
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Insect Sci., 14 June 2024
Sec. Invasive Insect Species
Volume 4 - 2024 | https://doi.org/10.3389/finsc.2024.1385884
 Alicia Fonseca-Muñoz1
Alicia Fonseca-Muñoz1 Gregorio Hernández-Salinas2
Gregorio Hernández-Salinas2 Salvador Ordaz-Silva3Imelda Virginia López-Sánchez3Jorge Luis Delgadillo-Ángeles3Evert Villanueva-Sánchez4Clemente Villanueva-Verduzco5Nadia S. Gómez-Domínguez6
Salvador Ordaz-Silva3Imelda Virginia López-Sánchez3Jorge Luis Delgadillo-Ángeles3Evert Villanueva-Sánchez4Clemente Villanueva-Verduzco5Nadia S. Gómez-Domínguez6 Carlos Granados-Echegoyen7*
Carlos Granados-Echegoyen7*Calliphoridae are widespread globally and can inhabit a variety of habitats. In this brief report, we assessed the appeal of Chrysomya rufifacies to resources that were previously inhabited by Lucilia sericata and L. cuprina, both common carrion colonizers. Two hundred adult male and female (1:1) C. rufifacies were released under controlled conditions into clear plastic cages (45 x 45 x 45 cm) with four types of substrates: beef-liver; liver with 100 third-instar larvae of L. cuprina; (III) liver containing 100 third-instar larvae of L. sericata; and liver containing 100 third-instar larvae of C. rufifacies. Each substrate was left in place for 24 hours at the end of a tube connected to the cage, where sticky traps were positioned to capture flies that might have been attracted to a specific substrate. The results indicate variations in the attraction of flies to different types of livers colonized by larvae of various species. It is suggested that flies may have specific preferences depending on the species of larvae present in the substrate. The liver without larvae was the preferred choice, while beef liver with C. rufifacies larvae was the least attractive. Results of statistical tests indicated that there is independence between attractiveness preference and the presence of C. rufifacies flies. Although there is a trend among certain levels of the variables in the correspondence analysis, these relationships are not statistically significant. However, they indicate specific patterns of association between different groups of flies and species of larvae. This study demonstrated that C. rufifacies does not show reduced attraction to any of the resources. A tetrahedron olfactometer device has been used for the first time in a behavioral study of C. rufifacies flies. This may enable future studies to enhance the understanding of fly behavior.
Several arthropods are attracted to carrion within minutes of death (1, 2). The attraction of these arthropods is influenced by various cues, including odor (1, 3). Many of the insects that colonize vertebrate carcasses are important for forensic investigations. Acceptance, i.e., oviposition, by these insects is an important part of the decomposition process (4). Colonization occurs in the natural cavities of the body, such as the nose, mouth, and ears. These are influenced by the nutritional value of resources (5), including feces (6), attractants such as chicken and beef livers (7), human blood, semen, and saliva (8), and others. Sometimes, predators are guided by cues indicating the presence of prey, which can alter their abundance and behavior (9). Some signals used by members of the same species and predators are excretions and secretions produced by blowfly larvae (10). Some adult blow flies (Diptera: Calliphoridae) do not rely on carrion for sustenance. Their competition is influenced by both density-dependent and density-independent factors, which occur during the larval stage (11).
The Calliphoridae family is distributed worldwide and is studied for its significance as forensic evidence associated with decomposing remains (1). The genera Calliphora, Cochliomyia, Chrysomya, and Lucilia are the most prevalent blowflies within the Calliphoridae family. In some regions of the world, Chrysomya rufifacies have been identified as the primary blowfly species that colonizes carrion (2, 12). The hairy maggot blowfly is found in tropical and subtropical regions worldwide, including Central and South America (13). The larvae of this species are predatory and cannibalistic during the second and third instars. These behaviors may provide competitive advantages over both larvae from different species and larvae from the same species (13). Such selection has influenced the behavior of other Chrysomya species competing for the same resources. Yang and Shiao (14) reported that female C. megacephala avoided laying eggs in the presence of C. rufifacies larvae; however, C. rufifacies showed a preference when C. megacephala larvae were present. Giao and Godoy (15) suggested that predation by other species may influence oviposition behavior in other calliphorids.
Other important calliphorids in forensic investigations include Lucilia sericata and L. cuprina. They are reported to be the first colonizers of corpses and are forensically important (1, 16). Sherman (17) and other studies have reported that they also cause myiasis. However, L. sericata larvae are also used for debriding necrotic ulcers in maggot therapy (18). Oviposition behavior varies among blowfly species. Limited research has been conducted on the relationship between other species and predators in their larval stage. There has been a growing interest in understanding the attraction of flies to a food source colonized by third-stage blowfly larvae. Forensic entomologists need to understand the behavioral ecology of adult blowflies (10). However, there is limited knowledge about attracting adults of C. rufifacies to third-stage larvae of L. sericata, C. rufifacies, and L. cuprina. This brief report aims to investigate the behavior of adult C. rufifacies towards a food source colonized by the larvae of these blowflies.
Blowflies were collected from a decomposing pig head in Santa María, Tule, Oaxaca, Mexico (17° 02’50” N, 96° 38’00” W). Maggots were collected and placed in 1000 mL plastic containers filled with sterile sawdust, which were then covered with white plastic sheets. The resulting adult flies were identified using the method described by Whitworth (19). They were then transferred to 30 x 30 x 30 cm plastic cages and kept under controlled conditions (26 ± 1°C, 70 ± 10% relative humidity, and a 12:12 photoperiod). Blowflies were fed 50 g of bovine liver, sugar, and water ad libitum for the first five days after emergence. Oviposition of the flies was induced by placing 50 grams of beef liver in gravid blowfly cages. The eggs were placed in 1-liter plastic jars filled with sawdust (20). After hatching, they were fed 300 grams of bovine liver. All flies were used from generations F2 to F4. The pupae were removed and transferred to 30 cm³ plastic cages to complete their development into adulthood.
Adult C. rufifacies flies were maintained in 30 cm³ plastic cages and provided with granulated sugar and water ad libitum. The flies were fed cow liver blood for the first 5 days after emerging. A total of 200 adult C. rufifacies flies, 7 days old, were aspirated from their cages using an aspirator (AC/DC Aspirator, BioQuip USA). The method used was modified from that described by Ma et al. (21)Tomberlin et al. (22); and Flint and Tomberlin (23). In the present study, we assembled and used a tetrahedron olfactometer device. We transferred these flies to a modified 45 x 45 x 45 cm clear plastic cage. The cage had four holes in the center of each wall. We placed a 4-inch diameter PVC tube with a perforated lid inside the cage, with each hole measuring 0.4 mm in diameter (Figure 1). Four types of substrates were studied: (I) liver; (II) liver containing 100 third instar larvae of L. cuprina; (III) liver containing 100 third instar larvae of L. sericata; (IV) liver containing 100 third instar larvae of C. rufifacies. To avoid positional bias, we allowed the cages to dry for 24-48 hours between experiments after thorough cleaning. Additionally, we rotated the tubes to different positions to ensure varied substrate exposure. We placed each substrate at the end of a tube connected to the cage and left it in position for 24 hours. Sticky traps (Trapper Max-Free, Bell Laboratories, Madison, WI, USA) were placed to facilitate the capture of flies exhibiting substrate preference. We recorded the number, sex, and gravid status of the captured flies.

Figure 1 A tetrahedron olfactometer device for quantification of study variables of C. rufifacies to various food substrates colonized by larvae of Lucilia sericata and Lucilia cuprina. (A) Zenithal perspective, (B) Frontal view.
A randomized experimental design was utilized. Five cages were set up as replications and the number of flies was counted twenty times in each cage for males, non-gravid females, and gravid females. The Kolmogorov-Smirnov test was used to assess the distribution of the data, while the chi-square test was used to assess the relationship between the variable’s “groups” and “species”. A simple correspondence analysis was conducted to assess the relationship between two variables or their levels and a multinomial logistic regression analysis was employed to model the behavior of the species to the variable’s “groups” and “species”. In all tests, a significance level of α = 0.05 was utilized with the JMP statistical software v7.0.
Variation in the attraction of flies to different types of livers colonized by larvae of different species is evident. This suggests that flies may have specific preferences depending on the species of larvae found in the substrate. Several factors could influence fly feeding preferences, including odor, texture, chemical composition, and the presence of associated microorganisms. We observed that Substrate I, the control treatment, appears to be the preferred choice of the flies, as it has the highest frequency and the highest attraction percentage. This could be because flies prefer the liver without larvae as a food source. In the case of beef liver with C. rufifacies larvae (Substrate IV), this treatment exhibited the lowest frequency and attraction percentage. It seems that flies are less attracted to beef liver colonized with C. rufifacies larvae compared to the other options. Similarly, beef liver with L. cuprina larvae (Substrate II) showed a slightly higher frequency and attraction than the C. rufifacies larvae treatment, but still lower than the control. The flies seem to exhibit a moderate preference for this type of liver colonized by L. cuprina larvae. For beef liver with L. sericata larvae (substrate III), similar to the treatment with C. rufifacies larvae, this treatment exhibits lower frequency and attractiveness compared to the control and the liver colonized by L. cuprina larvae. The data presented in the results provide an intriguing insight into the feeding preferences of flies regarding various types of beef livers colonized by larvae of three different species (Figure 2).
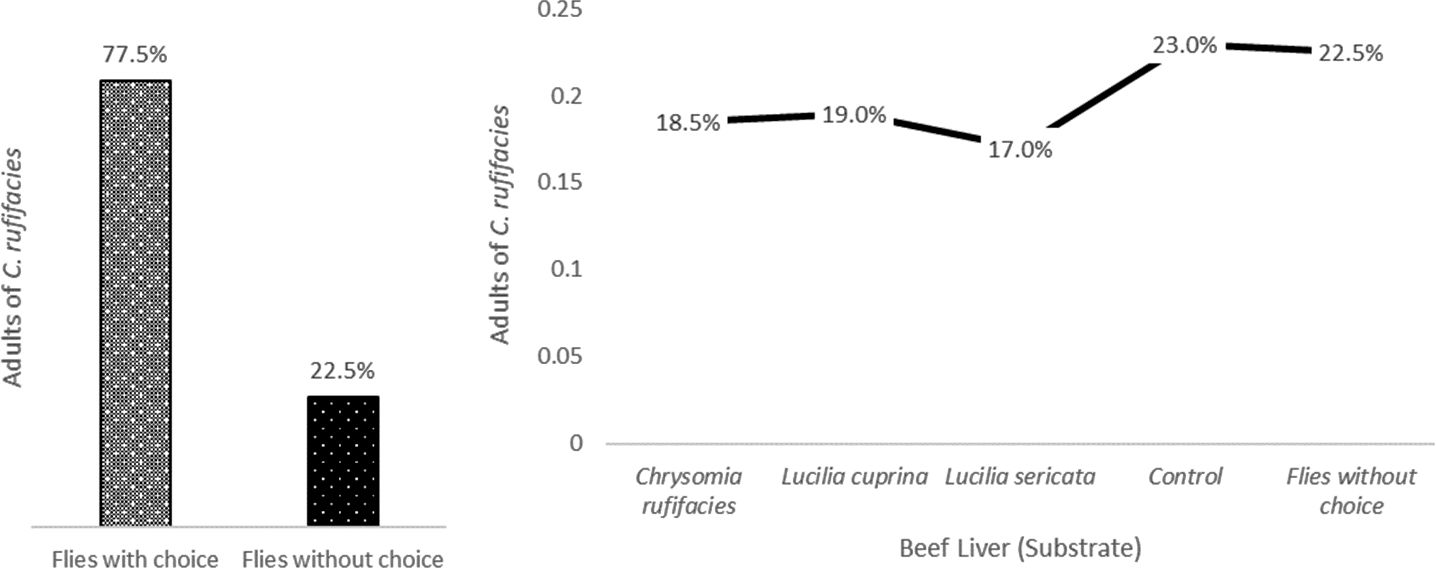
Figure 2 Attraction of Chrysomya rufifacies to Different Substrates Colonized by Lucilia sericata and Lucilia cuprina Larvae.
The results of the statistical test, based on the Chi-square test, indicate a p-value greater than 0.05. Therefore, variables are independent or have no dependence between variables. That is, one does not affect the other, and vice versa. The results of the statistical test indicate that the two variables, attraction (target choice) and C. rufifacies flies, are independent of each other (Table 1). In correspondence analysis, a specific trend is observed in the graph between certain levels of each variable to the other variable, which is indicated by ellipses. Although these relationships or dependencies are not statistically significant, it is observed that some groups of both variables are relatively close to each other, while others are further away from the center. For instance, the group of “gravid females” from one of the two variables is similar to the “L. sericata” group from the other variable. Therefore, it could be argued that while the statistical test results show no significant relationship or dependency between the two variables, a discernible tendency or pattern indicates that gravid females are commonly associated with the species L. sericata. In a similar vein, the “non-gravid females” exhibit a similar pattern, with two distinct groups or subgroups identified: one associated with the “beef liver (control)” and the other with L. cuprina. Therefore, the farther it is from the center, the greater its weight and potential influence on, or dependence on, other levels or groups of variables (Figure 3).
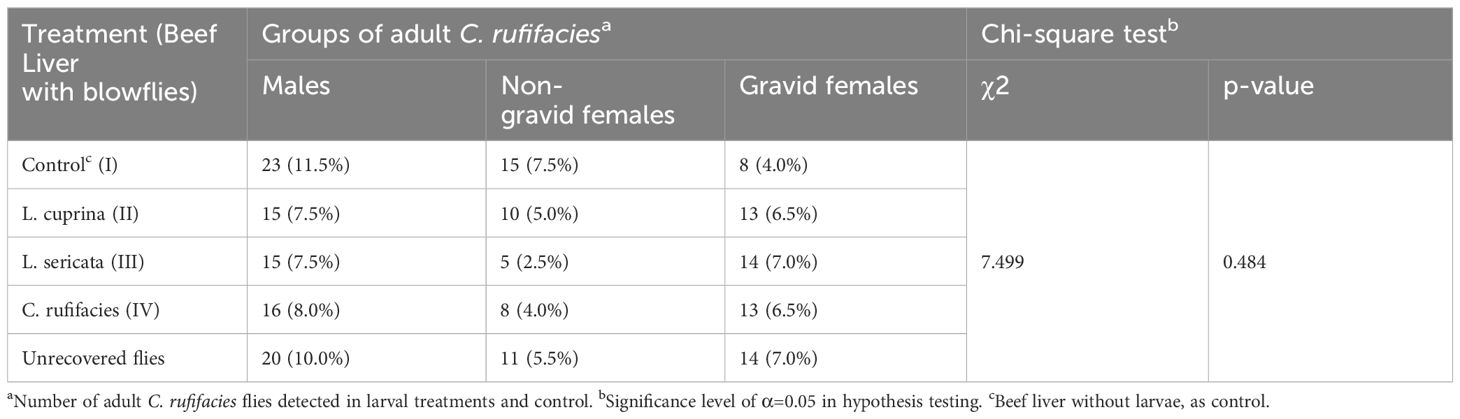
Table 1 Abundance (N) and relative abundance (%) of adult Chrysomya rufifacies, categorized by sex, attracted to resources colonized by Lucilia sericata and Lucilia cuprina larvae.
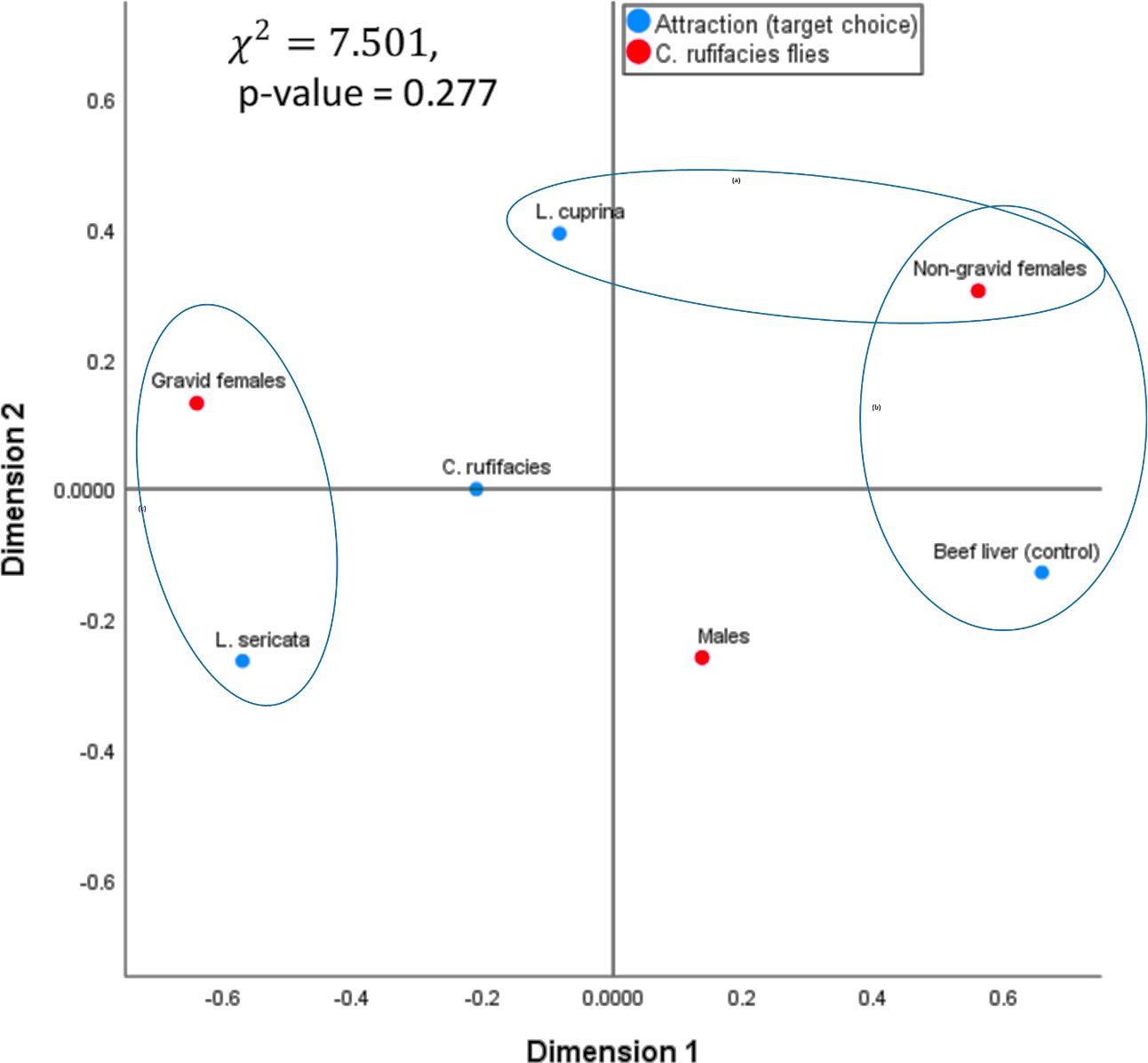
Figure 3 Correspondence analysis of C. rufifacies to various food substrates colonized by larvae of Lucilia sericata and Lucilia cuprina.
This study demonstrates that blowflies do not exhibit differences in attraction to food colonized by both species. However, this response was only observed in third-instar maggots and not in first or second-instars. Distance may also be a factor. Previous behavioral studies with C. rufifacies have shown that it preys on C. macellaria, possibly due to its weaker competitive abilities compared to C. rufifacies (24). However, there are other behavioral studies of adult C. rufifacies, especially concerning the presence of Calliphoridae species.
Wall and Fisher (25) discussed the utilization of their eyes by L. sericata for locating and seeking scents to lay eggs over short distances. However, some calliphorids responded to distances greater than 10 meters (26). In our study, we observed attraction at a short distance of about 45 centimeters, where adult C. rufifacies flies were attracted by the presence of third-instar larvae of blowfly species. There was no significant difference between females and males in terms of attractiveness to C. rufifacies for colonizing food resources. Gomes et al. (27) suggested that olfactory cues may stimulate blowflies to oviposit. However, visual cues are the key factors that influence flies’ decisions on where to oviposit.
Competition for resources can lead insects to develop various survival strategies, such as predation and cannibalism. The signals that indicate to a scavenger insect that the presence of a resource is beneficial to its offspring are not well understood in scavenging insects. We endorse the study conducted by Tomberlin et al. (28), which highlights the significance of C. rufifacies in forensic entomology due to its larval biology. The study reveals that during the initial larval stage, C. rufifacies feeds directly on carrion, while in the second and third stages, they act as facultative predators of larvae of other species. Therefore, our study aims to investigate the relationship between the presence of flies and the larvae of other species such as L. sericata and L. cuprina.
Yang and Shiao (14) reported that female C. megacephala avoids laying eggs in the presence of C. rufifacies larvae. However, C. rufifacies showed a preference for oviposition in the presence of C. megacephala larvae. The study did not demonstrate a preference for oviposition by C. rufifacies when in the presence of L. sericata and L. cuprina species compared to the control group. On the other hand, Shah and Sakhawat (29) demonstrated that calliphorids exhibit preferences for different colors and stages of meat. In our study, we utilized beef liver at 24 hours of decomposition, which elicited attraction at 26 ± 1°C, 70 ± 10% relative humidity, and a 12:12 photoperiod. Previous studies have shown that C. macellaria selectively oviposits in beef liver resources with C. rufifacies larvae more than with C. macellaria (10). However, in the present study, the oviposition rate of C. rufifacies was similar between L. cuprina and L. sericata.
Although the correlation coefficient indicates a low correlation between the groups and the species present, the simple correspondence analysis revealed a tendency for “gravid” flies to be associated with L. sericata larvae, while “males” were more commonly found in the “control” group. However, there was no significant tendency, dependence, or statistical association with the females of C. rufifacies or with the presence of L. cuprina larvae and C. rufifacies. In the study, 43% of males showed a stronger reaction in the liver without larvae of any species. This indicates that the most represented gravid females were attracted by larvae of L. sericata, with a 30.7% response rate. The presence of non-gravid females in all three treatments was consistent, ranging from 42% to 44%. However, the control group was smaller. The study revealed that adult C. rufifacies were more attracted to food that had been colonized by larvae of L. sericata, unlike their species. Studies conducted with other species (predator or prey) suggest that blowflies may exhibit escape or attraction behavior towards a previously colonized resource. This study demonstrated that C. rufifacies did not exhibit reduced attractiveness to such resources.
This brief communication is part of a project led by the primary author in collaboration with a multidisciplinary team of researchers in Mexico. The project aims to understand the biology and ecology of flies that are of forensic significance in the country. The purpose of this report is to conduct a more in-depth investigation into the chemical and physical characteristics of each type of substrate and their impact on the attraction of calliphorid flies. Our findings may offer insight into how these variables influence the preference of flies for a specific substrate.
Understanding the feeding preferences of flies is crucial for managing pests in agricultural, livestock, and urban settings. The results obtained could have practical implications for the development of more effective pest control strategies, such as the selection of baits or traps tailored to specific fly species. In addition, they provide a solid foundation for future research on the ecology and feeding behavior of flies, as well as the development of more effective pest control strategies, which will be detailed in future publications. This study demonstrated that C. rufifacies does not show reduced attraction to any of the resources.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
AF-M: Writing – review & editing, Writing – original draft, Visualization, Validation, Software, Resources, Project administration, Investigation, Conceptualization. GH-S: Writing – review & editing, Visualization, Validation, Supervision, Software. SO-S: Writing – review & editing, Visualization, Validation, Supervision, Software, Resources. IL-S: Writing – review & editing, Visualization, Validation, Supervision, Software. JD-Á: Writing – review & editing, Visualization, Validation, Supervision, Software. EV-S: Writing – review & editing, Visualization, Validation, Supervision, Software, Investigation. CV-V: Writing – review & editing, Visualization, Validation, Supervision, Software. NG-D: Formal analysis, Methodology, Software, Validation, Visualization, Writing – review & editing. CG-E: Writing – review & editing, Writing – original draft, Visualization, Resources, Project administration, Methodology, Investigation.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
We would like to express our gratitude to the Instituto Politécnico Nacional CIIDIR Oaxaca and the Laboratorio de Entomología Aplicada of the Universidad Autónoma de Campeche for their support during the preparation of this document. Finally, we would like to express our gratitude to all the researchers who contributed to this research project, as well as to the reviewers for their valuable suggestions on a previous version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Byrd J, Lord WD, Wallace JR, Tomberlin JK. Collection of entomological evidence during legal investigations. In: Byrd J, Castner J, editors. Forensic entomology: the utility of arthropods in legal investigations, 2nd ed. CRC Press, USA, Boca Raton (2009). p. 127–76. doi: 10.1201/NOE0849392153.ch3
2. Richards SL, Connelly CR, Day JF, Hope T, Ortiz R. Arthropods associated with carrion in a salt marsh habitat in southeastern florida. Fla. Entomol. (2015) 98:613–9. doi: 10.1653/024.098.0234
3. Forbes S, Carter DO. Processes and mechanisms of death and decomposition of vertebrate carrion. In: Benbow ME, Tomberlin JK, Tarone AM, editors. Carrion ecology, evolution, and their applications. CRC Press, Boca Raton, FL (2016). p. 13–30. doi: 10.1201/b18819-4
4. Ody H, Bulling MT, Barnes KM. Effects of environmental temperature on oviposition behavior in three blow fly species of forensic importance. Forensic Sci Int. (2017) 275:138–43. doi: 10.1016/j.forsciint.2017.03.001
5. Vanlaerhoven SL. Ecological theory and its application in forensic entomology. In: Byrd J, Castner J, editors. Forensic entomology: the utility of arthropods in legal investigations, 2nd ed. CRC Press, USA, Boca Raton (2009). p. 493–518.
6. Stofolano JG, Li M-F, Sutton JA, Yin C-M. Faeces feeding by adult Phormia regina (Diptera: Calliphoridae): impact on reproduction. Med Vet Entomol. (1995) 9:388–92. doi: 10.1111/j.1365-2915.1995.tb00010.x
7. Zhu JJ, Chaudhury MF, Tangtrakulwanich K, Skoda SR. Identification of oviposition attractants of the secondary screwworm, cochliomyia macellaria (F.) released from rotten chicken liver. J Chem Ecol. (2013) 39:1407–14. doi: 10.1007/s10886-013-0359-z
8. Durdle A, Mitchell RJ, van Oorschot RAH. The food preferences of the blow fly Lucilia cuprina Offered human blood, semen and saliva, and various nonhuman foods sources. J Forensic Sci. (2015) 61:99–103. doi: 10.1111/1556-4029.12912
9. Peckarsky BL, Abrams PA, Bolnick DI, Dill LM, Grabowski JH, Luttbeg B, et al. Revisiting the classics: considering nonconsumptive effects in textbook examples of predator–prey interactions. Ecol. (2008) 89:2416–25. doi: 10.1890/07-1131.1
10. Flores M, Crippen TL, Longnecker M, Tomberlin JK. Nonconsumptive Effects of Predatory Chrysomya rufifacies (Diptera: Calliphoridae) Larval Cues on Larval Cochliomyia macellaria (Diptera: Calliphoridae) Growth and Development. J Med Entomol. (2017) 54:1167–74. doi: 10.1093/jme/tjx104
11. Abrams PA. When does greater mortality increase population size? The long history and diverse mechanisms underlying the hydra effect. Ecol Lett. (2009) 12:462–74. doi: 10.1111/j.1461-0248.2009.01282.x
12. García-Espinosa F, Valdés M, Sánchez F, Yusseff S, Quintero M. Desarrollo larval y requerimiento calóricos de Chrysomya rufifacies (Díptera: Calliphoridae) durante primavera y verano en Torreón, Coahuila. Acta Zool Mex. (2012) 28:172–84.
13. Baumgartner DL. Review of chrysomya rufifacies (Diptera: calliphoridae). J Med Entomol. (1993) 30:338–52. doi: 10.1093/jmedent/30.2.338
14. Yang ST, Shiao S. Oviposition preferences of two forensically important blow fly species, Chrysomya megacephala and C. rufifacies (Diptera: Calliphoridae), and implications for postmortem interval estimation. J Med Entomol. (2012) 49:424–35. doi: 10.1603/ME11133
15. Giao J, Godoy W. Ovipositional behavior in predator and prey blowflies. J Insect Behav. (2007) 20:77–86. doi: 10.1007/s10905-006-9064-x
16. El-Bassiony GM. Blowflies (Diptera: Calliphoridae) as forensic indicators in Egypt with special reference to the development data of Lucilia Cuprina (Wiedemann). Sumerianz J Biotechnol. (2020) 3:43–51.
17. Sherman RA. Wound myiasis in urban and suburban United States. Arch Intern Med. (2000) 160:2004–14. doi: 10.1001/archinte.160.13.2004
18. Bazalinski D, Kózka M, Karnas M, Więch P. Effectiveness of chronic wound debridement with the use of larvae of Lucilia sericata. J Clin Med. (2019) 8:1845. doi: 10.3390/jcm8111845
19. Whitworth T. Keys to the genera and species of blow flies (Diptera: Calliphoridae) of America north of Mexico. Proc Entomol Soc Wash. (2006) 108:689–72. doi: 10.11646/zootaxa.2663.1.1
20. Lima T, Von Zuben CJ. Chrysomya megacephala (Fabricius)(Diptera: Calliphoridae) oviposition behavior in previous oviposition situation. Neotrop. Entomol. (2016) 45:612–7. doi: 10.1007/s13744-016-0412-z
21. Ma Q, Fonseca A, Liu W, Fields AT, Pimsler ML, Spindola AF, et al. Proteus mirabilis interkingdom swarming signals attract blow flies. ISME J. (2012) 6:1356–66. doi: 10.1038/ismej.2011.210
22. Tomberlin JK, Crippen TL, Tarone AM, Singh B, Adams K, Rezenom YH, et al. Interkingdom responses of flies to bacteria mediated by fly physiology and bacterial quorum sensing. Anim. Behav. (2012) 84:1449–56. doi: 10.1016/j.anbehav.2012.09.013
23. Flint CA, Tomberlin JK. Optimizing a dual-choice cube olfactometer design for measuring adult secondary screwworm (Diptera: calliphoridae) attraction to a resource. J Med Entomol. (2021) 58:994–1003. doi: 10.1093/jme/tjaa254
24. Brundage A, Bros S, Honda J. Seasonal and habitat abundance and distribution of some forensically important blow flies (Diptera: Calliphoridae) in Central California. Forensic Sci Int. (2011) 212:115–20. doi: 10.1016/j.forsciint.2011.05.023
25. Wall R, Fisher P. Visual and olfactory cue interaction in resource-location by the blowfly, Lucilia sericata. Physiol Entomol. (2001) 26:212–8. doi: 10.1046/j.0307-6962.2001.00237.x
26. Eisemann CH. Upwind flight by gravid Australian sheep blow flies Lucilia cuprina (Wiedemann) (Diptera: Calliphoridae) in response to stimuli from sheep. Bull Entomol Res. (1988) 78:273–9. doi: 10.1017/S0007485300013031
27. Gomes L, Gomes G, Casarin FE, da Silva IM, Sanches MR, von Zuben CJ, et al. Visual and olfactory factors interaction in resource-location by the blow fly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae), in natural conditions. Neotrop. Entomol. (2007) 36:633–9. doi: 10.1590/s1519-566x2007000500001
28. Tomberlin J, Albert A, Byrd J, Hall D. Interdisciplinary workshop yields new entomological data for forensic sciences: Chrysomya rufifacies (Diptera: Calliphoridae) established in North Carolina. J Med Entomol. (2006) 43:1287–8. doi: 10.1093/jmedent/43.6.1287
Keywords: non-attraction, Chrysomya, food resource, ecology behavior, Calliphoridae
Citation: Fonseca-Muñoz A, Hernández-Salinas G, Ordaz-Silva S, López-Sánchez IV, Delgadillo-Ángeles JL, Villanueva-Sánchez E, Villanueva-Verduzco C, Gómez-Domínguez NS and Granados-Echegoyen C (2024) Behavioral report of Chrysomya rufifacies in response to substrate infestation by Lucilia sericata and Lucilia cuprina using a tetrahedron olfactometer device. Front. Insect Sci. 4:1385884. doi: 10.3389/finsc.2024.1385884
Received: 14 February 2024; Accepted: 22 May 2024;
Published: 14 June 2024.
Edited by:
Osariyekemwen Uyi, University of Benin, NigeriaReviewed by:
Steve B. S. Baleba, International Centre of Insect Physiology and Ecology (ICIPE), KenyaCopyright © 2024 Fonseca-Muñoz, Hernández-Salinas, Ordaz-Silva, López-Sánchez, Delgadillo-Ángeles, Villanueva-Sánchez, Villanueva-Verduzco, Gómez-Domínguez and Granados-Echegoyen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlos Granados-Echegoyen, Z3JhbmFkb3MuZWNoZWdveWVuQHlhaG9vLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.