 Emile Brisson-Curadeau
Emile Brisson-Curadeau Allison Patterson
Allison Patterson Shannon Whelan
Shannon Whelan Thomas Lazarus†
Thomas Lazarus† Kyle H. Elliott
Kyle H. Elliott
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Mar. Sci. , 14 November 2017
Sec. Marine Conservation and Sustainability
Volume 4 - 2017 | https://doi.org/10.3389/fmars.2017.00357
This article is part of the Research Topic The Ecological Significance of Marine Top Predators: From Key Stone Species To Environmental Sentinels View all 5 articles
In 1987, David Cairns proposed that a gradient of colony-based measures on seabirds could be used to assess food supply in the ocean. Measures closely tied to the ocean, such as foraging trip duration, would be sensitive to small declines in food supply while measures more closely tied with the nest site, such as reproductive success, would be sensitive to large declines in food supply. The continual refinement of tracking devices holds the potential to clearly link variables measured via seabirds to food supply, possibly extending Cairns' hypothesis. Here, we review the various tests of Cairns' hypothesis, and demonstrate that those tests have had variable success, partly because of the complex and nonlinear relationships between food supply and colony-based measures. We summarize the metrics available from biologgers and argue that such devices can provide a more direct proxy of food supply. We conclude that Cairns' hypothesis can be extended to biologger-derived parameters and that seabird behavior can be used as an early warning signal for declining food supply.
Fish and other seafood provide important ecosystem services, including provisioning, ecotourism, and promoting ecosystem diversity (Holmlund and Hammer, 1999). Inaccurate or incomplete quantification of seafood populations can lead to overfishing and ecosystem collapse, ultimately impacting nearby human communities (Myers et al., 1997; Myers and Worm, 2003; Worm et al., 2006). Food supply metrics, like population trends or relative abundance of seafood on different spatial scales, are often at the center of interest for fisheries management, and are typically quantified from commercial catch data. Yet those data can be biased, may miss early life stages that are important for predictive models, and will miss components of ecosystems that may be essential for ecosystem function but not commercially valuable (Richards and Schnute, 1986; Rose and Kulka, 1999; Salthaug and Godø, 2000). Ship-based biological surveys can provide additional data, but are expensive, especially in remote areas, such as the Arctic, and only cover a single trajectory and a single point in space at each time point (Montevecchi, 1993; Furness and Camphuysen, 1997).
Seabirds range widely across oceans, yet return to a single central place, the colony, during the breeding season. In one afternoon at a seabird colony, a biologist can sample an area of ocean that would cost millions of dollars to investigate using a scientific vessel, obtaining much of the information needed for food supply assessment. Indeed, Cairns (1987) proposed that a gradient of colony-based measures of seabird breeding behavior could be used as indicators of food supply. He proposed that nest attendance would be most sensitive to variation in prey availability, followed by chick growth rates, reproductive success, and adult survival. In each case, the relationship would be nonlinear, such that there would be a small range of food supply over which each variable would be able to provide meaningful predictions.
Since Cairns' paper, biologgers (e.g., time-depth recorders, GPS loggers, geolocators, satellite tags) have become a common part of the seabird biologist “tool kit,” allowing for the measurement of several new variables (Jouventin and Weimerskirch, 1990; Ropert-Coudert et al., 2009; Garthe et al., 2011; Regular et al., 2013). These devices allow biologists to follow seabirds out to sea, with some of them (e.g., camera loggers, accelerometers, beak-opening sensors) directly documenting the very moment where a seabird encounters and captures (“samples”) its prey (Simeone and Wilson, 2003; Bost et al., 2008; Watanabe and Takahashi, 2013). Owing to the closer temporal and spatial proximity between the measured variable (seabird behavior) and the variable of interest (food supply), biologgers have the potential for the measured variable to provide more accurate estimates of the variable of interest, including those of the Cairns hypothesis. In particular, whereas Cairns' original parameters could be confounded at the colony by predation, disease, weather, and human disturbance, some biologger-derived parameters may be less directly impacted. Finally, biologgers allow researchers to indirectly assess food supply throughout the year at a higher temporal resolution, and to monitor the fine-scale spatial distribution of food supply. In this paper, we review the evidence in support of Cairns' hypothesis and examine the use of biologgers on seabirds to monitor food supply. We are careful to separate food supply (the underlying variable of interest) from fish stocks (measured by independent metrics meant to approximate food supply, such as fishing catch per unit effort and hydroacoustic surveys) and prey availability (the variable that seabirds directly respond to). As an example, the behavior of a surface-feeding seabird (e.g., black-legged kittiwake) might be used to infer capelin (Mallotus villosus) populations (food supply). Elsewhere, that inference's viability might have been tested by independent hydroacoustic surveys at one life stage (fish stocks). Nonetheless, only capelin near the surface are accessible to kittiwakes (prey availability). Our assumption is that all three variables are related to each other.
We reviewed 489 studies citing Cairns' (1987) seminal paper, or citing later follow-up reviews by Piatt et al. (2007a,b), that tested for a relationship between colony-based parameters (those that do not require the use biologgers) and at least one independent measure of food supply. We concentrated on food supply targeted by the fishing industry, therefore excluding zooplankton. We focused on three categories of colony-based parameters that were most commonly examined (foraging trip duration/colony attendance, chick growth rate/mass, and breeding success; Supplementary Material). Median sample size was 19 colony years (range: 8–47 years), and studies shorter than four years were not included in the review. Although Cairns (1987) predicted that seabirds would respond nonlinearly to variation in food supply, four out of fifteen studies only tested for linear correlations. Of the studies that tested for both linear and nonlinear effects, nonlinear relationships provided the best fits for 32 out of 42 colony-based parameters examined (linear: 3; no relationship: 7). In addition, we completed a non-comprehensive review of the literature to find examples of studies that used biologgers on seabirds to estimate food supply, to assess the potential benefits of biologgers to use seabirds as indicator of food supply.
Eleven studies looked at the effect of prey availability on breeding success, with all of them defining breeding success as the number of fledged chick per breeding pair. Cury et al. (2011) performed a comprehensive review of long-term seabird studies examining the relationship between breeding success and fish stocks; they found that seabird breeding success responded to changes in fish stocks when it was within the lower third of maximum fish stocks. This indicates that breeding success is less sensitive to variation in food supply than Cairns' original hypothesis (Figure 1A). The study by Cury et al. (2011) re-analyzed data from populations examined by other studies included in our review (e.g., Oro and Furness, 2002; Crawford et al., 2006; Piatt et al., 2007a). Although they found only asymptotic relationships (and one non-significant relationship) across species (Cury et al., 2011), the original studies reported the full range of functional responses to changes in fish stocks (no relationship, linear, asymptotic, step), with asymptotic relationships being the most common. There was also great variability in the explanatory power of these relationships (r2: 0.19–0.75). In some cases, the relationship between reproductive success and fish stocks was only significant when certain years with exceptionally low breeding success were excluded (e.g., Crawford et al., 2006; Wanless et al., 2007), indicating that other drivers of reproductive failure can confound this relationship, in addition to food availability.
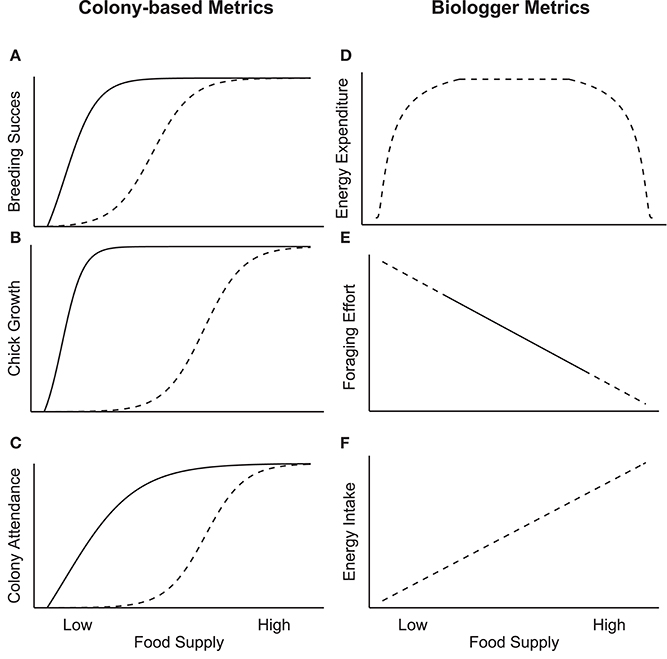
Figure 1. Theoretical (dashed lines) and observed (solid lines) relationships between food supply and colony-based parameters (A–C; adapted from Cairns, 1987) or biologging parameters (D–F). Cairns' original parameters are sensitive to low food supply, while biologging parameters are theoretically sensitive to a broader range of food supply.
The link between chick growth and prey availability was the subject of four studies. In five cases where an asymptote or step function was reported for chick growth rate/mass, the variation occurred within the lowest 25% of measured fish stocks, indicating that chick size is less sensitive to variation in food supply than Cairns' original hypothesis implied (Figure 1B). Three cases reported no relationship between chick growth rates/mass and fish stocks, one case reported a negative linear relationship (Österblom et al., 2006), and one case reported a weak positive relationship (Cohen et al., 2014). The explanatory power of these models was generally weak, with only one study reporting r2 values >0.5 (Piatt et al., 2007a). In studies that considered both chick growth rates and mass, the relationship with fish stocks was not always consistent; Suryan et al. (2006) found that fish stocks predicted chick growth rate but not mass, while Dänhardt and Becker (2011) found that fish stocks predicted chick mass but not growth rate at one colony and no relationship with fish stocks at another. Finally, one study examined feeding rates (Harding et al., 2007b), presumably representative of chick growth rates, but relationships were driven by differences between colonies monitored by sight or by video camera, with the possibility that observations by sight missed some feeding events. Chick growth measures are probably not well correlated with fish stock because breeding adults, especially older ones, likely compensate in years with low prey availability by increasing foraging effort.
Cairns (1987) predicted that measures of activity budgets and colony attendance would be sensitive to a broader range of variation in food supply than measures of reproductive success or chick growth rate (Figure 1C). A very limited number of cases—four studies looking at three colony/species combinations—examined foraging trip duration or colony attendance. The one study that considered foraging trip duration (Piatt et al., 2007a) reported a negative exponential relationship with fish stocks, where trip duration varied across the entire range of fish stocks. Three cases considered colony attendance, and a different relationship was reported for each species; only one species (common murre, Uria aalge) showed a strong relationship with fish stocks, where attendance during chick rearing varied in the lower 60% of observed fish stocks (Harding et al., 2007a,b; Piatt et al., 2007a). Furthermore, the relationship between colony attendance and fish stocks was driven by differences between colonies monitored by sight or by video camera (Harding et al., 2007a,b; Piatt et al., 2007a), and it is possible that visual observations missed some attendance periods. While there is suggestive evidence that foraging and attendance parameters are sensitive to changes in prey availability, the lack of studies that consider these parameters speaks to the difficulty of data collection without use of biologgers.
Our review found conflicting trends that challenge the utility of colony-based parameters as indicators of food supply. Studies that tested for relationships between colony-based measures and multiple prey types or age classes reported different trends, depending on the prey types or age classes considered (e.g., Oro and Furness, 2002; Wanless et al., 2007; Dänhardt and Becker, 2011; Santora et al., 2014). Thus, seabirds may be indicators of only a specific subset of food supply in some cases (e.g., certain species, age classes within species). However, seabirds may preferentially select specific prey or switch to alternate prey depending on conditions (Suryan et al., 2002), which could also obscure relationships with specific food supply. While Cairns' (1987) hypothesis specifically addresses total biomass, additional variation with food supply, including average prey mass (Österblom et al., 2006), timing of prey arrival (Wanless et al., 2007), and age class (Oro and Furness, 2002; Dänhardt and Becker, 2011), can complicate the relationship between food supply and colony-based parameters. Furthermore, studies examined colony-based measures at different breeding stages (i.e., incubation, chick-rearing), which could add further noise in the response to prey availability. A mismatch between the timing of measurements of fish stocks and seabird responses to prey availability could also obscure our ability to detect relationships even when they are present.
Responses to fish stocks varied between (e.g., Piatt et al., 2007a) and within species (e.g., Dänhardt and Becker, 2011). Differences in responses to variation in fish stocks among species may be related to foraging mode. For example, the relationships between fish stocks and foraging trip duration/colony attendance parameters were weaker in black-legged kittiwakes (Rissa tridactyla; surface foragers) than in common murres (divers), possibly because murres can reallocate resting time to foraging effort when food is scarce (Piatt et al., 2007a). Although breeding success of surface foragers is often considered more sensitive to low prey availability than that of diving foragers (Furness and Tasker, 2000), we found little evidence that the strength of the relationship is stronger for surface foragers than divers, especially as many of the studies reviewed focused on two species, black-legged kittiwakes or common murres. Furthermore, hydroacoustic and trawling surveys of food supply may better-represent prey available to diving birds, while aerial surveys (e.g., Suryan et al., 2006) are more representative of prey available to surface foragers. Interestingly, different colonies of the same species showed different responses to variation in fish stocks (common terns, Sterna hirundo: Dänhardt and Becker, 2011; black-legged kittiwakes: Piatt et al., 2007a vs. Suryan et al., 2006).
Although it is clear that variation in prey availability influences seabird behavior and breeding success, these relationships are generally weak and nonlinear. The colony-based parameters examined may identify coarse changes in food supply, but are poor predictors of smaller changes, and therefore of limited utility as indicators of changes in food supply.
Biologging provides new opportunities to use seabirds as indicators of marine food supply (Table 1). The consensus from the studies in Table 1 is that biologgers are useful tools that demonstrate how seabirds adjust their foraging behavior to fish stocks and distribution, sometimes showing strong correlations (Litzow and Piatt, 2003; Bertrand et al., 2012; Cohen et al., 2014; Boyd et al., 2015). Three studies found a linear correlation between fish stocks and foraging parameters (Litzow and Piatt, 2003; Bertrand et al., 2012; Cohen et al., 2014), which suggests that foraging distance from the colony, diving depth, and diving activity may be good candidate parameters for detection of changes in food supply. Most studies emphasized that because it is more difficult to collect fish stock data than seabird foraging behavior data, few can link the two—which is, after all, why seabirds are examined as potential indicators of food supply. Five studies collected data over five or fewer colony years, making it difficult to determine whether the relationship between bird behavior and fish stocks is linear or nonlinear (Monaghan et al., 1994; Suryan et al., 2006; Pichegru et al., 2010; Angel et al., 2015). Biologgers also record many foraging parameters, which were not all considered by each study; therefore, some studies might have missed a potentially important correlation.
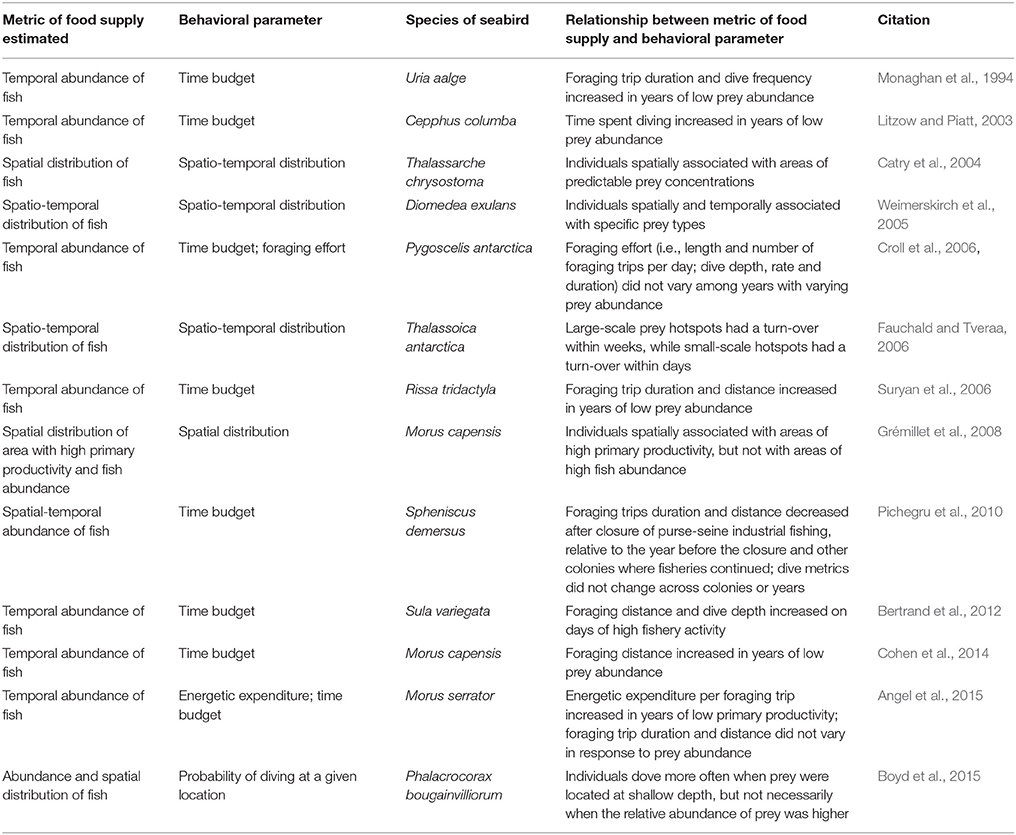
Table 1. Studies that used biologgers to link seabird foraging parameters to food supply (variation in time, space, or both).
Similar to the case for colony-based metrics, the link between fish stocks and seabird distribution likely varies among species (Cairns, 1987), which could explain differences among studies. Although several studies found correlations between the distribution of birds and prey (Catry et al., 2004; Weimerskirch et al., 2005; Fauchald and Tveraa, 2006), one study found no relationship between foraging effort and fish stocks (Croll et al., 2006), or only with primary productivity, rather than fish stocks (Grémillet et al., 2008). Thus, species may vary in their responses to food supply in both colony-based reproductive parameters, and biologger-derived foraging parameters. Furthermore, biologging also potentially suffers from the same bias as colony monitoring, as factors other than prey availability could also explain changes in behaviors. Most considerations described for colony-based measures, like differences among colonies, also likely play a role in the differences observed.
Even if some biologger-derived parameters were sometimes reported to be linearly correlated with fish stocks (Litzow and Piatt, 2003; Bertrand et al., 2012; Cohen et al., 2014), other parameters were not. Indeed, seabird energy expenditure is often thought to be at a ceiling over a large range of food supply (Elliott et al., 2014; Figure 1D). Years with low food supply may prevent birds from expending high energy while birds may not need to expend high energy expenditure during years of high food supply. Based on the relationships reported in Table 1, we propose an extension of Cairns' hypothesis that includes biologger-derived parameters (Figure 1D–F). In contrast to energy expenditure, foraging effort (i.e., number of dives, distance traveled, flight duration) was often reported to be linearly related to fish stocks (Table 1), at least over the range of fish stocks measured (Figure 1E). We argue that energy intake, or prey encounter rate, should be even more closely linked to fish stocks, and the relationship is likely to be linear given that encounter rate is likely proportional to abundance (Figure 1F). This relationship remains untested, but could likely be verified via accelerometers, beak-opening recorders, and esophagus temperature recorders (Simeone and Wilson, 2003; Hanuise et al., 2010; Watanabe and Takahashi, 2013).
The use of biologgers can provide an opportunity to extend the Cairns' hypothesis, by adding new parameters that may have a more linear relationship with fish stocks in at least some seabird species. Some of those parameters may be sensitive over the entire range of food supply, obviating the need for multiple parameters. A major advantage of biologging is the potential to estimate food supply over multiple spatial and temporal scales. Year-round biologger deployments (e.g., Daunt et al., 2014; Fayet et al., 2016) can document year-round variation in food supply over large scales. Repeated biologger deployments on the same individuals over multiple years (e.g., Daunt et al., 2014) will allow us to examine inter- and intra-individual responses to variation in food supply. Inter-species variation is also likely important. Such issues can be addressed with a larger sample size, which is now possible due to the reducing cost of biologgers.
Relatively few biologging studies compared biologger-derived measures of foraging behavior with independent measures of food supply, and those that did had small sample sizes relative to the studies of colony-based reproductive parameters. We recommend that biologgers become integral components of long-term seabird monitoring studies. With the aggregation of data over many years, tests of nonlinear relationships will become more robust.
We encourage the use of biologging devices with multiple channels, such as GPS-accelerometers. Accelerometers provide direct estimates of prey capture rates and energy expenditure over the entire range of food supply variation (Wilson et al., 2006; Watanabe and Takahashi, 2013; Stothart et al., 2016), and coupling GPS locations with accelerometer data could allow the modeling of energy landscapes, with net energy intake peaks corresponding to regions of high food supply. Regardless, biologgers provide a variety of potential foraging parameters, and we suggest that studies examine multiple parameters simultaneously.
Biologging improves our ability to quantify foraging behavior of seabirds, which in turn improves our ability to use seabirds as indicators of food supply. Thus, seabird behavioral parameters derived from biologging devices may act as an early warning signal for declining food supply, which is valuable for fisheries managers and marine biologists alike. The addition of biologging technology as a tool for studying marine birds has made the use of seabirds as indicators of food supply even more promising than in 1987.
KE wrote the first section, AP and SW wrote the second section, TL and EB-C wrote the third section. All individuals edited the final version. Final authorship order was determined from the success of their brackets in the 2017 March Mammal Madness tournament.
Funding was provided by the Canada Research Chair in Arctic Ecology (KE), CREATE-EI (AP), a CGS-M (EB-C), Discovery Grant (KE) and Strategic Research Grant (SW) from the Natural Sciences and Engineering Research Council of Canada, MITACS (TL) and Fonds de recherche du Quebec: Nature et technologies (EB-C).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2017.00357/full#supplementary-material
Angel, L. P., Barker, S., Berlincourt, M., Tew, E., Warwick-Evans, V., and Arnould, J. P. (2015). Eating locally: Australasian gannets increase their foraging effort in a restricted range. Biol. Open 4, 1298–1305. doi: 10.1242/bio.013250
Bertrand, S., Joo, R., Arbulu Smet, C., Tremblay, Y., Barbraud, C., and Weimerskirch, H. (2012). Local depletion by a fishery can affect seabird foraging. J. Appl. Ecol. 49, 1168–1177. doi: 10.1111/j.1365-2664.2012.02190.x
Bost, C. A., Jaeger, A., Huin, W., Koubbi, P., Halsey, L. G., Hanuise, N., et al. (2008). “Monitoring prey availability via data loggers deployed on seabirds: advances and present limitations,” in 5th World Fisheries Congress :Fisheries for Global Welfare and Environment, eds K. Tsukamoto, T. Kawamura, T. Takeuchi, T. D. Jr., Beard, and M. J. Kaise (Tokyo: Terra Publishers), 121–137.
Boyd, C., Castillo, R., Hunt, G. L., Punt, A. E., VanBlaricom, G. R., Weimerskirch, H., et al. (2015). Predictive modelling of habitat selection by marine predators with respect to the abundance and depth distribution of pelagic prey. J. Anim. Ecol. 84, 1575–1588. doi: 10.1111/1365-2656.12409
Catry, P., Phillips, R. A., Phalan, B., Silk, J. R., and Croxall, J. P. (2004). Foraging strategies of grey-headed albatrosses Thalassarche chrysostoma: integration of movements, activity and feeding events. Mar. Ecol. Prog. Ser. 261–273. doi: 10.3354/meps280261
Cohen, L. A., Pichegru, L., Grémillet, D., Coetzee, J., Upfold, L., and Ryan, P. G. (2014). Changes in prey availability impact the foraging behaviour and fitness of cape gannets over a decade. Mar. Ecol. Prog. Ser. 505, 281–293. doi: 10.3354/meps10762
Crawford, R. J., Barham, P. J., Underhill, L. G., Shannon, L. J., Coetzee, J. C., Dyer, B. M., et al. (2006). The influence of food availability on breeding success of African penguins Spheniscus demersus at Robben Island, South Africa. Biol. Conserv. 132, 119–125. doi: 10.1016/j.biocon.2006.03.019
Croll, D. A., Demer, D. A., Hewitt, R. P., Jansen, J. K., Goebel, M. E., and Tershy, B. R. (2006). Effects of variability in prey abundance on reproduction and foraging in chinstrap penguins (Pygoscelis antarctica). J. Zool. 269, 506–513. doi: 10.1111/j.1469-7998.2006.00090.x
Cury, P. M., Boyd, I. L., Bonhommeau, S., Anker-Nilssen, T., Crawford, R. J., Furness, R. W., et al. (2011). Global seabird response to forage fish depletion—one-third for the birds. Science 334, 1703–1706. doi: 10.1126/science.1212928
Dänhardt, A., and Becker, P. H. (2011). Herring and sprat abundance indices predict chick growth and reproductive performance of common terns breeding in the Wadden Sea. Ecosystems 14, 791–803. doi: 10.1007/s10021-011-9445-7
Daunt, F., Reed, T. E., Newell, M., Burthe, S., Phillips, R. A., Lewis, S., et al. (2014). Longitudinal bio-logging reveals interplay between extrinsic and intrinsic carry-over effects in a long-lived vertebrate. Ecology 95, 2077–2083. doi: 10.1890/13-1797.1
Elliott, K. H., Vaillant, M., Kato, A., Gaston, A. J., Ropert-Coudert, Y., Hare, J. F., et al. (2014). Age-related variation in energy expenditure in a long-lived bird within the envelope of an energy ceiling. J. Anim. Ecol. 83, 136–146. doi: 10.1111/1365-2656.12126
Fauchald, P., and Tveraa, T. (2006). Hierarchical patch dynamics and animal movement pattern. Oecologia 149, 383–395. doi: 10.1007/s00442-006-0463-7
Fayet, A. L., Freeman, R., Shoji, A., Kirk, H. L., Padget, O., Perrins, C. M., et al. (2016). Carry-over effects on the annual cycle of a migratory seabird: an experimental study. J. Anim. Ecol. 85, 1516–1527. doi: 10.1111/1365-2656.12580
Furness, R. W., and Camphuysen, K. C. (1997). Seabirds as monitors of the marine environment. ICES J. Mar. Sci. 54, 726–737. doi: 10.1006/jmsc.1997.0243
Furness, R. W., and Tasker, M. L. (2000). Seabird-fishery interactions: quantifying the sensitivity of seabirds to reductions in sandeel abundance, and identification of key areas for sensitive seabirds in the North Sea. Mar. Ecol. Prog. Ser. 202, 253–264. doi: 10.3354/meps202253
Garthe, S., Montevecchi, W. A., and Davoren, G. K. (2011). Inter-annual changes in prey fields trigger different foraging tactics in a large marine predator. Limnol. Oceanogr. 56, 802–812. doi: 10.4319/lo.2011.56.3.0802
Grémillet, D., Lewis, S., Drapeau, L., van Der Lingen, C. D., Huggett, J. A., Coetzee, J. C., et al. (2008). Spatial match–mismatch in the Benguela upwelling zone: should we expect chlorophyll and sea-surface temperature to predict marine predator distributions? J. Appl. Ecol. 45, 610–621. doi: 10.1111/j.1365-2664.2007.01447.x
Hanuise, N., Bost, C. A., Huin, W., Auber, A., Halsey, L. G., and Handrich, Y. (2010). Measuring foraging activity in a deep-diving bird: comparing wiggles, oesophageal temperatures and beak-opening angles as proxies of feeding. J. Exp. Biol. 213, 3874–3880. doi: 10.1242/jeb.044057
Harding, A. M., Piatt, J. F., and Schmutz, J. A. (2007a). Seabird behavior as an indicator of food supplies: sensitivity across the breeding season. Mar. Ecol. Prog. Ser. 352, 269–274. doi: 10.3354/meps07072
Harding, A. M. A., Piatt, J. F., Schmutz, J. A., Shultz, M. T., Van Pelt, T. I., Kettle, A. B., et al. (2007b). Prey density and the behavioral flexibility of a marine predator: the common murre (Uria aalge). Ecology 88, 2024–2033. doi: 10.1890/06-1695.1
Holmlund, C. M., and Hammer, M. (1999). Ecosystem services generated by fish populations. Ecol. Econ. 29, 253–268. doi: 10.1016/S0921-8009(99)00015-4
Jouventin, P., and Weimerskirch, H. (1990). Satellite tracking of wandering albatrosses. Nature 343, 746–748. doi: 10.1038/343746a0
Litzow, M. A., and Piatt, J. F. (2003). Variance in prey abundance influences time budgets of breeding seabirds: evidence from pigeon guillemots Cepphus columba. J. Avian Biol. 34, 54–64. doi: 10.1034/j.1600-048X.2003.02995.x
Monaghan, P., Walton, P., Wanless, S., Uttley, J. D., and Bljrns, M. D. (1994). Effects of prey abundance on the foraging behaviour, diving efficiency and time allocation of breeding guillemots Uria aalge. IBIS Int. J. Avian Sci. 136, 214–222. doi: 10.1111/j.1474-919X.1994.tb01087.x
Montevecchi, W. A. (1993). “Birds as indicators of change in marine prey stocks,” in Birds as Monitors of Environmental Change, eds R. W. Furness and J. J. D. Greenwood (Beckenham: Springer), 217–266.
Myers, R. A., Hutchings, J. A., and Barrowman, N. J. (1997). Why do fish stocks collapse? The example of cod in Atlantic Canada. Ecol. Appl. 7, 91–106. doi: 10.1890/1051-0761(1997)007[0091:WDFSCT]2.0.CO;2
Myers, R. A., and Worm, B. (2003). Rapid worldwide depletion of predatory fish communities. Nature 423, 280–283. doi: 10.1038/nature01610
Oro, D., and Furness, R. W. (2002). Influences of food availability and predation on survival of kittiwakes. Ecology 83, 2516–2528. doi: 10.1890/0012-9658(2002)083[2516:IOFAAP]2.0.CO;2
Österblom, H., Casini, M., Olsson, O., and Bignert, A. (2006). Fish, seabirds and trophic cascades in the Baltic Sea. Mar. Ecol. Prog. Ser. 323, 233–238. doi: 10.3354/meps323233
Piatt, J. F., Harding, A. M., Shultz, M., Speckman, S. G., van Pelt, T. I., Drew, G. S., et al. (2007a). Seabirds as indicators of marine food supplies: Cairns revisited. Mar. Ecol. Prog. Ser. 352, 221–234. doi: 10.3354/meps07078
Piatt, J. F., Sydeman, W. J., and Wiese, F. (2007b). A modern role for seabirds as indicators. Mar. Ecol. Prog. Ser. 352, 199–204. doi: 10.3354/meps07070
Pichegru, L., Grémillet, D., Crawford, R. J. M., and Ryan, P. G. (2010). Marine no-take zone rapidly benefits endangered penguin. Biol. Lett. 6, 498–501. doi: 10.1098/rsbl.2009.0913
Regular, P., Hedd, A., Montevecchi, W. A., Robertson, G. J., Storey, A. E., and Walsh, C. J. (2013). Why timing is everything: energetic costs and reproductive consequences of resource mismatch for a chick-rearing seabird. Ecosphere 5:155. doi: 10.1890/ES14-00182.1
Richards, L. J., and Schnute, J. T. (1986). An experimental and statistical approach to the question: is CPUE an index of abundance? Can. J. Fish. Aquat. Sci. 43, 1214–1227.
Ropert-Coudert, Y., Beaulieu, M., Hanuise, N., and Kato, A. (2009). Diving into the world of biologging. Endanger. Species Res. 10, 21–27. doi: 10.3354/esr00188
Rose, G. A., and Kulka, D. W. (1999). Hyperaggregation of fish and fisheries: how catch-per-unit- effort increased as the northern cod (Gadus morhua) declined. Can. J. Fish. Aquat. Sci. 56, 118–127. doi: 10.1139/f99-207
Santora, J. A., Schroeder, I. D., Field, J. C., Wells, B. K., and Sydeman, W. J. (2014). Spatio- temporal dynamics of ocean conditions and forage taxa reveal regional structuring of seabird–prey relationships. Ecol. Appl. 24, 1730–1747. doi: 10.1890/13-1605.1
Simeone, A., and Wilson, R. P. (2003). In-depth studies of Magellanic penguin (Spheniscus magellanicus) foraging: can we estimate prey consumption by perturbations in the dive profile? Mar. Biol. 143, 825–831. doi: 10.1007/s00227-003-1114-8
Stothart, M. R., Elliott, K. H., Wood, T., Hatch, S. A., and Speakman, J. R. (2016). Counting calories in cormorants: dynamic body acceleration predicts daily energy expenditure measured in pelagic cormorants. J. Exp. Biol. 219, 2192–2200. doi: 10.1242/jeb.130526
Suryan, R. M., Irons, D. B., Brown, E. D., Jodice, P. G., and Roby, D. D. (2006). Site-specific effects on productivity of an upper trophic-level marine predator: bottom-up, top-down, and mismatch effects on reproduction in a colonial seabird. Prog. Oceanogr. 68, 303–328. doi: 10.1016/j.pocean.2006.02.006
Suryan, R. M., Irons, D. B., Kaufman, M., Benson, J., Jodice, P. G., Roby, D. D., et al. (2002). Short-term fluctuations in forage fish availability and the effect on prey selection and brood-rearing in the black-legged kittiwake Rissa tridactyla. Mar. Ecol. Prog. Ser. 236, 273–287. doi: 10.3354/meps236273
Wanless, S., Frederiksen, M., Daunt, F., Scott, B. E., and Harris, M. P. (2007). Black-legged kittiwakes as indicators of environmental change in the North Sea: evidence from long- term studies. Prog. Oceanogr. 72, 30–38. doi: 10.1016/j.pocean.2006.07.007
Watanabe, Y. Y., and Takahashi, A. (2013). Linking animal-borne video to accelerometers reveals prey capture variability. Proc. Natl. Acad. Sci. U.S.A. 110, 2199–2204. doi: 10.1073/pnas.1216244110
Weimerskirch, H., Gault, A., and Cherel, Y. (2005). Prey distribution and patchiness: factors in foraging success and efficiency of wandering albatrosses. Ecology 86, 2611–2622. doi: 10.1890/04-1866
Wilson, R. P., White, C. R., Quintana, F., Halsey, L. G., Liebsch, N., Martin, G. R., et al. (2006). Moving towards acceleration for estimates of activity-specific metabolic rate in free- living animals: the case of the cormorant. J. Anim. Ecol. 75, 1081–1090. doi: 10.1111/j.1365-2656.2006.01127.x
Keywords: biologging, early warning signals, ecological indicators, marine birds, movement ecology
Citation: Brisson-Curadeau E, Patterson A, Whelan S, Lazarus T and Elliott KH (2017) Tracking Cairns: Biologging Improves the Use of Seabirds as Sentinels of the Sea. Front. Mar. Sci. 4:357. doi: 10.3389/fmars.2017.00357
Received: 19 August 2017; Accepted: 24 October 2017;
Published: 14 November 2017.
Edited by:
Filipe Rafael Ceia, University of Coimbra, PortugalReviewed by:
Alastair Martin Mitri Baylis, South Atlantic Environmental Research Institute, Falkland IslandsCopyright © 2017 Brisson-Curadeau, Patterson, Whelan, Lazarus and Elliott. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kyle H. Elliott, a3lsZS5lbGxpb3R0QG1jZ2lsbC5jYQ==
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.