 Hong Jin
Hong Jin Xinyue Meng
Xinyue Meng Jianwei Feng
Jianwei Feng
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol., 28 February 2025
Sec. Cancer Immunity and Immunotherapy
Volume 16 - 2025 | https://doi.org/10.3389/fimmu.2025.1560393
This article is part of the Research TopicCommunity Series in Immunological Precision Therapeutics: Integrating Multi-Omics Technologies and Comprehensive Approaches for Personalized Immune Intervention: Volume IIView all 7 articles
Breast cancer (BC) is the most common cancer in women and a leading cause of cancer-related mortality. Despite advances in screening and treatment, outcomes for advanced or recurrent BC remain poor, highlighting the need for new strategies. Recent research emphasizes the tumor microenvironment (TME), particularly tumor-associated macrophages (TAMs), as key drivers of tumor growth, metastasis, and resistance to therapy. The presence of M2-like TAMs in the TME promotes immune evasion and tumor progression across BC subtypes. This review summarizes TAMs classification, their role in BC, and emerging therapies targeting TAMs, including depletion, inhibition of recruitment, and reprogramming from pro-tumoral M2 to anti-tumoral M1 phenotypes. Targeting TAMs offers a promising strategy to improve BC treatment outcomes.
Breast cancer (BC) stands as the most frequently diagnosed malignant neoplasm among women and remains the foremost contributor to cancer‐related fatalities in the female population (1). Despite continuous refinements in screening approaches and therapeutic regimens, the prognosis for individuals with advanced or treatment‐resistant recurrent BC has fallen short of expectations. Developing more effective interventions aimed at decreasing relapse and improving clinical outcomes thus remains a critical imperative.
Recent investigations underscore the strong relationship between tumor and the tumor microenvironment (TME) (2–5). The TME denotes a sophisticated niche encompassing cells, signaling molecules, and the local physicochemical context, all of which modulate tumor cell proliferation (6), dissemination, progression (7–9), and responses to treatment (10–13).Within this specialized milieu, a variety of components—such as blood vessels, fibroblasts, neurons, inflammatory mediators, and immune populations including T cells, B cells, neutrophils, mast cells, and tumor‐associated macrophages (TAMs)—coexist around malignant cells (14, 15). Numerous stromal factors and cellular elements in the TME contribute to BC pathogenesis. Among these, TAMs represent a pivotal fraction of the microenvironment (16). By interacting with diverse cell populations, TAMs facilitate immune evasion and promote tumor progression (17, 18), ultimately influencing the clinical outlook for BC patients (19). They exert profound impacts on tumor growth, invasiveness, and therapy response (20). This review primarily discusses the roles and mechanisms of TAMs in the initiation and advancement of various BC subtypes, and further discusses emerging strategies that target these macrophages.
Most TAMs arise from bone marrow–derived monocytes, although a fraction are tissue-resident macrophages that infiltrate tumors from adjacent tissues. Once macrophages exit the circulation, their notable plasticity allows them to adopt distinct functions under various local microenvironmental stimuli. Commonly, they are categorized into two major phenotypes: classically activated M1 and alternatively activated M2 (21–23). The M1 subset is driven by Toll-like receptor (TLR) signaling, helper T cell 1 (Th1) cytokines, and interferon (IFN)-γ, which facilitates antigen presentation, amplifies cytotoxic functions, and participates in inflammatory responses as well as tumor immunosuppression. On the other hand, M2 macrophages arise in response to cytokines such as interleukin (IL)-4, IL-10, IL-13, and transforming growth factor-β (TGF-β), thereby restraining antitumor immunity, promoting tumor-associated vasculature formation, and advancing disease progression (24, 25).
At each phase of tumor progression, M1 and M2 macrophages coexist in varying ratios. During the initial stage, macrophages commonly favor the M1 phenotype to foster antitumor effects. However, as malignancies advance, M2 macrophages become dominant, forming the principal TAM subset in the TME (26). Moreover, M2 macrophages can be categorized into M2a, M2b, M2c, and M2d subtypes. Inflammatory factors are pivotal in inflammatory diseases progression and significantly influence the efficacy of therapies (27–30). For instance, Helper T cell 2 (Th2) cytokines (e.g., IL-4 and IL-13) prompt macrophages to acquire the M2a state, while immune complex signaling and TLR activation generate M2b cells, and IL-10 drives macrophages toward the M2c subtype (31). Macrophage polarization is tied to specialized metabolic pathways involving glucose, lipids, and glutamine. Evidence indicates that heightened glucose metabolism in TAMs triggers the buildup of various tumor-related metabolites, which facilitate the shift from the M1 to the M2 phenotype by modulating gene expression and signaling pathways, thereby granting TAMs a robust tumor-supportive capacity (32). These findings highlight that steering macrophages toward the antitumor M1 phenotype and limiting their transition into the tumor-promoting M2 state may offer an effective avenue for cancer treatment. M2 subtypes involve in BC progression. For instance,M2a macrophages differentiated in vitro through IL-4/IL-13 stimulation enhance the migratory and invasive capabilities of BC cells more prominently compared to M2b or M2c subtypes (33). M2b macrophages, activated through immune complex signaling and TLR pathways, contribute to immune modulation and inflammation, which can enhance metastasis and resistance in HER2-positive BC (34). M2c macrophages, driven by IL-10, are involved in immune suppression and tissue remodeling, facilitating the immune evasion of tumor cells in TNBC and enhancing the metastatic potential of cancer cells (34). M2d macrophages, characterized by their role in promoting angiogenesis and immune tolerance, support tumor growth and are particularly relevant in TNBC and HER2-overexpressing BC (34, 35).
Of the various BC subtypes, the luminal category—characterized by estrogen receptor (ER) and/or progesterone receptor (PR) positivity—predominates. These luminal cells generally respond well to endocrine therapy yet show diminished sensitivity to chemotherapy. M2-oriented TAMs produce CCI2, which recruits monocytes into the TME and activates the phosphatidylinositol 3 kinase/protein kinase B/mammalian target of rapamycin (PI3K/AKT/mTOR) pathway, thereby driving endocrine resistance and establishing an endocrine-resistant niche. Additionally, individuals with ER-positive tumors and elevated CCI2 levels experience notably shorter progression-free survival compared with those exhibiting lower CCI2 expression (36). In THP-1 cells, diminished CXCR4 expression profoundly suppresses the oncogenic MAPK pathway and curtails the migratory capacity of MCF-7 cells (37). Wang et al. (38) demonstrated that, following co-culture of TAMs with MCF-7 cells, secretion of bone morphogenetic protein 2 (BMP-2) by TAMs increased sharply, leading to pronounced microcalcification within BC cells and ultimately contributing to unfavorable patient outcomes.
An estimated 15%–20% of patients with breast carcinoma exhibit HER2 amplification, which predisposes them to higher rates of brain metastasis than those lacking HER2 overexpression, thereby creating a significant therapeutic challenge for this subgroup. In cancers displaying elevated HER2, TAMs can modulate the response to trastuzumab and consequently influence prognosis (39). In a humanized mouse model with human breast tumors overexpressing HER2, TAMs have been shown to regulate the antitumor actions of pertuzumab and trastuzumab via antibody-dependent phagocytosis (40). Among the cytokines secreted by tumor-associated macrophages (TAMs), interleukin-8 (IL-8) plays a pivotal role. In locally advanced HER2-positive breast cancer, IL-8 undermines the efficacy of lapatinib by activating the SRC/STAT3/ERK1/2 signaling cascade, which in turn stimulates the EGFR pathway. This interaction contributes to adverse clinical outcomes and the development of therapeutic resistance (41). Furthermore, matrix metalloproteinase 11 (MMP11) derived from macrophages promotes the migration of HER2+ tumor cells through the CCL2–CCR2 axis. Its pro-oncogenic influence may arise from inducing immunosuppression via upregulation of PD-L1 expression in BC cells (42).
Triple-negative breast cancer (TNBC) exhibits poor pathological differentiation, marked invasiveness, and frequent recurrence and metastasis. Moreover, it remains largely unresponsive to endocrine and targeted therapies, resulting in a more adverse prognosis than that of other BC subtypes (43). Within TNBC lesions, TAMs are abundantly recruited and correlate with unfavorable clinical outcomes (44). Hypoxia in the tumor microenvironment is crucial for driving metastatic progression. In in vitro experiments, when oxygen availability is limited, TAMs secrete the pro-tumor metastasis factor CCL22, thereby enhancing the metastatic potential of MDA-MB-231 cells (45). Furthermore, Zhang et al. (46) revealed that in M2-type TAMs, the Yes-associated protein 1 (YAP1) modulates the monocyte chemoattractant protein 1 (MCP-1)/CCR2 pathway, leading to increased metastasis of MDA-MB-231 and BT549 cells.
TAMs encourage neoplastic cell proliferation by secreting bioactive molecules (47). Through lipocalin 2 (Lcn2) secretion, TAMs augment TNBC cell proliferation and motility. Additionally, Lcn2 antagonists impede the expansion and migration of TNBC cells (48). Among the central chemokines released by TAMs is C-X-C motif chemokine ligand 8 (CXCL8). While it may not substantially alter the proliferative ability of mammary carcinoma cells, CXCL8 heightens the invasiveness and miration of both murine and human breast tumors, triggers epithelial–mesenchymal transition (EMT), and supports the self-renewal capacity of BC stem cells (BCSCs) (49). In addition, IL-10 produced by M2-subtype TAMs enhances the Janus kinase 2 (JAK2)/STAT3 cascade, which fosters proliferation, invasion, and migration of MDA-MB-231 cells while curtailing apoptotic processes (50). Furthermore, TAM-produced VEGF-C supports vascular normalization in breast malignancies, reducing pulmonary dissemination of tumor cells yet elevating lymph node metastases (51).
Cytokines released by TAMs, exemplified by interleukin-6, promote the expansion of BCSCs through the STAT3 pathway. This cascade also upregulates essential transcription factors, such as sex-determining region Y box protein 2 (SOX2), octamer-binding transcription factor 3/4 (OCT3/4), and the Nanog homeobox (NANOG), thereby affecting the proliferation, metastatic competence, and neovascularization of 4T1 cells (52). Meanwhile, M2-skewed macrophages secrete vascular endothelial growth factor, triggering elevated expression of prostate cancer-associated transcript 6 (PCAT6), which in turn engages the VEGFR/AKT/mTOR axis, facilitating angiogenesis in TNBC (53). Cheng et al. (54) identified substantial B7-H3 expression within TAMs in patients with triple-negative breast tumors, noting that B7-H3-overexpressing TAMs display potent pro-angiogenic effects, impede T-cell infiltration, and promote metastatic dissemination. IL-17A promotes OPN secretion by tumor cells, activating LYVE-1/JNK/c-Jun signaling to expand immunosuppressive LYVE-1+ macrophages (55).
Exosomes secreted by TAMs also contribute to malignant cell dissemination. In 4T1 cells, uptake of these vesicles elevates miR-223-3p, which drives pulmonary metastasis by targeting chromobox homolog 5 (CBX5) (56). Macrophage-derived exosomes promote BC cell proliferation, invasiveness, and metastatic potential both in vitro and in vivo by activating the STAT3 signaling pathway and regulating key target genes, including CyclinD1, MMP2, and MMP9 (57, 58). Moreover, M2 macrophage-derived exosomes can deliver miR-92b into MDA-MB-231 cells, where this microRNA promotes autophagy in TNBC cells by suppressing phosphatase and tensin homolog (PTEN) (59). Collectively, these findings indicate that TAMs critically affect TNBC progression via the release of cytokines, chemokines, and exosomes. TGF-β1 originating from TAMs modulates hepatic leukemia factor (HLF), recognized as a novel oncogenic mediator in TNBC. Consequently, this factor enhances ferroptosis resistance by inducing γ-glutamyl transferase 1 (γ-GGT1), thereby supporting tumor cell proliferation and elevating drug resistance (18).
In BRCA-associated TNBC, therapy with olaparib stimulates macrophage maturation and bolsters antitumor mechanisms, although its effect proves short-lived. Subsequent immunosuppressive cascades emerge, marked by upregulated PD-L1 and colony stimulating factor 1 (CSF1) receptor, alongside a strong dependence on lipid metabolism. TAMs relying on lipid-based pathways demonstrate immunosuppressive properties, thereby constraining the efficacy of PARP inhibitors and contributing to resistance (60).
Cross talk between TAMs and other immune cells contribute to immune evasion and progression in BC. Interaction between IL-15Rα+ TAMs and BC cells reduces CD8+ T cell recruitment in TME (61). Besides, TAMs inhibit the activity of CD8+ T cells by promoting collagen accumulation and altering the metabolic environment within BC (62). TAMs that express the transcription factor IRF8 play a significant role in inducing T cell exhaustion in the TME of BC (63). The density of tumor-associated neutrophils (TANs) was correlated with the presence of CD163+ M2-like TAMs. M1/M2 TAM ratio was elevated, TANs served as a negative prognostic indicator in BC (64). (Figure 1).
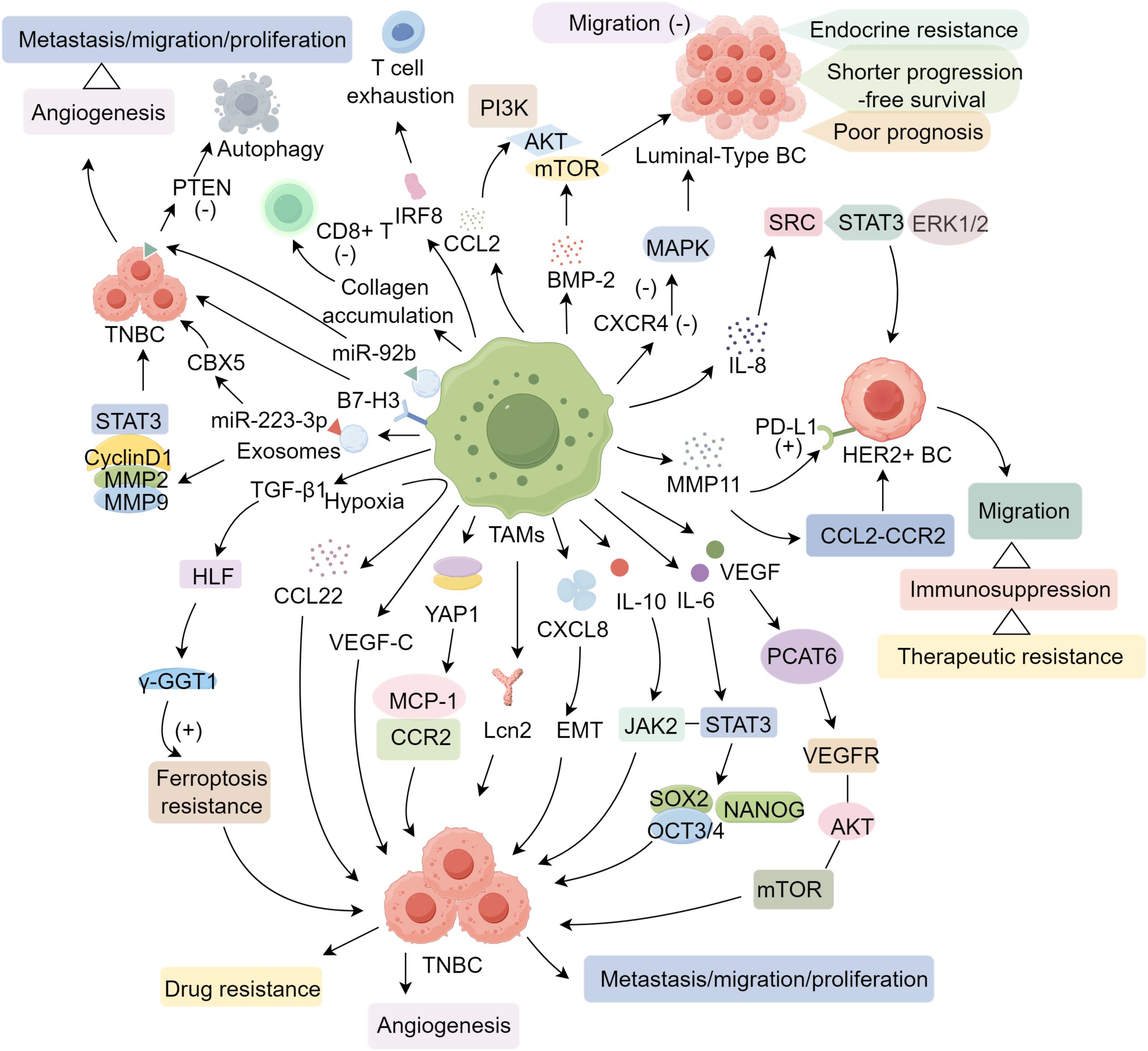
Figure 1. The mechanisms of TAMs in BC.
TAMs are pivotal in facilitating tumor cell growth, invasion, metastasis, and the formation of new blood vessels. Targeting and eliminating TAMs along with their precursor cells represents a significant strategy in anti-cancer therapies. Bisphosphonates (BP) are extensively utilized for managing patients with bone metastases. Evidence from clinical trials indicates that BP medications enhance the survival outcomes of BC patients. Additionally, both experimental models and clinical studies have shown that TAMs uptake BP, leading to their apoptosis or impaired functionality, thereby inhibiting tumor progression (65, 66). Chemotherapeutic drugs like docetaxel also suppress TAMs to manage tumor advancement. In a mouse model of orthotopic BC using the 4T1 cell line, the combined intraperitoneal administration of zoledronic acid (ZA) and docetaxel resulted in a significantly greater reduction in tumor growth and lung metastasis compared to ZA alone (67). Furthermore, doxycycline effectively depletes CD68+ TAMs. In the PyMT-Maclow BC mouse model, treatment with doxycycline decreased TAMs surrounding the tumor margins by 43%, thereby slowing tumor growth, reducing angiogenesis, and lowering the expression levels of genes that promote blood vessel formation (68). Recently, electroacupuncture treatment was shown to restore normal vascular function in TNBC through the reduction of glyoxalase-1 levels, which facilitated the polarization of TAMs toward the M1 phenotype (69).
Chemokines, cytokines, and complement components drive TAM recruitment in BC. CCL2 binding to CCR2 on monocytes facilitates TAM infiltration, making the CCL2/CCR2 pathway a key therapeutic target. Chitosan-DNA nanoparticles/siRNA targeting CCR2 (CNP/siCCR2) nanoparticles inhibit TAM recruitment, reduce tumor immunosuppression, and enhance chemotherapy efficacy in BC models (70). Elevated CCL2 and CCL5 in ER+ BC lead to macrophage infiltration and angiogenesis, which are reduced by anti-CCL2/CCL5 therapies (71).
CSF1 is crucial for macrophage proliferation and differentiation, with high levels linked to poor BC prognosis (72). Targeting CSF1 expression effectively diminishes TAM infiltration and enhances the chemosensitivity of BC. Pignatelli et al. (73) demonstrated that antibodies directed against the CSF1R impede the trans-endothelial migration of BC cells, proposing CSF1 as a potential therapeutic target for metastasis across multiple bBC subtypes. In murine BC models, the inhibition of CSF1/CSF1R signaling through either CSF1 monoclonal antibodies or the CSF1R tyrosine kinase inhibitor PLX3397 results in the depletion of TAMs and significantly postpones tumor recurrence following radiotherapy (74).
Angiogenesis is fundamental to both the initiation and progression of BC, with Vascular Endothelial Growth Factor (VEGF) being the principal angiogenic factor secreted by TAMs. Consequently, the VEGF/VEGFR2 axis has become a focal point of research; however, challenges such as therapeutic resistance, tumor evasion, recurrence, and potential harm to normal tissues present significant obstacles. Song et al. (75) reported that the administration of specific small interfering RNAs targeting VEGF and Placenta Growth Factor (PIGF) via nanoparticles to M2-polarized TAMs and BC cells as an immunotherapeutic approach markedly inhibits BC growth and pulmonary metastasis.
M1 macrophages exhibit tumor-suppressive properties, whereas TAMs, which share characteristics with M2 macrophages, promote tumor progression. Current research is focused on reprogramming macrophages toward anti-tumor phenotypes. A study by Guerriero et al. demonstrated that the class IIa histone deacetylase inhibitor (HDACi) TMP195 effectively reduced tumor-promoting macrophages while increasing anti-tumor macrophages in murine models. This shift led to a reduction in tumor growth and lung metastasis. Furthermore, TMP195, when combined with chemotherapy or immune checkpoint blockade, significantly enhanced the anti-tumor response, suggesting its potential as an adjuvant in cancer therapy (76). Emerging small-molecule HDAC inhibitors, such as entinostat, when used in combination with immune checkpoint inhibitors (ICIs), have been found to facilitate the transition of M2-type TAMs to the M1 phenotype, concurrently suppressing NF-κB and STAT3 signaling pathways, thereby curbing tumor progression (77). The chemotherapeutic agent cabazitaxel has the capacity to restore TAM-mediated programmed cell removal (prCR) in BC by promoting macrophage polarization toward the M1 type, activating the TLR/NF-κB axis, and enhancing the secretion of pro-inflammatory cytokines (78).
TLRs, essential pathogen recognition receptors on immune cells, play a pivotal role in immune modulation. In a murine model, the administration of a TLR3 ligand facilitated the transformation of M2-like TAMs back to the M1 phenotype through the IFN-α/β signaling pathway, thereby suppressing tumor growth (79). Lei et al. (80) found that the FAPα enzyme-activated vinca alkaloid prodrug ZGP-DAVLBH, a vascular disrupting agent, enhanced the secretion of granulocyte-macrophage colony-stimulating factor (GM-CSF) in both tumor tissues and the serum of tumor-bearing mice. This led to a phenotypic shift of TAMs from M2 to M1 and induced apoptosis in the doxorubicin-resistant BC cell line MCF-7/ADR. Additionally, Tasquinimod, an oral quinoline-3-carboxamide, was shown to convert immunosuppressive M2-type TAMs into pro-inflammatory M1-type TAMs in a 4T1 orthotopic BC model, resulting in inhibited tumor growth and reduced pulmonary metastasis (81).
Nanotechnology offers precise drug delivery to specific cellular targets. Targeting TAMs through nanocarriers can induce their polarization to the M1 phenotype, thereby modulating their phagocytic activity and ultimately regulating BC growth, metastasis, and invasion. Examples include hybrid nanoparticles encapsulating paclitaxel and baicalin (HPoNPC) (82), macrophage membrane-coated nanoparticles loaded with the TGF-β R1 inhibitor SD208 (Mφ-SDNP) (83), and organic polymer nanoparticles (CPTH) (84). These nanostructures effectively promote the shift of TAMs from M2 to M1, specifically target BC cells, and exhibit potent anti-tumor effects. Extracellular vesicles from Platycodon grandiflorum (PGEVs) promote TAM repolarization to M1, inhibiting TNBC progression (85). Recent strategy to enhance tumor oxygenation, converts immunosuppressive M2 to antitumor M1 macrophages offering promising in overcoming BC drug resistance (86). Matairesinol (MAT), a plant lignan, exhibits anticancer and immunomodulatory effects. It reduces M2a and M2d macrophage viability, promoting their shift to the M1 phenotype. Conditioned medium from MAT-treated macrophages induces apoptosis in TNBC cells, highlighting MAT’s potential as a novel anti-TNBC therapeutic (87).
TAMs are integral components of the BC TME, exerting profound influences on tumor progression, metastasis, and resistance to therapy. The dualistic nature of TAMs, characterized by their ability to adopt either pro-inflammatory M1 or anti-inflammatory M2 phenotypes, underscores their pivotal role in shaping the immune landscape of BC. The dominance of M2-like TAMs in various BC subtypes, including luminal, HER2-positive, and triple-negative forms, is closely associated with unfavorable clinical outcomes and reduced therapeutic efficacy. Therapeutic strategies targeting TAMs—ranging from their depletion and inhibition of recruitment to the reprogramming of their phenotypic states—offer multifaceted approaches to disrupt the pro-tumoral activities mediated by these cells. Advances in pharmacological agents, molecular inhibitors, and nanotechnology-based delivery systems have demonstrated potential in shifting TAMs toward a tumor-suppressive M1 phenotype, thereby enhancing the effectiveness of existing treatments and mitigating metastatic spread. Continued exploration of the intricate interactions between TAMs and BC cells is essential for the development of more effective, TAM-targeted therapies. Ultimately, integrating TAM modulation into comprehensive BC treatment regimens holds significant promise for improving patient prognoses and overcoming current therapeutic limitations.
HJ: Writing – original draft. XM: Writing – original draft, Writing – review & editing. JF: Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Brentnall AR, Cuzick J, Buist DSM, Bowles EJA. Long-term accuracy of breast cancer risk assessment combining classic risk factors and breast density. JAMA Oncol. (2018) 4:e180174. doi: 10.1001/jamaoncol.2018.0174
2. Xia Z, Chen S, He M, Li B, Deng Y, Yi L, et al. Editorial: Targeting metabolism to activate T cells and enhance the efficacy of checkpoint blockade immunotherapy in solid tumors. Front Immunol. (2023) 14:1247178. doi: 10.3389/fimmu.2023.1247178
3. Zhang X, Zhang P, Cong A, Feng Y, Chi H, Xia Z, et al. Unraveling molecular networks in thymic epithelial tumors: deciphering the unique signatures. Front Immunol. (2023) 14:1264325. doi: 10.3389/fimmu.2023.1264325
4. Deng Y, Shi M, Yi L, Naveed Khan M, Xia Z, Li X. Eliminating a barrier: Aiming at VISTA, reversing MDSC-mediated T cell suppression in the tumor microenvironment. Heliyon. (2024) 10:e37060. doi: 10.1016/j.heliyon.2024.e37060
5. Xie H, Xi X, Lei T, Liu H, Xia Z. CD8(+) T cell exhaustion in the tumor microenvironment of breast cancer. Front Immunol. (2024) 15:1507283. doi: 10.3389/fimmu.2024.1507283
6. Wang JF, Wang JS, Liu Y, Ji B, Ding BC, Wang YX, et al. Knockdown of integrin beta1 inhibits proliferation and promotes apoptosis in bladder cancer cells. Biofactors. (2025) 51:e2150. doi: 10.1002/biof.2150
7. Wang Y, Zhu H, Wang X. Prognosis and immune infiltration analysis of endoplasmic reticulum stress-related genes in bladder urothelial carcinoma. Front Genet. (2022) 13:965100. doi: 10.3389/fgene.2022.965100
8. Yang H, Li Z, Zhu S, Wang W, Zhang J, Zhao D, et al. Molecular mechanisms of pancreatic cancer liver metastasis: the role of PAK2. Front Immunol. (2024) 15:1347683. doi: 10.3389/fimmu.2024.1347683
9. Sun Z, Wang J, Fan Z, Yang Y, Meng X, Ma Z, et al. Investigating the prognostic role of lncRNAs associated with disulfidptosis-related genes in clear cell renal cell carcinoma. J Gene Med. (2024) 26:e3608. doi: 10.1002/jgm.v26.1
10. Desai SA, Patel VP, Bhosle KP, Nagare SD, Thombare KC. The tumor microenvironment: shaping cancer progression and treatment response. J Chemother. (2025) 2024:1–30. doi: 10.1080/1120009X.2023.2300224
11. Zhang P, Pei S, Wu L, Xia Z, Wang Q, Huang X, et al. Integrating multiple machine learning methods to construct glutamine metabolism-related signatures in lung adenocarcinoma. Front Endocrinol (Lausanne). (2023) 14:1196372. doi: 10.3389/fendo.2023.1196372
12. Zhang X, Zhuge J, Liu J, Xia Z, Wang H, Gao Q, et al. Prognostic signatures of sphingolipids: Understanding the immune landscape and predictive role in immunotherapy response and outcomes of hepatocellular carcinoma. Front Immunol. (2023) 14:1153423. doi: 10.3389/fimmu.2023.1153423
13. Wang X, Zhao Y, Strohmer DF, Yang W, Xia Z, Yu C. The prognostic value of MicroRNAs associated with fatty acid metabolism in head and neck squamous cell carcinoma. Front Genet. (2022) 13:983672. doi: 10.3389/fgene.2022.983672
14. Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. (2013) 19:1423–37. doi: 10.1038/nm.3394
15. Shee K, Yang W, Hinds JW, Hampsch RA, Varn FS, Traphagen NA, et al. Therapeutically targeting tumor microenvironment-mediated drug resistance in estrogen receptor-positive breast cancer. J Exp Med. (2018) 215:895–910. doi: 10.1084/jem.20171818
16. Mantovani A, Allavena P, Marchesi F, Garlanda C. Macrophages as tools and targets in cancer therapy. Nat Rev Drug Discovery. (2022) 21:799–820. doi: 10.1038/s41573-022-00520-5
17. Chen S, Lai SWT, Brown CE, Feng M. Harnessing and enhancing macrophage phagocytosis for cancer therapy. Front Immunol. (2021) 12:635173. doi: 10.3389/fimmu.2021.635173
18. Li H, Yang P, Wang J, Zhang J, Ma Q, Jiang Y, et al. HLF regulates ferroptosis, development and chemoresistance of triple-negative breast cancer by activating tumor cell-macrophage crosstalk. J Hematol Oncol. (2022) 15:2. doi: 10.1186/s13045-021-01223-x
19. Cortes M, Sanchez-Moral L, de Barrios O, Fernandez-Acenero MJ, Martinez-Campanario MC, Esteve-Codina A, et al. Tumor-associated macrophages (TAMs) depend on ZEB1 for their cancer-promoting roles. EMBO J. (2017) 36:3336–55. doi: 10.15252/embj.201797345
20. Shapouri-Moghaddam A, Mohammadian S, Vazini H, Taghadosi M, Esmaeili SA, Mardani F, et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol. (2018) 233:6425–40. doi: 10.1002/jcp.v233.9
21. Xiang X, Wang J, Lu D, Xu X. Targeting tumor-associated macrophages to synergize tumor immunotherapy. Signal Transduct Target Ther. (2021) 6:75. doi: 10.1038/s41392-021-00484-9
22. Cassetta L, Pollard JW. A timeline of tumour-associated macrophage biology. Nat Rev Cancer. (2023) 23:238–57. doi: 10.1038/s41568-022-00547-1
23. Boutilier AJ, Elsawa SF. Macrophage polarization states in the tumor microenvironment. Int J Mol Sci. (2021) 22:6995. doi: 10.3390/ijms22136995
24. Duan Z, Luo Y. Targeting macrophages in cancer immunotherapy. Signal Transduct Target Ther. (2021) 6:127. doi: 10.1038/s41392-021-00506-6
25. Amer HT, Stein U, El Tayebi HM. The monocyte, a maestro in the tumor microenvironment (TME) of breast cancer. Cancers (Basel). (2022) 14:5460. doi: 10.3390/cancers14215460
26. Rugo HS, Olopade OI, DeMichele A, Yau C, van ‘t Veer LJ, Buxton MB, et al. Adaptive randomization of veliparib-carboplatin treatment in breast cancer. N Engl J Med. (2016) 375:23–34. doi: 10.1056/NEJMoa1513749
27. Zhai X, Zhang H, Xia Z, Liu M, Du G, Jiang Z, et al. Oxytocin alleviates liver fibrosis via hepatic macrophages. JHEP Rep. (2024) 6:101032. doi: 10.1016/j.jhepr.2024.101032
28. Xiao J, Lin H, Liu B, Xia Z, Zhang J, Jin J. Decreased S1P and SPHK2 are involved in pancreatic acinar cell injury. biomark Med. (2019) 13:627–37. doi: 10.2217/bmm-2018-0404
29. Xiao J, Huang K, Lin H, Xia Z, Zhang J, Li D, et al. Mogroside II(E) inhibits digestive enzymes via suppression of interleukin 9/interleukin 9 receptor signalling in acute pancreatitis. Front Pharmacol. (2020) 11:859. doi: 10.3389/fphar.2020.00859
30. Zhang H, Xia T, Xia Z, Zhou H, Li Z, Wang W, et al. KIF18A inactivates hepatic stellate cells and alleviates liver fibrosis through the TTC3/Akt/mTOR pathway. Cell Mol Life Sci. (2024) 81:96. doi: 10.1007/s00018-024-05114-5
31. Basak U, Sarkar T, Mukherjee S, Chakraborty S, Dutta A, Dutta S, et al. Tumor-associated macrophages: an effective player of the tumor microenvironment. Front Immunol. (2023) 14:1295257. doi: 10.3389/fimmu.2023.1295257
32. Liu J, Cao X. Glucose metabolism of TAMs in tumor chemoresistance and metastasis. Trends Cell Biol. (2023) 33:967–78. doi: 10.1016/j.tcb.2023.03.008
33. Little AC, Pathanjeli P, Wu Z, Bao L, Goo LE, Yates JA, et al. IL-4/IL-13 stimulated macrophages enhance breast cancer invasion via rho-GTPase regulation of synergistic VEGF/CCL-18 signaling. Front Oncol. (2019) 9:456. doi: 10.3389/fonc.2019.00456
34. Zhang Q, Sioud M. Tumor-associated macrophage subsets: shaping polarization and targeting. Int J Mol Sci. (2023) 24:7493. doi: 10.3390/ijms24087493
35. Weng YS, Tseng HY, Chen YA, Shen PC, Al Haq AT, Chen LM, et al. MCT-1/miR-34a/IL-6/IL-6R signaling axis promotes EMT progression, cancer stemness and M2 macrophage polarization in triple-negative breast cancer. Mol Cancer. (2019) 18:42. doi: 10.1186/s12943-019-0988-0
36. Li D, Ji H, Niu X, Yin L, Wang Y, Gu Y, et al. Tumor-associated macrophages secrete CC-chemokine ligand 2 and induce tamoxifen resistance by activating PI3K/Akt/mTOR in breast cancer. Cancer Sci. (2020) 111:47–58. doi: 10.1111/cas.v111.1
37. Jaramillo-Valverde L, Levano KS, Capristano S, Tarazona DD, Cisneros A, Yufra-Picardo VM, et al. CXCR4 knockdown via CRISPR/CAS9 in a tumor-associated macrophage model decreases human breast cancer cell migration. Cureus. (2021) 13:e20842. doi: 10.7759/cureus.20842
38. Wang S, Jiang H, Zheng C, Gu M, Zheng X. Secretion of BMP-2 by tumor-associated macrophages (TAM) promotes microcalcifications in breast cancer. BMC Cancer. (2022) 22:34. doi: 10.1186/s12885-021-09150-3
39. Jaaskelainen MM, Tiainen S, Siiskonen H, Ahtiainen M, Kuopio T, Ronka A, et al. The prognostic and predictive role of tumor-infiltrating lymphocytes (FoxP3 + and CD8 +) and tumor-associated macrophages in early HER2 + breast cancer. Breast Cancer Res Treat. (2023) 201:183–92. doi: 10.1007/s10549-023-07017-8
40. Loi M, Salvatore G, Sottili M, Calosi L, Desideri I, Becherini C, et al. Tumor-associated macrophages (TAMs) modulate response to HER2-targeted agents in a humanized mouse model of breast cancer. Clin Transl Oncol. (2022) 24:1395–402. doi: 10.1007/s12094-022-02785-z
41. Ahmed S, Mohamed HT, El-Husseiny N, El Mahdy MM, Safwat G, Diab AA, et al. IL-8 secreted by tumor associated macrophages contribute to lapatinib resistance in HER2-positive locally advanced breast cancer via activation of Src/STAT3/ERK1/2-mediated EGFR signaling. Biochim Biophys Acta Mol Cell Res. (2021) 1868:118995. doi: 10.1016/j.bbamcr.2021.118995
42. Kang SU, Cho SY, Jeong H, Han J, Chae HY, Yang H, et al. Matrix metalloproteinase 11 (MMP11) in macrophages promotes the migration of HER2-positive breast cancer cells and monocyte recruitment through CCL2-CCR2 signaling. Lab Invest. (2022) 102:376–90. doi: 10.1038/s41374-021-00699-y
43. Liu Y, Zhu XZ, Xiao Y, Wu SY, Zuo WJ, Yu Q, et al. Subtyping-based platform guides precision medicine for heavily pretreated metastatic triple-negative breast cancer: The FUTURE phase II umbrella clinical trial. Cell Res. (2023) 33:389–402. doi: 10.1038/s41422-023-00795-2
44. Chen X, Yang M, Yin J, Li P, Zeng S, Zheng G, et al. Tumor-associated macrophages promote epithelial-mesenchymal transition and the cancer stem cell properties in triple-negative breast cancer through CCL2/AKT/beta-catenin signaling. Cell Commun Signal. (2022) 20:92. doi: 10.1186/s12964-022-00888-2
45. Zhao W, Zhou B, Sun J, Qiao H, Li J, Ma Q, et al. Macrophage CCL22 secretion under hypoxic conditions promotes the metastasis of triple-negative breast cancer. Proc Anticancer Res. (2022) 6:8–14. doi: 10.26689/par.v6i6.4493
46. Zhang Y, Fan Y, Jing X, Zhao L, Liu T, Wang L, et al. OTUD5-mediated deubiquitination of YAP in macrophage promotes M2 phenotype polarization and favors triple-negative breast cancer progression. Cancer Lett. (2021) 504:104–15. doi: 10.1016/j.canlet.2021.02.003
47. Shang L, Zhong Y, Yao Y, Liu C, Wang L, Zhang W, et al. Subverted macrophages in the triple-negative breast cancer ecosystem. BioMed Pharmacother. (2023) 166:115414. doi: 10.1016/j.biopha.2023.115414
48. Zivalj M, Van Ginderachter JA, Stijlemans B. Lipocalin-2: A nurturer of tumor progression and a novel candidate for targeted cancer therapy. Cancers (Basel). (2023) 15:5159. doi: 10.3390/cancers15215159
49. Nie G, Cao X, Mao Y, Lv Z, Lv M, Wang Y, et al. Tumor-associated macrophages-mediated CXCL8 infiltration enhances breast cancer metastasis: Suppression by Danirixin. Int Immunopharmacol. (2021) 95:107153. doi: 10.1016/j.intimp.2020.107153
50. Manore SG, Doheny DL, Wong GL, Lo H-W. IL-6/JAK/STAT3 signaling in breast cancer metastasis: biology and treatment. Front Oncol. (2022) 12:866014. doi: 10.3389/fonc.2022.866014
51. Banerjee K, Kerzel T, Bekkhus T, de Souza Ferreira S, Wallmann T, Wallerius M, et al. VEGF-C-expressing TAMs rewire the metastatic fate of breast cancer cells. Cell Rep. (2023) 42:113507. doi: 10.1016/j.celrep.2023.113507
52. Radharani NNV, Yadav AS, Nimma R, Kumar TVS, Bulbule A, Chanukuppa V, et al. Tumor-associated macrophage derived IL-6 enriches cancer stem cell population and promotes breast tumor progression via Stat-3 pathway. Cancer Cell Int. (2022) 22:122. doi: 10.1186/s12935-022-02527-9
53. Dong F, Ruan S, Wang J, Xia Y, Le K, Xiao X, et al. M2 macrophage-induced lncRNA PCAT6 facilitates tumorigenesis and angiogenesis of triple-negative breast cancer through modulation of VEGFR2. Cell Death Dis. (2020) 11:728. doi: 10.1038/s41419-020-02926-8
54. Cheng N, Bei Y, Song Y, Zhang W, Xu L, Zhang W, et al. B7-H3 augments the pro-angiogenic function of tumor-associated macrophages and acts as a novel adjuvant target for triple-negative breast cancer therapy. Biochem Pharmacol. (2021) 183:114298. doi: 10.1016/j.bcp.2020.114298
55. Zhao L, Wang Z, Tan Y, Ma J, Huang W, Zhang X, et al. IL-17A/CEBPbeta/OPN/LYVE-1 axis inhibits anti-tumor immunity by promoting tumor-associated tissue-resident macrophages. Cell Rep. (2024) 43:115039. doi: 10.1016/j.celrep.2024.115039
56. Wang Z, Zhang C, Guo J, Wang W, Si Q, Chen C, et al. Exosomal miRNA-223-3p derived from tumor associated macrophages promotes pulmonary metastasis of breast cancer 4T1 cells. Transl Oncol. (2023) 35:101715. doi: 10.1016/j.tranon.2023.101715
57. Munir MT, Kay MK, Kang MH, Rahman MM, Al-Harrasi A, Choudhury M, et al. Tumor-associated macrophages as multifaceted regulators of breast tumor growth. Int J Mol Sci. (2021) 22:6526. doi: 10.3390/ijms22126526
58. Yu X, Zhang Q, Zhang X, Han Q, Li H, Mao Y, et al. Exosomes from macrophages exposed to apoptotic breast cancer cells promote breast cancer proliferation and metastasis. J Cancer. (2019) 10:2892–906. doi: 10.7150/jca.31241
59. Liu CG, Chen J, Goh RMW, Liu YX, Wang L, Ma Z. The role of tumor-derived extracellular vesicles containing noncoding RNAs in mediating immune cell function and its implications from bench to bedside. Pharmacol Res. (2023) 191:106756. doi: 10.1016/j.phrs.2023.106756
60. Mehta AK, Cheney EM, Hartl CA, Pantelidou C, Oliwa M, Castrillon JA, et al. Targeting immunosuppressive macrophages overcomes PARP inhibitor resistance in BRCA1-associated triple-negative breast cancer. Nat Cancer. (2021) 2:66–82. doi: 10.1038/s43018-020-00148-7
61. Zhang W, Zhang Q, Yang N, Shi Q, Su H, Lin T, et al. Crosstalk between IL-15Ralpha(+) tumor-associated macrophages and breast cancer cells reduces CD8(+) T cell recruitment. Cancer Commun (Lond). (2022) 42:536–57. doi: 10.1002/cac2.12311
62. Tharp KM, Kersten K, Maller O, Timblin GA, Stashko C, Canale FP, et al. Tumor-associated macrophages restrict CD8(+) T cell function through collagen deposition and metabolic reprogramming of the breast cancer microenvironment. Nat Cancer. (2024) 5:1045–62. doi: 10.1038/s43018-024-00775-4
63. Nixon BG, Kuo F, Ji L, Liu M, Capistrano K, Do M, et al. Tumor-associated macrophages expressing the transcription factor IRF8 promote T cell exhaustion in cancer. Immunity. (2022) 55:2044–2058 e2045. doi: 10.1016/j.immuni.2022.10.002
64. Schmidt E, Distel L, Erber R, Buttner-Herold M, Rosahl MC, Ott OJ, et al. Tumor-associated neutrophils are a negative prognostic factor in early luminal breast cancers lacking immunosuppressive macrophage recruitment. Cancers (Basel). (2024) 16:3160. doi: 10.3390/cancers16183160
65. Van Acker HH, Anguille S, Willemen Y, Smits EL, Van Tendeloo VF. Bisphosphonates for cancer treatment: Mechanisms of action and lessons from clinical trials. Pharmacol Ther. (2016) 158:24–40. doi: 10.1016/j.pharmthera.2015.11.008
66. Junankar S, Shay G, Jurczyluk J, Ali N, Down J, Pocock N, et al. Real-time intravital imaging establishes tumor-associated macrophages as the extraskeletal target of bisphosphonate action in cancer. Cancer Discovery. (2015) 5:35–42. doi: 10.1158/2159-8290.CD-14-0621
67. Sun X, Gao D, Gao L, Zhang C, Yu X, Jia B, et al. Molecular imaging of tumor-infiltrating macrophages in a preclinical mouse model of breast cancer. Theranostics. (2015) 5:597–608. doi: 10.7150/thno.11546
68. Rumney RMH, Coffelt SB, Neale TA, Dhayade S, Tozer GM, Miller G. PyMT-Maclow: A novel, inducible, murine model for determining the role of CD68 positive cells in breast tumor development. PloS One. (2017) 12:e0188591. doi: 10.1371/journal.pone.0188591
69. Qi X, Lian Y, Fan Z, Wang H, Jiang H, He M, et al. Electroacupuncture normalized tumor vasculature by downregulating glyoxalase-1 to polarize tumor-associated macrophage to M1 phenotype in triple-negative breast cancer. Int Immunopharmacol. (2025) 147:113988. doi: 10.1016/j.intimp.2024.113988
70. Shen S, Zhang Y, Chen KG, Luo YL, Wang J. Cationic polymeric nanoparticle delivering CCR2 siRNA to inflammatory monocytes for tumor microenvironment modification and cancer therapy. Mol Pharm. (2018) 15:3642–53. doi: 10.1021/acs.molpharmaceut.7b00997
71. Svensson S, Abrahamsson A, Rodriguez GV, Olsson AK, Jensen L, Cao Y, et al. CCL2 and CCL5 are novel therapeutic targets for estrogen-dependent breast cancer. Clin Cancer Res. (2015) 21:3794–805. doi: 10.1158/1078-0432.CCR-15-0204
72. Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CM, Pryer N, et al. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell. (2014) 26:623–37. doi: 10.1016/j.ccell.2014.09.006
73. Pignatelli J, Goswami S, Jones JG, Rohan TE, Pieri E, Chen X, et al. Invasive breast carcinoma cells from patients exhibit MenaINV- and macrophage-dependent transendothelial migration. Sci Signal. (2014) 7:ra112. doi: 10.1126/scisignal.2005329
74. Shiao SL, Ruffell B, DeNardo DG, Faddegon BA, Park CC, Coussens LM. TH2-polarized CD4(+) T cells and macrophages limit efficacy of radiotherapy. Cancer Immunol Res. (2015) 3:518–25. doi: 10.1158/2326-6066.CIR-14-0232
75. Song Y, Tang C, Yin C. Combination antitumor immunotherapy with VEGF and PIGF siRNA via systemic delivery of multi-functionalized nanoparticles to tumor-associated macrophages and breast cancer cells. Biomaterials. (2018) 185:117–32. doi: 10.1016/j.biomaterials.2018.09.017
76. Guerriero JL, Sotayo A, Ponichtera HE, Castrillon JA, Pourzia AL, SChad S, et al. Class IIa HDAC inhibition reduces breast tumours and metastases through anti-tumour macrophages. Nature. (2017) 543:428–32. doi: 10.1038/nature21409
77. Sidiropoulos DN, Rafie CI, Jang JK, Castanon S, Baugh AG, Gonzalez E, et al. Entinostat decreases immune suppression to promote antitumor responses in a HER2+ Breast tumor microenvironment. Cancer Immunol Res. (2022) 10:656–69. doi: 10.1158/2326-6066.CIR-21-0170
78. Cao X, Li B, Chen J, Dang J, Chen S, Gunes EG, et al. Effect of cabazitaxel on macrophages improves CD47-targeted immunotherapy for triple-negative breast cancer. J Immunother Cancer. (2021) 9:e002022. doi: 10.1136/jitc-2020-002022
79. Vidyarthi A, Khan N, Agnihotri T, Negi S, Das DK, Aqdas M, et al. TLR-3 stimulation skews M2 macrophages to M1 through IFN-alphabeta signaling and restricts tumor progression. Front Immunol. (2018) 9:1650. doi: 10.3389/fimmu.2018.01650
80. Lei X, Chen M, Li X, Huang M, Nie Q, Ma N, et al. A vascular disrupting agent overcomes tumor multidrug resistance by skewing macrophage polarity toward the M1 phenotype. Cancer Lett. (2018) 418:239–49. doi: 10.1016/j.canlet.2018.01.016
81. Olsson A, Nakhle J, Sundstedt A, Plas P, Bauchet AL, Pierron V, et al. Tasquinimod triggers an early change in the polarization of tumor associated macrophages in the tumor microenvironment. J Immunother Cancer. (2015) 3:53. doi: 10.1186/s40425-015-0098-5
82. Guo C, Su Y, Wang B, Chen Q, Guo H, Kong M, et al. Novel polysaccharide building hybrid nanoparticles: remodelling TAMs to target ERalpha-positive breast cancer. J Drug Target. (2022) 30:450–62. doi: 10.1080/1061186X.2021.2020798
83. Kim J, Kim M, Yong SB, Han H, Kang S, Lahiji SF, et al. Engineering TGF-beta inhibitor-encapsulated macrophage-inspired multi-functional nanoparticles for combination cancer immunotherapy. Biomater Res. (2023) 27:136. doi: 10.1186/s40824-023-00470-y
84. Zhang L, Cui S, Ding N, Zhang J, Cui E, Xiang Q, et al. Tumor-associated macrophages regulating a polymer nanoplatform for synergistic treatment of breast tumors. ACS Appl Mater Interfaces. (2023) 15:34527–39. doi: 10.1021/acsami.3c05497
85. Yang M, Guo J, Li J, Wang S, Sun Y, Liu Y, et al. Platycodon grandiflorum-derived extracellular vesicles suppress triple-negative breast cancer growth by reversing the immunosuppressive tumor microenvironment and modulating the gut microbiota. J Nanobiotechnology. (2025) 23:92. doi: 10.1186/s12951-025-03139-x
86. Zhang LY, Chen XT, Li RT, Meng W, Huang GQ, Chen YJ, et al. Overcoming hypoxia-induced breast cancer drug resistance: a novel strategy using hollow gold-platinum bimetallic nanoshells. J Nanobiotechnology. (2025) 23:85. doi: 10.1186/s12951-025-03132-4
Keywords: breast cancer, macrophages, TAMs, tumor microenvironment, HER2, treatment
Citation: Jin H, Meng X and Feng J (2025) Mechanisms of tumor-associated macrophages in breast cancer and treatment strategy. Front. Immunol. 16:1560393. doi: 10.3389/fimmu.2025.1560393
Received: 14 January 2025; Accepted: 13 February 2025;
Published: 28 February 2025.
Edited by:
Zhiheng Lin, Shanghai University of Traditional Chinese Medicine, ChinaReviewed by:
Zhijia Xia, Affiliated Hospital of Guilin Medical University, ChinaCopyright © 2025 Jin, Meng and Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianwei Feng, Zmp3MTAwMUAxMjYuY29t; Xinyue Meng, bWVuZ3hpbnl1ZTg4QDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.