eutrophication and thermotolerant coliforms are observed in the upper reaches of the Caeté Estuary during the dry season, however, water quality might improve during the rainy season (Monteiro et al., 2016a,b). Water in shallow areas of the inner Guanabara Bay have the poorest quality, being hypertrophic, and sometimes hypoxic (Ribeiro and Kjerfve, 2002; Aguiar et al., 2011). There, fecal sterols in sediments indicate areas of severe and historical sewage contamination (Paranhos et al., 1998; Santos et al., 2008). Poor water quality represents a risk to the biota due to low dissolved oxygen (Braga et al., 2000) and pathogens.
Environmental and Fish Contamination With Petroleum Hidrocarbons and Persistent Organic Pollutants
A pipeline leak following an accident in 2000 was responsible for ~340,000 gallons of oil spilled into Guanabara Bay, severely polluting the upper reaches of the bay with polycyclic aromatic hydrocarbons (PAHs) (Gabardo et al., 2000; Farias et al., 2008; Massone et al., 2013). After two years, sediments in the inner bay were highly contaminated with PAHs, while water samples no longer showed toxicity effects (Silva et al., 2007a). After four years, PAHs decreased by 70%, showing that climatic variations over time can be efficient in hydrocarbons degradation (Farias et al., 2008) and that water circulation was somehow effective in the spill area. Another Diesel oil spill (60,000 L) accident occurred in 2005, and PAHs in tissues of C. parallelus, M. liza, G. genidens, Brevoortia aurea (Clupeidae), and M. furnieri peaked when rainfall inuced the runoff of adjacent streams and left the bay exposed to oil from other sources (Silva et al., 2007b; Soares-Gomes et al., 2010).
In Guanabara Bay, M. furnieri and M. liza presented relatively high levels of polychlorinated biphenyls (PCB) and low levels of polybrominated diphenyl ethers (PBDE) when compared to other ecosystems around the world (Gonçalves da Silva et al., 2013). Croakers presented the highest PCB and PBDE levels, while mullets showed intermediary levels. For C. parallelus, M. liza, T. lepturus, and Sardinella brasiliensis (Clupeidae) the levels of PCBs and organochlorine pesticides (OCs) are below the maximum limit established by the Food and Drug Administration (U.S. Environmental Protection Agency-EPA) and, apparently, pose low risk to human consumption (Silva et al., 2003).
Environmental and Fish Contamination With Metals
According to 210Pb dating, increases in heavy metal flux started at the beginning of the last century in Guanabara Bay (Baptista Neto et al., 2013). Sediments are affected by Cr, Pb, and Zn, being the northwestern bay and the Rio de Janeiro harbor the most polluted sectors (Abreu et al., 2016). Sources of metals are related to urban emissions, continental sources, biogeochemical processes, and a chlor-alkali plant at the western portion of the estuary that was responsible for mercury-contaminated effluents entering the bay (Wasserman et al., 2000; Cordeiro et al., 2015; Aguiar et al., 2016). Hg peaks in the western bay, while the northwestern bay is not severely contaminated (Machado et al., 2008). Dredging can pose additional risk of Hg contamination (Silveira et al., 2017), however mangrove forests are biogeochemical barriers accumulating metals and promoting the reduction of metal transport by tidal currents (Kehrig et al., 2003; Borges et al., 2007).
Metal contamination in waters and sediments were also assessed in the Amazon Complex (Lima et al., 2017), Sepetiba Bay (Fonseca et al., 2013), Paraguaçu Estuary (Hatje and Barros, 2012), and Santos Estuarine System (Kim et al., 2016). Sources are associated with mining, industry, natural igneous rock decomposition, reduced freshwater flow, farming, and non-treated sewage (Braga et al., 2000; Paraquetti et al., 2004; Genz et al., 2008).
Bagre spp., M. liza, M. furnieri, C. undecimalis (Kehrig et al., 1998, 2002; Baêta et al., 2006) and T. lepturus (Seixas et al., 2012) are still not severely contaminated with heavy metals in Guanabara Bay. Livers of M. furnieri, Bagre spp. and M. liza accumulates higher loads of Hg and methil-Hg, showing relationship with total length (Kehrig et al., 2002, 2009; Baêta et al., 2006). In Sepetiba Bay, M. liza, C. leiarchus, M. furnieri, and G. genidens had their muscles, gonads, and livers contaminated with heavy metal above maximum permissible levels established by the Brazilian Ministry of Health for human consumption (Lima Junior et al., 2002). Hg in Guanabara and Sepetiba Bays presented positive relationship with trophic levels, suggesting that Hg is being biomagnified through the food webs (Kehrig et al., 2002; Baêta et al., 2006; Bisi et al., 2012).
In the Goiana Estuary, Hg levels in muscles of T. lepturus increased with body size and weight. Hg levels decreased when rainfall increased, through biodilution (Costa M. F. et al., 2009; Barletta et al., 2012). Such seasonal pattern was also observed in Ciénaga Grande de Santa Marta Lagoon for E. plumieri and M. incilis (Alonso et al., 2000). However, Cd, Zn, and Cu in muscle of Ariopsis bonillai (Ariidae) were highest when salinity was lower (Campos, 1992). In the Santos Estuarine System, high levels of Hg in muscle of C. spixii (Azevedo et al., 2009, 2011) and above-limits for Cu and Zn in liver of M. curema (Fernandez et al., 2014) confirmed a high anthropogenic influence.
Subtropical Southwestern Atlantic
Paranaguá Estuarine Complex: South Brazil Tropical-Subtropical Transition
The interaction between seasons and estuarine habitat shifts (ecocline) structured fish movement in the Paranaguá Estuarine Complex (Barletta et al., 2008) (Figure S11). Seasonal variations in temperature and salinity influenced the spatio-temporal use of intertidal habitats for A. brasiliensis (Carvalho and Spach, 2015). Batoids are also distributed according to seasonal variation in the salinity gradient (Possatto et al., 2016). The species C. spixii, S. stellifer, Anchoa parva (Engraulidae), Achirus lineatus (Achiridae), and G. genidens were the most abundant taxa (Barletta et al., 2008). These species preferred the upper and middle estuaries during most of the year, except A. parva, which peaked in the lower estuary during the dry season. During the late-rainy season, when rainfall increases, fishes move seawards to the middle estuary.
Strong influence of the salinity ecocline on fish distribution was also observed in a multiple sampling survey (1993–2011) in the same estuary (Passos et al., 2013). Anchoa januaria, Anchoa tricolor (Engraulidae) A. brasiliensis and H. clupeola were the most abundant species in this system. The upper estuary was inhabited by the estuarine A. januaria, while marine fishes as A. brasiliensis, H. clupeola, and A. tricolor dominated the middle and lower estuaries.
The feeding ecology of Rhinobatos percellens (Rhinobatidae) (Carmo et al., 2015), Ctenogobius shufeldti (Gobidae) (Contente et al., 2012), and A. brasiliensis (Contente et al., 2011); the reproduction and recruitment of Etropus crossotus (Paralichthyidae) (Oliveira and Favaro, 2011); and the importance of the system as nursery for Epinephelus itajara (Serranidae) (Félix-Hackradt and Hackradt, 2008) and Manta birostris (Myliobatidae) (Medeiros et al., 2015) are other information available for this estuarine complex.
Guaratuba Bay Estuary: South Brazil/Tropical-Subtropical Transition
Salinity fluctuations, hydrodynamic variability, and geomorphology of the channel structured fish assemblages in Guaratuba Bay (Vendel et al., 2010; Figure S12). Salinity, transparency, and organic matter were the most important factors distinguishing habitats. A. januaria, A. lepidentostole, O. oglinum, A. brasiliensis, S. greeleyi, and A. lyolepis were all abundant species. The upper-inner estuary was characterized by A. januaria, while the lower-outer estuary was characterized by O. oglinum, A. lyolepis, A. lepidentostole, S. greeleyi, and A. brasiliensis.
In mangrove areas, Haemulopsis corvinaeformis (Haemulidae), and S. rastrifer were the most representative species (Chaves and Vendel, 1997; Chaves and Bouchereau, 1999; Chaves and Corrêa, 2000) and, E. argenteus, E. melanopterus, B. ronchus, and Citharichthys arenaceus (Paralichthydae) were common species (Chaves and Corrêa, 1998). High catches were recorded for winter months, when temperature decreased and salinity increased (Chaves and Bouchereau, 1999). However, mangroves are spawning areas for ~40% of fishes species, especially in summer and spring months, when maturation and spawning become evident (Chaves and Vendel, 1997; Chaves and Bouchereau, 2000). However, for other species as C. parallelus, spawning and hatching are associated to stronger marine influence, while juvenile recruitment associates to salinity declines during the rainy season, when river runoff increases (Chaves and Nogueira, 2013). Mangroves, salt marshes (Chaves and Vendel, 1996, 2008) and adjacent rivers (Costa P. V. et al., 2009) are also feeding grounds for several taxa.
Babitonga Bay Estuary: South Brazil/Subtropical
Depth and salinity structured the spatio-temporal variability of dominant species in Babitonga Bay (Figure S13). Engraulidae, Eucinostomus spp. and Mugil spp. dominated shallow fish assemblages in abundance, followed by A. brasiliensis, A. januaria, A. tricolor, H. clupeola, and Oligoplites saliens (Carangidae) (Vilar et al., 2011). The marine species H. clupeola, O. saliens, A. tricolor, and T. carolinus were characteristic of the outer bay, while estuarine-residents and marine-migrants (A. brasiliensis, A. januaria, S. greeleyi, C. spilopterus and D. rhombeus) were characteristic of the inner bay. Thespecies T. carolinus peaked in the warm rainy season; M. furnieri in the transition season; and O. saliens in the cold dry season.
The first occurrence of larval Microdesmus longipinnis (Microdesmidae) (Souza-Conceição et al., 2013) and the distribution of larvae of the invasive species Omobranchus punctatus (Blennidae) were assessed in Babitonga Bay (Costa et al., 2011). Sciaenidae larvae (B. ronchus, Cynoscion sp., M. furnieri) are abundant and distributed along the entire bay, and peak in abundance in spring and summer months (Costa et al., 2012). Larval Lycengraulis grossidens (Engraulidae) also exhibit spatio-temporal patterns within the Bay (Costa and Souza-Conceição, 2009).
Patos Lagoon Estuary: South Brazil/Subtropical
Salinity and temperature explained most of the variability in a long-term assessment of shallow-water estuarine-dependent fishes in the Patos Lagoon Estuary (Garcia et al., 2012; Figure S14). The westerly wind belt movement, or Southern Annular Mode, influenced variations occurring at a scale of 2 years, while rainfall anomalies due to ENSO events influenced inter-annual variability at scales of 3–7 years. Mugil liza, Brevoortia pectinata (Clupeidae), and M. furnieri occurred year-round. Mugil gaimardianus (Mugilidae), M. curema, and B. pectinata were associated with higher salinities and temperatures. Mugil liza and M. furnieri associated with lower salinities and temperatures.
Shallow-water marine fishes peaked in the 1995/1996 La Niña due to low precipitation rates and freshwater runoff; decreasing in the 1997/1998 El Niño when precipitation and river discharge increased (Garcia et al., 2001, 2004). Anchoa marinii (Engraulidae), A. brasiliensis, M. liza, M. platanus, M. curema, and Jenynsia multidentata (Anablepidae) were frequent species. Estuarine-resident fishes increased upstream in La Ninã situation, and showed opposite patterns in El Niño. Estuarine-dependent fishes also showed contrasting patterns (Garcia et al., 2001). The species A. brasiliensis and M. platanus peaked in shallow waters. Mugil liza peaked in La Niña and lowered in El Niño. During El Niño, sub-adults and adults of A. brasiliensis were absent from upstream shallow waters due to the high freshwater discharge (Garcia et al., 2001), while the freshwater fishes Astyanax eigenmaniorum, Oligosarcus jenynsii (Characidae), and Parapimelodus nigribarbis (Pimelodidae) dominated the upstream reaches (Garcia et al., 2001, 2003).
Age, growth, and reproductive aspects of Mycteroperca marginata (Epinephelidae) (Seyboth et al., 2011), Odontesthes argentinensis (Atherinopsidae) (Moresco and Bemvenuti, 2006), M. liza (Garbin et al., 2014), and Jenynsia multidentata (Anablepidae) (Garcia et al., 2004) were also studied for this system. New species belonging to the families Loricariidae (Carvalho et al., 2008; Rodriguez and Reis, 2008) and Gobiidae (Burns et al., 2010; Cheffe et al., 2010), and reports on the occurrence range and competition of the invasive Acestrorhynchus pantaneiro (Acetrorhynchidae) (Neuhaus et al., 2016) are also available for the lagoon. Long-term assessments of ichthyoplankton are also available and must be incorporated into conservation planning (Martins et al., 2007; Costa et al., 2016; Costa and Muelbert, 2017).
Population genetics revealed that many species in the lagoon and adjacent systems are in process of speciation, suggesting that managerial actions must consider different sub-populations (Beheregaray and Levy, 2000; Da Silva Cortinhas et al., 2016).
The ariid white sea catfish Genidens barbus was once an important fishery resource in the estuary, where the species had the most abundant population known in the past (Velasco et al., 2007). Therefore, studies on this species became important for the region. Description of growth parameters using length and age data (Velasco et al., 2007) and the use of estuarine carbon sources revealed by isotope analyses (Pereyra et al., 2016) report biological aspects for the species. Ecological aspects were revealed by otolith microchemistry and patterns of estuarine uses (Avigliano et al., 2015), inter-annual variability (Avigliano et al., 2017a), freshwater residence times (Avigliano et al., 2017b), nursery grounds and connectivity (Avigliano et al., 2016) are available for the species.
Micropogonis furnieri is a common species in the Patos Lagoon and represents an economically important fish for the artisanal fishery (Costa et al., 2015). Several aspects of the biology and ecology of this species is also available regarding the system. Feeding ecology studies using different aspects, as daily consumption (Figueiredo and Vieira, 2005), feeding strategies (Mendoza-Carranza and Vieira, 2008), and stable isotope inferences are available (Mont'Alverne et al., 2016). Otolith microchemistry was used to reveal aspects of growth and age of young individuals (Cavole and Haimovici, 2015). In addition, studies have emphasized the importance of ontogenetic approaches while studying fish movement, since early life stages of M. furnieri have multiple and complex habitat preferences, suggesting that essential fish habitats must be explored for species with complex life cycles to improve management and conservation planning (Costa et al., 2014, 2015).
Environmental Impacts in Subtropical West Atlantic Estuaries
Habitat Changes
The largest Latin America maritime terminal for agro-industrial products (e.g., soy beans) is located in the Paranaguá Estuarine Complex and Pontal do Felix port is a recent additional structure built in the upper estuary. Dredging operations for maintenance of the shipping channel have altered its geomorphology resulting in upstream intrusion of coastal waters and changes in fish communities (Barletta et al., 2016). During the dredging process, total mean density and biomass of C. spixii, Aspistor luniscutis (Ariidae) and G. genidens increased, while Menticirrhus americanus (Sciaenidae), S. rastrifer and C. leiarchus decreased significantly in the dredged channel. Ariid catfishes were favored by the damage of the benthic fauna, while other species as sciaenids disappeared. Future dredging operations must be avoided during reproductive (October to December) and recruitment (April to June) seasons of fishes, which occurs in the late-rainy season. Therefore, dredging in the Paranaguá Estuary should be performed during the dry season (July to September) to promote sustainable fishery practices (Barletta et al., 2016).
The expansion of the Rio Grande Port and jetties at the entrance of the Patos Lagoon Estuary has changed sediment deposition patterns (texture and distribution) in the estuarine and coastal regions (Cunha and Calliari, 2009; Silva et al., 2015). Moreover, the construction of a dam between the estuary and the Patos-Mirim lagoon to prevent the entrance of salt water resulted in the absence of estuarine and marine species upstream (Burns et al., 2006).
Sewage Contamination and Eutrophication
In Paranaguá Bay (Martins et al., 2010; Cunha et al., 2011; Brauko et al., 2016), Babitonga Bay (Martins et al., 2014) and Patos Lagoon (Martins et al., 2007), fecal steroids are higher close to ports and urban areas pointing to a significant sewage contamination of waters and sediments. In Guaratuba Bay, signs of eutrophication due to organic and inorganic matter inputs are still partially controlled by precipitation and tidal currents (Mizerkowski et al., 2012; Rodrigues et al., 2013). Phosphorus is higher in sediments from the upper estuary and in the transition between the upper and middle estuary (Cotovicz Junior et al., 2014). Increases in nutrient inputs, water temperature and salinity induce blooms of harmful diatoms in the bay, poisoning the biota and causing human intoxication (Tibiriçá et al., 2015).
Plastic Contamination
Contamination with plastic debris was assessed only in the Paranaguá Estuarine Complex, being more significant near urbanized areas in the upper and middle reaches. It is affected by local hydrological processes, with no seasonal or spatial trends along the salinity gradient (Possatto et al., 2015).
Environmental and Fish Contamination With Persistent Organic Pollutants
In November 2004, an oil tanker spilled methanol and bunker oil in front of Paranaguá harbor and the osmoregulation of A. brasiliensis was affected until seven months after the accident (Souza-Bastos and Freire, 2011). Tributyltin (TBT) and organic contaminants of antifouling paints in liver were reported for C. spixii, with higher levels closer to Paranaguá harbor (Santos et al., 2014). Furthermore, a model revealed that PAHs adsorbed in sediments and solubilized in water can be possibly transferred to fishes and biomagnified along the trophic chain of the estuarine complex (Froehner et al., 2011).
In Guanabara Bay, aliphatic hydrocarbons and linear alkylbenzenes inputs are related with geochemical processes. Higher loads were observed during summer months due to increased summer holidays (Dauner and Martins, 2015). OCs and PCBs had low concentrations due to low inputs from the Germany rivers discharging in the bay (Combi et al., 2013). In Babitonga Bay, PCBs and diclorodifeniltricloroetano (DDT) also presented high concentration at specific sites close to anthropogenic impacted areas of São Francisco harbor (Rizzi et al., 2017).
For the Patos Lagoon Estuary, hydrocarbon pollution is related to combustion of fossil fuels, release of oil, industrial and domestic effluents. Higher concentrations in sediments are observed near refineries and oil terminals, shipping lanes and sewage discharges (Medeiros et al., 2005; Garcia et al., 2010).
Environmental and Fish Contamination With Metals
Sediments in Paranaguá (Anjos et al., 2012) and Guaratuba Bays (Sanders et al., 2008) are contaminated with metals, sometimes above World Health Organization (WHO) critical limits. Metal contamination in water is widely reported for the Patos Lagoon Estuary (Windom et al., 1999; Mirlean et al., 2001; Costa and Wallner-Kersanach, 2013). Metals presented higher concentrations close to urban and industrial areas, being human inputs and geochemical anomalies the main identified sources of these pollutants (Niencheski et al., 1994; Sá et al., 2006; Anjos et al., 2012; Costa et al., 2013).
In the Paranaguá Estuarine Complex, muscles of C. spixii and G. genidens are contaminated by metals, with levels of Cr and As exceeding the permissible limits for seafood consumption (U.S. EPA) (Angeli et al., 2013). In Babitonga Bay, levels of Zn and Cr in muscles of E. brasilianus, C. paralellus, and M. platanus are still acceptable for human consumption (Bonatti et al., 2004). However, in the Patos Lagoon Estuary, Hg levels in M. furnieri, N. barba, G. genidens, and Odontesthes bonariensis (Atherinopsidae) were above background levels, but below critical levels for human consumption (U.S. EPA) (Niencheski et al., 2001; Kutter et al., 2009).
Warm Temperate Southwestern Atlantic
Río de La Plata Estuary: Subtropical/Temperate Transition
A comprehensive overview of fish diversity and the environmental influence on fish distribution and composition is available for the Río de La Plata Estuary (Jaureguizar et al., 2016; Figure S15). Salinity structured spatial patterns of larval fishes during austral late spring (December 1999) and summer (February 2000) (Berasategui et al., 2004). The horizontal salinity gradient, bottom salinity and water temperature are the main factors structuring species composition across seasons (Jaureguizar et al., 2003b, 2004; Lorenzo et al., 2011). Micropogonias furnieri, C. guatucupa, B. aurea, M. schmitti, and M. ancylodon are abundant species. The freshwater and shallow upper estuary was typified by P. valenciennesi and N. barbus (Jaureguizar et al., 2004). The middle estuary contained the highest densities of M. furnieri, B. aurea, M. ancylodon, where depths and salinities had intermediate values (Jaureguizar et al., 2004). Density of M. furnieri and M. ancylodon were higher in summer, while density of B. aurea increased during winter and autumn. The saltier and deeper lower estuary was dominated by C. guatucupa, M. schmitti. Larger catches of C. guatucupa were observed in autumn, while M. schmitti during winter and spring (Jaureguizar et al., 2004).
Studies regarding ontogenetic diet shifts in Urophycis brasiliensis (Phycidae) (Acuña-Plavan et al., 2007), first records of larval Elops smithi (Elopidae) (Machado et al., 2012) and adult E. melanopterus (Gerreidae) (Solari et al., 2010), phylogenetic approaches for the endemic Pimelodus albicans (Pimelodidae) (Vergara et al., 2008) and aspects of reproduction in B. aurea (Macchi and Acha, 2000) are available. However, most studies have focused on Sciaenidae species, especially the most abundant and commercially interesting ones (M. furnieri, C. guatucupa, Pogonias cromis, and Macrodon ancylodon).
The species P. cromis spawn between October and January (Macchi et al., 2002), M. ancylodon between October and March (Militelli and Macchi, 2006), C. guatucupa in March and December (Militelli and Macchi, 2006), and M. furnieri between November and March (Macchi et al., 2003). Young age-classes of C. guatucupa decreases seawards, with a reverse pattern in spring, while older age-classes inhabit marine coastal areas (Jaureguizar et al., 2006). Short-term changes in oceanographic conditions have greater influence structuring C. guatucupa populations than long-term variability (Jaureguizar and Raúl, 2009). During 1998 El Niño and 1999 La Niña, adults were associated with high salinities, when the area was dominated by wind driven inflow of seawater. During typical years (1994 and 2003), juveniles correlated with low salinities, when winds forced freshwater to flow seawards (Jaureguizar and Raúl, 2009).
Selective processes differentiate populations of M. furnieri between the estuary and adjacent coastal locations, and the influence of salinity and temperature in phenotypic cohesion can structure these populations (D'Anatro et al., 2011; D'Anatro, 2017). Larvae of M. furnieri are abundant during the warmest months, showing high predominance in the river-estuarine transition and positive correlation with bottom salinity horizontal gradient (Braverman et al., 2009). Ontogenetic approaches revealed a complex use of the estuary by M. furnieri, based on the bottom salinity gradient (Jaureguizar et al., 2003a, 2008). Spawning occurs in the innermost areas, near the upstream edge of the salinity wedge (Acha et al., 1999).
Pando Estuary: A Sub-estuary of Río de La Plata
For the Pando Sub-Estuary (Figure S16), M. furnieri, M. liza, P. orbignyanus, B. aurea, and Parapimelodus valenciennis (Pimelodidae) are the most abundant species (Acuña-Plavan et al., 2010; Gurdek and Acuña-Plavan, 2016). Marine migrants were correlated with higher salinity, while freshwater species peaked at lower salinity and estuarine species correlated with higher temperature. Significant correlations between abundance and temperature are related to synchronized events during species life cycles.
Length-weight relationship of 12 species, including the abundant ones, are available for this sub-estuary (Gurdek and Acuña-Plavan, 2014). For the most abundant species M. furnieri, the length-weight relationship was assessed along its ontogeny over an intra-annual cycle (Gurdek and Acuña-Plavan, 2016). This species produces the seasonal courtship/spawning sounds from November to March (spawning season), with a strong seasonal variability, appearing in October, peaking in January-March, and disappearing in April (Tellechea et al., 2011).
Mar Chiquita Coastal Lagoon: Temperate Southwest Atlantic, Argentina
In Mar Chiquita Lagoon (Figure S17)., B. aurea, O. argentinensis, M. liza, and M. furnieri are abundant species, especially in summer months (Bruno et al., 2013). Juveniles of Platanichthys platina and Ramnogaster arcuata (Clupeidae) are also frequent (González Castro et al., 2009). Marine fishes peaked in the lower estuary when temperature increased in summer and autumn. Brevoortia aurea was the most abundant species in the southern-lower portion, and O. argentinensis in the middle and northern-upper portion of this estuary (Bruno et al., 2013). Juvenile recruitment is ruled by seasonal onshore winds (Bruno et al., 2015). However, salinity and temperature structured fish assemblages (González Castro et al., 2009). Brevortia aurea peaks in higher salinity and temperatures. The species M. liza and O. argentinensis correlated with low salinities. Peaks of O. argentinensis occurr at lower temperatures, while M. furnieri peaks at higher temperatures. The occurrence of M. curema (González Castro et al., 2006) and T. carolinus (Díaz de Astarloa et al., 2000), both with tropical-subtropical affinities, in the temperate lagoon is related to the presence of warm neritic waters from the continental shelf originated in subantarctic waters of the Malvinas Current.
Anchoa marinii (Engraulidae) reproduces between December and April when temperature increases and mature females aggregates in adjacent coastal areas (López et al., 2015). The species M. liza migrates seawards in April-May and November-December to spawn (González Castro et al., 2011). Aggregations of mature females of B. aurea can be observed in October-November in the lower estuary (Lajud et al., 2016).
Bahía Blanca Estuary: Temperate Southwest Atlantic, Argentina
For the Bahía Blanca Estuary (Figure S18), the reproductive aspects of Sympterygia acuta (Rajidae) (Díaz-Andrade et al., 2009) and the feeding ecology of R. arcuata (Lopez Cazorla et al., 2011) are described. In addition, the influence of biotic and environmental factors on seasonal patterns of juvenile fishes in Spartina alterniflora saltmarsh and a contiguous tidal flat in the Bahía Blanca Estuary was also evaluated (Valiñas et al., 2012). However, fish research is focused on three economically important species: M. furnieri, C. guatucupa and P. orbignyanus (Blaber and Barletta, 2016).
The age structure of the population of C. guatucupa was revealed by otolith analyses (Lopez Cazorla, 2000), while for P. orbignyanus, age structure was revealed by scales reading and growth parameters (Lopez Cazorla, 2005). Seasonal diet shifts and ontogenetic changes in the feeding ecology of the three species (Lopez Cazorla and Forte, 2005; Sardiña and Lopez Cazorla, 2005a,b), as well as dietary overlaps between the two co-occurring sciaenid species (Sardiña and Lopez Cazorla, 2005c) were assessed within the system. Foraging activities of M. furnieri have direct and indirect effects on the granulometric composition and stability of bottom sediment of salt marshes (Molina et al., 2017).
Environmental Impacts in Warm Temperate Western Atlantic Estuaries
Habitat Changes
Habitat modifications in Río de La Plata Estuary include shoreline retreats due to deforestation, tourism activities, and land development (Cellone et al., 2016). Urban beach erosion and accretion have strong relationships with climate changes, sea level rise, and increased storminess (Gutiérrez et al., 2016).
Various ports, cities, livestock, agriculture and oil, chemical, and plastic industries are located in Bahía Blanca Estuary (Spetter et al., 2015). The estuary has one of the largest deep water ports in the country (Ingeniero White port), which is regularly dredged (Zilio et al., 2013). A project to deepening a navigation channel in the inner estuary to a depth of 13.5 m related to a natural gas provision have been pointed to cause physical and social consequences to the system. The loss of nursery services and of the jobs associated with fishing activities, cease of recreational activities during the dredging process, the loss of the coastal landscape, water pollution derived from sediment removing and the effects on groundwater are imminent possibilities (Zilio et al., 2013). Increased erosion of coastal terraces in response to rising sea level, increased land use in the harbor area and aggradation due to dredged spoils deposition are also observed in Bahía Blanca (Pratolongo et al., 2013).
Sewage Contamination and Eutrophication
The Río de La Plata Estuary is moderately eutrophic. Increases in freshwater runoff and nutrient loads, associated to a low potential to dilute and flush nutrients, suggest that the system is prone to worsening eutrophication conditions generating further dissolved oxygen stress and harmful algal blooms (Nagy et al., 2002). Sewage effluents pose a potential ecotoxicological risk to aquatic biota due to inputs of estrogens (Valdés et al., 2015). In the Bahía Blanca Estuary, sewage and industrial discharges receive poor or no treatment before reaching the system (Pierini et al., 2012). Microbiological contamination was revealed by the presence of Escherichia coli and Samonella spp. in water and sediments close to sewage discharges in the Rosales Harbor and tributaries discharging into the estuary (Spetter et al., 2015; Streitenberger and Baldini, 2016).
Environmental Contamination With Plastic
Plastic contamination has been assessed since 1999 by the National Direction of Aquatic Resources in the Río de La Plata Estuary and by the Ocean Conservancy's International Coastal Cleanup Day in Uruguayan aquatic systems. Most plastic debris comes from urban wastes, waterways, and marine traffic in Uruguayan waters (Lozoya et al., 2015). The bottom salinity front of the salt-wedge is the main barrier accumulating plastic debris upstream (Acha et al., 2003). In the southern coast of the system, 100% of freshwater fishes belonging to eleven species were contaminated with microplastics in their gut contents, especially fibers, with higher numbers near sewage discharges, where they varied from 30 to 89 fibers per stomach (Pazos et al., 2017).
Environmental and Fish Contamination With Petroleum Hidrocarbons Persistent Organic Pollutants
High levels of aliphatic and aromatic hydrocarbons in waters, sediments, soils and biota were observed after ~1,000 tons of oil spilled in coastal waters of Río de La Plata (Colombo et al., 2005a,b). Petroleum direct inputs and combustion, harbor activities, and vehicular emissions are the main sources of hydrocarbons to this system (Venturini et al., 2015). PCBs have highest concentrations near industrialized areas close to Buenos Aires, and transformers oil containing Aroclor 1,254–1,260 are the probable sources (Colombo et al., 2005c).
PAHs, PCBs, and PBDEs in sediments of the Bahía Blanca Estuary had higher concentrations near urban and industrial areas (Arias et al., 2010; Oliva et al., 2015; Tombesi et al., 2017). TBT and dibutyltin (DBT) concentrations were higher in sediments near Puerto Belgrano harbor (Argentina's Army), where there are intense shipyard activities (Delucchi et al., 2007). Levels of OCs were moderate, when compared to worldwide ranges, and showed a high correlation with precipitation in sites near agricultural fields (Arias et al., 2011).
In Río de La Plata, Cyprinus carpio (Cyprinidae), Mugil cephalus (Mugilidae), O. bonariensis, and Prochilodus lineatus (Prochilodontidae) showed moderate to high levels of aliphatic hydrocarbons and PCBs in their muscles with signs of bioaccumulation (Colombo et al., 2000, 2007a,b; Menone et al., 2000; Cappelletti et al., 2015). The highest concentrations were recorded in P. lineatus, especially near the urban center, exceeding guidelines for human consumption (U.S. EPA) (Colombo et al., 2000; Speranza et al., 2016). In the Bahía Blanca Estuary OCs contaminated C. guatucupa with a size-related bioaccumulation pattern (Lanfranchi et al., 2006). PAHs were reported to contaminate Odontesthes sp. in the system; and the global average for this pollutant indicated that Bahía Blanca is chronicaly polluted (Arias et al., 2009, 2010).
Environmental and Fish Contamination With Metals
Metal contamination in sediments and water were assessed in Río de La Plata Estuary (Tatone et al., 2013, 2015), Mar Chiquita Lagoon (Marcovecchio et al., 2001; Beltrame et al., 2009), and Bahía Blanca Estuary (Botté et al., 2007; Grecco et al., 2011). Sources and highs levels of metals are related to polluted discharges from agricultural, urban and industrial sources (Camilión et al., 2003; Marcovecchio et al., 2016; Santucci et al., 2017).
In Río de La Plata, Pterodoras granulosus (Doradidae), P. lineatus (Villar et al., 2001), M. platanus, and M. furnieri are contaminated with metals with signs of bioaccumulation. Hg is the most important contaminant for M. furnieri, however values are still below the international standards (U.S. EPA) recommended for suspending human consumption (Corrales et al., 2016). Hg levels in edible muscle of fishes from Mar Chiquita Lagoon are also safely below recommendation standards (Marcovecchio et al., 2001; Marco et al., 2006). In the Bahía Blanca Estuary, low levels of Cd and Zn were observed in top predators M. schmitti and Halaelurus bivius (elasmobranchii), however Hg levels exceeded the international standards (Marcovecchio et al., 1986, 1988a,b, 1991) and also showed signs of bioaccumulation. Livers of Brevortia aurea, O. argentinensis, M. furnieri, C. guatucupa, M. schmitti, and P. orbignyanus are also contaminated with metals. At least one sample of each species presented concentrations of Mn and Cr hazardous to humans (La Colla et al., 2017).
Eastern Pacific Estuaries
Tropical Eastern Pacific
Bahía málaga estuary: Colombia/Panamá Bight Ecoregion
At least 237 species inhabit the Bahía Málaga Estuary (Figure S19) (Artunduaga, 1978; Rubio, 1984a,b; Castillo, 1986; Castellanos-Galindo et al., 2006). The combination of tidal and dial cycles explained shifts in fish communities, but a biomass decrease was notable when rainfall increased at the end of the annual cycle (Castellanos-Galindo and Krumme, 2013). Spatial and temporal patterns of larval fish community along the main channel of the estuary was also assessed during an annual cycle (Medina-Contreras et al., 2014). Seriola sp. (Carangidae) and Cetengraulis mysticetus (Engraulidae) were the most abundant larvae. Salinity or temperature were not correlated with larval density, but larvae varied greatly among months, suggesting a strong influence of seasonality in the bay (Medina-Contreras et al., 2014).
The composition and diversity of intertidal fishes were assessed in Isla Palma, in the lower estuary (Castellanos-Galindo et al., 2005). Freshwater fishes are known to reach Isla Palma due to high rainfall rates influenced by the Intertropical Convergence Zone (Cordoba and Giraldo, 2014). Clupeidae were the most abundant family, in terms of numbers, inhabiting intertidal mangrove habitats in the innermost portion of the estuary, however Lutjanidae, Tetraodontidae and Ariidae dominated in weight (Castellanos-Galindo and Krumme, 2013).
Studies on the feeding ecology of Centropomus unionensis (Centropomidae) (Mancilla and Rubio, 1992), and diet, growth and reproduction of Lutjanus guttatus (Lutjanidae) (Suárez and Rubio, 1992a,b) are also available. According to a trophic flow model performed in the inner portion of the bay, the very low salinity throughout the year is responsible for the low number of primary and secondary consumers (e.g., zooplankton, crustaceans and some fishes) in the mangrove system. Therefore, zoobenthivorous (snappers, catfishes) and detritivorous (mullets) fishes dominate in biomass, when compared to piscivorous and zooplanktivorous fishes (Castellanos-Galindo et al., 2017).
Rio palmar and rio javita estuaries: Ecuador/Guayaquil Ecoregion
Fish communities were compared between the dry and the wet season in mangrove creeks and main channel of Palmar and Javita rivers (Shervette et al., 2007; Figure S20). The diversity of species belonging to the families Gobiidae, Gerreidae and Engraulidae is high in the mangrove system of Palmar Estuary, while species of Carangidae, Engrauliae, and Gerreidae are diverse in the Javita main channel. Significant differences were detected among areas and seasons for both systems, and the percent of mangroves and mean depth influenced fish communities. Although these estuaries present low species diversity compared with other tropical estuarine systems (Blaber, 2000), they still provide important nursery habitats for many fishes of commercial and ecological importance (Shervette et al., 2007).
Environmental Impacts in Tropical Eastern Pacific Estuaries
Habitat Changes
Not more than 4,000 people, distributed in small villages, live in Bahía Málaga (Castellanos-Galindo and Krumme, 2013). However, over the last 25 years, Bahía Málaga has faced anthropogenic changes relative to the construction of a naval base in the mid 1980's and an increased tourism activity nearby (Castellanos-Galindo et al., 2011). This resulted in mangrove loss, but no specific estimates are available. Governmental agencies plan to construct a deep-water commercial harbor, what might be an important threat to marine and estuarine fish diversity. Nevertheless, La Plata (~6,791 ha) and La Sierpe (~25,178 ha) areas have recently been declared protected areas by local environmental agencies.
Sewage Contamination and Eutrophication
Water quality assessments asserted that Bahía Málaga is still in good conditions with no signs of eutrophication or changes in physic-chemical parameters (Betancourt Portela et al., 2011). However, further chemical pollution assessments are necessary for the region, including their fate for the local biota.
Temperate Eastern Pacific
Valdivia River Estuarine System: Chile/Araucanian/Chiloense Ecoregion
A study on the migratory patterns of Galaxias maculatus (Galaxiidae) from larval to adult stage revealed a strong association with seasonal changes in salinity along the Valdivia estuary (Hugo, 1973; Figure S21). Another study in Corral Bay detected that higher densities of larvae were observed near the tidal front, and differences were related to tidal intrusion of salt water (Vargas et al., 2003). According to patterns of circulation, Corral bay is a source of young fish larvae. Strangomera bentincki (Clupeidae), Odontesthes regia laticlavia (Atherinopsidae), Gobiesox marmoratus (Gobiesocidae), and Hypsoblennius sordidus (Blenniidae) were the most abundant larval species.
The seasonal influence on diatery ontogenetic shifts of the Chilean silverside O. regia was assessed in the Valdivia Estuarine System and this species is acknowledged as a selective omnivorous predator (Fierro et al., 2014). In the Corral bay, the feeding ecology of Myxodes viridis (Clinidae) was also assessed in different depths and tidal cycles (Ochoa-Muñoz et al., 2013).
Environmental Impacts in Temperate Eastern Pacific Estuaries
Environmental Contamination With Persistent Organic Pollutants
PAHs in sediments of Corral Bay showed a medium pollution rate with a temporal variation, with a substantial increase observed from March to September during 2000 (Palma-Fleming et al., 2004). Aliphatic hydrocarbons were at low to medium contamination rate with no temporal variation, and inputs are relative to the petroleum hydrocarbon-diesel fraction and biogenic hydrocarbons (Palma-Fleming et al., 2012). Such impacts are related to control of prague, fluvial wood transportation, wood and paper industries, shipyards, fishing industries, and salmon conditioning jails.
Environmental Contamination With Metals
Dramatic changes in the Valdivia River Estuarine System are related to wastewater and solid emissions from industrial or domestic activities, aerial emissions, farming/agriculture, and oil spills (Palma-Fleming et al., 2012). Cd level within the Corral Bay was lower than in other Pacific coastal areas, but increased upstream rivers as salinity decreased (Pinochet et al., 1995). Cu and As in sediments were above standards of the Sediment Quality Guidelines (U.S. EPA) in all sampling points of Corral Bay, being an indicative of toxicological effects for the biota (Palma-Fleming et al., 2012).
Discussion
The Importance of the Ecocline and Seasonality Concepts to Study Fish Movement and Environmental Quality Assessments
Estuaries were long poorly understood because of their complex natural processes and only recently, over the past four decades, fully recognized as a key coastal ecosystem (Elliot and Whitefield, 2011). Therefore, studies regarding the estuarine ecocline concept and the influence of environmental variability on fish movement and environmental changes are still missing even in large and important systems of the Western Atlantic and along all eastern Pacific coast of SA (Blaber and Barletta, 2016). The influence of the estuarine ecocline on fish movement were discussed in the Amazon, Caeté, Guanabara, Santos, Guaratuba, Babitonga, Pando and Mar Chiquita systems. In Ciénaga Grande, Goiana, Paraguaçu, Sepetiba, Paranaguá and Patos Lagoon systems, not only fishes, but also environmental quality (water and sediments) were assessed regarding the influence of the ecocline. However, the main concern is that in most surveys, fishes and environmental quality were short-term assessed, and/or across limited spatial scales, regardless the annual variability of the salinity gradient, as in Cienaga Grande Lagoon (Rueda and Defeo, 2003), Paraguaçu Estuary (Hatje and Barros, 2012), Guaratuba Bay (Vendel et al., 2010), and Río de La Plata (Jaureguizar et al., 2003b). Therefore, ecological patterns, biological behavior and physico-chemical processes might still be obfuscated by the lack of annual cycles, long-term, and full lenght approaches. Thus, efforts for conservation, recovery and sustainable use of estuarine resources were lead by poorly informed managerial actions (Barletta et al., 2010).
Systems in the tropical Northwestern Atlantic have recently gained attention. Lists of species are available for the Atrato River Delta (Correa-Herrera et al., 2016, 2017) and Orinoco River Delta, on the Caribbean coast (Cervigón, 1985; Blaber, 2000; Blaber and Barletta, 2016) (Figure 1, Table 1). In the Atrato Delta, spatio-temporal patterns of fish larvae and their densities, comparable to microplastics available in the water, were also assessed (Correa-Herrera et al., 2017). In Northeast Brazil, few fish and human impacts studies are available for the Parnaíba River Delta (Oliveira, 1974; Watanabe L. A. et al., 2014; Ribeiro et al., 2017), Potengi River Estuary (Oliveira et al., 2011; Buruaem et al., 2013; Souza et al., 2016) and Itamaracá Estuarine System (Ekau et al., 2001; Vasconcelos Filho et al., 2003, 2010). In Southern Brazil, the Laguna Estuarine Complex has intense artisanal fisheries activities and an industrial fleet focused on mullets for most of the year (Barletta et al., 2017b). However, fish research and human impacts along the ecocline were never evaluated.
“An ecocline represents a boundary of progressive change between two systems, representing the response to the gradual difference in one major environmental factor acting at a different scales and influencing the total differences within the gradient” (Attrill and Rundle, 2002). Within any estuarine system, the salinity is the major environmental factor referred in the ecocline concept (Barletta and Dantas, 2016). Hence, researchers have proposed that the proper management of estuaries are dependent upon reliable biological and abiotic data encompassing multiple aspects of space and seasonality along the estuarine gradient (Machado et al., 2016; Barletta et al., 2017a; Underwood et al., 2017).
Regarding fish research, accurate sampling design using monthly surveys replicated along different reaches and encompassing several aspects of seasonality are available for the Caeté Estuary (Barletta et al., 2005), Goiana Estuary (Dantas et al., 2013) and Paranaguá Estuarine Complex (Barletta et al., 2008, 2016). These provided reliable data on fish movement over annual cycles on which managerial reccomendations must be build. In European Atlantic estuaries (Vetemaa et al., 2006; Martinho et al., 2007), as well as in North-American Atlantic estuaries (Love and May, 2007; Granados-Dieseldorff and Baltz, 2008), accurate sampling designs for fish research are also available as reference to be replicated in any estuarine system.
In addition, the spatio-temporal variability of the estuarine ecocline influences patterns of use of essential habitats by the different ontogenetic phases of a fish species. Therefore, the ontogenetic approach is another important point of view while studying fish movement (Acuña-Plavan et al., 2007; Ferreira et al., 2019). Very few studies worldwide address ontogeny, such as in estuaries from Australia (Taylor et al., 2006), North America (Stehlik and Meise, 2000), South Africa (Harris et al., 1999), and Asia (Lin et al., 2007). In SA, fish ontogeny was assessed in Río de La Plata (Acuña-Plavan et al., 2007), Pando (Gurdek and Acuña-Plavan, 2016), Bahía Blanca (Lopez Cazorla and Forte, 2005; Sardiña and Lopez Cazorla, 2005a,b), and Goiana (Dantas et al., 2010, 2015; Ramos et al., 2016) estuaries. These studies asserted that the ecological unit is not the species in their own, but their different ontogenetic phases. Costa et al. (2014) and Ferreira et al. (2016, 2019) emphasized that the different phases of a species can have multiple and complex habitat preferences over spatio-temporal scales, and essential habitats must be explored to improve management and conservation planning.
Regarding environmental and fish contamination, accurate samplings are available for microplastics in the Goiana Estuary (Lima et al., 2014; Ferreira et al., 2016, 2019; Silva et al., 2018), for metals in Guanabara Bay (Baêta et al., 2006; Cordeiro et al., 2015) and for POPs in Río de La Plata (Colombo et al., 2000; Venturini et al., 2015). However, for most SA systems, the bioavailability and contamination of fishes with POPs, metals and microplastics were never assessed along the ecocline. Rainfall variability is a strong controller of metal and POP levels in fish tissues, since drought periods can enhance contamination and high precipitation rates diminishes the contaminants through biodilution (Costa M. F. et al., 2009; Barletta et al., 2012; Bisi et al., 2012). Fish contamination is higher near urban, industrial or rural areas and, sometimes, values exceeded the permissible limits for human consumption (U.S. EPA) (Marcovecchio et al., 2001; Delucchi et al., 2007; Fernandez et al., 2014). Moreover, the bioaccumulation and biomagnification of metals and POPs are occurring in the trophic web of most estuaries (Marcovecchio et al., 1986; Cappelletti et al., 2015). In the Negombo Estuary (Sri Lanka), for example, metal contamination of M. cephalus was year-round categorized along spatial locations and seasons providing a reliable design that can be replicated worldwide using different pollutants (Mendis et al., 2015).
Moreover, metals and POPs are somewhat controlled by other abiotic factors and their proper assessment are indeed also needed. The biogeochemistry and bioavailability of metals and POPs in estuaries are controlled by suspended particulate matter loads and dissolved oxygen levels, which in turn are strongly controlled by the salinity gradient (Janeiro et al., 2008; Bayen, 2012; Costa et al., 2012). Freshwater inputs from rivers and streams; effluents from the agriculture fields; untreated domestic and industrial sewages; oil spill and release; and combustion of fossil fuel are probable sources of metals and POPs contamination for fishes, but are hardly ever assessed (Costa M. F. et al., 2009; Barletta et al., 2012; Venturini et al., 2015). For this reason, all these sources still need further attention in SA coastal systems.
On the other hand, the inadequate disposal practices and fishery activities along river basins and coastal areas are the most obvious source of plastic pollution (Lima et al., 2014). This problem become worse when large plastics fragment into smaller particles (<5 mm), increasing the chances of interactions even during the earliest phases of a fish life cycle (Lima et al., 2015, 2016). Microplastics present higher concentrations within semi-enclosed environments and the river basin is recognized as one of the main source of microplastic inputs into estuaries, where they become ubiquitous over the year (Lima et al., 2014; Cheung et al., 2016; Lebreton et al., 2017); Vendel et al., 2017.
The problems regarding the interaction of microplastics and fishes are available for the Goiana (Ferreira et al., 2016, 2019; Silva et al., 2018) and Río de La Plata (Pazos et al., 2017) estuaries. The most complete scenario is reported for the Goiana Estuary, where the distribution patterns of microplastics were first reported (Lima et al., 2014). The estuarine ecocline acts as a control of microplastics in drier months and exporter of microplastics to the marine environment in rainy months, when runoff increases seawards (Lima et al., 2014; Lebreton et al., 2017). Microplastics have comparable densities with ichthyoplankton, emphasizing a high concentration in the main channel (Lima et al., 2015). Ingestion of plastic filaments is widespread and affect different ontogenetic phases of demersal fishes (Possatto et al., 2011; Dantas et al., 2012; Ramos et al., 2012; Ferreira et al., 2016, 2019; Silva et al., 2018). Every estuarine system in SA is experiencing this same problem of contamination due to generalized poor disposal practices (Costa and Barletta, 2015). However, “there have been very few papers describing multivariate tests of spatial or temporal patterns of microplastics” (Underwood et al., 2017).
Disruption of Estuarine Ecoclines Caused by Human Modifications in Estuarine Morphology
Human interventions in estuarine geomorphology lead to alterations in the natural inflow of salt water, and disrupt the ecocline along the whole system. Such problem is worse under the influence of industrial activities due to several months of dredging to the maintenance of waterways and artificial channels or the damming of major rivers, which increase saline intrusion, changing fish communities (Barletta et al., 2016, 2017b; Prestrelo and Monteiro-Neto, 2016). Impacts of dredging has been widely discussed worldwide (Wilber and Clarke, 2001; Nayar et al., 2003; Güt and Curran, 2017). The loss of nursery services and of the jobs associated with fishing activities are the main consequences of dredging or damming. Wasserman et al. (2016) asserted that the main concern for the interpretation of such impacts is the absence of studies using local hydrodynamics.
In SA, dredging activities were discussed in Sepetiba Bay (Araújo et al., 2016), Guanabara Bay (Silveira et al., 2017), Bahía Blanca Estuary (Zilio et al., 2013), and Paranaguá Estuarine Complex (Barletta et al., 2016). The damming of the São Francisco Estuary, for example, decreased the magnitude of its flow from 2,846 m3 s−1 to 800 m3 s−1, leading to increased saline intrusion (Barletta et al., 2017b). Water transposition from the river basin to the adjacent semi-arid region to serve rural populations, industrial, and tourism activities during severe droughts has been widely criticized for its potential impacts (Barletta et al., 2017b). In the Rhine-Meuse estuary, for example, it is suggested that the restoration of tidal and river dynamics in polders are the best options for the ecological rehabilitation of one of the most important wetlands in the Netherlands (Storm et al., 2005; Slater, 2016). However, possibilities for a natural recovery of the São Francisco Estuary are far from any perspective, since background and novel information on human-driven changes are absent. Further investigations are needed since the saline intrusion in the current stagnant estuary has increased, changing ecological functions for fishes. Fortunately, larger systems have shown the capacity to withstand human modifications. However, “the full recovery of coastal marine and estuarine ecosystems from over a century of degradation can take a minimum of 15–20 years for attainment of the original biotic composition and diversity may lag far beyond that period” (Borja et al., 2010).
Estuarine Conservation and Recommendations
The avoidance of governmental institutions in using the basic estuarine concepts has been debated for decades (Dauvin and Ruellet, 2009). Researches using ecocline concepts are increasing in quantity and quality (Blaber, 2000; Barletta et al., 2016; Reis et al., 2016). However, the time elapsed to compile scientific information and the establishment of plans is the main challenge. Despite the variety of coastal ecosystems in SA, the number of effective Marine Protected Areas is still insufficient, and conservation measures are not implemented outside these areas to guarantee the conservation of connected habitats.
The lack of basic sanitation is the worst concern in SA (Costa and Barletta, 2016). In riverside settlements, uncontrolled sewage discharges along the whole course of rivers only increases at estuaries. Several nano- and biotechnologies are alternatives to reduce contaminants in effluents and offer a potential treatment of surface water, groundwater and wastewater contaminated by toxic metal ions, organic and inorganic solutes, and microorganisms (Wang et al., 2010; Qu et al., 2013; Martínez-Huitle and Ferro, 2016).
Plans aiming at estuarine conservation must consider year-rounded cycles of retention and flush of environmental contaminants along ecoclines, as well as their interactions with fishes (Barletta et al., 2016). However, the majority of studies consider only limited spatial patterns based on the distance to point-sources of impacts. The lack of temporal assessments also leads to misinterpretation of whether estuaries are able to withstand human modifications and when (and if) estuaries will recover from unpredictable climatic events. Withal, environmental quality assessment have evolved when biological measures in fishes were integrated to ecological relevance, providing a new support for management, and monitoring schemes (Duarte et al., 2017).
An example of successful monitoring of is through the use of the Integrated Biomarker Response index (IBR), which can indicate different sources of anthropogenic contamination in aquatic environments using fish as bioindicator. Larger biomarker responses are often found in the most contaminated sites (Duarte et al., 2017). In the Estuarine-Lagoon Complex of Iguape-Cananéia (Southeast Brazil), the spatio-temporal changes in parameters such as oxidative stress, biotransformation, genotoxicity, and histopathological alterations in Atherinella brasiliensis correlated to sediment pollution with metals, PAHs and pharmaceuticals and personal hygiene products (PPCPs) in areas of greater human presence (Salgado et al., 2018). The IBR corroborated with these results, indicating that the Cananéia City has the worst environmental quality (Salgado et al., 2018). The metabolic enzyme activities, protein content and lipid peroxidation were analyzed in muscle and liver of Ramnogaster arcuata (Clupeidae) to assess the correlation with PAHs levels in tissues. IBR was significant to assess PAH toxicity and highlighted R. arcuata as a good bioindicator in the Bahía Blanca Estuary (Ronda et al., 2018). These multi-biomarker approaches help in the comprehension of ecosystem health aiming at a more comprehensive assessment of environmental quality (Duarte et al., 2017). Moreover, comparative measures of IBR before and after any type of managerial or conservative action can help to assure the success or failure of such actions. Therefore, further studies regarding fish indices should be performed as a tool to assess environmental quality in SA estuaries.
Another problem faced in SA is the overexploitation of estuarine fishery resoruces. Along the Amazon River basin, multispecies fishery follows a clear seasonal pattern of river hydrological cycles (Isaac et al., 2016). However, hydrological alterations due to the construction of dams, in addition to deforestation and climate change affect the seasonal and annual dynamics of these fisheries (Isaac et al., 2016; Pinaya et al., 2016). Overexploitation of fishery resources is known since the early 80's, when traditional communities shift from labor in multiple resources (agriculture, fishing, and small-scale stock raising) to concentrate in commercial fishing (McGrath et al., 1993). Although captures per unit effort remained stable over time, body lengths of most-caught species were below length at first maturation (Castello et al., 2011). Fishery was characterized as moderately exploited, with few profitable species being overexploited. For the region, management of fisheries can lead to increased yields through the enforcement of minimum size of capture and closed seasons of catches (Castello et al., 2011).
Landings at Guanabara Bay, accounted to ~19,000 tons (US$ 4.8 million) between 2001 and 2002 (Jablonski et al., 2006). Small pelagic fishes (Atlantic anchoveta and Brazilian sardinella) and demersal fishes (croakers, mullets and catfishes) comprised the main catches, with densities compatible with commercial fisheries (Jablonski et al., 2006). However, fishermen associations have complained that fish abundance and fishery income has decreased since the oil spill in January 2000 (Jablonski et al., 2006). Unless regional industries implement regulations to avoid environmental degradation and measures to treat effluents, pollution will continue to threaten the development of artisanal fishing (Bessa et al., 2004).
State of poverty and lack of power of the artisanal fishermen are pointed as the main factor making this group invisible in the formulation of public policies, which favors industrial fishing (Galli et al., 2007). In the Rio de LA Plata Estuary, for example, the overexploitation of M. furnieri was attributed to improvements of the industrial fleet (Horta and Defeo, 2012). In Bahia Blanca Estuary, Cynoscion guatucupa is the most important resource captured by the artisanal fishery fleet. However, overexploitation and collapses in stocks are also related to increasing industrial operations in open waters adjacent to the estuary (Lopez Cazorla et al., 2014). Thus, the recognition of the socio-economic importance of artisanal fishing through the participation in decisions to the sustainability of their activities, improvement of the fishermen income through fish processing and selling at the landing points are frames needed to change this situation (Jablonski et al., 2006; Galli et al., 2007).
The conservation of coastal ecosystems and traditional livelihoods are ruled by different policies to preserve natural resources through the co-management perspective (ICMbio, 2012; Barletta et al., 2017b). Limited mesh sizes, closed periods, establishment of fishing seasons; no-take-zones, quotas for common resources; efforts to reduce/eliminate wastewater disposal, control uptake of freshwater by damming and control salinity intrusion by dredging can be highlighted as priorities for SA estuaries.
Re-establish operation licenses and accurately analyse risk assessments of activities next to river basins and sourround estuaries are important steps to prevent habitat loss (Lotze et al., 2006; Huang et al., 2014). Mangrove deforestation, for example, is of major concern in most estuarine systems along the Ecuadorian coast, especially due to aquaculture, and construction of shrimp ponds (Shervette et al., 2007; Hamilton and Stankwitz, 2012). It is asserted that shrimp aquaculture is responsible for 80% of carbon lost due to mangrove deforestation (Hamilton and Lovette, 2015). From an estimated 362,000 ha of mangrove forest, almost 50% had been lost, being the Muisne region the most impacted area (Zhengyun et al., 2003). In the Chone Estuary, for example, the establishment of shrimp farms, covering 51,919,128 m2 was responsible to reduce mangrove area from 42,377,182 m2 in 1968 to 14,654,255 m2 in 2006 (Hamilton and Stankwitz, 2012). In the Grande Estuary, most estuarine area is dedicated to shrimp farms, accounting for a mangrove loss of 47%. On the other hand, in Rio Hondo and Cayapas-Mataje, only 16 and 10% of mangrove loss due to shrimp farming was recorded by 2008 (Hamilton and Stankwitz, 2012). The maintenance of shrimp ponds needs to be managed and integrated to mangrove functioning, as an example of using mangroves as nutrient filter of pond effluents prior to the return of water back to the estuary (Twilley et al., 1998). Moreover, successful actions aiming at the recovery of deforested and degraded mangrove has been presented for some systems in SA. Replanting of mangroves in the Acaraú River (Paula et al., 2016) and Paraiba do Sul River estuaries (Rezende et al., 2015); and salt marshes in the Patos Lagoon (Tagliani et al., 2007) are acknowledged as a good approach to recover degraded wetlands and restablish biodiversity (Rezende et al., 2015).
Biodiversity is also threatened due to invasive species, which in turn are introduced by water ballast and aquaculture escaping. Estuaries and coasts are susceptible to introductions of non-native species since they are centers for the vessel activities. The introduction of these species may cause niche overlaps, when the invasive species uses the same resources as native species (Neuhaus et al., 2016). Although this sharing of resources may not affect the survival of the species, introductions and invasions of piscivorous predators, for example, can impact not only the species richness of native fish community but may also cause changes over all trophic levels, altering the equilibrium of the system (Neuhaus et al., 2016). This might also lead to impacts on native fishery resources, diseases, and parasites occurrences (Tomás et al., 2012). In Brazil, several case studies relate the introduction of non-native carnivorous fish to a decrease in local diversity. Examples are the introduction of O. beta in the Santos Estuary (Tomás et al., 2012), A. pantaneiro in the Patos Lagoon Estuary (Neuhaus et al., 2016) and O. punctatus in Babitonga Bay (Costa et al., 2011). Thus, studies assessing the overlapping food niches and, consequently, the degree of competition in the presence of invasive species, are important for the conservation of native species.
The estuarine ecocline has a great influence in the connectivity among river basins and coastal waters, and estuaries are recognized as important ecosystems for providing biological and geochemical demands to both these systems (Able, 2005; Barletta et al., 2010; Watanabe K. et al., 2014). Conservation issues aiming to improve resilience must onset before the ecosystem modifications overpass their capacity to maintain its natural resources and services (Williams and Crutzen, 2013). This is the case of the environmental tragedy at Doce River basin (southeast Brazil), on November 2015, where the disruption of a containment dam of mining tailings (Fundão/Samarco) contaminated the river with 50 million.m−3 of mud flood (Barletta et al., 2017b). It caused an immeasurable impact along hundreds of km of river bed toward the coasts of more than one Brazilian state. Lives and livelihood losses, kill of tons of fishes (Barletta et al., 2017b), contamination of sediments, potable and surface waters with metals (Segura et al., 2016; Gomes et al., 2017) are among the many accounted impacts. This is just one case of environmental accident affecting a SA coastal system, where co-management, practices of emergency and risk assessment protocols have traditionally less and less attention. Therefore, potentially, every system can share the fate of Doce River.
All these issues are of major concern for the South Eastern Pacific estuaries, since the lack of estuarine research lead to a poor understand of what is the real status of the systems. This might be attributed to socio-economic problems. Most countries have a lower development indicator and the percentage of gross domestic product destined to research and development is lowest in countries such as Peru, Colombia, Ecuador and Chile, when compared to Brazil and Argentine, for example (Cociocca and Delgado, 2017). In addition, problems such as guerrillas, narco-traffickers, decaying economy due to populism, inequality, and educational options has been of major concern for these contries. Therefore, financial support is hardly ever destined to research and development (Cociocca and Delgado, 2017). Moreover, the commoditization of shrimp farmings in Ecuador has destroyed thousands of hectares of mangroves (Zhengyun et al., 2003), but concomitantly has generated $ 1 billion annually, leading to government negligencies (Hamilton, 2012). In Peru, a research Agenda of priorities for coastal systems has only recently gained attention due to the workshop “Advancing Green Growth in Peru” in 2016 (McKinley et al., 2018). Nevertheless, Brazil and Argentina has been also suffering due to economic instability, corruption, crime and narco-traffickers and the future of research is an imminent concern (Cociocca and Delgado, 2017).
According to all recorded damages, SA estuaries have almost never recovered to background situations, despite time and efforts. The constant habitat modifications that were relatively slow for nearly 400 years, have increased since after Second World War and industrialization (and urbanization) in the continent, worsening environmental quality and reducing fish yields for coastal zones. If anthropogenic interferences keep with their course and speed, regardless all reported environmental concerns, natural resources and services provided by estuaries will face the most severe degradation in the near future. Further research to support managerial planning and action should, therefore, not only be scientific-based on the continent-wide natural diversity, but also need to consider the social importance of estuaries for traditional fishers and other populations, whose dependence upon these systems goes beyond livelihoods.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge financial support from Conselho Nacional de Desenvolvimento Científico e Tecnológico through grant (CNPq-Proc.404931/2016-2); Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) through scholarship (FACEPE/BFP-0130-1.08/15). MB is CNPq Fellow.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00237/full#supplementary-material
References
Able, K. W. (2005). A re-examination of fish estuarine dependence: evidence for connectivity between estuarine and ocean habitats. Estuar. Coast. Shelf Sci. 64, 5–7. doi: 10.1016/j.ecss.2005.02.002
Abreu, I. M., Cordeiro, R. C., Soares-Gomes, A., Abessa, D. M. S., Maranho, L. A., and Santelli, R. E. (2016). Ecological risk evaluation of sediment metals in a tropical Euthrophic Bay, Guanabara Bay, Southeast Atlantic. Mar. Pollut. Bull. 109, 435–445. doi: 10.1016/j.marpolbul.2016.05.030
Acha, E. M., Acha, E. M., Mianzan, H. W., Mianzan, H. W., Iribarne, O., Iribarne, O., et al. (2003). The role of the Río de la Plata bottom salinity front in accumulating debris. Mar. Pollut. Bull. 46, 197–202. doi: 10.1016/S0025-326X(02)00356-9
Acha, E. M., Mianzan, H., Lasta, C. A., and Guerrero, R. A. (1999). Estuarine spawning of the whitemouth croaker Micropogonias furnieri (Pisces: Sciaenidae), in the Río de la Plata, Argentina. Mar. Freshw. Res. 50, 57. doi: 10.1071/MF98045
Acuña-Plavan, A., Passadore, C., and Gimenez, L. (2010). Fish Assemblage in a temperate estuary on the Uruguayan coast: seasonal variation and environmental influence. Braz. J. Oceanogr. 58, 299–314. doi: 10.1590/S1679-87592010000400005
Acuña-Plavan, A., Sellanes, J., Rodríguez, L., and Burone, L. (2007). Feeding ecology of Urophycis brasiliensis on the Uruguayan coast of the Río de la Plata estuary. J. Appl. Ichthyol. 23, 231–239. doi: 10.1111/j.1439-0426.2007.00855.x
Aguiar, V. M. C., and Braga, E. S. (2007). Seasonal and tidal variability of phosphorus along a salinity gradient in the heavily polluted estuarine system of Santos/São Vicente - São Paulo, Brazil. Mar. Pollut. Bull. 54, 464–471. doi: 10.1016/j.marpolbul.2006.11.001
Aguiar, V. M. C., Lima, M. N., Abuchacra, R. C., Abuchacra, P. F. F., Neto, J. A. B., Borges, H. V., et al. (2016). Ecological risks of trace metals in Guanabara Bay, Rio de Janeiro, Brazil: an index analysis approach. Ecotoxicol. Environ. Saf. 133, 306–315. doi: 10.1016/j.ecoenv.2016.07.012
Aguiar, V. M. d. C., Neto, J. A. B., and Rangel, C. M. (2011). Eutrophication and hypoxia in four streams discharging in Guanabara Bay, RJ, Brazil, a case study. Mar. Pollut. Bull. 62, 1915–1919. doi: 10.1016/j.marpolbul.2011.04.035
Almeida, M. P., Barthem, R. B., Viana, A. S., and Charvet-Almeida, P. (2009). Factors affecting the distribution and abundance of freshwater stingrays (Chondrichthyes: Potamotrygonidae) at Marajó Island, mouth of the Amazon River. Panam. J. Aquat. Sci. 4, 1–11
Alonso, D., Pineda, P., Olivero, J., González, H., and Campos, N. (2000). Mercury levels in muscle of two fish species and sediments from the Cartagena Bay and the Cienaga Grande de Santa Marta, Colombia. Environ. Pollut. 109, 157–163. doi: 10.1016/S0269-7491(99)00225-0
Angeli, J. L. F., Trevizani, T. H., Ribeiro, A., Machado, E. C., Figueira, R. C. L., Markert, B., et al. (2013). Arsenic and other trace elements in two catfish species from Paranaguá Estuarine Complex, Paraná, Brazil. Environ. Monit. Assess. 185, 8333–8342. doi: 10.1007/s10661-013-3176-5
Anjos, V. E., Machado, E. C., and Grassi, M. T. (2012). Biogeochemical behavior of arsenic species at Paranaguá Estuarine Complex, Southern Brazil. Aquat. Geochem. 18, 407–420. doi: 10.1007/s10498-012-9161-8
Araruna, J. T. A. Jr., Campos, T. M. P., and Pires, P. J. M. (2014). Sediment characteristics of an impacted coastal bay: Baía de Guanabara, Rio de Janeiro, Brazil. J. Coast. Res. 71, 41–47. doi: 10.2112/SI71-005.1
Araújo, F. G., Azevedo, M. C. C., and Guedes, A. P. P. (2016). Inter-decadal changes in fish communities of a tropical bay in southeastern Brazil. Reg. Stud. Mar. Sci. 3, 107–118. doi: 10.1016/j.rsma.2015.06.001
Araújo, F. G., Azevedo, M. C. C., Silva, M. A., Pessanha, A. L. M., Gomes, I. D., and Cruz-Filho, A. G. (2002). Environmental influences on the demersal fish assemblages in the Sepetiba Bay, Brazil. Estuaries 25, 441–450. doi: 10.1007/BF02695986
Araújo, F. G., Pinto, S. M., Neves, L. M., and Azevedo, M. C. C. (2017). Inter-annual changes in fish communities of a tropical bay in southeastern Brazil: what can be inferred from anthropogenic activities? Mar. Pollut. Bull. 114, 102–113. doi: 10.1016/j.marpolbul.2016.08.063
Arias, A. H., Pereyra, M. T., and Marcovecchio, J. E. (2011). Multi-year monitoring of estuarine sediments as ultimate sink for DDT, HCH, and other organochlorinated pesticides in Argentina. Environ. Monit. Assess. 172, 17–32. doi: 10.1007/s10661-010-1315-9
Arias, A. H., Spetter, C. V., Freije, R. H., and Marcovecchio, J. E. (2009). Polycyclic Aromatic Hydrocarbons in water, mussels (Brachidontes sp., Tagelus sp.) and fish (Odontesthes sp.) from Bahía Blanca Estuary, Argentina. Estuar. Coast. Shelf Sci. 85, 67–81. doi: 10.1016/j.ecss.2009.06.008
Arias, A. H., Vazquez-Botello, A., Tombesi, N., Ponce-Vélez, G., Freije, H., and Marcovecchio, J. (2010). Presence, distribution, and origins of Polycyclic Aromatic Hydrocarbons (PAHs) in sediments from Bahía Blanca estuary, Argentina. Environ. Monit. Assess. 160, 301–314. doi: 10.1007/s10661-008-0696-5
Artunduaga, E. (1978). Consideraciones sobre el núcleo de pescadores de Málaga en el Pacífico colombiano. Divulgación Pesquera Inst. Desarro. Recur. Nat. Renovables Bogotá 13, 1–14.
Assis, D. A. S., Dias-Filho, V. A., Magalhães, A. L. B., and Brito, M. F. G. (2017). Establishment of the non-native fish Metynnis lippincottianus (Cope 1870) (Characiformes: Serrasalmidae) in lower São Francisco River, northeastern Brazil. Stud. Neotrop. Fauna Environ. 52, 228–238. doi: 10.1080/01650521.2017.1348057
Attrill, M. J., and Rundle, S. D. (2002). Ecotone or ecocline: ecological boundaries in estuaries. Estuar. Coast. Shelf Sci. 55, 929–936. doi: 10.1006/ecss.2002.1036
Avigliano, E., Carvalho, B., Velasco, G., Tripodi, P., Vianna, M., and Volpedo, A. V. (2016). Nursery areas and connectivity of the adults anadromous catfish (Genidens barbus) revealed by otolith-core microchemistry in the south-western Atlantic Ocean. Mar. Freshw. Res. 68, 931–940. doi: 10.1071/MF16058
Avigliano, E., Carvalho, B., Velasco, G., Tripodi, P., and Volpedo, A. V. (2017a). Inter-annual variability in otolith chemistry of catfish Genidens barbus from South-western Atlantic estuaries. J. Mar. Biol. Assoc. U.K. 98, 855–865. doi: 10.1017/S0025315417000212
Avigliano, E., Leisen, M., Romero, R., Carvalho, B., Velasco, G., Vianna, M., et al. (2017b). Fluvio-marine travelers fromSA: cyclic amphidromy and freshwater residency, typical behaviors in Genidens barbus inferred by otolith chemistry. Fish. Res. 193, 184–194. doi: 10.1016/j.fishres.2017.04.011
Avigliano, E., Velasco, G., and Volpedo, A. V. (2015). Use of lapillus otolith microchemistry as an indicator of the habitat of Genidens barbus from different estuarine environments in the southwestern Atlantic Ocean. Environ. Biol. Fishes 98, 1623–1632. doi: 10.1007/s10641-015-0387-3
Azevedo, J. S., Braga, E. S., Favaro, D. T., Perretti, A. R., Rezende, C. E., and Souza, C. M. M. (2011). Total mercury in sediments and in Brazilian Ariidae catfish from two estuaries under different anthropogenic influence. Mar. Pollut. Bull. 62, 2724–2731. doi: 10.1016/j.marpolbul.2011.09.015
Azevedo, J. S., Serafim, A., Company, R., Braga, E. S., Fávaro, D. I., and Bebianno, M. J. (2009). Biomarkers of exposure to metal contamination and lipid peroxidation in the benthic fish Cathorops spixii from two estuaries in South America, Brazil. Ecotoxicology 18, 1001–1010. doi: 10.1007/s10646-009-0370-x
Baêta, A. P., Kehrig, H. A., Malm, O., and Moreira, I. (2006). Total mercury and methylmercury in fish from a tropical estuary. Environ. Toxicol. 1, 183–192. doi: 10.2495/ETOX060181
Baptista Neto, J. A., Peixoto, T. C. S., Smith, B. J., Mcalister, J. J., Patchineelam, S. M., Patchineelam, S. R., et al. (2013). Geochronology and heavy metal flux to Guanabara Bay, Rio de Janeiro state: a preliminary study. An. Acad. Bras. Cienc. 85, 1317–1327. doi: 10.1590/0001-3765201394612
Barbosa, N. D., Rocha, R. M., and Lucena Frédou, F. (2012). The reproductive biology of Plagioscion squamosissimus (Heckel, 1840) in the Pará River estuary (Amazon Estuary). J. Appl. Ichthyol. 28, 800–805. doi: 10.1111/j.1439-0426.2012.02040.x
Barletta, M., Amaral, C. S., Corrêa, M. F. M., Guebert, F., Dantas, D. V., Lorenzi, L., et al. (2008). Factors affecting seasonal variations in demersal fish assemblages at an ecocline in a tropical-subtropical estuary. J. Fish Biol. 73, 1314–1336. doi: 10.1111/j.1095-8649.2008.02005.x
Barletta, M., Barletta-Bergan, A., Saint-Paul, U., and Hubold, G. (2003). Seasonal changes in density, biomass, and diversity of estuarine fishes in tidal mangrove creeks of the lower Caeté Estuary (northern Brazilian coast, east Amazon). Mar. Ecol. Prog. Ser. 256, 217–228. doi: 10.3354/meps256217
Barletta, M., Barletta-Bergan, A., Saint-Paul, U., and Hubold, G. (2005). The role of salinity in structuring the fish assemblages in a tropical estuary. J. Fish Biol. 66, 45–72. doi: 10.1111/j.1095-8649.2004.00582.x
Barletta, M., Cysneiros, F. J. A., and Lima, A. R. A. (2016). Effects of dredging operations on the demersal fish fauna of a South American tropical–subtropical transition estuary. J. Fish Biol. 89, 890–920. doi: 10.1111/jfb.12999
Barletta, M., and Dantas, D. V. (2016). “Environmental gradientes,” in Encyclopedia of Estuaries, ed M. J. Kennish (New Jersey, NJ: Springer), 237–242.
Barletta, M., Jaureguizar, A. J., Baigun, C., Fontoura, N. F., Agostinho, A. A., Almeida-Val, V. M. F., et al. (2010). Fish and aquatic habitat conservation in South America: a continental overview with emphasis on neotropical systems. J. Fish Biol. 76, 2118–2176. doi: 10.1111/j.1095-8649.2010.02684.x
Barletta, M., Lima, A. R. A., and Costa, M. F. (2019). Distribution, sources and consequences of nutrients, persistent organic pollutants, metals and microplastics in South American estuaries. Sci. Total Environ. 651, 1199–1218. doi: 10.1016/j.scitotenv.2018.09.276
Barletta, M., Lima, A. R. A., Costa, M. F., and Dantas, D. V. (2017a). “Estuarine ecoclines and the associated fauna: ecological information as the basis for ecosystem conservation,” in Coastal Wetlands: Alteration and Remediation, eds. C. W. Finkl and C. Makowski (Cham: Springer International Publishing AG), 479–512. doi: 10.1007/978-3-319-56179-0_16
Barletta, M., Lima, A. R. A., Dantas, D. V., Oliveira, I. M., Neto, J. R., Fernandes, C. A. F., et al. (2017b). “How can accurate landing stats help in designing better fisheries and environmental management for Western Atlantic estuaries?,” in Coastal Wetlands: Alteration and Remediation, eds. C. W. Finkl and C. Makowski (Cham: Springer International Publishing AG), 631–703. doi: 10.1007/978-3-319-56179-0_20
Barletta, M., Lucena, L. R. R., Costa, M. F., Barbosa-Cintra, S. C. T., and Cysneiros, F. J. A. (2012). The interaction rainfall vs. weight as determinant of total mercury concentration in fish from a tropical estuary. Environ. Pollut. 167, 1–6. doi: 10.1016/j.envpol.2012.03.033
Barletta-Bergan, A., Barletta, M., and Saint-Paul, U. (2002a). Structure and seasonal dynamics of larval fish in the Caete River Estuary in North Brazil. Estuar. Coast. Shelf Sci. 54, 193–206. doi: 10.1006/ecss.2001.0842
Barletta-Bergan, A., Barletta, M., and Saint-Paul, U. (2002b). Community structure and temporal variability of ichthyoplankton in North Brazilian mangrove creeks. J. Fish Biol. 61, 33–51. doi: 10.1111/j.1095-8649.2002.tb01759.x
Barros, A. C., and Uhl, C. (1995). Logging along the Amazon River and estuary: patterns, problems and potential. For. Ecol. Manag. 77, 87–105. doi: 10.1016/0378-1127(95)03574-T
Barthem, R. B., de Brito Ribeiro, M. C. L., and Petrere, M. (1991). Life strategies of some long-distance migratory catfish in relation to hydroelectric dams in the Amazon Basin. Biol. Conserv. 55, 339–345. doi: 10.1016/0006-3207(91)90037-A
Bayen, S. (2012). Occurrence, bioavailability and toxic effects of trace metals and organic contaminants in mangrove ecosystems: a review. Envir Intern. 48, 84–101. doi: 10.1016/j.envint.2012.07.008
Beheregaray, L. B., and Levy, J. A. (2000). (Teleostei, Atherinopsidae): evidence for speciation in an estuary of Southern Brazil. Copeia 2000, 441–447. doi: 10.1643/0045-8511(2000)000[0441:PGOTSO]2.0.CO;2
Beltrame, M. O., De Marco, S. G., and Marcovecchio, J. E. (2009). Dissolved and particulate heavy metals distribution in coastal lagoons. a case study from Mar Chiquita Lagoon, Argentina. Estuar. Coast. Shelf Sci. 85, 45–56. doi: 10.1016/j.ecss.2009.04.027
Berasategui, A. D., Acha, E. M., and Fernández Araoz, N. C. (2004). Spatial patterns of ichthyoplankton assemblages in the Río de la Plata Estuary (Argentina-Uruguay). Estuar. Coast. Shelf Sci. 60, 599–610. doi: 10.1016/j.ecss.2004.02.015
Bessa, E. S., D'Avignon, A., Valle, R., and Teixeira, M. G. (2004). Integrating economic development and the environment: artisan fishing production in Guanabara Bay, Rio de Janeiro. Environ. Manag. 34, 332–340. doi: 10.1007/s00267-003-2530-8
Betancourt Portela, J. M., Sánchez Díazgranados, J. G., Mejía Ladino, L. M., and Cantera Kintz, J. R. (2011). Calidad de las aguas superficiales de bahía málaga, pacífico colombiano. Acta Biol. Colomb. 16, 175–192. doi: 10.15446/abc
Bisi, T. L., Lepoint, G., Azevedo, A. D. F., Dorneles, P. R., Flach, L., Das, K., et al. (2012). Trophic relationships and mercury biomagnification in Brazilian tropical coastal food webs. Ecol. Indic. 18, 291–302. doi: 10.1016/j.ecolind.2011.11.015
Blaber, S. J. M. (2000). Tropical Estuarine Fishes: Ecology, Exploitation and Conservation. Malden, MA: Blackwell Science. 372.
Blaber, S. J. M., and Barletta, M. (2016). A review of estuarine fish research in South America: what has been achieved and what is the future for sustainability and conservation? J. Fish Biol. 89, 537–568. doi: 10.1111/jfb.12875
Bonatti, M., Furlan, S., Manente, S., and Perin, G. (2004). Study of the toxicity of marine sediments of babitonga Bay – Brazil. J. Coast. Res. 21:39
Borba, T. A. C., and Rollnic, M. (2016). Runoff quantification on Amazonian Estuary based on hydrodynamic model. J. Coast. Res. 75, 43–47. doi: 10.2112/SI75-009.1
Borges, A. C., Dias, J. C., Machado, W., Patchineelam, S. R., and Sella, S. M. (2007). Distribuição espacial de ferro, cobre e chumbo em sedimentos de manguezal em um gradiente de degradação na Baía de Guanabara (Estado do Rio de Janeiro). Quim. Nova 30, 66–69. doi: 10.1590/S0100-40422007000100015
Borja, Á., Dauer, D. M., Elliott, M., and Simenstad, C. A. (2010). Medium-and long-term recovery of estuarine and coastal ecosystems: patterns, rates and restoration effectiveness. Estua. Coast. 33, 1249–1260. doi: 10.1007/s12237-010-9347-5
Botté, S. E., Freije, R. H., and Marcovecchio, J. E. (2007). Dissolved heavy metal (Cd, Pb, Cr, Ni) concentrations in surface water and porewater from Bahía Blanca estuary tidal flats. Bull. Environ. Contam. Toxicol. 79, 415–421. doi: 10.1007/s00128-007-9231-6
Braga, E. S., Bonetti, C. V. D. H., Burone, L., and Bonetti Filho, J. (2000). Eutrophication and bacterial pollution caused by industrial and domestic wastes at the Baixada Santista Estuarine System - Brazil. Mar. Pollut. Bull. 40, 165–173. doi: 10.1016/S0025-326X(99)00199-X
Brauko, K. M., Muniz, P., Martins, C. D. C., and Lana, P. C. (2016). Assessing the suitability of five benthic indices for environmental health assessment in a large subtropical South American estuary. Ecol. Indic. 64, 258–265. doi: 10.1016/j.ecolind.2016.01.008
Braverman, M. S., Acha, E. M., Gagliardini, D. A., and Rivarossa, M. (2009). Distribution of whitemouth croaker (Micropogonias furnieri, Desmarest 1823) larvae in the Río de la Plata estuarine front. Estuar. Coast. Shelf Sci. 82, 557–565. doi: 10.1016/j.ecss.2009.02.018
Brito, M. F. G., Bartolette, R., D'ávilla, T., Gomes, M. V. T., and Dias-filho, V. A. (2016). Reappearance of matrinxã Brycon orthotaenia (Characiformes: Bryconidae) in the lower São Francisco river, Brazil. AACL Bioflux 9, 949–953.
Bruno, D. O., Barbini, S. A., Díaz de Astarloa, J. M., and Martos, P. (2013). Fish abundance and distribution patterns related to environmental factors in a choked temperate coastal lagoon (Argentina). Brazil. J. Oceanogr. 61, 43–53. doi: 10.1590/S1679-87592013000100005
Bruno, D. O., Cousseau, M. B., Díaz de Astarloa, J. M., and Acha, E. M. (2015). Recruitment of juvenile fishes into a small temperate choked lagoon (Argentina) and the influence of environmental factors during the process. Sci. Mar. 79, 43–55. doi: 10.3989/sciamar.04085.02A
Burns, M. D. M., Garcia, A. M., and Vieira, J. P. (2010). Pisces, Perciformes, Gobiidae, Ctenogobius stigmaticus (Poey, 1860): new species record at patos lagoon estuary, state of rio grande do Sul, Brazil. Check List 6, 56–57. doi: 10.15560/6.1.056
Burns, M. D. M., Garcia, A. M., Vieira, J. P., Bemvenuti, M. A., Marques, D. M. L. M., and Condini, V. (2006). Evidence of habitat fragmentation affecting fish movement between the Patos and Mirim coastal lagoons in southern Brazil. Neotrop. Ichthyol. 4, 69–72. doi: 10.1590/S1679-62252006000100006
Buruaem, L. M., Araujo, G. S., Rosa, P. A., Nicodemo, S. C., Porto, V. F., Fonseca, J. R., et al. (2013). Assessment of sediment toxicity from the Areia Branca off-shore harbour and the Potengi river estuary (RN), northeastern Brazil. Panam. J. Aquat. Sci. 8, 312–326
Camilión, M. C., Manassero, M. J., Hurtado, M. A., and Ronco, A. E. (2003). Copper, lead and zinc distribution in soils and sediments of the south-western coast of the Río de La Plata estuary. J. Soils Sedim. 3, 213–220. doi: 10.1065/jss2003.04.073
Campos, N. H. (1992). Concentraciones de metales traza en Ariopsis bonillai (Pisces: Siluriformes) de Santa Marta, Caribe colombiano. Rev. Biol. Trop. 40, 179–183.
Cappelletti, N., Speranza, E., Tatone, L., Astoviza, M., Migoya, M. C., and Colombo, J. C. (2015). Bioaccumulation of dioxin-like PCBs and PBDEs by detritus-feeding fish in the Rio de la Plata estuary, Argentina. Environ. Sci. Pollut. Res. 22, 7093–7100. doi: 10.1007/s11356-014-3935-z
Carmo, W. P. D., Bornatowski, H., Oliveira, E. C., and Fávaro, L. L. (2015). Diet of the chola guitarfish, Rhinobatos percellens (Rhinobatidae), in the paranaguá estuarine complex. An. Acad. Bras. Cienc. 87, 721–731. doi: 10.1590/0001-3765201520140121
Carvalho, B. M., and Spach, H. L. (2015). Habitat use by Atherinella brasiliensis (Quoy and Gaimard, 1825) in intertidal zones of a subtropical estuary, Brazil. Acta Sci. Biol. Sci. 37, 177–184. doi: 10.4025/actascibiolsci.v37i2.25205
Carvalho, D. G., and Baptista Neto, J. A. (2016). Microplastic pollution of the beaches of Guanabara Bay, Southeast Brazil. Ocean Coast. Manag. 128, 10–17. doi: 10.1016/j.ocecoaman.2016.04.009
Carvalho, T. P., Lehmann, A. P, Pereira, E. H. L., and Reis, R. E. (2008). A new species of Hisonotus (Siluriformes: Loricariidae: Hypoptopomatinae) from the Laguna dos Patos Basin, Southern Brazil. Copeia 2008, 510–516. doi: 10.1643/CI-07-130
Castellanos-Galindo, G. A., Caicedo-Pantoja, J. A., Mejía-Ladino, L. M., and Rubio, E. (2006). Peces marinos y estuarinos de Bahía Málaga, Valle del Cauca, Pacífico colombiano Gustavo. Biota Colomb. 7, 263–282
Castellanos-Galindo, G. A., Cantera, J., Valencia, N., Giraldo, S., Peña, E., Kluger, L. C., et al. (2017). Modeling trophic flows in the wettest mangroves of the world: the case of Bahía Málaga in the Colombian Pacific coast. Hydrobiologia 803, 13–27. doi: 10.1007/s10750-017-3300-6
Castellanos-Galindo, G. A., Cantera, J. R., Espinosa, S., and Mejía-Ladino, L. M. (2011). Use of local ecological knowledge, scientist's observations and grey literature to assess marine species at risk in a tropical eastern Pacific estuary. Aquat. Conserv. Mar. Freshw. Ecosyst. 21, 37–48. doi: 10.1002/aqc.1163
Castellanos-Galindo, G. A., Giraldo, A., and Rubio, E. A. (2005). Community structure of an assemblage of tidepool fishes on a tropical eastern Pacific rocky shore, Colombia. J. Fish Biol. 67, 392–408. doi: 10.1111/j.0022-1112.2005.00735.x
Castellanos-Galindo, G. A., and Krumme, U. (2013). Tidal, diel and seasonal effects on intertidal mangrove fish in a high-rainfall area of the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser. 494, 249–265. doi: 10.3354/meps10512
Castello, L., McGrath, D. G., and Beck, P. S. A. (2011). Resource sustainability in small-scale fisheries in the Lower Amazon floodplains. Fish. Res. 110, 356–364. doi: 10.1016/j.fishres.2011.05.002
Castillo, L. (1986). “Lista anotada de peces,” en Impacto ecológico en Bahía Málaga a raíz del desarrollo de la Base Naval del Pacífico y la carretera de acceso, Tomo VI, eds J. Cantera, H. von Prahl (Cali: Cenipacífico), 397–487.
Castro, L. T., Santos-Martínez, A., and Acero, P. A. (1999). Reproducción de Bairdiella ronchus (Pisces: Sciaenidae) en la Ciénaga Grande de Santa Marta, Caribe Colombiano. Rev. Biol. Trop. 47, 553–559.
Castro, M. S., Bonecker, A. C. T., and Valentin, J. L. (2005). Seasonal variation in fish larvae at the entrance of Guanabara Bay, Brazil. Braz. Arch. Biol. Technol. 48, 121–128. doi: 10.1590/S1516-89132005000100016
Castro, R. O., Silva, M. L., Marques, M. R. C., and Araújo, F. V. (2016). Evaluation of microplastics in Jurujuba Cove, Niterói, RJ, Brazil, an area of mussels farming. Mar. Pollut. Bull. 110, 555–558. doi: 10.1016/j.marpolbul.2016.05.037
Cavole, L. M., and Haimovici, M. (2015). The use of otolith microstructure in resolving issues of ageing and growth of young Micropogonias furnieri from southern Brazil. Mar. Biol. Res. 11, 933–943. doi: 10.1080/17451000.2015.1031799
Cellone, F., Carol, E., and Tosi, L. (2016). Coastal erosion and loss of wetlands in the middle Río de la Plata estuary (Argentina). Appl. Geogr. 76, 37–48. doi: 10.1016/j.apgeog.2016.09.014
Cervigón, F. (1985). “La octiofauna de las aguas costeiras estuarinas del Delta del Rio Orinoco en la costa Atlántica Occidental, Caribe,” in Fish Community Ecology in Estuaries and Coastal Lagoons: Towards an Ecosystem Integration, ed A. Yáñez-Arancibia (Mexico City: UNAM Press), 57–78.
Chaves, P., and Bouchereau, J.-L. (1999). Biodiversité et dynamique des peuplements ichtyiques de la mangrove de Guaratuba, Brésil. Oceanol. Acta 22, 353–364. doi: 10.1016/S0399-1784(99)80057-7
Chaves, P., and Bouchereau, J.-L. (2000). Use of mangrove habitat for reproductive activity by the fish assemblage in the Guaratuba Bay, Brazil. Oceanol. Acta 23, 273–280. doi: 10.1016/S0399-1784(00)00130-4
Chaves, P. T., and Nogueira, A. B. (2013). Salinity influence on development and weight- length relationship of the fat snook. Bol. Inst. Pesca 39, 423–432.
Chaves, P. T., and Vendel, A. L. (2008). Análise comparativa da alimentação de peixes (Teleostei) entre ambientes de marisma e de manguezal num estuário do sul do Brasil (Baía de Guaratuba, Paraná). Rev. Bras. Zool. 25, 10–15. doi: 10.1590/S0101-81752008000100002
Chaves, P. T. C., and Corrêa, C. E. (2000). Temporary use of a coastal ecosystem by the fish, Pomadasys corvinaeformis (Perciformes: Haemulidae), at Guaratuba Bay, Brazil. Rev. Bras. Oceanogr. 48, 1–7. doi: 10.1590/S1413-77392000000100001
Chaves, P. T. C., and Corrêa, M. F. M. (1998). Composição ictiofaunística da área de manguezal da Baía de Guaratuba, Paraná, Brasil. Rev. Bras. Zool. 15, 195–202. doi: 10.1590/S0101-81751998000100017
Chaves, P. T. C., and Vendel, A. L. (1996). Aspectos da alimentação de Genidens genidens (Valenciennes) (Siluriformes, Ariidae) na Baía de Guaratuba, Paraná. Rev. Bras. Zool. 13, 669–675.
Chaves, P. T. C., and Vendel, A. L. (1997). Reprodução de Stellifer rastrifer (Jordan) (Teleostei, Sciaenidae) na Baía de Guaratuba, Paraná, Brasil. Rev. Bras. Zool. 14, 81–89. doi: 10.1590/S0101-81751997000100008
Cheffe, M. M., Lanés, L. E. K., Volcan, M. V., Burns, M. D., and de, M (2010). Pisces, Perciformes, Gobiidae, Evorthodus lyricus (Girard, 1858): new record from Patos-Mirim Lagoon System, state of Rio Grande do Sul, southern Brazil. Check List 6, 676–678. doi: 10.15560/6.4.676
Cheung, P. K., Cheung, L. T. O., and Fok, L. (2016). Seasonal variation in the abundance of marine plastic debris in the estuary of a subtropical macro-scale drainage basin in South China. Sci. Total Environ. 562, 658–665. doi: 10.1016/j.scitotenv.2016.04.048
Cociocca, D. R., and Delgado, G. (2017). The reality of scientific research in Latin America; an insider's perspective. Cell Stress Chaperones 22, 847–852. doi: 10.1007/s12192-017-0815-8
Coimbra, M. R. M., Lima, A. P. S., Oliveira, K. K. C., and Severi, W. (2017). Microsatellite assessment of the genetic diversity in indigenous populations of curimba (Prochilodus argenteus) in the São Francisco river (Brazil). Conserv. Genet. 18, 965–975. doi: 10.1007/s10592-017-0947-5
Colombo, J. C., Barreda, A., Bilos, C., Cappelletti, N., Demichelis, S., Lombardi, P., et al. (2005a). Oil spill in the Río de la Plata estuary, Argentina: 1. Biogeochemical assessment of waters, sediments, soils and biota. Environ. Pollut. 134, 277–289. doi: 10.1016/j.envpol.2004.02.032
Colombo, J. C., Barreda, A., Bilos, C., Cappelletti, N., Migoya, M. C., and Skorupka, C. (2005b). Oil spill in the Río de la Plata estuary, Argentina: 2-Hydrocarbon disappearance rates in sediments and soils. Environ. Pollut. 134, 267–276. doi: 10.1016/j.envpol.2004.07.028
Colombo, J. C., Bilos, C., Lenicov, M. R., Colautti, D., Landoni, P., Brochu, C., et al. (2000). Detritivorous fish contamination in the Río de la Plata estuary: a critical accumulation pathway in the cycle of anthropogenic compounds. Can. J. Fish. Aquat. Sci. 57, 1139–1150. doi: 10.1139/f00-031
Colombo, J. C., Cappelletti, N., Barreda, A., Migoya, M. C., and Skorupka, C. N. (2005c). Vertical fluxes and accumulation of PCBs in coastal sediments of the Río de la Plata estuary, Argentina. Chemosphere 61, 1345–1357. doi: 10.1016/j.chemosphere.2005.03.090
Colombo, J. C., Cappelletti, N., Migoya, M. C., and Speranza, E. (2007a). Bioaccumulation of anthropogenic contaminants by detritivorous fish in the Río de la Plata estuary: 1-Aliphatic hydrocarbons. Chemosphere 68, 2128–2135. doi: 10.1016/j.chemosphere.2007.02.001
Colombo, J. C., Cappelletti, N., Migoya, M. C., and Speranza, E. (2007b). Bioaccumulation of anthropogenic contaminants by detritivorous fish in the Río de la Plata estuary: 2-Polychlorinated biphenyls. Chemosphere 69, 1253–1260. doi: 10.1016/j.chemosphere.2007.05.073
Combi, T., Taniguchi, S., Figueira, R. C. L., de Mahiques, M. M., and Martins, C. C. (2013). Spatial distribution and historical input of Polychlorinated Biphenyls (PCBs) and Organochlorine Pesticides (OCPs) in sediments from a subtropical estuary (Guaratuba Bay, SW Atlantic). Mar. Pollut. Bull. 70, 247–252. doi: 10.1016/j.marpolbul.2013.02.022
Contente, R. F., Stefanoni, M. F., and Spach, H. L. (2011). Feeding ecology of the Brazilian silverside Atherinella brasiliensis (Atherinopsidae) in a sub-tropical estuarine ecosystem. J. Mar. Biol. Assoc. U.K. 91, 1197–1205. doi: 10.1017/S0025315410001116
Contente, R. F., Stefanoni, M. F., and Spach, H. L. (2012). Feeding ecology of the American freshwater goby Ctenogobius shufeldti (Gobiidae, Perciformes) in a sub-tropical estuary. J. Fish Biol. 80, 2357–2373. doi: 10.1111/j.1095-8649.2012.03300.x
Cordeiro, R. C., Machado, W., Santelli, R. E., Figueiredo, A. G., Seoane, J. C. S., Oliveira, E. P., et al. (2015). Geochemical fractionation of metals and semimetals in surface sediments from tropical impacted estuary (Guanabara Bay, Brazil). Environ. Earth Sci. 74, 1363–1378. doi: 10.1007/s12665-015-4127-y
Cordoba, D. F., and Giraldo, A. (2014). Ensamble íctico en corrientes de agua dulce de Isla Palma (Bahía Málaga, Pacífico colombiano). Boletín Científico Museos Hist. Nat. 18, 111–124.
Corrêa, B., and Vianna, M. (2016). Spatial and temporal distribution patterns of the silver mojarra Eucinostomus argenteus (Perciformes: Gerreidae) in a tropical semi-enclosed bay. J. Fish Biol. 89, 641–660. doi: 10.1111/jfb.12843
Corrales, D., Acuña, A., Salhi, M., Saona, G., and Brugnoli, E. (2016). Copper, zinc, mercury and arsenic content in Micropogonias furnieri and Mugil platanus of the Montevideo coastal zone, Río de la Plata. Braz. J. Oceanogr. 64, 57–66. doi: 10.1590/S1679-87592016105406401
Correa-Herrera, T., Barletta, M., Lima, A. R. A., Jiménez-Segura, L. F., and Arango-Sánchez, L. B. (2017). Spatial distribution and seasonality of ichthyoplankton and anthropogenic debris in a river delta in the Caribbean Sea. J. Fish Biol. 90, 1356–1387. doi: 10.1111/jfb.13243
Correa-Herrera, T., Jiménez-Segura, L. F., and Barletta, M. (2016). Fish species from a micro-tidal delta in the Caribbean Sea. J. Fish Biol. 89, 863–875. doi: 10.1111/jfb.12860
Costa, L. D. F., Oliveira Casartelli, M. R., and Wallner-Kersanach, M. (2013). Labile copper and zinc fractions under different salinity conditions in a shipyard area in the patos lagoon estuary, south of Brazil. Quim. Nova 36, 1089–1095. doi: 10.1590/S0100-40422013000800002
Costa, L. D. F., and Wallner-Kersanach, M. (2013). Assessment of the labile fractions of copper and zinc in marinas and port areas in Southern Brazil. Environ. Monit. Assess. 185, 6767–6781. doi: 10.1007/s10661-013-3063-0
Costa, M. D. P., and Muelbert, J. H. (2017). Long-term assessment of temporal variability in spatial patterns of early life stages of fishes to facilitate estuarine conservation. Mar. Biol. Res. 13, 74–87. doi: 10.1080/17451000.2016.1213397
Costa, M. D. P., Muelbert, J. H., Moraes, L. E., Vieira, J. P., and Castello, J. P. (2014). Estuarine early life stage habitat occupancy patterns of whitemouth croaker Micropogonias furnieri (Desmarest, 1830) from the Patos Lagoon, Brazil. Fish. Res. 160, 77–84. doi: 10.1016/j.fishres.2013.10.025
Costa, M. D. P., Muelbert, J. H., Vieira, J. P., and Castello, J. P. (2015). Dealing with temporal variation and different life stages of whitemouth croaker Micropogonias furnieri (Actinopterygii, Sciaenidae) in species distribution modeling to improve essential estuarine fish habitat identification. Hydrobiologia 762, 195–208. doi: 10.1007/s10750-015-2348-4
Costa, M. D. P., Possingham, H. P., and Muelbert, J. H. (2016). Incorporating early life stages of fishes into estuarine spatial conservation planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 26, 1013–1030. doi: 10.1002/aqc.2584
Costa, M. D. P., Schwingel, P. R., Souza-Conceição, J. M., and Spach, H. L. (2012). Distribuição espaço-temporal de larvas de Sciaenidae em um estuário subtropical (Santa Catarina, Brasil). Braz. J. Aquat. Sci. Technol. 16, 51–59. doi: 10.14210/bjast.v16n2.p51-59
Costa, M. D. P., and Souza-Conceição, J. M. (2009). Composição e abundância de ovos e larvas de peixes na baia da Babitonga, Santa Catarina, Brasil. Panam. J. Aquat. Sci. 4, 372–382.
Costa, M. D. P., Souza-Conceição, J. M., Schwingel, P. R., and Spach, H. L. (2011). Assessment of larval distribution of invasive Omobranchus punctatus (Valenciennes, 1836) (Pisces: Blenniidae) in a subtropical estuary (Southern Brazil). Aquat. Invasions 6, 33–38. doi: 10.3391/ai.2011.6.S1.008
Costa, M. F., Barbosa, S. C. T., Barletta, M., Dantas, D. V., Kehrig, H. A., Seixas, T. G., et al. (2009). Seasonal differences in mercury accumulation in Trichiurus lepturus (Cutlassfish) in relation to length and weight in a Northeast Brazilian estuary. Environ. Sci. Pollut. Res. 16, 423–430. doi: 10.1007/s11356-009-0120-x
Costa, M. F., and Barletta, M. (2015). Microplastics in coastal and marine environments of the western tropical and sub-tropical Atlantic Ocean. Environ. Sci. Processes Impacts 17, 1868–1879. doi: 10.1039/c5em00158g
Costa, M. F., and Barletta, M. (2016). Special challenges in the conservation of fishes and aquatic environments of South America. J. Fish Biol. 89, 4–11. doi: 10.1111/jfb.12970
Costa, M. F., Landing, W. M., Kehrig, H. A., Barletta, M., Holmes, C. D., Barrocas, P. R. G., et al. (2012). Mercury in tropical and subtropical coastal environments. Environ. Res.119, 88–100. doi: 10.1016/j.envres.2012.07.008
Costa, P. V., Silva, U. A. T., Ventura, R., Ostrensky, A., and Angelo, L. (2009). Fish predation on brachyuran larvae and juveniles in the Pinheiros river, Guaratuba Bay, Paraná, Brazil. Zoologia 26, 231–240. doi: 10.1590/S1984-46702009000200005
Cotovicz Junior, L. C., Machado, E. D. C., Brandini, N., Zem, R. C., and Knoppers, B. A. (2014). Distributions of total, inorganic and organic phosphorus in surface and recent sediments of the sub-tropical and semi-pristine Guaratuba Bay estuary, SE Brazil. Environ. Earth Sci. 72, 373–386. doi: 10.1007/s12665-013-2958-y
Cunha, C. L. N., Scudelari, A. C., Gonçalves, J. E., and Mercuri, E. G. F. (2011). Assessment of sanitary sewage pollution in the Paranaguá Estuarine Complex of Paranaguá, Brazil using environmental modeling. J. Coast. Res. 64, 912–916.
Cunha, R. M. P., and Calliari, L. J. (2009). Natural and antropic geomorphological changes in the inlet of Patos Lagoon before and after its fixation. J. Coast. Res. 2009, 708–712.
Da Silva Cortinhas, M. C., Kersanach, R., Proietti, M., Dumont, L. F. C., D'Incao, F., Lacerda, A. L. F., et al. (2016). Genetic structuring among silverside fish (Atherinella brasiliensis) populations from different Brazilian regions. Estuar. Coast. Shelf Sci. 178, 148–157. doi: 10.1016/j.ecss.2016.06.007
D'Anatro, A. (2017). Correlation between environmental factors, a life history trait, phenotypic cohesion, and gene flow levels in natural populations of Micropogonias furnieri: is salinity the main factor driving divergence? Estuar. Coast. 40, 1717–1731. doi: 10.1007/s12237-017-0234-1
D'Anatro, A., Pereira, A. N., and Lessa, E. P. (2011). Genetic structure of the white croaker, Micropogonias furnieri Desmarest 1823 (Perciformes: Sciaenidae) along Uruguayan coasts: contrasting marine, estuarine, and lacustrine populations. Environ. Biol. Fishes 91, 407–420. doi: 10.1007/s10641-011-9799-x
Dantas, D. V., Barletta, M., and Costa, M. F. (2015). Feeding ecology and seasonal diet overlap between Stellifer brasiliensis and Stellifer stellifer in a tropical estuarine ecocline. J. Fish Biol. 86, 707–733. doi: 10.1111/jfb.12592
Dantas, D. V., Barletta, M., Costa, M. F., Barbosa-Cintra, S. C. T., Possatto, F. E., Ramos, J. A. A., et al. (2010). Movement patterns of catfishes (Ariidae) in a tropical semi-arid estuary. J. Fish Biol. 76, 2540–2557. doi: 10.1111/j.1095-8649.2010.02646.x
Dantas, D. V., Barletta, M., and Ferreira, M. (2013). Seasonal diet shifts and overlap between two sympatric catfishes in an estuarine nursery. Estuar. Coast. 36, 237–256. doi: 10.1007/s12237-012-9563-2
Dantas, D. V., Barletta, M., Lima, A. R. A., de Assis Almeida Ramos, J., da Costa, M. F., and Saint-Paul, U. (2012). Nursery habitat shifts in an estuarine ecosystem: patterns of use by sympatric catfish species. Estuar. Coast. 35, 587–602. doi: 10.1007/s12237-011-9452-0
Dauner, A. L. L., and Martins, C. C. (2015). Spatial and temporal distribution of aliphatic hydrocarbons and linear alkylbenzenes in the particulate phase from a subtropical estuary (Guaratuba Bay, SW Atlantic) under seasonal population fluctuation. Sci. Total Environ. 536, 750–760. doi: 10.1016/j.scitotenv.2015.07.091
Dauvin, J.-C., and Ruellet, T. (2009). The estuarine quality paradox: is it possible to define an ecological quality status for specific modified and naturally stressed estuarine ecosystems? Mar. Pollut. Bull. 58, 38–47. doi: 10.1016/j.marpolbul.2008.11.008
Day, J. W., Yáñez-Arancibia, A., Kemp, W. M., and Crump, B. C. (eds.). (2012). Introduction to Estuarine Ecology, in Estuarine Ecology, 2nd Edn. Hoboken, NJ: John Wiley and Sons, Inc. doi: 10.1002/9781118412787.ch1
Delucchi, F., Tombesi, N. B., Freije, R. H., and Marcovecchio, J. E. (2007). Butyltin compounds in sediments of the Bahía Blanca estuary, Argentina. Environ. Monit. Assess. 132, 445–451. doi: 10.1007/s10661-006-9547-4
Díaz de Astarloa, J. M., Figueroa, D. E., Cousseau, M. B., and Barragán, M. (2000). Occurrence of Trachinotus carolinus (Carangidae) in laguna costera Mar Chiquita, with comments on other occasionally recorded fishes in Argentinean waters. Bull. Mar. Sci. 66, 399–403.
Díaz-Andrade, M. C., Galíndez, E., and Estecondo, S. (2009). The ovary of the bignose fanskate Sympterygia acuta Garman, 1877 (Chondrichthyes, Rajidae) in the Bahía Blanca estuary, Argentina: morphology and reproductive features. Braz. J. Biol. 69, 405–413. doi: 10.1590/S1519-69842009000200025
Duarte, I. A., Reis-Santos, P., França, S., Cabral, H., and Fonseca, V. F. (2017). Biomarker responses to environmental contamination in estuaries: a comparative multi-taxa approach. Aquat. Toxicol. 189, 31–41. doi: 10.1016/j.aquatox.2017.05.010
Duponchelle, F., Pouilly, M., Pécheyran, C., Hauser, M., Renno, J. F., Panfili, J., et al. (2016). Trans-Amazonian natal homing in giant catfish. J. Appl. Ecol. 53, 1511–1520. doi: 10.1111/1365-2664.12665
Duque, G., and Acero, P. A. (2003). Food habits of Anchovia clupeoides (Pisces: Engraulidae) in the Ciénaga Grande de Santa Marta, Colombian Caribbean. Gulf Mex. Sci. 21, 1–9. doi: 10.18785/goms.2101.01
Duque-Nivia, G., Acero, P. A., Santos-Martinez, A., and Rubio, E. R. (1996). Food habits of the species of the genus Oligoplites (Carangidae) from the Cienaga Grande de Santa Marta-Colombian Caribbean. Cybium 20, 251–260.
Ekau, W., Westhaus-ekau, P., Macêdo, S. J., and Dorrien, C. V. (2001). The larval fish fauna of the “Canal de Santa Cruz” esturay in Northeast Brazil. Trop. Oceanogr. Online 29, 117–128. doi: 10.5914/tropocean.v29i2.3028
Elliot, M., and Whitefield, A. K. (2011). Challenging paradigms in estuarine ecology and management. Estuar. Coast. Shelf Sci. 94, 306–314. doi: 10.1016/j.ecss.2011.06.016
Elliott, M., and Quintino, V. (2007). The estuarine quality paradox, environmental homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 54, 640–645. doi: 10.1016/j.marpolbul.2007.02.003
Farias, C. O., Hamacher, C., Wagener, A. L. R., and Scofield, A. L. (2008). Origin and degradation of hydrocarbons in mangrove sediments (Rio de Janeiro, Brazil) contaminated by an oil spill. Org. Geochem. 39, 289–307. doi: 10.1016/j.orggeochem.2007.12.008
Félix-Hackradt, F. C., and Hackradt, C. W. (2008). Populational study and monitoring of the goliath grouper, Epinephelus itajara (Lichtenstein, 1822), in the coast of Paraná, Brazil. Nat. Conservação 6, 141−156.
Fernandez, W. S., Dias, J. F., Boufleur, L. A., Amaral, L., Yoneama, M. L., and Dias, J. F. (2014). Bioacumulation of trace elements in hepatic and renal tissues of the white mullet Mugil curema Valenciennes, 1836 (Actinopterygii, Mugilidae) in two coastal systems in southeastern Brazil. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 318, 94–98. doi: 10.1016/j.nimb.2013.05.103
Ferreira, G. V. B., Barletta, M., and Lima, A. R. A. (2019). Use of estuarine resources by top predator fishes. How do ecological patterns affect rates of contamination by microplastics? Sci. Total Environ. 655, 292–304. doi: 10.1016/j.scitotenv.2018.11.229
Ferreira, G. V. B., Barletta, M., Lima, A. R. A., Dantas, D. V., Justino, A. K. S., and Costa, M. F. (2016). Plastic debris contamination in the life cycle of Acoupa weakfish (Cynoscion acoupa) in a tropical estuary. ICES J. Mar. Sci. J. Cons. 73, 2695–2707. doi: 10.1093/icesjms/fsw108
Fierro, P., Bertran, C., Martinez, D., Valdovinos, C., and Vargas-Chacoff, L. (2014). Ontogenetic and temporal changes in the diet of the Chilean silverside Odontesthes regia (Atherinidae) in Southern Chile. Cah. Biol. Mar. 55, 323–332.
Figueiredo, G. M., and Vieira, J. P. (2005). Diel feeding, daily food consumption and the predatory impact of whitemouth croaker (Micropogonias furnieri) in an estuarine environment. Mar. Ecol. 26, 130–139. doi: 10.1111/j.1439-0485.2005.00048.x
Fonseca, E. F., Baptista Neto, J. A., and Silva, C. G. (2013). Heavy metal accumulation in mangrove sediments surrounding a large waste reservoir of a local metallurgical plant, Sepetiba Bay, SE, Brazil. Environ. Earth Sci. 70, 643–650. doi: 10.1007/s12665-012-2148-3
Franco, A. C. S., Brotto, D. S., Zee, D. M. W., and Neves dos Santos, L. (2014). Reproductive biology of Cetengraulis edentulus (Cuvier, 1829), The major fishery resource in Guanabara Bay, Brazil. Neotrop. Ichthyol. 12, 819–826. doi: 10.1590/1982-0224-20130205
Franco, A. C. S., Ramos Chaves, M. C. N., Castel-Branco, M. P. B., and Neves dos Santos, L. (2016). Responses of fish assemblages of sandy beaches to different anthropogenic and hydrodynamic influences. J. Fish Biol. 89, 921–938. doi: 10.1111/jfb.12889
Frederico, R. G., Farias, I. P., Araújo, M. L. G., Charvet-Almeida, P., and Alves-Gomes, J. A. (2012). Phylogeography and conservation genetics of the Amazonian freshwater stingray Paratrygon aiereba Müller and Henle, 1841 (Chondrichthyes: Potamotrygonidae). Neotrop. Ichthyol. 10, 71–80. doi: 10.1590/S1679-62252012000100007
Froehner, S., MacEno, M., and MacHado, K. S. (2011). Predicting bioaccumulation of PAHs in the trophic chain in the estuary region of Paranagua, Brazil. Environ. Monit. Assess. 174, 135–145. doi: 10.1007/s10661-010-1444-1
Gabardo, I. T., Meniconi, M. F. G., Falcão, L. V., Vital, N. A. A., and Pereira, R. C. L. (2000). Hydrocarbon and ecotoxicity in seawater and sediment samples of Guanabara Bay after the oil spill in January 2000. Int. Oil Spill Conf. 2001, 941–950. doi: 10.7901/2169-3358-2001-2-941
Galli, E., Peipke, E., and Wennersten, R. (2007). When the traditional meets the modern: the sustainability of the artisanal fishing in Guanabara Bay, Brazil. Int. J. Environ. Sustain. Dev. 6, 373–388. doi: 10.1504/IJESD.2007.016241
Galloway, T. S., Cole, M., and Lewis, C. (2017). Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol Evol. 1:0016. doi: 10.1038/s41559-017-0116
Garbin, T., Castello, J. P., and Kinas, P. G. (2014). Age, growth, and mortality of the mullet Mugil liza in Brazil's southern and southeastern coastal regions. Fish. Res. 149, 61–68. doi: 10.1016/j.fishres.2013.09.008
Garcia, A. M., Garcia, A. M., Vieira, J. P., Vieira, J. P., Winemiller, K. O., Winemiller, K. O., et al. (2004). Reproductive cycle and spatiotemporal variation in abundance of the one-sided livebearer. Sites J. 20Th Century Contemp. French Stud. 515, 39–48. doi: 10.1023/B:HYDR.0000027316.59258.a0
Garcia, A. M., Raseira, M. B., Vieira, J. P., Winemiller, K. O., and Grimm, A. M. (2003). Spatiotemporal variation in shallow-water freshwater fish distribution and abundance in a large subtropical coastal lagoon. Environ. Biol. Fishes 68, 215–228. doi: 10.1023/A:1027366101945
Garcia, A. M., Vieira, J. P., and Winemiller, K. O. (2001). Dynamics of the shallow-water fish assemblage of the Patos Lagoon estuary (Brazil) during cold and warm ENSO episodes. J. Fish Biol. 59, 1218–1238. doi: 10.1006/jfbi.2001.1734
Garcia, A. M., Vieira, J. P., Winemiller, K. O., Moraes, L. E., and Paes, E. T. (2012). Factoring scales of spatial and temporal variation in fish abundance in a subtropical estuary. Mar. Ecol. Prog. Ser. 461, 121–135. doi: 10.3354/meps09798
Garcia, M. R., Mirlean, N., Baisch, P. R., and Caramão, E. B. (2010). Assessment of polycyclic aromatic hydrocarbon influx and sediment contamination in an urbanized estuary. Environ. Monit. Assess. 168, 269–276. doi: 10.1007/s10661-009-1110-7
Genz, F., Lessa, G. C., and Cirano, M. (2008). Vazão mínima para estuários: um estuado de caso no Rio Paraguaçu/BA. Rev. Bras. Recur. Hídricos 13, 73–82. doi: 10.21168/rbrh.v13n3.p73-82
Genz, F., and Luz, L. D. (2012). Distinguishing the effects of climate on discharge in a tropical river highly impacted by large dams. Hydrol. Sci. J. 57, 1020–1034. doi: 10.1080/02626667.2012.690880
Giarrizzo, T., and Krumme, U. (2007). Spatial differences and seasonal cyclicity in the intertidal fish fauna from four mangrove creeks in a salinity zone of the Curuçá Estuary, North Brazil. Bol. Mar. Sci. 80, 739–754.
Giarrizzo, T., Krumme, U., and Wosniok, W. (2010). Size-structured migration and feeding patterns in the banded puffer fish Colomesus psittacus (Tetraodontidae) from north brazilian mangrove creeks. Mar. Ecol. Prog. Ser. 419, 157–170. doi: 10.3354/meps08852
Giarrizzo, T., and Saint-Paul, U. (2008). Ontogenetic and seasonal shifts in the diet of the pemecou sea catfish Sciades herzbergii (Siluriformes: Ariidae), from a macrotidal mangrove creek in the Curuçá estuary, Northern Brazil. Rev. Biol. Trop. 56, 861–873. doi: 10.15517/rbt.v56i2.5629
Giarrizzo, T., Schwamborn, R., and Saint-Paul, U. (2011). Utilization of carbon sources in a northern Brazilian mangrove ecosystem. Estuar. Coast. Shelf. Sci. 95, 447–457. doi: 10.1016/j.ecss.2011.10.018
Giarrizzo, T., Silva de Jesus, A. J., Lameira, E. C., Araújo de Almeida, J. B., Isaac, V., and Saint-Paul, U. (2006). Weight-length relationships for intertidal fish fauna in a mangrove estuary in Northern Brazil. J. Appl. Ichthyol. 22, 325–327. doi: 10.1111/j.1439-0426.2006.00671.x
Goch, Y. G. F., Krumme, U., Saint-Paul, U., and Zuanon, J. A. S. (2005). Seasonal and diurnal changes in the fish fauna composition of a mangrove lake in the Caeté estuary, north Brazil. Amazoniana 18, 299–315.
Gomes, L. E. O., Correa, L. B., Sá, F., Neto, R. R., and Bernardino, A. F. (2017). The impacts of the Samarco mine tailing spill on the Rio Doce estuary, Eastern Brazil. Mar. Pollut. Bull. 120, 28–36. doi: 10.1016/j.marpolbul.2017.04.056
Gonçalves da Silva, S. F., Brüning, I. M. R. A., Montone, R. C., Taniguchi, S., Cascaes, M. J., Dias, P. S., et al. (2013). Polybrominated Diphenyl Ethers (PBDES) and Polychlorinated Biphenyls (PCBS) in mussels and two fish species from the estuary of the Guanabara Bay, Southeastern Brazil. Bull. Environ. Contam. Toxicol. 91, 261–266. doi: 10.1007/s00128-013-1026-3
González Castro, M., Díaz de Astarloa, J. M., and Cousseau, M. B. (2006). First record of a tropical affinity mullet, Mugil curema (Mugilidae), in a temperate southwestern Atlantic coastal lagoon. Cybium 30, 90–91.
González Castro, M., Díaz de Astarloa, J. M., Cousseau, M. B., Figueroa, D. E., Delpiani, S. M., Bruno, D. O., et al. (2009). Fish composition in a south-western Atlantic temperate coastal lagoon: spatial–temporal variation and relationships with environmental variables. J. Mar. Biol. Assoc. U.K. 89:593. doi: 10.1017/S0025315409003002
González Castro, M., Macchi, G. J., and Cousseau, M. B. (2011). Studies on reproduction of the mullet Mugil platanus Güünther, 1880 (Actinopterygii, Mugilidae) from the Mar Chiquita coastal lagoon, Argentina: Similarities and differences with related species. Ital. J. Zool. 78, 343–353. doi: 10.1080/11250003.2010.549154
Granados-Dieseldorff, P., and Baltz, D. M. (2008). Habitat use by nekton along a stream-order gradient in a Louisiana estuary. Estuar. Coasts 31, 572–583. doi: 10.1007/s12237-008-9051-x
Grecco, L. E., Gómez, E. A., Botté, S. E., Marcos, Á. O., Marcovecchio, J. E., and Cuadrado, D. G. (2011). Natural and anthropogenic heavy metals in estuarine cohesive sediments: geochemistry and bioavailability. Ocean Dyn. 61, 285–293. doi: 10.1007/s10236-010-0354-7
Gurdek, R., and Acuña-Plavan, A. (2014). Weight-length relationships of 12 fish species from the Pando tidal creek estuary (subsystem of the Río de la Plata, Uruguay). J. Appl. Ichthyol. 30, 426–427. doi: 10.1111/jai.12241
Gurdek, R., and Acuña-Plavan, A. (2016). Intra-annual length-weight relationships for juveniles of Micropogonias furnieri (Desmarest, 1823) in a sub-estuarine system from Uruguay. Panam. J. Aquat. Sci. 11, 165–169.
Güt, J. A., and Curran, M. C. (2017). Assessment of fish assemblages before dredging of the shipping channel near the mouth of the Savannah River in coastal Georgia. Estuar. Coasts 40, 251–267. doi: 10.1007/s12237-016-0139-4
Gutiérrez, O., Panario, D., Nagy, G. J., Bidegain, M., and Montes, C. (2016). Climate teleconnections and indicators of coastal systems response. Ocean Coast. Manag. 122, 64–76. doi: 10.1016/j.ocecoaman.2016.01.009
Hamilton, S. E. (2012). “The commodification of ecuador's estuary environments from 1970 – 2006,” in Congress of the Latin American Studies Association, San Francisco, May 23-26, 2012.21 pp. doi: 10.13140/RG.2.1.2918.8324
Hamilton, S. E., and Lovette, J. (2015). Ecuador's mangrove forest carbon stocks: a spatiotemporal analysis of living carbon holdings and their depletion since the advent of commercial aquaculture. PLoS ONE 10:e0118880. doi: 10.1371/journal.pone.0118880
Hamilton, S. E., and Stankwitz, C. (2012). Examining the relationship between international aid and mangrove deforestation in coastal Ecuador from 1970 to 2006. J. Land. Use. Sci. 7, 177–202. doi: 10.1080/1747423X.2010.550694
Harris, S. A., Cyrus, D. P., and Beckley, L. E. (1999). The larval fish assemblage in nearshore coastal waters off the St Lucia Estuary, South Africa. Estuar. Coast. Shelf Sci. 49, 789–811. doi: 10.1006/ecss.1999.0554
Hatje, V., and Barros, F. (2012). Overview of the 20th century impact of trace metal contamination in the estuaries of Todos os Santos Bay: past, present, and future scenarios. Mar. Pollut. Bull. 64, 2603–2614. doi: 10.1016/j.marpolbul.2012.07.009
Hegg, J. C., Giarrizzo, T., and Kennedy, B. P. (2015). Diverse early life-history strategies in migratory Amazonian catfish: implications for conservation and management. PLoS ONE 10, 1–19. doi: 10.1371/journal.pone.0129697
Horta, S., and Defeo, O. (2012). The spatial dynamics of the whitemouth croaker artisanal fishery in Uruguay and interdependencies with the industrial fleet. Fish. Res. 125–126, 121–128. doi: 10.1016/j.fishres.2012.02.007
Huang, J., Huang, Y., and Zhang, Z. (2014). Coupled effects of natural and anthropogenic controls on seasonal and spatial variations of river water quality during baseflow in a coastal watershed of southeast china. PLoS ONE 9:e91528. doi: 10.1371/journal.pone.0091528
Hugo, C. C. (1973). Migration of Galaxias maculatus (Jenyns) (Galaxiidae, pisces) in valdivia estuary, chile. Hydrobiologia 43, 301–312. doi: 10.1007/BF00015353
ICMbio (2012). Chico Mendes Institute for Biodiversity Conservation. Managerial Plan for the Marine Extractive Reserve of Caeté-Taperaçu (PA). Available online at: http://www.icmbio.gov.br/portal/unidadesdeconservacao/biomas-brasileiros/marinho/unidades-de-conservacao-marinho/2107-resex-marinha-de-caete-taperacu (accessed July 4, 2017).
Isaac, V. J., Castello, L., Santos, P. R. B., and Ruffino, M. L. (2016). Seasonal and interannual dynamics of river-floodplain multispecies fisheries in relation to flood pulses in the Lower Amazon. Fish. Res. 183, 352–359. doi: 10.1016/j.fishres.2016.06.017
Jablonski, S., Azevedo, A. F., and Moreira, L. H. A. (2006). Fisheries and conflicts in Guanabara Bay, Rio de Janeiro, Brazil. Brazil. Arch. Biol. Technol. 49, 79–91. doi: 10.1590/S1516-89132006000100010
Janeiro, J., Fernandes, E., Martins, F., and Fernandes, R. (2008). Wind and freshwater influence over hydrocarbon dispersal on Patos Lagoon, Brazil. Mar. Pollut. Bull. 56, 650–665. doi: 10.1016/j.marpolbul.2008.01.011
Jaureguizar, A. J., Bava, J., Carozza, C. R., and Lasta, C. A. (2003a). Distribution of whitemouth croaker Micropogonias furnieri in relation to environmental factors at the Río de la Plata estuary, Soh America. Mar. Ecol. Prog. Ser. 255, 271–282. doi: 10.3354/meps255271
Jaureguizar, A. J., Menni, R., Bremec, C., Mianzan, H., and Lasta, C. (2003b). Fish assemblage and environmental patterns in the Río de la Plata estuary. Estuar. Coast. Shelf. Sci. 56, 921–933. doi: 10.1016/S0272-7714(02)00288-3
Jaureguizar, A. J., Menni, R., Guerrero, R., and Lasta, C. (2004). Environmental factors structuring fish communities of the Río de la Plata estuary. Fish. Res. 66, 195–211. doi: 10.1016/S0165-7836(03)00200-5
Jaureguizar, A. J., Militelli, M. I., and Guerrero, R. (2008). Distribution of Micropogonias furnieri at different maturity stages along an estuarine gradient and in relation to environmental factors. J. Mar. Biol. Assoc. U.K. 88, 175–181. doi: 10.1017/S0025315408000167
Jaureguizar, A. J., and Raúl, G. (2009). Striped weakfish (Cynoscion guatucupa) population structure in waters adjacent to Rio de la Plata, environmental influence on its inter-annual variability. Estuar. Coast. Shelf. Sci. 85, 89–96. doi: 10.1016/j.ecss.2009.04.013
Jaureguizar, A. J., Ruarte, C., and Guerrero, R. A. (2006). Distribution of age-classes of striped weakfish (Cynoscion guatucupa) along an estuarine-marine gradient: correlations with the environmental parameters. Estuar. Coast. Shelf. Sci. 67, 82–92. doi: 10.1016/j.ecss.2005.10.014
Jaureguizar, A. J., Solari, A., Cortés, F., Milessi, A. C., Militelli, M. I., Camiolo, M. D., et al. (2016). Fish diversity in the Río de la Plata and adjacent waters: an overview of environmental influences on its spatial and temporal structure. J. Fish Biol. 89, 569–600. doi: 10.1111/jfb.12975
Kehrig, H. A., Costa, M., Moreira, I., and Malm, O. (2002). Total and methylmercury in a Brazilian estuary, Rio de Janeiro. Mar. Pollut. Bull. 44, 1018–1023. doi: 10.1016/S0025-326X(02)00140-6
Kehrig, H. A., Malm, O., and Moreira, I. (1998). Mercury in a widely consumed fish Micropogonias furnieri (Demarest, 1823) from four main Brazilian estuaries. Sci. Total Environ. 213, 263–271. doi: 10.1016/S0048-9697(98)00099-0
Kehrig, H. A., Pinto, F. N., Moreira, I., and Malm, O. (2003). Heavy metals and methylmercury in a tropical coastal estuary and a mangrove in Brazil. Org. Geochem. 34, 661–669. doi: 10.1016/S0146-6380(03)00021-4
Kehrig, H. D. A., Seixas, T. G., Palermo, E. A., Baêta, A. P., Castelo-Branco, C. W., Malm, O., et al. (2009). The relationships between mercury and selenium in plankton and fish from a tropical food web. Environ. Sci. Pollut. Res. 16, 10–24. doi: 10.1007/s11356-008-0038-8
Kim, B. S. M., Salaroli, A. B., Ferreira, P. A. L., Sartoretto, J. R., Mahiques, M. M., and Figueira, R. C. L. (2016). Spatial distribution and enrichment assessment of heavy metals in surface sediments from Baixada Santista, Southeastern Brazil. Mar. Pollut. Bull. 103, 333–338. doi: 10.1016/j.marpolbul.2015.12.041
Kutter, V. T., Mirlean, N., Baisch, P. R. M., Kutter, M. T., and Silva, E. (2009). Mercury in freshwater, estuarine, and marine fishes from Southern Brazil and its ecological implication. Environ. Monit. Assess. 159, 35–42. doi: 10.1007/s10661-008-0610-1
La Colla, N. S., Botté, S. E., Oliva, A. L., and Marcovecchio, J. E. (2017). Tracing Cr, Pb, Fe, and Mn occurrence in the Bahía Blanca estuary through commercial fish species. Chemosphere 175, 286–293. doi: 10.1016/j.chemosphere.2017.02.002
Lajud, N. A., Martín, J., de Astarloa, D., and González-Castro, M. (2016). Reproduction of Brevoortia aurea (Spix and Agassiz, 1829) (Actinopterygii: Clupeidae) in the Mar Chiquita Coastal Lagoon, Buenos Aires, Argentina. Neotrop. Ichthyol. 14:e150064. doi: 10.1590/1982-0224-20150064
Lanfranchi, A. L., Menone, M. L., Miglioranza, K. S. B., Janiot, L. J., Aizpún, J. E., and Moreno, V. J. (2006). Striped weakfish (Cynoscion guatucupa): a biomonitor of organochlorine pesticides in estuarine and near-coastal zones. Mar. Pollut. Bull. 52, 74–80. doi: 10.1016/j.marpolbul.2005.08.008
Lebreton, L. C. M., van der Zwet, J., Damsteeg, J.-W., Slat, B., Andrady, A., and Reisser, J. (2017). River plastic emissions to the world's oceans. Nat. Commun. 8:15611. doi: 10.1038/ncomms15611
Lima Junior, R. G. S., Araújo, F. G., Maia, M. F., and Braz Pinto, A. S. S. (2002). Evaluation of heavy metals in fish of sepetiba and ilha grande bays, Rio de Janeiro, Brazil. Envir. Res. Sec. 89, 171–179. doi: 10.1006/enrs.2002.4341
Lima, A. R. A., Barletta, M., and Costa, M. F. (2015). Seasonal distribution and interactions between plankton and microplastics in a tropical estuary. Estuar. Coast. Shelf Sci. 165, 213–225. doi: 10.1016/j.ecss.2015.05.018
Lima, A. R. A., Barletta, M., Costa, M. F., Ramos, J. A. A., Dantas, D. V., Melo, P. A. M. C., et al. (2016). Changes in the composition of ichthyoplankton assemblage and plastic debris in mangrove creeks relative to moon phases. J. Fish Biol. 89, 619–640. doi: 10.1111/jfb.12838
Lima, A. R. A., Barletta, M., Dantas, D. V., Possato, F. E., Ramos, J. A. A., and Costa, M. F. (2012). Early development and allometric shifts during the ontogeny of a marine catfish (Cathorops spixii-Ariidae). J. Appl. Ichthyol. 28, 217–225. doi: 10.1111/j.1439-0426.2011.01903.x
Lima, A. R. A., Barletta, M., Dantas, D. V., Ramos, J. A. A., and Costa, M. F. (2013). Early development of marine catfishes (Ariidae): from mouth brooding to the release of juveniles in nursery habitats. J. Fish Biol. 82, 1990–2014. doi: 10.1111/jfb.12128
Lima, A. R. A., Costa, M. F., and Barletta, M. (2014). Distribution patterns of microplastics within the plankton of a tropical estuary. Environ. Res. 132, 146–155. doi: 10.1016/j.envres.2014.03.031
Lima, M. W., Santos, M. L. S., Faial, K. C. F., Freitas, E. S., Lima, M. O., Pereira, J. A. R., et al. (2017). Heavy metals in the bottom sediments of the Furo of Laura estuary, Eastern Amazon, Brazil. Mar. Pollut. Bull. 118, 403–406. doi: 10.1016/j.marpolbul.2017.02.073
Lin, H. J., Kao, W. Y., and Wang, Y. T. (2007). Analyses of stomach contents and stable isotopes reveal food sources of estuarine detritivorous fish in tropical/subtropical Taiwan. Estuar. Coast. Shelf. Sci. 73, 527–537. doi: 10.1016/j.ecss.2007.02.013
Lopez Cazorla, A. (2000). Age structure of the population of weakfish Cynoscion guatucupa (Cuvier) in the Bahia Blanca waters, Argentina. Fish. Res. 46, 279–286. doi: 10.1016/S0165-7836(00)00152-1
Lopez Cazorla, A. (2005). On the age and growth of flounder Paralichthys orbignyanus (Jenyns,1842) in Bahiía Blanca Estuary, Argentina. Hydrobiologia 537, 81–87. doi: 10.1300/J028v11n01_06
Lopez Cazorla, A., and Forte, S. (2005). Food and feeding habits of flounder Paralichthys orbignyanus (Jenyns, 1842) in Bahía Blanca Estuary, Argentina. Hydrobiologia 549, 251–257. doi: 10.1007/s10750-005-5446-x
Lopez Cazorla, A., Molina, J. M., and Ruarte, C. (2014). The artisanal fishery of Cynoscion guatucupa in Argentina: exploring the possible causes of the collapse in Bahía Blanca estuary. J. Sea Res. 88, 29–35. doi: 10.1016/j.seares.2013.12.016
Lopez Cazorla, A., Pettigrosso, R. E., Tejera, L., and Camina, R. (2011). Diet and food selection by Ramnogaster arcuata (Osteichthyes, Clupeidae). J. Fish Biol. 78, 2052–2066. doi: 10.1111/j.1095-8649.2011.02995.x
López, S., Mabragaña, E., Díaz De Astarloa, J. M., and González-Castro, M. (2015). Reproductive studies of Anchoa marinii Hildebrand, 1943 (Actinopterygii: Engraulidae) in the nearby-coastal area of Mar Chiquita coastal lagoon, Buenos Aires, Argentina. Neotrop. Ichthyol. 13, 221–228. doi: 10.1590/1982-0224-20140083
Lorenzo, M. I., Dìaz de Astarloa, J. M., Norbis, W., and Cousseau, M. B. (2011). Long term fish assemblages as units of management in a temperate estuary (Rio De La Plata - SW Atlantic ocean). Brazilian J. Oceanogr. 59, 43–59. doi: 10.1590/S1679-87592011000100004
Lotze, H. K., Lenihan, H. S., Bourque, B. J., Bradbury, R. H., Cooke, R. G., Kay, M. C., et al. (2006). Depletion, degradation, and recovery potential of estuaries and coastal seas. Science 312, 1806–1809. doi: 10.1126/science.1128035
Loureiro, S. N., Mourão, M Jr., and Giarrizzo, T. (2017). Length–weight relationships for seven fish species from Marajó Bay, Amazon estuary, northern Brazil. J. Appl. Ichthyol. 33, 620–622. doi: 10.1111/jai.13281
Love, J. W., and May, E. B. (2007). Relationships between fish assemblage structure and selected environmental factors in Maryland's Coastal Bays. Northeast. Nat. 14, 251–268. doi: 10.1656/1092-6194(2007)14[251:rbfasa]2.0.co;2
Lozoya, J. P., Carranza, A., Lenzi, J., Machín, E., Teixeira De Mello, F., González, S., et al. (2015). Management and research on plastic debris in Uruguayan Aquatic Systems: update and perspectives. J. Integr. Coast. Zo. Manag.—Rev. Gestão Costeira Integr. 15, 377–393. doi: 10.5894/rgci583
Macchi, G. J., and Acha, E. M. (2000). Spawning frequency and batch fecundity of Brazilian menhaden, Brevoortia aurea, in the Rio de la Plata estuary off Argentina and Uruguay. Fish. Bull. 98, 283–289.
Macchi, G. J., Acha, E. M., and Lasta, C. A. (2002). Reproduction of black drum (Pogonias cromis) in the Rio de la Plata estuary, Argentina. Fish. Res. 59, 83–92. doi: 10.1016/S0165-7836(01)00410-6
Macchi, G. J., Acha, E. M., and Militelli, M. I. (2003). Seasonal egg production of whitemouth croaker (Micropogonias furnieri) in the Rio de la Plata estuary, Argentina-Uruguay. Fish. Bull. 101, 332–342.
Machado, A. A. S., Spence, K., Kloas, W., Toffolon, M., and Zarfl, C. (2016). Metal fate and effects in estuaries: a review and conceptual model for better understanding of toxicity. Sci Total Envir. 541, 268–281. doi: 10.1016/j.scitotenv.2015.09.045
Machado, I., Vera, M., Calliari, D., and Rodríguez-Graña, L. (2012). First record of an Elops smithi (Pisces: Elopidae) larva in a South American subtropical-temperate estuary. Mar. Biodivers. Rec. 5:e108. doi: 10.1017/S1755267212000917
Machado, W., Santelli, R. E., Loureiro, D. D., Oliveira, E. P., Borges, A. C., Ma, V. K., et al. (2008). Mercury accumulation in sediments along an eutrophication gradient in Guanabara Bay, Southeast Brazil. J. Braz. Chem. Soc. 19, 569–575. doi: 10.1590/S0103-50532008000300028
Mancilla, S., and Rubio, E. (1992). “Biología alimentaria del róbalo de giba Centropomus unionensis (Brocourt 1868) (Pisces: Centropomidae) en áreas aledañas a la Bahía de Buenaventura y Málaga,” in Comisión Colombiana de Oceanografía, Memorias VIII Seminario Nacional de las Ciencias y las Tecnologías del Mar (Oxford: Blackwell), 940–951.
Marco, S. G., Botté, S. E., and Marcovecchio, J. E. (2006). Mercury distribution in abiotic and biological compartments within several estuarine systems from Argentina: 1980-2005 period. Chemosphere 65, 213–223. doi: 10.1016/j.chemosphere.2006.02.059
Marcovecchio, J. E., Andrade, S., Ferrer, L. D., Asteasuain, R. O., De Marco, S. G., Gavio, M. A., et al. (2001). Mercury distribution in estuarine environments from Argentina: the detoxification and recovery of salt marshes after 15 years. Wetl. Ecol. Manag. 9, 317–322. doi: 10.1023/A:1011860618461
Marcovecchio, J. E., Botté, S. E., and Fernández Severini, M. D. (2016). Distribution and behavior of zinc in estuarine environments: an overview on Bahía Blanca estuary (Argentina). Environ. Earth Sci. 75:1168. doi: 10.1007/s12665-016-5942-5
Marcovecchio, J. E., Moreno, V. J., and Pérez, A. (1986). Bio-magnification of total mercury in Bahia Blanca estuary shark. Mar. Pollut. Bull. 17, 276–278. doi: 10.1016/0025-326X(86)90064-0
Marcovecchio, J. E., Moreno, V. J., and Perez, A. (1988a). Determination of heavy metal concentrations in biota of Bahia Blanca, Angentina. Sci. Total Environ. 75, 181–190. doi: 10.1016/0048-9697(88)90031-9
Marcovecchio, J. E., Moreno, V. J., and Perez, A. (1988b). The sole, Paralichthys sp., as an indicator species for heavy metal pollution in the bahia blanca estuary, Argentina. Sci. Total Environ. 75, 191–199. doi: 10.1016/0048-9697(88)90032-0
Marcovecchio, J. E., Moreno, V. J., and Pérez, A. (1991). Metal accumulation in tissues of sharks from the Bahía Blanca estuary, Argentina. Mar. Environ. Res. 31, 263–274. doi: 10.1016/0141-1136(91)90016-2
Martínez-Huitle, C. A., and Ferro, S. (2016). Electrochemical oxidation of organic pollutants for the wastewater treatment: direct and indirect processes. Chem. Soc. Rev. 35, 1324–1340. doi: 10.1039/b517632h
Martinho, F., Leitão, R., Neto, J. M., Cabral, H. N., Marques, J. C., and Pardal, M. A. (2007). The use of nursery areas by juvenile fish in a temperate estuary, Portugal. Hydrobiologia 587, 281–290. doi: 10.1007/s10750-007-0689-3
Martins, C. C., Braun, J. A. F., Seyffert, B. H., Machado, E. C., and Fillmann, G. (2010). Anthropogenic organic matter inputs indicated by sedimentary fecal steroids in a large South American tropical estuary (Paranaguá estuarine system, Brazil). Mar. Pollut. Bull. 60, 2137–2143. doi: 10.1016/j.marpolbul.2010.07.027
Martins, C. C., Cabral, A. C., Barbosa-Cintra, S. C. T., Dauner, A. L. L., and Souza, F. M. (2014). An integrated evaluation of molecular marker indices and linear alkylbenzenes (LABs) to measure sewage input in a subtropical estuary (Babitonga Bay, Brazil). Environ. Pollut. 188, 71–80. doi: 10.1016/j.envpol.2014.01.022
Martins, C. D. C., Fillmann, G., and Montone, R. C. (2007). Natural and anthropogenic sterols inputs in surface sediments of Patos Lagoon, Brazil. J. Braz. Chem. Soc. 18, 106–115. doi: 10.1590/S0103-50532007000100012
Martins, D. M. F., Chagas, R. M., Melo Neto, J. O., and Méllo Júnior, A. V. (2011). Impactos da construção da usina hidrelétrica de Sobradinho no regime de vazões no Baixo São Francisco. Rev. Bras. Eng. Agrícola e Ambient. 15, 1054–1061. doi: 10.1590/S1415-43662011001000010
Massone, C. G., Wagener, A. L. R., Abreu, H. M., and Veiga, Á. (2013). Revisiting hydrocarbons source appraisal in sediments exposed to multiple inputs. Mar. Pollut. Bull. 73, 345–354. doi: 10.1016/j.marpolbul.2013.05.043 Baseline
McGrath, D. G., Castro, F., Futemma, C., Amaral, B. D., and Calabria, J. (1993). Fisheries and the evolution of resource management on the lower Amazon floodplain. Hum. Ecol. 21, 167–195. doi: 10.1007/BF00889358
McKinley, E., Aller-Rojas, O., Hattam, C., Germond-Duret, C., Martín, I. V. S., Hopkins, C. R., et al. (2018). Charting the course for a blue economy in Peru: a research agenda. Environ. Dev. Sustain. 23, 1–23. doi: 10.1007/s10668-018-0133-z
Medeiros, A. M., Luiz, O. J., and Domit, C. (2015). Occurrence and use of an estuarine habitat by giant manta ray Manta birostris. J. Fish Biol. 86, 1830–1838. doi: 10.1111/jfb.12667
Medeiros, P. M., Bícego, M. C., Castelao, R. M., Del Rosso, C., Fillmann, G., and Zamboni, A. J. (2005). Natural and anthropogenic hydrocarbon inputs to sediments of Patos Lagoon Estuary, Brazil. Environ. Int. 31, 77–87. doi: 10.1016/j.envint.2004.07.001
Medeiros, P. R. P., Knoppers, B. A., Cavalcante, G. H., and Souza, W. F. L. (2011). Changes in nutrient loads (N, P, and Si) in the Sao Francisco Estuary after the construction of dams. Brazilian Arch. Biol. Technol. 54, 387–397. doi: 10.1590/S1516-89132011000200022
Medina-Contreras, D., Cantera, J., Escarria, E., and Mejía-Ladino, L. M. (2014). Distribution and density of ichthyoplankton in the Bahía Málaga estuary, Pacific coast of Colombia (September 2009-February 2010). Boletín Investig. Mar. Y Costeras 43, 107–119.
Mendis, B. R. C., Najim, M. M. M., Kithsiri, H. M. P., and Azmy, S. A. M. (2015). Bioaccumulation of heavy metals in the selected commercially important edible fish species gray mullet (Mugil cephalus) from Negombo Estuary. J. Environ. Prof. Sri Lanka 4, 1–9. doi: 10.4038/jepsl.v4i2.7858
Mendoza-Carranza, M., and Vieira, J. (2008). Whitemouth croaker Micropogonias furnieri (Desmarest, 1823) feeding strategies across four southern Brazilian estuaries. Aquat. Ecol. 42, 83–93. doi: 10.1007/s10452-007-9084-4
Menone, M. L., Aizpún De Moreno, J. E., Moreno, V. J., Lanfranchi, A. L., Metcalfe, T. L., and Metcalfe, C. D. (2000). PCBs and organochlorines in tissues of silverside (Odontesthes bonariensis) from a coastal lagoon in Argentina. Arch. Environ. Contam. Toxicol. 38, 202–208. doi: 10.1007/s002449910027
Militelli, M. I., and Macchi, G. J. (2006). Spawning and fecundity of striped weakfish, Cynoscion guatucupa, in the Río de la Plata estuary and adjacent marine waters, Argentina-Uruguay. Fish. Res. 77, 110–114. doi: 10.1016/j.fishres.2005.08.004
Mirlean, N., Baraj, B., Niencheski, L. F., Baisch, P., and Robinson, D. (2001). The effect of accidental sulphuric acid leaking on metal distributions in estuarine sediment of patos lagoon. Mar. Pollut. Bull. 42, 1114–1117. doi: 10.1016/S0025-326X(01)00099-6
Mizerkowski, B. D., Machado, E. C., Brandini, N., Nazario, M. G., and Bonfim, K. V. (2012). Environmental water quality assessment in Guaratuba bay, state of Paraná, southern Brazil. Brazilian J. Oceanogr. 60, 109–115. doi: 10.1590/S1679-87592012000200001
Molina, L. M., Valiñas, M. S., Pratolongo, P. D., Elias, R., and Perillo, G. M. E. (2017). Effect of “Whitemouth Croaker” (Micropogonias furnieri, Pisces) on the stability of the sediment of Salt Marshes - an Issue to be resolved. Estuar. Coasts 40, 1–13. doi: 10.1007/s12237-017-0237-y
Mont'Alverne, R., Jardine, T. D., Pereyra, P. E. R., Oliveira, M. C. L. M., Medeiros, R. S., Sampaio, L. A., et al. (2016). Elemental turnover rates and isotopic discrimination in a euryhaline fish reared under different salinities: implications for movement studies. J. Exp. Mar. Bio. Ecol. 480, 36–44. doi: 10.1016/j.jembe.2016.03.021
Monteiro, D. P., Giarrizzo, T., and Isaac, V. (2009). Feeding ecology of juvenile dog snapper Lutjanus jocu (Bloch and Shneider, 1801) (Lutjanidae) in intertidal mangrove creeks in Curuçá estuary (Northern Brazil). Brazil. Arch. Biol. Technol. 52, 1421–1430. doi: 10.1590/S1516-89132009000600014
Monteiro, M. C., Jiménez, J. A., and Pereira, L. C. C. (2016a). Natural and human controls of water quality of an Amazon estuary (Caeté-PA, Brazil). Ocean Coast. Manag. 124, 42–52. doi: 10.1016/j.ocecoaman.2016.01.014
Monteiro, M. C., Pereira, L. C. C., and Jimenez, J. A. (2016b). The trophic status of an Amazonian Estuary under anthropogenic pressure (Brazil). J. Coast. Res. 75, 98–102. doi: 10.2112/SI75-020.1
Moresco, A., and Bemvenuti, M. A. (2006). Biologia reprodutiva do peixe-rei Odontesthes argentinensis (Valenciennes) (Atherinopsidae) da região marinha costeira do sul do Brasil. Rev. Bras. Zool. 23, 1168–1174. doi: 10.1590/S0101-81752006000400025
Mourão, K. R., Fredou, T., and Lucena Fredou, F. (2015). Spatial and seasonal variation of the ichthyofauna and habitat use in the inner portion of the Brazilian Amazon Estuary. Bol. Do Inst. Pesca 41, 529–545
Mourão, K. R. M., Ferreira, V., and Lucena-Frédou, F. (2014). Composition of functional ecological guilds of the fish fauna of the internal sector of the amazon estuary, pará, Brazil. An. Acad. Bras. Cienc. 86, 1783–1800. doi: 10.1590/0001-3765201420130503
Mulato, I. P., Corrêa, B., and Vianna, M. (2015). Time-space distribution of Micropogonias furnieri (Perciformes, sciaenidae) in a tropical estuary in southeastern Brazil. Bol. do Inst. Pesca 41, 1–18.
Nagy, G. J., Gómez-Erache, M., López, C. H., and Perdomo, A. C. (2002). Distribution patterns of nutrients and symptoms of eutrophication in the Río de la Plata estuarine system. Hydrobiologia 475, 125–139. doi: 10.1023/A:1020300906000
Nayar, S., Goh, B. P. L., Chou, L. M., and Reddy, S. (2003). In situ microcosms to study the impact of heavy metals resuspended by dredging on periphyton in a tropical estuary. Aquat. Toxicol. 64, 293–306. doi: 10.1016/S0166-445X(03)00062-6
Nestler, J. M., Pompeu, P. S., Goodwin, R. A., Smith, D. L., Silva, L. G. M., Baigún, C. R. M., et al. (2012). The river machine: a template for fish movement and habitat, fluvial geomorphology, fluid dynamics and biogeochemical cycling. River Res. Appl. 28, 490–503. doi: 10.1002/rra.1567
Neuhaus, E. B., Antonetti, D. A., and Schulz, U. H. (2016). The invasive fish Acestrorhynchus pantaneiro Menezes, 1992 in Southern Brazil: occurrence and food niche overlap with two native species. J. Appl. Ichthyol. 32, 1107–1112. doi: 10.1111/jai.13147
Niencheski, L. F., Windom, H. L., Baraj, B., Wells, D., and Smith, R. (2001). Mercury in fish from patos and Mirim Lagoons, Southern Brazil. Mar. Pollut. Bull. 42, 1403–1406. doi: 10.1016/S0025-326X(01)00219-3
Niencheski, L. F., Windom, H. L., and Smith, R. (1994). Distribution of particulate trace metal in Patos Lagoon estuary (Brazil). Mar. Pollut. Bull. 28, 96–102. doi: 10.1016/0025-326X(94)90545-2
Ochoa-Muñoz, M. J., Valenzuela, C. P., Toledo, S., Bustos, C. A., and Landaeta, M. F. (2013). Feeding of a larval clinid fish in a microtidal estuary from southern Chile. Rev. Biol. Mar. Oceanogr. 48, 45–57. doi: 10.4067/S0718-19572013000100005
Oliva, A. L., Quintas, P. Y., La Colla, N. S., Arias, A. H., and Marcovecchio, J. E. (2015). Distribution, sources, and potential ecotoxicological risk of Polycyclic Aromatic Hydrocarbons in surface sediments from Bahía Blanca Estuary, Argentina. Arch. Environ. Contam. Toxicol. 69, 163–172. doi: 10.1007/s00244-015-0169-0
Oliveira, A. M. E. (1974). Ictiofauna das águas estuarinas do Rio Parnaíba (Brasil). Arq. Ciências do Mar 14, 41–45.
Oliveira, E. C., and Favaro, L. F. (2011). Reproductive biology of the flatfish Etropus crossotus (Pleuronectiformes: Paralichthyidae) in the Paranaguá estuarine complex, Paraná State, subtropical region of Brazil. Neotrop. Ichthyol. 9, 795–805. doi: 10.1590/S1679-62252011005000043
Oliveira, M. F., Santos Costa, E. F. D., Freire, F., Oliveira, J. E. L., and Luchiari, A. C. (2011). Some aspects of the biology of white mullet, Mugil curema (Osteichthyes, Mugilidae), in the northeastern region, Brazil. Panam. J. Aquat. Sci. 6, 138–147.
Palma-Fleming, H., Asencio, P. A. J, and Gutierrez, E. (2004). Polycyclic aromatic hydrocarbons in sediments and mussels of Corral Bay, south central Chile. J. Environ. Monitir. 6, 229–233. doi: 10.1039/b307018b
Palma-Fleming, H., Quiroz, R. E, Campillay, C., Figueroa, M., Varas, A., et al. (2012). Temporal and spatial trends of total Aliphatic Hydrocarbons of diesel range and trace elements in sediments and mussels of the Corral Bay Area, Valdivia, South Central Chile. J. Chil. Chem. Soc. 57, 1074–1082. doi: 10.4067/S0717-97072012000200003
Paranhos, R., Pereira, A. P., and Mayr, L. M. (1998). Diel variability of water quality in a tropical polluted bay. Environ. Monit. Assess. 50, 131–141. doi: 10.1023/A:1005855914215
Paraquetti, H. H. M., Ayres, G. A., De Almeida, M. D., Molisani, M. M., and De Lacerda, L. D. (2004). Mercury distribution, speciation and flux in the Sepetiba Bay tributaries, SE Brazil. Water Res. 38, 1439–1448. doi: 10.1016/j.watres.2003.11.039
Passos, A. C., Contente, R. F., Abbatepaulo, F. V., Spach, H. L., Vilar, C. C., Joyeux, J. C., et al. (2013). Analysis of fish assemblages in sectors along a salinity gradient based on species, families and functional groups. Br. J. Oceanogr. 61, 251–264. doi: 10.1590/S1679-87592013000400006
Passos, C. V. B., Fabré, N. N., Malhado, A. C. M., Batista, V. S., and Ladle, R. J. (2016). Estuarization increases functional diversity of demersal fish assemblages in tropical coastal ecosystems. J. Fish Biol. 89, 847–862. doi: 10.1111/jfb.13029
Paula, A. L. S., Lima, B. K. S., and Maia, R. C. (2016). The recovery of a degraded mangrove in Ceará through the production of Laguncularia racemosa (L.) C.F. Gaertn. (Combretaceae) and Avicennia sp. Stapf ex Ridl (Acanthaceae) seedlings. Rev Árvore 40, 377–385. doi: 10.1590/0100-67622016000300001
Pazos, R. S., Maiztegui, T., Colautti, D. C., Paracampo, A. H., and Gómez, N. (2017). Microplastics in gut contents of coastal freshwater fish from Río de la Plata estuary. Mar. Pollut. Bull. 122, 85–90. doi: 10.1016/j.marpolbul.2017.06.007
Pereira, T. S., Moreira, Í. T. A., Oliveira, O. M. C., Rios, M. C., Filho, W. A. C. S., Almeida, M., et al. (2015). Distribution and ecotoxicology of bioavailable metals and as in surface sediments of Paraguaçu estuary, Todos os Santos Bay, Brazil. Mar. Pollut. Bull. 99, 166–177. doi: 10.1016/j.marpolbul.2015.07.031
Pereyra, P. E. R., Mont'Alverne, R., and Garcia, A. M. (2016). Carbon primary sources and estuarine habitat use by two congeneric ariid catfishes in a subtropical coastal lagoon. Zoologia 33, 1–7. doi: 10.1590/S1984-4689zool-20150075
Pessanha, A. L. M., and Araújo, F. G. (2003). Spatial, temporal and diel variations of fish assemblages at two sandy beaches in the Sepetiba Bay, Rio de Janeiro, Brazil. Estuar. Coast. Shelf Sci. 57, 817–828. doi: 10.1016/S0272-7714(02)00411-0
Pierini, J. O., Streitenberger, M. E., and Baldini, M. D. (2012). Evaluation of faecal contamination in Bahia Blanca estuary (Argentina) using a numerical model. Rev. Biol. Mar. Oceanogr. 47, 193–202. doi: 10.4067/S0718-19572012000200003
Pinaya, W. H. D., Lobon-Cervia, F. J., Pita, P., Souza, R. B., Freire, J., and Isaac, V. J. (2016). Multispecies fisheries in the lower amazon river and its relationship with the regional and global climate variability. PLoS ONE 11:e0157050. doi: 10.1371/journal.pone.0157050
Pinochet, H., De Gregori, I., Delgado, D., Gras, N., Muñoz, L., Bruhn, C., et al. (1995). Cadmium and copper in bivalves mussels and associated bottom sediments and waters from corral bay-chile. Environ. Technol. 16, 539–548. doi: 10.1080/09593331608616295
Possatto, F. E., Barletta, M., Costa, M. F., Ivar do Sul, J. A., and Dantas, D. V. (2011). Plastic debris ingestion by marine catfish: an unexpected fisheries impact. Mar. Pollut. Bull. 62, 1098–1102. doi: 10.1016/j.marpolbul.2011.01.036
Possatto, F. E., Broadhurst, M. K., Spach, H. L., Winemiller, K. O., Millar, R. B., Santos, K. M., et al. (2016). Mapping the spatio-temporal distribution of threatened batoids to improve conservation in a subtropical estuary. J. Fish Biol. 89, 1098–1104. doi: 10.1111/jfb.12972
Possatto, F. E., Spach, H. L., Cattani, A. P., Lamour, M. R., Santos, L. O., Cordeiro, N. M. A., et al. (2015). Marine debris in a world heritage listed Brazilian estuary. Mar. Pollut. Bull. 91, 548–553. doi: 10.1016/j.marpolbul.2014.09.032
Pratolongo, P., Mazzon, C., Zapperi, G., Piovan, M. J., and Brinson, M. M. (2013). Land cover changes in tidal salt marshes of the Bahía Blanca estuary (Argentina) during the past 40 years. Estuar. Coast. Shelf Sci. 133, 23–31. doi: 10.1016/j.ecss.2013.07.016
Prestrelo, L., and Monteiro-Neto, C. (2016). Before–after environmental impact assessment of an artificial channel opening on a south-western Atlantic choked lagoon system. J. Fish Biol. 89, 735–752. doi: 10.1111/jfb.13012
Qu, X., Alvarez, P. J. J., and Li, Q. (2013). Applications of nanotechnology in water and wastewater treatment. Water Res. 47, 3931–3946. doi: 10.1016/j.watres.2012.09.058
Ramos, J. A. A., Barletta, M., and Costa, M. F. (2012). Ingestion of nylon threads by gerreidae while using a tropical estuary as foraging grounds. Aquat. Biol. 17, 29–34. doi: 10.3354/ab00461
Ramos, J. A. A., Barletta, M., Dantas, D. V., and Costa, M. F. (2016). Seasonal and spatial ontogenetic movements of Gerreidae in a Brazilian tropical estuarine ecocline and its application for nursery habitat conservation. J. Fish Biol. 89, 696–712. doi: 10.1111/jfb.12872
Ramos, J. A. A., Barletta, M., Dantas, D. V., Lima, A. R. A., and Costa, M. F. (2011). Influence of moon phase on fish assemblages in estuarine mangrove tidal creeks. J. Fish Biol. 78, 344–354. doi: 10.1111/j.1095-8649.2010.02851.x
Reis, R. E., Albert, J. S., Dario, F. D., Mincarones, M. M., Petry, P., and Rocha, L. A. (2016). Fish biodiversity and conservation in South America. J. Fish Biol. 89, 12–47. doi: 10.1111/jfb.13016
Reis-Filho, J. A., Nunes, J. D. A. C. D. C., and Ferreira, A. (2010). Estuarine ichthyofauna of the Paraguaçu River, Todos os Santos Bay, Bahia, Brazil. Biota Neotrop. 10, 301–312. doi: 10.1590/S1676-06032010000400034
Reis-Filho, J. A., and Santos, A. C. A. (2014). Effects of substratum type on fish assemblages in shallow areas of a tropical estuary. Mar. Ecol. 35, 456–470. doi: 10.1111/maec.12102
Rezende, C. E., Kahn, J. R., Passareli, L., and Vásquez, W. F. (2015). An economic valuation of mangrove restoration in Brazil. Ecol. Econ. 120, 296–302. doi: 10.1016/j.ecolecon.2015.10.019
Ribeiro, C. H. A., and Kjerfve, B. (2002). Anthropogenic influence on the water quality in Guanabara Bay, Rio de Janeiro, Brazil. Reg. Environ. Chang. 3, 13–19. doi: 10.1007/s10113-001-0037-5
Ribeiro, J. S., de Oliveira, F. C. R., and Ederli, N. B. (2017). Short communication: first report of nematodes parasitizing the four-eyed-fish, Anableps anableps (Pisces, Cyprinodontiformes). Parasitol. Res. 116, 2249–2254. doi: 10.1007/s00436-017-5528-5
Ríos-Pulgarín, M. I., Barletta, M., and Mancera-Rodriguez, N. J. (2016). Hydrological cycle effects on the aquatic community in a Neotropical stream of the Andean piedmont during the 2007–2010 ENSO events. J. Fish Biol. 89, 131–156. doi: 10.1111/jfb.12885
Rizzi, J., Taniguchi, S., and Martins, C. C. (2017). Polychlorinated Biphenyls (PCBs) and Organochlorine Pesticides (OCPs) in sediments from an urban- and industrial-impacted subtropical estuary (Babitonga Bay, Brazil). Mar. Pollut. Bull. 119, 390–395. doi: 10.1016/j.marpolbul.2017.03.032
Rodrigues, M. L., Santos, R. B., Santos, E. J. S., Pereira, S. M., Oliveira, A., and Soares, E. C. (2017). Biologia populacional da carapeba listrada, Eugerres brasilianus (Cuvier, 1830), próximo à foz do Rio São Francisco (Brasil). Bol. do Inst. Pesca 43, 152–163. doi: 10.20950/1678-2305.2017v43n2p152
Rodrigues, S. K., Abessa, D. M. S., and Machado, E. C. (2013). Geochemical and ecotoxicological assessment for estuarine surface sediments from Southern Brazil. Mar. Environ. Res. 91, 68–79. doi: 10.1016/j.marenvres.2013.02.005
Rodrigues-Barreto, N., Shimada-Brotto, D. V., Guterres-Giordano, R., Andrade-Bertoncini, A., and Santos, L. N. (2017). The rocky reef fishes of Vermelha Beach, a marine estuarine transitional zone at Guanabara Bay, Brazil. Lat. Am. J. Aquat. Res. 45, 33–40. doi: 10.3856/vol45-issue1-fulltext-4
Rodriguez, M. S., and Reis, R. E. (2008). Taxonomic review of Rineloricaria (Loricariidae: Loricariinae) from the Laguna dos Patos drainage, Southern Brazil, with the descriptions of two new species and the recognition of two species groups. Copeia 2008, 333–349. doi: 10.1643/CI-06-218
Ronda, A. C., Oliva, A. L., Arias, A. H., Orazi, M. M., and Marcovecchio, J. E. (2018). Biomarker responses to polycyclic aromatic hydrocarbons in the native fish Ramnogaster arcuata, South America. Int. J. Environ. Res. 13, 77–89. doi: 10.1007/s41742-018-0155-2
Rubio, E. A. (1984a). Estudio taxonómico preliminar de la ictiofauna de la Bahía de Málaga, Colombia. Cespedesia 13, 97–111.
Rubio, E. A. (1984b). Estudio taxonómico preliminar de la ictiofauna de Bahía Málaga (Pacífico colombiano). An. Inst. Invest. Mar. Punta de Betin 14, 157–173.
Rueda, M. (2001). Spatial distribution of fish species in a tropical estuarine lagoon: a geostatistical appraisal. Mar. Ecol. Prog. Ser. 222, 217–226. doi: 10.3354/meps222217
Rueda, M., and Defeo, O. (2003). Spatial structure of fish assemblages in a tropical estuarine lagoon: combining multivariate and geostatistical techniques. J. Exp. Mar. Bio. Ecol. 296, 93–112. doi: 10.1016/S0022-0981(03)00319-8
Rueda, M., and Santos-Martínez, A. (1999). Population dynamics of the striped mojarra Eugerres plumieri from the Cienaga Grande de Santa Marta, Colombia. Fish. Res. 42, 155–166. doi: 10.1016/S.0165-7836(99)00033-8
Sá, F., Machado, E. C., Angulo, R. J., Veiga, F. A., and Brandini, N. (2006). Arsenic and heavy metals in sediments near Paranaguá Port, Southern Brazil. J. Coast. Res. 39, 1066–1068.
Salgado, L. D., Marques, A. E. M. L., Kramer, R. D., Oliveira, F. G., Moretto, S. L., Lima, B. A., et al. (2018). Integrated assessment of sediment contaminant levels and biological responses in sentinel fish species Atherinella brasiliensis from a sub-tropical estuary in south Atlantic. Chemosphere 219, 15–27. doi: 10.1016/j.chemosphere.2018.11.204
Sanders, C. J., Santos, I. R., Silva-Filho, E. V., and Patchineelam, S. R. (2008). Contrasting mercury and manganese deposition in a mangrove-dominated estuary (Guaratuba Bay, Brazil). Geo-Marine Lett. 28, 239–244. doi: 10.1007/s00367-008-0104-8
Santos, D. M., Santos, G. S., Cestari, M. M., Oliveira Ribeiro, C. A., Assis, H. C. S., Yamamoto, F., et al. (2014). Bioaccumulation of butyltins and liver damage in the demersal fish Cathorops spixii (Siluriformes, Ariidae). Environ. Sci. Pollut. Res. 21, 3166–3174. doi: 10.1007/s11356-013-2280-y
Santos, E. S., Carreira, R. S., and Knoppers, B. A. (2008). Sedimentary sterols as indicators of environmental conditions in Southeastern Guanabara Bay, Brazil. Brazilian J. Oceanogr. 56, 97–113. doi: 10.1590/S1679-87592008000200003
Santos, J. A. P., Schmiegelow, J. M. M., Rotundo, M. M., and Barrella, W. (2015). Composition and temporal variation of the fish assemblages in upper Santos Estuarine System, São Paulo, Brazil. Bol. Inst. Pesca 41, 945–959.
Santos, N. B., Rocha, R. M., and Lucena Fredóu, F. (2010). Reproductive biology of Plagioscion magdalenae (Teleostei: Sciaenidae) (Steindachner, 1878) in the bay of Marajo, Amazon Estuary, Brazil. Neotrop. Ichthyol. 8, 333–340. doi: 10.1590/S1679-62252010000200012
Santos, S. R., Andrade, A. C., Verani, J. R., and Vianna, M. (2015). Population explosion of the burrfish Chilomycterus spinosus spinosus (Diodontidae, Tetraodontiformes) in a eutrophic tropical estuary. Mar. Biol. Res. 11, 955–964. doi: 10.1080/17451000.2015.1045000
Santucci, L., Carol, E., Borzi, G., and García, M. G. (2017). Hydrogeochemical and isotopic signature of surface and groundwater in a highly industrialized sector of the Rio de la Plata coastal plain (Argentina). Mar. Pollut. Bull. 120, 387–395. doi: 10.1016/j.marpolbul.2017.05.007
Sardiña, P., and Lopez Cazorla, A. (2005a). Trophic ecology of the whitemouth croaker, Micropogonias furnieri (Pisces: Sciaenidae), in south-western Atlantic waters. J. Mar. Biol. Assoc. 85, 405–413. doi: 10.1017/S0025315405011331h
Sardiña, P., and Lopez Cazorla, A. (2005c). Feeding interrelationships and comparative morphology of two young sciaenids co-occurring in South-western Atlantic waters. Hydrobiologia 548, 41–49. doi: 10.1007/s10750-005-3643-2
Sardiña, P., and Lopez Cazorla, A. C. (2005b). Feeding habits of the juvenile striped weakfish, Cynoscion guatucupa Cuvier 1830, in Bahía Blanca estuary (Argentina): seasonal and ontogenetic changes. Hydrobiologia 532, 23–38. doi: 10.1007/s10750-004-8769-0
Schwamborn, R., and Giarrizzo, T. (2015). Stable isotope discrimination by consumers in a tropical mangrove food web: how important are variations in C/N ratio? Estuar. Coast. 38, 813–825. doi: 10.1007/s12237-014-9871-9
Segura, F. R., Nunes, E. A., Paniz, F. P., Paulelli, A. C. C., Rodrigues, G. B., Braga, G. Ú. L., et al. (2016). Potential risks of the residue from Samarco's mine dam burst (Bento Rodrigues, Brazil). Environ. Pollut. 218, 813–825. doi: 10.1016/j.envpol.2016.08.005
Seixas, T. G., Moreira, I., Malm, O., and Kehrig, H. A. (2012). Mercury and selenium in a top-predator fish, Trichiurus lepturus (Linnaeus, 1758), from the tropical Brazilian coast, Rio de Janeiro. Bull. Environ. Contam. Toxicol. 89, 434–438. doi: 10.1007/s00128-012-0680-1
Seyboth, E., Condini, M. V., Albuquerque, C. Q., Varela, A. S., Velasco, G., Vieira, J. P., et al. (2011). Age, growth, and reproductive aspects of the dusky grouper Mycteroperca marginata (Actinopterygii: Epinephelidae) in a man-made rocky habitat in southern Brazil. Neotrop. Ichthyol. 9, 849–856. doi: 10.1590/S1679-62252011005000038
Shervette, V. R., Aguirre, W. E., Blacio, E., Cevallos, R., Gonzalez, M., Pozo, F., et al. (2007). Fish communities of a disturbed mangrove wetland and an adjacent tidal river in Palmar, Ecuador. Estuar. Coast. Shelf Sci. 72, 115–128. doi: 10.1016/j.ecss.2006.10.010
Silva, A. M. F., Lemes, V. R. R., Barretto, H. H. C., Oliveira, E. S., Alleluia, I. B., and Paumgartten, F. J. R. (2003). Polychlorinated Biphenyls and Organochlorine Pesticides in edible fish species and dolphins from Guanabara Bay, Rio de Janeiro, Brazil. Bull. Environ. Contam. Toxicol. 70, 1151–1157. doi: 10.1007/s00128-003-0102-5
Silva, D. R. Jr., Paranhos, R., and Vianna, M. (2016). Spatial patterns of distribution and the influence of seasonal and abiotic factors on demersal ichthyofauna in an estuarine tropical bay. J. Fish Biol. 89, 821–846. doi: 10.1111/jfb.13033
Silva, J. D. B., Barletta, M., Lima, A. R. A., and Ferreira, G. V. B. (2018). Use of resources and microplastic contamination throughout the life cycle of grunts (Haemulidae) in a tropical estuary. Environ. Pollut. 242, 1010–1021. doi: 10.1016/j.envpol.2018.07.038
Silva, D. R Jr., Carvalho, D. M. T., and Vianna, M. (2013). The catfish Genidens genidens (Cuvier, 1829) as a potential sentinel species in Brazilian estuarine waters. J. Appl. Ichthyol. 29, 1297–1303. doi: 10.1111/jai.12280
Silva, P. D., Lisboa, P. V., and Fernandes, E. H. (2015). Changes on the fine sediment dynamics after the port of rio grande expansion. Adv. Geosci. 39, 123–127. doi: 10.5194/adgeo-39-123-2015
Silva, T. F., Azevedo, D. A., and Neto, F. R. A. (2007a). Distribution of polycyclic aromatic hydrocarbons in surface sediments and waters from guanabara Bay, Rio de Janeiro, Brazil. J. Braz. Chem. Soc. 18, 628–637. doi: 10.1590/S0103-50532007000300021
Silva, T. F., Azevedo, D. A., and Neto, F. R. A. (2007b). Polycyclic Aromatic Hydrocarbons in fishes and sediments from the Guanabara Bay, Brazil. Environ. Forensics 8, 257–264. doi: 10.1080/15275920701506433
Silveira, A. E. F., Nascimento, J. R., Sabadini-Santos, E., and Bidone, E. D. (2017). Screening-level risk assessment applied to dredging of polluted sediments from Guanabara Bay, Rio de Janeiro, Brazil. Mar. Pollut. Bull. 118, 368–375. doi: 10.1016/j.marpolbul.2017.03.016
Slater, F. M. (2016). Ecotones, ecoclines and eco-perturbations: the aquatic flora and fauna of the S'Albufera Natural Park, Majorca, a contribution and review. Mediterr. J. Biosci. 1, 120–127
Soares-Gomes, A., Neves, R. L., Aucélio, R., Van Der Ven, P. H., Pitombo, F. B., Mendes, C. L. T., et al. (2010). Changes and variations of Polycyclic Aromatic Hydrocarbon concentrations in fish, barnacles and crabs following an oil spill in a mangrove of Guanabara Bay, Southeast Brazil. Mar. Pollut. Bull. 60, 1359–1363. doi: 10.1016/j.marpolbul.2010.05.013
Solari, A., Garcia, M. L., and Jaureguizar, J. A. (2010). First record of Eucinostomus melanopterus, Bleeker 1863 (Perciformes, Gerreidae) from the Río de la Plata estuary. J. Appl. Ichthyol. 26, 485–487. doi: 10.1111/j.1439-0426.2009.01323.x
Souza, F. M., Brauko, K. M., Gilbert, E. R., Martins, C. C., Lana, P. C., and Camargo, M. G. (2016). Complex spatial and temporal variation of subtropical benthic macrofauna under sewage impact. Mar. Environ. Res. 116, 61–70. doi: 10.1016/j.marenvres.2016.02.008
Souza-Bastos, L. R., and Freire, C. A. (2011). Osmoregulation of the resident estuarine fish Atherinella brasiliensis was still affected by an oil spill (Vicuña tanker, Paranaguá Bay, Brazil), 7months after the accident. Sci. Total Environ. 409, 1229–1234. doi: 10.1016/j.scitotenv.2010.08.035
Souza-Conceição, J. M., Costa, M. D. P., Spach, H. L., and Schwingel, P. R. (2013). Microdesmus longipinnis (Gobioidei, Microdesmidae): Ocorrência, abundância e amostragem em um estuário subtropical. Neotrop. Biol. Conserv. 8, 111–114. doi: 10.4013/nbc.2013.82.07
Speranza, E. D., Colombo, M., Tatone, L. M., Cappelletti, N., Migoya, M. C., and Colombo, J. C. (2016). Fatty acid alterations in the detritivorous Prochilodus lineatus promoted by opportunistic feeding on sewage discharges in the Río de la Plata estuary. J. Fish Biol. 89, 2024–2037. doi: 10.1111/jfb.13104
Spetter, C. V., Buzzi, N. S., Fernández, E. M., Cuadrado, D. G., and Marcovecchio, J. E. (2015). Assessment of the physicochemical conditions sediments in a polluted tidal flat colonized by microbial mats in Bahía Blanca Estuary (Argentina). Mar. Pollut. Bull. 91, 491–505. doi: 10.1016/j.marpolbul.2014.10.008
Stehlik, L., and Meise, C. (2000). Diet of winter flounder in a New Jersey estuary: ontogenetic change and spatial variation. Estuaries 23, 381–391. doi: 10.2307/1353330
Storm, C., van der Velden, J. A., and Kuijpers, J. W. M. (2005). From nature conservation towards restoration of estuarine dynamics in the heavily modified Rhine-Meuse estuary, The Netherlands. Arch. Hydrobiol. 155, 305–318. doi: 10.1127/lr/15/2003/305
Streitenberger, M. E., and Baldini, M. D. (2016). Aporte de los afluentes a la contaminación faecal del estuario de Bahía Blanca, Argentina. Rev. Int. Contam. Ambient. 32, 243–248. doi: 10.20937/RICA.2016.32.02.10
Suárez, A., and Rubio, E. (1992a). Aspectos Sobre Crecimiento y Ciclo Sexual de Lutjanus guttatus (Pisces: Lutjanidae) en Bahía Málaga. Pacífico colombiano. Comisión Colombiana de Oceanografía, memorias VIII Seminario Nacional de las Ciencias y las Tecnologías del Mar. 924–939.
Suárez, A., and Rubio, E. (1992b). Hábitos Alimenticios de los Juveniles de Lutjanus guttatus (Pisces: Lutjanidae) en Bahía Málaga. Pacífico colombiano. Comisión Colombiana de Oceanografía, memorias VIII Seminario Nacional de las Ciencias y las Tecnologías del Mar. 913–923.
Tagliani, P. R. A., Asmus, M. L., Tagliani, C. R. A., Polette, M., Costa, C. S. B., and Sala, E. (2007). Integrated coastal zone management in the Patos Lagoon Estuary (South Brazil): state of art. WIT Trans. Ecol. Environ. 103, 679–686. doi: 10.2495/WRM070631
Tatone, L. M., Bilos, C., Skorupka, C. N., and Colombo, J. C. (2013). Trace metals in settling particles from the sewage impacted buenos aires coastal area in the Río de la Plata Estuary, Argentina. Bull. Environ. Contam. Toxicol. 90, 318–322. doi: 10.1007/s00128-012-0914-2
Tatone, L. M., Bilos, C., Skorupka, C. N., and Colombo, J. C. (2015). Trace metal behavior along fluvio-marine gradients in the Samborombón Bay, outer Río de la Plata estuary, Argentina. Cont. Shelf Res. 96, 27–33. doi: 10.1016/j.csr.2015.01.007
Taylor, M. D., Stewart Fielder, D., and Suthers, I. M. (2006). Spatial and ontogenetic variation in the diet of wild and stocked mulloway (Argyrosomus japonicus, Sciaenidae) in Australian Estuaries. Estuaries Coast. 29, 785–793. doi: 10.1007/bf02786529
Tellechea, J. S., Bouvier, D., and Norbis, W. (2011). Spawning sounds in whitemouth croaker (Sciaenidae): seasonal and daily cycles. Bioacoustics 20, 159–168. doi: 10.1080/09524622.2011.9753641
Tibiriçá, C. E. J. A., Fernandes, L. F., and Mafra, L. L. Jr. (2015). Seasonal and spatial patterns of toxigenic species of Dinophysis and Pseudo-nitzschia in a subtropical brazilian estuary. Br. J. Oceanogr. 63, 17–32. doi: 10.1590/S1679-87592015071906301
Tomás, A. R. G., Tutui, S. L. S., Fagundes, L., and Souza, M. R. (2012). Opsanus beta: an invasive fish species in the Santos Estuary, Brazil. Bol. Inst. Pesca 38, 349–355.
Tombesi, N., Pozo, K., Álvarez, M., Pribylová, P., Kukučka, P., Audy, O., et al. (2017). Tracking Polychlorinated Biphenyls (PCBs) and Polybrominated Diphenyl Ethers (PBDEs) in sediments and soils from the southwest of Buenos Aires Province, Argentina (South eastern part of the GRULAC region). Sci. Total Environ. 575, 1470–1476. doi: 10.1016/j.scitotenv.2016.10.013
Twilley, R. R., Gottfried, R. R., Rivera-Monroy, V. H., Zhang, W., Armijos, M. M., and Bodero, A. (1998). An approach and preliminary model of integrating ecological and economic constraints of environmental quality in the Guayas River estuary, Ecuador. Environ. Sci. Policy 1, 271–288. doi: 10.1016/S1462-9011(98)00012-4
Underwood, A. J., Chapman, M. G., and Browne, M. A. (2017). Some problems and practicalities in design and interpretation of samples of microplastic waste. Anal. Methods 9, 1332–1345. doi: 10.1039/C6AY02641A
Valdés, M. E., Marino, D. J., Wunderlin, D. A., Somoza, G. M., Ronco, A. E., and Carriquiriborde, P. (2015). Screening concentration of E1, E2 and EE2 in sewage effluents and surface waters of the “Pampas” region and the “Río de la Plata” estuary (Argentina). Bull. Environ. Contam. Toxicol. 94, 29–33. doi: 10.1007/s00128-014-1417-0
Valiñas, M. S., Molina, L. M., Addino, M., Montemayor, D. I., Acha, E. M., and Iribarne, O. O. (2012). Biotic and environmental factors affect Southwest Atlantic saltmarsh use by juvenile fishes. J. Sea Res. 68, 49–56. doi: 10.1016/j.seares.2011.12.001
Vargas, C. A., Araneda, S. E., and Valenzuela, G. (2003). Influence of tidal phase and circulation on larval fish distribution in a partially mixed estudary, Corral Bay, Chile. J. Mar. Biol. Assoc. 83, 217–222. doi: 10.1017/S0025315403006994h
Vasconcellos, R. M., Araújo, F. G., Santos, J. N. S., and Silva, M. A. (2010). Short-term dynamics in fish assemblage structure on a sheltered sandy beach in Guanabara Bay, Southeastern Brazil. Mar. Ecol. 31, 506–519. doi: 10.1111/j.1439-0485.2010.00375.x
Vasconcelos Filho, A. L., Neumann-Leitão, S., Eskinazi-Leça, E., and Oliveira, A. M. E. (2010). Hábitos alimentares de peixes consumidores secundários do Canal de Santa Cruz, Pernambuco, Brasil. Trop. Oceanogr. Online 38, 121–128. doi: 10.5914/tropocean.v38i2.5166
Vasconcelos Filho, A. L., Neumann-Leitão, S., Eskinazi-Leça, E., Schwamborn, R., Oliveira, A. M. E., and Paranaguá, M. N. (2003). Trophic interactions between fish and other compartment communities in a tropical estuary in Brazil as indicator of environmental quality. Adv. Ecol. Sci. 18, 173–183. doi: 10.2495/ECO030161
Velasco, G., Reis, E. G., and Vieira, J. P. (2007). Calculating growth parameters of Genidens barbus (Siluriformes, Ariidae) using length composition and age data. J. Appl. Ichthyol. 23, 64–69. doi: 10.1111/j.1439-0426.2006.00793.x
Vendel, A. L., Bessa, F., Alves, V. E. N., Amorim, A. L. A., Patrício, J., and Palma, A. R. T. (2017). Widespread microplastic ingestion by fish assemblages in tropical estuaries subjected to anthropogenic pressures. Mar. Pollut. Bull. 117, 448–455. doi: 10.1016/j.marpolbul.2017.01.081
Vendel, A. L., Bouchereau, J.-L., and Chaves, P. T. (2010). Environmental and subtidal fish assemblage relationships in two different Brazilian coastal estuaries. Br. Arch. Biol. Technol. 53, 1393–1406. doi: 10.1590/S1516-89132010000600016
Venekey, V., and de Melo, T. P. G. (2016). Nematodes as indicators of shrimp farm impact on an amazonian estuary (Curuçá, Pará, Brazil). Br. J. Oceanogr. 64, 75–88. doi: 10.1590/S1679-87592016108206401
Venturini, N., Bícego, M. C., Taniguchi, S., Sasaki, S. T., García-Rodríguez, F., Brugnoli, E., et al. (2015). A multi-molecular marker assessment of organic pollution in shore sediments from the Río de la Plata Estuary, SW Atlantic. Mar. Pollut. Bull. 91, 461–475. doi: 10.1016/j.marpolbul.2014.06.056
Vergara, J., Azpelicueta, M. D. L. M., and Garcia, G. (2008). Phylogeography of the Neotropical catfish Pimelodus albicans (Siluriformes: Pimelodidae) from Río de La Plata basin, South America, and conservation remarks. Neotrop. Ichthyol. 6, 75–85. doi: 10.1590/S1679-62252008000100009
Vetemaa, M., Eschbaum, R., Verliin, A., Albert, A., Eero, M., Lillemägi, R., et al. (2006). Annual and seasonal dynamics of fish in the brackish-water Matsalu Bay, Estonia. Ecol. Freshw. Fish 15, 211–220. doi: 10.1111/j.1600-0633.2006.00134.x
Viana, A., and Lucena Frédou, F. (2014). Ichthyofauna as bioindicator of environmental quality in an industrial district in the amazon estuary, Brazil. Br. J. Biol. 74, 315–324. doi: 10.1590/1519-6984.16012
Vilar, C. C., Spach, H. L., and Souza-Conceição, J. M. (2011). Fish assemblage in shallow areas of Baía da Babitonga, Southern Brazil: structure, spatial and temporal patterns. Panam. J. Aquat. Sci. 6, 303–319.
Villar, C., Stripeikis, J., Colautti, D., D'Huicque, L., Tudino, M., and Bonetto, C. (2001). Metals contents in two fishes of different feeding behaviour in the Lower Parana River and Rio de la Plata Estuary. Hydrobiologia 457, 225–233. doi: 10.1023/a:1012285820526
Wang, L., Min, M., Li, Y., Chen, P., Chen, Y., Liu, Y., et al. (2010). Cultivation of green algae Chlorella sp. in different wastewaters from municipal wastewater treatment plant. Appl. Biochem. Biotech. 162, 1174–1186. doi: 10.1007/s12010-009-8866-7
Wasserman, J. C., Freitas-Pinto, A. A. P., and Amouroux, D. (2000). Mercury concentrations in sediment profiles of a degraded tropical coastal environment. Environ. Technol. 21, 297–305. doi: 10.1080/09593332108618117
Wasserman, J. C., Wasserman, M. A. V., Barrocas, P. R. G., and Almeida, A. M. (2016). Predicting pollutant concentrations in the water column during dredging operations: implications for sediment quality criteria. Mar. Pollut. Bull. 108, 24–32. doi: 10.1016/j.marpolbul.2016.05.005
Watanabe, K., Kasai, A., Antonio, E. S., Suzuki, K., Ueno, M., and Yamashita, Y. (2014). Influence of salt-wedge intrusion on ecological processes at lower trophic levels in the Yura Estuary, Japan. Estuar. Coast. Shelf Sci. 139, 67–77. doi: 10.1016/j.ecss.2013.12.018
Watanabe, L. A., Vallinoto, M., Neto, N. A., Muriel-Cunha, J., Saint-Paul, U., Schneider, H., et al. (2014). The past and present of an estuarine-resident fish, the “four-eyed fish” Anableps anableps (Cyprinodontiformes, Anablepidae), revealed by mtDNA sequences. PLoS ONE 9:e101727. doi: 10.1371/journal.pone.0101727
Wilber, D. H., and Clarke, D. G. (2001). Biological effects of suspended sediments: a review of suspended sediment impacts on fish and shellfish with relation to dredging activities in estuaries. North Am. J. Fish. Manag. 21, 855–875. doi: 10.1577/1548-8675(2001)0212.0.CO
Williams, J., and Crutzen, P. J. (2013). Perspectives on our planet in the Anthropocene. Environ. Chem. 10, 269–280. doi: 10.1071/EN13061
Windom, H. L., Niencheski, L. F., and Smith, R. G. (1999). Biogeochemistry of nutrients and trace metals in the estuarine region of the Patos Lagoon (Brazil). Estuar. Coast. Shelf Sci. 48, 113–123. doi: 10.1006/ecss.1998.0410
Zhengyun, Z., Zhixian, S., Qiaoying, Z., and Aiying, S. (2003). The current status of world protection for mangrove forest. Chin. J. Oceanol. Limnol. 21, 261–269. doi: 10.1007/BF02842842
Keywords: estuarine ecocline, estuarine conservation, human modification, contamination of aquatic habitats, fish ecology, environmental quality
Citation: Barletta M and Lima ARA (2019) Systematic Review of Fish Ecology and Anthropogenic Impacts in South American Estuaries: Setting Priorities for Ecosystem Conservation. Front. Mar. Sci. 6:237. doi: 10.3389/fmars.2019.00237
Received: 14 January 2019; Accepted: 16 April 2019;
Published: 09 May 2019.
Edited by:
Christos Dimitrios Arvanitidis, Hellenic Centre for Marine Research (HCMR), GreeceReviewed by:
Alan Whitfield, South African Institute for Aquatic Biodiversity, South AfricaJosé Lino Vieira De Oliveira Costa, Universidade de Lisbon, Portugal
Copyright © 2019 Barletta and Lima. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mário Barletta, YmFybGV0dGFAdWZwZS5icg==