 P.S. Soumia
P.S. Soumia Dhananjay V. Shirsat
Dhananjay V. Shirsat Vadivelu Karuppaiah
Vadivelu Karuppaiah Pratap A. Divekar
Pratap A. Divekar Vijay Mahajan1
Vijay Mahajan1
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Insect Sci., 28 February 2025
Sec. Insect Systematics
Volume 5 - 2025 | https://doi.org/10.3389/finsc.2025.1536160
Onion (Allium cepa Linnaeus) is an important vegetable crop valued for its nutritional properties and economics worldwide. Onion cultivation faces serious threats from pests and diseases, particularly onion thrips (Thrips tabaci), which cause substantial yield losses. Recently, Black thrips (Thrips parvispinus), an invasive key pest of chili, have been reported to cause severe damage in onion crop and is likely to devastate the onion cultivation in near future. Therefore, this study was conducted to address the knowledge gap concerning the genetic basis and evolutionary history of T. tabaci and T. parvispinus through sequencing of their mitochondrial genomes. T. tabaci and T. parvispinus were collected from different locations in Maharashtra, India, and reared in the laboratory. The mitochondrial genomes of T. tabaci and T. parvispinus were sequenced to a length of 15,277 and 15,285 bp, respectively. Both genomes exhibited similar gene organization with regard to thirteen protein-coding genes and two rRNA genes. T. tabaci contained 19 tRNA genes whereas T. parvispinus contained 18 tRNA genes. The evolutionary positions of T. tabaci and T. parvispinus within the Thysanoptera order were elucidated through phylogenetic analysis of the mitogenomes of 15 thrips species. These findings provide crucial insights into the genetic makeup and evolutionary dynamics of both the thrips species, thereby aiding the development of novel and sustainable pest management strategies to mitigate their impacts on crops in the changing climate scenario.
Onion (Allium cepa L.) is widely acknowledged as an important vegetable crop worldwide, extensively grown and consumed for its culinary and nutritional value, and plays a vital role in the economies of many countries (1). Onion cultivation is particularly widespread in India, which is one of the world’s leading producer and exporter (2). However, onion production faces numerous challenges due to the prevalence of various pests and diseases, which lead to substantial yield losses (3). Globally, onion growers are concerned about onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae) (4, 5). In India, this pest is considered of national significance, causing 30% to 40% yield loss in onion crops (6, 7). Thrips infestation not only weakens the plants but also makes them susceptible to secondary infections and diseases, which exacerbates economic impacts on farmers (8, 9). These pests are known vectors for various plant viruses, including the iris yellow spot virus (8, 10), and they also transmit diseases like Stemphylium leaf blight and bacterial leaf blight (3, 11–13). Virus-vector relationship have been well documented in the case of thrips; however, their interaction with fungal pathogens remains largely unexplored (Saini et al., 2024). Moreover, thrips usually exhibit genetic heterogeneity, which might be due to the spatial variation in insecticide efficacy (14). Based on mitochondrial DNA sequences, T. tabaci has been classified into three biotypes: one associated with tobacco and two associated with leek (L1 and L2) (15). Also, due to the anticipated increase in temperature, the lifecycle of T. tabaci is likely to shorten, leading to multiple generations within a single crop season (16). Currently, onion growers rely on chemical pesticides to manage these pests, but it often seems futile due to their overlapping generations, concealed feeding behavior, and growing insecticide resistance (17, 18).
Similarly, Thrips parvispinus (Southeast Asian thrips or black thrips), are known to infest a variety of host plants, including vegetables, ornamentals, and field crops. Recently, T. parvispinus has been found infesting onion crops (19), which could pose a substantial threat in the near future (20). The inclusion of T. parvispinus in the list of onion pests reveals a potential gap in our understanding of thrips species interactions and their impact on onions. A comprehensive understanding of the biology, genetics, and evolutionary relationships of thrips is required to tackle these pests in onion and to devise appropriate management strategies. Recent advancements in molecular biology have enabled researchers to explore the genetic makeup of various organisms, providing insight into their evolutionary histories and ultimately helping in devising novel pest management strategies.
The increasing interest in mitochondrial genomes for phylogenetic studies has led to a surge in published mitogenome sequences, particularly amongst insects. Although the complete mitochondrial genomes of several thrips species, such as Thrips imagines (21), Frankliniella occidentalis (22), Frankliniella intonsa (23), Scirtothrips dorsalis (24), Anaphothrips obscurus (25), Thrips palmi (26), Dendrothrips minowai (27), Thrips hawaiiensis (28), Thrips parvispinus (29) and Aptinothrips stylifer (30), have been sequenced, comprehensive research specifically focusing on onion thrips is unexplored. Insect mitogenomes are small, circular, and consist of 37 genes: 13 protein-coding genes (PCGs), two ribosomal RNA genes (rRNAs), and 22 transfer RNA genes (tRNAs), along with a large A+T-rich control region (CR) that regulates transcription and replication (31, 32). Their maternal inheritance, conserved gene content, and rapid evolutionary rate make mitogenomes valuable molecular markers for evolutionary research (33). The significant variation in mitochondrial genome organization observed within the subfamily Thripinae is highly unusual and contrasts with patterns seen in most other animals. The reasons behind the rapid evolution of mitochondrial genomes in Thripinae, as well as the evolutionary dynamics of mitochondrial genomes in other thrips, remain to be explored (25). In this context, the present study on the complete mitogenome of T. tabaci and its phylogenetic implications is of great significance. Furthermore, comparative studies of different thrips mitogenomes will help in elucidating the evolutionary patterns and population dynamics within the Thysanoptera order. Therefore, the study aims to address the knowledge gap by presenting the complete mitochondrial genome sequences of T. tabaci and T. parvispinus. This will offer valuable insights into their evolutionary relationships, population structure, and genetic diversity, ultimately aiding in devising an effective pest management strategy.
Adults of onion thrips were collected from onion plants and initially reared in the laboratory on French beans at the ICAR-Directorate of Onion and Garlic Research (ICAR-DOGR) in Pune, Maharashtra, India (latitude: 18.84°N, longitude: 73.88°E, 616.29 meters above sea level). After completing their life cycle, adults emerged from individual eggs, were collected and used for further analysis. Meanwhile, Black thrips from chili plants in a farmer’s field (latitude: 18.87°N, longitude: 74.05°E, elevation: 667.84 meters) were collected and preserved in 99% ethanol for further analysis. The species identity of these specimens was confirmed through DNA barcoding of the COX-1 gene, and their sequence information has been submitted in the NCBI GeneBank database with accession numbers PP980527 and PP982736 for T. tabaci and T. parvispinus respectively.
Single adults of T. tabaci and T. parvispinus were macerated in liquid nitrogen, and total genomic DNA was extracted using the DNeasy Blood and Tissue Kit (QIAGEN, Germany). The integrity of isolated total DNA was visualized on 1% agarose gel and further quantified using a nanodrop (Bio-Rad, Hercules, California, USA). Mitochondrial DNA was then synthesized from the total DNA using the REPLI-g Mitochondrial DNA Kit (QIAGEN, Germany).
The genome library was constructed with the QIASeq FX DNA kit (QIAGEN, Germany), and sequencing was performed on an Illumina NextSeq 2000 platform using 300-cycle paired-end chemistry, generating primary FASTQ data. These FASTQ files were assessed for total bases, read counts, GC%, Q30, and uncertain base percentages. Reads of high quality were obtained by eliminating adaptor contamination, ambiguous reads, and junk sequences using the fastp tool (v0.12.4) (34). Subsequently, BWA MEM (v0.7.17) was used to align the cleaned reads to the reference sequence (35). Protein-coding and RNA genes were identified from the consensus sequence with SAM tools’ mpileup (36). A de novo assembly using a de Bruijn graph approach was performed to construct longer DNA contigs, and the mitogenome was assembled using the MEGAHIT tool (37) as part of the MitoZ package (38). The quality of assembly was evaluated using the Quast tool (39).
Gene annotation for the mitogenome was conducted with the Prokka annotation tool (40) on the Proksee web server (https://proksee.ca/) (41), producing a circular genome map, GC concentration, and GC skew. For tRNA gene structure prediction and mitogenome assessment, MITOS2 (42) at the Galaxy Europe Web Server (https://usegalaxy.eu/) was used.
The relative synonymous codon usage (RSCU), codon usage, and base composition (A+T contents) of the PCGs were analyzed using MEGA Software (v11.0.13) (43). The GC skewness was computed using the formula GC skew = (G-C)/(G+C); whereas the formula AT skew = (A-T)/(A+T) was used for determining the AT skewness (44). Intergenic spacers and gene overlaps were manually determined. The complete mitochondrial genomes of T. tabaci and T. parvispinus were submitted to the NCBI genome database under accession numbers PQ197393 and PQ197392, respectively.
The evolutionary studies related to T. tabaci and T. parvispinus with the metagenome of 14 other thrips species were analyzed using the NCBI retrieved sequence information. The damsel bug, Alloeorhynchus bakeri, was used as an outgroup species. The nucleotide sequences of 13 protein coding genes of 16 thrips species and one outgroup species were aligned individually using MAFFT 7, the Database of Aligned Structure Homologue (DASH) was utilized to incorporate homologous structures based on amino acid codons (45). Followed by removal of the ambiguously aligned sites, the aligned amino acid sequences were then converted to the nucleic acid sequence. For phylogenetic analysis the sequences of 13 PCGs were concatenated in following order: nad5, nad4, nad4L, nad6, cox1, nad3, cox2, cox3, atp6, atp8, nad1, nad2 and cytB in a single sequence of each species. All the 17 sequences were aligned using CLustalW tool (46). The phylogenetic tree was constructed using the Maximum Likelihood method in MEGA 11 software (43), employing the general time reversible model with gamma distribution (GTR+G) and bootstrap values from 500 iterations.
The circular genome of T. tabaci is 15,277 bp long (Figure 1A), comprising 34 sequence elements (such as 13 PCGs, 19 tRNAs, and 2 rRNA-coding genes). Strand localization analysis revealed 19 genes located on the H-strand (+), whereas 15 genes on the L-strand (–) (Table 1). Gene length in the T. tabaci mitochondrion was 400 bp on average, with minimum and maximum lengths of 57 bp (trnS1) and 1695 bp (nad5), respectively. Base compositions of the complete mitochondrial genome of T. tabaci were 41.3%, 34.8%, 12%, and 11.9% for A, T, G, and C nucleotides. The 13 PCGs and two rRNAs had AT contents ranging from 69.80% to 82.50%, whereas the overall mitogenome had 76.10%. In contrast, the GC content varied from 17.50% to 30.20% for the 13 PCGs and 2 rRNAs, whereas the overall mitogenome had 23.90%.
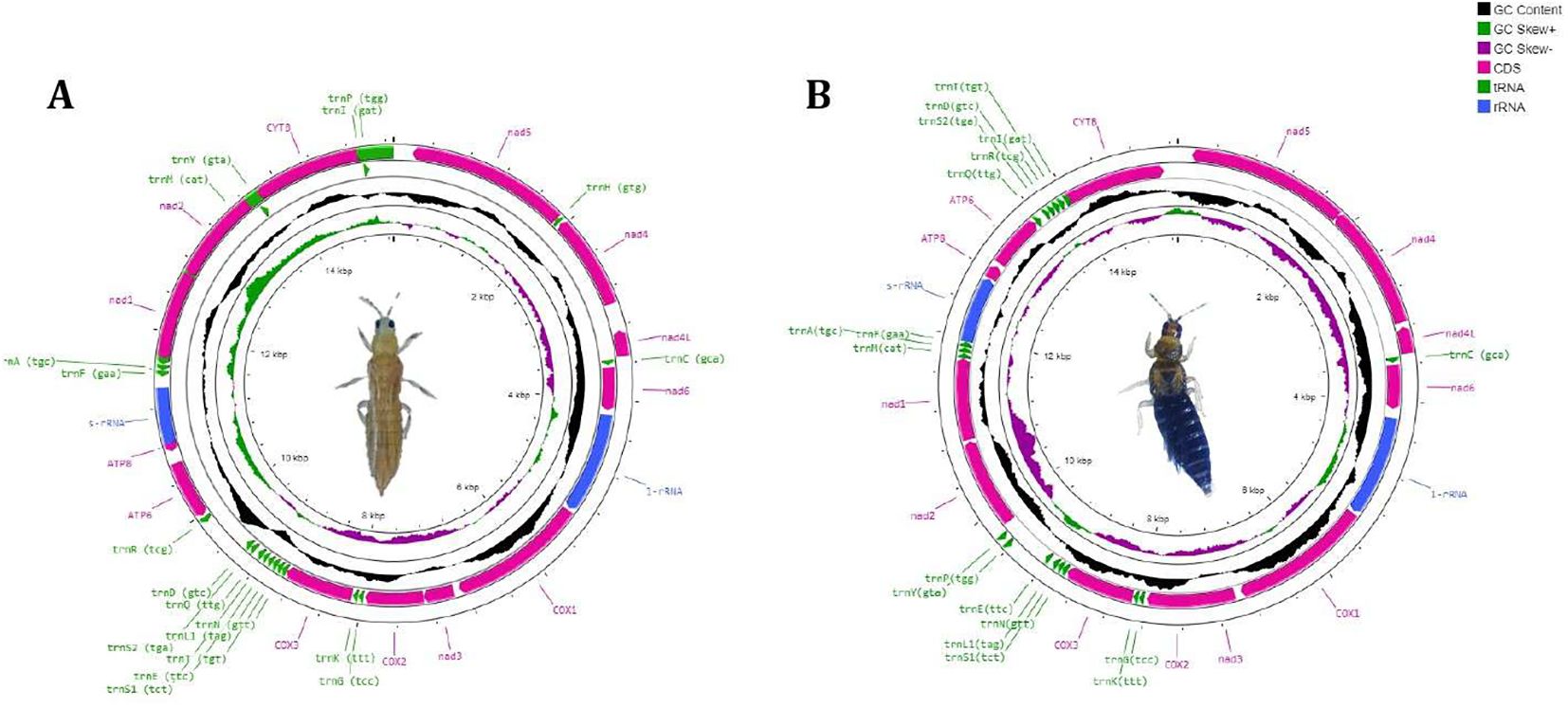
Figure 1. Circular genome map of (A) T. tabaci and (B) T. parvispinus showing mitogenome sequence features.
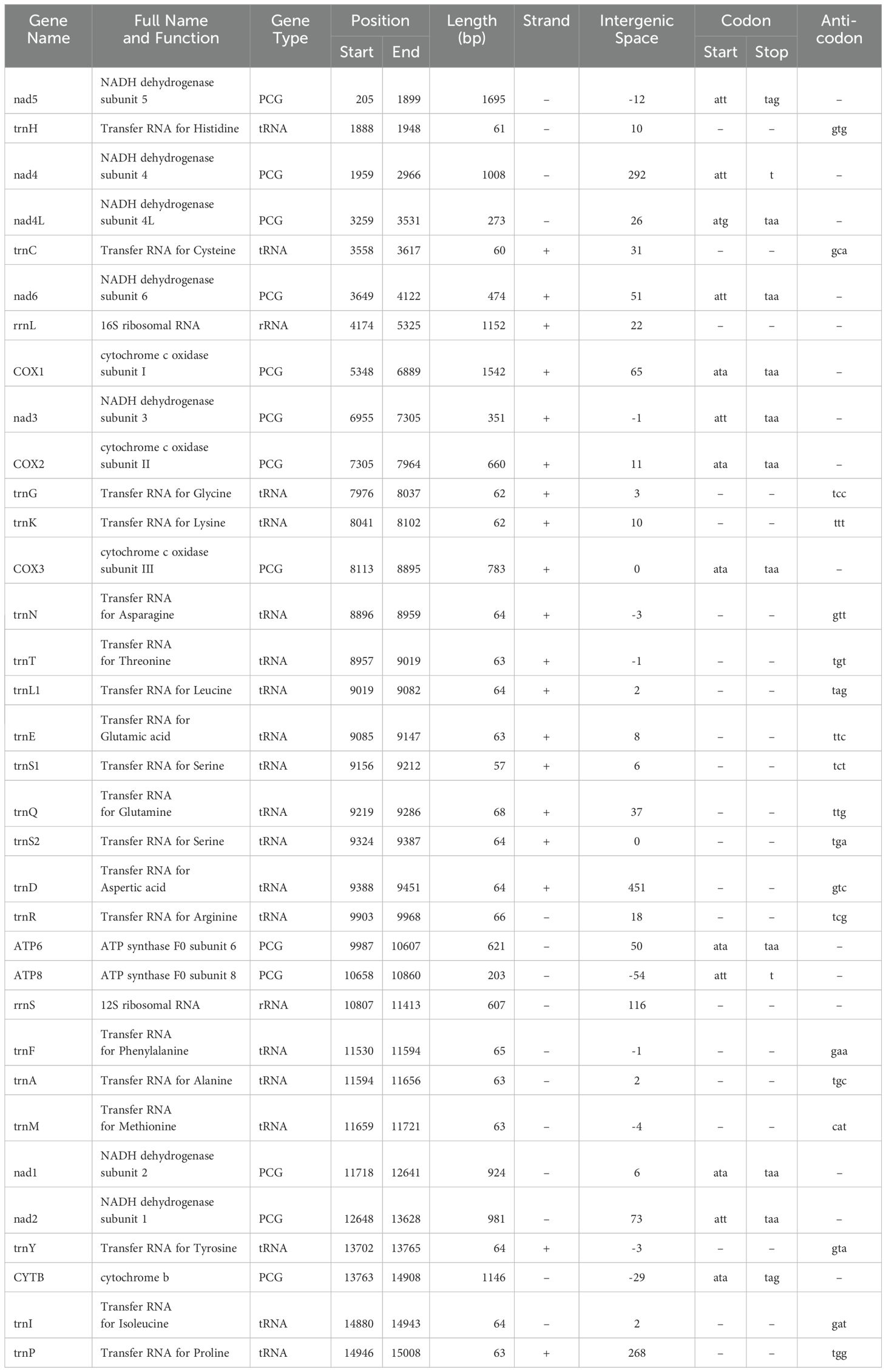
Table 1. Summary of Thrips tabaci mitochondrial genome.
The AT skewness for the 13 PCGs ranged from 0.426 to 0.033, higher than that of the rRNA genes, rrnS (-0.193) and rrnL (0.180) (Table 2). The 1560 bp of intergenic nucleotides were spread across 23 locations, with individual spacer lengths ranging from 2 to 451 bp. The longest intergenic spacer (451 bp) was found between the trnD and trnR genes. There were overlaps between nine genes, whose lengths varied from 1 to 54 bp. The atp8 and rrnS genes had the lengthiest overlap, 54 bp, as shown in Table 1 and Figure 1A.
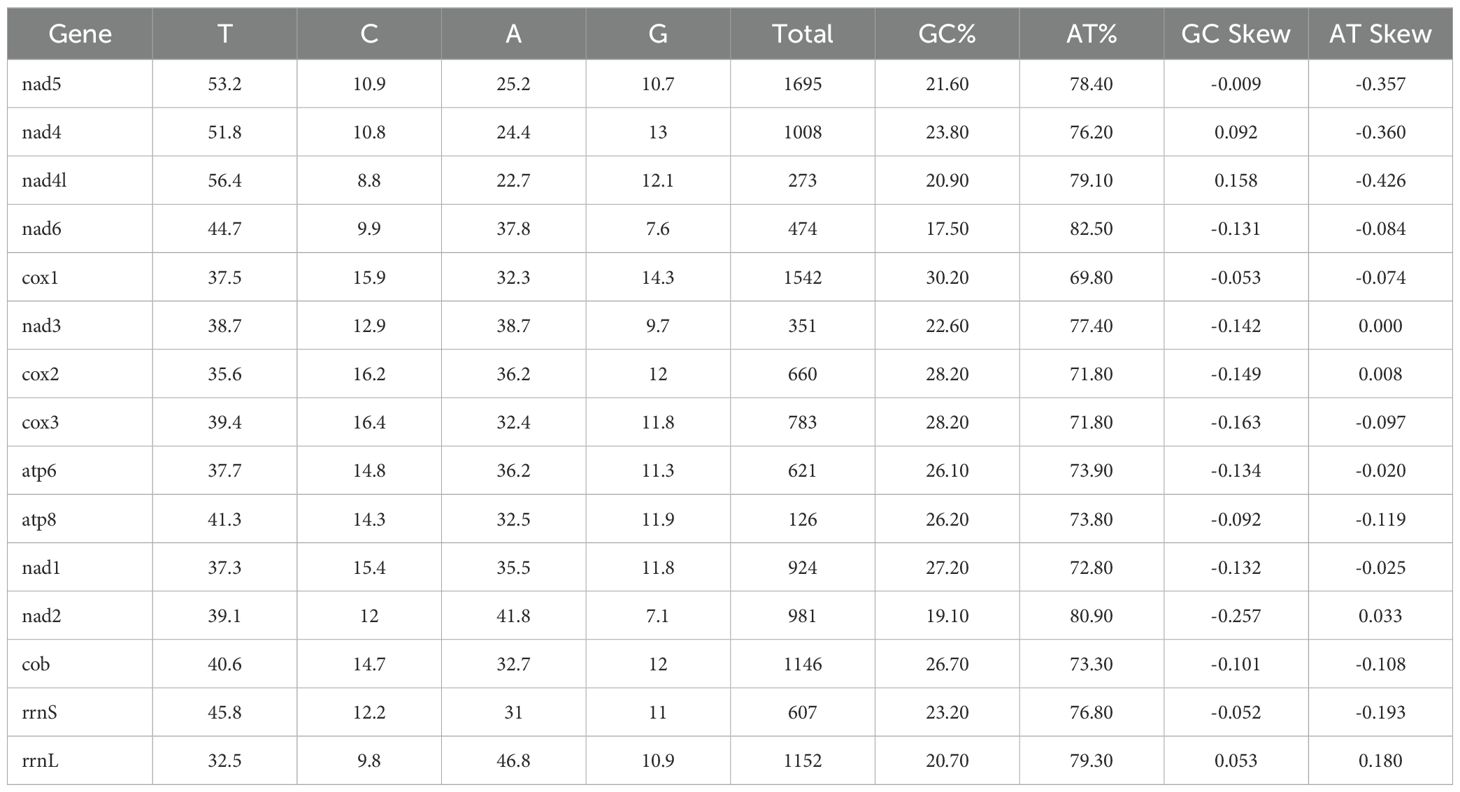
Table 2. Nucleotide composition of Thrips tabaci mitogenome.
Likewise, the circular mitogenome of T. parvispinus is 15,285 bp, comprising 13 PCGs, 18 tRNA-coding genes, and 2 rRNA-coding genes, 28 of which are located on the H-strand (+) the remaining 5 on the L-strand (–). Gene length in the T. parvispinus mitochondrion was 428 bp on average, with minimum and maximum lengths of 61 bp (trnS1) and 1695 bp (nad5), respectively (Figure 1B, Table 3). Our result was in accordance with Pakrashi et al. (29), revealing a mitogenome length of 15,067 bp. The mitogenome of T. parvispinus had base compositions of 43.3% A, 30% T, 10% G, and 11.7% C, with an overall AT content of 73.3% and GC content of 21.7%. The AT content across the 13 PCGs and 2 rRNAs ranged from 72.8% to 84%, while the GC content varied from 17% to 27.2%. AT skewness was higher in the PCGs (0.064 to 0.410) compared to the rRNAs (0.161 for rrnS and 0.187 for rrnL) (Table 4). We identified 1,039 bp of intergenic nucleotides across 24 locations, with the longest spacer (347 bp) between the trnE and trnP genes. Seven genes had overlaps, with the largest (35 bp) between the COX2 and ND4 genes (Table 3, Figure 1B).
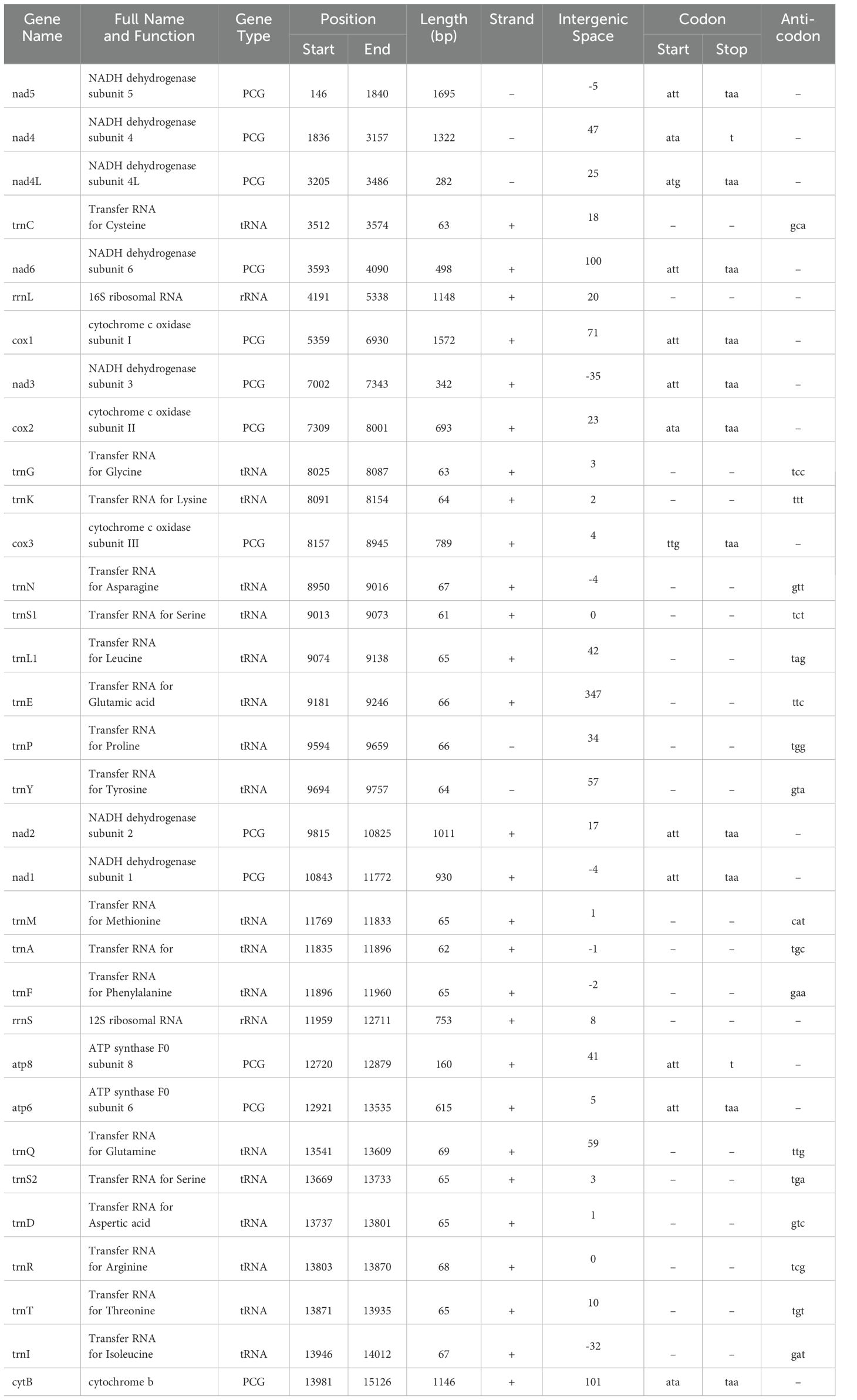
Table 3. Summary of Thrips parvispinus mitochondrial genome.
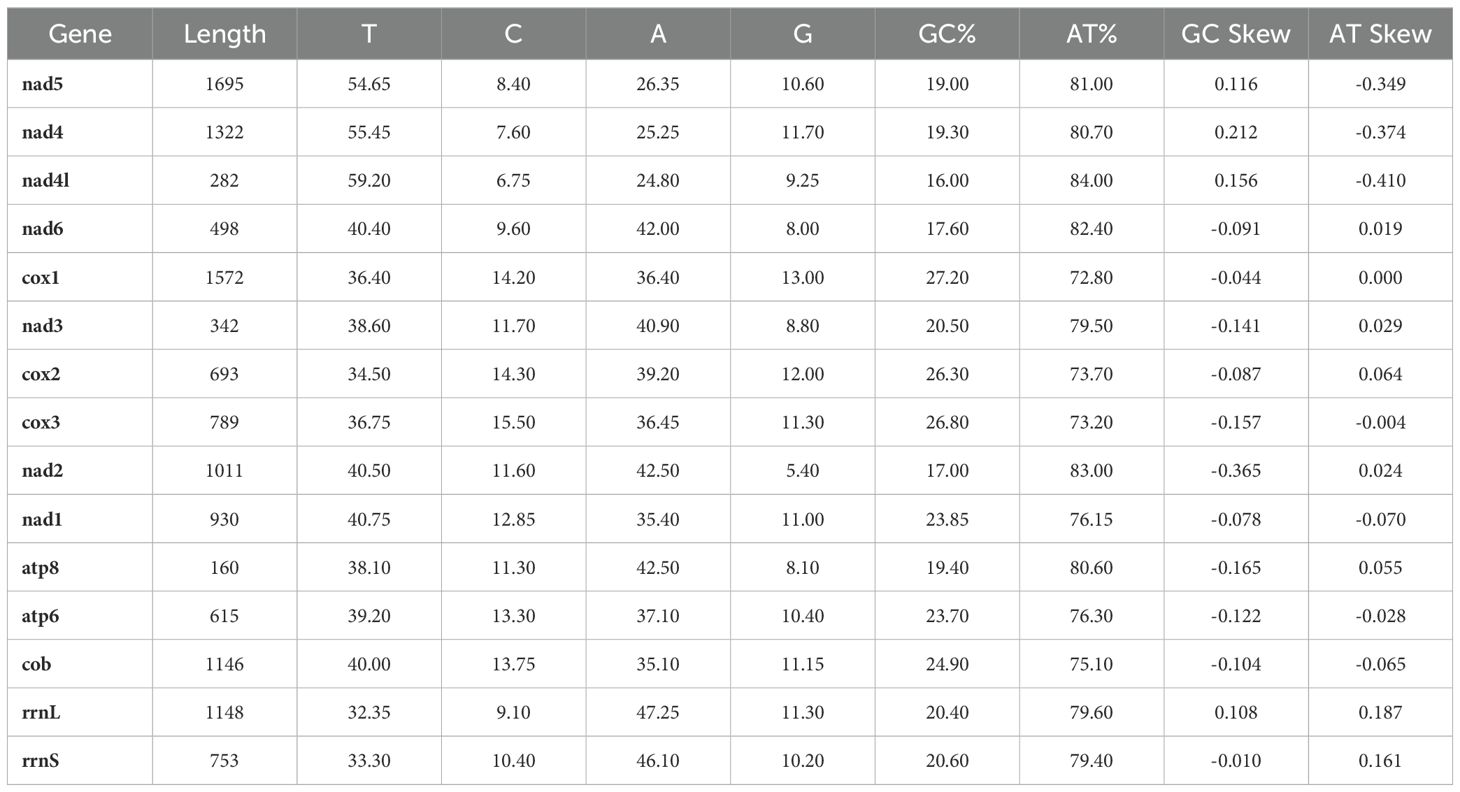
Table 4. Nucleotide composition of Thrips parvispinus mitochondrial genome.
The mitochondrial genomes of T. tabaci and T. parvispinus exhibit distinct characteristics in their PCGs. The total lengths of the 13 PCGs were 10,584 bp for T. tabaci and 11,055 bp for T. parvispinus. In T. tabaci, the coding sequences ranged from 126 bp (atp8) to 1,542 bp (cox1), with genes such as cox1, cox2, cox3, nad3, and nad6 located on the H-strand, while others like nad1, nad2, nad4, nad4L, nad5, atp6, atp8, and cytB were on the L-strand. Conversely, in T. parvispinus, the coding sequences ranged from 160 bp (atp8) to 1,695 bp (nad5), with most genes situated on the H-strand, except nad4, nad4L, and nad5, which were on the L-strand.
Three start codons (ATT, ATA, and ATG) were identified in the T. tabaci mitogenome. ATT was adopted by genes like nad2, nad3, nad4, nad5, nad6, and atp8, whereas genes like cox1, cox2, cox3, atp6, nad2, and CytB used ATA. However, ATG was used as the start codon by the nad4L gene. In contrast, T. parvispinus utilized four start codons, with ATT being the most prevalent across 8 genes (cox1, atp6, atp8, nad1, nad2, nad3, nad5, and nad6), while 3 genes (nad4, cox2, and cytB) used ATA as their start codon. The ATG and TTG start codons are used by nad4L and cox3, respectively. The ATN start codon is the most common starting codon in most thrips and other insect species (26, 47–49). For stop codons, T. tabaci primarily used TAA, but nad5 and cytB used TAG, and nad4 and atp8 used an incomplete stop codon “T”. Similarly, T. parvispinus mostly used TAA, with nad4 and atp8 also adopting the incomplete “T”. Incomplete termination codons are common in other thrips species (26, 47) and insect mitochondrial genomes and are presumed to be restored through post-transcriptional polyadenylation (48, 49).
The RSCU and amino acid usage in the PCGs of T. tabaci and T. parvispinus are summarized in Figure 2 and Table 5. In T. tabaci mitogenome, leucine, phenylalanine, isoleucine, serine, lysine, tyrosine, and valine were the most frequent amino acids, whereas tryptophan, methionine, and cyanine were the rarest. In general, UUU, followed by UUA, AUU, AUA, and AAA, was the most frequently used codon. However, in the T. parvispinus mitogenome, isoleucine, phenylalanine, leucine, asparagine, serine, lysine, and tyrosine were the most frequently used amino acids, whereas tryptophan, cyanine, and methionine were the rarest, with UUU, followed by AUU, UUA, AAU, and AUA being the most frequently used codons (Table 5).
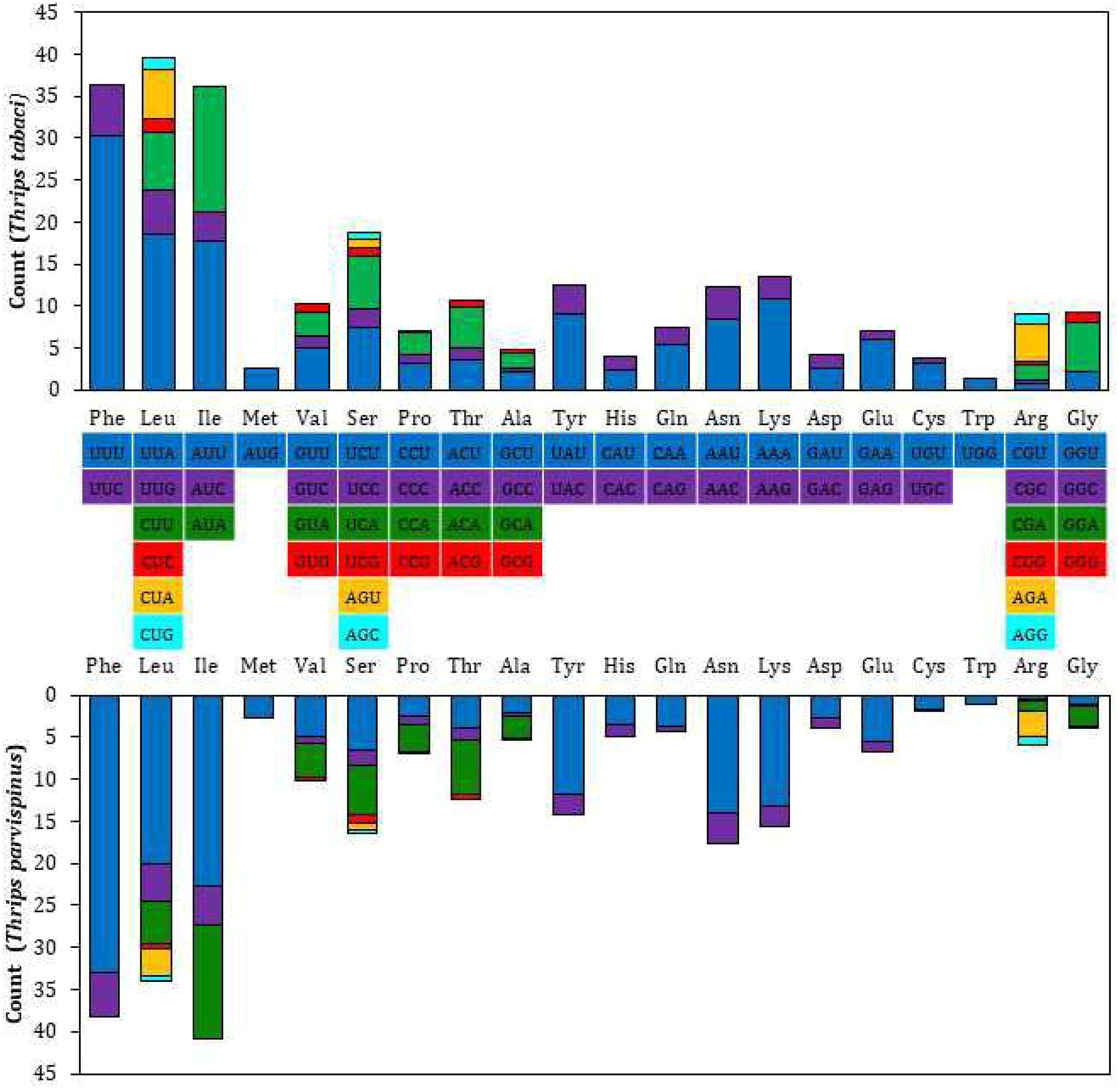
Figure 2. Relative synonymous codon use (RSCU) of 13 PCG’s of the T. tabaci and T. parvispinus.
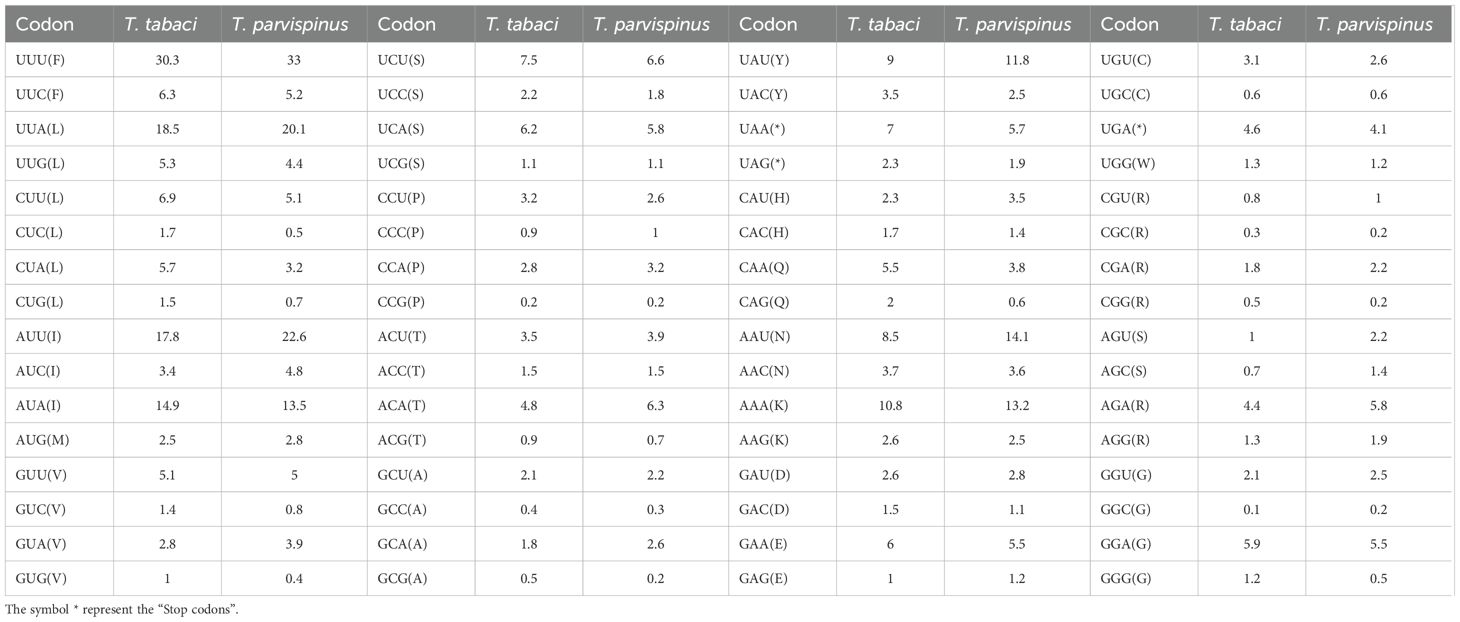
Table 5. Amino acid usage in the 13 PCG’s of the T. tabaci and T. parvispinus.
The circular mitochondrial genome of T. tabaci contains 19 tRNA genes ranging in size from 57 to 68 bp. In the mitogenome, 13 of the tRNA genes are located on the H-strand (+), while the remaining six are located on the L-strand (–) (Figure 1A and Table 1). The 19 tRNA-coding genes of T. tabaci mitochondrial genome collectively comprised 1,200 bp, representing 7.85% of the entire mitogenome. In T. parvispinus mitogenome, 18 tRNA-coding genes ranging in size from 61 to 69 bp were detected. All of the tRNA-coding genes of T. parvispinus were located on the H-strand (+) with the exception of two genes encoding proline (Pro/P) and tyrosine (Tyr/Y) located on the L-strand (-) of mitogenome (Figure 1B and Table 3). These 18 tRNA-coding genes of T. parvispinus comprise a 1,170-bp region that covers 7.65% of the mitogenome. The characteristic clover leaf secondary structures of tRNA genes were identified in the mitogenomes of both thrips species, with the exception of trnS1, which lacks a dihydrouridine arm (Figures 3, 4). Three genes encoding tRNAs for leucine (Leu/L2), tryptophan (Trp/W), and valine (Val/V) were lacking in the mitogenome of T. tabaci; however, in addition to these three genes, histidine (His/H) was missing in the T. parvispinus mitogenome.
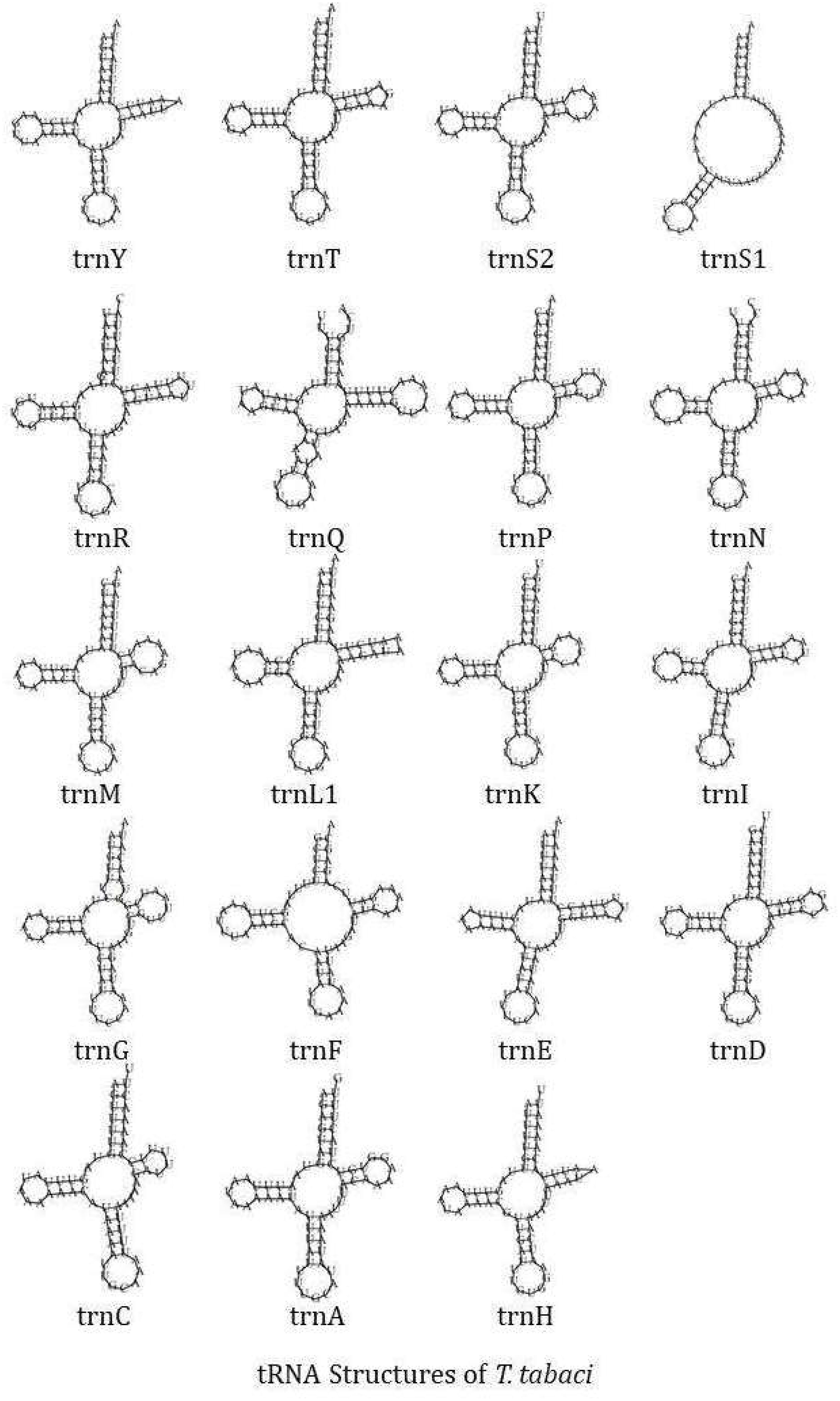
Figure 3. tRNA structures of the T. tabaci.
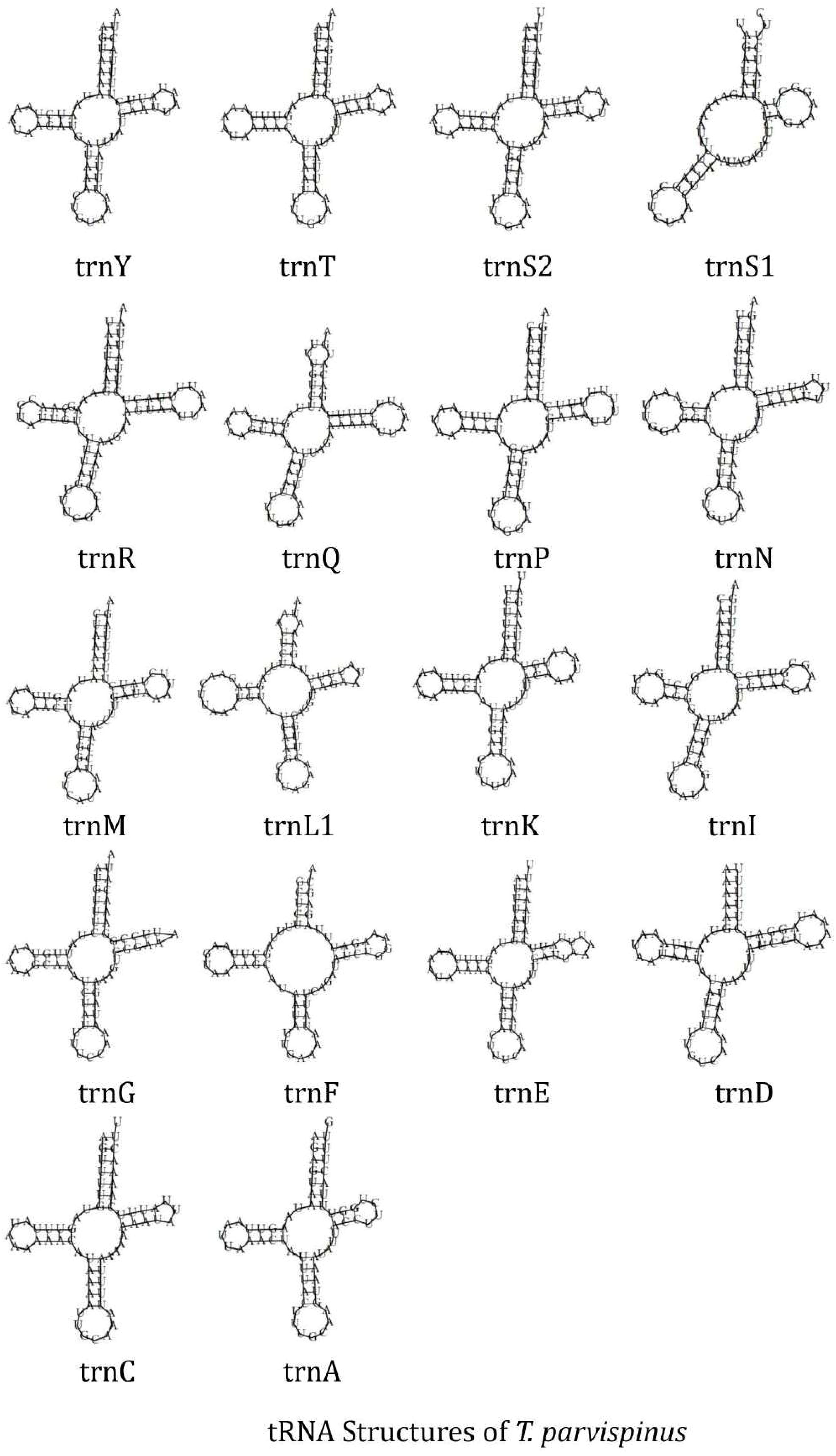
Figure 4. tRNA structures of the T. parvispinus.
The rRNA genes rrnL and rrnS collectively constitute 11.51% of the total mitogenome. The rrnS gene (607 bp long with 76.80% A+T content) was situated between the atp8 and tRNA-Phe genes. The rrnL gene (1,152 bp long with 79.30% A+T content) was located between the nad6 and cox1 genes of the T. tabaci mitogenome. However, in the T. parvispinus mitogenome, both the rRNA-coding genes rrnL (1148 bp) and rrnS (753 bp) were found on the H-strand (+) and covered 12.43% (1901 bp) of the whole mitochondrial genome.
Species within the same family, Thripidae, T. palmi, T. imagines, T. hawaiiensis, F. intonsa, F. occidentalis, Dendrothrips minowai, Pseudodendrothrips mori, Neohydatothrips samayunkur, Scirtothrips dorsalis, Anaphothrips obscurus, were grouped together with T. tabaci and T. parvispinus, The species from other thrips family such as Stenurothripidae, Aeolothripidae and Phlaeothripidae were grouped in another cluster. The T. parvispinus was closely related to T. hawaiiensis with 98 bootstrap value. However, T. tabaci was slightly outgrouped from T. hawaiiensis. The F. intonsa, and F. occidentalis were cluster separately from species for genus Thrips (Figure 5). The other cluster included Holarthothrips indicus of the Stenurothripidae family grouped with 51 Bootstrap value with Franklinothrips vespiformis of the Aeolothripidae family, Gynaikothrips uzeli, and Haplothrips aculeatus of the Phlaeothripidae family. The damsel bug, Alloeorhynchus bakeri, was used in the dataset as an outgroup species.
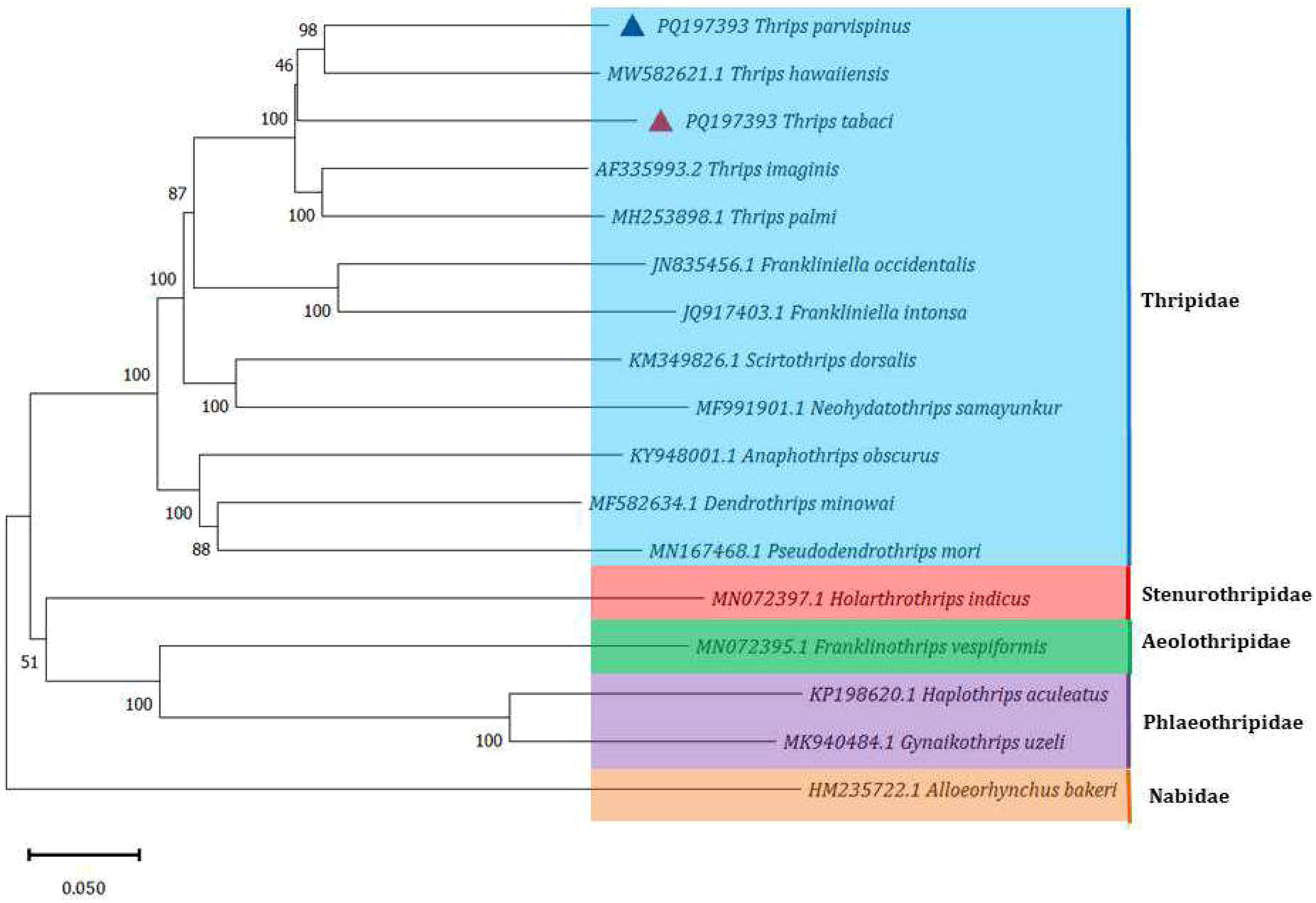
Figure 5. Phylogenetic analysis of 16 Thrips species, with the damsel bug, Alloeorhynchus bakeri as outgroup. (T. tabaci and T. parvispinus marked with red and blue triangles respectively are from current study).
The T. tabaci and T. parvispinus mitochondrial genome contains one control in both species, each of which is 451 bp and 347 bp, respectively. In T. tabaci, it is located between the trnD and trnR genes; however, in T. parvispinus, it is situated between the trnE and trnP genes. The percentages of AT in the control T. tabaci and T. parvispinus regions were 62.7% and 76.7%, respectively. T-stretch, GAnT motif, ATnC motif, tandem repeats, and stem loops were found in the control region of the T. tabaci mitogenome. However, the T-stretch followed by the TATA box, GAnT motif, tandem repeats, and stem loop were found in the T. parvispinus mitogenome.
The T. tabaci mitochondrial genomes contain 22 intergenic spacer regions of length 1109 bp that vary in length from 2 to 292 bp. There were 13 major intergenic spacers >10 bp in length observed in the circular genome. The longest intergenic spacer regions (292 bp) were located between nad4 and nad4L genes. However 9 overlapping sequences were found in T. tabaci mitochondrial genome, ranging in size from 1 to 54 bp. Similar to T. tabaci, T. parvispinus mitochondrial genome contained 22 intergenic spacer regions of total 692 bp, varying in length from 1 to 101 bp. There were 14 major intergenic spacers >10 bp in length observed in the T. parvispinus mitogenome. The cytB genes were followed by the longest intergenic spacer region of 101 bp. The 7 overlapping sequences were present in T. tabaci mitochondrial genome, ranging from 1 to 35 bp in size. Amid the nad4 and nad4L genes of the T. tabaci mitogenome, there were 292 bp noncoding nucleotides; however, T. parvispinus revealed 47 bp.
The circular mitogenome of T. tabaci and T. parvispinus from the present study revealed 15,277 bp and 15,285 bp. respectively. A similar trend was observed in most of the thrips species reported earlier. For instance, T. imagines (15,407 bp) (21), T. palmi (15,333 bp) (26), Frankliniella intonsa (15,215 bp) (23), Frankliniella occidentalis (14,889 bp) (22), Scriptothrips dorsalis (15,343 bp) (24), and Anaphothrips obscurus (14,890 bp) (25).
The gene order among these thrips species showed variations, especially within the subfamily Thripinae. In T. tabaci and T. parvispinus the several tRNA genes from both T. tabaci and T. parvispinus have been translocated, showing the variation and arrangement of the gene. Similar variation in the order of the mitogenome genes of the all thrips species was observed. This variation can provide insights into evolutionary processes and mechanisms of genetic diversity. It also highlights the complexity of mitochondrial inheritance and their adaptation to various environments at the genetic level (47). These variations might affect their physiology, behavior, and ecological interactions, making them an interesting subject for evolutionary studies (48). Additionally, understanding these evolutionary patterns can help in pest control strategies, as some thrips species are significant agricultural pests.
The RSCU data analysis revealed that Lysine followed by phenylalanine, leucine, isoleucine, tyrosine, and serine, are the most frequently used amino acids, which is common in most of the thrips species (23, 25, 26). The characteristic clover leaf secondary structures of tRNA genes were identified in the mitogenomes of both thrips species, with the exception of trnS1, which lacks a dihydrouridine (DHU) arm, which is common in most insect species. The DHU arm in the trnS1 secondary structure was missing in marigold thrips (Neohydatothrips samayunkur) (49), green semilooper (Chrysodeixis acutaI) (50), and Indian dammer bee, Tetragonula iridipennis (51).
In both the thrips species, some tRNA-coding genes, namely Leucine (Leu/L2), Tryptophan (Trp/W), and Valine (Val/V) were missing in T. tabaci, and in addition to these three, Histidin (His/H) is missing in T. parvispinus mitogenome. The presence of duplicate copies of tRNA in some thrips species and missing tRNA genes in Gynaikothrips has been reported by Tyagi et al. (52). However in many insect species mitochondrialt tRNA genes reported as lost or missing, on manual annotation found to have unusual secondary structures and contain many nucleotide mismatches (53). In some of the insect species, the truncated tRNA was observed, which formed during their evolution (54). The truncation of tRNA genes poses problems in locating and annotating them due to a high level of nucleotide mismatches (55). In the absence of a well-paired acceptor stem, the 3’ end is not clearly defined. The region downstream from the anticodon stem is extremely variable in sequence and length (54).
The 7 overlapping sequences were present in the T. tabaci mitochondrial genome, ranging from 1 to 35 bp in size. Amid the nad4 and nad4L genes of the T. tabaci mitogenome, there were 292 bp noncoding nucleotides; however, T. parvispinus revealed 47 bp, as in Aeolothrips xinjiangensis (148 bp) (48). Most of the thrips mitogenomes exhibited overlaps of 1-21 bp (mostly around 7 bp) between the nad4 and nad4L. In insect mitogenomes, the nd4 to nd4L regions are transcribed into polycistronic mRNA with either overlaps or no intergenic spacers between them (56–58). However, such a long intergenic spacer might split this polycistronic mRNA into two monocistronic mRNAs (48).
The phylogenetic analysis revealed that the genera Thrips and Frankliniella are closely related, as they cluster together. This close relationship is attributed to their shared homology of paired ctenidia on abdominal segments V-VIII (59). T. tabaci, T. palmi, F. accidentalis, and F. intonsa are known vectors of topoviruses (47). These species cluster together, suggesting they share similar genomic characteristics.
As like the genomic characteristics, the biological similarities are also present in T. tabaci and T. parvispinus. Field emission scanning electron microscopy (FESEM) analysis of both thrips species identified similar types of sensilla, including sensilla basiconica (SBI, SBII, SBIII), sensilla chaetica (SChI, SChII), sensilla trichodea (ST), sensilla campaniformia (SCa), and sensilla cavity (SCav); however, variations in the length of these sensilla were observed between the two species. Also, some morphological characters have the variation in both the thrips species, such as antennae (seven-segmented with forked sensorium on third, and fourth segments), ctenidia (paired ctenidia were present in 5th–8th abdominal segments laterally), and pronotum (two pairs of posteroangular setae) (60).
Mitochondrial genome data have been widely used for phylogenetic, evolutionary studies, and population genetics in insects (61). Among eukaryotes, Thysanoptera, along with other minor paraenopteran insect orders, is regarded as a model for rapid mitochondrial genome evolution (24). In the Thysanoptera order, thrips display exceptional interspecific variation in mitogenomic structure, making them an ideal model for studying mitochondrial evolution (48). In the current study, the several tRNA genes from both T. tabaci and T. parvispinus have been translocated; however, the protein-coding genes and most of the tRNA genes have a similar arrangement, representing that these two species have a low rate of the rearrangement among them, which may be originating from the most recent common ancestors of these thrips species. Similar results were reported by Yan et al. (23) in flower thrips, Frankliniella intonsa, and other thrips species. Mitochondrial DNA plays a crucial regulatory role in insect adaptation to environmental changes, including insecticide resistance (62). Many insecticides target mitochondrial functions, such as oxidative phosphorylation. Mutations in mtDNA can lead to changes in these functions, resulting in resistance to the insecticides (63). Mutations in the mitochondrial-encoded Cytb gene have been implicated in resistance to the novel acaricide Bifenazate in Tetranychus urticae and Panonychus citri (64). Furthermore, a newly identified mutation in the Cytb gene of T. urticae has been associated with resistance to the miticide Acequinocyl (65). However, in the current study, no such mutation was detected in the mitochondrial genes. Also, no report was found in any thrips species to date detecting insecticidal resistance in the thrips species due to mutation in the mitochondrial gene.
The phylogenetic analysis includes species from different thrips families, such as Thripidae, Aeolothripidae, Phlaeothripidae, and Stenurothripidae, showing the phylogenetic diversity among these groups. The diversity within these groups may contribute to the complexity and lower bootstrap values in some parts of the tree. The thrips species clustered together such as T. hawaiiensis and T. parvispinus, with bootstrap values 98, have closely related species T. tabaci with lowest bootstrap value 46, suggesting lower confidence in their relationships. The thrips species in phylogenetic study other than the Thripidae family, Holarthrothrips indicus of Stenurothripidae family, and Franklinothrips vespiformis of Aeolothripidae, Gynaikothrips uzeli and Haplothrips aculeatus of the family Phlaeothripidae shown the lower boot strap values, 51. The lower bootstrap values of these species could be because of the low rate of genetic flow between these species (66).
Complete mitochondrial genome sequences of T. tabaci and T. parvispinus provide critical insights into their genetic makeup and evolutionary relationships. The mitochondrial genomes of T. tabaci and T. parvispinus were found to be 15,277 bp and 15,285 bp in size, respectively, with similar gene organization, including 13 protein-coding genes, 2 rRNA genes, and 19 tRNA genes in T. tabaci, and 18 tRNA genes in T. parvispinus. The high AT content observed in both species reflects their typical insect mitochondrial genome characteristics, which are essential for understanding their genetic diversity and evolutionary adaptations. Phylogenetic analysis revealed the evolutionary positions of T. tabaci and T. parvispinus within the Thysanoptera order, highlighting the genetic makeup and taxonomy of these species with other thrips species. The findings emphasize the significance of understanding the genetic basis of onion thrips, which can aid in developing targeted pest management strategies to mitigate their impact on onion production. The detailed mitochondrial genome sequences and phylogenetic analyses presented in this study provide a valuable resource for further research on thrips biology, genetics, and pest management.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
The manuscript presents research on animals that do not require ethical approval for their study.
PS: Conceptualization, Data curation, Formal analysis, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. DS: Data curation, Formal analysis, Methodology, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. VK: Data curation, Formal analysis, Methodology, Resources, Supervision, Writing – original draft, Writing – review & editing. PD: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. VM: Funding acquisition, Resources, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The author(s) declare that funding for this study, was received from the in-house project of ICAR-Directorate of Onion and Garlic Research, Pune, India with Project code: 052/S/IPP/DOGR/21-26/CPT/003.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sharma K, Mahato N, Nile SH, Lee ET, Lee YR. Economical and environmentally-friendly approaches for usage of onion (Allium cepa L.) waste. Food Funct. (2016) 7:3354–69. doi: 10.1039/C6FO00251J
2. Gulati A, Wardhan H, Sharma P. Tomato, onion and potato (TOP) value chains. In: Gulati A, Ganguly K, Wardhan H, editors. Agricultural value chains in India: ensuring competitiveness, inclusiveness, sustainability, scalability, and improved finance. Singapore: Springer Nature Singapore (2022). p. 33–97. Available at: https://library.oapen.org/handle/20.500.12657/52430 (Accessed October 10, 2024).
3. Leach A, Reiners S, Nault B. Challenges in integrated pest management: A case study of onion thrips and bacterial bulb rot in onion. Crop Prot. (2020) 133:105123. doi: 10.1016/j.cropro.2020.105123
4. Nawrocka B, Vidal S. Economic importance and the control method of Thrips tabaci Lind. on onion. IOBC WPRS Bull. (2003) 26:321–4.
5. Diaz-Montano J, Fuchs M, Nault BA, Fail J, Shelton AM. Onion thrips (Thysanoptera: Thripidae): a global pest of increasing concern in onion. J economic entomology. (2011) 104:1–13. doi: 10.1603/EC10269
6. Gill HK, Garg H, Gill AK, Gillett-Kaufman JL, Nault BA. Onion thrips (Thysanoptera: Thripidae) biology, ecology, and management in onion production systems. J Integrated Pest Manage. (2015) 6:6. doi: 10.1093/jipm/pmv006
7. Soumia PS, Karuppaiah V, Singh M. Managing Thrips tabaci: a pest of national significance. Indian Horticulture. (2017) 62:55–6. https://epubs.icar.org.in/index.php/IndHort/article/view/87714 (Accessed October 10, 2024).
8. Khandagale K, Roylawar P, Randive P, Karuppaiah V, Soumia PS, Shirsat D, et al. Isolation and expression profiling of insecticidal lectins from wild alliums against onion thrips (Thrips tabaci lindeman). Proc Natl Acad Sciences India Section B: Biol Sci. (2022) 92:451–9. doi: 10.1007/s40011-022-01360-4
9. Iglesias L, Havey MJ, Nault BA. Management of onion thrips (Thrips tabaci) in organic onion production using multiple IPM tactics. Insects. (2021) 12:207. doi: 10.3390/insects12030207
10. Gawande SJ, Anandhan S, Ingle A, Roylawar P, Khandagale K, Gawai T, et al. Microbiome profiling of the onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae). PloS One. (2019) 14:e0223281. doi: 10.1371/journal.pone.0223281
11. Leach A, Hay F, Harding R, Damann KC, Nault B. Relationship between onion thrips (Thrips tabaci) and Stemphylium vesicarium in the development of Stemphylium leaf blight in onion. Ann Appl Biol. (2020) 176:55–64. doi: 10.1111/aab.12558
12. Grode A, Chen S, Walker ED, Szendrei Z. Onion thrips (Thysanoptera: Thripidae) feeding promotes infection by Pantoea ananatis in onion. J economic entomology. (2017) 110:2301–7. doi: 10.1093/jee/tox273
13. Grode AS, Brisco-McCann E, Wiriyajitsonboom P, Hausbeck MK, Szendrei Z. Managing onion thrips can limit bacterial stalk and leaf necrosis in Michigan onion fields. Plant Dis. (2019) 103:938–43. doi: 10.1094/PDIS-07-18-1271-RE
14. Khan AA, Kanwal H, Batool S, Hassan Z, Shah JM, Aatif HM, et al. Impact of Predators and Some Commercial Insecticides on Thrips tabaci Lindeman and Amrasca biguttula Ishida Populations in Bt Cotton under Arid Climate. Pakistan J Zoology. (2022) 54:137–44. doi: 10.17582/journal.pjz/20200512190543
15. Khan F, Roy MC, Kim Y. Thelytokous reproduction of onion thrips, Thrips tabaci Lindeman 1889, infesting welsh onion and genetic variation among their subpopulations. Insects. (2022) 13:78. doi: 10.3390/insects13010078
16. Karuppaiah V, Maruthadurai R, Das B, Soumia PS, Gadge AS, Thangasamy A, et al. Predicting the potential geographical distribution of onion thrips, Thrips tabaci in India based on climate change projections using MaxEnt. Sci Rep. (2023) 13:7934. doi: 10.1038/s41598-023-35012-y
17. Morse JG, Hoddle MS. Invasion biology of thrips. Annu Rev entomology. (2006) 51:67–89. doi: 10.1146/annurev.ento.51.110104.151044
18. Martin NA, Workman PJ, Butler RC. Insecticide resistance in onion thrips (Thrips tabaci)(Thysanoptera: Thripidae). New Z J Crop Hortic Sci. (2003) 31:99–106. doi: 10.1080/01140671.2003.9514242
19. Saini S, Raj K, Saini AK. Effect of Thrips parvispinus injury in enhancing onion purple blotch severity. Plant Dis Res. (2022) 37:246–6.
20. Saini S, Raj K, Saini AK, Kumar R, Saini A, Khan A, et al. Unravelling the synergistic interaction of Thrips tabaci and newly recorded, Thrips parvispinus with Alternaria porri (Ellis.) Cif., inciting onion purple blotch. Front Microbiol. (2024) 15:1321921. doi: 10.3389/fmicb.2024.1321921
21. Shao R, Barker SC. The highly rearranged mitochondrial genome of the plague thrips, Thrips imaginis (Insecta: Thysanoptera): convergence of two novel gene boundaries and an extraordinary arrangement of rRNA genes. Mol Biol Evol. (2003) 20:362–70. doi: 10.1093/molbev/msg045
22. Yan D, Tang Y, Xue X, Wang M, Liu F, Fan J. The complete mitochondrial genome sequence of the western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae) contains triplicate putative control regions. Gene. (2012) 506:117–24. doi: 10.1016/j.gene.2012.06.022
23. Yan D, Tang Y, Hu M, Liu F, Zhang D, Fan J. The mitochondrial genome of Frankliniella intonsa: insights into the evolution of mitochondrial genomes at lower taxonomic levels in Thysanoptera. Genomics. (2014) 104:306–12. doi: 10.1016/j.ygeno.2014.08.003
24. Dickey AM, Kumar V, Morgan JK, Jara-Cavieres A, Shatters RG, McKenzie CL, et al. A novel mitochondrial genome architecture in thrips (Insecta: Thysanoptera): extreme size asymmetry among chromosomes and possible recent control region duplication. BMC Genomics. (2015) 16:1–7. doi: 10.1186/s12864-015-1672-4
25. Liu H, Li H, Song F, Gu W, Feng J, Cai W, et al. Novel insights into mitochondrial gene rearrangement in thrips (Insecta: Thysanoptera) from the grass thrips, Anaphothrips obscurus. Sci Rep. (2017) 7:4284. doi: 10.1038/s41598-017-04617-5
26. Chakraborty R, Tyagi K, Kundu S, Rahaman I, Singha D, Chandra K, et al. The complete mitochondrial genome of Melon thrips, Thrips palmi (Thripinae): Comparative analysis. PloS One. (2018) 13:e0199404. doi: 10.1371/journal.pone.0199404
27. Chen SC, Jiang HY, Peng P, Hu X, Lin Q, Wang XQ. The complete mitochondrial genome of the stick tea thrips Dendrothrips minowai (Thysanoptera: Thripidae). Mitochondrial DNA Part B. (2018) 3:58–9. doi: 10.1080/23802359.2017.1419099
28. Wang Y, Wu C, Xie Y, Liu H, Zhang H. Complete mitochondrial genome sequence for the Thrips hawaiiensis (Thysanoptera: Thripidae). Mitochondrial DNA Part B. (2021) 6:2092–4. doi: 10.1080/23802359.2021.1942268
29. Pakrashi A, Patidar A, Singha D, Kumar V, Tyagi K. Comparative analysis of the two suborders of Thysanoptera and characterization of the complete mitochondrial genome of Thrips parvispinus. Arch Insect Biochem Physiol. (2023) 2(114):1–15. doi: 10.1002/arch.22010
30. Li C, Gao Y, Wang D, Dang L. The extraordinary rearrangement of mitochondrial genome of the wheat pest, Aptinothrips stylifer and the mitochondrial phylogeny of Thripidae (Thysanoptera). Arch Insect Biochem Physiol. (2024) 115:e22086. doi: 10.1002/arch.22086
31. Wolstenholme DR. Genetic novelties in mitochondrial genomes of multicellular animals. Curr Opin Genet Dev. (1992) 2:918–25. doi: 10.1016/S0959-437X(05)80116-9
32. Cameron SL. Insect mitochondrial genomics: implications for evolution and phylogeny. Annu Rev entomology. (2014) 59:95–117. doi: 10.1146/annurev-ento-011613-162007
33. Krzywinski J, Grushko OG, Besansky NJ. Analysis of the complete mitochondrial DNA from Anopheles funestus: an improved dipteran mitochondrial genome annotation and a temporal dimension of mosquito evolution. Mol Phylogenet Evol. (2006) 39:417–23. doi: 10.1016/j.ympev.2006.01.006
34. Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. (2018) 34:i884–90. doi: 10.1093/bioinformatics/bty560
35. Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics. (2009) 25:1754–60. doi: 10.1093/bioinformatics/btp324
36. Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The sequence alignment/map format and SAMtools. Bioinformatics. (2009) 25:2078–9. doi: 10.1093/bioinformatics/btp352
37. Li D, Liu CM, Luo R, Sadakane K, Lam TW. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics. (2015) 31:1674–6. doi: 10.1093/bioinformatics/btv033
38. Meng G, Li Y, Yang C, Liu S. MitoZ: a toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. (2019) 47:e63–3. doi: 10.1093/nar/gkz173
39. Gurevich A, Saveliev V, Vyahhi N, Tesler G. QUAST: quality assessment tool for genome assemblies. Bioinformatics. (2013) 29:1072–5. doi: 10.1093/bioinformatics/btt086
40. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. (2014) 30:2068–9. doi: 10.1093/bioinformatics/btu153
41. Grant JR, Enns E, Marinier E, Mandal A, Herman EK, Chen CY, et al. Proksee: in-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. (2023) 51:W484–92. doi: 10.1093/nar/gkad326
42. Bernt M, Donath A, Jühling F, Externbrink F, Florentz C, Fritzsch G, et al. MITOS: improved de novo metazoan mitochondrial genome annotation. Mol Phylogenet Evol. (2013) 69:313–9. doi: 10.1016/j.ympev.2012.08.023
43. Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. (2021) 38:3022–7. doi: 10.1093/molbev/msab120
44. Perna NT, Kocher TD. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J Mol Evol. (1995) 41:353–8. doi: 10.1007/BF00186547
45. Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. (2013) 30:772–80. doi: 10.1093/molbev/mst010
46. Li KB. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics. (2003) 19:1585–6. doi: 10.1093/bioinformatics/btg192
47. Inoue T, Sakurai T. The phylogeny of thrips (Thysanoptera: Thripidae) based on partial sequences of cytochrome oxidase I, 28S ribosomal DNA and elongation factor-1 α and the association with vector competence of tospoviruses. Appl entomology zoology. (2007) 42:71–81. doi: 10.1303/aez.2007.71
48. Liu Q, He J, Song F, Tian L, Cai W, Li H. Positive correlation of the gene rearrangements and evolutionary rates in the mitochondrial genomes of thrips (Insecta: Thysanoptera). Insects. (2022) 13:585. doi: 10.3390/insects13070585
49. Kumar V, Tyagi K, Kundu S, Chakraborty R, Singha D, Chandra K. The first complete mitochondrial genome of marigold pest thrips, Neohydatothrips samayunkur (Sericothripinae) and comparative analysis. Sci Rep. (2019) 9:191. doi: 10.1038/s41598-018-37889-6
50. Soumia PS, Shirsat DV, Krishna R, Choudhary JS, Naaz N, P.a. and Singh M. Unfolding the mitochondrial genome structure of green semilooper (Chrysodeixis acuta Walker): An emerging pest of onion (Allium cepa L.). PloS One. (2022) 17:e0273635. doi: 10.1371/journal.pone.0273635
51. Karuppaiah V, Gadge AS, Shirsat DV, Soumia PS, Mainkar P, Kumar S, et al. The complete mitochondrial genome of the Indian dammer bee, Tetragonula iridipennis, and the phylogenomics of Meliponini. Front Ecol Evol. (2023) 11:1171242. doi: 10.3389/fevo.2023.1171242
52. Tyagi K, Chakraborty R, Cameron SL, Sweet AD, Chandra K, Kumar V. Rearrangement and evolution of mitochondrial genomes in Thysanoptera (Insecta). Sci Rep. (2020) 10:695. doi: 10.1038/s41598-020-57705-4
53. Xue XF, Deng W, Qu SX, Hong XY, Shao R. The mitochondrial genomes of sarcoptiform mites: are any transfer RNA genes really lost? BMC Genomics. (2018) 19:1–11. doi: 10.1186/s12864-018-4868-6
54. Beckenbach AT, Joy JB. Evolution of the mitochondrial genomes of gall midges (Diptera: Cecidomyiidae): rearrangement and severe truncation of tRNA genes. Genome Biol Evol. (2009) 1:278–87. doi: 10.1093/gbe/evp027
55. Masta SE, Boore JL. Parallel evolution of truncated transfer RNA genes in arachnid mitochondrial genomes. Mol Biol Evol. (2008) 25:949–59. doi: 10.1093/molbev/msn051
56. Margam VM, Coates BS, Hellmich RL, Agunbiade T, Seufferheld MJ, Sun W, et al. Mitochondrial genome sequence and expression profiling for the legume pod borer Maruca vitrata (Lepidoptera: Crambidae). PloS One. (2011) 6:e16444. doi: 10.1016/j.ijbiomac.2018.11.017
57. Gao S, Ren Y, Sun Y, Wu Z, Ruan J, He B, et al. PacBio full-length transcriptome profiling of insect mitochondrial gene expression. RNA Biol. (2016) 13:820–5. doi: 10.1080/15476286.2016.1197481
58. Ma Y, Zheng BY, Zhu JC, van Achterberg C, Tang P, Chen XX. The first two mitochondrial genomes of wood wasps (Hymenoptera: Symphyta): Novel gene rearrangements and higher-level phylogeny of the basal hymenopterans. Int J Biol macromolecules. (2019) 123:1189–96. doi: 10.1016/j.ijbiomac.2018.11.017
59. Mound LA. (2002). The Thrips and Frankliniella genus groups: the phylogenetic significance of ctenidia, in: Thrips and Tospoviruses: Proceedings of the 7th International Symposium on Thysanoptera, pp. 379–86. Australian National Insect Collection, Canberra, Australia.
60. Pandi A, Malaichamy K, Samuel KJ, Selvaraj D, Kasivelu G, Kolanthasamy E. Unveiling thrips morphology: A comparative analysis of microscopy-based ultrastructural, morphological, and molecular characterization of Thrips tabaci and Thrips parvispinus in onion. . Microscopy Res technique. (2024) 87:3003–15. doi: 10.1002/jemt.24653
61. Singh D, Kabiraj D, Sharma P, Chetia H, Mosahari PV, Neog K, et al. The mitochondrial genome of Muga silkworm (Antheraea assamensis) and its comparative analysis with other lepidopteran insects. PloS One. (2017) 12:e0188077. doi: 10.1371/journal.pone.0188077
62. Jiang SD, Wang L, Wang L, Sun J, Wang JJ, Wei DD. Mitochondrial coding genes mediate insecticide tolerance in the oriental fruit fly, Bactrocera dorsalis (Hendel). Pesticide Biochem Physiol. (2024) 199:105763. doi: 10.1016/j.pestbp.2023.105763
63. Guven C, Sevgiler Y, Taskin E. Pyrethroid insecticides as the mitochondrial dysfunction inducers. Mitochondrial Dis. (2018) 293:293–322. doi: 10.5772/intechopen.80283
64. Van Leeuwen T, Vontas J, Tsagkarakou A, Dermauw W, Tirry L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: a review. Insect Biochem Mol Biol. (2010) 40:563–72. doi: 10.1016/j.ibmb.2010.05.008
65. Fotoukkiaii SM, Mermans C, Wybouw N, Van Leeuwen T. Resistance risk assessment of the novel complex II inhibitor pyflubumide in the polyphagous pest Tetranychus urticae. J Pest Sci. (2020) 93:1085–96. doi: 10.1007/s10340-020-01213-x
Keywords: invasive pest, mitochondrial genome, phylogeny, Thrips parvispinus, Thrips tabaci
Citation: Soumia PS, Shirsat DV, Karuppaiah V, Divekar PA and Mahajan V (2025) Unravelling the complete mitochondrial genomes of Thrips tabaci Lindeman and Thrips parvispinus Karny (Thysanoptera: Thripidae) and their phylogenetic implications. Front. Insect Sci. 5:1536160. doi: 10.3389/finsc.2025.1536160
Received: 28 November 2024; Accepted: 10 February 2025;
Published: 28 February 2025.
Edited by:
Tiago Fernandes Carrijo, Universidade Federal do ABC, BrazilReviewed by:
Menglin Wang, University of California, Berkeley, United StatesCopyright © 2025 Soumia, Shirsat, Karuppaiah, Divekar and Mahajan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: P.S. Soumia, c291bWlhcHNAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.