 Carlos Hermosilla1*
Carlos Hermosilla1* Jörg Hirzmann1
Jörg Hirzmann1 Liliana M. R. Silva1
Liliana M. R. Silva1 Sandra Scheufen2Ellen Prenger-Berninghoff2
Sandra Scheufen2Ellen Prenger-Berninghoff2 Christa Ewers2Vreni Häussermann3,4Günter Försterra3,4Sven Poppert5
Christa Ewers2Vreni Häussermann3,4Günter Försterra3,4Sven Poppert5 Anja Taubert1
Anja Taubert1- 1Institute of Parasitology, Justus Liebig University Giessen, Giessen, Germany
- 2Institute for Hygiene and Infectious Diseases of Animals, Justus Liebig University Giessen, Giessen, Germany
- 3Huinay Scientific Field Station, Puerto Montt, Chile
- 4Pontificia Universidad Católica de Valparaíso, Valparaíso, Chile
- 5Swiss Tropical and Public Health Institute, Basel, Switzerland
Present study aimed to characterize gastrointestinal parasites and culturable bacteria from free-living South American sea lions (Otaria flavescens) inhabiting waters of Comau Fjord, Patagonia, Chile. Therefore, a total of 28 individual fecal samples were collected from sea lions within their natural marine habitat during several diving expeditions. Using classical parasitological techniques, study revealed infections with five different gastrointestinal parasite genera. In addition, bacterial cultures showed presence of at least 28 different bacterial genera. Referring to parasites, protozoan, and metazoan species were found with some of them bearing anthropozoonotic potential and/or pathogenic impact for these marine mammals. As such, four of identified parasite genera harbored zoonotic potential (i.e., Entamoeba, Balantidium, Diphyllobothrium, Anisakis) and one genus (Parafilaroides) represented a specific lungworm of marine pinnipeds. Proglottids from fecal samples showed high morphological homology to “Diphyllobothrium” scoticum (Rennie and Reid, 1912; Meggitt, 1924), which was found in Antarctic sea leopards (Hydrurga leptonyx; Phocidae), but contained eggs of smaller size. Molecular characterization revealed 97–100% identity to a new “Diphyllobothrium” species which was recently isolated from a Californian sea lion (Zalophus californianus; Otariidae) in San Francisco. As such, O. flavescens represents a new host record for this parasite species. Furthermore, potential zoonotic bacteria (i.e., Clostridium, Escherichia, Vibrio, Yersinia, Salmonella) were identified amongst others in O. flavescens indicating a reservoir role for these pinnipeds in marine ecosystem. Current data should be considered as a baseline study for future monitoring surveys on anthropozoonotic pathogens circulating in wild free-living sea lions and their possible impact on public health issues and marine wildlife.
Introduction
South American sea lions (Otaria flavescens) belonging to family Otariidae are common carnivorous pinnipeds living along Eastern and Western coasts of South America. They are endemic in Argentina, Peru, South Brazil, and Chile (Vaz-Ferreira, 1982; Crespo, 1988; Túnez et al., 2008; Aznar et al., 2012; Hernández-Orts et al., 2013; Pereira et al., 2013). Along Chilean coastal shores, including Southern and Northern Patagonian fjords (Haussermann et al., 2014), more than 200 colonies of free-living South American sea lions were described.
So far, abundant reports on ecology, feeding, behavior, reproduction, sightings, life history parameters, as well as population dynamics of this otariid species are available (Crespo et al., 1997, 1999; Lima and Páez, 1997; Alonso et al., 2000; Naya et al., 2002; Suarez et al., 2005; Haussermann et al., 2014). Several reports have further focused on endogenous helminth fauna and microbiota of South American sea lions, comprising single parasite and bacterial species records, taxonomy, and population studies on some of these parasitoses and bacterial diseases (Cattan et al., 1976; George-Nascimento and Carvajal, 1981; Zdzitowircki, 1986; Hernández-Orts et al., 2013). Nonetheless, scarce data have been reported on gastrointestinal protozoan parasites occurring in free-living South American sea lions within their natural marine environment (Hermosilla et al., 2016b). Same holds true for studies on marine mammal microbiota, despite their crucial ecological roles in oceanic ecosystem (Bik et al., 2016). Being apex predators, South American sea lions are likely to play a role as indicators of ocean health as reported elsewhere for other marine mammals (Hunt et al., 2013).
Although recent conservation efforts have allowed South American sea lion populations to slowly recover, they still are classified as least concern and would profit from a better understanding of acute, chronic or subclinical effects of parasitic and bacterial diseases for their long-term survival and preservation (Glad et al., 2010; Hermosilla et al., 2016b). In this context, pinniped-associated gastrointestinal microbiota (Glad et al., 2010) and parasites (Cattan et al., 1976; Hernández-Orts et al., 2013; Hermosilla et al., 2016b) are well-known to play a critical role in sea lion health, nutrition, differentiation of host tissue, reproduction, colonization resistance and adequate maturation of host innate and adaptive immune system (Bik et al., 2016). In healthy individuals, gut microbiota are robust and resistant to perturbations and maintain their composition in normal range to sustain homeostasis, symbiosis and proper immunity (Ohno, 2015).
The present study therefore aimed to investigate gastrointestinal parasite diversity and to identify culturable bacterial species in fecal samples being collected without any disturbance of animals during several diving expeditions at Comau Fjord of Northern Patagonia, Chile. Current findings expand knowledge on parasitic and bacterial diversity in these marine mammals and further discuss impacts of zoonotic pathogens on ocean and public health, since many of these pathogens most probably are also present in marine-derived products, such as bivalves, gastropods, crustaceans, cephalopods and fishes, foreseen for human consumption.
Materials and Methods
Study Area, Sample Collection, Coproscopical, and Bacterial Analyses
South American sea lions (O. flavescens) were investigated in a colony allocated at 11th Region of Chile (Aysén) in Comau Fjord (42.39°S, 72.44°W), Northern Patagonia, Chile. The colony was composed of approximately 200 animals comprising males, females and puppies. General topography of study area was composed of rocky shores, where sea lion colony was allocated, surrounded by waters containing robust bull kelps (Durvillaea antarctica) as dominant seaweed in this fjord. Waters in front of colony were shallow and maximum depth in opened water was approximately 30 m.
For sample collection, boat- and underwater-based surveys during scuba dives in front of sea lion colony were conducted in summer of 2014. This unique non-invasive research methodology not only allows underwater behavioral observations of sea lions (Figure 1A) which are otherwise impossible to monitor, but additionally provides easy access to sea lions’ fecal material without any animal disturbance as demonstrated recently for scat collections of free-swimming dolphins (Kleinertz et al., 2014) and large baleen whales (Hermosilla et al., 2016a; de Vos et al., 2018).

Figure 1. (A) Scat collection of South American sea lions (Otaria flavescens) (n = 28) was performed by scuba dives under water in front of sea lion colony of Comau Fjord, Patagonia, Chile, without disturbance of animals. (B) Illustration of an Anisakis spp. egg and (C) a Balandium spp. cyst (scale bar 20 μm) identified in sea lion fecal samples.
In this study, a total of 28 individual fecal samples were collected during underwater survey activities. In total 7 scuba dives were performed (with durations of 15–20 min each) to collect all scat samples. Whenever sea lion defecation occurred, scat samples were immediately collected in 15 ml plastic tubes (Falcon). Thereafter, a fecal swap was taken and transferred into special tubes containing Amies transport medium with charcoal (Nerbe, Winsen, Germany) for bacterial cultivation (Hermosilla et al., 2018) and rest of scat material was fixed in 70% ethanol for further parasitological diagnosis. All swap- as well as fecal-samples were stored at 4°C until further diagnosis within the vessel of Huinay Scientific Field Station (Chile) responsible for sea lion scat collection and thereafter transferred to Germany. Spontaneously shed helminth stages were collected during scuba dive episodes and fixed in 70% ethanol. Scat samples as well as helminth specimens were then transferred to Huinay Scientific Field Station, Puerto Montt, Chile, and stored at 4°C until further parasitic and bacteriological analyses. Parasitological as well as bacteriological analyses were conducted at the Institute of Parasitology and at the Institute for Hygiene and Infectious Diseases of Animals, respectively, both institutions belonging to Faculty of Veterinary Medicine of the Justus Liebig University Giessen, Giessen, Germany.
Coproscopical parasitic analyses included standard sodium acetate acetic acid formalin (SAF) technique according to Yang and Scholten (1977) for detection of parasite eggs, cysts, sporocysts, and oocysts within fecal samples as described elsewhere (Hermosilla et al., 2013). In addition, carbol fuchsin-stained faecal smears (CFS) were carried out for the detection of Cryptosporidium spp. oocysts according to Heine (1982). Moreover, a commercially available coproantigen ELISA (ProSpecT®, Oxoid) was performed for detection of Giardia- and Cryptosporidium-antigens in fecal samples. Parasitological identification of helminth species, cysts, oocysts, and eggs was based on morphological criteria referring to previous reports (Anderson, 1992; Kleinertz et al., 2014; Hermosilla et al., 2016a,b).
For bacterial analyses, fecal samples were streaked on blood agar containing 5% defibrinated sheep blood (E. Merck, Darmstadt, Germany) and water-blue metachrome-yellow lactose agar (acc. to Gassner, E. Merck). Plates were incubated at 37°C and analyzed after 24 and 48 h. Colonies were sub-cultured and pure cultures were identified using standard morphological and biochemical methods as well as with MALDI-TOF mass spectrometry using the Microflex LT/SH® instrument according to the manufacturer’s instructions (Bruker Daltonics, Bremen, Germany) (Bisping and Amtsberg, 1988; Burkhardt, 1992; Carter et al., 1995). For Clostridium (C.) perfringens detection, samples were streaked on Zeissler agar (E. Merck, Damstadt, Germany) and incubated at 37°C overnight under anaerobic conditions in a jar using AnaeroGenTM gas sachets (Oxoid, Wesel, Germany). After streaking on agar plates, fecal material was placed in Müller-Kauffmann Novobiocin Tetrathionate broth (E. Merck) with iodine as well as in Rappaport-Vassiliadis enrichment broth (Oxoid) which was incubated at 37 and 43°C, respectively. Ten μl of each broth was cultured on water-blue metachrome-yellow lactose agar as well as on Brilliance Salmonella agar containing Salmonella Selective Supplement (Oxoid, Wesel) after 24 and 48 h of broth incubation. Suspect Salmonella colonies were sub-cultured on water-blue metachrome-yellow lactose agar and analyzed after 24 h of incubation by MALDI-TOF mass spectrometry. Salmonella serovars determination was performed at the National Reference Laboratory for the Analysis and Testing of Zoonosis (Salmonella) at the German Federal Institute of Risk Assessment (Berlin, Germany).
All fecal sampling procedures were conducted in accordance to Institutional Ethic Commissions of Huinay Scientific Field Station (Chile) and of Justus Liebig University Giessen (Germany), and in accordance to current Chilean Animal Protection Laws.
Morphological Analyses and Staining of Cestodes
Two cestode strobilas – one collected from a rock of seal colony and another retrieved from a fecal sample (sea lion ID F07) – were investigated microscopically for further species identification. Strobilas preserved and shipped in 70% ethanol were hydrated for 1 day in water at 4°C with several changes. Complete fragments, sagittal, median sagittal and transverse sections of strobila were examined using stereomicroscopy (Leica) and were documented by a digital camera and CellSens® software (both Olympus). Some strobila fragments and thick transverse sections – cut with a razor blade – were stained with iron acetocarmine (Merck) following protocol of Georgiev et al. (1986) and mounted in DePeX. For a detailed analysis of reproductive organs, strobila fragments (1 cm) were fixed in 4% formaldehyde, embedded in paraplast and 10 μm sagittal and transverse sections were stained with hematoxylin and eosin (HE). HE staining was performed with Harris hematoxylin (Thermo Fisher Scientific) and 0.25% Eosin Y (Merck) in 80% ethanol following a xylene-free protocol which uses 1.7% dish washer solution at 90°C as deparaffinizing agent and over-drying at 60°C instead of dehydration (Negi et al., 2013). Microscopic measurements were performed using a digital camera and CellSens® software (Olympus).
For Diphyllobothriidea species identification, keys and checklists (Dailey, 1975; Delyamure et al., 1985; Felix, 2013; Hernandez-Orts et al., 2015; Kuchta and Scholz, 2017), expedition reports on cestode collections from pinnipeds (Fuhrmann, 1921; Markowski, 1952b; Yurakhno and Maltsev, 1997) and primary literature on individual Diphyllobothrium species from seals (Yurakhno and Maltsev, 1994; Hernández-Orts et al., 2015) were reviewed. Species classification follows most recent revision of Diphyllobothriidae family with polyphyletic Diphyllobothrium species considered incertae sedis indicated by inverted commas (Waeschenbach et al., 2017).
Molecular Analyses of Cestodes
To identify sea lion cestodes sequence data of ribosomal DNA region – 18S, ITS1, 5.8S, ITS2, partial 28S – and complete mitochondrial cytochrome oxidase subunit 1 gene (cox1) were generated and compared to GenBank database entries. Genomic DNA was isolated from cestode proglottids using DNeasy blood and tissue kit (Qiagen, Hilden, Germany). The rDNA region was amplified using primer combinations NF1 (Porazinska et al., 2009)/D3A (Nunn, 1992) and ZX-1 (Waeschenbach et al., 2007)/L2230 (Lockyer et al., 2003). The cox1 gene was amplified using following primer combinations: Cox1F/Cox1R (Wicht et al., 2010), COI Dice1F/Dice11R (Van Steenkiste et al., 2015), and Diphyllnad/Diphyllrrnl (Mercado et al., 2010). PCRs were performed in 50 μl reactions using HOT FIREPol Blend Master Mix 7.5 mM MgCl2 (Solis BioDyne, Tartu, Estonia), 200 nM of forward and reverse primers each, and 100 ng of worm DNA under the following conditions: 15 min 95°C initial denaturation, 35 cycles 20 s 95°C, 30 s 54°C, 2.5 min 72°C and 5 min 72°C final extension. For amplification of partial cox1 with primers Dice1F and Dice11R a touchdown-protocol was performed (Van Steenkiste et al., 2015). Amplicons were gel-purified, cloned, and sequenced by an external service provider (LGC Genomics GmbH, Berlin, Germany). Complete sequences of rDNA and mitochondrial region were assembled from overlapping amplicons and analyzed by BLAST search against GenBank database. Sequences are available under accession numbers KY945917 (rDNA, gravid proglottid), KY945922 (cox1, gravid proglottid) and MF893274 (cox1, immature proglottid fecal sample F07), respectively.
For phylogenetic analysis, a BLAST search in GenBank database was conducted with amplified sequences and a dataset of high-scoring Diphyllobothriidae taxa was chosen. For each taxon, homologous sequences of 3′-region of 18S rDNA and 5′-region (D2-D3) of 28S rDNA were concatenated and aligned by MAFFT 7 (Katoh and Standley, 2013). Only those taxa were considered with both sequence regions available from GenBank (Supplementary Table S1). Aligned sequences corresponded to nucleotides 1451-1650 (3′ 18S) and nucleotides 3851–4700 (5′ 28S D2-D3) of rDNA sequence from gravid proglottid (KY945917). A phylogenetic tree was constructed using Bayesian analysis (MrBayes 3.2) and TreeDyn from Phylogeny.fr website (Dereeper et al., 2008).
Results
Parasite Infections
Parasitological analyses of South American sea lion fecal samples revealed five different parasite taxa comprising two protozoan and three metazoan taxa. Metazoan parasites consisted of cestodes (one species) and nematodes (two species). Neither acanthocephalan nor trematode eggs were detected in current survey. A list with parasite stages and their respective prevalence is represented in Table 1. Overall, most prevalent metazoan parasitic stages found in sea lion colony were eggs of Diphyllobothriidae gen. sp. (44.8%), followed by eggs of Anisakis spp. (34.5%; Figure 1B) and at much lower prevalence Parafilaroides spp. larvae (3.4%). Most prevalent protozoan parasitic stages found in sea lion samples were cysts of Balantidium spp. (13.8%; Figure 1C) followed by cysts of Entamoeba spp. (3.4%). Neither oocysts/antigens of Cryptosporidium nor cysts/antigens of Giardia were found. Referring to parasite genus level, these parasitological findings include one new host records, i.e., Entamoeba, for O. flavescens.
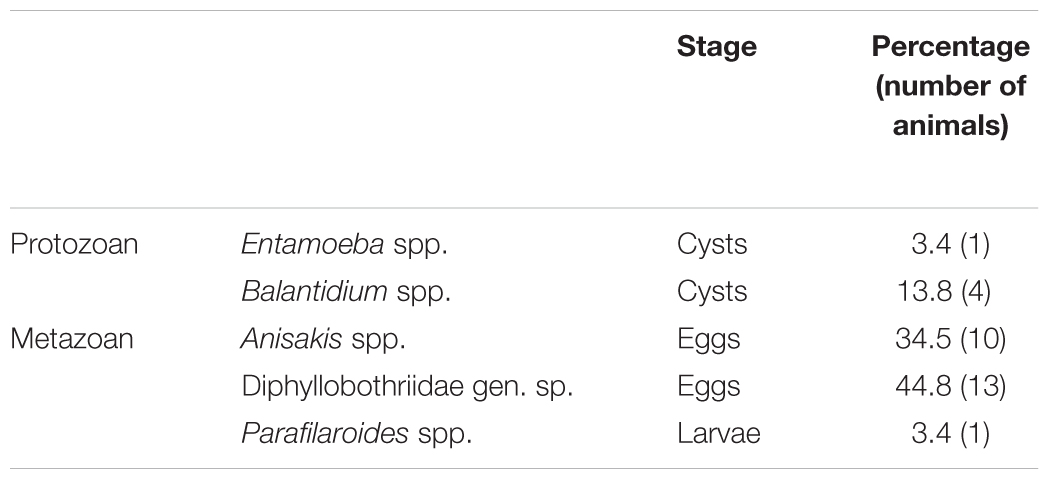
Table 1. Gastrointestinal parasites identified in South American sea lion (Otaria flavescens) fecal samples with respective percentages.
Important to note is that some of gastrointestinal endoparasites detected in South American sea lions bear anthropozoonotic potential, such as Balantidium, Entamoeba, Diphyllobothriidae (Adenocephalus) and Anisakis.
Morphology of “Diphyllobothrium” Proglottids
Two scolex-less “Diphyllobothrium” strobilas, a large one (length 50 cm) with gravid proglottids from a rock near colony (Figure 2A) and a short one with immature proglottids isolated from a fecal sample (Figure 2D) were collected. Sequencing cox1 gene of both strobilas confirmed that these belong to same species and proved that large strobila also originated from sea lions. Both strobilas were segmented, slightly craspedote (Figure 2C) with proglottids wider than long. Gravid proglottids had a mean width of 7233 μm and a length of 1037 μm (min-max: 7077–7418 μm × 825–1385 μm; n = 11); immature proglottid was much smaller with a mean width of 2500 μm. Gravid strobila was fleshy thick (1.5 mm; Figures 2F,I; Immature: Figure 2E) with a striking bulged, pocket-like appearance on ventral surface around uterine openings (Figure 2H). As shown in unstained longitudinal sections, these pockets correspond to the large distal uterine sac, densely packed with hundreds of eggs (Figure 2F). Eggs liberated from uterus were unembryonated, oval, thick-walled, operculated and with a knob on opposite pole (Figure 2G). Their mean size was 57.4 μm × 40.9 μm (min–max: 54.1–61.3 μm × 38.5–44.1; n = 22). Uterine openings were slit-like and situated in a transverse groove, irregularly alternating lateral to midline of strobila (Figures 2B,C). Vitteline glands (max. size 175 μm × 30 μm) were seen in cortical, lateral parts of proglottids in iron acetocarmine stained strobila fragments and thick transverse sections, leaving a free space around median region of uterus openings and genital pores (Figures 2C,J). Numerous calcareous corpuscles (18 μm × 9 μm, pale violet stained) were seen in dorsal and ventral cortical parenchyma (Figure 2L). Testes were located between well-developed (100 μm thick) dorsal and ventral longitudinal muscles (Figure 2K) arranged in a single layer with 15–20 testes seen in transverse sections on each side of segment (Figure 2J). The ovary in posterior part of proglottid was followed by a tube-like uterus, which made 5–6 loops on each side (Figure 2C) terminating in a large uterine sac lined with villous tissue (Figure 2L). In HE-stained sagittal sections, eggs have been lost during processing but a few numbers of bile-colored eggs and their pattern in villous tissue were still seen. The receptaculum seminis seen in median longitudinal sections was filled with sperms (Figure 2L). Male genital organs were located anterior from uterine sac. The vas deferens led to a muscular seminal vesicle completely filled with sperms followed by cirrus-sac with coiled cirrus (Figure 2L). Size and shape of genital apparatus was of taxonomic value according to Markowski (1952b). Cirrus-sac wall appeared less muscular than wall of seminal vesicle. In all median sagittal sections, cirrus-sac was longer than seminal vesicle.
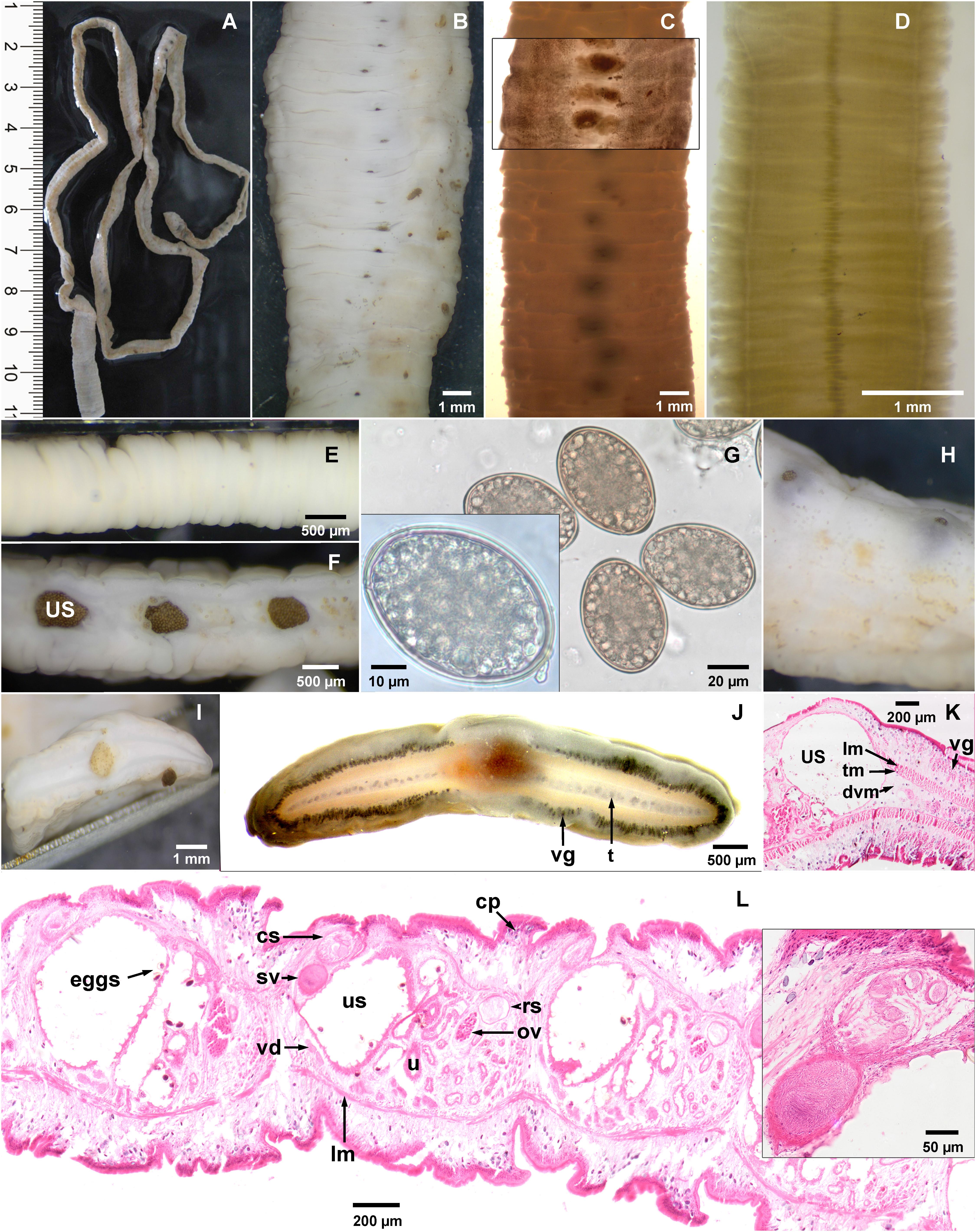
Figure 2. Morphology of Diphyllobothrium scoticum-like strobila collected from South American sea lions (Otaria flavescens) of Comau Fjord, Chile. (A–C,F–L) gravid strobila, (D,E) immature strobila. (B–D) Ventral view of gravid (B,C) and immature strobila (D) showing variation in size and shape and irregularly alternating uterine openings. (E) Lateral (immature strobila) and (F) median sagittal view. (G) Eggs with operculum and knob. (H) Ventral view of species-specific pocket-like terminal uterine sac. (I,J) Transverse thick sections: (J) showing 15–18 testes in single layer on each side and vitelline glands (iron acetocarmine staining). (K) Transverse section showing vitelline glands and muscle layers (HE staining). (L) Median sagittal section showing male and female genital organs; eggs in uterine sac were lost during processing. lm, tm, dvm longitudinal, transverse, dorso ventral muscles; cp, calcareous particles; t, testes; vd, vas deferens; sv, seminal vesicle; cs, cirrus sac; ov, ovary; rs, receptaculum seminis; u, uterus; us, uterine sac; vg, vitelline glands.
However, two differences of present specimens compared to D. scoticum are obvious. Firstly eggs obtained from gravid proglottids from O. flavescens are considerably smaller, and secondly cirrus sac and seminal vesicle have a reverse size ratio (Table 2). For further detailed characterization we performed additionally molecular analysis.
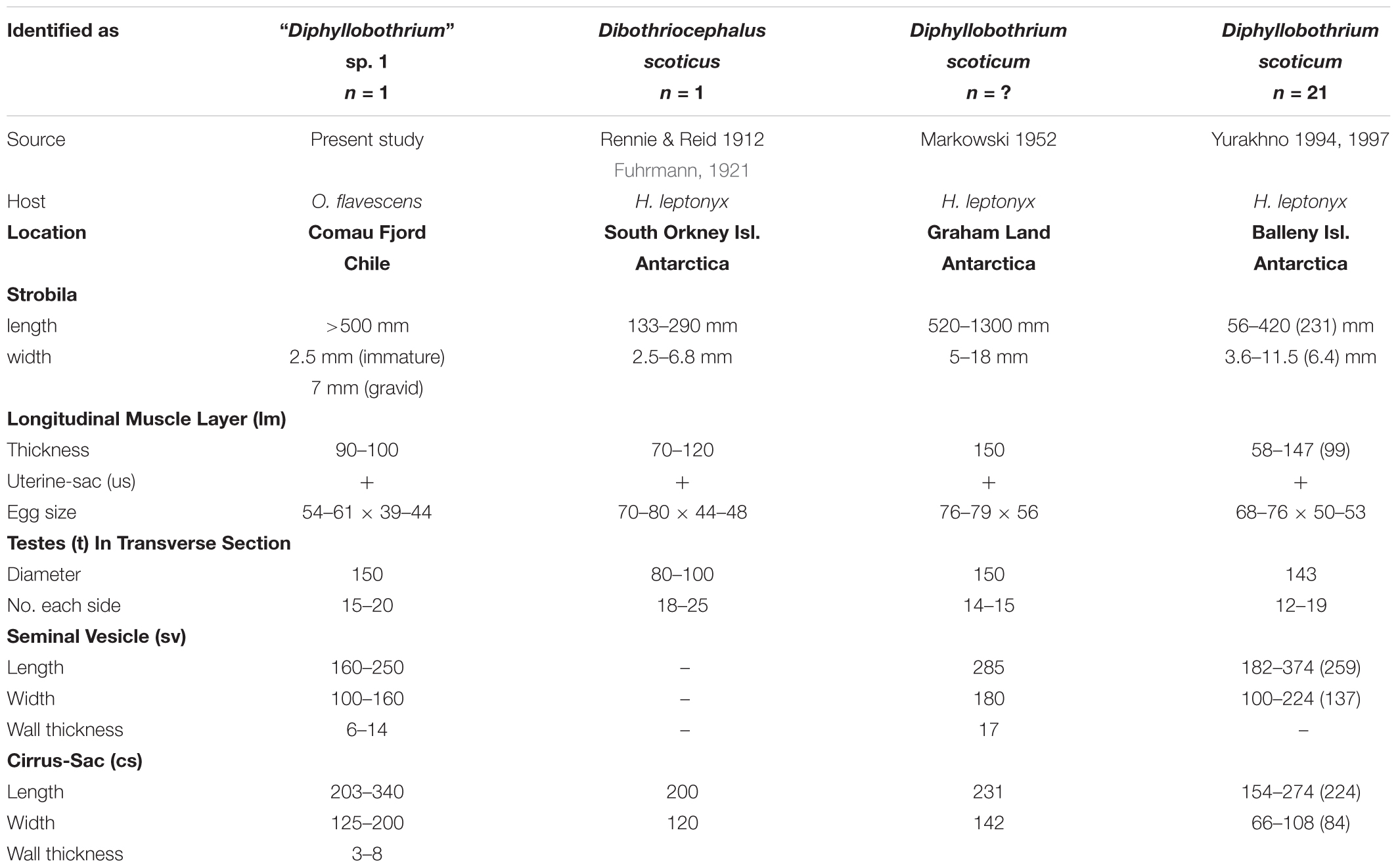
Table 2. Morphometric data of “Diphyllobothrium” sp. 1 from Otaria flavescens compared to “Dip”. scoticum from H. leptonyx based on literature [μm; (mean)].
Molecular and Phylogenetic Analyses of “Diphyllobothrium” Scoticum-Like Strobilas
In order to confirm species identity of two strobilas, we amplified and compared a fragment of mitochondrial genome from both specimens including complete cox1- and partial nad3- and 16S-sequences. Complete amplicons were 3017 bp for both strobilas with an identity of 99.4% (2999/3017 bp) – clearly beyond the threshold of 9% determined for cox1 in delimitation of platyhelminth species (Hebert et al., 2003; Vilas et al., 2005) – confirming that both were haplotypes of same species. A BLAST search of this sequence toward GenBank resulted in a closely related sequence (KY552890) of 97% identity (1519/1566 bp) belonging to an unidentified Diphyllobothrium sp. 1 from a California sea lion (Waeschenbach et al., 2017), whereas sequences from other morphologically identified diphyllobothridean species exhibited a lower homology of 89% identity or less (e.g., Dib. dentriticus KY552847: 89%, 841/949 bp; Dib. latus AB269325: 86%, 2586/3022 bp; A. pacificus KR269744: 83%, 1295/1566 bp). Thus, the two “Diphyllobothrium” scoticum-like strobilas found were supposed to be conspecific with Diphyllobothrium sp. 1.
For further molecular analysis we amplified rDNA region – 18S, ITS1, 5.8S, ITS2 and partial 28S – from the gravid strobila; 5994 bp. The 18S sequence was 100% identical with the Californian isolate (KY552794; 1994 bp) and partial 28S sequence was almost identical having only a 4 bp deletion (KY552829; 1481/1485 bp). This confirms supposed species identity based on cox1 sequence, which showed more variation (97% identity) because mitochondrial genes evolve faster than nuclear ones. In conclusion, “D.” scoticum-like strobilas from Chilean South American sea lions were identified as “Diphyllobothrium” sp. 1.
To investigate taxonomic position and relationship to other Diphyllobothriidae genera and species a concatenated sequence of 18S 3′-region and 28S 5′-region was used for phylogenetic analysis. In inferred phylogenetic tree (Figure 3) the “Diphyllobothrium” sp. 1 species clusters with A. pacificus, “D.” scoticum collected from a Southern elephant seal (Mirounga leonina) of Macquarie island (island between Tasmania and Antarctica) and another unidentified Diphyllobothriidae gen. n. sp. of a plerocercoid from an Antarctic fish (Trematomus bernacchii). The latter two and “D.” scoticum were new sequences recently added to GenBank (Waeschenbach et al., 2017) for which no morphological descriptions were reported at time of writing this manuscript.
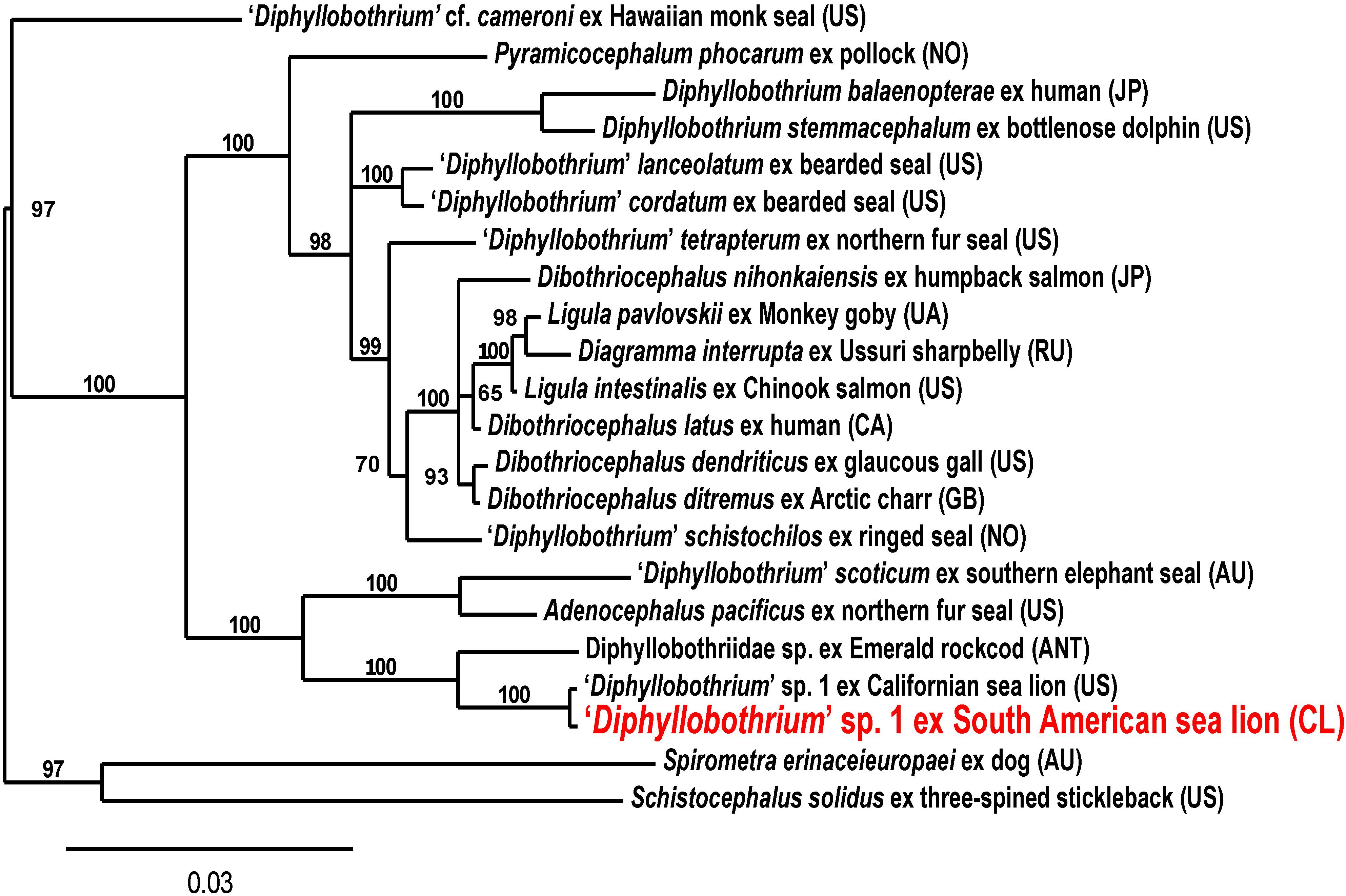
Figure 3. Phylogenetic tree of “Diphyllobothrium” sp. 1 from South American sea lions (Otaria flavescens) and related Diphyllobothriidae based on concatenated partial 18S and 28S rDNA regions using Bayesian analysis. Branch support as posterior probabilities in percent; scale bar: number of substitutions per site; host and country after the species name (Accession Nos. Supplementary Table S1).
Bacterial Findings
Bacteriological analyses of fecal samples of South American sea lions revealed a wide bacterial diversity. A complete list with bacterial species and their respective frequencies is illustrated in Table 3. At least 45 different bacterial species could be cultured from sea lion samples with Escherichia coli (82.1%) and Clostridium perfringens (64.3%) being most predominant ones. Of six detected Salmonella (S.) isolates, four were identified as serotype S. Cerro and two as S. Pensacola. To authors knowledge no report exists, so far, on serotype Pensacola in marine mammal sources. Furthermore, Yersinia enterocolitica could be detected in one fecal sample in this study.

Table 3. Bacterial species isolated from South American sea lion (Otaria flavescens) samples with respective percentages.
Discussion
The usefulness of current applied non-invasive collection method, which clearly left free-swimming sea lions unmolested in their natural environment, was reinforced (Hermosilla et al., 2015; de Vos et al., 2018). Overall, five different parasite taxa were here found in individual sea lion samples thereby covering a rather narrow range of parasites when compared to other studies (Hernández-Orts et al., 2013; Hermosilla et al., 2016b). Eggs of Diphyllobothriidae gen. sp. showed highest prevalence with 44.8%. Based on present analyses of cestode strobilas, these eggs most probably belong to “Diphyllobothrium” sp. 1 and A. pacificus – the latter being quite abundant in South American sea lions (Torres et al., 2012; Hernández-Orts et al., 2013, 2015). So far, three diphyllobothriidean tapeworms were reported in Chile: two freshwater species [Dibothriocephalus latus (syn. Diphyllobothrium latum) and Dib. dendriticus (syn. Dip. dendriticum)] and one marine species [i.e., Adenocephalus pacificus (syn. Dip. pacificum)] (Torres et al., 2012) originating from stranded sea lions from Patagonia (Hernández-Orts et al., 2013) and Valdivia (Hermosilla et al., 2016b) but at lower prevalences of 26.8 and 13%, respectively.
Several morphological features used to differentiate diphyllobothriidean species are intraspecific variables, which necessitate molecular characterization for species verification as discussed for A. pacificus by Hernández-Orts et al. (2015). Surprisingly, morphology of strobilas from O. flavescens was almost identical to “Dip.” scoticum previously only identified in sea leopards of Antarctica. “Dip.” scoticum is characterized by a species-specific uterine sac also present in species from O. flavescens. The only obvious differences were the smaller size of eggs and size proportion of cirrus sac versus seminal vesicle. Diphyllobothriid species egg size was previously shown to be intraspecific and host-related highly variable (Hernandez-Orts et al., 2015; Leštinová et al., 2016; Yamasaki et al., 2016). However, maximum length of eggs from “Diphyllobothrium” sp. 1 (54–61 μm) was clearly shorter than minimal length of “Dip.” scoticum eggs (68–79 μm). Suspected findings of D. scoticum outside of Antarctica were reported, interestingly in O. flavescens [Baylis and Hamilton (1934) and Markowski (1952a) from Falkland islands; Hernández-Orts et al. (2011); Hernández-Orts et al. (2013) from Argentina Northern Patagonia], but later these cases were identified as A. pacificus or remained undefined (Baer et al., 1967; Hernández-Orts et al., 2015). In order to exclude misidentification we also performed a molecular characterization of two proglottids. During preparation of this publication a sequence from a diphyllobothriidean species of Zalophus californianus from San Francisco was published by Waeschenbach et al. (2017) with 100% identity to present isolates, but apart from scolex no digitals/drawings were included. Additionally, a sequence assigned to “Dip.” scoticum was published by same authors and a second new species not assigned to a genus. However, unexpected to our morphological findings, sequence identity to “Dip.” scoticum sequence was not higher than to other diphyllobothriids (cox1: 84%, 462/551; KY552883); but were more closely related in phylogenetic analysis. According to authors, “Dip.” scoticum was collected from a Southern elephant seal (M. leonina) of sub-antarctic Macquarie island (Tasmania, Australia) (Johnston, 1937). More importantly, diphyllobothriasis still represents a neglected fish-borne zoonosis not only in Chile (Mercado et al., 2010; Torres et al., 2012) but also worldwide (Curtis and Bylund, 1991; Chai et al., 2005; Scholz et al., 2009; Kuchta et al., 2015). Humans become infected by ingestion of raw or undercooked fish carrying plerocercoid-stages of Diphyllobothrium. In summary, morphology of proglottids of present “Diphyllobothrium” species were most consistent with descriptions of D. scoticum. This Antarctic cestode species was firstly collected by Rennie and Reid (1912) during Scotia Scottish National Antarctic expedition (1902–1904) and described as Dibothriocephalus scoticus from leopard seals (Hydrurga leptonyx) and later renamed as Diphyllobothrium scoticum by Meggitt (1924). Most detailed descriptions are given by Fuhrmann (1921) from collections of Scotia, Markowski (1952b) from several worms found in four leopard seals of British Graham Land expedition (1934–37) and Yurakhno and Maltsev (1994) with data summarized from 21 specimens from leopard seals of Balleny island. Johnston (1937) also found these cestodes in sea leopards in the Aurora collection of the Australasian Antarctic expedition (1911–14) but did not provide a further description. Uterine sac is a typical species-specific feature of this species but larger than in species D. lobodoni from crabeater seal (Lobodon carcinophagus) of the Pacific region of Antarctica.
Anisakidae gen. sp. eggs were also found at a high prevalence of 34.5%. Based on a reported high frequency of occurrence in South American sea lions (George-Nascimento and Carvajal, 1980; Hernández-Orts et al., 2013), these eggs most probably originated from the anthropozoonotic ascarid nematodes Contracaecum, Pseudoterranova, or Anisakis. These marine ascarids usually parasitize stomach and small intestine, either freely within the gastrointestinal lumen or firmly attached to gut mucosa thereby inducing focal inflammation (Young and Lowe, 1969; Geraci and St Aubin, 1987; Hermosilla et al., 2016b). Occasionally, deep mucosal penetration by larval or adult stages can lead to severe ulcers, gastritis, enteritis and even intestinal wall perforation (Young and Lowe, 1969; McClelland, 1980; Geraci and St Aubin, 1987). In humans, allergic reactions against Anisakis simplex major allergen were recently reported (Martinez-Aranguren et al., 2014; Garcia Alonso et al., 2015).
Third most prevalent parasite found in sea lions was Balantidium (13.8%). Within genus Balantidium, B. coli is the only species nowadays considered as pathogenic for diverse terrestrial mammals including humans (Ponce-Gordo et al., 2011; Hassell et al., 2013; Yin et al., 2015; Zanzani et al., 2016). Balantidium infections were recently recorded in free-swimming fin whales from Azores Archipelago, Portugal (Hermosilla et al., 2016a), and in wild South American sea lions in the city of Valdivia, Chile (Hermosilla et al., 2016b), indicating that Balantidium is most probably circulating in different oceanographic areas. Interestingly, B. coli infections were also reported in Chilean humans and pigs (Palomino and Donckaster, 1971; Letonja et al., 1975).
Referring to current bacterial findings, Mycobacterium pinnipedii (tuberculosis), Edwardsiella tarda and Clostridium sp. infections were already demonstrated to circulate in wild populations of South American sea lions (Konagaya et al., 2006; de Amorim et al., 2014; Fernandez et al., 2014). Clostridium spp., especially C. perfringens, was one of most abundant bacteria isolated in this study. Clostridiales were identified as part of microbiota of different marine mammal species before assuming that they may be involved in chitin degradation (Konagaya et al., 2006; Greig et al., 2014; Delport et al., 2016; Medeiros et al., 2016; Soverini et al., 2016). C. perfringens is also regarded as a common microorganism in marine environments (Miller et al., 2006). Overall, high abundance of Clostridiales might be due to environmental and dietary consistence. However, different case reports also indicate their pathogenic potential in marine mammals. As such, C. perfringens-septicemia in a stranded common dolphin in California (Danil et al., 2014), C. perfringens-induced colonic rupture in a captive Californian sea lion (Van Bonn, 1995) and cases of myositis and skin lesions in different marine mammals (Greenwood and Taylor, 1978; Buck et al., 1987; Lang et al., 2014) were documented. Furthermore, the potential of Clostridiales to causing severe wound infections in humans is also well known (Stevens et al., 2012).
Edwardsiella tarda, which was detected in 32.1% of current samples, was described as a constituent of marine mammal microbiota (González-Fuentes et al., 2010). There is no data on its pathogenic potential for marine mammals, but E. tarda worldwide is an economically important fish pathogen (Park et al., 2012). As zoonotic opportunistic pathogen, it might causes fatal human water- or food-borne infections leading to gastroenteritis, endocarditis, empyema, hepatobiliary infections, peritonitis, intra-abdominal abscesses, osteomyelitis, wound infections, meningitis, or bacteremia (Hirai et al., 2015).
In line to other marine mammal species (Buck et al., 2006; Lockwood et al., 2006; Schaefer et al., 2009; Greig et al., 2014; Delport et al., 2016), most abundant bacterial species in this study was E. coli. This bacteria forms part of physiological intestinal microbiota (Greig et al., 2014) but is also described as an opportunistic pathogen causing septicemia (Steiger et al., 1989; Higgins, 2000; Carrasco et al., 2011) and wound or umbilical infections (Lockwood et al., 2006; Lang et al., 2014). Pathogenic E. coli O157:H7 serotypes were not yet detected in marine mammals but opportunistic water-borne human infections have to be considered.
As zoonotic pathogens, Salmonella serotypes (serotype Cerro and Pensacola) were isolated from six animals (21.4%) in this study. Several Salmonella serotypes from a variety of marine mammal species were described before (Gilmartin et al., 1979; Thornton et al., 1998; Fenwick et al., 2004; Stoddard et al., 2008; Davison N. et al., 2010; Davison N. J. et al., 2010; Carrasco et al., 2011; Berardi et al., 2014; Baily et al., 2016), but serotype S. Cerro was only once reported in sea lions from New Zealand (Fenwick et al., 2004). Whilst serotype S. Cerro is frequently isolated from dairy cows, serotype Pensacola is infrequently described in human samples (Edwards et al., 1948). Past studies revealed that cross infections between marine and terrestrial mammals might occur, especially based on pinniped amphibious lifestyle in contrast to cetaceans. Thus, detection of same serotypes in humans and pinnipeds might result from shared coastal environments. Due to public health concerns, incidence of this pathogen should be monitored in future.
Another microorganism of zoonotic risk, Yersinia enterocolitica, was only detected in one animal. Infrequent isolation of this pathogen was also described in dolphins (Buck et al., 2006). Human infections are usually food-borne causing a wide range of clinical symptoms, commonly gastrointestinal disorders (Bancerz-Kisiel and Szweda, 2015). The marine bacteria Vibrio spp. were as well isolated from one animal in current study. Most members of this genus are non-pathogenic but a few species are able to cause wound infections or gastrointestinal disease in humans (Colwell, 2006). In line to other studies (Higgins, 2000; Buck et al., 2006; Lockwood et al., 2006; Morris et al., 2011), Pseudomonas spp. (other than P. aeruginosa) were also here detected. However, to reveal pathogenic potential of Y. enterocolitica-, Vibrio spp.-, and Pseudomonas spp.-isolates, further analyses on species identification, serotyping and molecular detection of virulence genes are necessary. Similarly, further investigations are required to reveal importance of all described bacteria in health status of marine mammals.
Conclusion
In conclusion, this survey adds data on new anthropozoonotic parasite and bacterial records to wild South American sea lions of Patagonia, Chile, and calls for more integrated research to avoid exposure of humans and pinnipeds to these circulating invasive pathogens. Regular monitoring programs are required to identify variations in incidence or prevalence of pathogens, in order to detect on time emergent diseases before observing consequences at population level. It will be of particular importance not only to include national authorities for public health issues but also biologists/ecologists responsible for preservation of these threatened marine mammal species in South America.
Author Contributions
CH, EP-B, CE, SP, and AT designed the project, coordinated and planned field studies. VH and GF collected sea lion scat samples in Patagonia, Chile. CH, JH, LS, SS, and EP-B carried out parasitological- and bacteriological-analyses. JH performed molecular characterization and phylogeny analysis of Diphyllobothrium sp. 1. CH, JH, LS, EP-B, CE, and AT prepared the manuscript. All authors reviewed the manuscript.
Funding
This work was partially funded by the Institute of Parasitology (JLU Giessen), the Institute for Hygiene and Infectious Diseases of Animals (JLU Giessen), and Huinay Scientific Field Station (Chile).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer GP and handling Editor declared their shared affiliation at the time of the review.
Acknowledgments
We deeply acknowledge the Chilean National Service of Fishing and Aquaculture (SERNAPESCA) for allowing Huinay Scientific Field Station the collection of scat samples and sending of samples to Germany. This is number 164 of Huinay Scientific Field Station. We would like to express our gratitude to Agnes Mohr, Birgit Reinhardt, and Christine Henrich (Institute of Parasitology, JLU Giessen, Germany) for their technical assistance in coprological/molecular analyses. This study is dedicated to Prof. Dr. Vladimir Hermosilla, ecologist and former Director of the Chilean Antarctic Institute (INACH), who emphasized in protecting threatened marine mammal species as well as Antarctic marine waters of Chile.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2018.00459/full#supplementary-material
TABLE S1 | Data set of 18S and 28S sequences used for phylogenetic analysis (if not indicated, sequences refer to Waeschenbach et al., 2017).
References
Alonso, M. K., Crespo, E. A., Pedraza, S. N., Garcia, N. A., and Coscarella, M. A. (2000). Food habits of the South American sea lion, Otaria flavescens, off Patagonia, Argentina. Fish. Bull. 98, 250–263.
Anderson, R. C. (1992). Nematode Parasites of Vertebrates: Their Development and Transmission. Cambridge: CABI Pub.
Aznar, F. J., Hernandez-Orts, J., Suarez, A. A., Garcia-Varela, M., Raga, J. A., and Cappozzo, H. L. (2012). Assessing host-parasite specificity through coprological analysis: a case study with species of Corynosoma (Acanthocephala: Polymorphidae) from marine mammals. J. Helminthol. 86, 156–164. doi: 10.1017/s0022149x11000149
Baer, J. G., Miranda, H., Fernandez, W., and Medina, J. (1967). Human diphyllobothriasis in Peru. Z. Parasitenkd. 28, 277–289. doi: 10.1007/BF00260267
Baily, J. L., Foster, G., Brown, D., Davison, N. J., Coia, J. E., Watson, E., et al. (2016). Salmonella infection in grey seals (Halichoerus grypus), a marine mammal sentinel species: pathogenicity and molecular typing of Salmonella strains compared with human and livestock isolates. Environ. Microbiol. 18, 1078–1087. doi: 10.1111/1462-2920.13219
Bancerz-Kisiel, A., and Szweda, W. (2015). Yersiniosis - a zoonotic foodborne disease of relevance to public health. Ann. Agric. Environ. Med. 22, 397–402. doi: 10.5604/12321966.1167700
Baylis, H. A., and Hamilton, J. E. (1934). The southern sea lion, Otaria byronia (De Blainville). Discovery Rep. 8, 306.
Berardi, T., Shapiro, K., Byrne, B. A., and Miller, W. (2014). Prevalence and characterization of Salmonella shed by captive and free-range California sea lions (Zalophus californianus) from a rehabilitation center and three state reserves along the California coast. J. Zoo. Wildl. Med. 45, 527–533. doi: 10.1638/2013-0197r1.1
Bik, E. M., Costello, E. K., Switzer, A. D., Callahan, B. J., Holmes, S. P., Wells, R. S., et al. (2016). Marine mammals harbor unique microbiotas shaped by and yet distinct from the sea. Nat. Commun. 7:10516. doi: 10.1038/ncomms10516
Bisping, W., and Amtsberg, G. (1988). Colour Atlas for the Diagnosis of Bacterial Pathogens in Animals. Hoboken, NJ: John Wiley & Sons, Limited.
Buck, J. D., Shepard, L. L., and Spotte, S. (1987). Clostridium perfringens as the cause of death of a captive Atlantic bottlenosed dolphin (Tursiops truncatus). J. Wildl. Dis. 23, 488–491. doi: 10.7589/0090-3558-23.3.488
Buck, J. D., Wells, R. S., Rhinehart, H. L., and Hansen, L. J. (2006). Aerobic microorganisms associated with free-ranging bottlenose dolphins in coastal Gulf of Mexico and Atlantic Ocean waters. J. Wildl. Dis. 42, 536–544. doi: 10.7589/0090-3558-42.3.536
Carrasco, S. E., Burek, K. A., Beckmen, K. B., Oaks, J. L., Davis, M. A., Baker, K. N., et al. (2011). Aerobic oral and rectal bacteria of free-ranging Steller sea lion pups and juveniles (Eumetopias jubatus) in Alaska. J. Wildl. Dis. 47, 807–820. doi: 10.7589/0090-3558-47.4.807
Carter, G. R., Roberts, A. W., and Chengappa, M. M. (1995). Essentials of Veterinary Microbiology. Baltimore: Williams & Wilkins.
Cattan, P. E., Babero, B. B., and Torres, D. (1976). The helminth fauna of Chile: IV. Nematodes of the genera Anisakis Dujardin, 1845 and Phocanema Myers, 1954 in relation with gastric ulcers in a South American Sea Lion, Otaria byronia. J. Wildl. Dis. 12, 511–515. doi: 10.7589/0090-3558-12.4.511
Chai, J. Y., Darwin Murrell, K., and Lymbery, A. J. (2005). Fish-borne parasitic zoonoses: status and issues. Int. J. Parasitol. 35, 1233–1254. doi: 10.1016/j.ijpara.2005.07.013
Colwell, R. (2006). “A global and historical perspective of the genus Vibrio,” in The biology of Vibrios, eds F. Thompson, B. Austin, and J. Swings (Washington, DC: ASM Press), 3–26.
Crespo, E. A. (1988). Dinámica Poblacional del lobo Marino del sur Otaria Flavescens (Shaw, 1800) en el norte del litoral patagónico. Ph.D. thesis, Universidad Nacional de Buenos Aires, ıBuenos Aires.
Crespo, E. A., Pedraza, S. N., Dans, S. L., Koen-Alonzo, M., Reyes, L. M., García, N. A., et al. (1997). Direct and indirect effects of the highseas fisheries on the marine mammal populations in the northern and central Patagonian coast. J. Northwest Atl. Fish. Sci. 22, 189–207. doi: 10.2960/J.v22.a15
Crespo, E. A., Schiavini, A. C. M., Pérez, F. H., and Cappozzo, H. L. (1999). “Distribution, abundance and seasonal changes of South American fur seals, Arctocephalus australis, along the coasts of Argentina,” in Proceedings of the 13th Annual Conference of the European Cetacean Society, eds P. G. H. Evans and J. A. Raga (Valencia: European Cetacean Society), 26–27.
Curtis, M. A., and Bylund, G. (1991). Diphyllobothriasis: fish tapeworm disease in the circumpolar north. Arctic Med. Res. 50, 18–24.
Dailey, M. D. (1975). The distribution and intraspecific variation of helminth parasites in pinnipeds. Rapports et Proces-verbaux des Réunions Conseil International pour l’Éxplotation de la Mer 169, 338–352.
Danil, K., St Leger, J. A., Dennison, S., Bernaldo de Quiros, Y., Scadeng, M., Nilson, E., et al. (2014). Clostridium perfringens septicemia in a long-beaked common dolphin Delphinus capensis: an etiology of gas bubble accumulation in cetaceans. Dis. Aquat. Organ. 111, 183–190. doi: 10.3354/dao02783
Davison, N., Barnett, J., Rule, B., Chappell, S., and Wise, G. (2010). Group B Salmonella in lungworms from a harbour porpoise (Phocoena phocoena). Vet. Rec. 167, 351–352. doi: 10.1136/vr.c4495
Davison, N. J., Simpson, V. R., Chappell, S., Monies, R. J., Stubberfield, E. J., Koylass, M., et al. (2010). Prevalence of a host-adapted group B Salmonella enterica in harbour porpoises (Phocoena phocoena) from the south-west coast of England. Vet. Rec. 167, 173–176. doi: 10.1136/vr.c3760
de Amorim, D. B., Casagrande, R. A., Alievi, M. M., Wouters, F., De Oliveira, L. G., Driemeier, D., et al. (2014). Mycobacterium pinnipedii in a stranded South American sea lion (Otaria byronia) in Brazil. J. Wildl. Dis. 50, 419–422. doi: 10.7589/2013-05-124
de Vos, A., Faux, C. E., Marthick, J., Dickinson, J., and Jarman, S. N. (2018). New determination of prey and parasite species for Northern Indian Ocean Blue Whales. Front. Mar. Sci. 5:104. doi: 10.3389/fmars.2018.00104
Delport, T. C., Power, M. L., Harcourt, R. G., Webster, K. N., and Tetu, S. G. (2016). Colony location and captivity influence the gut microbial community composition of the Australian Sea Lion (Neophoca cinerea). Appl. Environ. Microbiol. 82, 3440–3449. doi: 10.1128/aem.00192-16
Delyamure, S. L., Skryabin, A. S., and Serdiukov, A. M. (1985). Diphyllobothriata—flatworm parasites of man, mammals and birds. Princ. Cestodol. 9, 1–200.
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny. fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36(Suppl. 2), W465–W469. doi: 10.1093/nar/gkn180
Edwards, P. R., Bruner, D. W., and Moran, A. B. (1948). Further studies on the occurrence and distribution of Salmonella types in the United States. J. Infect. Dis. 83, 220–231. doi: 10.1093/infdis/83.3.220
Felix, J. (2013). Reported Incidences of Parasitic Infections in Marine Mammals from 1892 to 1978. Morrisville, NC: Lulu Press.
Fenwick, S. G., Duignan, P. J., Nicol, C. M., Leyland, M. J., and Hunter, J. E. (2004). A comparison of Salmonella serotypes isolated from New Zealand sea lions and feral pigs on the Auckland Islands by pulsed-field gel electrophoresis. J. Wildl. Dis. 40, 566–570. doi: 10.7589/0090-3558-40.3.566
Fernandez, A., Villanueva, M. P., Gonzalez, M., Fernandez, F., Latif, F., Flores, S. N., et al. (2014). Adhesive and invasive capacities of Edwardsiella tarda isolated from South American sea lion. Braz. J. Microbiol. 45, 1095–1099. doi: 10.1590/S1517-83822014000300044
Fuhrmann, O. (1921). Die Cestoden der Deutschen Südpolar Expedition 1901–1903. Deutsche Südpolar Expedition, 1901–1903 (1920), 16. Zoologie 8, 469–524.
Garcia Alonso, M., Caballero, M. L., Umpierrez, A., Lluch-Bernal, M., Knaute, T., and Rodriguez-Perez, R. (2015). Relationships between T cell and IgE/IgG4 epitopes of the Anisakis simplex major allergen Ani s 1. Clin. Exp. Allergy 45, 994–1005. doi: 10.1111/cea.12474
George-Nascimento, M., and Carvajal, J. (1980). [New records of anisakid nematodes from Chilean marine fauna]. Bol. Chil. Parasitol. 35, 15–18.
George-Nascimento, M., and Carvajal, J. (1981). [Helminth parasites of the South American sea lion Otaria flavescens from the Gulf of Arauco. Chile]. Bol. Chil. Parasitol. 36, 72–73.
Georgiev, B. B., Biserkov, V., and Genov, T. (1986). In toto staining method for cestodes in acetocarmine. Helminthologia 23, 279–281.
Geraci, J. R., and St Aubin, D. J. (1987). Effects of parasites on marine mammals. Int. J. Parasitol. 17, 407–414. doi: 10.1016/0020-7519(87)90116-0
Gilmartin, W. G., Vainik, P. M., and Neill, V. M. (1979). Salmonellae in feral pinnipeds off the Southern California coast. J. Wildl. Dis. 15, 511–514. doi: 10.7589/0090-3558-15.4.511
Glad, T., Kristiansen, V. F., Nielsen, K. M., Brusetti, L., Wright, A. D., and Sundset, M. A. (2010). Ecological characterisation of the colonic microbiota in arctic and sub-arctic seals. Microb. Ecol. 60, 320–330. doi: 10.1007/s00248-010-9690-x
González-Fuentes, M., Latif, F., Fernández, F., Villanueva, M. P., Ulloa, J., and Fernández, H. (2010). Species of the family Enterobacteriaceae in feces of South American sea lion, Otaria flavescens, settled in the Valdivia River. Rev. Biol. Mar. Oceanogr. 45, 331–334.
Greenwood, A. G., and Taylor, D. C. (1978). Clostridial myositis in marine mammals. Vet. Rec. 103, 54–55. doi: 10.1136/vr.103.3.54
Greig, D. J., Gulland, F. M., Smith, W. A., Conrad, P. A., Field, C. L., Fleetwood, M., et al. (2014). Surveillance for zoonotic and selected pathogens in harbor seals Phoca vitulina from central California. Dis. Aquat. Organ. 111, 93–106. doi: 10.3354/dao02762
Hassell, J. M., Blake, D. P., Cranfield, M. R., Ramer, J., Hogan, J. N., Noheli, J. B., et al. (2013). Occurrence and molecular analysis of Balantidium coli in mountain gorilla (Gorilla beringei beringei) in the Volcanoes National Park, Rwanda. J. Wildl. Dis. 49, 1063–1065. doi: 10.7589/2013-02-041
Haussermann, V., Forsterra, G., and Plotnek, E. (2014). Sightings of marine mammals and birds in the Comau Fjord, Northern Patagonia, between 2003 and mid 2012. Spixiana 35, 247–262.
Hebert, P. D. N., Ratnasingham, S., and de Waard, J. R. (2003). Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc. Biol. Sci. 270, S96–S99. doi: 10.1098/rsbl.2003.0025
Heine, J. (1982). An easy technique for the demonstration of cryptosporidia in feces. J. Vet. Med. B Infect. Dis. Vet. Public Health 29, 324–327.
Hermosilla, C., Hirzmann, J., Silva, L. M. R., Brotons, J. M., Cerdà, M., Prenger-Berninghoff, E., et al. (2018). Occurrence of anthropozoonotic parasitic infections and faecal microbes in free-ranging sperm whales (Physeter macrocephalus) from the Mediterranean Sea. Parasitol. Res. 117, 2531–2541. doi: 10.1007/s00436-018-5942-3
Hermosilla, C., Navarro, M., and Taubert, A. (2013). “Gastrointestinal endoparasite fauna of ‘urban’ sea lions (Otaria flavenscens) in Valdivia, Chile (in German),” in Tagung der DVG-Fachgruppe “Parasitologie uns Parasitäre Krankheiten”, ed. D. V. Gesellschaft (Germany: Giessen), 134.
Hermosilla, C., Silva, L. M., Kleinertz, S., Prieto, R., Silva, M. A., and Taubert, A. (2016a). Endoparasite survey of free-swimming baleen whales (Balaenoptera musculus, B. physalus, B. borealis) and sperm whales (Physeter macrocephalus) using non/minimally invasive methods. Parasitol. Res. 115, 889–896. doi: 10.1007/s00436-015-4835-y
Hermosilla, C., Silva, L. M., Navarro, M., and Taubert, A. (2016b). Anthropozoonotic Endoparasites in Free-Ranging “Urban” South American Sea Lions (Otaria flavescens). J. Vet. Med. 2016:7507145. doi: 10.1155/2016/7507145
Hermosilla, C., Silva, L. M., Prieto, R., Kleinertz, S., Taubert, A., and Silva, M. A. (2015). Endo- and ectoparasites of large whales (Cetartiodactyla: Balaenopteridae, Physeteridae): overcoming difficulties in obtaining appropriate samples by non- and minimally-invasive methods. Int. J. Parasitol. Parasites Wildl. 4, 414–420. doi: 10.1016/j.ijppaw.2015.11.002
Hernández-Orts, J. S., Montero, F. E., Juan-Garcia, A., Garcia, N. A., Crespo, E. A., Raga, J. A., et al. (2013). Intestinal helminth fauna of the South American sea lion Otaria flavescens and fur seal Arctocephalus australis from northern Patagonia. Argentina J. Helminthol. 87, 336–347. doi: 10.1017/s0022149x12000454
Hernández-Orts, J. S., Montero, F. E., Leonardi, M. S., Raga, J. A., Aznar, F. J., and Crespo, E. A. (2011). Intestinal helminths from the South American sea lion, Otaria flavescens, from North Patagonia, Argentina. J. Helminthol. 87, 336–347. doi: 10.1017/S0022149X12000454
Hernández-Orts, J. S., Scholz, T., Brabec, J., Kuzmina, T., and Kuchta, R. (2015). High morphological plasticity and global geographical distribution of the Pacific broad tapeworm Adenocephalus pacificus (syn. Diphyllobothrium pacificum): Molecular and morphological survey. Acta Trop. 149, 168–178. doi: 10.1016/j.actatropica.2015.05.017
Hernandez-Orts, J. S., Viola, M. N. P., Garcia, N. A., Crespo, E. A., Gonzalez, R., Garcia-Varela, M., et al. (2015). A checklist of the helminth parasites of marine mammals from Argentina. Zootaxa 3936, 301–334. doi: 10.11646/zootaxa.3936.3.1
Hirai, Y., Asahata-Tago, S., Ainoda, Y., Fujita, T., and Kikuchi, K. (2015). Edwardsiella tarda bacteremia. A rare but fatal water- and foodborne infection: review of the literature and clinical cases from a single centre. Can. J. Infect. Dis. Med. Microbiol. 26, 313–318. doi: 10.1155/2015/702615
Hunt, K. E., Moore, M. J., Rolland, R. M., Kellar, N. M., Hall, A. J., Kershaw, J., et al. (2013). Overcoming the challenges of studying conservation physiology in large whales: a review of available methods. Conserv. Physiol. 1:cot006. doi: 10.1093/conphys/cot006
Johnston, T. H. (1937). “Cestoda,” in Australasian Antarctic expedition 1911–14, ed. T.H. Johnston. Sci Rep. Ser C Zool. Bot. 10, 15–74.
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kleinertz, S., Hermosilla, C., Ziltener, A., Kreicker, S., Hirzmann, J., Abdel-Ghaffar, F., et al. (2014). Gastrointestinal parasites of free-living Indo-Pacific bottlenose dolphins (Tursiops aduncus) in the Northern Red Sea. Egypt. Parasitol. Res. 113, 1405–1415. doi: 10.1007/s00436-014-3781-4
Konagaya, Y., Tsuchiya, C., and Sugita, H. (2006). Purification and characterization of chitinases from Clostridium sp. E-16 isolated from the intestinal tract of the South American sea lion (Otaria flavescens). Lett. Appl. Microbiol. 43, 187–193. doi: 10.1111/j.1472-765X.2006.01926.x
Kuchta, R., and Scholz, T. (2017). “Diphyllobothriidea,” in Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth, eds J. N. Caira and K. Jensen (Lawrence, KS: University of Kansas),167–189.
Kuchta, R., Serrano-Martínez, M. E., and Scholz, T. (2015). Pacific broad tapeworm Adenocephalus pacificus as a causative agent of globally reemerging diphyllobothriosis. Emerg. Infect. Dis. 21:1697. doi: 10.3201/eid2110.150516
Lang, B., Eskens, U., Hartmann, A., Braun, V., Schmidt, M. J., and von Hegel, G. (2014). [Focal meningitis in a California sea lion (Zalophus californianus)-diagnostic procedure and pathology]. Tierarztl. Prax. Ausg. G. Grosstiere Nutztiere 42, 368–372. doi: 10.15653/tpg-140301
Leštinová, K., Soldánová, M., Scholz, T., and Kuchta, R. (2016). Eggs as a suitable tool for species diagnosis of causative agents of human diphyllobothriosis (Cestoda). PLoS Negl. Trop. Dis. 10:e0004721. doi: 10.1371/journal.pntd.0004721
Letonja, T., Henriquez, A., Reyes, G., and Zapata, L. (1975). Prevalence of Balantidium coli infections in swine from Santiago, Chile (author’s transl). Bol. Chil. Parasitol. 30, 88–89.
Lima, M., and Páez, E. (1997). Demography and population dynamics of South American fur seals. J. Mammal. 78, 914–920. doi: 10.2307/1382951
Lockwood, S. K., Chovan, J. L., and Gaydos, J. K. (2006). Aerobic bacterial isolations from harbor seals (Phoca vitulina) stranded in Washington: 1992-2003. J. Zoo Wildl. Med. 37, 281–291. doi: 10.1638/05-035.1
Lockyer, A. E., Olson, P. D., and Littlewood, D. T. J. (2003). Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): implications and a review of the cercomer theory. Biol. J. Linn. Soc. 78, 155–171. doi: 10.1046/j.1095-8312.2003.00141.x
Markowski, S. (1952a). The cestodes of pinnipeds in the Arctic and other regions. J. Helminthol. 26, 171–214.
Markowski, S. (1952b). The cestodes of seals from the Antarctic. Bull. Br. Mus. Nat. Hist. D. Zool. 1, 123–150. doi: 10.5962/bhl.part.21631
Martinez-Aranguren, R. M., Gamboa, P. M., Garcia-Lirio, E., Asturias, J., Goikoetxea, M. J., and Sanz, M. L. (2014). In vivo and in vitro testing with rAni s 1 can facilitate diagnosis of Anisakis simplex allergy. J. Investig. Allergol. Clin. Immunol. 24, 431–438.
McClelland, G. (1980). Phocanema decipiens: pathology in seals. Exp. Parasitol. 49, 405–419. doi: 10.1016/0014-4894(80)90075-2
Medeiros, A. W., Giongo, A., Valdez, F. P., Blaese de Amorin, D., Tavares, M., d’Azevedo, P. A., et al. (2016). Characterization of the faecal bacterial community of wild young South American (Arctocephalus australis) and Subantarctic fur seals (Arctocephalus tropicalis). FEMS Microbiol. Ecol. 92:fiw029. doi: 10.1093/femsec/fiw029
Meggitt, F. J. (1924). The Cestodes of Mammals. Available at: https://www.cabdirect.org/cabdirect/abstract/19272902351
Mercado, R., Yamasaki, H., Kato, M., Muñoz, V., Sagua, H., Torres, P., et al. (2010). Molecular identification of the Diphyllobothrium species causing diphyllobothriasis in Chilean patients. Parasitol. Res. 106, 995–1000. doi: 10.1007/s00436-010-1765-6
Miller, W. A., Miller, M. A., Gardner, I. A., Atwill, E. R., Byrne, B. A., Jang, S., et al. (2006). Salmonella spp., Vibrio spp., Clostridium perfringens, and Plesiomonas shigelloides in marine and freshwater invertebrates from coastal California ecosystems. Microb. Ecol. 52, 198–206. doi: 10.1007/s00248-006-9080-6
Morris, P. J., Johnson, W. R., Pisani, J., Bossart, G. D., Adams, J., Reif, J. S., et al. (2011). Isolation of culturable microorganisms from free-ranging bottlenose dolphins (Tursiops truncatus) from the southeastern United States. Vet. Microbiol. 148, 440–447. doi: 10.1016/j.vetmic.2010.08.025
Naya, D. E., Arim, M., and Vargas, R. (2002). Diet of South American fur seals (Arctocephalus australis) in Isla de Lobos, Uruguay. Mar. Mammal. Sci. 18, 734–745. doi: 10.1111/j.1748-7692.2002.tb01070.x
Negi, A., Puri, A., Gupta, R., Chauhan, I., Nangia, R., and Sachdeva, A. (2013). Biosafe alternative to xylene: a comparative study. J. Oral Maxillofac. Pathol. 17, 363–366. doi: 10.4103/0973-029x.125199
Nunn, G. B. (1992). Nematode Molecular Evolution : An Investigation of Evolutionary Patterns Among Nematodes Based Upon DNA Sequences. Ph.D. thesis, University of Nottingham, Nottingham.
Ohno, H. (2015). Impact of commensal microbiota on the host pathophysiology: focusing on immunity and inflammation. Semin. Immunopathol. 37, 1–3. doi: 10.1007/s00281-014-0472-2
Palomino, H., and Donckaster, R. (1971). Clinical and epidemiological study of a case of human balantidiasis. Bol. Chil. Parasitol. 26, 44–45.
Park, S. B., Aoki, T., and Jung, T. S. (2012). Pathogenesis of and strategies for preventing Edwardsiella tarda infection in fish. Vet. Res. 43:67. doi: 10.1186/1297-9716-43-67
Pereira, E. M., Muller, G., Secchi, E., Pereira, J. Jr., and Valente, A. L. (2013). Digenetic trematodes in South American sea lions from southern Brazilian waters. J. Parasitol. 99, 910–913. doi: 10.1645/ge-3216.1
Ponce-Gordo, F., Fonseca-Salamanca, F., and Martinez-Diaz, R. A. (2011). Genetic heterogeneity in internal transcribed spacer genes of Balantidium coli (Litostomatea. Ciliophora). Protist 162, 774–794. doi: 10.1016/j.protis.2011.06.008
Porazinska, D. L., Giblin-Davis, R. M., Faller, L., Farmerie, W., Kanzaki, N., Morris, K., et al. (2009). Evaluating high-throughput sequencing as a method for metagenomic analysis of nematode diversity. Mol. Ecol. Resour. 9, 1439–1450. doi: 10.1111/j.1755-0998.2009.02611.x
Rennie, J., and Reid, A. (1912). XXII.—The Cestoda of the Scottish National Antarctic Expedition. Earth Env. Sci. T. R. Soc. 48, 441–453. doi: 10.1017/S0080456800002945
Schaefer, A. M., Goldstein, J. D., Reif, J. S., Fair, P. A., and Bossart, G. D. (2009). Antibiotic-resistant organisms cultured from Atlantic bottlenose dolphins (Tursiops truncatus) inhabiting estuarine waters of Charleston, SC and Indian River Lagoon, FL. Ecohealth 6, 33–41. doi: 10.1007/s10393-009-0221-5
Scholz, T., Garcia, H. H., Kuchta, R., and Wicht, B. (2009). Update on the human broad tapeworm (genus Diphyllobothrium), including clinical relevance. Clin. Microbiol. Rev. 22, 146–160. doi: 10.1128/cmr.00033-08
Soverini, M., Quercia, S., Biancani, B., Furlati, S., Turroni, S., Biagi, E., et al. (2016). The bottlenose dolphin (Tursiops truncatus) faecal microbiota. FEMS Microbiol. Ecol. 92:fiw055. doi: 10.1093/femsec/fiw055
Steiger, G. H., Calambokidis, J., Cubbage, J. C., Skilling, D. E., Smith, A. W., and Gribble, D. H. (1989). Mortality of harbor seal pups at different sites in the inland waters of Washington. J. Wildl. Dis. 25, 319–328. doi: 10.7589/0090-3558-25.3.319
Stevens, D. L., Aldape, M. J., and Bryant, A. E. (2012). Life-threatening clostridial infections. Anaerobe 18, 254–259. doi: 10.1016/j.anaerobe.2011.11.001
Stoddard, R. A., DeLong, R. L., Byrne, B. A., Jang, S., and Gulland, F. M. (2008). Prevalence and characterization of Salmonella spp. among marine animals in the Channel Islands, California. Dis. Aquat. Organ. 81, 5–11. doi: 10.3354/dao01905
Suarez, A. A., Sanfelice, D., Cassini, M. H., and Cappozzo, H. L. (2005). Composition and seasonal variation in the diet of the South American sea lion (Otaria flavescens) from Quequén, Argentina. Lat. Am. J. Aquat. Mamm. 42, 163–174. doi: 10.5597/lajam00079
Thornton, S. M., Nolan, S., and Gulland, F. M. (1998). Bacterial isolates from California sea lions (Zalophus californianus), harbor seals (Phoca vitulina), and northern elephant seals (Mirounga angustirostris) admitted to a rehabilitation center along the central California coast, 1994-1995. J. Zoo Wildl. Med. 29, 171–176.
Torres, P., Leyan, V., and Puga, S. (2012). Prevalence, intensity, and abundance of infection and pathogenesis caused by diphyllobothriosis in vulnerable, native fish and introduced trout in Lake Panguipulli. Chile. J. Wildl. Dis. 48, 937–950. doi: 10.7589/2011-08-235
Túnez, J. I., Cappozzo, H. L., and Cassini, M. H. (2008). Regional factors associated with the distribution of South American fur seals along the Atlantic coast of South America. Ices J. Mar. Sci. 65, 1733–1738. doi: 10.1093/icesjms/fsn168
Van Steenkiste, N., Locke, S. A., Castelin, M., Marcogliese, D. J., and Abbott, C. L. (2015). New primers for DNA barcoding of digeneans and cestodes (Platyhelminthes). Mol. Ecol. Resour. 15, 945–952. doi: 10.1111/1755-0998.12358
Vaz-Ferreira, R. (1982). “Otaria flavescens (Shaw), South American sea lion,” in Mammals of the world, (Rome: FAO Fisheries Series 5), 447–495.
Vilas, R., Criscione, C. D., and Blouin, M. S. (2005). A comparison between mitochondrial DNA and the ribosomal internal transcribed regions in prospecting for cryptic species of platyhelminth parasites. Parasitology 131, 839–846. doi: 10.1017/S0031182005008437
Waeschenbach, A., Brabec, J., Scholz, T., Littlewood, D. T. J., and Kuchta, R. (2017). The catholic taste of broad tapeworms - multiple routes to human infection. Int. J. Parasitol. 47, 831–843. doi: 10.1016/j.ijpara.2017.06.004
Waeschenbach, A., Webster, B. L., Bray, R. A., and Littlewood, D. T. (2007). Added resolution among ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with complete small and large subunit nuclear ribosomal RNA genes. Mol. Phylogenet. Evol. 45, 311–325. doi: 10.1016/j.ympev.2007.03.019
Wicht, B., Yanagida, T., Scholz, T., Ito, A., Jiménez, J. A., and Brabec, J. (2010). Multiplex PCR for differential identification of broad tapeworms (Cestoda: Diphyllobothrium) infecting humans. J. Clin. Microbiol. 48, 3111–3116. doi: 10.1128/JCM.00445-10
Yamasaki, H., Kumazawa, H., Sekikawa, Y., Oda, R., Hongo, I., Tsuchida, T., et al. (2016). First confirmed human case of Diphyllobothrium stemmacephalum infection and molecular verification of the synonymy of Diphyllobothrium yonagoense with D. stemmacephalum (Cestoda: Diphyllobothriidea). Parasitol. Int. 65, 412–421. doi: 10.1016/j.parint.2016.06.003
Yang, J., and Scholten, T. (1977). A fixative for intestinal parasites permitting the use of concentration and permanent staining procedures. Am. J. Clin. Pathol. 67, 300–304. doi: 10.1093/ajcp/67.3.300
Yin, D. M., Lv, C. C., Tan, L., Zhang, T. N., Yang, C. Z., Liu, Y., et al. (2015). Prevalence of Balantidium coli infection in sows in Hunan province, subtropical China. Trop. Anim. Health Prod. 47, 1637–1640. doi: 10.1007/s11250-015-0904-6
Young, P. C., and Lowe, D. (1969). Larval nematodes from fish of the subfamily Anisakinae and gastro-intestinal lesions in mammals. J. Comp. Pathol. 79, 301–313. doi: 10.1016/0021-9975(69)90043-7
Yurakhno, M. V., and Maltsev, V. N. (1994). Diphyllobothrium lobodoni sp. n.(Cestoda: Diphyllobothriidae)–a parasite of the crabeater seal. Parazitologiia 28, 270–275.
Yurakhno, M. V., and Maltsev, V. N. (1997). An infection of seals from Antarctica with cestodes. Parazitologiia 31, 81–89.
Keywords: Otaria flavescens, Diphyllobothrium scoticum, Balantidium, Entamoeba, Clostridium
Citation: Hermosilla C, Hirzmann J, Silva LMR, Scheufen S, Prenger-Berninghoff E, Ewers C, Häussermann V, Försterra G, Poppert S and Taubert A (2018) Gastrointestinal Parasites and Bacteria in Free-Living South American Sea Lions (Otaria flavescens) in Chilean Comau Fjord and New Host Record of a Diphyllobothrium scoticum-Like Cestode. Front. Mar. Sci. 5:459. doi: 10.3389/fmars.2018.00459
Received: 27 June 2018; Accepted: 14 November 2018;
Published: 13 December 2018.
Edited by:
Francesca Carella, Università degli Studi di Napoli Federico II, ItalyReviewed by:
Amy C. Hirons, Nova Southeastern University, United StatesVanessa Labrada Martagón, Universidad Autónoma de San Luis Potosí, Mexico
Gianluca Polese, Università degli Studi di Napoli Federico II, Italy
Copyright © 2018 Hermosilla, Hirzmann, Silva, Scheufen, Prenger-Berninghoff, Ewers, Häussermann, Försterra, Poppert and Taubert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlos Hermosilla, Q2FybG9zLlIuSGVybW9zaWxsYUB2ZXRtZWQudW5pLWdpZXNzZW4uZGU=