 Bryan Wilson
Bryan Wilson Oliver Müller
Oliver Müller Eva-Lena Nordmann
Eva-Lena Nordmann Lena Seuthe
Lena Seuthe Gunnar Bratbak
Gunnar Bratbak Lise Øvreås
Lise Øvreås- 1Marine Microbiology Research Group, University of Bergen, Bergen, Norway
- 2Department of Arctic and Marine Biology, UiT - The Arctic University of Norway, Tromsø, Norway
As the global climate changes, the higher latitudes are seen to be warming significantly faster. It is likely that the Arctic biome will experience considerable shifts in ice melt season length, leading to changes in photoirradiance and in the freshwater inputs to the marine environment. The exchange of nutrients between Arctic surface and deep waters and their cycling throughout the water column is driven by seasonal change. The impacts, however, of the current global climate transition period on the biodiversity of the Arctic Ocean and its activity are not yet known. To determine seasonal variation in the microbial communities in the deep water column, samples were collected from a profile (1-1000 m depth) in the waters around the Svalbard archipelago throughout an annual cycle encompassing both the polar night and day. High-throughput sequencing of 16S rRNA gene amplicons was used to monitor prokaryote diversity. In epipelagic surface waters (<200 m depth), seasonal diversity varied significantly, with light and the corresponding annual phytoplankton bloom pattern being the primary drivers of change during the late spring and summer months. In the permanently dark mesopelagic ocean depths (>200 m), seasonality subsequently had much less effect on community composition. In summer, phytoplankton-associated Gammaproteobacteria and Flavobacteriia dominated surface waters, whilst in low light conditions (surface waters in winter months and deeper waters all year round), the Thaumarchaeota and Chloroflexi-type SAR202 predominated. Alpha-diversity generally increased in epipelagic waters as seasonal light availability decreased; OTU richness also consistently increased down through the water column, with the deepest darkest waters containing the greatest diversity. Beta-diversity analyses confirmed that seasonality and depth also primarily drove community composition. The relative abundance of the eleven predominant taxa showed significant changes in surface waters in summer months and varied with season depending on the phytoplankton bloom stage; corresponding populations in deeper waters however, remained relatively unchanged. Given the significance of the annual phytoplankton bloom pattern on prokaryote diversity in Arctic waters, any changes to bloom dynamics resulting from accelerated global warming will likely have major impacts on surface marine microbial communities, those impacts inevitably trickling down into deeper waters.
1. Introduction
Polar regions are vulnerable and most sensitive to global climate change. Therefore, there is an increasing research focus needed on these high latitude environments. During the annual cycle, the poles undergo some of the most extreme environmental changes on the planet, from that of the subzero permanently dark winter to the relative warmth and perpetual daylight of the summer. The extensive biodiversity of these regions is understandably well-adapted to these periodic shifts but accelerated atmospheric warming is irrefutably altering conditions here and therefore needs to be studied in detail. The Arctic is warming three times faster than the global mean warming rate (Trenberth et al., 2007) and the extent and thickness of sea ice in the polar oceans is steadily decreasing (Chen et al., 2009), at a rate of up to ten percent per year (Comiso et al., 2008). Recent decades have also seen the summer melt season increase in length (Markus et al., 2009) and the percentage of thin first-year ice increase as compared with thicker multiyear ice (MYI) (Comiso, 2012), such that the late summer Arctic Ocean may be ice-free before the end of the twenty-first century (Boe et al., 2009) or sooner still (Kerr, 2012). Some of these striking environmental changes occurring in the Arctic are related to the inflow of Atlantic water to the Arctic Ocean. The West Spitsbergen Current passing through the eastern Fram Strait is the most significant inflow to the Arctic Ocean and it has intensified over the last decades (Schauer et al., 2004). Increased inflow of the warm and highly saline Atlantic Water affects water column stability and is also probably one of the main drivers of the recent sea ice loss north of Svalbard (Onarheim et al., 2014; Randelhoff et al., 2015). The cumulative consequence of these effects has been the exposure of polar seas to increasing levels of solar radiation (Perovich et al., 2007). Furthermore, enhanced permafrost thawing (Romanovsky et al., 2010) in concert with the profound influence of several large river systems (an hydrology peculiar to the Arctic) (Anderson, 2002) and a greater erosion of exposed coastlines (Lantuit et al., 2012), is leading to an increased terrigenous input of carbon to the Arctic Ocean (Frey and McClelland, 2009). All of which has the potential to radically impact the primary production and successive trophic levels of the polar marine environment (Anderson and Macdonald, 2015).
The greater part of high latitude research has been carried out in the Arctic (predominantly due to the relative logistical ease of working in the region when compared with Antarctica) and the Svalbard archipelago in particular has become a key site for Arctic marine studies, particularly with regards to its being the confluence of both the Arctic and Atlantic Oceans (Hop et al., 2002, 2006; Svendsen et al., 2002). The ocean around the Western coastline of Svalbard is a sea ice-associated pelagic ecosystem (Svendsen et al., 2002) and as with other high latitude locations, seasonal variations in light (and thus in primary production) are more pronounced here than elsewhere. The extreme seasonality of these environmental drivers has revealed a number of trends particular to the polar regions. A single major spring bloom along the retreating ice edge accounts for >50% of the annual primary production around Svalbard and in the Northern Barents Sea (Sakshaug, 2004). By late summer, this develops into a successional post-bloom stage, comprising different phytoplankton populations (Sherr et al., 2003) and it follows, their different associated successional heterotrophic prokaryote consortia, in particular the Flavobacteriia and Gammaproteobacteria classes (Alonso-Sáez et al., 2008; Teeling et al., 2012). This phenomenon of the polar phytoplankton blooms (Williams et al., 2013) followed by the heterotrophic bacterial populations also seemingly drives the annual disappearance of the chemolithoautotrophic marine Archaea from surface waters (Kalanetra et al., 2009; Alonso-Sáez et al., 2012; Pedneault et al., 2014) and the subsequent seasonal fluctuations in prokaryote diversity (Murray et al., 1998; Ghiglione and Murray, 2012; Grzymski et al., 2012; Ladau et al., 2013).
A significant fraction of the phytoplankton primary production sinks out of these surface waters (Reigstad et al., 2008), contributing a single annual major input of organic carbon and energy to the microbial communities residing in dark mesopelagic and deep waters. This subsurface realm dominates the global ocean biome and whereas the Arctic Ocean is the shallowest of the five major oceanic divisions, still its average depth is >1000 m deep. These aphotic zones are characterized by higher pressures, lower temperatures and higher inorganic nutrient concentrations than the photic surface waters above (Arístegui et al., 2009; Orcutt et al., 2011). Yet it is these physicochemical factors, in addition to their remoteness from the surface wind effects and solar irradiation that affect the upper layers so, that also determines their characteristic stability (Orcutt et al., 2011). The waters at these depths contain the largest pool of microbes in aquatic systems (Whitman et al., 1998) and play a major role in ocean biogeochemistry, comprising extraordinarily high genetic and metabolic diversity (Arístegui et al., 2009). The marine snow (primarily dissolved and particulate organic matter) produced by the spring and summer phytoplankton blooms in the stratified epipelagic zone is transported down during winter into the mesopelagic zone by convective mixing and subduction after cooling of the sea surface (Arístegui et al., 2009; Grzymski et al., 2012). Chemolithoautotrophic processes (such as archaeal ammonia oxidation) then come into play during the dark winter months (Grzymski et al., 2012) and the resulting nitrate buildup fuels the subsequent phytoplankton spring bloom (Connelly et al., 2014). However, should suggested models of a freshening Arctic be correct (Comeau et al., 2011), surface Arctic basin waters in a warming world may become increasingly stratified, such that the vertical flux of nutrients between deeper waters and the epipelagic zone may be reduced; primary productivity would consequently be lessened and this annual biogeochemical cycle, so essential for Arctic Ocean productivity, would inevitably be disrupted (Tremblay et al., 2008).
As the majority of Arctic studies of marine microbial communities have either been carried out in the more amenable spring and summer seasons or in shallow waters, the primary objective of the present study was to expand upon these data. More specifically, we wished to identify the key mediators of the prokaryotic microbial community in the Atlantic water inflow to the Arctic Ocean during the light-driven summers vs. the dark winter night, seasons characterized by the massive variation in availability of fresh photosynthesis-derived carbon. Additionally, we intended to compare the cold, deep and dark mesopelagic ocean with the cold, shallow and dark surface waters above, to gain more insight into the driving mechanisms resulting from such environmental conditions. High-throughput sequencing technologies have previously highlighted the extreme microbial seasonality of the polar regions (Kirchman et al., 2010; Christman et al., 2011; Connelly et al., 2014). In this study we implement the same technologies to sequence reverse-transcribed total RNA (with its significantly shorter life span than DNA) to provide a timely snapshot of the more metabolically-active fraction of the marine microbial community.
2. Materials and Experimental Methods
2.1. Sampling
Samples were taken from various transects bisecting the West Spitsbergen Current along the coast of Svalbard, a Norwegian archipelago in the Arctic Ocean (Figure 1) from the research vessels RV Lance and RV Helmer Hanssen, operating under either the MicroPolar and Carbon Bridge projects (Table 1). Samples (25–50 L) representative of the water column profile were collected from a range of water masses (defined in Paulsen et al., 2016) between 1m and 1000 m (Table 1) using Niskin bottles mounted on a rosette deployed from the vessels. Water samples were filtered through 0.22 μm Sterivex Filter Units (Merck-Millipore, MA, USA) via a peristaltic pump and frozen at −80°C immediately.
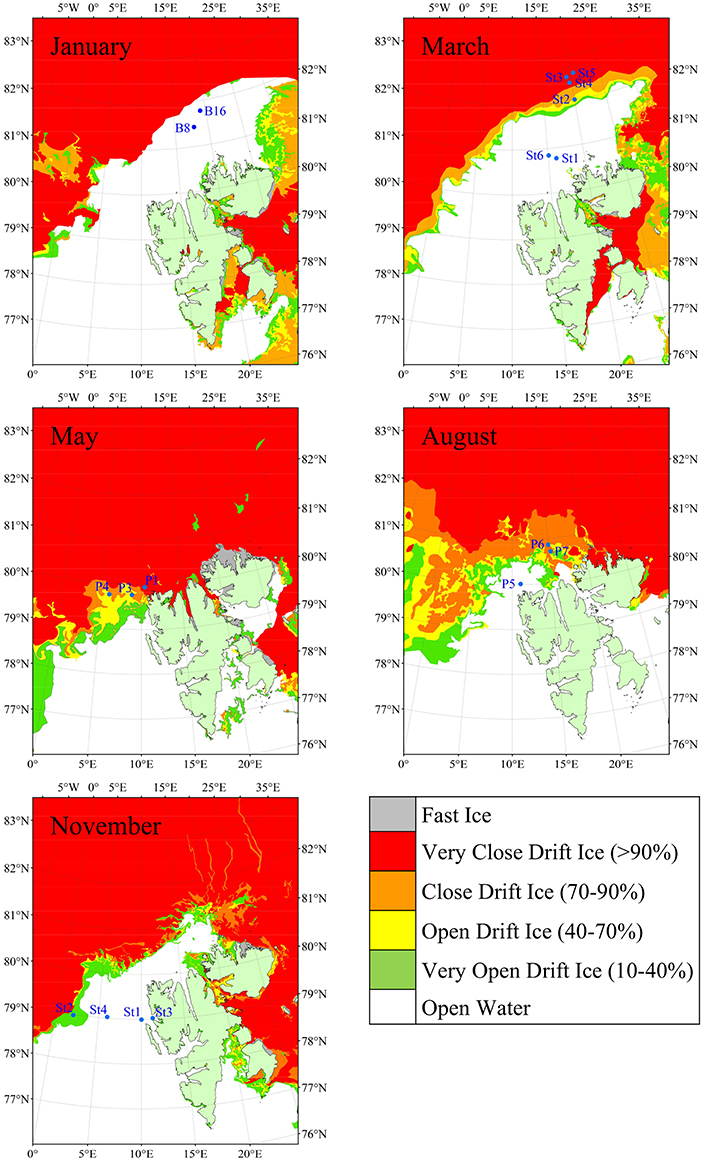
Figure 1. Map showing sampling locations and sea ice extent around the Svalbard Archipelago. Maps were generated using the Global Self-consistent Hierarchical High-resolution Geography (GSHHG) data from the National Geospatial-Intelligence Agency (NGA), distributed under the GNU Lesser General Public License (LGPL). Ice data were provided by the Norwegian Ice Service (MET Norway) for the following dates; January 10, March 7, May 23, August 12, and November 7, 2014.
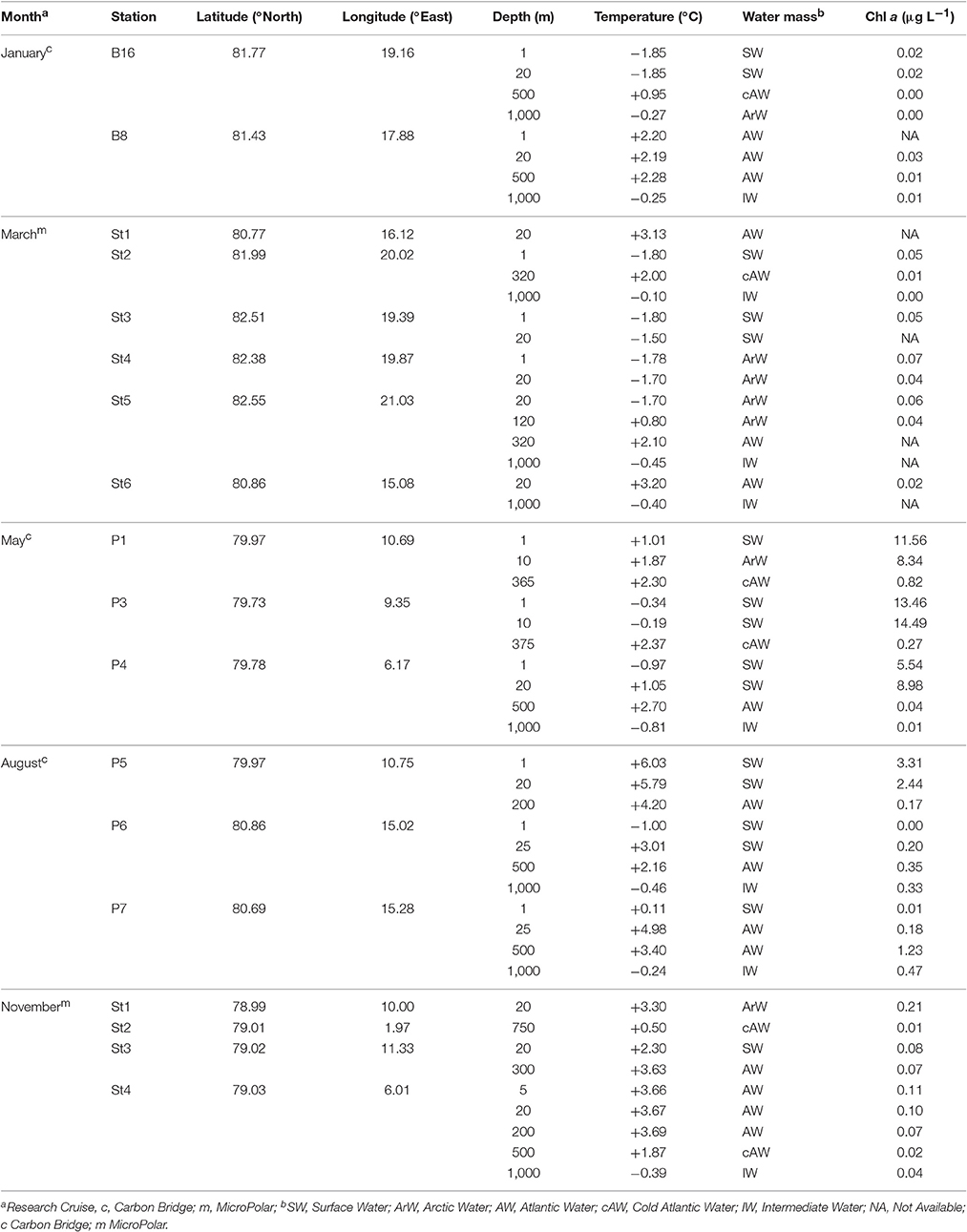
Table 1. Chemical and biological parameters for samples taken from the Arctic Ocean around the Svalbard Archipelago in 2014.
2.2. Chlorophyll a Measurement
The concentration of chlorophyll (chl) a was determined fluorometrically (Parson et al., 1984). Sample water was filtered onto triplicate Whatman GF/F glass-fiber filters (Sigma-Aldrich, MO, USA). The chl a on the filters was immediately extracted in 5 mL methanol (>99.8% v/v) at 4°C in the dark for 12 h without grinding. The fluorescence of the extracts was measured with a fluorometer (Model 10-AU, Turner Designs, CA, USA), calibrated with pure chl a (Sigma-Aldrich).
2.3. RNA Extraction
RNA were extracted directly from filters with the AllPrep DNA/RNA Mini Kit (Qiagen, CA, USA) using a protocol modified from the manufacturer's instructions. Briefly, filters were thawed on ice and an extraction buffer (comprising 990 μL Buffer RLT Plus and 10 μL β-mercaptoethanol per filter) prepared. Extraction buffer (1 mL) was added to each filter and filters vortexed vertically for 2 min, inverted and vortexed for a further 2 min. Lysate was removed from the filter using a 10 mL syringe and transferred to a 1.5 mL microfuge tube. Lysate (700 μL) was loaded on to an AllPrep DNA spin column and centrifuged for 30 s at 8000 × g, saving the flow-through for subsequent RNA extraction. Centrifugation steps were repeated for any remaining lysate volume, as necessary.
One volume 70% (v/v) ethanol was added to the flow-through, mixed by pipetting and 700 μL transferred to an RNeasy spin column in a 2 mL microfuge tube, before centrifugation for 15 s at 8000 × g. Centrifugation was repeated for the remaining liquid volume as necessary, discarding the flow-through each time. Buffer RW1 (700 μL) was added to the RNeasy spin column and the tube centrifuged again for 15 s at 8000 × g and the flow-through discarded. Buffer RPE (500 μL) was added to the RNeasy spin column and the tube centrifuged for 15 s at 8000 × g, discarding the flow-through. Buffer RPE (500 μL) was again added to the RNeasy spin column and the tube centrifuged for 2 min at 8000 × g. The RNeasy spin column was placed in a fresh 2 mL microfuge tube and centrifuged for 1 min at full speed. The RNeasy spin column was placed in a fresh 1.5 mL microfuge tube and 50 μL RNase-free water added to the spin column, before centrifugation for 1 min at 8000 × g. RNase-free water (50 μL) was added to the spin column again and the centrifugation repeated. The eluate (100 μL) was stored at −20°C.
The quantity and quality of RNA were assessed using a Qubit 3.0 Fluorometer (Thermo Fisher Scientific Inc., MA, USA) and by agarose gel electrophoresis.
2.4. Reverse Transcription and PCR for High-Throughput Sequencing (HTS) Library Construction
RNA (10 ng) was treated with the DNA-free DNA Removal kit (Invitrogen, CA, USA), prior to reverse transcription using the SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen), as per the manufacturer's instructions. The V4 region of the 16S rRNA gene was amplified from cDNA using a two-step nested PCR approach. In the first step, triplicate samples were amplified using primers 519F (5′-CAGCMGCCGCGGTAA-3′) (Øvreås et al., 1997) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) (Caporaso et al., 2011). The reaction mixture consisted of 10 μL HotStarTaq Master Mix (Qiagen), 500 nM of each primer, 10 ng of cDNA and nuclease-free water to bring the total volume to 20 μL. Reactions were initially denatured for 15 min at 95°C, followed by 25 cycles of denaturation at 95°C for 20 s, primer annealing at 55°C for 30 s and extension at 72°C for 30 s, followed by a final extension step of 72°C for 7 min. Triplicate amplicons were pooled and purified using the DNA Clean & Concentrator-5 kit (Zymo Research Corporation, CA, USA), as per the manufacturer's instructions, and quantified using a Qubit 3.0 Fluorometer.
In the second step, pooled amplicons were amplified by nested PCR using MID-tagged primers 519F and 806R in a reaction mixture comprising 25 μL HotStarTaq Master Mix, 500 nM of each primer, 50 ng of pooled DNA, and nuclease-free water to bring the total volume to 50 μL. Reactions were initially denatured for 15 min at 95°C, followed by 15 cycles of denaturation at 95°C for 20 s, primer annealing at 62°C for 30 s and extension at 72°C for 30 s. This was followed by a final extension step of 72°C for 7 min. The quantity and quality of RNA was assessed by agarose gel electrophoresis. Amplicons were purified using Agencourt AMPure XP Beads (Beckman Coulter Inc., CA, USA) and quantified again using a Qubit 3.0 Fluorometer and by agarose gel electrophoresis. MID-tagged amplicons were then pooled in equimolar amounts for library construction.
Libraries were sent to the Norwegian Sequencing Centre (Oslo, Norway) for HTS on a MiSeq platform (Illumina, CA, USA) using the MiSeq Reagent Kit v2 (Illumina).
2.5. Bioinformatic Analyses
16S rRNA gene sequences were processed using a custom BioPython (Cock et al., 2009) script incorporating various bioinformatic tools, along with the QIIME pipeline (Version 1.8.0) (Caporaso et al., 2010b). Briefly, FASTQ files were quality end-trimmed at a phred quality score ≥20 using Sickle (Version 1.33) (Joshi and Fass, 2011) and PhiX contaminants and adapters removed using Bowtie 2 (Langmead and Salzberg, 2012) and cutadapt (Martin, 2011), respectively. Paired-end reads were merged using PANDAseq (Masella et al., 2012) and all reads <200 bp removed. The remaining sequence reads were checked for chimeras with the identify chimeric seqs and filter fasta scripts in QIIME, using usearch61 (Edgar, 2010) and the ChimeraSlayer (Haas et al., 2011) reference database (Gold.fa) found in the Broad Microbiome Utilities suite (http://microbiomeutil.sourceforge.net/). The pick de novo otus script in QIIME (using default parameters) was used for de novo OTU picking (using uclust (Edgar, 2010) and a sequence similarity threshold of 97%), taxonomy assignment (using PyNAST, Caporaso et al., 2010a) at 90% sequence similarity against the Greengenes core reference alignment database (Release 13_8) (DeSantis et al., 2006), and finally, the assembly of a table of OTU abundances with taxonomic identifiers for each OTU. OTUs were grouped by different taxonomic levels using the summarize taxa and plot taxa summary QIIME scripts. Rarefaction curves and Chao1 values were calculated using QIIME's alpha rarefaction script, whilst principal coordinate analysis plots used the beta diversity through plots script. All statistical figures were produced using the R Software Environment (R Core Team, 2013). High-throughput sequencing data were submitted to the European Nucleotide Archive (ENA) under Accession Number PRJEB19605.
3. Results
3.1. Environmental Data
The dataset comprised fifty two samples in total, with the numbers of samples taken on each cruise (Table 1) varying with prevailing conditions and ship access time. Summer and winter conditions in the Arctic Ocean around Svalbard are very different (Figure 1) and much of the archipelago is ice-bound during the year, with the dark polar night persisting for almost three months. The study area (Table 1) is hydrographically characterized by Atlantic water masses, either as pure Atlantic water [AW; T (temperature) > 2°C and S (salinity) > 34.92; (Walczowski, 2013) and references therein], or as modified colder water masses, such as cold Atlantic Water (cAW) with 0 < T <2°C (S > 34.9) and Intermediate Water (IW) T <0°C (S > 3 4.9) (de Steur et al., 2014). Arctic water (ArW) was found at some stations and depths. Not all water classified here as ArW necessarily originated from the central Arctic Ocean. This may instead have been water that had undergone freshening and cooling processes, and hence had a density ρθ > 27.7 kg m−3 and S <34.92 (or 34.9 when cooler than 2°C). These are also the physical characteristics for ArW in the central Arctic Ocean. Cold surface water (SW; ρθ > 27.7 kg m−3 and S < 34.92) was encountered at stations within the marginal ice zone, created by sea ice melt. Sea ice extended furthest North during the winter months, and furthest South during May and August, prohibiting sampling north of Svalbard during the summer months. Consequently, most stations sampled in May and August were covered by ice, while all stations sampled during March and November were situated in open waters.
Concentration of chl a (a proxy for phytoplankton biomass) showed a clear seasonal cycle in phytoplankton, with lower concentrations throughout the water column during the dark winter months and March, higher concentrations in surface waters in May, and intermediate concentrations in August (Table 1). Detailed investigations of the phytoplankton showed a major shift from communities dominated by smaller-celled zooplankton during winter and March, to spring bloom communities dominated by large-celled phytoplankton, such as diatoms and Phaeocystis colonies in May (M. Reigstad, Personal Communication). In August, the phytoplankton communities were diverse and dominated again by smaller cells. The community shift from May to August reflects the general shift from nitrate-based phytoplankton communities in May to phytoplankton communities based on regenerated production in August (M. Reigstad, Personal Communication). In the deep mesopelagic waters, there was much less variation in biological and chemical properties than surface waters over the course of the year.
3.2. Prokaryote Diversity
After quality trimming and chimera removal, the complete 16S rRNA dataset (targeting the V4 region) comprised 7 902 016 sequence reads from 52 samples, with on average 127 452 reads per sample, totalling 470 567 OTUs (75.7% of these [356 512 OTUs] were singletons). Prokaryote communities were dominated by taxa typical of marine environments, including the bacterial classes Alphaproteobacteria, Gammaproteobacteria and Chloroflexi-type SAR202 and the archaeal class Thaumarchaeota (Figure 2). The relative abundances of these major classes were seen to vary significantly with season and depth. The Thaumarchaeota, Chloroflexi-type SAR202, AB16 (Marine Group A, originally SAR406) and Deltaproteobacteria were only observed to predominate in waters where light availability was low (in surface waters during the winter months and in deeper waters the year round). Representation of the Gammaproteobacteria and Flavobacteriia however was greatest when light availability and phytoplankton levels were highest (in epipelagic waters in the summer months). In the darker months (January, March and November), the relative abundances of the Alphaproteobacteria were recorded at similar levels irrespective of sample depth. In May however, levels were substantially higher in deeper waters, whilst August saw an abrupt shift to similarly high levels in surface waters.
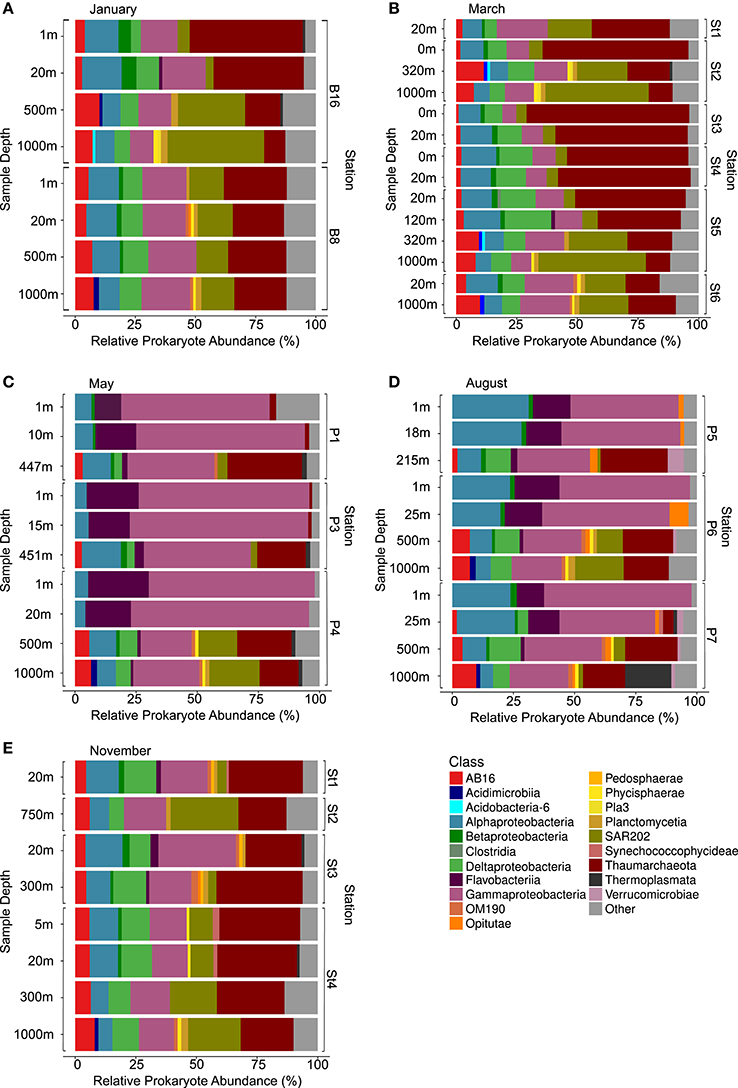
Figure 2. Taxonomic diversity and relative abundance at the class level of prokaryotes from samples collected from the Arctic Ocean at different times, locations and depths. Sequences were taxonomically assigned to the Greengenes database (DeSantis et al., 2006) using QIIME (Caporaso et al., 2010b). For clarity of presentation, unassigned sequences and classes comprising <1% of the total number of sequences within a sample were simply classified as “Other” (gray), whilst sequences affiliated with phylum Cyanobacteria class Chloroplast were removed. (A) January, (B) March, (C) May, (D) August, and (E) November.
Principal coordinate analyses (PCoA) on unweighted UniFrac distances (Figure 3) indicated that light availability and depth (of which light availability is a factor) primarily drove the phylogenetic beta-diversity across prokaryote communities, with three clearly separated clusters; one cluster (Figure 3A) containing communities from deep and dark mesopelagic waters, predominantly those samples collected from January, March and November; a second cluster (Figure 3B) comprising communities from samples collected from shallow and light epipelagic waters in May and August only; and a third cluster (Figure 3C), an admixture of communities from shallow and dark epipelagic waters in January, March and November and mesopelagic waters from May and August. Prokaryote alpha-diversity generally decreased as light availability increased to a maximum in the summer (Figure 4 and Figure S1). Within each sampling period, Chao-1 estimates of OTU richness increased with increasing depth, with deeper waters containing a greater richness than surface waters, with maximum richness over the year observed in the deepest January samples. In surface waters, richness decreased until a minimum in August, before increasing again as light availability decreased during the late autumn.
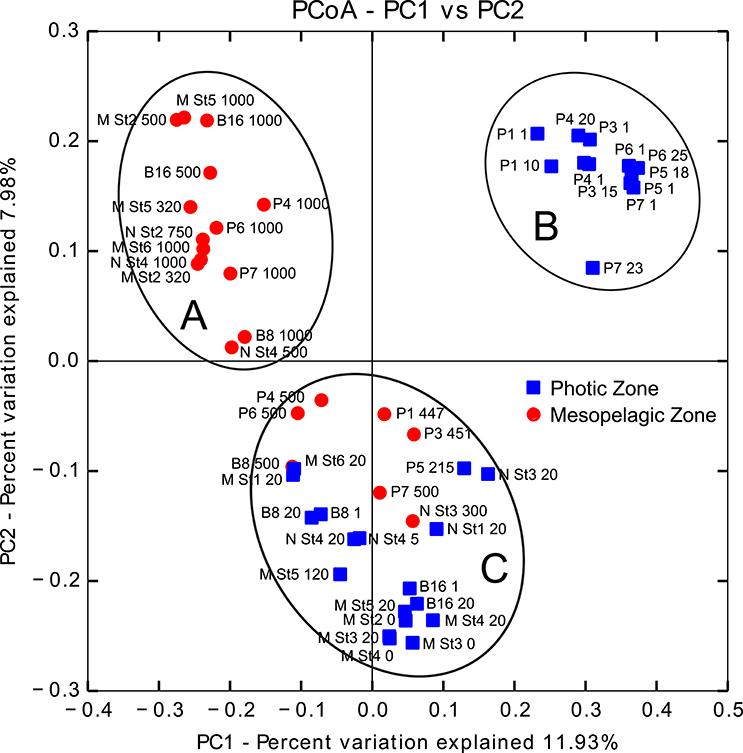
Figure 3. Principal Coordinate Analysis (PCoA) of unweighted UniFrac distances of prokaryote 16S OTUs in putative Epipelagic (<200 m depth; blue squares) and Mesopelagic (200–1000 m depth; red circles) Zones of the Arctic Ocean. Text in the figure at each point refers to sampling station and depth; B8–16 (January), M St1–6 (March), P1–4 (May), P5–7 (August), and N St1–4 (November). PC, principal coordinate. Ellipses around the clusters labeled (A–C) were added to the figure manually for descriptive purposes only.
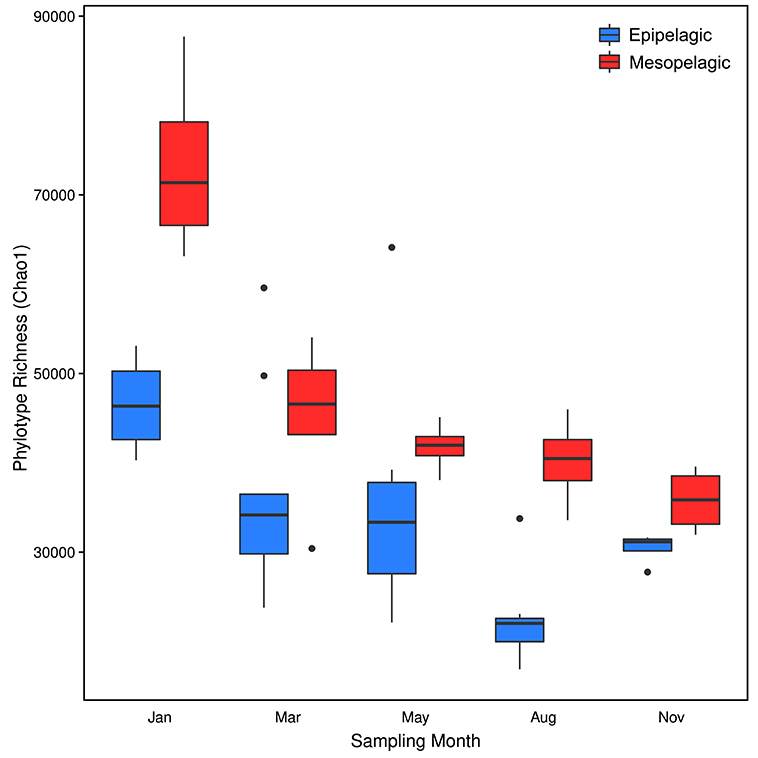
Figure 4. Chao1 estimates of prokaryote 16S OTU richness in putative Epipelagic (<200 m depth) and Mesopelagic (200–1000m depth) Zones of the Arctic Ocean. The ends of the box represent the 25th and 75th percentiles, the whiskers represent minimum and maximum range, the band inside each box represents the median and the black circles represent outliers.
The seasonal and depth variation in the relative abundance of eleven major prokaryote taxa (Figure 5) confirmed the impact of increased light availability and the resulting phytoplankton blooms on Arctic marine microbial populations in epipelagic waters. Generally, the dominant taxa in surface waters were seen to exhibit either a significant positive or negative population change in the summer months, whilst corresponding populations in deeper waters remained relatively unchanged. The relative abundance of 16S rRNAs derived from chloroplasts reached a maximum in May (Figure 5) and this was reflected by maxima in the Oceanospirillaceae, Alteromonadaceae and Flavobacteriaceae and minima in the archaeal family Cenarchaeaceae, Rhodospirillaceae, Nitrospinaceae and OM27, moving from March into May - in all these taxa, relative abundance increased substantially with the transition into the darker winter months. The family AEGEAN185 were unusual in that there was sharp decrease in the May relative abundance in deeper waters, as were the group SAR324 which saw concurrent minima in both surface and deeper waters. The second bloom in August, characterized by a different phytoplankton community to the earlier May one (E. S. Egge, Personal Communication), saw maxima in both the families Halomonadaceae and Rhodobacteraceae.
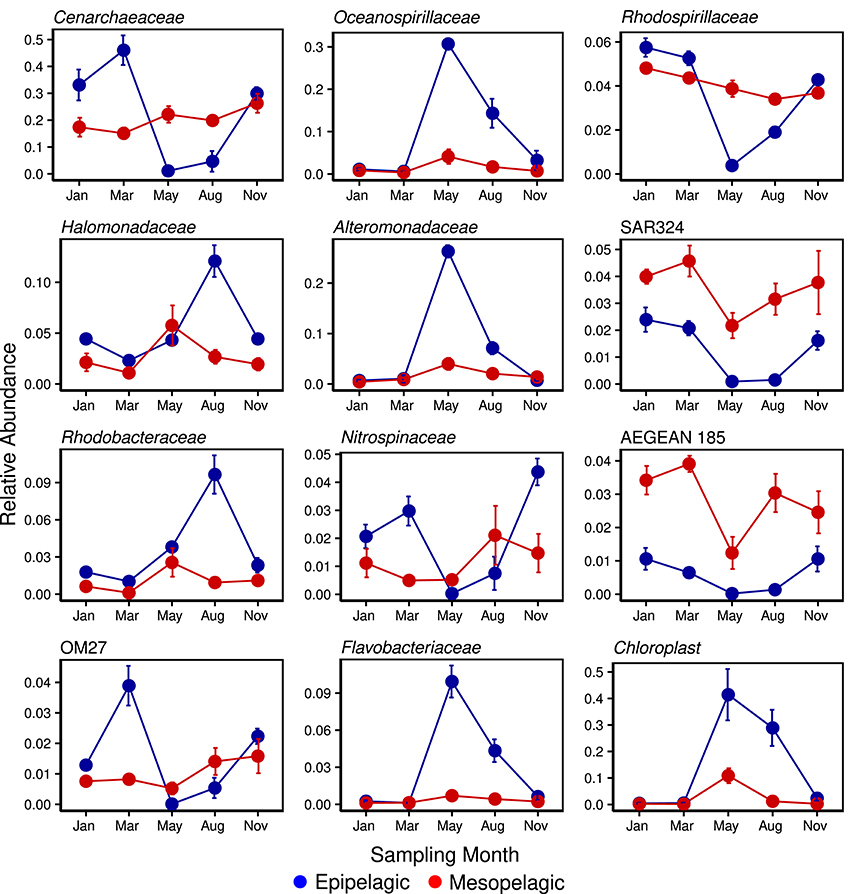
Figure 5. Temporal variability of selected strong indicator prokaryote 16S phylotypes (at the family level) in putative Epipelagic (<200 m depth) and Mesopelagic (200–1000 m depth) Zones of the Arctic Ocean.
4. Discussion
Recent advances in high-throughput sequencing technologies and software development for data management and processing have helped to shed light on the composition and seasonal dynamics of marine microbial communities, suggesting that many follow cyclical and predictable patterns (Fuhrman et al., 2006, 2015). Our study collected water samples in the seas around Svalbard from nominal depths ranging from 1 to 1000 m throughout the year, spanning the whole spectrum of environmental light conditions from the total darkness of the polar night to the perpetual illumination of the polar day. Our analysis of prokaryote 16S ribosomal RNA diversity using high-throughput sequencing confirmed that the microbial communities of Arctic waters during the polar winter and summer varied significantly. Additionally, this data suggested that seasonal and depth-related light availability and sea conditions and the associated phytoplankton blooms are the primary drivers for successional changes in community composition in these waters. Phylogenetic diversity increased with decreasing illumination, with regards to both seasonality and water depth, with the greatest richness to be found in the deepest and darkest water samples. The phytoplankton bloom and post-bloom stages dominated surface water communities in May and August, respectively, and saw corresponding increases in the relative abundance of bloom-associated copiotrophic organisms related to the Gammaproteobacteria and Flavobacteriia. The chemolithotrophic Thaumarchaeota and Chloroflexi-type SAR202 dominated deep aphotic waters all year round but varied significantly in surface waters with varying light levels, proliferating in the dark winter months and diminishing in the well-lit summer months. Whether these taxa were responding directly to the changes in light availability or nutrient composition, or were outcompeted by the resulting phytoplankton blooms and successional prokaryotes is not known. The objectives of this research were to assess these microbial communities in time and space and investigate how these organisms respond down through the water column to the extreme variations in surface environmental conditions during the polar year.
We observed distinct seasonal fluctuations in prokaryote diversity, comprising high richness in the darker autumn and winter months and lower richness in the late spring and summer (Gilbert et al., 2012; El-Swais et al., 2015), similar to other surveys of surface waters in high latitude marine ecosystems (Murray et al., 1998; Murray and Grzymski, 2007; Ghiglione and Murray, 2012; Grzymski et al., 2012; Ladau et al., 2013). Phylotype richness peaked in January and then decreased through the year (Figure 4) until the annual minimum in August, coincident with the late summer phytoplankton post-bloom, before increasing again in the late autumn. Cyclic annual patterns of prokaryote community structure have been observed in waters off the Antarctic Peninsula (Murray et al., 1998; Church et al., 2003; Murray and Grzymski, 2007) and had sampling in our study persisted, we predict that phylotype richness would likely have peaked again in January (Fuhrman et al., 2015). Rarefaction curves (Figure S1) also indicated that there was a distinct change in the number of unique phylotypes with season, which contrasts with seasonal pyrosequencing data for the Western Arctic (Kirchman et al., 2010). However, the authors do concede that the depth of their sequencing efforts may not have captured the complete diversity of these communities. Samples in the current study were rarified to 62500 sequences (tenfold higher than that of Kirchman et al., 2010) and this was sufficient to illustrate seasonal differences in community diversity. Whilst rarefaction curves for the May and August surface samples approached an asymptote, showing that coverage by these libraries was high, sequencing efforts for the winter months were typically undersaturated, as evidenced by the continued upwards curve of the January samples even at 170 000 reads (data not shown).
There was a clear trend in the variation in relative proportions of the two prokaryote domains in surface waters over the annual cycle, with the Archaea increasing to maxima in the winter months and decreasing to almost undetectable levels in the summer months (Figure 5). This was reflected in reverse by the Bacteria, mirroring seasonal observations of near-surface prokaryote communities in the Western Arctic (Alonso-Sáez et al., 2008). In our study, the Proteobacteria were the most abundant bacterial phylum throughout the year and of these, it was the class Gammaproteobacteria that predominated, particularly in the summer months, along with class Flavobacteriia of the phylum Bacteroidetes (Figure 2). The high relative abundance of these taxa in coastal waters around Svalbard (Zeng et al., 2013) has been associated with the release of dissolved organic matter following a phytoplankton bloom (Ghiglione and Murray, 2012; De Corte et al., 2013; Buchan et al., 2014; El-Swais et al., 2015) and they are seen to dominate Arctic MYI communities (Bowman et al., 2012). The Gammaproteobacteria are typically seen to increase toward the summer months (Alonso-Sáez et al., 2008) and of the strong indicator phylotypes determined during the study year, three of the top five (Oceanospirillaceae, Halomondaceae and Alteromonadaceae) were members of this class (Figure 5). Members of both the Oceanospirillaceae and Alteromonadaceae are known to be r-strategist, broad substrate generalists and are frequently seen to be closely-associated with phytoplankton blooms (Teeling et al., 2012; Buchan et al., 2014; El-Swais et al., 2015). However, in comparison with the abundance of physiological and genomic studies of bloom-associated Bacteroidetes and Alphaproteobacteria strains, there is a relative paucity of data regarding the Gammaproteobacteria (Buchan et al., 2014).
Recent whole genome analyses of various members of the Bacteroidetes have confirmed long-held assumptions of a preference for and selective advantage of the phylum when growing on complex organic matter (Abell and Bowman, 2005; Teeling et al., 2012; Fernández-Gómez et al., 2013; Williams et al., 2013), such as that typically produced by marine phytoplankton (Passow, 2002). Consequently, Bacteroidetes and phytoplankton are often found in close association in polar waters (Grossart et al., 2005; Piquet et al., 2011; Williams et al., 2013) with the relative abundance of the former significantly correlated with the emergence of the late spring blooms (Alonso-Sáez et al., 2008). During May of 2014, the Flavobacteriaceae were notable by their prolific increase in relative abundance from negligible levels in March, congruent with the spike in chloroplast 16S rRNA abundance (Figure 5) and chl a maxima (Table 1). Secondary bacterial production is typically correlated with chl a concentration (Buchan et al., 2014). The Flavobacteriaceae are one of the most commonly found groups in polar ecosystems (Abell and Bowman, 2005), particularly in the summer months (Grzymski et al., 2012; Williams et al., 2012), frequently comprising the majority of Bacteroidetes sequences in these environments (Ghiglione and Murray, 2012; Williams et al., 2013) and amongst the first groups to respond to phytoplankton blooms (Williams et al., 2013). Previously, the Flavobacteriia have been found at peak abundance during the decay phase of a bloom (Riemann et al., 2000; Pinhassi et al., 2004) but this was not the case in our post-bloom August samples (Figure 2). Proteorhodopsins, which support photoheterotrophic growth, have however been found in flavobacterial isolates (Gómez-Consarnau et al., 2007) previously and this may explain the peak abundance observed during the higher photoirradiance conditions of May. It has been suggested that increased numbers of Bacteroidetes would be found in the water column during spring and summer as a consequence of increased melting sea ice, either by seeding, as persistent members of sea ice biota (Bowman et al., 2012; Lo Giudice et al., 2012) or as a result of growth on organic matter released by the thawing (Piquet et al., 2011). As an abundance of Flavobacteriial proteins involved in oxidative stress protection have also been recovered from Antarctic metaproteomes, this suggests that the group may also exhibit a higher tolerance to the high solar irradiance found in polar spring and summer waters (Williams et al., 2013) and may indeed come to play a more dominant role in the surface layers of an ice-free Arctic.
A notable outcome immediately apparent from the 16S rRNA phylogenetic diversity data was the glaring disparity in the relative abundance of Alphaproteobacteria, when compared with previously published studies which saw them dominate polar waters (Alonso-Sáez et al., 2008; Manganelli et al., 2009; Bowman et al., 2012; Williams et al., 2013; Zeng et al., 2013). Closer examination revealed that the ubiquitous SAR11 were notable by their unusually low representation in the dataset, with their relative abundance no higher than 2.5% during the entirety of the study. The SAR11 typically comprise the greater part of observed Alphaproteobacteria in global marine communities (Morris et al., 2002), and more pertinently in the Arctic (Alonso-Sáez et al., 2008; Kirchman et al., 2010; Bowman et al., 2012). In a previous study in Svalbard coastal waters (De Corte et al., 2013), SAR11 were only detected at the 16S rDNA level, none being detected in the 16S rRNA; in the present investigation however, similarly low levels of SAR11 were discerned in both 16S rDNA and rRNA (data not shown). In silico analysis of the universal prokaryotic primer set 519F-806R (Øvreås et al., 1997; Caporaso et al., 2011) used in this study suggested a low-binding efficiency of the reverse primer with the SAR11 cluster, an inherent flaw of the primers confirmed in recent studies (Apprill et al., 2015; Parada et al., 2016). Preliminary shotgun metagenomic data for this same polar time-series did reveal a significantly higher abundance of SAR11 (up to 28% of total prokaryote abundance; data not shown) and so, we can assume that the SAR11 are likely underrepresented in this 16S tag amplicon data set.
Members of the Alphaproteobacterial families Rhodospirillaceae and Rhodobacteraceae were however highly abundant in Arctic waters over the year, with both taxa appearing to be significantly correlated (negatively and positively, respectively) with different phytoplankton bloom stages (Figure 5). The Rhodobacteraceae were the most abundant OTUs recorded in late summer seas off Svalbard (Zeng et al., 2013) and the genus Roseobacter within this family has been seen to peak with chl a maximum in the Western Arctic (Alonso-Sáez et al., 2008), which is assumed to relate to this taxon's common association with phytoplankton blooms (González et al., 2000; Pinhassi et al., 2004; West et al., 2008; Buchan et al., 2014). In the current study however, Roseobacter species were not detected at the Genus level using the Greengenes database (DeSantis et al., 2006) but comparison of unassigned Rhodobacteraceae OTUs with BLAST confirms that they were present, albeit not at the levels seen in similar studies during the polar summer (Grzymski et al., 2012).
The Arctic Ocean has the greatest freshwater input of any ocean (Carmack, 2007) and so as might be expected, the Betaproteobacteria, one of the most prevalent groups in freshwaters, were present in the surface marine samples all year round (Figure 2) albeit at low levels; it is the corresponding absence of rivers in Antarctica which supports their recorded low levels in Southern Oceans (Ghiglione et al., 2012). Interestingly, we observed that the relative abundance of Betaproteobacteria in surface water samples decreased as a function of distance either from the coast or from the pack ice. A number of large glacial fjords flow into the Western coastal waters off Svalbard (Figure 1) and the Betaproteobacteria have been retrieved from Kongsfjorden there (Zeng et al., 2009, 2013; Piquet et al., 2010). The group have also been seen to predominate in Arctic pack ice summer melt pools (Brinkmeyer et al., 2004), which might further seed ocean waters upon thawing. In more temperate latitudes, the abundance of Betaproteobacteria decreases with increasing salinity (Garneau et al., 2009) but in the colder waters of the Arctic, they may persist further offshore (Pedrós-Alió et al., 2015). With climate change bringing with it increased freshwater inputs to the Arctic Ocean, we may also see the relative abundance of the Betaproteobacteria in marine microbial communities rise.
The lower diversity of surface waters during summer appears to be related to both taxonomic and methodological factors; whilst the spike in carbon and nutrient concentrations following a phytoplankton bloom inevitably leads to the proliferation of a few specific bacterial groups (Buchan et al., 2014), the elevated levels of phytoplankton in surface waters can often overwhelm efforts to sample representatively. Indeed, during this study, filters were frequently blocked by phytoplankton cell debris and consequently, without preventative measures, chloroplast 16S rRNAs would routinely comprise >90% of amplicon tag sequences, thereby reducing the sequencing coverage of targetted prokaryotes. Accordingly, prokaryote richness in surface waters during the winter months (and in deeper waters year round) was considerably higher, potentially also due to the more complex composition of the dissolved organic carbon pool (Alonso-Sáez et al., 2008) in darker waters not dominated by just a few bloom-related taxa.
Whilst light availability is thought to be the main driving factor in the epipelagic zone (Giovannoni and Stingl, 2005), conditions within deep dark waters are far from homogenous (Hewson et al., 2006; Teira et al., 2006; Galand et al., 2010). The oceans comprise regional water masses, defined by their distinct temperature and salinity properties, and these circulate around the globe at different spatial scales. Our sampling area off the west coast of Svalbard is notable as being the confluence of North Atlantic and Arctic waters and depending on sample season, location and depth, the deep water column may comprise a number of different water masses (Table 1). A study of the deep Arctic Ocean suggested that these water masses may act as physical barriers to microbial dispersal and that communities within these water masses may therefore have a distinct biogeography (Galand et al., 2010). Preliminary analyses of our data however did not seem to suggest a relationship between water mass and community composition (data not shown), with the effect of light and depth being much more significant (Figure 3).
Microbial life in the mesopelagic zone is quite different from that of the epipelagic zone (Orcutt et al., 2011) and whilst the absence of light is the most obvious difference, the deep oceans are also typically colder and at a higher pressure (increasing by approximately 10 MPa km−1 depth) than surface waters. Within each seasonal sample in our study, there was a clear difference in prokaryote composition between epipelagic and mesopelagic communities (Figure 2) and as might be expected, these differences were much more pronounced during the lighter months of May and August, driven by the massive change in surface communities during the phytoplankton blooms. In the darker months, communities were much more similar with depth and were dominated throughout the water column by the Thaumarchaeota and Chloroflexi-type SAR202. Seasonal changes had little effect on mesopelagic communities, with the moderate variations in prokaryote diversity congruent with the relatively unchanging nature of the deep marine environment. However, we do see a significant difference in phylotype richness in mesopelagic waters between January and the other months and one reason for this might very well be January's placing in the polar year, in the middle of the dark Arctic winter. The phytoplankton bloom results in a single annual pulse of nutrients and in the successional development of mesozooplankton and heterotrophic prokaryotes, the bulk of the simple compounds will be degraded relatively rapidly in the photic surface waters. What is left sinks as marine snow, which continues to be metabolized by the prokaryote community on its way down through the water column. By the time much of this matter reaches the mesopelagic depths, several months after the initial deposition from surface waters, what remains is likely to be diverse relcalcitrant compounds. The primary factor differentiating the deep ocean from surface waters is that metabolic reactions are based only on chemical redox reactions (rather than photosynthetic processes) and much more variably coupled temporally, spatially and functionally (Orcutt et al., 2011). The more diverse chemical processes ongoing in the deep ocean will therefore likely require more functionally (and phylogenetically) diverse organisms than in surface waters, the richness of which seemingly peaks in the winter months.
Unmitigated atmospheric warming will undoubtedly affect gross changes in the marine environment and its phytoplankton populations through increased photoirradiation and nutrients made available by thawing sea ice and permafrost (Li et al., 2009; Doney et al., 2012; El-Swais et al., 2015), respectively. Whilst the phytoplankton blooms are restricted to the well-lit upper layers of the ocean, the effects of their prolific but short-lived activity are seen throughout the deep ocean beneath. As these summer blooms are inextricably linked to the deep and dark activities of the winter blooms of chemolithoautotrophic organisms, a change in one system will inevitably be manifested in the other, potentially via more subtle effects on specific taxonomic or functional prokaryote groups. As major contributors to the marine nitrogen cycle (Nicol and Schleper, 2006) and the dominant chemolithoautotrophs in polar waters, the role of the Thaumarchaeota (previously classified as members of the phylum Crenarchaeota Brochier-Armanet et al., 2008) is well-established (Fuhrman et al., 1992; DeLong et al., 1994; Karner et al., 2001). More recently, the cyclical rise (during winter) and decline (in summer) of the group in the surface waters of polar regions has received some attention (Murray et al., 1998, 1999; Church et al., 2003; Alonso-Sáez et al., 2008, 2012; Grzymski et al., 2012; Williams et al., 2012) and indeed, our data confirmed these findings. The almost total disappearance of the Thaumarchaeota from the photic zone during the summer months (Figure 5) is therefore suggestive of photoinhibition of ammonia oxidation (Guerrero and Jones, 1996; Murray et al., 1998; Mincer et al., 2007; Schleper and Nicol, 2010; Merbt et al., 2012), or has been more recently posited, a sensitivity to reactive oxygen species produced as a result of photosynthesis (Tolar et al., 2016). The reasons may however be multifactorial and it is thought that the Archaea may also be outcompeted by phytoplankton (Murray et al., 1998; Ward, 2000, 2005; Church et al., 2003; Herfort et al., 2007; Smith et al., 2014) and Bacteria (which are much more active in the uptake of the labile bloom-produced substrates Alonso-Sáez et al., 2008; Kalanetra et al., 2009), or even subjected to selective viral infection (Labonté et al., 2015). The proportional abundance of Thaumarchaeota has been correlated with ammonium concentrations (Herfort et al., 2007; Kirchman et al., 2007) and their peak abundance in winter surface waters has been hypothesized to result from mixing with deep water masses in Antarctic seas (Kalanetra et al., 2009; Grzymski et al., 2012); however, in areas of the Arctic Ocean where the water column tends to remain stratified during the winter (Forest et al., 2011), recent data suggests that the increase is in fact due to growth and proliferation of surface water Thaumarchaeota populations in situ (Alonso-Sáez et al., 2012). As the polar winter precludes photosynthesis and consequently, a source of labile organic matter, the autotrophic Thaumarchaeota are ideally adapted to bloom in these otherwise limiting conditions (Pedrós-Alió et al., 2015). Interestingly, in both Arctic (Alonso-Sáez et al., 2012) and Antarctic (Kalanetra et al., 2009; Grzymski et al., 2012) winter surface waters, a single Thaumarchaeotal OTU was seen to dominate archaeal 16S rRNA and ammonia monooxygenase amoA gene libraries and we saw a similar dominance in Arctic winter surface waters; however, as we continued down through the water profile, this dominant OTU gradually yields to another Thaumarchaeotal OTU, which ultimately dominates deeper waters. These shallow and deep cladal differences have been described in ammonia-oxidizing Archaea (AOA) previously, based primarily in differences in their amoA genes (Francis et al., 2005; Sintes et al., 2013; Pedneault et al., 2014; among others) but also other metabolic genes (Sintes et al., 2013; Luo et al., 2014; Villanueva et al., 2015).
Any change in Thaumarchaeota populations may have significant impacts on certain of the phytoplankton populations. In unlit surface waters, the Thaumarchaeota oxidize ammonium to nitrate, which in turn promotes phytoplankton growth come the lighter months; however, as the larger diatoms are thought to outcompete picophytoplankton for nitrate (Stolte and Riegman, 1995), diatom populations may be proportionally more affected by a loss of Thaumarchaeota (Comeau et al., 2011). Conversely, a warming freshening Arctic will be increasingly stratified and there will be less mixing with the nutrient-rich depths, potentially favoring smaller picophytoplankton (Li et al., 2009).
Taxa showing similar seasonal dynamics to the chemolithotrophic Thaumarchaeota in surface waters were the Deltaproteobacteria-affiliated SAR324, Nitrospinaceae and OM27 (Figure 5), all of which saw drops in relative abundance during the summer months. The nitrite-oxidizing genus Nitrospina has been observed in winter-only samples from both Antarctic (Grzymski et al., 2012) and Arctic (Alonso-Sáez et al., 2010) waters, as well as in temperate waters (El-Swais et al., 2015) and has also been correlated with amoA-containing Thaumarchaeota in Monterey Bay (Mincer et al., 2007). Whilst we recorded generally higher levels of all three taxa in winter surface and year-round deeper waters, active populations were still detected in surface waters in summer months, albeit at much lower levels. The Chloroflexi-type SAR202 cluster is also typical and highly abundant in mesopelagic waters (Morris et al., 2004; Varela et al., 2008; Arístegui et al., 2009; Dobal-Amador et al., 2016). Recent studies have shown the Chloroflexi to be well adapted to deeper oligotrophic waters and to efficiently utilize recalcitrant organic compounds uptake, such as those found in the mesopelagic zone (Yilmaz et al., 2016).
5. Conclusions
This high-throughput sequencing study of a polar year in the Arctic Ocean revealed the driving force of light and phytoplankton blooms on the marine prokaryote community, and in particular the Thaumarchaeota, in both surface and deep ocean waters. Investigations such as this one and others discussed in the text are fundamentally important as a historic record of the current microbial state of the polar oceans and an indicator of the ongoing rate of change (Ducklow et al., 2009). Additionally, we believe that it is crucial to consider the deeper water column in these studies, as part of a whole, dynamic and interconnected marine system. As a complement to this study, we are also investigating shotgun metagenomic and metatranscriptomic data for these same seasonal samples, which will greatly improve the resolution of these preliminary results. We suggest that the future progress of the field would benefit greatly from repeated longer-term investigations, in concert with the continued improvement of omics tools and in this manner, we hope that these data may be used to support the overwhelming physical evidence of our changing global climate.
Author Contributions
BW analyzed data, prepared figures and tables, and wrote the paper. OM, EN, LS, GB, and LØ collected, optimized, and processed samples. OM, LS, GB, and LØ were involved in project description, design of experiment, discussion, and interpreting data.
Funding
The MicroPolar project (also responsible for the cruises during March and November) was funded by the Research Council of Norway (RCN 225956). We would also like to thank the Carbon Bridge project (RCN 226415) for allowing us to participate during their cruises in January, May, and August. LØ is also affiliated with the University Centre in Svalbard (UNIS) and was funded, along with OM, by “Microorganisms in the Arctic: Major drivers of biogeochemical cycles and climate change” (RCN 227062). LS was funded through the Carbon Bridge project (RCN 226415).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank crew members of the Norwegian Research Vessels RV Helmer Hanssen and RV Lance for their assistance in sampling expeditions and all colleagues in the UiB Marine Microbiology group and collaborators abroad who contributed to the research effort.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmars.2017.00095/full#supplementary-material
References
Abell, G. C. J., and Bowman, J. P. (2005). Ecological and biogeographic relationships of class Flavobacteria in the Southern Ocean. FEMS Microbiol. Ecol. 51, 265–277. doi: 10.1016/j.femsec.2004.09.001
Alonso-Sáez, L., Galand, P. E., Casamayor, E. O., Pedrós-Alió, C., and Bertilsson, S. (2010). High bicarbonate assimilation in the dark by Arctic bacteria. ISME J. 4, 1581–1590. doi: 10.1038/ismej.2010.69
Alonso-Sáez, L., Sánchez, O., Gasol, J. M., Balagué, V., and Pedrós-Alió, C. (2008). Winter-to-summer changes in the composition and single-cell activity of near-surface Arctic prokaryotes. Environ. Microbiol. 10, 2444–2454. doi: 10.1111/j.1462-2920.2008.01674.x
Alonso-Sáez, L., Waller, A. S., Mende, D. R., Bakker, K., Farnelid, H., Yager, P. L., et al. (2012). Role for urea in nitrification by polar marine Archaea. Proc. Natl. Acad. Sci. U.S.A. 109, 17989–17994. doi: 10.1073/pnas.1201914109
Anderson, L. G., and Macdonald, R. W. (2015). Observing the Arctic Ocean carbon cycle in a changing environment. Polar Res. 34:26891. doi: 10.3402/polar.v34.26891
Anderson, L. G. (2002). “DOC in the Arctic Ocean,” in Biogeochemistry of Marine Dissolved Organic Matter, eds D. A. Hansell and C. A. Carlson (New York, NY: Elsevier), 665–683.
Apprill, A., McNally, S. P., Parsons, R., and Weber, L. (2015). Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 75, 129–137. doi: 10.3354/ame01753
Arístegui, J., Gasol, J. M., Duarte, C. M., and Herndl, G. J. (2009). Microbial oceanography of the dark ocean's pelagic realm. Limnol. Oceanogr. 54, 1501–1529. doi: 10.4319/lo.2009.54.5.1501
Boe, J., Hall, A., and Qu, X. (2009). September sea-ice cover in the Arctic Ocean projected to vanish by 2100. Nat. Geosci. 2, 341–343. doi: 10.1038/ngeo467
Bowman, J. S., Rasmussen, S., Blom, N., Deming, J. W., Rysgaard, S., and Sicheritz-Ponten, T. (2012). Microbial community structure of Arctic multiyear sea ice and surface seawater by 454 sequencing of the 16S RNA gene. ISME J. 6, 11–20. doi: 10.1038/ismej.2011.76
Brinkmeyer, R., Glöckner, F. O., Helmke, E., and Amann, R. (2004). Predominance of β-proteobacteria in summer melt pools on Arctic pack ice. Limnol. Oceanogr. 49, 1013–1021. doi: 10.4319/lo.2004.49.4.1013
Brochier-Armanet, C., Boussau, B., Gribaldo, S., and Forterre, P. (2008). Mesophilic crenarchaeota: proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 6, 245–252. doi: 10.1038/nrmicro1852
Buchan, A., LeCleir, G. R., Gulvik, C. A., and González, J. M. (2014). Master recyclers: features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Micro. 12, 686–698. doi: 10.1038/nrmicro3326
Caporaso, J. G., Bittinger, K., Bushman, F. D., DeSantis, T. Z., Andersen, G. L., and Knight, R. (2010a). PyNAST: a flexible tool for aligning sequences to a template alignment. Bioinformatics 26, 266–267. doi: 10.1093/bioinformatics/btp636
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010b). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Caporaso, J. G., Lauber, C., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U.S.A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Carmack, E. (2007). The alpha/beta ocean distinction: a perspective on freshwater fluxes, convection, nutrients and productivity in high-latitude seas. Deep-Sea Res. II 54, 2578–2598. doi: 10.1016/j.dsr2.2007.08.018
Chen, J. L., Wilson, C. R., Blankenship, D., and Tapley, B. D. (2009). Accelerated Antarctic ice loss from satellite gravity measurements. Nat. Geosci. 2, 859–862. doi: 10.1038/ngeo694
Christman, G. D., Cottrell, M. T., Popp, B. N., Gier, E., and Kirchman, D. L. (2011). Abundance, diversity, and activity of ammonia-oxidizing prokaryotes in the coastal Arctic Ocean in summer and winter. Appl. Environ. Microb. 77, 2026–2034. doi: 10.1128/AEM.01907-10
Church, M. J., Delong, E. F., Ducklow, H. W., Karner, M. B., Preston, C. M., and Karl, D. M. (2003). Abundance and distribution of planktonic Archaea and Bacteria in the waters west of the Antarctic Peninsula. Limnol. Oceanogr. 48, 1893–1902. doi: 10.4319/lo.2003.48.5.1893
Cock, P. J. A., Antao, T., Chang, J. T., Chapman, B. A., Cox, C. J., Dalke, A., et al. (2009). Biopython: freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 25, 1422–1423. doi: 10.1093/bioinformatics/btp163
Comeau, A. M., Li, W. K. W., Tremblay, J.-E., Carmack, E. C., and Lovejoy, C. (2011). Arctic Ocean microbial community structure before and after the 2007 record sea ice minimum. PLoS ONE 6:e27492. doi: 10.1371/journal.pone.0027492
Comiso, J. C. (2012). Large decadal decline of the Arctic multiyear ice cover. J. Clim. 25, 1176–1193. doi: 10.1175/JCLI-D-11-00113.1
Comiso, J. C., Parkinson, C. L., Gersten, R., and Stock, L. (2008). Accelerated decline in the Arctic sea ice cover. Geophys. Res. Lett. 35:L01703. doi: 10.1029/2007GL031972
Connelly, T. L., Baer, S. E., Cooper, J. T., Bronk, D. A., and Wawrik, B. (2014). Urea uptake and carbon fixation by marine pelagic bacteria and archaea during the Arctic summer and winter seasons. Appl. Environ. Microb. 80, 6013–6022. doi: 10.1128/AEM.01431-14
De Corte, D., Sintes, E., Yokokawa, T., and Herndl, G. J. (2013). Comparison between MICRO-CARD-FISH and 16S rRNA gene clone libraries to assess the active versus total bacterial community in the coastal Arctic. Environ. Microbiol. Rep. 5, 272–281. doi: 10.1111/1758-2229.12013
de Steur, L., Hansen, E., Mauritzen, C., Beszczynska-Möller, A., and Fahrbach, E. (2014). Impact of recirculation on the East Greenland current in fram strait: results from moored current meter measurements between 1997 and 2009. Deep-Sea Res. I 92, 26–40. doi: 10.1016/j.dsr.2014.05.018
DeLong, E. F., Wu, K. Y., Prézelin, B. B., and Jovine, R. V. M. (1994). High abundance of archaea in antarctic marine picoplankton. Nature 371, 695–697. doi: 10.1038/371695a0
DeSantis, T. Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E. L., Keller, K., et al. (2006). Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microb. 72, 5069–5072. doi: 10.1128/AEM.03006-05
Dobal-Amador, V., Nieto-Cid, M., Guerrero-Feijoo, E., Hernando-Morales, V., Teira, E., and Varela, M. M. (2016). Vertical stratification of bacterial communities driven by multiple environmental factors in the waters (0-5000 m) off the Galician coast (NW Iberian margin). Deep-Sea Res. I 114, 1–11. doi: 10.1016/j.dsr.2016.04.009
Doney, S. C., Ruckelshaus, M., Duffy, J. E., Barry, J. P., Chan, F., English, C. A., et al. (2012). Climate change impacts on marine ecosystems. Ann. Rev. Mar. Sci. 4, 11–37. doi: 10.1146/annurev-marine-041911-111611
Ducklow, H. W., Doney, S. C., and Steinberg, D. K. (2009). Contributions of long-term research and time-series observations to marine ecology and biogeochemistry. Ann. Rev. Mar. Sci. 1, 279–302. doi: 10.1146/annurev.marine.010908.163801
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
El-Swais, H., Dunn, K. A., Bielawski, J. P., Li, W. K. W., and Walsh, D. A. (2015). Seasonal assemblages and short-lived blooms in coastal north-west Atlantic Ocean bacterioplankton. Environ. Microbiol. 17, 3642–3661. doi: 10.1111/1462-2920.12629
Fernández-Gómez, B., Richter, M., Schüler, M., Pinhassi, J., Acinas, S. G., González, J. M., et al. (2013). Ecology of marine bacteroidetes: a comparative genomics approach. ISME J. 7, 1026–1037. doi: 10.1038/ismej.2012.169
Forest, A., Tremblay, J.-E., Gratton, Y., Martin, J., Gagnon, J., Darnis, G., et al. (2011). Biogenic carbon flows through the planktonic food web of the Amundsen Gulf (Arctic Ocean): a synthesis of field measurements and inverse modeling analyses. Prog. Oceanogr. 91, 410–436. doi: 10.1016/j.pocean.2011.05.002
Francis, C. A., Roberts, K. J., Beman, J. M., Santoro, A. E., and Oakley, B. B. (2005). Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. U.S.A. 102, 14683–14688. doi: 10.1073/pnas.0506625102
Frey, K. E. and McClelland, J. W. (2009). Impacts of permafrost degradation on arctic river biogeochemistry. Hydrol. Process 23, 169–182. doi: 10.1002/hyp.7196
Fuhrman, J. A., Cram, J. A., and Needham, D. M. (2015). Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Micro. 13, 133–146. doi: 10.1038/nrmicro3417
Fuhrman, J. A., Hewson, I., Schwalbach, M. S., Steele, J. A., Brown, M. V., and Naeem, S. (2006). Annually reoccurring bacterial communities are predictable from ocean conditions. Proc. Natl. Acad. Sci. U.S.A. 103, 13104–13109. doi: 10.1073/pnas.0602399103
Fuhrman, J. A., McCallum, K., and Davis, A. A. (1992). Novel major archaeabacterial group from marine plankton. Nature 356, 148–149. doi: 10.1038/356148a0
Galand, P. E., Potvin, M., Casamayor, E. O., and Lovejoy, C. (2010). Hydrography shapes bacterial biogeography of the deep Arctic Ocean. ISME J. 4, 564–576. doi: 10.1038/ismej.2009.134
Garneau, M.-E., Vincent, W. F., Terrado, R., and Lovejoy, C. (2009). Importance of particle-associated bacterial heterotrophy in a coastal Arctic ecosystem. J. Mar. Syst. 75, 185–197. doi: 10.1016/j.jmarsys.2008.09.002
Ghiglione, J.-F., Galand, P. E., Pommier, T., Pedrós-Alió, C., Maas, E. W., Bakker, K., et al. (2012). Pole-to-pole biogeography of surface and deep marine bacterial communities. Proc. Natl. Acad. Sci. U.S.A. 109, 17633–17638. doi: 10.1073/pnas.1208160109
Ghiglione, J. F. and Murray, A. E. (2012). Pronounced summer to winter differences and higher wintertime richness in coastal Antarctic marine bacterioplankton. Environ. Microbiol. 14, 617–629. doi: 10.1111/j.1462-2920.2011.02601.x
Gilbert, J. A., Steele, J. A., Caporaso, J. G., Steinbruck, L., Reeder, J., Temperton, B., et al. (2012). Defining seasonal marine microbial community dynamics. ISME J. 6, 298–308. doi: 10.1038/ismej.2011.107
Giovannoni, S. J., and Stingl, U. (2005). Molecular diversity and ecology of microbial plankton. Nature 437, 343–348. doi: 10.1038/nature04158
Gómez-Consarnau, L., González, J. M., Coll-Lladó, M., Gourdon, P., Pascher, T., Neutze, R., et al. (2007). Light stimulates growth of proteorhodopsin-containing marine Flavobacteria. Nature 445, 210–213. doi: 10.1038/nature05381
González, J. M., Simó, R., Massana, R., Covert, J. S., Casamayor, E. O., Pedrós-Alió, C., et al. (2000). Bacterial community structure associated with a dimethylsulfoniopropionate-producing North Atlantic algal bloom. Appl. Environ. Microb. 66, 4237–4246. doi: 10.1128/AEM.66.10.4237-4246.2000
Grossart, H.-P., Levold, F., Allgaier, M., Simon, M., and Brinkhoff, T. (2005). Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 7, 860–873. doi: 10.1111/j.1462-2920.2005.00759.x
Grzymski, J. J., Riesenfeld, C. S., Williams, T. J., Dussaq, A. M., Ducklow, H., Erickson, M., et al. (2012). A metagenomic assessment of winter and summer bacterioplankton from Antarctica Peninsula coastal surface waters. ISME J. 6, 1901–1915. doi: 10.1038/ismej.2012.31
Guerrero, M. A. and Jones, R. D. (1996). Photoinhibition of marine nitrifying bacteria. 1. Wavelength-dependent response. Mar. Ecol. Prog. Ser. 141, 183–192. doi: 10.3354/meps141183
Haas, B. J., Gevers, D., Earl, A. M., Feldgarden, M., Ward, D. V., Giannoukos, G., et al. (2011). Chimeric 16s rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 21, 494–504. doi: 10.1101/gr.112730.110
Herfort, L., Schouten, S., Abbas, B., Veldhuis, M. J. W., Coolen, M. J. L., Wuchter, C., et al. (2007). Variations in spatial and temporal distribution of Archaea in the North Sea in relation to environmental variables. FEMS Microbiol. Ecol. 62, 242–257. doi: 10.1111/j.1574-6941.2007.00397.x
Hewson, I., Steele, J. A., Capone, D. G., and Fuhrman, J. A. (2006). Remarkable heterogeneity in meso- and bathypelagic bacterioplankton assemblage composition. Limnol. Oceanogr. 51, 1274–1283. doi: 10.4319/lo.2006.51.3.1274
Hop, H., Falk-Petersen, S., Svendsen, H., Kwasniewski, S., Pavlov, V., Pavlova, O., et al. (2006). Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Prog. Oceanogr. 71, 182–231. doi: 10.1016/j.pocean.2006.09.007
Hop, H., Pearson, T., Hegseth, E. N., Kovacs, K., Wiencke, C., Kwasniewski, S., et al. (2002). The marine ecosystem of Kongsfjorden, Svalbard. Polar Res. 21, 167–208. doi: 10.1111/j.1751-8369.2002.tb00073.x
Joshi, N. A., and Fass, J. N. (2011). Sickle: A Sliding-Window, Adaptive, Quality-Based Trimming Tool for FastQ Files. Available online at: https://github.com/najoshi/sickle
Kalanetra, K. M., Bano, N., and Hollibaugh, J. T. (2009). Ammonia-oxidizing Archaea in the Arctic Ocean and Antarctic coastal waters. Environ. Microbiol. 11, 2434–2445. doi: 10.1111/j.1462-2920.2009.01974.x
Karner, M. B., DeLong, E. F., and Karl, D. M. (2001). Archaeal dominance in the mesopelagic zone of the Pacific Ocean. Nature 409, 507–510. doi: 10.1038/35054051
Kerr, R. A. (2012). Ice-Free Arctic Sea may be years, not decades, a way. Science 337:1591. doi: 10.1126/science.337.6102.1591
Kirchman, D. L., Cottrell, M. T., and Lovejoy, C. (2010). The structure of bacterial communities in the western Arctic Ocean as revealed by pyrosequencing of 16S rRNA genes. Environ. Microbiol. 12, 1132–1143. doi: 10.1111/j.1462-2920.2010.02154.x
Kirchman, D. L., Elifantz, H., Dittel, A. I., Malmstrom, R. R., and Cottrell, M. T. (2007). Standing stocks and activity of Archaea and bacteria in the western Arctic Ocean. Limnol. Oceanogr. 52, 495–507. doi: 10.4319/lo.2007.52.2.0495
Labonté, J. M., Swan, B. K., Poulos, B., Luo, H., Koren, S., Hallam, S. J., et al. (2015). Single-cell genomics-based analysis of virus-host interactions in marine surface bacterioplankton. ISME J. 9, 2386–2399. doi: 10.1038/ismej.2015.48
Ladau, J., Sharpton, T. J., Finucane, M. M., Jospin, G., Kembel, S. W., O'Dwyer, J., et al. (2013). Global marine bacterial diversity peaks at high latitudes in winter. ISME J. 7, 1669–1677. doi: 10.1038/ismej.2013.37
Langmead, B. and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lantuit, H., Overduin, P. P., Couture, N., Wetterich, S., Aré, F., Atkinson, D., et al. (2012). The Arctic coastal dynamics database: a new classification scheme and statistics on Arctic permafrost coastlines. Estuar. Coast 35, 383–400. doi: 10.1007/s12237-010-9362-6
Li, W. K. W., McLaughlin, F. A., Lovejoy, C., and Carmack, E. C. (2009). Smallest Algae Thrive As the Arctic Ocean Freshens. Science 326:539. doi: 10.1126/science.1179798
Lo Giudice, A., Caruso, C., Mangano, S., Bruni, V., De Domenico, M., and Michaud, L. (2012). Marine bacterioplankton diversity and community composition in an Antarctic Coastal environment. Microb. Ecol. 63, 210–223. doi: 10.1007/s00248-011-9904-x
Luo, H., Tolar, B. B., Swan, B. K., Zhang, C. L., Stepanauskas, R., Ann Moran, M., et al. (2014). Single-cell genomics shedding light on marine Thaumarchaeota diversification. ISME J. 8, 732–736. doi: 10.1038/ismej.2013.202
Manganelli, M., Malfatti, F., Samo, T. J., Mitchell, B. G., Wang, H., and Azam, F. (2009). Major role of microbes in carbon fluxes during austral winter in the southern drake passage. PLoS ONE 4:e6941. doi: 10.1371/journal.pone.0006941
Markus, T., Stroeve, J. C., and Miller, J. (2009). Recent changes in Arctic sea ice melt onset, freezeup, and melt season length. J. Geophys. Res. Oceans 114:C12024. doi: 10.1029/2009JC005436
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17, 10–12. doi: 10.14806/ej.17.1.200
Masella, A. P., Bartram, A. K., Truszkowski, J. M., Brown, D. G., and Neufeld, J. D. (2012). PANDAseq: PAired-eND assembler for illumina sequences. BMC Bioinformatics 13:31. doi: 10.1186/1471-2105-13-31
Merbt, S. N., Stahl, D. A., Casamayor, E. O., Martí, E., Nicol, G. W., and Prosser, J. I. (2012). Differential photoinhibition of bacterial and archaeal ammonia oxidation. FEMS Microbiol. Lett. 327, 41–46. doi: 10.1111/j.1574-6968.2011.02457.x
Mincer, T. J., Church, M. J., Taylor, L. T., Preston, C., Karl, D. M., and DeLong, E. F. (2007). Quantitative distribution of presumptive archaeal and bacterial nitrifiers in monterey bay and the North Pacific Subtropical Gyre. Environ. Microbiol. 9, 1162–1175. doi: 10.1111/j.1462-2920.2007.01239.x
Morris, R. M., Rappé, M. S., Connon, S. A., Vergin, K. L., Siebold, W. A., Carlson, C. A., et al. (2002). SAR11 clade dominates ocean surface bacterioplankton communities. Nature 420, 806–810. doi: 10.1038/nature01240
Morris, R. M., Rappé, M. S., Urbach, E., Connon, S. A., and Giovannoni, S. J. (2004). Prevalence of the Chloroflexi-related SAR202 bacterioplankton cluster throughout the mesopelagic zone and deep ocean. Appl. Environ. Micro. 70, 2836–2842. doi: 10.1128/AEM.70.5.2836-2842.2004
Murray, A. E., and Grzymski, J. J. (2007). Diversity and genomics of Antarctic marine micro-organisms. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 362, 2259–2271. doi: 10.1098/rstb.2006.1944
Murray, A. E., Preston, C. M., Massana, R., Taylor, L. T., Blakis, A., Wu, K., et al. (1998). Seasonal and spatial variability of bacterial and archaeal assemblages in the coastal waters near anvers island, antarctica. Appl. Environ. Microb. 64, 2585–2595.
Murray, A. E., Wu, K., Moyer, C., Karl, D. M., and DeLong, E. F. (1999). Evidence for circumpolar distribution of planktonic Archaea in the Southern Ocean. Aquat. Microb. Ecol. 18, 263–273. doi: 10.3354/ame018263
Nicol, G. W. and Schleper, C. (2006). Ammonia-oxidising Crenarchaeota: important players in the nitrogen cycle? Trends Microbiol. 14, 207–212. doi: 10.1016/j.tim.2006.03.004
Onarheim, I. H., Smedsrud, L. H., Ingvaldsen, R. B., and Nilsen, F. (2014). Loss of sea ice during winter north of Svalbard. Tellus A 66:23933. doi: 10.3402/tellusa.v66.23933
Orcutt, B. N., Sylvan, J. B., Knab, N. J., and Edwards, K. J. (2011). Microbial ecology of the dark ocean above, at, and below the seafloor. Microbiol. Mol. Biol. Rev. 75, 361–422. doi: 10.1128/MMBR.00039-10
Øvreås, L., Forney, L., Daae, F. L., and Torsvik, V. (1997). Distribution of bacterioplankton in meromictic lake saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microb. 63, 3367–3373.
Parada, A. E., Needham, D. M., and Fuhrman, J. A. (2016). Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 18, 1403–1414. doi: 10.1111/1462-2920.13023
Parson, T. R., Maita, Y., and Lalli, C. M. (1984). “4.3 - fluorometric determination of chlorophylls,” in A Manual of Chemical and Biological Methods for Seawater Analysis, eds T. R. Parson, Y. Maita, and C. M. Lalli (Amsterdam: Pergamon), 107–109.
Passow, U. (2002). Transparent exopolymer particles (TEP) in aquatic environments. Progr. Oceanogr. 55, 287–333. doi: 10.1016/S0079-6611(02)00138-6
Paulsen, M., Doré, H., Garczarek, L., Seuthe, L., Müller, O., Sandaa, R.-A., et al. (2016). Synechococcus in the Atlantic gateway to the Arctic Ocean. Front. Mar. Sci. 3:191. doi: 10.3389/fmars.2016.00191
Pedneault, E., Galand, P., Potvin, M., Tremblay, J.-E., and Lovejoy, C. (2014). Archaeal amoA and ureC genes and their transcriptional activity in the Arctic Ocean. Sci. Rep. 4:4661. doi: 10.1038/srep04661
Pedrós-Alió, C., Potvin, M., and Lovejoy, C. (2015). Diversity of planktonic microorganisms in the Arctic Ocean. Progr. Oceanogr. 139, 233–243. doi: 10.1016/j.pocean.2015.07.009
Perovich, D. K., Light, B., Eicken, H., Jones, K. F., Runciman, K., and Nghiem, S. V. (2007). Increasing solar heating of the Arctic Ocean and adjacent seas, 1979-2005: attribution and role in the ice-albedo feedback. Geophys. Res. Lett. 34:L19505. doi: 10.1029/2007GL031480
Pinhassi, J., Sala, M. M., Havskum, H., Peters, F., Guadayol, O., Malits, A., and Marrasé, C. (2004). Changes in bacterioplankton composition under different phytoplankton regimens. Appl. Environ. Microb. 70, 6753–6766. doi: 10.1128/AEM.70.11.6753-6766.2004
Piquet, A. M.-T., Bolhuis, H., Meredith, M. P., and Buma, A. G. (2011). Shifts in coastal Antarctic marine microbial communities during and after melt water-related surface stratification. FEMS Microbiol. Ecol. 76, 413–427. doi: 10.1111/j.1574-6941.2011.01062.x
Piquet, A. M.-T., Scheepens, J. F., Bolhuis, H., Wiencke, C., and Buma, A. G. J. (2010). Variability of protistan and bacterial communities in two Arctic fjords (Spitsbergen). Polar Biol. 33, 1521–1536. doi: 10.1007/s00300-010-0841-9
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Randelhoff, A., Sundfjord, A., and Reigstad, M. (2015). Seasonal variability and fluxes of nitrate in the surface waters over the Arctic shelf slope. Geophys. Res. Lett. 42, 3442–3449. doi: 10.1002/2015GL063655
Reigstad, M., Riser, C. W., Wassmann, P., and Ratkova, T. (2008). Vertical export of particulate organic carbon: attenuation, composition and loss rates in the northern Barents sea. Deep-Sea Res. II 55, 2308–2319. doi: 10.1016/j.dsr2.2008.05.007
Riemann, L., Steward, G. F., and Azam, F. (2000). Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl. Environ. Microb. 66, 578–587. doi: 10.1128/AEM.66.2.578-587.2000
Romanovsky, V. E., Smith, S. L., and Christiansen, H. H. (2010). Permafrost thermal state in the polar Northern Hemisphere during the international polar year 2007-2009: a synthesis. Permafrost Periglac 21, 106–116. doi: 10.1002/ppp.689
Sakshaug, E. (2004). “Primary and secondary production in the Arctic seas,” in The Organic Carbon Cycle in the Arctic Ocean, eds R. Stein and R. W. MacDonald (Berlin; Heidelberg: Springer), 57–81.
Schauer, U., Fahrbach, E., Osterhus, S., and Rohardt, G. (2004). Arctic warming through the fram strait: oceanic heat transport from 3 years of measurements. J. Geophys. Res. Oceans 109:C06026. doi: 10.1029/2003JC001823
Schleper, C., and Nicol, G. W. (2010). “Ammonia-oxidising archaea physiology, ecology and evolution,” in Advances in Microbial Physiology, Vol. 57, ed R. K. Poole (Cambridge: Academic Press), 1–41.
Sherr, E. B., Sherr, B. F., Wheeler, P. A., and Thompson, K. (2003). Temporal and spatial variation in stocks of autotrophic and heterotrophic microbes in the upper water column of the central Arctic Ocean. Deep-Sea Res. I 50, 557–571. doi: 10.1016/S0967-0637(03)00031-1
Sintes, E., Bergauer, K., De Corte, D., Yokokawa, T., and Herndl, G. J. (2013). Archaeal amoA gene diversity points to distinct biogeography of ammonia-oxidizing Crenarchaeota in the ocean. Environ. Microbiol. 15, 1647–1658. doi: 10.1111/j.1462-2920.2012.02801.x
Smith, J. M., Chavez, F. P., and Francis, C. A. (2014). Ammonium uptake by phytoplankton regulates nitrification in the sunlit ocean. PLoS ONE 9:e108173. doi: 10.1371/journal.pone.0108173
Stolte, W. and Riegman, R. (1995). Effect of phytoplankton cell size on transient-state nitrate and ammonium uptake kinetics. Microbiology 141, 1221–1229. doi: 10.1099/13500872-141-5-1221
Svendsen, H., Beszczynska-Møller, A., Hagen, J., Lefauconnier, B., Tverberg, V., Gerland, S., et al. (2002). The physical environment of kongsfjorden-krossfjorden, an Arctic fjord system in Svalbard. Polar Res. 21, 133–166. doi: 10.1111/j.1751-8369.2002.tb00072.x
Teeling, H., Fuchs, B. M., Becher, D., Klockow, C., Gardebrecht, A., Bennke, C. M., et al. (2012). Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science 336, 608–611. doi: 10.1126/science.1218344
Teira, E., Lebaron, P., van Aken, H., and Herndl, G. J. (2006). Distribution and activity of Bacteria and Archaea in the deep water masses of the North Atlantic. Limnol. Oceanogr. 51, 2131–2144. doi: 10.4319/lo.2006.51.5.2131
Tolar, B. B., Powers, L. C., Miller, W. L., Wallsgrove, N. J., Popp, B. N., and Hollibaugh, J. T. (2016). Ammonia oxidation in the ocean can be inhibited by nanomolar concentrations of hydrogen peroxide. Front. Mar. Sci. 3:237. doi: 10.3389/fmars.2016.00237
Tremblay, J.-É., Simpson, K., Martin, J., Miller, L., Gratton, Y., Barber, D., et al. (2008). Vertical stability and the annual dynamics of nutrients and chlorophyll fluorescence in the coastal, southeast Beaufort sea. J Geophys. Res. Oceans 113:C07S90. doi: 10.1029/2007JC004547
Trenberth, K. E., Jones, P. D., Amenhem, P., Bojariu, R., Easterling, D., Tank, A. K., et al. (2007). “Observations: atmospheric surface and climate change,” in Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, eds S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Avery, M. Tignor, and H. L. Miller (Cambridge, UK: Cambridge University Press), 235–336.
Varela, M. M., Van Aken, H. M., and Herndl, G. J. (2008). Abundance and activity of Chloroflexi-type SAR202 bacterioplankton in the meso- and bathypelagic waters of the (sub)tropical Atlantic. Environ. Microbiol. 10, 1903–1911. doi: 10.1111/j.1462-2920.2008.01627.x
Villanueva, L., Schouten, S., and Sinninghe Damsté, J. S. (2015). Depth-related distribution of a key gene of the tetraether lipid biosynthetic pathway in marine Thaumarchaeota. Environ. Microbiol. 17, 3527–3539. doi: 10.1111/1462-2920.12508
Walczowski, W. (2013). Frontal structures in the West Spitsbergen current margins. Ocean Sci. 9, 957–975. doi: 10.5194/os-9-957-2013
Ward, B. B. (2000). “Nitrification and the marine nitrogen cycle,” in Microbial Ecology of the Oceans, ed D. L. Kirchman (New York, NY: Wiley-Liss), 427–453.
Ward, B. B. (2005). Temporal variability in nitrification rates and related biogeochemical factors in monterey bay, california, U.S.A. Mar. Ecol. Prog. Ser. 292, 97–109. doi: 10.3354/meps292097
West, N. J., Obernosterer, I., Zemb, O., and Lebaron, P. (2008). Major differences of bacterial diversity and activity inside and outside of a natural iron-fertilized phytoplankton bloom in the Southern Ocean. Environ. Microbiol. 10, 738–756. doi: 10.1111/j.1462-2920.2007.01497.x
Whitman, W. B., Coleman, D. C., and Wiebe, W. J. (1998). Prokaryotes: the unseen majority. Proc. Natl. Acad. Sci. U.S.A. 95, 6578–6583. doi: 10.1073/pnas.95.12.6578
Williams, T. J., Long, E., Evans, F., DeMaere, M. Z., Lauro, F. M., Raftery, M. J., et al. (2012). A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters. ISME J. 6, 1883–1900. doi: 10.1038/ismej.2012.28
Williams, T. J., Wilkins, D., Long, E., Evans, F., DeMaere, M. Z., Raftery, M. J., et al. (2013). The role of planktonic Flavobacteria in processing algal organic matter in coastal East Antarctica revealed using metagenomics and metaproteomics. Environ. Microbiol. 15, 1302–1317. doi: 10.1111/1462-2920.12017
Yilmaz, P., Yarza, P., Rapp, J. Z., and Glöckner, F. O. (2016). Expanding the world of marine bacterial and archaeal clades. Front. Microbiol. 6:1524. doi: 10.3389/fmicb.2015.01524
Zeng, Y., Zheng, T., and Li, H. (2009). Community composition of the marine bacterioplankton in Kongsfjorden (Spitsbergen) as revealed by 16S rRNA gene analysis. Polar Biol. 32, 1447–1460. doi: 10.1007/s00300-009-0641-2
Keywords: Arctic, marine microbiology, seasonality, nutrient cycling, depth, climate change, phytoplankton bloom, mesopelagic
Citation: Wilson B, Müller O, Nordmann E-L, Seuthe L, Bratbak G and Øvreås L (2017) Changes in Marine Prokaryote Composition with Season and Depth Over an Arctic Polar Year. Front. Mar. Sci. 4:95. doi: 10.3389/fmars.2017.00095
Received: 01 October 2016; Accepted: 20 March 2017;
Published: 13 April 2017.
Edited by:
Connie Lovejoy, Laval University, CanadaReviewed by:
Nico Salmaso, Fondazione Edmund Mach, ItalyDolors Vaque, Consejo Superior de Investigaciones Científicas, Spain
Copyright © 2017 Wilson, Müller, Nordmann, Seuthe, Bratbak and Øvreås. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bryan Wilson, YnJ5YW4ud2lsc29uQHVpYi5ubw==