
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mar. Sci., 16 November 2017
Sec. Deep-Sea Environments and Ecology
Volume 4 - 2017 | https://doi.org/10.3389/fmars.2017.00368
This article is part of the Research TopicAnthropogenic Disturbances in the Deep SeaView all 18 articles
 Chris Hauton1*
Chris Hauton1* Alastair Brown1
Alastair Brown1 Sven Thatje1
Sven Thatje1 Nélia C. Mestre2
Nélia C. Mestre2 Maria J. Bebianno2
Maria J. Bebianno2 Inês Martins3
Inês Martins3 Raul Bettencourt3
Raul Bettencourt3 Miquel Canals4
Miquel Canals4 Anna Sanchez-Vidal4
Anna Sanchez-Vidal4 Bruce Shillito5
Bruce Shillito5 Juliette Ravaux5
Juliette Ravaux5 Magali Zbinden5
Magali Zbinden5 Sébastien Duperron5
Sébastien Duperron5 Lisa Mevenkamp6
Lisa Mevenkamp6 Ann Vanreusel6
Ann Vanreusel6 Cristina Gambi7
Cristina Gambi7 Antonio Dell'Anno7
Antonio Dell'Anno7 Roberto Danovaro8Vikki Gunn9
Roberto Danovaro8Vikki Gunn9 Phil Weaver9
Phil Weaver9In January 2017, the International Seabed Authority released a discussion paper on the development of Environmental Regulations for deep-sea mining (DSM) within the Area Beyond National Jurisdiction (the “Area”). With the release of this paper, the prospect for commercial mining in the Area within the next decade has become very real. Moreover, within nations' Exclusive Economic Zones, the exploitation of deep-sea mineral ore resources could take place on very much shorter time scales and, indeed, may have already started. However, potentially toxic metal mixtures may be released at sea during different stages of the mining process and in different physical phases (dissolved or particulate). As toxicants, metals can disrupt organism physiology and performance, and therefore may impact whole populations, leading to ecosystem scale effects. A challenge to the prediction of toxicity is that deep-sea ore deposits include complex mixtures of minerals, including potentially toxic metals such as copper, cadmium, zinc, and lead, as well as rare earth elements. Whereas the individual toxicity of some of these dissolved metals has been established in laboratory studies, the complex and variable mineral composition of seabed resources makes the a priori prediction of the toxic risk of DSM extremely challenging. Furthermore, although extensive data quantify the toxicity of metals in solution in shallow-water organisms, these may not be representative of the toxicity in deep-sea organisms, which may differ biochemically and physiologically and which will experience those toxicants under conditions of low temperature, high hydrostatic pressure, and potentially altered pH. In this synthesis, we present a summation of recent advances in our understanding of the potential toxic impacts of metal exposure to deep-sea meio- to megafauna at low temperature and high pressure, and consider the limitation of deriving lethal limits based on the paradigm of exposure to single metals in solution. We consider the potential for long-term and far-field impacts to key benthic invertebrates, including the very real prospect of sub-lethal impacts and behavioral perturbation of exposed species. In conclusion, we advocate the adoption of an existing practical framework for characterizing bulk resource toxicity in advance of exploitation.
Whilst the technological challenges of mineral recovery from deep water are significant (e.g., Collins et al., 2013; Glasby et al., 2015), they are no longer regarded as insurmountable obstacles to the exploitation of seabed resources by deep-sea mining (DSM) (Hannington et al., 2017). Consequently, the start—and subsequent expansion—of DSM will soon become contingent on: (a) a viable economic assessment (see Petersen et al., 2016; Hannington et al., 2017) and, (b) an appropriate legislative framework.
Legislatively, under the UN Convention on the Law of the Sea (UNCLOS), legal responsibility for mineral exploitation in the “Area Beyond National Jurisdiction” (or the “Area”) rests with the International Seabed Authority (ISA). Early in 2017 the ISA published draft Environmental Regulations for deep-sea resource exploitation within the Area (ISA, 2017). This draft code advocates the Precautionary Principle in the initial pursuit of deep-sea mineral resources and a framework of Adaptive Management to monitor and regulate exploitation by licensed contractors. However, these two concepts present challenges for the ISA, as have been recently reviewed by Jaeckel (2016) and Le et al. (2017). These reviews have argued that key challenges remain with the identification of environmental risks of DSM, relevant to the application of the Precautionary Principle, and in the development of appropriate monitoring technologies with which to support the Adaptive Management of exploitation.
Recently, great progress has been made in constraining the toxic risks of mineral extraction under the unique conditions of high pressure and low temperature, as well as presenting these risks in an operationally appropriate manner to support decisions on exploitation licensing. At this key time for the development of DSM exploitation legislation, this synthesis paper provides a summation of the scientific community's latest understanding of the challenges in quantifying the absolute toxic risks of metal exposure to benthic organisms (meiofauna to megafauna). In part, this synthesis presents the key conclusions of the “Ecotoxicology” working group of the EC FP7 Project “Managing Impacts of Deep Sea Resource Exploitation” (MIDAS; https://www.eu-midas.net/), placed within the wider context offered by the research developments of other teams.
Deep-sea ore deposits comprise complex mixtures of potentially toxic elements including, for example: copper, cadmium, zinc, and lead, as well as rare earth elements, such as lanthanum and yttrium (Petersen et al., 2016). These ore deposits form in distinct geological settings, as has been extensively and recently reviewed (Petersen et al., 2016). In situ, the surface of these ore deposits are weathered and present minimal toxic risk. Indeed, nodules and massive sulfides can provide valuable substrate for a diverse faunal assemblage (e.g., Wang et al., 2013; Amon et al., 2016; Vanreusel et al., 2016). DSM, whether of seafloor massive sulfides (SMS), polymetallic nodules, or cobalt-rich crusts, may break up these ore deposits and may release toxic concentrations of metals into the environment at distinct phases of the mining cycle, affecting organisms within different marine compartments (Figure 1). Metals may be released in different physical phases (in solution, as ground particulates of different size and shape, or adsorbed onto the surface of particulates (see: Fallon et al., 2017 and references therein), which will affect their bioavailability to organisms. For example, metals in solution may be taken up across permeable tissues such as gill or gut surfaces, whilst particulate or adsorbed metals might be taken into the gut of deposit feeders. In the longer term, metal accumulation in tissues presents a further toxic risk for predatory species higher in the food chain, as they can consume contaminated prey items.
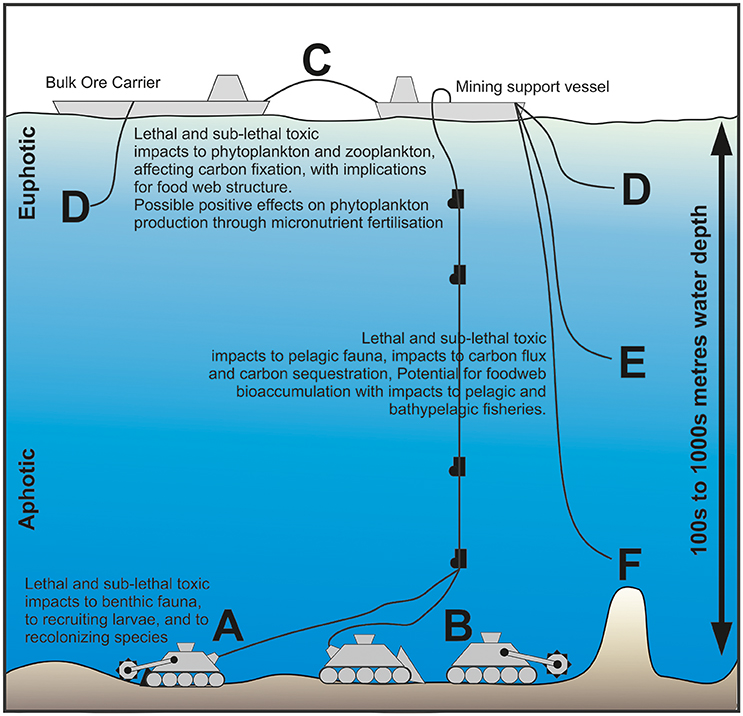
Figure 1. Schematic representation of potential deep-sea mining operations (not to scale), identifying stages at which toxic metals may be released to the environment. Metals may be released in particulate or dissolved phases associated with localized plumes from mining collectors (A), or individually controlled mining tools and collectors (B) operating in tandem. Dewatering ore slurry at the mining support vessel or transportation barge (C) may introduce toxic concentrations of metals into the euphotic zone, impacting photosynthetic primary producers or zooplankton communities and potentially affecting the sequestration of carbon dioxide in the surface ocean, as well as pelagic food webs driving the flux of carbon to the sea floor. Mid-water discharge (D,E), whilst away from the productive euphotic zone, may impact vertically migrating zooplankton—again altering the vertical flux of carbon to the sea floor. Whilst the discharge of water near the seabed (F) will limit mid-water impacts, the release of thermally altered and buoyant water may disperse contaminants over a much larger area than that immediately impacted by the mining tools.
It is expected that the mining of massive sulfides or cobalt-rich crusts will involve crushed ore being pumped as a slurry from the seafloor mining tool, or via a collector after stockpiling on the sea bed (Figure 1B), to the surface mining support vessel via a riser pipe (Hoagland et al., 2010). Whilst polymetallic nodules may be collected intact (Figure 1A), nodules in the Clarion-Clipperton Fracture Zone in the Pacific Ocean are friable and readily crumble with handling. Consequently, for all resources, there is a risk that extraction will release metals in dissolved and fine particulate phases into the water column as a plume over potential scales of 100–1,000 s of km2 (Oebius et al., 2001). Current operator plans include the transfer of bulk ore from the mining support vessel to a barge for transfer to shore and processing (Figure 1C) and this slurry will require dewatering before and/or after transfer (Hoagland et al., 2010). Dewatering the ore slurry at the surface may also release metals into the marine environment in the return water (Figure 1D). Releasing return water into the ocean at the surface or mid-depth (Figures 1D,E, respectively) will disperse potential toxicants widely (100–1,000 s of km) and may impact organisms within the water column. Return water, potentially at above-ambient temperature, that is discharged above the seafloor may disperse as a thermally buoyant plume over a large area, carrying toxicants with it (Figure 1F). The impacts of these potential metal releases must be understood for their potential to cause “serious harm” to deep sea marine ecosystems, the avoidance of which is a fundamental legal remit of the ISA (Levin et al., 2016).
There is increasing evidence of the need for incorporation of an Ecosystem Service (ES) approach into the environmental management of the deep sea, and specifically DSM (Danovaro et al., 2017; Le et al., 2017, respectively). Since the publication of the Millennium Ecosystem Assessment in 2005, the role of ecosystems in providing resources and services for the benefit of human kind has been recognized, and this is applicable to terrestrial as well as shallow and deep-sea ecosystems (Le et al., 2017). Consideration of ES incorporates a holistic view of species' distributions, abundance and diversity (Ecosystem Structures), as well measures of species' activity (Ecosystem Functions/Supporting Services) (Le et al., 2017; see also Figure 2).
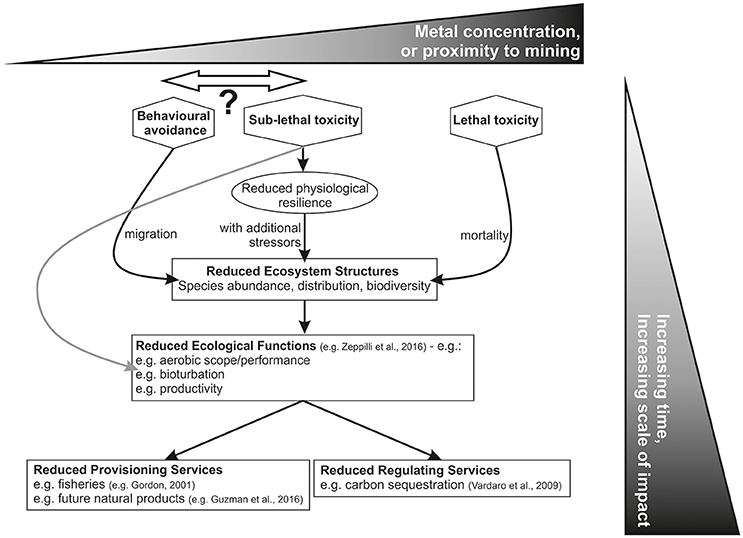
Figure 2. Representation of how behavioral, sub-lethal and lethal impacts of metal exposure on individual organisms can, over time or with increasing magnitude of the metal release, scale to produce impacts on Ecosystem Structures, Ecological Functions and thence to Ecosystem Services (adapted from Le et al., 2017). Shown from left to right is the effect of increasing metal concentration, or increased proximity to the metal release. Behavioral modifications or sub-lethal effects will occur at low metal concentrations or far from a mining site whilst lethal impacts at high metal concentrations or at locations close to metal release will occur. Sub-lethal exposure can result in a reduction in organism performance that cause direct impacts to Ecological Function (gray line) or, in combination with other physiological stressors (e.g., hypoxia, restricted diet, post-spawning) can result in organism mortality that cause direct impacts to Ecosystem Structures. Note that the relative positions and relative order of behavioral and sub-lethal effects is variable.
Whilst, our knowledge of deep sea ecosystems and the diversity of life that they contain has been rapidly growing in recent years (Ramirez-Llodra et al., 2010), it is clear that we have only just begun to constrain the true diversity of life throughout the deep ocean. Deep sea communities are typified by species with slow growth and delayed maturity; characteristics which make them particularly susceptible to anthropogenic impacts or other environmental perturbation. These joint considerations: a current paucity of available data on deep sea community structure and the life history characteristics of known deep sea species, mean that there is a high probability that, should large scale deep sea mineral exploitation be prosecuted in the near future, we may in fact irreversibly impact unique biological and genetic resources even before they have been identified (Thornburg et al., 2010; Harden-Davies, 2017).
Abyssal benthic environments support biologically diverse species assemblages of meio-, macro- and megafauna (note, microfauna are explicitly not considered in this synthesis). These biologically diverse communities are comprised of populations with low overall abundance as a result of the absent primary production over the vast majority of the abyssal sea floor, and a complete reliance upon periodic phytodetrital nutrient supply from the surface ocean thousands of meters away. This unique placement renders them energy limited (Smith et al., 2008), therefore restricting overall species abundance.
Nonetheless, recent studies in the Clarion Clipperton Fracture Zone have identified diverse epifaunal communities of filtering feeding organisms, including hexactinellid sponges, crinoids, hydroids, and soft corals, that are reliant on the nodules as substrate for attachment (Amon et al., 2016; Vanreusel et al., 2016). Dominant mobile megafauna associated with abyssal nodule fields include key benthic bioturbating groups, such as holothurians (sea cucumbers) and ophiuroids (brittlestars) (Amon et al., 2016).
In contrast, biological communities that colonize massive sulfides at hydrothermal vents are potentially supplied with a source of energy to support chemosynthetic primary production. These systems can also occur at shallower depths, receiving greater carbon flux from the surface ocean (Van Dover, 2000). As a result, active vent communities are normally comprised of a restricted diversity of fast-growing species that can occur at locally very high abundance. Faunal assemblages typically include crustaceans, molluscs and polychaete worms although with distinct biogeographic provinces (e.g., Van Dover et al., 2002), many of which indirectly rely on the vent fluid as an energy resource (Boschen et al., 2016; and references therein). Hydrothermal vent communities can show high endemism with high rates of speciation recorded; certain taxa being geographically restricted to a single, or small number of active vent sites (e.g., Wolff, 2005). As a consequence, whilst a single vent system might be seen as a productive and robust system, it may represent the only instance of that community within the deep ocean. Moreover, inactive, or relic, vents that might be first targeted for the exploitation of SMS, may support a more diverse faunal assemblage, including organisms that are not tolerant of vent conditions (e.g., relatively warmer waters and higher metal concentrations) or that do not rely on the vent fluids as a source of energy (Marsh et al., 2012; Sarrazin et al., 2015).
Water column, or pelagic communities, that might be impacted by plume release at mid-depths (Figures 1D,E; Ellis, 2001) mediate the transfer of energy and organic carbon from primary producers in the euphotic zone to the sea floor (Robison, 2009), but also from hydrothermal plumes on the deep sea floor (Phillips, 2017). Already studies have demonstrated that metals can exert a toxic effect to marine phyto- and zooplankton species (e.g., Hirota, 1981; Hu, 1981; Moraitouapostolopoulou and Verriopoulos, 1982; Caroppo et al., 2006; Fuchida et al., 2017) and can lead to metal accumulation (bioaccumulation) in higher trophic levels of food chains (Amiard Triquet et al., 1993). In contrast, others have noted that some metals (including for example: copper, zinc, iron) represent essential micronutrients and that long-term exposure to low concentrations might create positive effects in pelagic communities through the improved function of some planktonic species (e.g., Loka Bharathi et al., 2005).
The bathypelagic zone (below ~1,000 m and above 4,000 m) has been regarded as the largest ecosystem on the planet (Robison, 2009) and represents ~75% of the volume of the ocean (Ramirez-Llodra et al., 2010). In addition to deep water fish that are exploited commercially (Gordon, 2001), this ecosystem supports a diverse assemblage of gelatinous and other zooplankton, which also play a pivotal role in the sequestration of carbon from the surface ocean to the deep sea floor, in so doing contributing to the global regulation of atmospheric carbon dioxide and thence the Earth's climate (Packard and Gomez, 2013; Teuber et al., 2014).
These different deep water habitats provide considerable Ecosystem Services for humankind; services that need to be afforded protection in the long term as they are both unique and also non-restorable (Le et al., 2017; see Figure 2). Indeed, the provision of Ecosystem Services has been argued as one standard for assessing “serious harm” in the context of DSM Environmental Impact Assessments (EIAs) (Le et al., 2017), because these services link environmental health to human well-being. Provisioning Services include supporting fisheries for human nutrition (e.g., Gordon, 2001) and novel natural products (e.g., Guzman et al., 2016), whilst Regulating Services include the absorption of carbon from the atmosphere and its subsequent sequestration with the deep ocean interior and within the seabed (e.g., Vardaro et al., 2009). At the sea floor diverse and stable biological communities represent a biodiversity resource which provides a reservoir of biological and genetic resources that might present the source of future natural products of further benefit to human kind (Harden-Davies, 2017).
The continued provision of Ecosystem Structures and Functions by marine communities in the vicinity of DSM requires the species assemblage to persist into the future (Zeppilli et al., 2016). Aside from the direct removal of settlement surface (or substrate) on which larvae may settle, the introduction of excess metals into the environment may exert a toxic effect on the resident fauna or may lead to organisms avoiding those areas exposed to high metal concentrations (Figure 2; see below). In either case, the disappearance of organisms from a system, or a reduction in the performance of a species or organism within a system, may result in a reduction in biodiversity and potentially increase ecosystem instability, which will present a risk to the continued provision of Ecosystem Services into the future.
However, it should be borne in mind that impacts to Ecosystem Services may represent a relatively slowly responding metric, one that integrates complex interactive effects (not just metal toxicity) over large spatial scales, and may therefore not be relied upon to provide an early or sensitive indication of toxic effects from released metals. It can be argued that the identification of any significant adverse impact, impacts that may precede any change in Ecosystem Services, should also constitute “serious harm” to the marine environment, and therefore should be appropriately quantified before large-scale mineral exploitation is licensed.
For many commonly-studied metals, existing acute toxicological data [lethal concentrations (LC50) and effective concentrations (EC50)] are available, but only for shallow-water biological species (e.g., Crompton, 1997). These data identify concentrations of metals which are either lethal, or “effective,” for 50% of the exposed population over a designated period, conventionally 72 or 96 h. Alternatively, more recent toxicological studies have adopted a variable exposure duration that matches a desired sub-lethal endpoint (Simpson et al., 2017). The US EPA ECOTOXicology Database (ECOTOX; https://cfpub.epa.gov/ecotox/) is an online resource that summarizes all available metadata included within each ecotoxicology publication, and this database is updated quarterly. However, interrogation of this database at the end of 2013, at the start of the MIDAS project, identified that no data were available for any deep-sea taxa.
Through DSM, the toxic effects of metals will act potentially at in situ deep-sea temperatures and pressures (high pressure—up to 60 MPa, low temperature—down to 2°C), which are very different from those of laboratory exposures reported in the ECOTOX database (conventionally set at standard conditions of a temperature of 20°C and a pressure of 0.1 MPa) (Mestre et al., 2014). Brown et al. (2017a) have contrasted the toxic limits of metals in solution at low temperature (10°C) and high hydrostatic pressure (10 MPa) with those recorded under standard conditions of temperature and pressure (20°C and 0.1 MPa). These initial experiments made use of the shallow-water shrimp Palaemon varians as an experimental model; a species that has a close phylogenetic relationship to deep-water hydrothermal-vent shrimp species (Tokuda et al., 2006). These initial experiments showed that both copper and cadmium toxicity were significantly reduced at low temperatures at 96 h, but that the effects of high hydrostatic pressure were more complex. Whilst copper toxicity was significantly increased at high hydrostatic pressure, cadmium toxicity was not. Consequently, copper toxicity was lower than cadmium toxicity at 20°C but greater than cadmium toxicity at 10°C, and remained greater than cadmium toxicity at 10.0 MPa (Brown et al., 2017a).
Similar results were found in an acute copper toxicity study with the experimental model organism Halomonhystera disjuncta GD1 (Mevenkamp et al., 2017); a nematode that is phylogenetically closely related to the deep-sea species H. hermesi (Van Campenhout et al., 2014; Tchesunov et al., 2015) that inhabits cold-seep ecosystems. In H. disjuncta GD1, cold temperatures (10°C) reduced copper toxicity, whilst toxicity was increased when nematodes were exposed to high hydrostatic pressure (10 MPa) (Mevenkamp et al., 2017).
Whilst these observations of temperature-mediated toxicity are not new (e.g., Lewis and Horning, 1991; Heugens et al., 2003; Khan et al., 2006; Prato et al., 2008; Barbieri et al., 2013), to the extent that authors have suggested the use of a correction factor in applying LC50 values for different temperature environments (Wang et al., 2014), the moderating effects of temperature may not be consistent within the real world (see Chapman et al., 2006). Moreover data indicate that the determination of any “temperature correction factor” will be different at high hydrostatic pressure for each metal in question and would have to be empirically determined for each case, and potentially for each biological species (see also: Kiffney and Clements, 1996; Gonzalez-Rey et al., 2007; Wang et al., 2014; for example).
A further issue of using existing data to regulate mining activity is that many assessments of metal toxicity are based on a single metal presented at a single oxidation state. Mineral ores represent complex mixtures of metals that are site-specific (e.g., Hering, 1971; Scott, 2001; Glasby et al., 2015; Petersen et al., 2016) and that change with mineral weathering (Koschinsky et al., 2003). It is therefore extremely difficult to predict the exact toxic potential of a mineral resource in situ from laboratory studies on single metals, or even metal mixtures (see also the recent discussion of Belzunce-Segarra et al., 2015).
As evidence of this complexity, laboratory studies of the toxicity of copper and cadmium mixtures in the decapod crustacean Palaemon varians were assessed by testing deviation from independent addition reference model (IA) predictions (see Jonker et al., 2005; Pan et al., 2015). Brown et al. (2017a) assessed 96-h lethal toxicity in a mixture of copper and cadmium based on the predicted LC29 of both metals (derived from the modeled mortality responses to individual metals in each temperature/pressure treatment), and compared with mortality in a control treatment and in individual LC29 exposures to copper and cadmium. IA predicted that mortality in a binary mixture with LC29 of both constituents would be 50%; however, the interaction of copper and cadmium appeared potentiating. Most importantly, whilst temperature did not significantly affect the interaction of the two metals when exposed in combination, high hydrostatic pressure significantly increased the toxic effect. These data emphasize the potential key role of pressure in mediating the toxic effect of plumes generated through DSM (Brown et al., 2017a).
The complexity of metal mixture toxicity has recently been reviewed (Pan et al., 2015, and references therein) and a detailed discussion is not presented here. Nonetheless, it is clear from our studies and those of other groups, that the scientific community is not in a position to reliably predict the combined toxic effect of metal mixtures in situ at mining sites. Ultimately, there are fundamental differences in metal uptake from solution, and the subsequent toxic effect in organisms, that are mediated by low temperature and high pressure. These findings confound the principle of regulating DSM on the basis of exposure thresholds established under standard conditions. We conclude that existing toxicological data of metals in solution established at standard laboratory conditions should not necessarily be applied to the context of DSM.
We argue that to continue to establish lethal limits for single metals, or simple combinations of metals, in the laboratory will only produce incremental progress in our understanding of “real world” toxicity of mineral resources. If the scientific community continues with this approach, progress in this field will be too slow to be incorporated usefully into recommendations for contractors (see also, the arguments in Jager et al., 2006). As an alternative, we propose that it will be necessary to assess the “bulk toxicity” of each mineral deposit to identify a priori the potential toxic risk of each mineral resource to be mined within a license area (e.g., Harris et al., 2014; Fallon et al., 2017; Simpson et al., 2017; see also discussion of the “Weight Of Evidence” approach below). In practice, it actually may not be necessary to quantify the individual toxicity of each metal within each resource (although assessment of mineral composition is undoubtedly integral to resource classification; e.g., International Council on Mining Metals, 2013). It may only be necessary to determine—under controlled, ecologically relevant, conditions—the bulk lethal toxicity of that resource for a number of different locally-relevant biological proxy organisms. However, assessments of bulk toxicity should be considered for all relevant life cycle stages of the proxy organisms, including larval stages which are known to be more sensitive (e.g., Rainbow, 2007; Simpson and Batley, 2007; Casado-Martinez et al., 2009; Camusso et al., 2012; Hedouin et al., 2016), and should consider all appropriate physical phases for the metal (e.g., in solution/aqueous, as particulates of relevant size and shape, or adsorbed onto the surface of particulates; see below). A similar approach could be adopted to determine the bulk lethal toxicity of any return waters from surface dewatering before any discharge takes place, potentially as part of a contractor's Environmental Impact Assessment (ISA, 2017).
Any metals released during the mining cycle will occur in different physical states (Simpson and Spadaro, 2016). Metals may enter solution/aqueous phase and be taken up across the gills, body wall and digestive tracts of exposed animals. Alternatively, crushed mineral particles, and dissolved metals that may adsorb onto sediment particles or flocculates may be ingested; this may particularly be the case for metals released during dewatering of ore slurry (for example Campana et al., 2012, 2013; Camusso et al., 2012). These various exposure routes have implications to predictions of toxicity as well, not least because the vast majority of data listed within databases such as the US EPA ECOTOX database are based on organisms exposed to metals in solution.
For example: Simpson and Spadaro (2016) have identified that metals in solution have greater potential toxicity than in particulate phases. Exposure to particulate copper, in the form of chalcopyrite (100% CuFeS2) or chalcocite (80% Cu2S), resulted in a reduced absolute mortality in both bivalves Spisula trigonella and benthic amphipods Melita plumulosa compared to copper in solution. Similarly, Farkas et al. (2017) have recently reported minimal toxicity in the pelagic copepod Calanus finmarchicus. exposed to 5 g l−1 of the fine-grained fraction of marble processing tailings. However, Farkas et al. (2017) do note that this pollutant did constitute a sub-lethal energetic burden to exposed copepods that took these fine-grained particulates into the gut (energetic implications and sub-lethal impacts are considered further in section Sub-Lethal Impacts of Chronic Exposure Should Be Considered).
These recent studies reinforce the necessity to determine the bulk toxicity of the mined resource in all relevant phases prior to the extraction of metals under an exploitation license.
The holistic view of organisms within their ecosystem providing the basis for Ecosystem Structures, Ecological Functions and Services (see earlier discussion, and Le et al., 2017), is in marked contrast to the paradigm of acute “lethal toxicity.” As discussed, acute toxicity is conventionally assessed in terms of the, rather artificial, construct of “96-h LC50.” However, DSM—particularly of polymetallic nodules—may continue within a license block for years to decades and the potential anthropogenic impacts may take place at unprecedented temporal and spatial scales (Glover and Smith, 2003; Selck et al., 2017). Key seabed organisms at considerable distances from the mined site may be subject to chronic metal exposures that are orders of magnitude lower than a lethal dose, but for very extended periods (Newman, 2010; Simpson et al., 2017).
A considerable body of research has identified the mechanisms by which metals in the marine environment might exert toxic effects in marine species (e.g., Hollenberg, 2010 and references in that special issue; Wu et al., 2016; Figure 3). Briefly, redox-active metals including iron, copper, cobalt, chromium and nickel, can catalyze the formation of reactive oxygen and reactive nitrogen species that can bind with lipids and cause lipid peroxidation and damage to cell membranes (Stohs and Bagchi, 1995; Ayala et al., 2014). Redox-inactive metals, including cadmium and zinc, also exert toxicity by binding with the sulfhydryl groups of proteins (Valko et al., 2016). Excess metals may also lead to the production of reactive species, including superoxide (), peroxides (ROOR), singlet oxygen, peroxynitrite (ONOO−), hydroxyl radicals (OH•) and nitric oxide (NO), which may also produce damage in animal tissues (Stocker and Keaney, 2004; Aitken and Roman, 2008; Martinez-Finley and Aschner, 2011). In response, organisms have multiple different mechanisms for the detoxification of reactive oxygen or nitrogen species to minimize damage (Figure 3). For example, organisms can produce thiol-rich metal binding proteins (e.g., metallothioneins; e.g., Hardivillier et al., 2004) that can either be sequestered as inclusion bodies or, if forming soluble complexes, can be secreted from the cells. Redox-inactive metals can be bound to glutathione to form non-toxic metal complexes. Reactive species are variously detoxified by enzymes including: superoxide dismutase (SOD), catalase (CAT), and peroxidases (PEROXs) (e.g., Company et al., 2004, 2006a,b, 2007, 2008; Bebianno et al., 2005; Gonzalez-Rey et al., 2008). Recently, Auguste et al. (2016), Martins et al. (2017) and Mestre et al. (2017) have assessed the potential sub-lethal impacts of exposure to dissolved metals specifically in a range of deep sea species, including molluscs and echinoderms that do not inhabit metal-rich environments, and molluscs and crustaceans from hydrothermal-vent habitats; habitats that are naturally metal rich. For example, copper concentrations from the mussel Bathymodiolus azoricus habitat on the Mid-Atlantic Ridge (MAR) have been recorded between 0.17 μM at ~850 m water depth at Menez Gwen to 2 μM at ~2,300 m water depth at Rainbow (Kádár et al., 2005) hydrothermal vent sites.
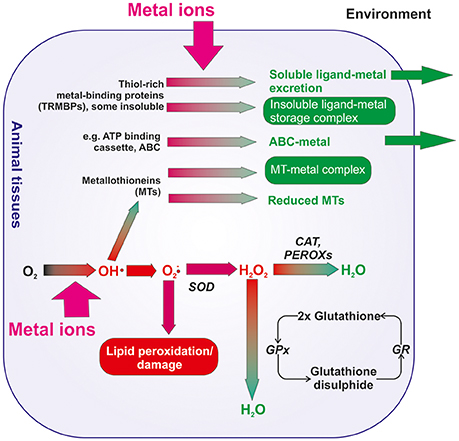
Figure 3. Simplified synthesis of the routes of metal toxicity and detoxification in animal tissues. Metals in the environment that enter animal tissues can be sequestered by various metal binding proteins, including metallothioneins and other thiol-rich peptides, and either stored or excreted from those tissues. Metallothioneins can also detoxify hydroxyl radicals within cells. Redox active metals can drive the formation of superoxide anions (O2-•) via hydroxyl radicals (OH•) that, if unregulated, can lead to lipid peroxidation and damage to cellular membranes. Superoxides can be enzymatically converted to hydrogen peroxide by superoxide dismutase (SOD). Hydrogen peroxide (H2O2) is enzymatically converted to water by catalases and peroxidases (CAT and PEROXs, respectively) or by the action of glutathione peroxidase (GPx) and glutathione disulphide reductase (GR). Synthesis adapted from: Stocker and Keaney (2004), Aitken and Roman (2008), and Martinez-Finley and Aschner (2011).
Auguste et al. (2016) established the effects of copper exposure on the expression of tissue metallothioneins and lipid peroxidation as well as effects of the activity of key antioxidant enzymes in different tissues of the hydrothermal vent shrimp Rimicaris exoculata collected from the TAG hydrothermal vent field on the MAR and maintained at 30 MPa and 10°C. Shrimp were held in solutions of either ~25 or ~250 μg l−1 copper for 72 h, which were at least two orders of magnitude less than the 72-h LC50 concentration of ~35,000 μg l−1 for Palaemon varians at 0.1 MPa and 10°C reported by Brown et al. (2017a). Auguste et al. (2016) demonstrated that even shrimp that have evolved to live in the metal-rich environment of a hydrothermal vent field (R. exoculata) are sensitive to copper exposure in solution. Similarly, copper exposure in hydrothermal vent mussels Bathymodioulus azoricus collected from the “Lucky Strike” hydrothermal vent site on the MAR and maintained at 17.5 MPa at ~6°C for 96 h caused elevated lipid peroxidation at copper concentrations in excess of 300 μg l−1, indicating lipid membrane damage within these tissues.
In these studies, however, data showed that effects of sub-lethal metal exposure were not consistent across all tissues within each organism. In the mussel B. azoricus, although lipid peroxidation was identified in several tissues, gills were more affected whilst mantle and digestive gland were found to be more resilient (Martins et al., 2017). This differential response may be related to the prominent role of the gill in harboring chemosynthetic symbionts and located at the interface between the internal milieu and external vent environment. Similarly, whilst the induction of metallothionein was significant in Rimicaris exoculata gill tissue after 72 h, this was not the case in muscle tissue (Auguste et al., 2016). In this latter case the differential response seen in different tissues of the shrimp was considered to result from differences in the route of uptake of the metal over the time course of the experiment.
Clearly, deep-sea species do have the ability to upregulate detoxification pathways in response to metal exposure and this can occur at relatively low concentrations of metals in solution. However, even with the upregulation of detoxification pathways, damage to tissues evidenced as lipid peroxidation can still be observed. Importantly, it is also clear that even those deep-sea species that inhabit metal-rich environments of active hydrothermal vents are physiologically responsive to metals in solution, although the relative sensitivity of vent and “off vent” species, and how this sub-lethal toxic stress might be phenotypically manifest in deep sea species, remains to be fully constrained.
As discussed, cellular protective mechanisms certainly respond to metal exposure, with upregulated metallothioneins, glutathione, molecular chaperones, antioxidants and/or cellular repair pathways increasing basal metabolic demand (Figure 3; see also Calow, 1989, 1991; Cherkasov et al., 2006; Ivanina and Sokolova, 2008; Ivanina et al., 2008; Sokolova and Lannig, 2008). Whilst successful detoxification might allow for organisms to survive chronic exposure to metals in solution and associated with sediment plumes, the expression of these pathways consumes energy. Basal metabolic rates will increase in association with detoxification and damage repair, as extensively reviewed by Sokolova et al. (2012). Elevated basal metabolic rate will ultimately reduce the energy available for other processes including growth, reproduction and locomotion (e.g., associated with foraging). These impacts may have significant further implications for the role and persistence of species (see Figure 2), even outside of the immediate mining footprint, that should be incorporated into the assessment of exploitation license applications. This is especially the case when the area being considered is adjacent to existing areas of exploitation. For example, in resource/food limited environments such as abyssal plains (Smith et al., 2008), or in environments that are routinely hypoxic (Peña et al., 2010), organisms may not have sufficient energy reserves or aerobic scope to meet the metabolic demand of continued tissue detoxification. Alternatively, at key times of the year—for example periods of reproduction or spawning—benthic organisms may be very susceptible to exposure to metal concentrations that would “normally” be considered to be sub-lethal (e.g., Martins et al., 2011; Levin et al., 2016).
As reviewed above, the maintenance of Ecosystem Services within a system impacted by mining is dependent on the species that remain, and in their individual performance and ecological role—itself an integration of physiology and behavior—in that system (Simpson et al., 2017).
Detailed electrophysiological studies have demonstrated that the vent shrimp Mirocaris exoculata and the phylogenetically close shallow-water shrimp Palaemon elegans have the sensory ability to detect hydrothermal fluid cues like the hydrogen sulfide, as well as food solutions (Machon et al., 2016). They therefore at least possess the potential to detect and respond to changes in their immediate environment, either to remain in close proximity to vent fluids or to move away from high concentrations of metals.
These sensory abilities extend beyond the crustacean taxa. Bivalve molluscs and gastropod snails can withdraw into their shells and either hold the two valves tightly shut or close their opercula opening to avoid detrimental conditions (e.g., Kádár et al., 2001). For example, the fresh water clam Corbicula fluminea can close its valves in response to threshold concentrations of 5.6 or 19.5 μg l−1 copper for response times of 300 and 30 min, respectively (Jou et al., 2016). Indeed, Hartmann et al. (2016) have argued that mussel behavior could be used as a biomarker for toxicological applications, and a similar case has been made for gastropod molluscs exhibiting avoidance behaviors when presented with repellent solutions (Hagner et al., 2015).
Of direct relevance to abyssal polymetallic nodule mining, Brown et al. (2017b) have reported consistent avoidance behaviors in echinoderms presented with copper contaminated sediments. In 96-h laboratory exposures at 4°C the shallow-water holothurian Holothuria forskali consistently avoided sediments contaminated with copper at concentrations of 5 mg l−1 by climbing onto the side of the treatment tank (Brown et al., 2017b). These behaviors resulted in no significant induction of antioxidant enzyme activity. These behaviors were mirrored by the abyssal holothurian Amperima sp. exposed to copper-contaminated sediments at a depth of 4,167 m in the Peru Basin (Brown et al., 2017b) and were also sufficient to avoid induction of antioxidant enzymes in the bulk tissue extracts.
These data demonstrate that macro- and megafaunal species have the sensory capacity to detect metals in the environment and that, in at least some species, this can result in the expression of avoidance behaviors to help protect the organism from toxic effects. It can be concluded that mobile species exposed to contaminated plumes have the potential to demonstrate chronic impacts by moving away from areas of contamination during exploitation. These migrations may produce long-lived direct and indirect (through species interactions) changes in the biological community structure at sites far field from the immediate mining site, creating downstream effects on system Ecological Functions and ultimately Ecosystem Services (Figure 2).
Moreover, avoidance behaviors do not necessarily constitute a permanent solution to toxicant exposure in marine organisms. For example, valve closure in molluscs represents only a temporary avoidance behavior as bivalve molluscs must open their valves periodically to irrigate their gills with oxygenated water (Byrne et al., 1991). Also, for many benthic species, including those at bathyal and abyssal depths, the speed at which a behavioral response can be elicited may be too slow to represent a viable means to avoid toxicant exposure (Ward et al., 2013). In addition, the potential variability in the “typical” behavioral response to toxicant exposure, which may impede their application as biomarker responses, has been identified by other research teams (e.g., Garcia-March et al., 2008; Melvin et al., 2017). Behavioral avoidance is also not possible for sedentary or attached species. As recently reported (e.g., Amon et al., 2016; Vanreusel et al., 2016), polymetallic nodules support a diverse assemblage of sedentary epibenthic fauna, including sponges and soft corals, which cannot move or otherwise avoid environmental exposure. These sessile fauna create vertical habitat for mobile species (Buhl-Mortensen et al., 2010) and their removal from the abyssal system through mining would create downstream impacts for other mobile fauna, impacting Ecosystem Structures and Functions.
Based on current information, including the results of the MIDAS Project Ecotoxicology working group, we argue that it is not possible to predict a priori the absolute toxicity, and therefore identify exposure thresholds, of mining different seabed resources at bathyal and abyssal depths. Indeed, to propose absolute exposure limits based on an incomplete understanding of the mineral resource composition, exposure route and duration, and biological species involved, would be scientifically flawed and would run counter to the ethos of the Precautionary Principle recently advocated by the ISA.
However, it is possible to adopt a Weight Of Evidence (WOE) approach to quantify the risk associated with mining a particular resource. WOE approaches have been advocated for quantifying the toxic risk of exposure to metals in sediments in the absence of absolute data on lethal or effective concentrations (Chapman et al., 1998). Since this time the use of the WOE has been refined to address further uncertainties in the methodology and recommendations on their refinement have since been advocated (e.g., Batley et al., 2002). They have since found application in the assessment of risk in terrestrial (e.g., Milton et al., 2003; Semenzin et al., 2008) and aquatic systems (e.g., Schiesari et al., 2007; Martín-Díaz et al., 2008; Kellar et al., 2014).
Using this approach, multiple Lines Of Evidence (LOE), including a characterization of the mineral resource (chemical composition, grain size), the accumulation of metals within organism tissues, the organism tissue biomarker response, and additional bioassays (MICROTOX, or similar), it is possible to quantify the risk associated with mining (Piva et al., 2011; Benedetti et al., 2012, 2014; Regoli et al., 2014; Bebianno et al., 2015; Mestre et al., 2017).
Applying this approach, with multiple LOE to locally relevant “canary” species in the vicinity of the mining site or validated shallow-water ecotoxicological proxy species, would provide a mechanism for regulators and contractors to develop a holistic overview of the toxic risk of mining and resource. Appropriate “canary species” could be identified as either: (a) biomass dominants within the local biological community (e.g., abyssal sponges or holothurians, or hydrothermal vent shrimp and mussels), or (b) key species necessary to maintain Ecosystem Services (e.g., abyssal holothurians, abyssal meiofauna), or (c) species which could be captured and caged for routine monitoring during exploitation (e.g., bivalve molluscs such as Bathymodiolus azoricus, which is phylogenetically close to the deeper-dwelling B. puteoserpentis, and which naturally inhabits hydrothermal vent environments but which can survive at atmospheric pressure rendering it tractable for laboratory experiments). Ultimately, the precise selection of appropriate canary species will be habitat dependent, and still requires much more comprehensive datasets of the resident biological communities in potential mining locations, particularly in the case of nodule fields at abyssal depths.
Using the WOE approach it will be possible to identify high-risk resources or high-risk communities within a license block during contractor exploration or contractor Environmental Impact Assessments prior to exploitation licenses being issued by the regulating authority—the ISA in the case of the mining within the Area. Exploitation of these high-risk areas could be restricted until appropriate mitigation is in place (through operating procedure, or through mining tool design) or could be used to identify lower risk areas within a license block that could be prioritized for exploitation.
The WOE approach has been applied and validated within the shallow waters of Portmán Bay, Spain during the MIDAS project (Mestre et al., 2017). Nevertheless, further testing and validation of the WOE is required before this approach can be universally recommended to the ISA for use with the Area. The full implications of adopting an a priori WOE approach at bathyal and abyssal depths should be tested in the field, potentially by adopting independent scientific advisors/agencies into a flexible and staged implementation of the Adaptive Management of pilot mining or as part of the first exploitation contracts issued by the ISA (Jaeckel, 2016).
In conclusion, the combined results of the EC MIDAS Ecotoxicology workgroup and other teams reviewed here identify the important, but as yet unpredictable, interacting roles of temperature and pressure in mediating the toxicity of metals to marine invertebrate fauna. As a consequence of these interactions, combined with the variable metal composition of different mineral reserves, it is not possible to predict acute thresholds for marine invertebrates that may be exposed to toxic concentrations of metals during DSM operations. Moreover, we conclude that acute exposure limits represent an artificial measure of possible impact, which do not identify sub-lethal and longer-term chronic responses that may disrupt Ecological Functions and Ecosystem Services within the deep sea. Our data indicate that future regulation of DSM exploitation should consider the potential for the perturbation of normal behaviors of deep-sea species, changes which may also affect the provision of Ecosystem Services in the long term. In order to quantify the toxic risk of mineral exploitation, we advocate the incorporation of existing “Weight of Evidence” approaches into environmental impact assessments. We identify the potential for these approaches to be further developed and validated through continued collaboration of scientific researchers and mining contractors during any future pilot mining operations, according to the principles of Adaptive Management.
CH, ST, MB, IM, MC, BS, AS-V, CG, AD, RD, VG, PW conceived the project and secured funding. All authors contributed to the science described in the manuscript. CH led manuscript production with contributions and comments from all authors.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The research leading to these results has received funding from the European Union Seventh Framework Programme (FP7/2007-2013) under the MIDAS project, grant agreement n° 603418. This publication reflects only the views of the authors; the European Commission is not liable for any use that may be made of the information contained herein. Martins is financed by SFRH/BPD/73481/2010 grant and had the support of Fundação para a Ciência e Tecnologia (FCT), through the strategic project UID/MAR/04292/2013 granted to MARE. Brown is supported through an IMarEST Stanley Gray Fellowship. Canals and Sanchez-Vidal acknowledge the support received from the Spanish funded projects NUREIEV (ref. CTM2013-44598-R) and NUREIEVA (ref. CTM2016-75953-C2-1-R), including ship time. The authors acknowledge the assistance of three reviewers in revising the manuscript. NM and MB acknowledge the support from FCT through the grant UID/MAR/00350/2013 attributed to CIMA, University of Algarve.
Aitken, R. J., and Roman, S. D. (2008). Antioxidant systems and oxidative stress in the testes. Oxid. Med. Cell. Longev. 1, 15–24. doi: 10.4161/oxim.1.1.6843
Amiard Triquet, C., Jeantet, A. Y., and Berthet, B. (1993). Metal transfer in marine food chains – bioaccumulation and toxicity. Acta Biol. Hungarica 44, 387–409.
Amon, D. J., Ziegler, A. F., Dahlgren, T. G., Glover, A. G., Goineau, A., Gooday, A. J., et al. (2016). Insights into the abundance and diversity of abyssal megafauna in a polymetallic-nodule region in the eastern Clarion-Clipperton Zone. Sci. Rep. 6:30492. doi: 10.1038/srep30492
Auguste, M., Mestre, N., Rocha, T. L., Cardoso, C., Cueff-Gauchard, V., Le Bloa, S., et al. (2016). Development of an ecotoxicological protocol for the deep-sea fauna using the hydrothermal vent shrimp Rimicaris exoculata. Aquat. Toxicol. 175, 277–285. doi: 10.1016/j.aquatox.2016.03.024
Ayala, A., Muñoz, M. F., and Argüelles, S. (2014). Lipid peroxidation: production, metabolism, and signalling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014:360438. doi: 10.1155/2014/360438
Barbieri, E., Branco, J. O., Santos, M. D. F., and Hidalgo, K. R. (2013). Effects of cadmium and zinc on oxygen consumption and ammonia excretion of the sea-bob shrimp, according to the temperature. Boletim Instit. Pesca 39, 299–309. doi: 10.20950/1678-2305.2013v39n3p299
Batley, G. E., Burton, G. A., Chapman, P. M., and Forbes, V. E. (2002). Uncertainties in sediment quality weight-of-evidence (WOE) assessments Human Ecol. Risk Assess. 8, 1517–1547. doi: 10.1080/20028091057466
Bebianno, M. J., Company, R., Serafim, A., Camus, L., Cosson, R. P., and Fiala-Medioni, A. (2005). Antioxidant systems and lipid peroxidation in Bathymodiolus azoricus from Mid-Atlantic Ridge hydrothermal vent fields. Aquat. Toxicol. 75, 354–373. doi: 10.1016/j.aquatox.2005.08.013
Bebianno, M. J., Pereira, C. G., Rey, F., Cravo, A., Duarte, D., D'Errico, G., et al. (2015). Integrated approach to assess ecosystem health in harbor areas. Sci. Tot. Environ. 514, 92–107. doi: 10.1016/j.scitotenv.2015.01.050
Belzunce-Segarra, M. J., Simpson, S. L., Amato, E. D., Spadaro, D. A., Hamilton, I. L., Jarolimek, C. V., et al. (2015). The mismatch between bioaccumulation in field and laboratory environments: interpreting the differences for metals in benthic bivalves. Environ. Poll. 204, 48–57. doi: 10.1016/j.envpol.2015.03.048
Benedetti, M., Ciaprini, F., Piva, F., Onorati, F., Fattorini, D., Notti, A., et al. (2012). A multidisciplinary weight of evidence approach toward polluted sediments: integrating sediment chemistry, bioavailability, biomarkers responses and bioassays. Environ. Int. 38, 17–28. doi: 10.1016/j.envint.2011.08.003
Benedetti, M., Gorbi, S., Fattorini, D., Pacitti, D., D'Errico, G., Piva, F., et al. (2014). Environmental hazards from natural seepage: integrated classification of risk from sediment chemistry, bioavailability and biomarkers responses in sentinel species. Environ. Poll. 185, 116–126. doi: 10.1016/j.envpol.2013.10.023
Boschen, R. E., Rowden, A. A., Clark, M. R., Pallentin, A., and Gardner, J. P. A. (2016). Seafloor massive sulfide deposits support unique megafaunal assemblages: implications for seabed mining and conservation. Mar. Environ. Res. 115, 78–88. doi: 10.1016/j.marenvres.2016.02.005
Brown, A., Thatje, S., and Hauton, C. (2017a). The effects of temperature and hydrostatic pressure on metal toxicity: insights into toxicity in the deep sea. Environ. Sci. Technol. 51, 10222–10231. doi: 10.1021/acs.est.7b02988
Brown, A., Wright, R., Mevenkamp, L., and Hauton, C. (2017b). A comparative experimental approach to ecotoxicology in shallow-water and deep-sea holothurians. Aquat. Toxicol. 191, 10–16. doi: 10.1016/j.aquatox.2017.06.028
Buhl-Mortensen, L., Vanreusel, A., Gooday, A. J., Levin, L. A., Priede, I. G., Buhl-Mortensen, P., et al. (2010). Biological structures as a source of habitat heterogeneity and biodiversity on the deep ocean margins. Mar. Ecol. 31, 21–50. doi: 10.1111/j.1439-0485.2010.00359.x
Byrne, R. A., Shipman, B. N., Smatresk, N. J., Dietz, T. H., and McMahon, R. F. (1991). Acid-base-balance during emergence in the fresh-water bivalve Corbicula-fluminea. Physiol. Zool. 64, 748–766. doi: 10.1086/physzool.64.3.30158205
Calow, P. (1991). Physiological costs of combating chemical toxicants–ecological implications. Comp. Biochem. Physiol. 100C, 3–6. doi: 10.1016/0742-8413(91)90110-F
Campana, O., Blasco, J., and Simpson, S. L. (2013). Demonstrating the appropriateness of developing sediment quality guidelines based on sediment geochemical properties. Environ. Sci. Technol. 47, 7483–7489. doi: 10.1021/es4009272
Campana, O., Spadaro, D. A., Blasco, J., and Simpson, S. L. (2012). Sublethal effects of copper to benthic invertebrates explained by changes in sediment properties and dietary exposure. Environ. Sci. Technol. 46, 6835–6842. doi: 10.1021/es2045844
Camusso, M., Polesello, S., Valsecchi, S., and Vignati, D. A. L. (2012). Importance of dietary uptake of trace elements in the benthic deposit feeding Lumbriculus variegatus. TrAC Trends Anal. Chem. 36, 103–112. doi: 10.1016/j.trac.2012.02.010
Caroppo, C., Stabili, L., Aresta, M., Corinaldesi, C., and Danovaro, R. (2006). Impact of heavy metals and PCBs on marine picoplankton. Environ. Toxicol. 21, 541–551. doi: 10.1002/tox.20215
Casado-Martinez, M. C., Smith, B. D., DelValls, T. A., and Rainbow, P. S. (2009). Pathways of trace metal uptake in the lugworm Arenicola marina. Aquat. Toxicol. 92, 9–17. doi: 10.1016/j.aquatox.2008.12.010
Chapman, P. M., McDonald, B. G., Kickham, P. E., and McKinnon, S. (2006). Global geographic differences in marine metals toxicity. Mar. Poll. Bull. 52, 1081–1084. doi: 10.1016/j.marpolbul.2006.05.004
Chapman, P. M., Wang, F. Y., Janssen, C., Persoone, G., and Allen, H. E. (1998). Ecotoxicology of metals in aquatic sediments: binding and release, bioavailability, risk assessment, and remediation. Can. J. Fish Aquat. Sci. 55, 2221–2243. doi: 10.1139/f98-145
Cherkasov, A. S., Ringwood, A. H., and Sokolova, I. M. (2006). Combined effects of temperature acclimation and cadmium exposure on mitochondrial function in eastern oysters Crassostrea virginica Gmelin (Bivalvia: Ostreidae). Environ. Toxicol. Chem. 25, 2461–2469. doi: 10.1897/05-584R.1
Collins, P. C., Croot, P., Carlsson, J., Colaço, A., Grehan, A., Hyeong, K., et al. (2013). A primer for the Environmental Impact Assessment of mining at seafloor massive sulphide deposits. Mar. Pol. 42, 198–209. doi: 10.1016/j.marpol.2013.01.020
Company, R., Serafim, A., Bebianno, M. J., Cosson, R., Shillito, B., and Fiala-Medioni, A. (2004). Effect of cadmium, copper and mercury on antioxidant enzyme activities and lipid peroxidation in the gills of the hydrothermal vent mussel Bathymodiolus azoricus. Mar. Environ. Res. 58, 377–381. doi: 10.1016/j.marenvres.2004.03.083
Company, R., Serafim, A., Cosson, R., Camus, L., Shillito, B., Fiala-Medioni, A., et al. (2006a). The effect of cadmium on antioxidant responses and the susceptibility to oxidative stress in the hydrothermal vent mussel Bathymodiolus azoricus. Mar. Biol. 148, 817–825. doi: 10.1007/s00227-005-0116-0
Company, R., Serafim, A., Cosson, R., Fiala-Médioni, A., Dixon, D., and Bebianno, M. J. (2006b). Temporal variation in the antioxidant defence system and lipid peroxidation in the gills and mantle of hydrothermal vent mussel Bathymodiolus azoricus. Deep-Sea Res. I 53, 1101–1116. doi: 10.1016/j.dsr.2006.05.008
Company, R., Serafim, A., Cosson, R., Fiala-Médioni, A., Dixon, D. R., and Bebianno, M. J. (2007). Adaptation of the antioxidant defence system in hydrothermal-vent mussels (Bathymodiolus azoricus) transplanted between two Mid-Atlantic Ridge sites. Mar. Ecol. 28, 93–99. doi: 10.1111/j.1439-0485.2006.00125.x
Company, R., Serafim, A., Cosson, R. P., Fiala-Médioni, A., Camus, L., Colaço, A., et al. (2008). Antioxidant biochemical responses to long-term copper exposure in Bathymodiolus azoricus from Menez-Gwen hydrothermal vent. Sci. Total Environ. 389, 407–417. doi: 10.1016/j.scitotenv.2007.08.056
Danovaro, R., Aguzzi, J., Fanelli, E., Billett, D., Gjerde, K., Jamieson, A., et al. (2017). Towards a global ecosystem-based strategy for the deep ocean. Science 355, 452–454. doi: 10.1126/science.aah7178
Ellis, D. V. (2001). A review of some environmental issues affecting marine mining. Mar. Geores. Geotech. 19, 51–63. doi: 10.1080/10641190109353804
Fallon, E. K., Petersen, S., Brooker, R. A., and Scott, T. B. (2017). Oxidative dissolution of hydrothermal mixed-sulphide ore: an assessment of current knowledge in relation to seafloor massive sulphide mining. Ore Geol. Rev. 86, 309–337. doi: 10.1016/j.oregeorev.2017.02.028
Farkas, J., Altin, D., Hammer, K. M., Hellstrom, K. C., Booth, A. M., and Hansen, B. H. (2017). Characterisation of fine-grained tailings from a marble processing plant and their acute effects on the copepod Calanus finmarchicus. Chemosphere 169, 700–708. doi: 10.1016/j.chemosphere.2016.11.118
Fuchida, S., Yokoyama, A., Fukuchi, R., Ishibashi, J., Kawagucci, S., Kawachi, M., et al. (2017). Leaching of metals and metalloids from hydrothermal ore particulates and their effects on marine phytoplankton. ACS Omega 7, 3175–3182. doi: 10.1021/acsomega.7b00081
Garcia-March, J. R., Solsona, M. A. S., and Garcia-Carrascosa, A. M. (2008). Shell gaping behaviour of Pinna nobilis L., 1758: circadian and circalunar rhythms revealed by in situ monitoring. Mar. Biol. 153, 689–698. doi: 10.1007/s00227-007-0842-6
Glasby, G. P., Li, J., and Sun, Z. (2015). Deep-sea nodules and Co-rich Mn crusts. Mar. Geosci. Geotech. 33, 72–78. doi: 10.1080/1064119X.2013.784838
Glover, A. G., and Smith, C. R. (2003). The deep-sea floor ecosystem: current status and prospects of anthropogenic change by the year 2025. Environ. Conserv. 30, 219–241. doi: 10.1017/S0376892903000225
Gonzalez-Rey, M., Serafim, A., Company, R., and Bebianno, M. J. (2007). Adaptation to metal toxicity: a comparison of hydrothermal vent and coastal shrimps. Mar. Ecol. 28, 100–107. doi: 10.1111/j.1439-0485.2006.00126.x
Gonzalez-Rey, M., Serafim, A., Company, R., Gomes, T., and Bebianno, M. J. (2008). Detoxification mechanisms in shrimp: comparative approach between hydrothermal vent fields and estuarine environments. Mar. Environ. Res. 66, 35–37. doi: 10.1016/j.marenvres.2008.02.015
Gordon, J. D. M. (2001). Deep-water fisheries at the Atlantic Frontier. Cont. Shelf Res. 21, 987–1003. doi: 10.1016/S0278-4343(00)00121-7
Guzman, E. A., Xu, Q. L., Pitts, T. P., Mitsuhashi, K. O., Baker, C., Linley, P. A., et al. (2016). Leiodermatolide, a novel marine natural product, has potent cytotoxic and antimitotic activity against cancer cells, appears to affect microtubule dynamics, and exhibits antitumor activity. Int. J. Can. 139, 2116–2126. doi: 10.1002/ijc.30253
Hagner, M., Kuoppala, E., Fagernäs, L., Tiilikkala, K., and Setälä, H. (2015). Using the Copse snail Arianta arbustorum (Linnaeus) to detect repellent compounds and the quality of wood vinegar. Int. J. Environ. Res. 9, 53–60. doi: 10.22059/IJER.2015.873
Hannington, M., Petersen, S., and Krätschell, A. (2017). Subsea mining moves closer to shore. Nat. Geosci. 10, 158–159. doi: 10.1038/ngeo2897
Harden-Davies, H. (2017). Deep-sea genetic resources: new frontiers for science and stewardship in areas beyond national jurisdiction. Deep-Sea Res. II. 137, 504–513. doi: 10.1016/j.dsr2.2016.05.005
Hardivillier, Y., Leignel, V., Denis, F., Uguen, G., Cosson, R., and Laulier, M. (2004). Do organisms living around hydrothermal vent sites contain specific metallothioneins? The case of the genus Bathymodiolus (Bivalvia, Mytilidae). Comp. Biochem. Physiol. 139C, 111–118. doi: 10.1016/j.cca.2004.09.012
Harris, C. A., Scott, A. P., Johnson, A. C., Panter, G. H., Sheahan, D., Roberts, M., et al. (2014). Principles of sound ecotoxicology. Environ. Sci. Technol. 48, 3100–3111. doi: 10.1021/es4047507
Hartmann, J. T., Beggel, S., Auerswald, K., Stoeckle, B. C., and Geist, J. (2016). Establishing mussel behavior as a biomarker in ecotoxicology. Aquat. Toxicol. 170, 279–288. doi: 10.1016/j.aquatox.2015.06.014
Hedouin, L. S., Wolf, R. E., Phillips, J., and Gates, R. D. (2016). Improving the ecological relevance of toxicity tests on scleractinian corals: influence of season, life stage, and seawater temperature. Environ. Poll. 213, 240–253. doi: 10.1016/j.envpol.2016.01.086
Hering, N. (1971). Deep-sea ores. I. Past and future equipment and techniques for detecting manganese deposits. Mar. Tech. 2, 155–162.
Heugens, E. H. W., Jager, T., Creyghton, R., Kraak, M. H. S., Hendriks, A. J., Van Straalen, N. M., et al. (2003). Temperature-dependent effects of cadmium on Daphnia magna: acclimation vs. sensitivity. Environ. Sci. Technol. 37, 2145–2151. doi: 10.1021/es0264347
Hirota, J. (1981). Potential effects of deep sea minerals mining on macro zooplankton in the North Equatorial Pacific. Mar. Mining 3, 19–58.
Hoagland, P., Beaulieu, S., Tivey, M. A., Eggert, R. G., German, C., Glowka, L., et al. (2010). Deep-sea mining of seafloor massive sulphides. Mar. Pol. 34, 728–732. doi: 10.1016/j.marpol.2009.12.001
Hollenberg, P. F. (2010). Introduction: mechanisms of metal toxicity special issue. Chem. Res. Toxicol. 23, 292–293. doi: 10.1021/tx900456p
Hu, V. J. H. (1981). Ingestion of deep sea mining discharge by 5 species of tropical copepods. Water Air Soil Poll. 15, 433–440. doi: 10.1007/BF00279425
International Council on Mining and Metals (2013). Committee for Mineral Reserves International Reporting Standards (CRISCO) International Reporting Template for the Public Reporting of Exploration results, Mineral Resources and Mineral Reserves. London, UK.
International Seabed Authority (ISA) (2017). A Discussion Paper on the Development and Drafting of Regulations on Exploitation for Mineral Resources in the Area (Environmental Matters). Kingston Jamaica.
Ivanina, A. V., Cherkasov, A. S., and Sokolova, I. M. (2008). Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin. J. Exp. Biol. 211, 577–586. doi: 10.1242/jeb.011262
Ivanina, A. V., and Sokolova, I. M. (2008). Effects of cadmium exposure on expression and activity of P-glycoprotein in eastern oysters, Crassostrea virginica Gmelin. Aquat. Toxicol. 88, 19–28. doi: 10.1016/j.aquatox.2008.02.014
Jaeckel, A. (2016). Deep seabed mining and adaptive management: the procedural challenges for the International Seabed Authority. Mar. Pol. 70, 205–211. doi: 10.1016/j.marpol.2016.03.008
Jager, T., Heugens, E. H. W., and Kooijman, S. A. L. M. (2006). Making sense of ecotoxicological test results: towards application of process-based models. Ecotoxicology 15, 305–314. doi: 10.1007/s10646-006-0060-x
Jonker, M. J., Svendsen, C., Bedaux, J. J., Bongers, M., and Kammenga, J. E. (2005). Significance testing of synergistic/antagonistic, dose level-dependent, or dose-ratio dependent effects in mixture dose-response analysis. Environ. Toxicol. Chem. 24, 2701–2713. doi: 10.1897/04-431R.1
Jou, L.-J., Chen, B.-C., Chen, W.-Y., and Liao, C.-M. (2016). Sensory determinants of valve rhythm dynamics provide in situ biodetection of copper in aquatic environments. Environ. Sci. Poll. Res. 23:5374–5389. doi: 10.1007/s11356-015-5735-5
Kádár, E., Costa, V., Martins, I., Santos, R. S., and Powell, J. J. (2005). Enrichment in trace metals (Al, Mn, Co, Cu, Mo, Cd, Fe, Zn, Pb and Hg) of macro-invertebrate habitats at hydrothermal vents along the Mid-Atlantic Ridge. Hydrobiology 548, 191–205. doi: 10.1007/s10750-005-4758-1
Kádár, E., Salanki, J., Jugdaohsingh, R., Powell, J. J., McCrohan, C. R., and White, K. N. (2001). Avoidance responses to aluminium in the freshwater bivalve Anodonta cygnea. Aquat. Toxicol. 55, 137–148. doi: 10.1016/S0166-445X(01)00183-7
Kellar, C. R., Hassell, K. L., Long, S. M., Myers, J. H., Golding, L., Rose, G., et al. (2014). Ecological evidence links adverse biological effects to pesticide and metal contamination in an urban Australian watershed. J. Appl. Ecol. 51, 426–439. doi: 10.1111/1365-2664.12211
Khan, M. A. Q., Ahmed, S. A., Catalin, B., Khodadoust, A., Ajayi, O., and Vaughn, M. (2006). Effect of temperature on heavy metal toxicity to juvenile crayfish, Orconectes immunis (Hagen). Environ. Toxicol. 21, 513–520. doi: 10.1002/tox.20213
Kiffney, P. M., and Clements, W. H. (1996). Effects of metals on stream macroinvertebrate assemblages from different altitudes. Ecol. Appl. 6, 472–491. doi: 10.2307/2269384
Koschinsky, A., Winkler, A., and Fritsche, U. (2003). Importance of different types of marine particles for the scavenging of heavy metals in the deep-sea bottom water. App. Geochem. 18, 693–710. doi: 10.1016/S0883-2927(02)00161-0
Le, J. T., Levin, L. A., and Carson, R. T. (2017). Incorporating ecosystem services into environmental management of deep-seabed mining. Deep-Sea Res II. 137, 486–803. doi: 10.1016/j.dsr2.2016.08.007
Levin, L. A., Mengerink, K., Gjerde, K. M., Rowden, A. A., Van Dover, C. L., Clark, M. R., et al. (2016). Defining “serious harm” to the marine environment in the context of deep-seabed mining. Mar. Pol. 74, 245–259. doi: 10.1016/j.marpol.2016.09.032
Lewis, P. A., and Horning, W. B. (1991). Differences in acute toxicity test-results of 3 reference toxicants on Daphnia at 2 temperatures. Environ. Toxicol. Chem. 10, 1351–1357. doi: 10.1002/etc.5620101014
Loka Bharathi, P. A., Nagender Nath, B., Ingole, B. S., Parthiban, G., Matondkar, S. G. P., Raghukumar, C., et al. (2005). Impacts on surface productivity during sediment dispersal experiment in Central Indian Basin. Mar. Geores. Geotechnol. 23, 315–330. doi: 10.1080/10641190500446672
Machon, J., Ravaux, J., Zbinden, M., and Lucas, P. (2016). New electroantennography method on a marine shrimp in water. J. Exp. Biol. 219, 3696–3700. doi: 10.1242/jeb.140947
Marsh, L., Copley, J. T., Huvenne, V. A. I., Linse, K., Reid, W. D. K., Rogers, A. D., et al. (2012). Microdistribution of faunal assemblages at deep-sea hydrothermal vents in the Southern Ocean. PLoS ONE 10:e48348. doi: 10.1371/journal.pone.0048348
Martín-Díaz, M. L., DelValls, T. A., Riba, I., and Blasco, J. (2008). Integrative sediment quality assessment using a biomarker approach: review of 3 years of field research. Cell. Biol. Toxicol. 24, 513–526. doi: 10.1007/s10565-008-9062-1
Martinez-Finley, E. J., and Aschner, M. (2011). Revelations from the nematode Caenorhabditis elegans on the complex interplay of metal toxicological mechanisms. J. Toxicol. 2011:895236. doi: 10.1155/2011/895236
Martins, I., Bettencourt, R., Colaço, A., Sarradin, P.-M., Santos, R. M., and Cosson, R. P. (2011). The influence of nutritional conditions on metal uptake by the mixotrophic dual symbiosis harboring vent mussel Bathymodiolus azoricus (Cosel et al. 1999). Comp. Biochem. Physiol. 153C, 40–52. doi: 10.1016/j.cbpc.2010.08.004
Martins, I., Goulart, J., Martins, E., Morales-Román, R., Marín, S., Riou, V., et al. (2017). Physiological impacts of acute Cu exposure on deep-sea vent mussel Bathymodiolus azoricus under a deep-sea mining activity scenario. Aquat. Toxicol. 193, 40–49. doi: 10.1016/j.aquatox.2017.10.004
Melvin, S. D., Petit, M. A., Duvignacq, M. C., and Sumpter, J. P. (2017). Towards improved behavioural testing in aquatic toxicology: acclimation and observation times are important factors when designing behavioural tests with fish. Chemosphere 180, 430–436. doi: 10.1016/j.chemosphere.2017.04.058
Mestre, N. C., Calado, R., and Soares, A. M. V. M. (2014). Exploitation of deep-sea resources: the urgent need to understand the role of high pressure in the toxicity of chemical pollutants to deep-sea organisms. Environ. Poll. 185, 369–371. doi: 10.1016/j.envpol.2013.10.021
Mestre, N. C., Rocha, T. L., Canals, M., Cardoso, C., Danovaro, R., Dell'Anno, A., et al. (2017). Environmental hazard assessment of a marine mine tailings deposit site and potential implications for deep-sea mining. Environ. Poll. 228, 169–178. doi: 10.1016/j.envpol.2017.05.027
Mevenkamp, L., Brown, A., Kordas, A., Thatje, S., Hauton, C., and Vanreusel, A. (2017). Hydrostatic pressure and temperature affect the tolerance of the free-living marine nematode Halomonhystera disjuncta to acute copper exposure. Aquat. Toxicol. 192C, 178–118. doi: 10.1016/j.aquatox.2017.09.016
Milton, A., Cooke, J. A., and Johnson, M. S. (2003). Accumulation of lead, zinc, and cadmium in a wild population of Clethrionomys glareolus from an abandoned lead mine. Arch. Environ. Contam. Toxicol. 44, 405–411. doi: 10.1007/s00244-002-2014-5
Moraitouapostolopoulou, M., and Verriopoulos, G. (1982). Individual and combined toxicity of 3 heavy-metals, Cu, Cd and Cr for the marine copepod Tisbe-holothuriae. Hydrobiology 87, 83–87. doi: 10.1007/BF00016665
Oebius, H. U., Becker, H. J., Rolinski, S., and Jankowski, J. A. (2001). Parametrization and evaluation of marine environmental impacts produced by deep-sea manganese nodule mining. Deep-Sea Res. II. 48, 3453–3467. doi: 10.1016/S0967-0645(01)00052-2
Packard, T. T., and Gomez, M. (2013). Modeling vertical carbon flux from zooplankton respiration. Prog. Ocean. 110, 59–60. doi: 10.1016/j.pocean.2013.01.003
Pan, J. J., Pan, J.-F., and Diao, M. Y. (2015). Trace metal mixture toxicity in aquatic organisms reviewed from a biotoxicity perspective. Hum. Ecol. Risk Assess. 21, 2155–2169. doi: 10.1080/10807039.2015.1032211
Peña, M. A., Katsev, S., Oguz, T., and Gilbert, D. (2010). Modelling dissolved oxygen dynamics and hypoxia. Biogeosciences 7, 933–957. doi: 10.5194/bg-7-933-2010
Petersen, S., Krätschell, A., Augustin, N., Jamieson, J., Hein, J. R., and Hannington, M. D. (2016). News from the seabed – Geological characteristics and resource potential of deep-sea mineral resources. Mar. Pol. 70, 175–187. doi: 10.1016/j.marpol.2016.03.012
Phillips, B. T. (2017). Beyond the vent: new perspectives on hydrothermal plumes and pelagic biology. Deep-Sea Res. II. 137, 480–485. doi: 10.1016/j.dsr2.2016.10.005
Piva, F., Ciaprini, F., Onorati, F., Benedetti, M., Fattorini, D., Ausili, A., et al. (2011). Assessing sediment hazard through a Weight Of Evidence approach with bioindicator organisms: a practical model to elaborate data from sediment chemistry, bioavailability, biomarkers and ecotoxicological bioassays. Chemosphere 83, 475–485. doi: 10.1016/j.chemosphere.2010.12.064
Prato, E., Scardicchio, C., and Biandolino, F. (2008). Effects of temperature on the acute toxicity of cadmium to Corophium insidiosum. Environ. Monitor. Assess. 136, 161–166. doi: 10.1007/s10661-007-9672-8
Rainbow, P. S. (2007). Trace metal bioaccumulation: models, metabolic availability and toxicity. Environ. Int. 33, 576–582. doi: 10.1016/j.envint.2006.05.007
Ramirez-Llodra, E., Brandt, A., Danovaro, R., De Mol, B., Escobar, E., German, C. R., et al. (2010). Deep, diverse and definitely different: unique attributes of the world's largest ecosystem. Biogeosciences 7, 2851–2899. doi: 10.5194/bg-7-2851-2010
Regoli, F., Pellegrini, D., Cicero, A. M., Nigro, N., Benedetti, M., Gorbi, S., et al. (2014). A multidisciplinary weight of evidence approach for environmental risk assessment at the Costa Concordia wreck: integrative indices from Mussel Watch. Mar. Environ. Res. 96, 92–104. doi: 10.1016/j.marenvres.2013.09.016
Robison, B. H. (2009). Conservation of deep pelagic biodiversity, Conserv. Biol. 23, 847–858. doi: 10.1111/j.1523-1739.2009.01219.x
Sarrazin, J., Legendre, P., de Busserolles, F., Fabri, M. C., Guilini, K., Ivanenko, V. N., et al. (2015). Biodiversity patterns, environmental drivers and indicator species on a high-temperature hydrothermal edifice, Mid-Atlantic Ridge. Deep-Sea Res. II. 121, 177–192. doi: 10.1016/j.dsr2.2015.04.013
Schiesari, L., Grillitsch, B., and Grillitsch, H. (2007). Biogeographic biases in research and their consequences for linking amphibian declines to pollution. Conserv. Biol. 21, 465–471. doi: 10.1111/j.1523-1739.2006.00616.x
Scott, S. D. (2001). Deep ocean mining. Geosci. Can. 28, 87–96. Available online at: https://journals.lib.unb.ca/index.php/GC/article/view/4078
Selck, H., Adamsen, P. B., Backhaus, T., Banta, G. T., Bruce, P. K. H., Burton, G. A., et al. (2017). Assessing and managing multiple risks in a changing world–the Roskilde recommendations. Environ. Toxicol. Chem. 36, 7–16. doi: 10.1002/etc.3513
Semenzin, E., Critto, A., Rutgers, M., and Marcomini, A. (2008). Integration of bioavailability, ecology and ecotoxicology by three lines of evidence into ecological risk indexes for contaminated soil assessment. Sci Tot. Environ. 389, 71–86. doi: 10.1016/j.scitotenv.2007.08.032
Simpson, S. L., and Batley, G. E. (2007). Predicting metal toxicity in sediments: a critique of current approaches. Integr. Environ. Assess. Manag. 3, 18–31. doi: 10.1002/ieam.5630030103
Simpson, S. L., Ho, K. T., and Campana, O. (2017). “Sediment toxicity testing.,” in Marine Ecotoxicology: Current Knowledge and Future Issues, eds J. Blasco, O. Campana, P. Chapman, and M. Hampel (Amsterdam: Elsevier), 197–235.
Simpson, S. L., and Spadaro, D. A. (2016). Bioavailability and chronic toxicity of metal sulfide minerals to benthic marine invertebrates: implications for deep sea exploration, mining and tailings disposal. Environ. Sci. Technol. 50, 4061–4070. doi: 10.1021/acs.est.6b00203
Smith, C. R., De Leo, F. C., Bernardino, A. F., Sweetman, A. K., and Martinez Arbizu, P. (2008). Abyssal food limitation, ecosystem structure and climate change. Trends Ecol. Evol. 23, 518–528. doi: 10.1016/j.tree.2008.05.002
Sokolova, I. M., Frederich, M., Bagwe, R., Lannig, G., and Sukhotin, A. A. (2012). Energy homeostasis as an integrative tool for assessing limits of environmental stress tolerance in aquatic invertebrates. Mar. Environ. Res. 79, 1–15. doi: 10.1016/j.marenvres.2012.04.003
Sokolova, I. M., and Lannig, G. (2008). Interactive effects of metal pollution and temperature on metabolism in aquatic ectotherms: implications of global climate change. Clim. Res. 37, 181–201. doi: 10.3354/cr00764
Stocker, R., and Keaney, J. F. Jr. (2004). Role of oxidative modifications in atherosclerosis. Phys. Rev. 84, 1381–1478. doi: 10.1152/physrev.00047.2003
Stohs, S. J., and Bagchi, D. (1995). Oxidative mechanisms in the toxicity of metal-ions. Free Rad. Biol. Med. 18, 321–336. doi: 10.1016/0891-5849(94)00159-H
Tchesunov, A. V., Portnova, D. A., and Van Campenhout, J. (2015). Description of two free-living nematode species of Halomonhystera disjuncta complex (Nematoda: Monhysterida) from two peculiar habitats in the sea. Helgoland Mar. Res. 69, 57–85. doi: 10.1007/s10152-014-0416-1
Teuber, L., Schukat, A., Hagen, W., and Auel, H. (2014). Trophic interactions and life strategies of epi- to bathypelagic calanoid copepods in the tropical Atlantic Ocean. J. Plank. Res. 36, 1109–1123. doi: 10.1093/plankt/fbu030
Thornburg, C. C., Zabriskie, T. M., and McPhail, K. L. (2010). Deep-sea hydrothermal vents: potential hot spots for natural products discovery? J. Nat. Prod. 73, 489–499. doi: 10.1021/np900662k
Tokuda, G., Yamada, A., Nakano, K., Arita, N., and Yamasaki, H. (2006). Occurrence and recent long-distance dispersal of deep-sea hydrothermal vent shrimps. Biol. Lett. 2, 257–260. doi: 10.1098/rsbl.2005.0420
Valko, M., Jomova, K., Rhodes, C. J., Kuča, K., and Musílek, K. (2016). Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 90, 1–37. doi: 10.1007/s00204-015-1579-5
Van Campenhout, J., Derycke, S., Tchesunov, A. V., Portnova, D., and Vanreusel, A. (2014). The dominant Håkon Mosby mud volcano nematode is genetically differentiated from its shallow–water relatives and shows genetic structure within the mud volcano. J. Zool. Syst. Evol. Res. 52, 203–216. doi: 10.1111/jzs.12054
Van Dover, C. L. (2000). The Ecology of Deep-sea Hydrothermal Vents. Princeton, NJ: Princeton University Press.
Van Dover, C. L., German, C. R., Speer, K. G., Parson, L. M., and Vrijenhoek, R. C. (2002). Evolution and biogeography of deep-sea vent and seep invertebrates. Science 295, 1253–1257. doi: 10.1126/science.1067361
Vanreusel, A., Hilario, A., Ribeiro, P. A., Menot, L., and Arbizu, P. M. (2016). Threatened by mining, polymetallic nodules are required to preserve abyssal epifauna. Sci. Rep. 6:26808. doi: 10.1038/srep26808
Vardaro, M. F., Ruhl, H. A., and Smith, K. L. (2009). Climate variation, carbon flux, and bioturbation in the abyssal North Pacific. Limnol. Oceanogr. 54, 2081–2088. doi: 10.4319/lo.2009.54.6.2081
Wang, T. W., Chan, T. Y., and Chan, B. K. K. (2013). Diversity and community structure of decapod crustaceans at hydrothermal vents and nearby deep-water fishing grounds off Kueishan Island, Taiwan: a high biodiversity deep-sea area in the NW Pacific. Bull. Mar. Sci. 89, 505–528. doi: 10.5343/bms.2012.1036
Wang, Z., Kwok, K. W. H., Lui, G. C. S., Zhou, G. J., Lee, J. S., Lam, M. H. W., et al. (2014). The difference between temperate and tropical saltwater species' acute sensitivity to chemicals is relatively small. Chemosphere 105, 31–43. doi: 10.1016/j.chemosphere.2013.10.066
Ward, D., Simpson, S. L., and Jolley, D. F. (2013). Slow avoidance response to contaminated sediments elicits sub-lethal toxicity to benthic invertebrates Enviro. Sci. Technol. 47, 5947–5953. doi: 10.1021/es400152a
Wolff, T. (2005). Composition and endemism of the deep-sea hydrothermal vent fauna. Cah. Biol. Mar. 46, 97–104.
Wu, X. Y., Cobbina, S. J., Mao, G. H., Xu, H., Zhang, Z., and Yang, L. Q. (2016). A review of toxicity and mechanisms of individual and mixtures of heavy metals in the environment. Environ. Sci. Poll. Res. 23, 8244–8259. doi: 10.1007/s11356-016-6333-x
Keywords: deep-sea mining, toxicology, manganese nodules, hydrothermal vents, adaptive management
Citation: Hauton C, Brown A, Thatje S, Mestre NC, Bebianno MJ, Martins I, Bettencourt R, Canals M, Sanchez-Vidal A, Shillito B, Ravaux J, Zbinden M, Duperron S, Mevenkamp L, Vanreusel A, Gambi C, Dell'Anno A, Danovaro R, Gunn V and Weaver P (2017) Identifying Toxic Impacts of Metals Potentially Released during Deep-Sea Mining—A Synthesis of the Challenges to Quantifying Risk. Front. Mar. Sci. 4:368. doi: 10.3389/fmars.2017.00368
Received: 10 August 2017; Accepted: 31 October 2017;
Published: 16 November 2017.
Edited by:
Jeroen Ingels, Florida State University, United StatesReviewed by:
Diva Joan Amon, Natural History Museum, United KingdomCopyright © 2017 Hauton, Brown, Thatje, Mestre, Bebianno, Martins, Bettencourt, Canals, Sanchez-Vidal, Shillito, Ravaux, Zbinden, Duperron, Mevenkamp, Vanreusel, Gambi, Dell'Anno, Danovaro, Gunn and Weaver. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chris Hauton, Y2gxMEBub2Muc290b24uYWMudWs=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.