 Charlotte Berkström
Charlotte Berkström Myron Papadopoulos
Myron Papadopoulos Narriman Saleh Jiddawi3
Narriman Saleh Jiddawi3 Lina Mtwana Nordlund
Lina Mtwana Nordlund
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 22 March 2019
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 6 - 2019 | https://doi.org/10.3389/fmars.2019.00130
This article is part of the Research TopicSmall-Scale and Artisanal Fisheries: Insights and Approaches for Improved Governance and Management in a Globalized ContextView all 13 articles
In developing countries where data and resources are lacking, the practical relevance of local ecological knowledge (LEK) to expand our understanding of the environment, has been highlighted. The potential roles of the LEK varies from direct applications such as gathering environmental information to a more participative involvement of the community in the management of resources they depend on. Fishers’ LEK could therefore be useful in order to obtain information on how to advance management of coastal fisheries. Many targeted fish species migrate between habitats to feed, spawn or recruit, connecting important habitats within the seascape. LEK could help provide answers to questions related to this connectivity and the identification of fish habitat use, and migrations for species and areas where such knowledge is scarce. Here we assess fishers’ LEK on connectivity between multiple habitats within a tropical seascape, investigate the differences in LEK among fisher groups and the coherence between LEK and conventional scientific knowledge (CSK). The study was conducted in 2017 in Zanzibar, Tanzania, a tropical developing country. One hundred and thirty-five semi-structured interviews were conducted in six different locations focusing on fish migrations, and matching photos of fish and habitats. Differences between fisher groups were found, where fishers traveling further, exposed to multiple habitats, and who fish with multiple gears had a greater knowledge of connectivity patterns within the seascape than those that fish locally, in single habitats and with just one type of gear. A high degree of overlap in LEK and CSK was found, highlighting the potential benefits of a collaboration between scientists and fishers, and the use of LEK as complementary information in the management of small-scale fisheries.
Small-scale fisheries are critically important for the provision of food security and sustained livelihoods, especially in developing tropical countries (FAO, 2012; Unsworth et al., 2018b). However, many marine coastal systems are intensely and synergistically affected by human activities and fish stocks have declined globally at an alarming rate, calling for management actions (Pauly and Zeller, 2016). Many fisheries appear to be failing in achieving yields or conservation goals where healthier oceans supporting more fish, feeding more people, and improving livelihoods are prioritized (Karr et al., 2017; Unsworth et al., 2018a).
Within the marine conservation community there is considerable interest in combining local and scientific knowledge to achieve management objectives. However, few studies have examined the merits and caveats of local ecological knowledge (LEK) or have shown how combining both knowledge systems would result in better management outcomes (Hamilton et al., 2012). In developing countries, where data and resources often are lacking, authors have highlighted the practical relevance of LEK in order to obtain useful information (Taylor et al., 2011; Silvano and Begossi, 2012; Thornton and Scheer, 2012). Since biologists do not always have the means or funds of gathering knowledge on ecological systems directly, the classical approach to management of natural resources, which is solely based on scientific knowledge, is destined to be unsuccessful (Davis and Ruddle, 2010). Although rare, examples suggest that the inclusion of LEK and the involvement of local fishers, increases the chances of success (Ruddle, 1995; Shephard et al., 2007; Nenadovic et al., 2012). Conventional scientific knowledge (CSK) is gained from data collected according to a scientific design and theoretically interpreted (Mackinson, 2001; Gaspare et al., 2015). LEK, on the other hand, is accumulated over one’s lifetime from observations and hands-on experience in interacting with ecological systems and utilizing natural resources for one’s livelihood (Olsson and Folke, 2001). Another aspect of LEK, which can also be denoted as indigenous or traditional ecological knowledge (IEK or TEK), is that it is also a cumulative body of knowledge that transcends generations, through cultural transmission and can often be associated with elders within the local community (Berkes et al., 2000; Johannes et al., 2000; García-Quijano, 2007; Davis and Ruddle, 2010). Fishers can provide novel information on the biology and ecology of species and help answer questions related to the identification of fish habitat use, nursery areas and migrations of species where such knowledge is scarce (Begossi et al., 2016). Le Fur et al. (2011) demonstrated that fishers in West Africa were able, collectively, to develop maps of nursery locations including specific details for each estuary. Moreover, fishers identified periods during which mature adults migrated toward spawning grounds and periods of juvenile recruitment. This information is crucial in fisheries management and can also be used in the establishment of marine protected areas (MPAs), particularly to determine the location and size of protection to maximize conservation, biodiversity, and fishery benefits. LEK was also compared with scientifically gathered data showing that the two data sets were similar (Le Fur et al., 2011), highlighting collaboration between scientists and fishermen and the use of LEK as complementary information.
Tropical seascapes are comprised of a mosaic of habitats including mangroves, seagrass meadows, macroalgal beds and coral reefs (Ogden, 1988). Many coral reef fishes, targeted by the local fishers, migrate to seagrass and mangrove areas to feed during dusk or dawn or during tidal fluctuations (Dorenbosch et al., 2004; Figure 1a; Unsworth et al., 2007). Similarly, many fishes utilize these adjacent habitats as nursery areas before migrating to coral reefs as adults (Berkström et al., 2013a; Figure 1b). These migrations transfer nutrients and energy between the ecosystems within the seascape and contribute to a shifting biomass that accumulates within the organisms throughout their different life stages (Berkström et al., 2012; Hyndes et al., 2014). Several species also undergo reproductive migrations, gathering in large schools in spawning areas (Claydon, 2004). The connectivity between different habitats where the species cover their full life cycle is important for the replenishment of fish stocks and the provisioning of ecosystem services vital to local human populations. Research on seascape connectivity suggests that connectivity can effectively increase the resilience of marine ecosystem functions and services (Mumby, 2006; Olds et al., 2013) and has recently been highlighted as important in the management of aquatic resources (Berkström et al., 2012; Nagelkerken et al., 2015; Sheaves et al., 2015; Olds et al., 2016). Although the tropical seascape supports a high biomass of fish in total, species-specific biomass is relatively low, causing artisanal fisheries to target several fish species by using many types of gears (Garcia-Quijano, 2015). Tropical fishers have thus adapted to this by incorporating different fishing methods across local habitats in order to try and maintain high levels of yields. Also, with fish stocks depleting, fishers have to move further to exploit more productive fishing grounds (García-Quijano, 2007). Since LEK is acquired by an individual’s hands-on-experience and observations of the environment in which they work, heterogeneity of ecological knowledge between fishers can arise between different groups of fishers (Crona, 2006; Crona and Bodin, 2006). Furthermore, Davis and Wagner (2003) highlighted the importance of identifying “experts” when researching LEK, in order to be able to use the most reliable and comprehensive LEK in fisheries management. The present study therefore sets out to distinguish whether there are differences in LEK between different groups of fishers that: (i) utilize single and multiple habitats, (ii) fish locally (within 5 km of their village) or distantly (>5 km away from their village), (iii) use different types of fishing gears, and (iv) fish in ancestral fishing grounds or not. Furthermore, LEK is compared with CSK on connectivity from the same area. It is hypothesized that fishers utilizing multiple habitats, move to fish, use multiple gears and fish in ancestral fishing grounds will have more comprehensive LEK than those that fish in single habitats, fish locally, use single gears and fish in non-ancestral fishing grounds.
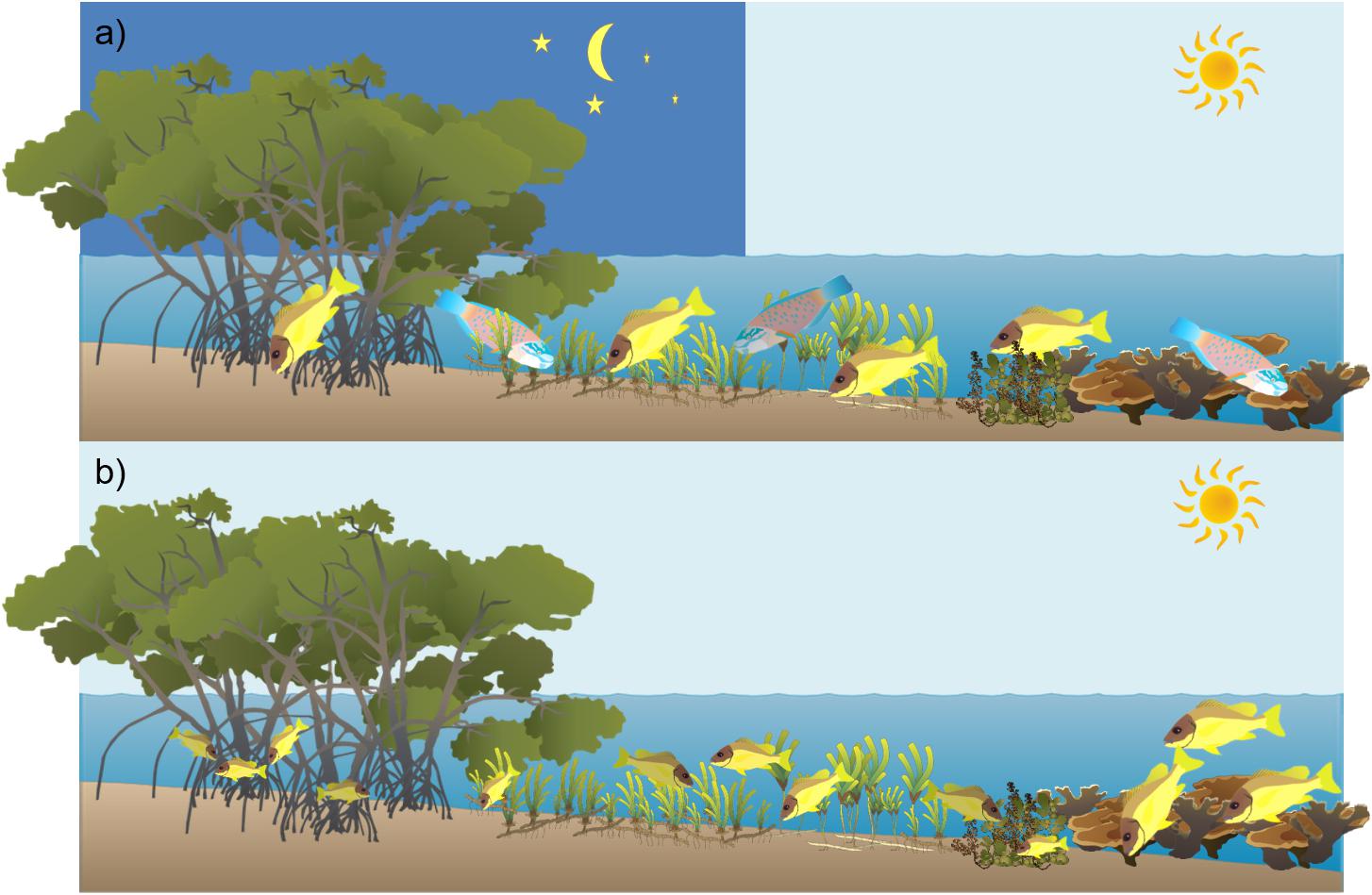
Figure 1. Schematic illustration adopted from Berkström (2012) showing (a) dial and tidal foraging migrations between coral reef, macroalgae, seagrass, and mangrove habitats and (b) ontogenetic migration of juvenile coral reef fish between the above-mentioned habitats within tropical seascapes. Image symbol courtesy of the Integration and Application Network, University of Maryland and Stina Tano.
The study was conducted on Unguja Island within the Zanzibar archipelago, Tanzania, off the coast of East Africa. It is the main island of the archipelago and is most commonly referred to as Zanzibar. Zanzibar is surrounded by rich marine resources from the Western Indian Ocean (WIO), where small-scale artisanal fishing and tourism take place. The fishery applies a variety of fishing techniques targeting a large number of species (Jiddawi and Öhman, 2002). The tropical seascape around Zanzibar is comprised of multiple habitats including mangrove forests, seagrass meadows, macroalgal beds, and coral reefs (Berkström et al., 2012; Khamis et al., 2017). It experiences large tidal fluctuations of up to 4m and is subjected to the northeast (kaskazi) and the southeast (kusi) monsoon seasons (McClanahan, 1988). The study was conducted in six locations: two sites located in the north-west part of the island, two sites in Menai Bay, the south-west part of the island and two sites on the eastern side of the island (Figure 2).
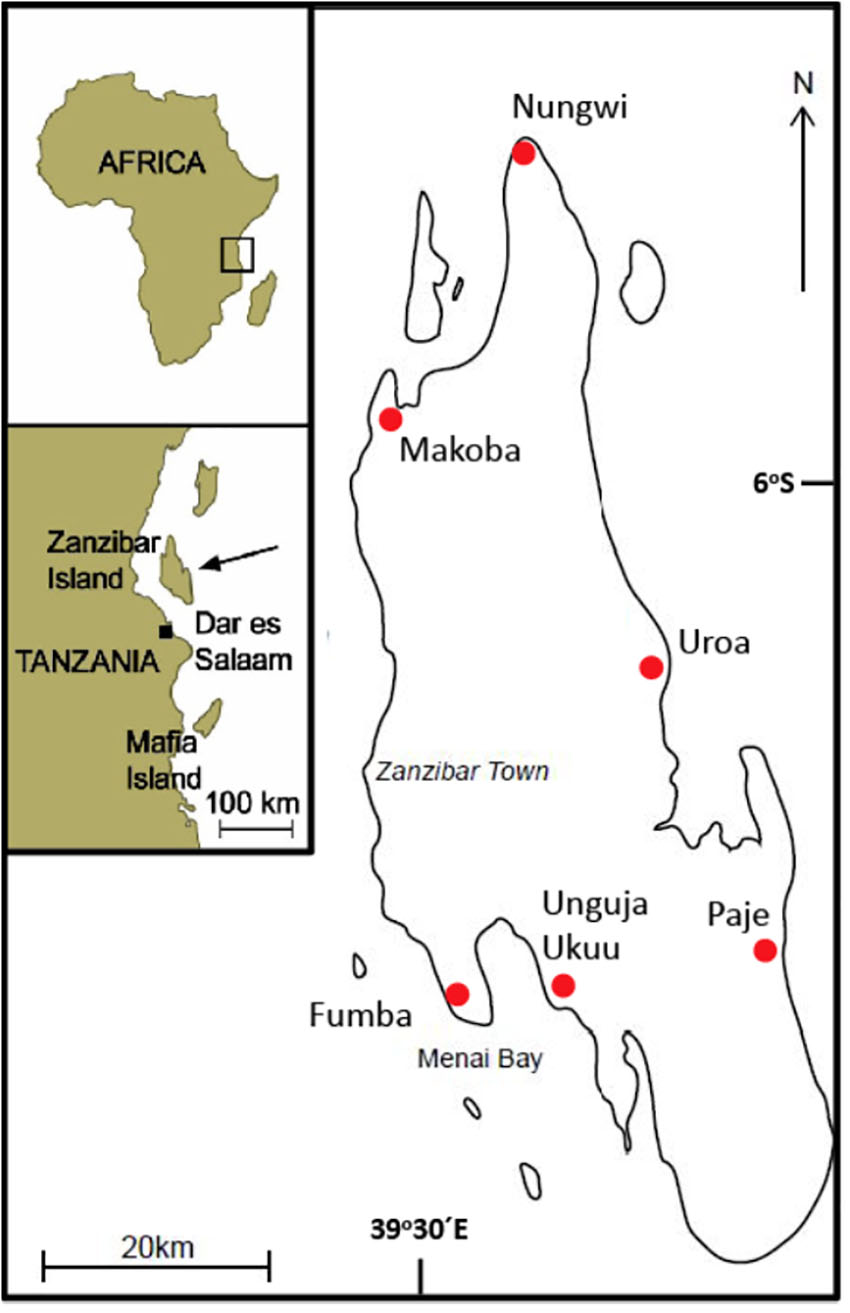
Figure 2. Map of Zanzibar off the east coast of Tanzania. The locations of the six sites, where interviews were conducted, are indicated with red markers.
Data on LEK was collected through semi-structured interviews with local fishers between September and November 2017. The interviews were conducted in Uroa, Ungunja Ukuu, Paje, Fumba, Nungwi, and Makoba (Figure 2). Fumba and Unguja Ukuu were chosen because both these locations are situated in Menai Bay, where scientific information on connectivity has previously been collected (Berkström et al., 2012; Berkström et al., 2013b; Tano et al., 2017). These areas were also chosen because they are comprised of multiple habitats (mangroves, seagrass, macroalgae, and coral reef).
A questionnaire was used for gathering information from local fishers. Interviews were conducted in Kiswahili via an interpreter and after conducting interviews in each village the answers were translated to English by the same interpreter. For each site, a beach recorder was used to find fishers willing to be interviewed. A minimum of 20 interviews were performed at each site. First, questions were asked to gather the demographics of the respondents. Second, questions were asked to gather data on LEK about habitat use and connectivity of selected species of fish. Three general questions regarding different types of fish migrations between habitats (diurnal/feeding, spawning, and ontogenetic) were asked. This section also contained pictures of fish species (juveniles and adults) and different habitats for the respondent to match the fish species to the habitats in which they are found. An array of fish species was included that either use single or multiple habitats. Toward the end, an open dialogue was held to better understand the level of ecological knowledge that the respondent possessed. Lastly, respondents were asked how they gained their knowledge that they demonstrated in the interview.
There were four variables of interest; type of fisher (local or distant), habitat usage (single or multiple), ancestry (if forefathers fished in the area), and gear usage. Based on the distance they moved to fishing grounds, fishers were classified as either local (<5 km), or distant (>5 km). Ancestry described if the respondents have been fishing in an area for generations or are new to the area. Gear usage was divided into five categories: multiple gears and the individual single gears dema traps, handlines, nets, and spears/sticks. Fishers that used either drag nets, seine nets, gill nets or mosquito nets or a combination of nets were classified under the general term “nets.” Fishers were also classified as either using a single habitat to fish in or multiple habitats to fish in. Fishers who said that they used multiple habitats, but where the second habitat was “open ocean” were changed to single habitat users.
Connectivity knowledge was assessed by asking three questions regarding diurnal/feeding, ontogenetic and spawning migrations. Respondents were asked if they knew of fish that move between habitats to feed, spawn or live in as juveniles, and were also asked to give examples. The more “yes” answers to the three questions represented a higher knowledge on connectivity and were scored (0–3). A score of “0” represented that all three questions were answered with a “no.”
Ecological data on habitat use and connectivity by fish in Menai Bay (Berkström et al., 2012, 2013b; Tano et al., 2017) was used to compare CSK data with LEK data by local fishers. Four habitats within the tropical seascape were in focus: (1) coral reefs, (2) seagrass meadows, (3) macroagal beds, and (4) mangroves. However, it was challenging to be certain that the local fishers were distinguishing between seagrass and macroalgae, therefore the two habitats were combined and referred to as submerged aquatic vegetation (SAV). Habitat scores were allocated to each fish species, which corresponded to the number of habitats used by each fish species. LEK habitat scores represented that of which the fishers were aware of and CSK habitat scores represented that of which the scientific community were aware of. Mean LEK habitat score was calculated for each fish species by averaging all the respondent’s answers for each fish species. The total number of fishers that mentioned that a particular fish species was present in one of the three habitats (coral, SAV, and mangrove) was also recorded. If more than 25% of fishers stated that a particular fish species was seen in a habitat, then that fish species was deemed to occur there. The fish species might occur in that habitat if 10–25% of fishers stated that they do. If less than 10% of fishers stated that they do occur, they were deemed not to occur there. In order to verify that the LEK data for habitat score can be counted on, an index of inaccuracy was created (Supplementary Figure S1).
Difference in LEK scores between type of fisher, fisher’s habitat usage and gear choice were analyzed with permutational multivariate analysis of variance (PERMANOVA). The assumptions of normality were not met so data was fourth root transformed. The PERMANOVA was performed on unbalanced data, although PERMANOVAs are robust in dealing with unbalanced data (Anderson, 2001). However, to make sure that differences found were not due to unbalanced data, data points were randomly taken out by using the “RANDBETWEEN (1;135)” function in Excel to get equal data sets for the different groups of fishers. PERMANOVA tests were rerun with the reduced, equal sample sizes. The results were similar, confirming that all of the data could be used in the analysis. The PERMANOVA test was performed using 999 permutations under a reduced model. A non-metric multi-dimensional scaling (nMDS) ordination with Euclidean dissimilarity index was performed in order to see patterns in the multivariable data. A Wilcoxon signed-rank test with continuity correction was used to compare the differences between the mean habitat scores for the different knowledge sources (CSK and LEK) and the different subcategories of LEK (migratory and local fishers, multiple, and single habitat users).
In total, 135 fishers were interviewed. The respondents were all male and between the ages of 17 and 75 years. On average, there were more respondents in the age class 25–34 years of age. Eighty-four percent of respondents had a formal educational background, whether it was primary education (23%), secondary education (59%), or tertiary education (2%). Most of the respondents had children (74%) and out of those respondents; 1–3 children (41%), 4–6 children (31%), or 7+ children (28%). For fishing gear, handlines and nets were more commonly used by fishers, as well as combinations of different fishing gears. Out of the total number of respondents, there were more fishers that utilized multiple habitats (n = 101) than a single habitat (n = 21). There were also more fishers that fished in non-local fishing grounds (i.e., distant fishers, n = 72) than fishers that fished locally (n = 50). Respondents’ knowledge of their environment was gained mainly through: hands-on experience (63%), experienced and shared knowledge (29%), and fishing seminars and formal education (8%) (Figure 3).
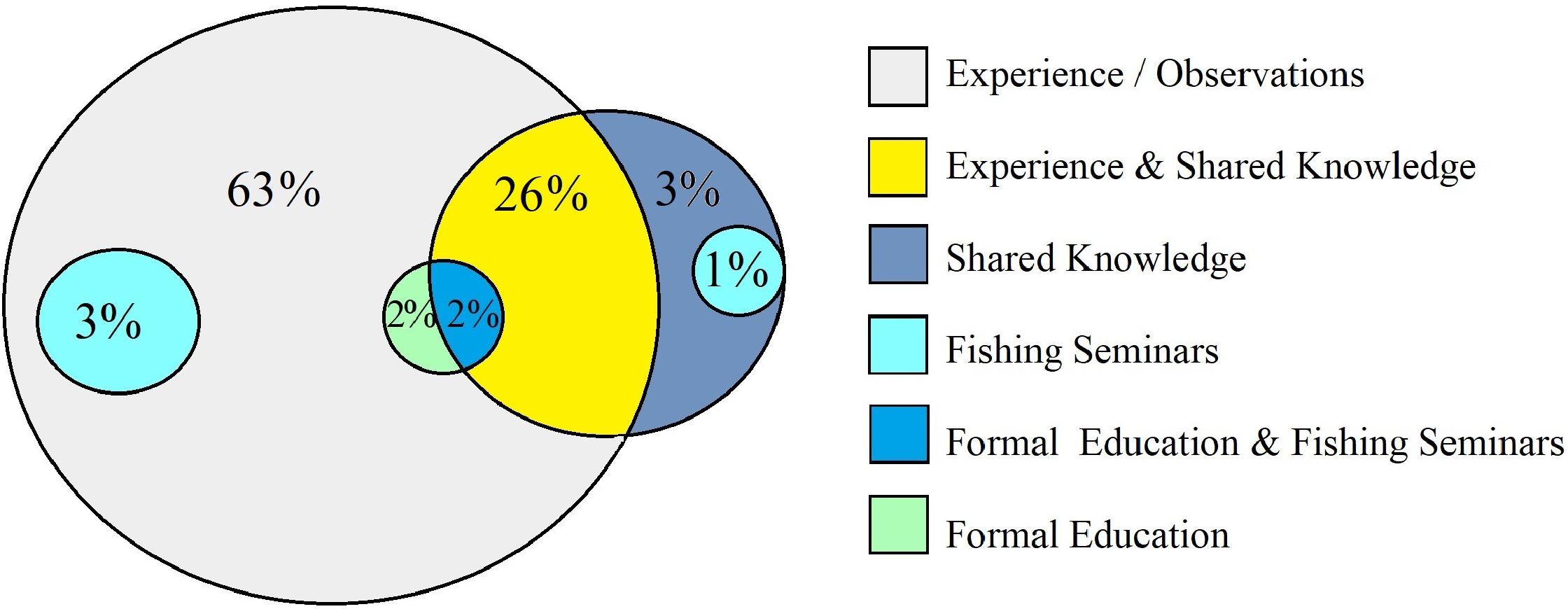
Figure 3. Venn diagram showing the methods of knowledge acquisition on fish ecology/biology by fishers.
There were differences between fishers with regards to type of fisher (distant/local), habitat usage (single/multiple), and gear usage. More than half of the respondents received the highest LEK score that can be allocated. There were significant differences in LEK scores between multiple and single habitat users [F(1,39) = 14.427; p = 0.005] and between fishers using different gears [F(4,39) = 3.679; p = 0.027, Table 1]. On average, fishers using single habitats had higher LEK scores than those fishing in multiple habitats. Dema trap fishers had the lowest LEK scores, whereas the other fishing gear users had similar LEK scores. Local fishers generally had higher LEK scores compared to distant fishers, however there was no significant difference found between the two types of fishers [F(1,39) = 3.777; p = 0.061, Table 1]. There was no significant difference found for ancestry [F(1,39) = 0.609; p = 0.64] and no interactions were found between the different variables (Table 1). On average, fishers knew less about spawning migrations compared to diurnal and ontogenetic migrations. A similar trend was seen across the different types of fishers, habitat usage, ancestry and gear usage.
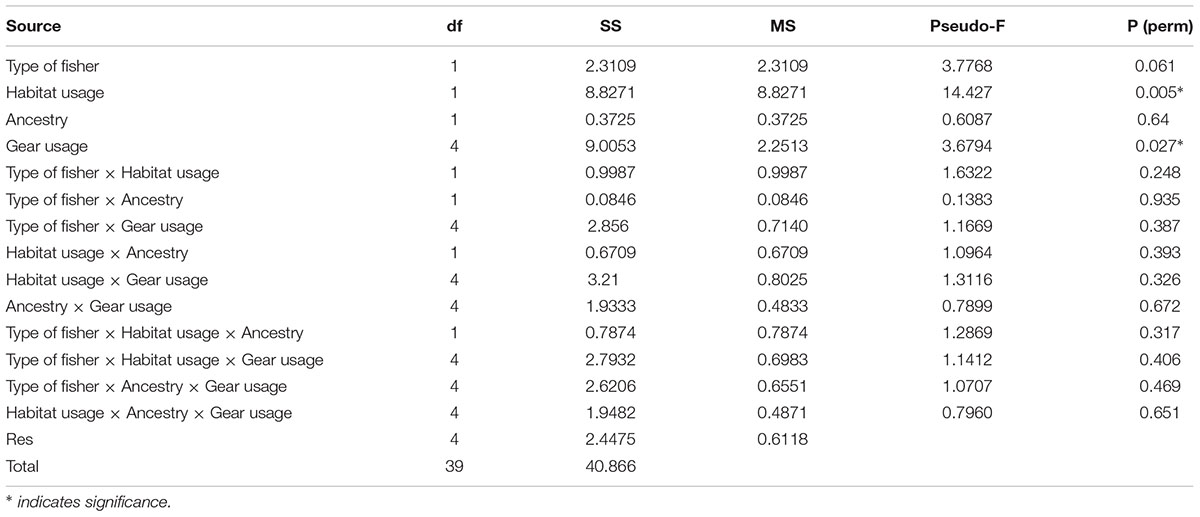
Table 1. A PERMANOVA table based on Euclidean dissimilarity for LEK data between different groups of fishers in Zanzibar, Tanzania.
There were significant differences found between CSK and LEK (Table 2). On average, CSK had higher habitat scores than LEK (Supplementary Table S1). There was a significant difference between CSK and LEK in multiple, single, local, and distant fishers (Table 2). It was also found that distant fishers had higher scores than local fishers and that multiple habitat users had higher scores than single habitat users (Table 2).
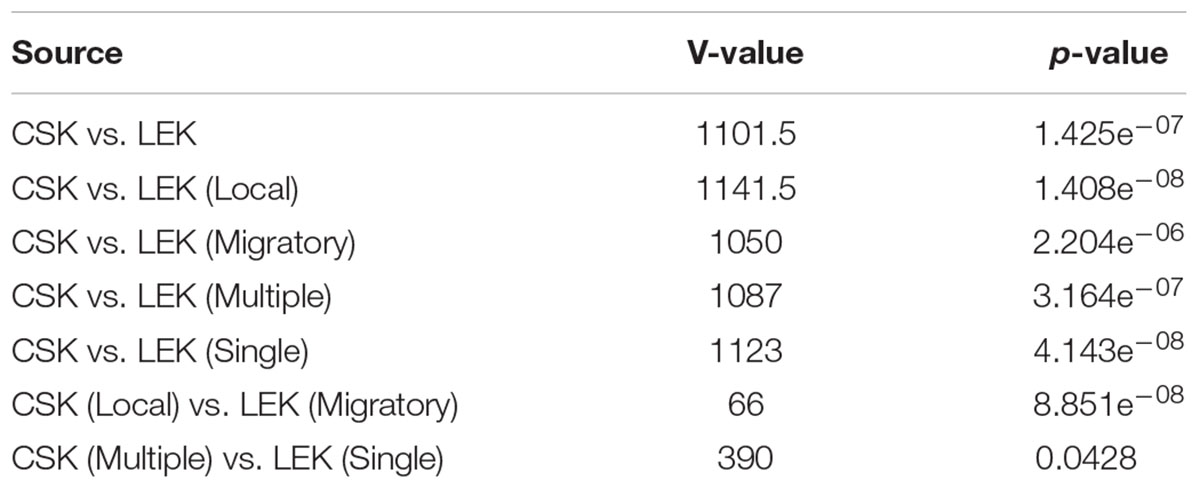
Table 2. A table showing results from a Wilcoxon signed-rank test with continuity correction comparing mean habitat scores between local ecological knowledge (LEK) and conventional science knowledge (CSK).
Local ecological knowledge and Conventional scientific knowledge corresponded with each other regarding the fish species that utilized coral reef habitats (Figure 4). LEK stated that SAV habitats were utilized by all the fish species that were shown, which did not correspond to CSK for some of the fish species. Most of the species that occur in mangroves, known by CSK, did not correspond with LEK (Figure 4). However, for the fish species Sphyraena flavicauda (barracuda) both CSK and LEK stated that it occurred in mangroves (Supplementary Table S1).
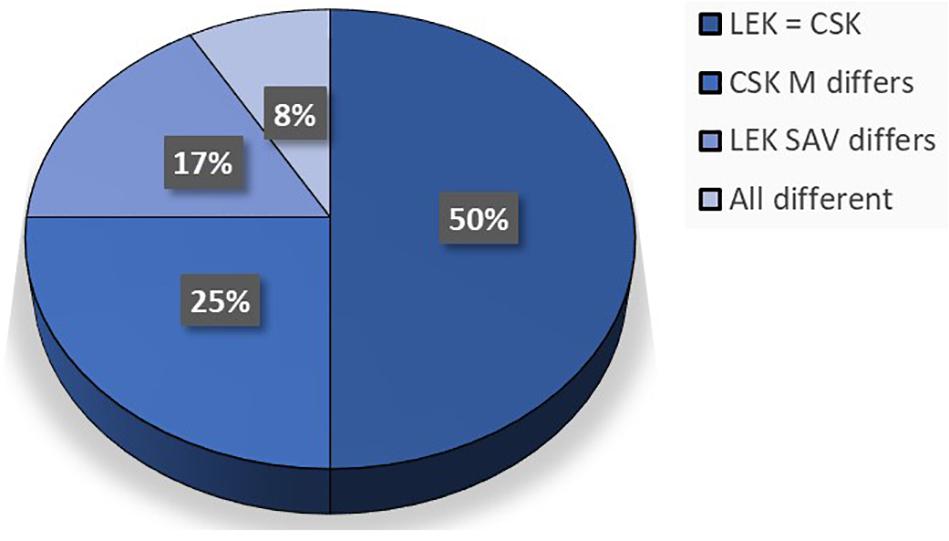
Figure 4. Pie chart depicting the relation between LEK and CSK regarding fish habitat use. The habitats that are used by different species of fish data obtained from scientific literature (CSK) (Berkström et al., 2012; Tano et al., 2017) compared with that obtained from interviewing local fishers (LEK) in Zanzibar, Tanzania. Fishers were shown pictures of different fish species and different habitats, and were asked to point to habitats that each individual fish species were found in. C, corals; SAV, submerged aquatic vegetation; M, mangroves. “CSK M” refers to scientific data regarding the mangrove habitat, “LEK SAV” refers to local fisher’s data regarding the SAV habitat. “All different” means that more than one LEK habitat did not match the CSK habitats.
A large majority of fishers demonstrated high knowledge of fish migrations between various habitats around Zanzibar. Knowledge on feeding and ontogenetic migrations were the highest while less was known about spawning migrations. At least half of the fishers had similar knowledge about connectivity as that of scientists. However, LEK on connectivity, differed between different groups of fishers. To our knowledge, this is one of the first studies to specifically assess fishers’ LEK on connectivity between multiple habitats within a tropical seascape. However, other studies have touched upon the topic of connectivity related to spawning migrations (Robinson et al., 2004; Silvano et al., 2006), fish habitat use (García-Quijano, 2007; Valdés-Pizzini and García-Quijano, 2009; Silvano et al., 2010; Gaspare et al., 2015), and comparison of habitat maps created by local fishers with satellite images (Aswani and Lauer, 2006b; Aswani and Vaccaro, 2008; Lauer and Aswani, 2008, 2010; Selgrath et al., 2016). The results from the present study may be highly valuable due to the lack of studies from Africa, with only 2–8% of the published articles on marine LEK from this region (Thornton and Scheer, 2012). The majority of studies have focused on North America and Oceania. Furthermore, tropical seascapes have a high diversity of target species for which the biological and ecological knowledge is limited (Silvano et al., 2006).
Overall, there were differences between all of the fisher groups except for those that fish in ancestral fishing grounds or not. When asked whether fish migrate to feed, spawn or during ontogeny, significant differences in LEK were found between fishers that fish in multiple and single habitats, and between fishers that use different types of gears. In contrast to what was hypothesized, single habitat users had higher LEK scores, meaning they knew more about fish migrations, than that of multiple habitat users. It was thought that multiple habitat users would have greater knowledge on seascape connectivity than single users, due to the fishers interacting with many different habitats within the seascape when fishing and at the same time developing localized knowledge based on those interactions, experiences, and observations. A possible reason for this result may be due to social intergroup dynamics, where ecological knowledge on fish migrations and fishers’ own experiences might be shared freely between the two different groups. Crona and Bodin (2006) found that fishers, despite fishing in different areas, had similar knowledge to each other due to frequent relations. However, at a more detailed level, multiple habitat users did have on average a greater knowledge of fish habitat usage than single habitat users when asked to match photos of individual fish species with photos of habitats. Even though the knowledge is shared between the two groups of fishers, the knowledge that is conveyed might consist of general behaviors of fish rather than detailed information on fish ecology, which may explain observed differences. A similar consensus can be seen in fishers that are grouped by fishing gears used. Fishers fishing with basket traps (dema) on average knew the least about fish migrations, which may be explained by the nature of how the fishing gear is utilized. Fishers can have a minimum of 5 basket traps and a maximum of 10 basket traps left at different sites in an area for long hours in order to increase catch probability (Jiddawi and Öhman, 2002). Since this method of fishing allows fishers to leave the trap at a particular time and then return to the trap after a few hours, fishers might not be very observant of fish behaviors during this period, unlike fishers that use gears that require the fisher to be present and vigilant at all times during the fishing period (e.g., fishers that use nets, handlines, spear/sticks and a combination of gears). Furthermore, handline and multiple gear fishers typically catch coral reef dwelling fishes that also utilize multiple habitats, e.g., fishes from the family Lutjanidae and Lethrinidae (Jiddawi and Öhman, 2002) and may therefore have a greater knowledge on the ecology of these families. A similar result was reported by Crona (2006), where groups of fishers were distinct from each other based on the type of gear that was utilized. Deep-sea fishers and seine-net fishers had the broadest concept on fish migrations, and in extension seine-net fishers also acknowledged the population dynamics of sea urchins, declining seagrass meadows and fish abundances (Crona, 2006).
When asked general questions about fish migrations, there were no significant differences found between fishers that fished locally and distantly, and those who fished in ancestral fishing grounds and those who did not. However, when asked to match photos of fish species with habitats in which they are found, significant differences were found between local, and distant fishers. As hypothesized, distant fishers knew more about connectivity, in terms of fish migrations, than local fishers on this more detailed level. This is in accordance with Crona and Bodin (2006), who also found that distant fishers were the most knowledgeable. This may be explained by distant fishers moving from one seascape to another, acquiring information on more fish species with varying habitat requirements compared to fishers that are more restricted to areas adjacent to their villages (Jiddawi and Öhman, 2002). Their experience of interacting with these multiple seascapes in different areas and with the fish species could add to their knowledge on fish ecology and on connectivity in general. On the other hand, this may differ in different seascape settings depending on which habitats are available within the local fishing grounds. If all habitats are present within the local fishing grounds, local fishers may be expected to acquire a deeper knowledge on connectivity than distant fishers since they spend more fishing time within their local seascape and less time traveling.
Tropical fish migrations occur on a daily, seasonal or annual basis and within an individual fisher’s lifetime (Berkström et al., 2012). Hence, the result of having no difference found between fishers that fished in ancestral fishing grounds and those who did not may be due to fishers observing the different types of migrations (diurnal/feeding, spawning, and ontogenetic) that fish undergo over their lifetime and do not necessarily depend on ancestral knowledge to know whether fish migrate or not.
Since conventional science is currently the presiding epistemological knowledge system that is widely used in resource management plans (Davis and Ruddle, 2010), LEK on connectivity was compared with CSK from the same area. CSK was collected in Zanzibar by scientists during a number of field trips where fish communities and habitats were studied directly during diving and snorkeling excursions (Berkström et al., 2012; Berkström et al., 2013b; Tano et al., 2017). LEK was found to correspond with CSK in 50% of the respondents’ answers, while LEK and CSK were completely different in only 8% of answers. This indicates that, even though there was a significant difference found between the two different epistemological knowledge systems due to the different approaches of acquiring ecological knowledge, both LEK and CSK can be used to complement each other. This was in accordance with other studies comparing LEK and CSK on fish biology and ecology in Tanzania (Gaspare et al., 2015), Brazil (Silvano et al., 2006, 2010; Silvano and Begossi, 2012; Begossi et al., 2016; Lima et al., 2017), and the Philippines (Selgrath et al., 2016). Multiple research papers advocate that local knowledge should be used in conjunction with scientific knowledge (Berkes et al., 2000; Johannes et al., 2000; Garcia-Quijano, 2015). However, Davis and Ruddle (2010) emphasized the point that LEK needs to be assessed for accuracy and validated with CSK. In the present study, the main discrepancy was between LEK and CSK on fish in mangrove habitats for 25% of the fish species. For these, LEK indicated that fish did not use mangrove habitats while CSK indicated that they did. This discrepancy may be explained by the fact that none of the fishers that were interviewed actively fished in mangrove habitats. Fishers would not have had observational experience with mangrove dwelling or migratory fishes and hence reflecting the lack of knowledge regarding this habitat. As García-Quijano (2007) stated: “fishers’ knowledge and experience are based on thousands of hours “sampling” local ecosystems with their fishing gear” and hence the lack of connectivity knowledge in mangrove habitats is likely due to the lack of “sampling” this habitat with their fishing gear.
Regarding SAV habitats (seagrass and macroalgae), LEK stated that fish utilized SAV habitats and CSK stated the opposite for 17% of the fish species. This may be due to fishers grouping seagrass and macroalgae together under the general idea that they are vegetation growing underwater and not distinguishing between the two. Furthermore, scientific surveys of fish in the seascape are likely to underestimate the number of fish species present due to field sampling only capturing a snapshot in time. For the fish species Gerres oyena (common silver-biddy), fishers overwhelmingly underscored the habitats that this particular fish uses, due to majority of fishers indicating that this particular species used sand habitats as its primary habitat. Although Berkström et al. (2012, 2013b) and Tano et al. (2017) (from which the scientific fish data was taken) only looked at fish species that utilized coral, seagrass, macroalgae and/or mangrove habitats, Allen and Erdmann (2012) concurred that common silver-biddy do indeed utilize sandy bottoms in sheltered waters near coral reefs. The fishers also indicated that the species might occur in coral and SAV habitats, which was similar to that of CSK stating that they do occur in those habitats.
Tropical local resource users are constantly feeling the ramifications of increased anthropogenic pressures on the tropical seascape and are experiencing decreased fish stocks and catching smaller sized fish (Jiddawi and Öhman, 2002). Fisheries management is, however, complex and often lacks accurate and viable data due to the costs and inaccessibility of areas to biologists (Garcia-Quijano, 2015). The present study highlighted that local resource users are knowledgeable in ecological processes and more importantly are familiar with fish migrations and the key habitats that different species of fish utilize, which could be used in fisheries and seascape management as well as providing valuable information for the design of MPAs. Information on fish connectivity within the seascape will facilitate in the siting of MPAs, which habitats to include, and what size and spacing is needed to maintain healthy fish populations (Johannes et al., 2000). A few studies in Turks and Caicos Islands (Close and Brent Hall, 2006) and in the Solomon Islands (Aswani and Lauer, 2006a,b; Lauer and Aswani, 2008) have coupled LEK and benthic mapping with a geographical information system (GIS) to aid in fisheries management and the implementation of MPAs. Aswani and Lauer (2006a) showed how indigenous people assisted in the design of MPAs by identifying marine habitats and related resident taxa on aerial photos which were then incorporated into a GIS database along with dive surveys from the same area. Converting fishers’ knowledge and socioecological behavior into geo-spatial data, aids in designing and implementing resource management strategies in a cost-effective and participatory way, bridging the gap between LEK and CSK (Aswani and Lauer, 2006b).
Although some progress toward the inclusion of LEK and bottom-up management systems have occurred, fishers’ knowledge has long been ignored by scientists, policy-makers, and governance institutions (Hind, 2015). Some of this lack of inclusion is likely an issue of utility. Natural scientists have found it difficult to integrate a knowledge culture, which is often qualitative and in non-standard format and different from their own (Soto, 2006). However, Valdés-Pizzini and García-Quijano (2009) showed that Puerto Rican fishers think in the same ecological way as fisheries scientists and managers by coupling fish species to different habitats. Furthermore, Begossi et al. (2016) found that fishers in Brazil classify fish taxonomy by the generic level and in a similar hierarchical fashion as that of scientist, analogous to the Linnean classification of genus. The fishers were also able to give detailed knowledge on fish diets, facilitating scientists with gaps in food-web ecology. Similarly, the fishers in the present study demonstrated relevant ecological knowledge by matching fish species to habitats and acknowledging fish migrations. As fisheries science and management evolve, current perceptions about fishers’ knowledge and their role in this area need to change (Baelde, 2007). LEK may not only fill scientific knowledge gaps, complementing CSK, but also contribute to higher success in fisheries management by making local resource users feel important and included in the process. The inclusion of LEK can also improve the political position of small-scale fishers which is often a disadvantaged stakeholder group for access to coastal resources (García-Quijano, 2007). Carmack and Macdonald (2008) argued that where focus and scale of inquiry is the same, collaborative research should take science and LEK as equals. This “conscience” approach assumes that both CSK and LEK is valid within its own set of rules and neither replaces the other. In this way the joint enquiry will have a joint focus on what is important for the local natural resource users.
In conclusion, it was deemed that the local fishers of Zanzibar had a high knowledge of connectivity which led more toward a general understanding of connectivity than an in-depth knowledge of connectivity, which scientists possess. However, the knowledge that local users possess (LEK) would aid fisheries management with valuable information regarding fish ecology and behaviors when used in conjunction with the knowledge gained from scientists (CSK). Results suggest differences between fisher groups, where fishers traveling further, exposed to multiple habitats, and fish with multiple gears have a greater knowledge on connectivity patterns within the seascape than those that fish locally, in single habitats and with just one type of gear. This should be recognized when finding “experts” within the local fishing community to consult in fisheries management. We suggest that a co-management approach to devising and implementing management proposals that incorporates both epistemological knowledge systems of LEK and CSK regarding seascape connectivity, would increase the effectiveness of natural resource management in aquatic environments.
The datasets generated for this study are available on request to the corresponding author.
All participants provided written informed consent to participate in this study. In Tanzania, at the time of the study, no ethics approval was required. No ethics approval was required in Sweden based on the nature of the interview data and the fact that the data was collected and analyzed outside of Sweden.
All authors conceived the research. MP conducted the fieldwork and analyzed the data. CB and MP wrote the draft. LN and NJ provided edits and feedback.
This work was provided by the Swedish Research Council VR, Grant: E0344801 and 2015-05902. LN acknowledges funding from the Swedish Research Council Formas Grant: 2014-1288.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor declared a past co-authorship with one of the authors NJ.
We wish to thank the individual fishers and fishing communities of Uroa, Ungunja Ukuu, Paje, Fumba, Nungwi, and Makoba for their valuable knowledge. We also wish to thank our interpreter Khamis for his help during data collection and Maria Eggertsen for statistical advice.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2019.00130/full#supplementary-material
Allen, G. R., and Erdmann, M. V. (2012). Reef Fishes of the East Indies: Volumes I-III, Tropical Reef Research. Perth: Universitiy of Hawai’i Press.
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Aswani, S., and Lauer, M. (2006a). Benthic mapping using local aerial photo interpretation and resident taxa inventories for designing marine protected areas. Environ. Conserv. 33, 263–273. doi: 10.1017/S0376892906003183
Aswani, S., and Lauer, M. (2006b). Incorporating Fishermen’s local knowledge and behavior into geographical information systems (GIS) for designing marine protected areas in Oceania. Hum. Organ. 65, 81–102. doi: 10.17730/humo.65.1.4y2q0vhe4l30n0uj
Aswani, S., and Vaccaro, I. (2008). Lagoon ecology and social strategies: habitat diversity and ethnobiology. Hum. Ecol. 36, 325–341. doi: 10.1007/s10745-007-9159-9
Baelde, P. (2007). “Using fisher’s knowledge goes beyond filling gaps in scientific knowledge: analysis of Australian experiences,” in Fishers’s Knowledge in Fisheries Science and Management, eds N. Haggan, B. Neis, and I. G. Baird (Paris: UNESCO).
Begossi, A., Salivonchyk, S., Lopes, P. F. M., and Silvano, R. A. M. (2016). Fishers’ knowledge on the coast of Brazil. J. Ethnobiol. Ethnomed. 12:20. doi: 10.1155/2018/7963401
Berkes, F., Colding, J., and Folke, C. (2000). Rediscovery of traditional eological knowledge as adaptive management. Ecol. Appl. 10, 1251–1262. doi: 10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Berkström, C. (2012). Ecological Connectivity in East African Seascapes. Ph.D. thesis, Stockholm University, Stockholm.
Berkström, C., Gullström, M., Lindborg, R., Mwandya, A. W., Yahya, S. A. S., Kautsky, N., et al. (2012). Exploring ‘knowns’ and ‘unknowns’ in tropical seascape connectivity with insights from East African coral reefs. Estuar. Coast. Shelf Sci. 107, 1–21. doi: 10.1016/j.ecss.2012.03.020
Berkström, C., Jörgensen, T. L., and Hellström, M. (2013a). Ecological connectivity and niche differentiation between two closely related fish species in the mangrove-seagrass-coral reef continuum. Mar. Ecol. Prog. Ser. 477, 201–215. doi: 10.3354/meps10171
Berkström, C., Lindborg, R., Thyresson, M., and Gullström, M. (2013b). Assessing connectivity in a tropical embayment: fish migrations and seascape ecology. Biol. Conserv. 166, 43–53. doi: 10.1016/j.biocon.2013.06.013
Carmack, E., and Macdonald, R. (2008). Water and ice-related phenomena in the coastal region of the Beaufort Sea: some parallels between native experience and western science. Arctic 61, 265–280.
Claydon, J. (2004). Spawning aggregatins of coral reef fishes: characteristics, hypotheses, threats and management. Oceanogr. Mar. Biol. Annu. Rev. 42, 265–302. doi: 10.1201/9780203507810.ch7
Close, C. H., and Brent Hall, G. (2006). A GIS-based protocol for the collection and use of local knowledge in fisheries management planning. J. Environ. Manage. 78, 341–352. doi: 10.1016/j.jenvman.2005.04.027
Crona, B., and Bodin, Ö (2006). What you know is who you know? Communication patterns among resource users as a prerequisite for co-management. Ecol. Soc. 11:7. doi: 10.5751/ES-01793-110207
Crona, B. I. (2006). Supporting and enhancing development of heterogeneous ecological knowledge among resource users in a Kenyan seascape. Ecol. Soc. 11:32. doi: 10.5751/ES-01712-110132
Davis, A., and Ruddle, K. (2010). Constructing confidence: rational skepticism and systematic enquiry in local ecological knowledge research. Ecol. Appl. 20, 880–894. doi: 10.1890/09-0422.1
Davis, A., and Wagner, J. R. (2003). Who Knows? On the importance of identifying “experts” when researching local ecological knowledge. Hum. Ecol. 31, 463–489. doi: 10.1023/a:1025075923297
Dorenbosch, M., Verweij, M. C., Nagelkerken, I., Jiddawi, N., and van der Velde, G. (2004). Homing and daytime tidal movements of juvenile snappers (Lutjanidae) between shallow-water nursery habitats in Zanzibar, Western Indian Ocean. Environ. Biol. Fish. 70, 203–209. doi: 10.1023/B:EBFI.0000033336.10737.f5
Garcia-Quijano, C. (2015). “Ecosystem-based knowledge and reasoning in tropical, multi-species, small-scale fishers’ LEK: what can fishers’ LEK contribute to coastal ecological science and management?,” in Fisher’s Knowledge and the Ecosystem Approach to Fisheries: Applications, Experiences and Lessons in Latin America, eds J. Fischer, J. Jorgensen, H. Josupeit, D. Kalikoski, and C. Lucas (Rome: FAO), 19–40.
García-Quijano, C. G. (2007). Fishers’ knowledge of marine species assemblages: bridging between scientific and local ecological knowledge in Southeastern Puerto Rico. Am. Anthropol. 109, 529–536. doi: 10.1525/aa.2007.109.3.529
Gaspare, L., Bryceson, I., and Kulindwa, K. (2015). Complementarity of fishers’ traditional ecological knowledge and conventional science: contributions to the management of groupers (Epinephelinae) fisheries around Mafia Island, Tanzania. Ocean Coast. Manage. 114, 88–101. doi: 10.1016/j.ocecoaman.2015.06.011
Hamilton, R., de Mitcheson, Y. S., and Aguilar-Perera, A. (2012). “The role of local ecological knowledge in the conservation and management of reef fish spawning aggregations,” in Reef Fish Spawning Aggregations: Biology, Research and Management, eds Y. S. de Mitcheson and P. L. Colin (Berlin: Springer), 331–369. doi: 10.1007/978-94-007-1980-4_10
Hind, E. J. (2015). A review of the past, the present, and the future of fishers’ knowledge research: a challenge to established fisheries science. ICES J. Mar. Sci. 72, 341–358. doi: 10.1093/icesjms/fsu169
Hyndes, G. A., Nagelkerken, I., McLeod, R. J., Connolly, R. M., Lavery, P. S., and Vanderklift, M. A. (2014). Mechanisms and ecological role of carbon transfer within coastal seascapes. Biol. Rev. 89, 232–254. doi: 10.1111/brv.12055
Jiddawi, N. S., and Öhman, M. C. (2002). Marine fisheries in Tanzania. AMBIO 31, 518–527. doi: 10.1579/0044-7447-31.7.518
Johannes, R. E., Freeman, M. M. R., and Hamilton, R. J. (2000). Ignore fishers’ knowledge and miss the boat. Fish Fish. 1, 257–271. doi: 10.1111/j.1467-2979.2000.00019.x
Karr, K. A., Fujita, R., Carcamo, R., Epstein, L., Foley, J. R., Fraire-Cervantes, J. A., et al. (2017). Integrating science-based co-management, partnerships, participatory processes and stewardship incentives to improve the performance of small-scale fisheries. Front. Mar. Sci. 4:345. doi: 10.3389/fmars.2017.00345
Khamis, Z. A., Kalliola, R., and Käyhkö, N. (2017). Geographical characterization of the Zanzibar coastal zone and its management perspectives. Ocean Coast. Manage. 149, 116–134. doi: 10.1016/j.ocecoaman.2017.10.003
Lauer, M., and Aswani, S. (2008). Integrating indigenous ecological knowledge and multi-spectral image classification for marine habitat mapping in Oceania. Ocean Coast. Manage. 51, 495–504. doi: 10.1016/j.ocecoaman.2008.04.006
Lauer, M., and Aswani, S. (2010). Indigenous knowledge and long-term ecological change: detection, interpretation, and responses to changing ecological conditions in Pacific Island communities. Environ. Manage. 45, 985–997. doi: 10.1007/s00267-010-9471-9
Le Fur, J., Guilavogui, A., Teitelbaum, A., and Rochet, M.-J. (2011). Contribution of local fishermen to improving knowledge of the marine ecosystem and resources in the Republic of Guinea, West Africa. Can. J. Fish. Aquat. Sci. 68, 1454–1469. doi: 10.1139/f2011-061
Lima, M. S. P., Oliviera, J. E. L., de Nóbrega, M. F., and Lopes, P. F. M. (2017). The use of Local Ecological Knowledge as a complementary approach to understand the temporal and spatial patterns of fishery resources distribution. J. Ethnobiol. Ethnomed. 13:30. doi: 10.1186/s13002-017-0156-9
Mackinson, S. (2001). Integrating local and scientific knowledge: an example in fisheries science. Environ. Manage. 27, 533–545. doi: 10.1007/s0026702366
McClanahan, T. R. (1988). Seasonality in East Africa’s coastal waters. Mar. Ecol. Prog. Ser. 44, 191–199. doi: 10.3354/meps044191
Mumby, P. J. (2006). Connectivity of reef fish between mangroves and coral reefs: algorithms for the design of marine reserves at seascape scales. Biol. Conserv. 128, 215–222. doi: 10.1016/j.biocon.2005.09.042
Nagelkerken, I., Sheaves, M., Baker, R., and Connolly, R. M. (2015). The seascape nursery: a novel spatial approach to identify and manage nurseries for coastal marine fauna. Fish Fish. 16, 362–371. doi: 10.1111/faf.12057
Nenadovic, M., Johnson, T., and Wilson, J. (2012). Implementing the western gulf of Maine area closure: the role and perception of fishers’ ecological knowledge. Ecol. Soc. 17:20. doi: 10.5751/ES-04431-170120
Ogden, J. C. (1988). “The influence of adjacent systems on the structure and function of coral reefs,” in Proceedings of the Sixth International Coral Reef Symposium, Vol. 1, Australia, 123–129.
Olds, A. D., Albert, S., Maxwell, P. S., Pitt, K. A., and Connolly, R. M. (2013). Mangrove-reef connectivity promotes the effectiveness of marine reserves across the western Pacific. Glob. Ecol. Biogeogr. 22, 1040–1049. doi: 10.1111/geb.12072
Olds, A. D., Connolly, R. M., Pitt, K. A., Pittman, S. J., Maxwell, P. S., Huijbers, C. M., et al. (2016). Quantifying the conservation value of seascape connectivity: a global synthesis. Glob. Ecol. Biogeogr. 25, 3–15. doi: 10.1111/geb.12388
Olsson, P., and Folke, C. (2001). Local ecological knowledge and institutional dynamics for ecosystem management: a study of lake Racken watershed, Sweden. Ecosystems 4, 85–104. doi: 10.1007/s100210000061
Pauly, D., and Zeller, D. (2016). Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 7:10244. doi: 10.1038/ncomms10244
Robinson, J., Isidore, M., Marguerite, M. A., Öhman, M. C., and Payet, R. J. (2004). Spatial and temporal distribution of reef fish spawning aggregations in the Seychelles - an interview-based survey of artisanal fisheries. Western Indian Ocean J. Mar. Sci. 3, 63–69.
Ruddle, K. (1995). “The role of validated local knowledge in the restoration of fisheries property rights: the example of the New Zealand Maori,” in Property Rights in a Social and Ecological Context. Case Studies and Design Applications, eds S. Hanna and M. Munasinghe (Washington DC: The World Bank), 11–120.
Selgrath, J. C., Roelfsema, C., Gergel, S. E., and Vincent, A. C. J. (2016). Mapping for coral reef conservation: comparing the value of participatory and remote sensing approaches. Ecosphere 7:e01325. doi: 10.1002/ecs2.1325
Sheaves, M., Baker, R., Nagelkerken, I., and Connolly, R. M. (2015). True value of estuarine and coastal nurseries for fish: incorporating complexity and dynamics. Estuar. Coasts 38, 401–414. doi: 10.1007/s12237-014-9846-x
Shephard, S., Connolly, P., Hareide, N.-R., and Rogan, E. (2007). Establishing stakeholder connections for management of the Irish orange roughy fishery. ICES J. Mar. Sci. 64, 841–845. doi: 10.1093/icesjms/fsl034
Silvano, M., Azevedo, R., and Begossi, A. (2010). What can be learned from fishers? An integrated survey of fishers’ local ecological knowledge and bluefish (Pomatomus saltatrix) biology on the Brazilian coast. Hydrobiologia 637, 3–18. doi: 10.1007/s10750-009-9979-2
Silvano, R. A. M., and Begossi, A. (2012). Fishermen’s local ecological knowledge on Southeastern Brazilian coastal fishes: contributions to research, conservation, and management. Neotrop. Ichthyol. 10, 133–147. doi: 10.1590/S1679-62252012000100013
Silvano, R. A. M., MacCord, P. F. L., Lima, R. V., and Begossi, A. (2006). When does this fish spawn? Fishermen’s local knowledge of migration and reproduction of Brazilian coastal fishes. Environ. Biol. Fish. 76, 371–386. doi: 10.1007/s10641-006-9043-2
Soto, C. G. (2006). Socio-Cultural Barriers to Applying Fishers’ Knowledge in Fishereies Management: An Evaluation of Litterature Cases. Burnaby: Simon Fraser University.
Tano, S. A., Eggertsen, M., Wikström, S. A., Berkström, C., Buriyo, A. S., and Halling, C. (2017). Tropical seaweed beds as important habitats for juvenile fish. Mar. Freshw. Res. 68, 1921–1934. doi: 10.1071/MF16153
Taylor, R. B., Morrison, M. A., and Shears, N. T. (2011). Establishing baselines for recovery in a marine reserve (Poor Knights Islands, New Zealand) using local ecological knowledge. Biol. Conserv. 144, 3038–3046. doi: 10.1016/j.biocon.2011.09.009
Thornton, T. F., and Scheer, A. M. (2012). Collaborative engagement of local and traditional knowledge and science in marine environments: a review. Ecol. Soc. 17:8. doi: 10.5751/es-04714-170308
Unsworth, R. K. F., Bell, J. J., and Smith, D. J. (2007). Tidal fish connectivity of reef and seagrass habitats in the Indo-Pacific. J. Mar. Biol. Assoc. 87, 1287–1296. doi: 10.1017/S002531540705638X
Unsworth, R. K. F., McKenzie, L. J., Nordlund, L. M., and Cullen-Unsworth, L. C. (2018a). A changing climate for seagrass conservation? Curr. Biol. 28, R1229–R1232. doi: 10.1016/j.cub.2018.09.027
Unsworth, R. K. F., Nordlund, L. M., and Cullen-Unsworth, L. C. (2018b). Seagrass meadows support global fisheries production. Conserv. Lett. 12:e12566. doi: 10.1111/conl.12566
Keywords: small-scale fisheries, seascape, fish migrations, data-poor, participatory research, coral reef, mangrove, seagrass
Citation: Berkström C, Papadopoulos M, Jiddawi NS and Nordlund LM (2019) Fishers’ Local Ecological Knowledge (LEK) on Connectivity and Seascape Management. Front. Mar. Sci. 6:130. doi: 10.3389/fmars.2019.00130
Received: 12 November 2018; Accepted: 04 March 2019;
Published: 22 March 2019.
Edited by:
Beatrice Irene Crona, Royal Swedish Academy of Sciences, SwedenReviewed by:
Fabio Fiorentino, Istituto per le Risorse Biologiche e le Biotecnologie Marine (IRBIM), ItalyCopyright © 2019 Berkström, Papadopoulos, Jiddawi and Nordlund. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Charlotte Berkström, Y2hhcmxvdHRlLmJlcmtzdHJvbUBzdS5zZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.