 Shengnan Yang1,2†
Shengnan Yang1,2† Fuxiao Liu
Fuxiao Liu- 1College of Veterinary Medicine, Qingdao Agricultural University, Qingdao, China
- 2Qingdao Center for Animal Disease Control & Prevention, Qingdao, China
A Commentary on
Inactivated rabies virus vectored MERS-Coronavirus vaccine induces protective immunity in mice, camels, and alpacas
By Chi H, Wang Y, Li E, Wang X, Wang H, Jin H, Han Q, Wang Z, Zhu A, Sun J, Zhuang Z, Zhang L, Ye J, Feng N, Hu M, Gao Y, Zhao J, Zhao Y, Yang S and Xia X (2022) Front. Immunol. 13: 823949. doi: 10.3389/fimmu.2022.823949
Introduction
A research article, named Inactivated Rabies Virus Vectored MERS-Coronavirus Vaccine Induces Protective Immunity in Mice, Camels, and Alpacas (doi: 10.3389/fimmu.2022.823949), was recently published in Frontiers in Immunology (Section: Vaccines and Molecular Therapeutics) (1). In this study, Chi et al. constructed a chimeric rabies virus (RABV) that expressed a genetically modified S1 gene from the Middle East respiratory syndrome coronavirus (MERS-CoV), and then evaluated its potential of virus-vectored vaccine after inactivation in different animals. This study demonstrated that the inactivated S1-expressing RABV was a promising vaccine candidate against MERS-CoV for camelids. Here, we would like to express our scientific opinions on this study.
Middle East respiratory syndrome
MERS is a severe infectious disease caused by MERS-CoV, initially identified in Saudi Arabia in 2012 (2). Typical signs clinically include fever, cough and shortness of breath in humans. MERS-CoV is a zoonotic virus, which has been identified in dromedary camels in several Member States in the Middle East, Africa and South Asia (3). There are a number of candidate vaccines that have been reported against MERS-CoV, including nucleic acid vaccine (4), subunit vaccine (5), nanoparticle vaccine (6), virus-vectored vaccine (7), and even live-attenuated vaccine (8). Most of the candidate vaccines have been designed using the MERS-CoV S protein, especially the S1 subunit.
Development of RABV-vectored vaccines
RABV virion is a bullet-shaped particle, containing a single-stranded, negative-sense RNA genome, coding for five proteins in order: nucleoprotein (N), phosphoprotein (P), matrix protein (M), glycoprotein (G), and RNA-dependent RNA polymerase (L). The RABV can be genetically modified using reverse genetics, whereby a foreign sequence can be inserted into the RABV genome for rescuing a replication-competent chimeric virus (9). This genetically modified RABV, if demonstrated to be able of expressing a foreign antigen that induces immune responses in vivo, would play a potential role in the development of RABV-vectored vaccines, such as live-attenuated vaccines (10, 11), inactivated vaccines (12, 13) and replication-deficient vaccines (14, 15).
Characteristics of chimeric RABV for developing inactivated vaccine
The inactivated RABV-vectored vaccine (IRVV) is a killed version of antigen-expressing RABV, rescued from its cDNA clone using reverse genetics. The development of an IRVV involves the construction of a chimeric RABV cDNA clone, which contains a foreign gene for virus recovery. More importantly, it must be ensured that the target antigen can be incorporated into the envelope of RABV virion (16). The foreign sequence-containing genome and the foreign antigen-containing virion are schematically shown in Figures 1A, B, respectively. Some RABV-expressed antigens will be processed, transported to the cell surface, and finally, as membrane-spanning proteins, embedded into the cellular envelope. Along with budding of RABV virion, the foreign antigen can be incorporated into the viral envelope (13, 17).
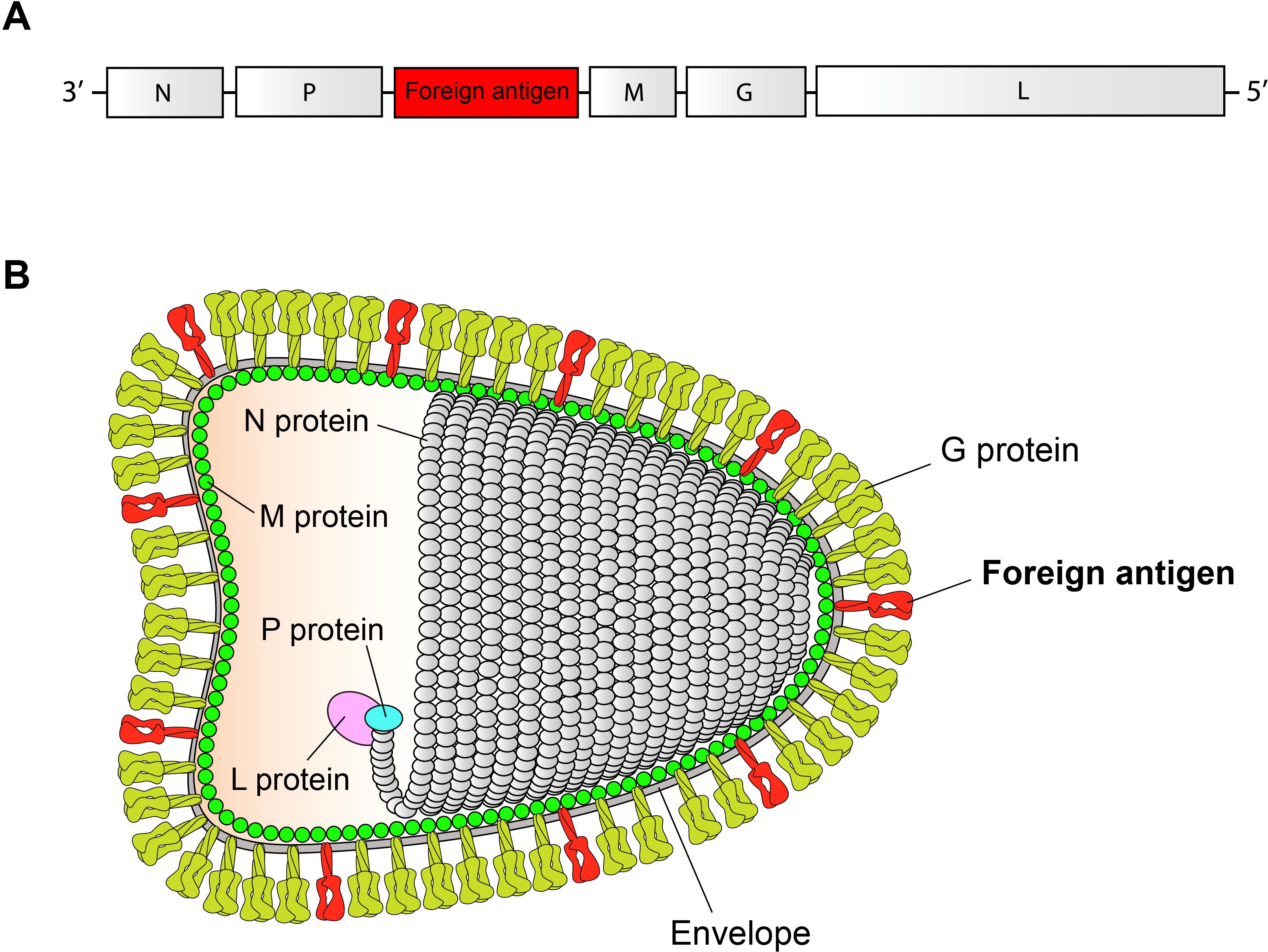
Figure 1. Schematic representations of chimeric RABV genome and virion. Chimeric RABV genome (A). The chimeric genome contains a single transcription unit of foreign antigen. Chimeric RABV virion (B). The foreign antigen, as a membrane-spanning protein, is embedded into the RABV envelope.
Development of anti-MERS-CoV IRVV
In the article published by Chi et al., the sequence of MERS-CoV S1 subunit was infused with the transmembrane domain (TM) of human CD4 and the cytoplasmic domain (CD) of RABV G protein. The fusion fragment, MERSS1-TM-CD, as a single transcription unit was inserted into the RABV (SRV9 strain) cDNA clone for rescuing a chimeric RABV, rSRV9-MERSS1. The transmission electron microscopy exhibited that the viable virus was successfully rescued using reverse genetics. The indirect immunofluorescence assay confirmed that the S1 subunit was expressed and transported to the cell surface. Subsequently, the rSRV9-MERSS1 stock was harvested, inactivated by the β-propiolactone, and then purified by the ultracentrifugation on a discontinuous sucrose gradient.
Further, Chi et al. carried out in vivo tests using three different animals: mice, camels, and alpacas. The test in mice revealed that the inactivated rSRV9-MERSS1 induced not only the robust, specific responses of MERS-CoV antibodies, but also the CD8+ T cell-specific responses. Moreover, the immunization alleviated virus replication and quickened virus clearance in MERS-CoV-infected mice. In addition, humoral immunities against MERS-CoV and RABV were identified in rSRV9-MERSS1-vaccinated camels and alpacas. The immune sera had a wide range of cross-neutralizing antibody responses against three MERS-CoV clades in camels and alpacas. Furthermore, MERS-CoV-specific variable domains of heavy-chain-only antibody were isolated from vaccinated alpacas and demonstrated to have robust therapeutic, prophylactic efficacies in a genetically modified mouse model.
Discussion
Compared with the live-attenuated vaccine, the IRVV shows a good safety profile in vivo, as evidenced by neither virus mutation nor virulence reversion occurring in IRVV-vaccinated animals. A chimeric RABV, albeit chemically inactivated, can even completely retain its immunogenicity, eliciting not only the anti-RABV immune response, but also more significantly, high-level antibodies against a target pathogen. In the study conducted by Chi et al., the rSRV9-MERSS1-inoculated mice, camels and alpacas were independently demonstrated to be able of secreting MERS-CoV-specific antibodies, implying its ability to inhibit the MERS-CoV infection in animals.
It has been widely demonstrated that inactivated chimeric RABVs have promising potentials in developing IRVVs (1, 12, 13, 17–23), whereas there are still a few disadvantages to them. For example, IRVVs may be less efficient than live-attenuated RABV-vectored vaccines in immunogenicity. The latter can elicit so potent immune response that a single dose is sufficient for the vaccination of animals, whereas the former are generally involved in the prime–boost vaccination (24, 25). In order to obtain MERS-CoV-specific antibodies at a high level, mice, camels and alpacas were separately subjected to the prime–boost vaccination in the study conducted by Chi et al. The test of MERS-CoV challenge showed that the rSRV9-MERSS1-based vaccination reduced MERS-CoV replication and accelerated its clearance in the lungs of genetically modified mice. Unfortunately, neither camels nor alpacas were subjected to the test of MERS-CoV challenge.
MERS-CoV has been still regarded as an emerging virus. Although various anti-MERS-CoV candidate vaccines have been recently reported, none of them has been commercially available as yet. In the field of virus-vectored vaccines, Chi et al. carried out their study that was valuable for guiding the development of IRVV. In conclusion, the inactivated rSRV9-MERSS1 was safe for the vaccine-inoculated animals, and induced potent immune responses in vivo. Therefore, this study will pave the way for the construction of virus-vectored vaccines against MERS-CoV in future.
Author contributions
SY: Writing – original draft. HL: Writing – original draft. FL: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Qingdao Demonstration Project for People-benefit from Science and Techniques (Grant No.: 24-1-8-xdny-4-nsh).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Chi H, Wang Y, Li E, Wang X, Wang H, Jin H, et al. Inactivated rabies virus vectored MERS-coronavirus vaccine induces protective immunity in mice, camels, and alpacas. Front Immunol. (2022) 13:823949. doi: 10.3389/fimmu.2022.823949
2. Bermingham A, Chand MA, Brown CS, Aarons E, Tong C, Langrish C, et al. Severe respiratory illness caused by a novel coronavirus, in a patient transferred to the United Kingdom from the Middle East, September 2012. Euro Surveill. (2012) 17:20290. doi: 10.2807/ese.17.40.20290-en
3. Peiris M, Perlman S. Unresolved questions in the zoonotic transmission of MERS. Curr Opin Virol. (2022) 52:258–64. doi: 10.1016/j.coviro.2021.12.013
4. Almansour I, Jermy BR. Nucleic acid vaccine candidates encapsulated with mesoporous silica nanoparticles against MERS-CoV. Hum Vaccin Immunother. (2024) 20:2346390. doi: 10.1080/21645515.2024.2346390
5. Zhang N, Tang J, Lu L, Jiang S, Du L. Receptor-binding domain-based subunit vaccines against MERS-CoV. Virus Res. (2015) 202:151–9. doi: 10.1016/j.virusres.2014.11.013
6. Hashemzadeh A, Avan A, Ferns GA, Khazaei M. Vaccines based on virus-like nano-particles for use against Middle East Respiratory Syndrome (MERS) coronavirus. Vaccine. (2020) 38:5742–6. doi: 10.1016/j.vaccine.2020.07.003
7. Malczyk AH, Kupke A, Prüfer S, Scheuplein VA, Hutzler S, Kreuz D, et al. A highly immunogenic and protective middle east respiratory syndrome coronavirus vaccine based on a recombinant measles virus vaccine platform. J Virol. (2015) 89:11654–67. doi: 10.1128/JVI.01815-15
8. Gutiérrez-Álvarez J, Honrubia JM, Fernández-Delgado R, Wang L, Castaño-Rodríguez C, Zúñiga S, et al. Genetically engineered live-attenuated middle east respiratory syndrome coronavirus viruses confer full protection against lethal infection. mBio. (2021) 12:e00103–21. doi: 10.1128/mBio.00103-21
9. Osakada F, Callaway EM. Design and generation of recombinant rabies virus vectors. Nat Protoc. (2013) 8:1583–601. doi: 10.1038/nprot.2013.094
10. Xu T, Liu L, Shi C, Liu W, Wang M, Tian L, et al. A recombinant rabies virus expressing Echinococcus granulosus EG95 induces protective immunity in mice. Transbound Emerg Dis. (2022) 69:e254–e66. doi: 10.1111/tbed.14292
11. Kurup D, Wirblich C, Feldmann H, Marzi A, Schnell MJ. Rhabdovirus-based vaccine platforms against henipaviruses. J Virol. (2015) 89:144–54. doi: 10.1128/JVI.02308-14
12. Jiao C, Liu D, Jin H, Huang P, Zhang H, Li Y, et al. Immunogenicity evaluation of a bivalent vaccine based on a recombinant rabies virus expressing gB protein of FHV-1 in mice and cats. Vet J. (2024) 304:106096. doi: 10.1016/j.tvjl.2024.106096
13. Abreu-Mota T, Hagen KR, Cooper K, Jahrling PB, Tan G, Wirblich C, et al. Non-neutralizing antibodies elicited by recombinant Lassa-Rabies vaccine are critical for protection against Lassa fever. Nat Commun. (2018) 9:4223. doi: 10.1038/s41467-018-06741-w
14. Takayama-Ito M, Lim CK, Yamaguchi Y, Posadas-Herrera G, Kato H, Iizuka I, et al. Replication-incompetent rabies virus vector harboring glycoprotein gene of lymphocytic choriomeningitis virus (LCMV) protects mice from LCMV challenge. PloS Negl Trop Dis. (2018) 12:e0006398. doi: 10.1371/journal.pntd.0006398
15. Gomme EA, Faul EJ, Flomenberg P, McGettigan JP, Schnell MJ. Characterization of a single-cycle rabies virus-based vaccine vector. J Virol. (2010) 84:2820–31. doi: 10.1128/JVI.01870-09
16. Scher G, Schnell MJ. Rhabdoviruses as vectors for vaccines and therapeutics. Curr Opin Virol. (2020) 44:169–82. doi: 10.1016/j.coviro.2020.09.003
17. Shuai L, Wang X, Wen Z, Ge J, Wang J, Zhao D, et al. Genetically modified rabies virus-vectored Ebola virus disease vaccines are safe and induce efficacious immune responses in mice and dogs. Antiviral Res. (2017) 146:36–44. doi: 10.1016/j.antiviral.2017.08.011
18. Smith ME, Koser M, Xiao S, Siler C, McGettigan JP, Calkins C, et al. Rabies virus glycoprotein as a carrier for anthrax protective antigen. Virology. (2006) 353:344–56. doi: 10.1016/j.virol.2006.05.010
19. Rios S, Bhattachan B, Vavilikolanu K, Kitsou C, Pal U, Schnell MJ. The Development of a Rabies Virus-Vectored Vaccine against Borrelia burgdorferi, Targeting BBI39. Vaccines (Basel). (2024) 12:78. doi: 10.3390/vaccines12010078
20. Kurup D, Fisher CR, Scher G, Yankowski C, Testa A, Keshwara R, et al. Tetravalent rabies-vectored filovirus and lassa fever vaccine induces long-term immunity in nonhuman primates. J Infect Dis. (2021) 224:995–1004. doi: 10.1093/infdis/jiab014
21. Huang J, Wang W, Li H, Bai Y, Song Y, Jiao C, et al. Three in one: An effective and universal vaccine expressing heterologous tandem RBD trimer by rabies virus vector protects mice against SARS-CoV-2. Antiviral Res. (2024) 227:105905. doi: 10.1016/j.antiviral.2024.105905
22. Zhang H, Jin H, Yan F, Song Y, Dai J, Jiao C, et al. An inactivated recombinant rabies virus chimerically expressed RBD induces humoral and cellular immunity against SARS-CoV-2 and RABV. Virol Sin. (2023) 38:244–56. doi: 10.1016/j.virs.2022.12.006
23. Zhang S, Hao M, Feng N, Jin H, Yan F, Chi H, et al. Genetically modified rabies virus vector-based rift valley fever virus vaccine is safe and induces efficacious immune responses in mice. Viruses. (2019) 11:919. doi: 10.3390/v11100919
24. Keshwara R, Shiels T, Postnikova E, Kurup D, Wirblich C, Johnson RF, et al. Rabies-based vaccine induces potent immune responses against Nipah virus. NPJ Vaccines. (2019) 4:15. doi: 10.1038/s41541-019-0109-5
25. Keshwara R, Hagen KR, Abreu-Mota T, Papaneri AB, Liu D, Wirblich C, et al. A Recombinant Rabies Virus Expressing the Marburg Virus Glycoprotein Is Dependent upon Antibody-Mediated Cellular Cytotoxicity for Protection against Marburg Virus Disease in a Murine Model. J Virol. (2019) 93:e01865–18. doi: 10.1128/JVI.01865-18
Keywords: rabies virus, MERS-CoV, inactivated RABV-vectored vaccine, antibody, immune response
Citation: Yang S, Li H and Liu F (2025) Commentary: Inactivated rabies virus vectored MERS-Coronavirus vaccine induces protective immunity in mice, camels, and alpacas. Front. Immunol. 16:1549481. doi: 10.3389/fimmu.2025.1549481
Received: 21 December 2024; Accepted: 17 February 2025;
Published: 04 March 2025.
Edited by:
Jin Cui, Northeast Agricultural University, ChinaReviewed by:
Yongxin Hu, China Animal Health and Epidemiology Center, ChinaCopyright © 2025 Yang, Li and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fuxiao Liu, bGF1ZGF3bkAxMjYuY29t
†These authors have contributed equally to this work