 Elaine Cristina Pereira De Martinis1†
Elaine Cristina Pereira De Martinis1† Virgínia Farias Alves2†
Virgínia Farias Alves2† Marita Gimenez Pereira1
Marita Gimenez Pereira1 Leonardo Neves Andrade1
Leonardo Neves Andrade1 Nathália Abichabki1,3
Nathália Abichabki1,3 Anna Abramova3,4
Anna Abramova3,4 Mirjam Dannborg3,4,5
Mirjam Dannborg3,4,5 Johan Bengtsson-Palme3,4,5*
Johan Bengtsson-Palme3,4,5*- 1Ribeirão Preto School of Pharmaceutical Sciences, University of São Paulo, Ribeirão Preto, São Paulo, Brazil
- 2Faculdade de Farmácia, Universidade Federal de Goiás, Goiânia, Goiás, Brazil
- 3Division of Systems and Synthetic Biology, Department of Life Sciences, SciLifeLab, Chalmers University of Technology, Gothenburg, Sweden
- 4Centre for Antibiotic Resistance Research (CARe), Gothenburg, Sweden
- 5Department of Infectious Diseases, Institute of Biomedicine, University of Gothenburg, Gothenburg, Sweden
Recent advances in cell culturing and DNA sequencing have dramatically altered the field of human microbiome research. Three-dimensional (3D) cell culture is an important tool in cell biology, in cancer research, and for studying host-microbe interactions, as it mimics the in vivo characteristics of the host environment in an in vitro system, providing reliable and reproducible models. This work provides an overview of the main 3D culture techniques applied to study interactions between host cells and pathogenic microorganisms, how these systems can be integrated with high-throughput molecular methods, and how multi-species model systems may pave the way forward to pinpoint interactions among host, beneficial microbes and pathogens.
1 Introduction
The study of microbe-microbe interactions as well as microbial interactions with their hosts represents a grand challenge for science. The advancement of high throughput sequencing technologies has provided valuable tools to better understand microbial diversity and function in the human microbiome, especially when combined with the culturing of fastidious pathogens and members of human microbiota (1).
Animal-based models have played a critical role over the years for research on the microbiome, as well as in several other fields related to biological and biomedical sciences, since some animals – such as rodents – present a conserved common structural organization that allows the modeling of human conditions (2, 3). However, in recent years there has been increasing opposition to the excessive use of animals in experiments, especially for ethical reasons, which has led to a search for robust alternative models, such as in vitro, ex vivo, and in silico methods that can be used in scientific research as adjuncts to or in replacement of animal models (4).
Eukaryotic cell culture is an interesting alternative to animal models for many biomedical applications, but these methods are limited because they usually involve growing cell lines in monolayers, failing to mimic important tissue functions. To improve these models, cell lines can be grown in three-dimensional culture (3D), hence developing some of the typical tissue structures, such as expression of tight junction proteins and production of mucin in the case of intestinal cells (3, 5, 6). Moreover, different types of cell lines can be grown in 3D cultures, as outlined in Table 1, but their advantages and drawbacks must be taken into account to allow the best choice of model for each application.
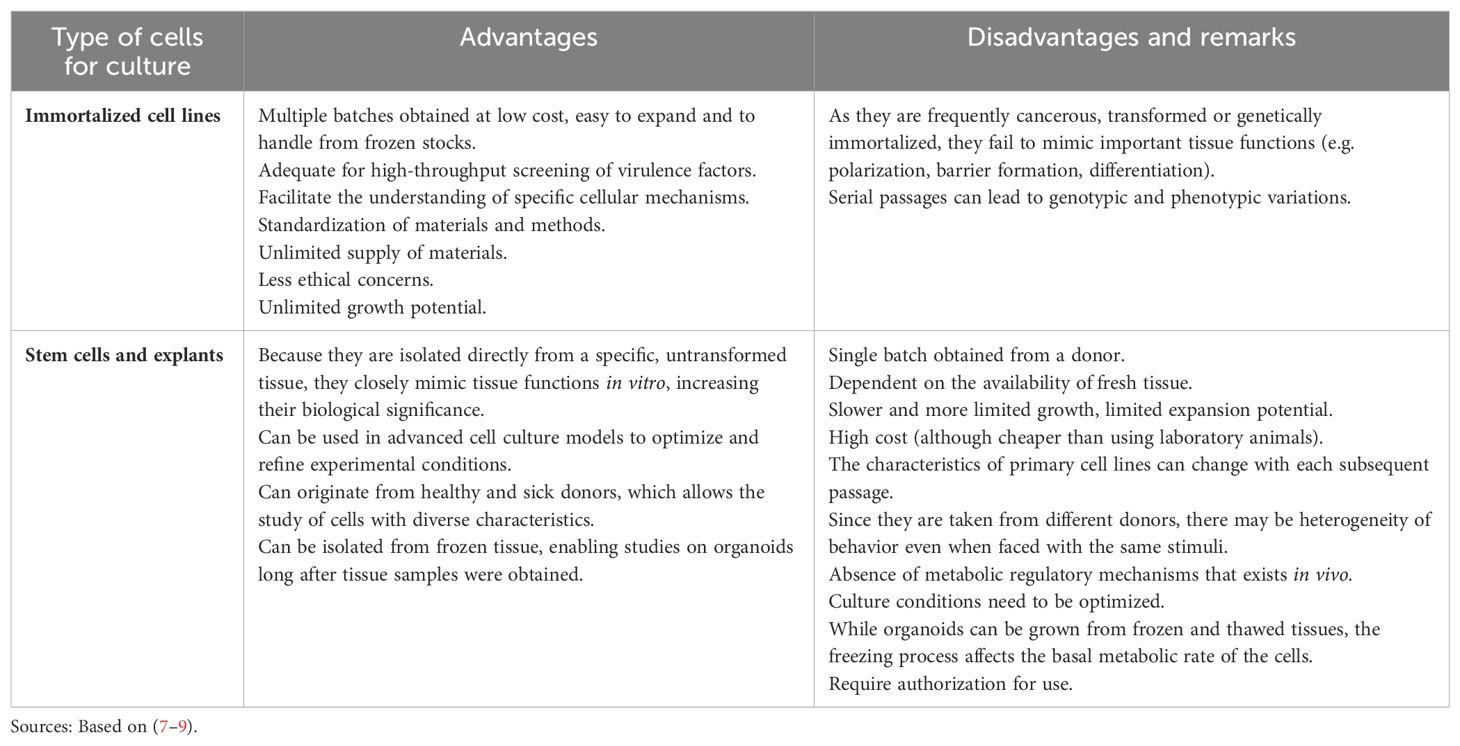
Table 1. Types of eukaryotic cells that can be used in three-dimensional models.
Three-dimensional cell culture has been applied in developmental, cellular, and cancer biology, as well as for studies of host-bacterial interactions since it mimics important characteristics that occur in vivo, including cell-cell and cell-extracellular matrix interactions in an in vitro system (6, 10, 11). Such 3D cultures represent a middle ground between monoculture experiments and animal models for the study of infectious diseases, especially if combined with high-throughput technologies. This combination aids in determination of host-specific immune responses and pathogen interactions, leading to new insights into both the pathogenesis and treatment of infections (12–14).
In view of the increasing accessibility and affordability of high-throughput techniques (e.g. transcriptomics, proteomics, and metabolomics), there are great opportunities for measuring the responses of 3D cultures in model systems at a very large scale, both on the eukaryotic tissue side and on the side of bacteria interacting with the host (14, 15). Thereby, experiments with 3D culture can improve the understanding of the host interactions with different microbial strains, as well as various drugs and chemicals.
To further increase the relevance of co-culture systems involving interactions between 3D-cultured human cells and bacteria one can introduce microbial communities rather than single species, to test for interactions in a multi-species context. However, as microbial communities are often highly complex and contain hundreds or thousands of species, the results of this approach are often hard to mechanistically interpret. To gain a better understanding of the mechanisms behind different interactions, microbial model communities can be used (16). Such microbial model communities offer a tradeoff allowing a reasonable degree of community complexity, but at the same time permit mapping a large part of the interactions between the microbes in the communities (and their host), which makes it possible to describe how the interactions are mediated and what genes are responsible for community behaviors.
Considering the importance of 3D eukaryotic cell culture models for studying host-microbe interactions, the main advantages and drawbacks of different systems available will be discussed in the next sections. Finally, the latest developments in 3D culture techniques combined with high-throughput molecular methods and multi-species model systems will be presented in the light of possible interactions among the host, beneficial microbes and pathogens.
2 Three-dimensional models for eukaryotic cell culture
The most recent advances in the development of technologies for 3D cultures of eukaryotic cells include multicellular spheroids, organoids, hydrogels, organ-on-a-chip platforms, and 3D bioimprints. Different principles and protocols for 3D cultures are used to recapitulate the morphology, as well as the functionality and microenvironmental characteristics of human tissues and organs, with different methods providing more realistic or reliable study systems for various purposes (Table 2). In particular for research on pathogenic bioagents, rotating wall vessel (RWV) bioreactors, extracellular matrix (ECM)-embedded/organoid models, and organ-on-a-chip (OAC) models have been used (15). RWV models have advanced to incorporate immune cells, allowing their role in host-microbe interactions to be elucidated (15). However, the choice of system depends on various factors, such as cost, technical complexity, and expertise needed to run experiments with the model.
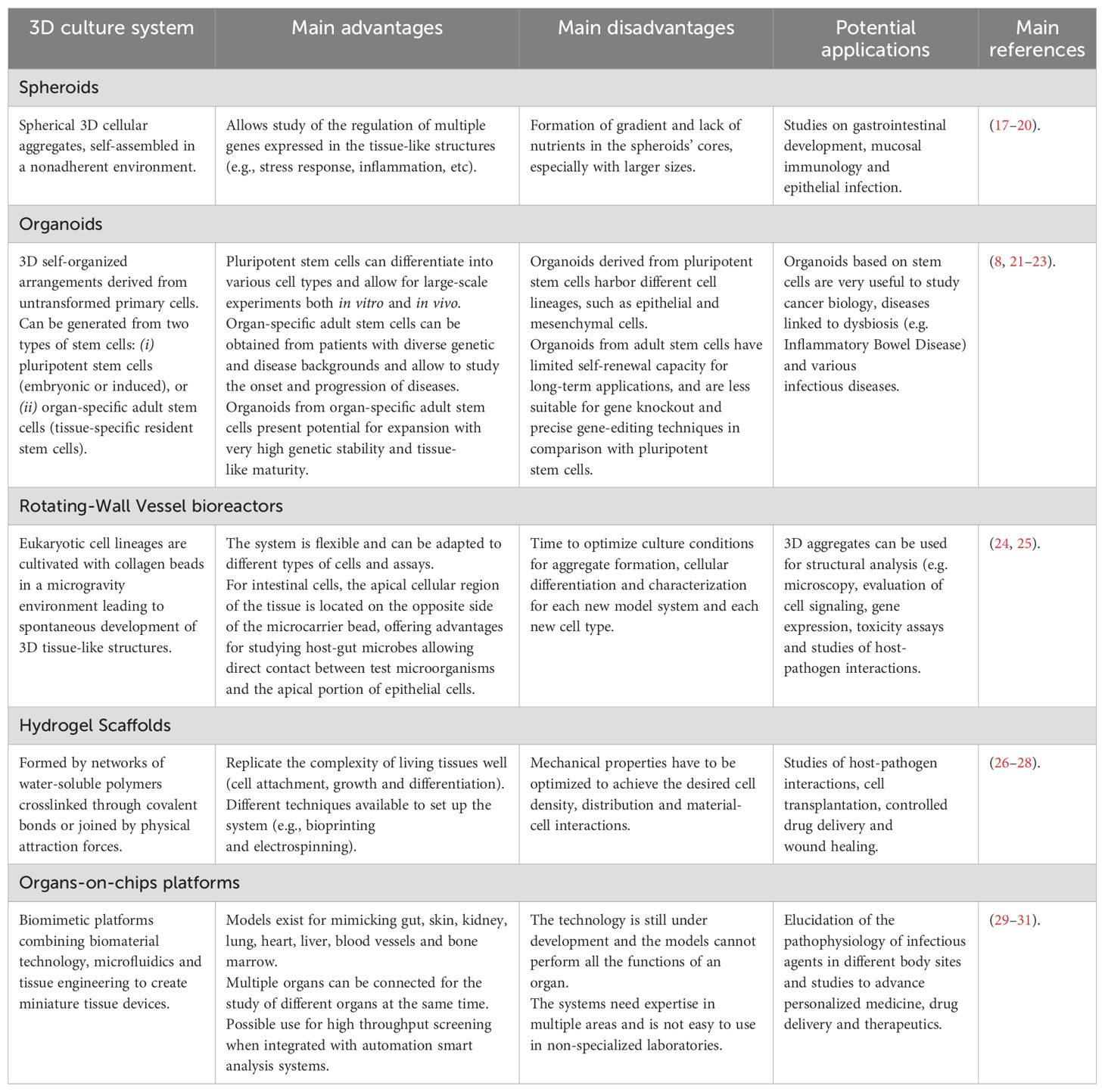
Table 2. Strengths, weaknesses, and applications of different 3D culture systems.
2.1 Spheroids
Spheroids are spherical 3D cellular aggregates, self-assembled in a nonadherent environment. The formation of spheroids occurs initially by the loose agglomeration of single cells from culture in suspension in a process of self-assembly. This is followed by the strong adhesion of initial cell aggregates promoted by homophilic binding between cadherins of peripheral cells, signal transduction by β-catenin and increased contact between adjacent cells promoted by actin, forming strong adhesive multi-cellular structures (20). There are different techniques for culturing spheroids, including pellet culture, liquid overlay, hanging drop, spinner culture, RWV, microfluidics, and magnetic levitation (20). Spheroids have positive regulation of multiple genes associated with the response to stress, inflammation, redox signaling, hypoxia, and angiogenesis (17, 18). This is advantageous for studying gastrointestinal development, mucosal immunology, and epithelial infection (19). However, the main disadvantage of this model is the formation of a diffusion gradient that occurs with increased spheroid size and can cause a lack of nutrients in the core of the spheroid (20). Some possible applications of spheroids include tumor models, tissue engineering, transplantation therapy, as well as drug screening (2, 32, 33). Figure 1 illustrates the steps of spheroid formation.

Figure 1. Scheme outlining the main steps and features in spheroid formation. Spheroids (3D) are derived from monolayer cultures (2D) but present higher degrees of cell-cell interaction and diffusion of fluids, leading to the expression of tissue-like properties (such as higher viability, proliferation and morphology, combined with less inflammation). Spheroids are adequate for large scale production by combining biomaterials with the cells, which can be used in systems with or without scaffolds, for different applications. Based on Laschke and Menger (32).
Some studies have reported on host-pathogen interactions using spheroids of eukaryotic cells. For example, Osswald et al. (34) observed that Salmonella enterica Typhimurium YB1 selectively localized, survived, and replicated in hypoxic areas within multicellular tumor spheroids (MCTS) from the HT-29 cell line. Furthermore, those authors reported that spores of Clostridium sporogenes, a strict anaerobe, germinated in the MCTS hypoxic areas. Such results indicate that spheroids can be an appropriate and reliable model to investigate live bacteria in their interactions with host tissues. In another study, Mukundan et al. (35) produced spheroids from THP-1 human monocyte/macrophage cells infected with mCherry-expressing Mycobacterium bovis BCG (Bacilli Camille Guérin) to mimic Mycobacterium tuberculosis infection and to create an HIV-TB (Human Immunodeficiency Virus - tuberculosis) co-infection model. In that study, the authors concluded that the spheroid system was useful for monitoring the kinetics of BCG growth, for studying HIV-TB co-infection and for tracking the anti-TB response of potential drug candidates. Broadly speaking, recent studies indicate spheroids prepared with eukaryotic cells are useful tools that deserve to be further explored to elucidate different mechanisms of host-microbe interactions.
2.2 Organoids
Organoids consist of 3D self-organized arrangements derived from untransformed primary cells, either Human Pluripotent Stem Cells (HPSC) or adult stem cells. These cells present the capacity to differentiate into various cell types, resembling natural properties of in vivo tissues, including protein expression, nutrient absorption, and barrier function (8, 36, 37). In organoid cultures, isolated stem cells receive, under specific culture conditions, combinations of selected growth factors to induce cell differentiation, so that the 3D structural organization, multicellularity, and functions of the in vivo target tissue/organ of interest are mimicked, hence they are also referred to as “organ in a dish” systems (8, 38–40). It is possible to produce organoids from virtually all human tissues, although brain and cardiac organoids can be only HPSC-derived, enabling the connection of cell culture and large-scale experiments in a variety of in vivo settings (21, 22) (Figure 2).

Figure 2. Schematic representation of organoid applications. 1 - Biobanking: organoids can be prepared from tissue resections after surgery and kept frozen in liquid nitrogen, which is important for studies on, e.g., personalized cancer therapy. 2 - Disease modelling: differentiated cells can be derived from human pluripotent stem cells and used for xenotransplantation in murines, serving as model phenotyping tool. 3 - Drug screening: organoids are appropriate for efficacy and toxicity testing. 4 - Host-microbe interactions: organoids are ideal models that recapitulate in vivo conditions for studies using human brain tissues (Zika virus) and intestinal cells (norovirus and rotavirus), among other infectious diseases. 5 - Multi-omics analysis: high-throughput characterization of healthy or diseased organoids can be done with regards to transcripts, proteins, metabolites, etc. 6 - Targeted gene editing: sequential mutations can be introduced into normal organoids to mimic different patient tumor formation scenarios (e.g., studies on colorectal carcinomas using CRISPR/Cas9). Figure based on Dutta et al. (39) and Hohwieler et al. (41).
An organoid generated from induced HPSC can harbor different cell lineages, such as epithelial and mesenchymal cells (8). Moreover, HPSC-derived organoids can be expanded to enable large-scale studies, such as drug screening and metabolic profiling (22). Organoids generated from adult tissue-specific stem cells consist of pure epithelial cells and generally present potential for expansion with very high genetic stability and tissue-like maturity (8, 42). However, these organoids have limited self-renewal capacity for long-term applications and are less suitable for gene knockout and precise gene-editing techniques, in comparison with HPSC-derived organoids (22). On the other hand, adult organoids can be derived from stem cells from a single individual, allowing the formation of an isogenic tissue with identical genes (42). Furthermore, adult organoids derived from different donors open up the possibility of developing induced pluripotent stem cells from patients with diverse genetic and disease backgrounds, which can leverage studies for understanding the onset and progression of those diseases, as well as potential treatments (22). This kind of approach, taking into consideration diverse genetic backgrounds, may help the study of diseases linked to dysbiosis, such as Inflammatory Bowel Disease (43, 44).
Organoids can be co-cultured (microinjected) with microorganisms to further simulate the tissue niche, thus constituting excellent platforms to evaluate host-pathogen dynamics, as well as to assist in the development of effective control measures against pathogens. This is especially true because organoids mimic the natural site of interaction, the natural target cell of infection, as well as the host factors that contribute to the outcome of the infection (45). Although there are model limitations, such as deficiency of immune cells, vasculature and innervation, the use of organoid models brings future perspectives to study not only known infectious diseases, but also future emerging ones (46).
Respiratory organoids, such as lung alveolar, lung airway, and bronchial organoids, have been used as models for infections caused by a variety of agents, including bacteria, fungi, parasites, and viruses, providing valuable insights into the underlying host-pathogen interactions at the cellular and molecular levels (45, 47). The recent COVID-19 (coronavirus disease 19) pandemic, caused by the SARS-CoV-2 virus, greatly boosted the development of new human lung organoid models. These models have contributed to better understanding the physiopathology of the disease with regards to host response and cellular damage mediated by immune cells, as well as the identification of potential therapeutic targets and the discovery of novel drugs (22, 46).
Human brain organoids (HBOs) have provided new physiologically relevant systems for functional modeling of the brain, as they can mimic the dynamic spatiotemporal process of early brain development. Indeed, HBOs have been used to study infectious agents with brain tropism, such as Zika virus (ZIKV), herpes simplex virus (HSV-1), human cytomegalovirus (HCMV) and Toxoplasma gondii (48). Watanabe et al. (49) developed an HBO system similar to the human fetal brain in vivo, efficiently reproducing cortical and basal ganglia structures, with functional neurons exhibiting spontaneous network-like activities, as demonstrated by immunohistochemical, transcriptomic, and electrophysiological analyses. Those authors were able to model the teratogenic effects of the ZIKV in the developing brain, identifying susceptibility receptors and therapeutic compounds that could mitigate the destructive action of ZIKV. Krenn et al. (48) used HBOs to reproduce the microcephaly-like phenotype caused by ZIKV and HSV-1, two viruses with different structures, sizes and modes of replication (48). The authors observed that both viruses replicated efficiently in the HBO system, causing features of microcephaly, albeit with major differences in the underlying structural defects and transcriptional profiles, as well as differences in involvement of the antiviral system and sensitivity to IFN-I (interferon -1).
Human intestinal organoids (HIOs) have been used to study enteric viruses and bacterial infections, as well as to evaluate commensal intestinal microbes (42, 50). HIOs derived from induced HPSCs, in addition to the luminal cells, present the majority of other cell types present in the human intestine, mimicking the in vivo native tissue architecture with villus and crypt domains, providing an effective system for studying the intestinal epithelium and its interaction with various stimuli, including enteric pathogens (38, 51). Using HIOs, Engevik et al. (52) demonstrated that Clostridioides difficile, a major cause of antibiotic-induced diarrhea, showed a decreased expression of the core protein MUC2 that is secreted by Goblet cells, affecting the thick layer of mucus that acts as a physical barrier against pathogens (52). On the other hand, Schulte et al. (53) developed an HIO model that comprised epithelial and endothelial cell layers, as well as a collagen matrix and immune cells, to study the acute phase of S. Typhimurium infection. The model mimicked human gastroenteritis by restricting the pathogen to the epithelial compartment, providing an advantage over existing mouse models (53). Intestinal organoids derived directly from adult tissues and made up only of epithelial cells are called enteroids or colonoids, and provide a physiologically relevant platform that, in addition to allowing the investigation of altered intestinal function and drug efficacy, also enables evaluation of interactions with microbes (6, 54). Using human small intestinal explants and enteroids, Sheikh et al. (55) demonstrated that EatA, a secreted autotransporter protein produced by some Enterotoxigenic Escherichia coli (ETEC) strains, plays a crucial role in the development of infection by degrading the mucin barrier, promoting microbial access to enterocytes, followed by toxin release. Enteroid systems have been used to clarify the pathogenicity mechanisms of infections caused by viruses (e.g. Norovirus, Adenovirus, Rotavirus) and bacteria (e.g. Salmonella, ETEC, enterohemorrhagic E. coli - EHEC) in the gastrointestinal tract (51, 55, 56).
2.3 Rotating-Wall Vessel bioreactors
Insights into microbe-host interactions can also be obtained with the organotypic Rotating-Wall Vessel (RWV) bioreactors, also known as “microgravity” reactors, which provide physiological-like conditions of fluid-shear, level of oxygenation and nutrients, favoring the development of specialized features of in vivo tissues, including spontaneous differentiation into multiple epithelial cell types, polarization, and cell-cell interactions (15, 25). In the RWV bioreactor, cells are cultivated with collagen-coated spheres or microcarrier matrices, that allow the spontaneous development of cohesive 3D structures representative of the parental tissue (24, 25). An interesting feature of RWV-bioreactors platforms for the study of host-microbe interactions is that the apical cellular region of the epithelial tissue is located on the opposite side of the collagen coating, allowing a better mimicry of in vivo conditions, since the microorganisms directly encounter the apical portion of the epithelial cells (24).
RWV-derived intestinal models have been used for studying interactions between eukaryotic cells and a variety of pathogens, such as S. Typhimurium (including multidrug-resistant strains from ST313), S. Typhi, pathogenic E. coli (EPEC, EHEC), Cryptosporidium parvum, Listeria monocytogenes, and human enteroviruses, including coxsackievirus B and poliovirus (5, 15, 24, 57–64). For example, Barrila et al. (64) developed and validated a novel RWV-based 3D co-culture infection model for Salmonella, using colonic epithelial cells and macrophages. For that, the authors first activated the pro-monocytic cell line U937 on collagen-coated scaffolds, and next added the HT-29 epithelial cells to the RWV 3D model for culturing until optimal differentiation was achieved. Those authors then used different Salmonella strains for studies of host-pathogen interactions and observed pathovar-specific differences in colonization patterns and intracellular colocalization. Overall, that study highlighted the usefulness of the RWV 3D model for identifying microenvironmental factors important for regulating infection.
Carvalho et al. (5) studied the interactions of EPEC and EHEC with human intestinal cells grown in microgravity in a rotary 3D cell culture system. As in many studies, this study did not directly compare how the bacteria adapted to organoids versus monolayers, nor whether the organoids showed a more natural response to bacterial exposure than monolayer cells. That said, the authors did find that organoids expressed more normal markers of intestinal tissue than conventionally grown monolayers, although these experiments were performed in the absence of bacteria. Organoids also produced higher levels of intestinally expressed disaccharidases and alkaline phosphatase than monolayers. In addition, the organoid HCT-8 developed microvilli and desmosomes characteristic of normal tissue (5). This speaks in favor of the organoids being more representative and more differentiated than HCT-8 cells grown as monolayers, which also implies that this would be a more realistic host-pathogen model.
Besides being a good model for the study of host-pathogen interplay, RWV bioreactors can be useful in assessing possible interactions between beneficial microbes and eukaryotic cells. For example, Pereira et al. (24) used the RWV microgravity model to study interactions between Lactobacillus sakei 1 (a potential probiotic strain) and Caco-2 cells, carrying out assays of adhesion/invasion and transcriptomics to elucidate the genes that were up or downregulated in intestinal cells by the presence of the lactic acid bacterium.
2.4 Hydrogel scaffolds
Hydrogels are 3D networks of water-soluble polymers from natural or synthetic sources, crosslinked either through covalent bonds or joined by physical intra- and intermolecular attraction forces (26). Hydrogels made with permeable biocompatible materials have been used as scaffolds to replicate the structural complexity of living tissues, as they provide a microenvironment for cell attachment, growth, differentiation, and interactions (10, 65). Hydrogel scaffolds can be obtained by several techniques, such as bioprinting and electrospinning (26). Bioprinting is a process based on additive manufacturing from materials containing living cells. These materials (so called bioink) are based on cytocompatible hydrogel precursor formulations and their properties are essential for printability: structural resolution, shape fidelity, and cell survival. Figure 3 illustrates possible cell distributions obtained in bioprinting, based on a study by Hölz et al. (27), for modeling the influence of cell density and mechanical properties of matrices (shear and loss modulus) during hydrogel fabrication. According to Hölz et al. (27) higher cell density can result in reduced modulus of hydrogels. Those authors also observed that hydrogel samples with edge and central cell clusters were softer compared to the ones with randomly distributed cells, but interestingly cluster distribution had minimal impact on the accumulated cell loadings in general.

Figure 3. Simplified representative hydrogel models in bioprinting, with possible cell distribution within the matrix: (A) random, (B) eight corner clusters and (C) central cluster. Based on Hölzl et al. (27), with permission under a Creative Commons license (https://creativecommons.org/licenses/by/3.0/).
Overall, models based on numerical approaches are very useful for depicting the cellular mechanics within hydrogels, allowing for the prediction of mechanical properties to achieve the desired cell density, distribution, and material–cell interaction, resulting in a suitable matured bioprinted tissue construct.
Electrospinning of hydrogels can also be used to produce scaffolds (26). In short, electrospinning is a process that uses electrostatic forces to pull fibers from a polymer solution and create nanofibrous structures that mimic the characteristics of extracellular matrices, providing a natural environment for tissue formation (66, 67). This versatile technique has relatively low experimental complexity and allows the use of a wide variety of biocompatible and biodegradable polymers to produce electrospun nanofiber scaffolds with physicochemical properties that promote cell adhesion, proliferation, and differentiation (26, 66, 67).
Hydrogel scaffolds have recently been applied to mimic host-pathogen interactions. In a study by Huang et al. (68), a mucus-like hydrogel was prepared using alginate-mucin (ALG-MUC) polymers crosslinked with calcium chloride. The hydrogel was incorporated into an aqueous two-phase system co-culture platform containing polyethylene glycol and dextran, to simultaneously support the growth of a monolayer of mammalian cells (16-HBE or Caco-2) and pathogenic bacteria (respectively for experiments with Pseudomonas aeruginosa or Shigella flexneri). The authors argued the ALG-MUC hydrogel combined with the polymer-based liquid microbial scaffold was suitable for mimicking the complexity of the human microbiome niche (68).
Alzheimer et al. (12) proposed to use an extracellular matrix structure named Small Intestine Submucosa – “SISmuc” – to study infection by Campylobacter jejuni in Caco-2 cells. That hydrogel scaffold allowed the formation of a polarized epithelial barrier and to follow the infectious process. Furthermore, the 3D platform enabled the identification of isolate-specific infection phenotypes, as well as relevant outcomes of the infectious process. Moreover, it was revealed that a small RNA pair was involved in the pathogenesis of C. jejuni, which suggested “SISmuc” could be useful for studying gastrointestinal pathogen infection mechanisms (12).
Biomimetic bone scaffolds containing collagen and minerals can also be useful for studying other host-pathogen interactions, such as in the case of osteomyelitis – an inflammatory disease of the bone caused by a wide range of opportunistic pathogens. Parente et al. (13) developed an osteomyelitis model based on murine osteoblastic cells (MC3T3-E1) to study infection by Staphylococcus aureus. Those authors observed the cells were well adapted to the hydrogel scaffold culture condition and revealed important mechanisms of bacterial pathogenesis. Taking these examples together, it is clear that hydrogel scaffolds can be useful tools to investigate host-pathogen interactions of relevance for human health.
2.5 Organ-on-a-chip platforms
Organ-on-a-chip platforms are biomimetic tools that combine biomaterial technology, microfluidics, and tissue engineering to create miniature tissue devices, thereby allowing the recapitulation of the physiologically critical characteristics of specific human tissues and organs and their interactions on a small scale (29, 30). The chip is prepared with transparent and biocompatible polymers and contains continuously perfused chambers, where specific cell cultures can be deposited (29). Furthermore, the device has at least one channel with multiple interfaces (air-liquid, liquid-liquid), physical stimulation (mechanical shear or stretching forces), and mimics tissue architecture (29). Currently, organ-on-a-chip models have been developed addressing multiple organs, such as the gut, skin, kidney, lung, heart, liver, blood vessels, and bone marrow. Although there are still many challenges associated with these technologies, the innovative organ-on-a-chip approach is extremely useful for elucidating the pathophysiological characteristics of infectious processes caused by different bioagents in diverse body sites, bringing new insights into the immunological responses, as well as mimicking clinical responses to antimicrobial agents and spontaneous microbial evolution related to drug exposure (29, 69).
Over time, microengineering has advanced and evolved, allowing the use of this effective technology to build organ-specific microenvironments to reconstitute tissue structures, tissue-to-tissue interactions, and interfaces, enabling the study of dynamic mechanical and biochemical stimuli found in specific organs, and the targeting of cells to group them into suitable tissues (70).
Gut-on-a-chip models have proved to be powerful tools for studying commensal and intestinal pathogens. In a simplified way, the device development involves the design, manufacturing of parts, assembly, and examination, taking into account the anatomy of the designed organ to evaluate whether the flow speed and shear forces are in accordance with the physiological conditions of the human body (71). Kim et al. (72) created a biomimetic microdevice containing microfluidic channels separated by a flexible porous membrane, surrounded by an extracellular matrix, and lined by Caco-2 cells, imitating the human intestinal physiological structure. Additionally, conditions of fluid flow and peristalsis-like motions were created, allowing the development of a polarized epithelium, containing villi, and forming an integral barrier. Finally, the authors inoculated Lactobacillus rhamnosus GG, a probiotic strain, which was successfully cultivated without compromising epithelial protection, and improving the intestinal protective barrier (72). Gut-on-a-chip microfluidic devices have also been applied to study virus-host interactions, using the enteropathogen coxsackievirus B (CVB) (15, 73). Exposure of CVB to the apical surface of the epithelium led to successful viral replication, induction of cytopathic effects, and polarized (apical) release of pro-inflammatory cytokines (15, 73). Basal side infection led to a decrease in viral titers and lower cytopathic effects, with apical secretion of pro-inflammatory cytokines. Grassart et al. (74), used the intestine-chip technology to address Shigella flexneri intestinal colonization and demonstrated the invasion of enterocytes through the apical portion, leading to loss of mucosal integrity. Also, those authors observed that S. flexneri took advantage of the peristaltic-like movements to invade the epithelium, a feature which would not have been possible to replicate using conventional in vitro study models (74).
Organ-on-a-chip models have also been used to study other infection sites. For example, to evaluate host-Mycobacterium tuberculosis interactions, Thacker et al. (75) developed a lung microdevice containing alveolar epithelial cells, macrophages, microchannels to mimic air and blood flow, plus surfactants. Those authors observed that, in the presence of surfactants, M. tuberculosis grew very slowly, both in the lung and immune cells or even did not grow at all, depending on the culture conditions. The authors also reported that the growth of M. tuberculosis into macrophages was attenuated in the presence of lung surfactants due to the removal of virulence-associated lipids from the bacterial cell surface (75). Other organ-on-a-chip models have been proposed to evaluate aspects of intricate parasite-host relationships, such as a liver-chip to address hepatitis B virus infection (76), a spleen-chip to study Plasmodium falciparum infection (77), an alveolus-chip model to evaluate co-infection with S. aureus and influenza virus (78), and an intestine chip to monitor ETEC infection (79), among others.
3 High-throughput sequencing methods
3.1 Transcriptomics and proteomics
Two of the major methods to get insights into the response of host cells to bacteria and, vice versa, are large-scale sequencing of RNA (transcriptomics or RNA-seq) and high-throughput identification of the protein content of the cells (proteomics). Transcriptomics has the benefit of being a relatively cheap “catch-all” type of method, in which the expression of all genes in a tissue can be quantified simultaneously and compared between conditions. In eukaryotic cells, one can take advantage of the poly-A-tail of mRNA to exclude most of the other forms of RNA (predominantly rRNA) contained in the cell, allowing to target the sequencing efforts towards protein-coding mRNA specifically. This has also enabled spatial transcriptomics (80, 81), in which transcription can be measured and connected to precise regions of the tissue, with close to individual eukaryotic cell resolution. However, this is not possible for bacteria, as they lack the poly-A-tail. This also means that in mixed samples, only a small fraction of the reads derived from an RNA-seq experiment will correspond to bacterial transcripts encoding proteins, requiring very high sequencing depths to perform relevant comparisons between conditions. Therefore, proteomic approaches might be more desirable in many cases to allow for studies of expressed proteins in both host and bacterial cells. Proteomic methods can also be targeted towards specific cellular compartments, such as the cell membranes or the cytoplasm, enabling an even stronger focus on the processes of interest.
3.2 Metagenomics and metatranscriptomics
When sequencing is performed on multi-species communities rather than individual species, it is referred to as metagenomics or metatranscriptomics (and correspondingly metaproteomics for proteins). Metagenomic sequencing of DNA has revolutionized our understanding of the diversity of bacteria and their functions, as it allows characterization of the uncultivable fraction of microbial communities. Analogously, metatranscriptomics and metaproteomics provide better pictures of the genes being expressed in a microbial community, or its protein content, respectively. This has been highly useful in deciphering a variety of host-associated environments, most predominantly the human gut (82–87), but also other body sites, such as the skin (88), stomach (89), vagina (90) and nasopharynx (91). It has also been successfully applied to better understand microbial ecosystems in a multitude of other environments (92–96). Yet, despite the versatility of these techniques, there are limitations to what can be achieved using them. A major hurdle is in the annotation of the sequence data. In principle, a good and representative reference data set is required to draw relevant conclusions based on the sequenced reads. Here, the most prominent issue is usually a lack of reference genomes to compare the reads to. Such lack can result in a large fraction of the obtained reads going unannotated (97, 98) creating large degrees of uncertainty. This uncertainty disproportionately will be associated with microbial species that are less well studied, biasing the analysis towards bacteria that are already well-characterized, such as E. coli and other Proteobacteria. While the magnitude of this problem has decreased as more genomes have been sequenced, this is still a major issue when undertaking metagenomic or metatranscriptomic sequencing in many environments, including the human body.
Another concern with the analysis of both transcriptomics and proteomics data is that the underlying assumptions of the analysis methods work well in single species studies, and to some degree in studies of host-single bacterium interactions. In the latter case, one major concern is the sometimes dramatically different abundances of host vs. bacterial material in various sample types. This problem can relatively easily be overcome by normalization methods when only one bacterial species is present together with the host. However, as the bacterial community combined with the host cells gets more complex, the problem with differential abundances gets exacerbated, as species in the microbial community may be present at vastly different abundances, sometimes differing by several orders of magnitude. This means that a transcriptomic or proteomic study in mixed-species communities may only be able to detect changes in transcription or protein composition for the most abundant microorganism(s) (99). Sometimes, the host material almost completely obscures the picture even in single-species studies. Two in vivo examples of these problems are studies of transcriptomes in the gastric environment (100) and the vagina (101). Both these environments are most often dominated by a single bacterium (Helicobacter in the stomach and different Lactobacillus species in the vagina) together with a vast amount of host cells. This means that while it is possible to detect other bacteria present in these environments, it is generally very hard to identify genes with differential expression in any other species than the dominant one. This also complicates comparisons between samples, particularly in the vaginal example where different species can be dominant in different persons.
Currently, the only feasible options to overcome these issues are to adapt the experimental methods or to use specialized normalization methods. When the microbial community composition is known, as in the case with microbial model communities (see section 4), target capture methods aimed to “fish” out the genomes of interest from a sample can be used (102, 103). One can also use cell sorting techniques to selectively perform, e.g., transcriptomics on only certain species in a community (104). This practice also allows measuring expression responses in single cells. Cell sorting is greatly eased using fluorescent markers, which allows the separation of different species in a microbial community (105). As host cells are usually much larger than microbial cells, these can typically be separated based on cell size. Regardless of the community, one can also apply more general approaches such as depletion of host DNA or RNA (106), depletion of rRNA in the case of metatranscriptomics (107), as well as applying combinations of metagenomics and metatranscriptomics that allow to better control for differences in gene expression levels between species (108).
Many of the problems with community transcriptomics (metatranscriptomics) can be overcome if one focuses on the DNA level instead of on expressed RNA. However, that comes with the loss of information on what genes are actually expressed in specific settings. The upside of this approach is that the differences in abundance between different species can be less extreme at the genomic (metagenome) level than at the transcriptional level, allowing for better detection of species that are enriched under certain conditions. Yet, if the abundance levels between species are extremely different, metagenomics suffers from the same issues as metatranscriptomics and metaproteomics. Overall, metagenomics, metatranscriptomics and metaproteomics are all formidable techniques to get better insights into host-associated communities. That said, they also all come with limitations which need to be taken into account when interpreting the outcomes of these experiments.
3.3 Large-scale transposon mutagenesis
Metagenomics can measure selective effects on the abundances of microorganisms, genes, and encoded functions in microbial communities, while metatranscriptomics and metaproteomics offer insights into the short-term responses to changing conditions. A complementary approach to these techniques is transposon mutagenesis. This term encompasses several specific protocols, all based on the same principles, aiming to characterize and quantify selection effects on individual genes at a very large scale (109). In brief, these protocols are based around a transposon – a piece of DNA that encodes a transposase protein that allows the piece of DNA to insert itself elsewhere in the genome – carried on a plasmid, which can be triggered to insert itself at a random location of a bacterial chromosome (110–112). Often the transposon used is a modified mariner transposon containing restriction enzyme recognition sites at its ends (Figure 4). The way these constructs are created allows for following individual viable deletion mutants using high-throughput sequencing, where specific tags associated with the gene disrupted by the transposon can be mapped back to the genome and identified. The benefit of these protocols is that both generation and detection of mutants can be achieved with very high throughput, in both in vitro and in vivo systems. By utilizing paired designs, in which the same source library of mutants is used for both a condition (such as growth in the presence of host cells) and a corresponding control (such as growth without host cells), the fitness for a given mutant can be directly compared between conditions. The paired setup allows for accurate and sensitive detection of mutants in genes important for growth under certain conditions, and specialized statistical packages exist for the analysis of such paired data (113).

Figure 4. Overview of the INSeq protocol, used as an example of a transposon sequencing technique. In this protocol, PCR amplification of 16-17 bp of DNA flanking each disrupted gene allows the fitness of individual mutants to be calculated through DNA sequencing. Specifically, a recognition site for the restriction enzyme MmeI has been inserted into the inverted repeats of the transposon. By using a linear PCR with a biotinylated primer (B), followed by binding of the PCR products to streptavidin beads, MmeI can cleave the DNA flanking a disrupted gene upstream of the transposon, leaving 16-17 bp of the gene sequence attached to the bead. This can subsequently be used for a regular PCR step, by ligating another DNA sequence containing an Illumina sequencing adapter and the binding site for another PCR primer, which will selectively amplify the 16-17 bp regions of disrupted genes. These can then be sequenced and quantified, allowing the determination of fitness effects by specific gene knock-outs at a large scale.
Transposon mutagenesis and sequencing allow for the determination of the fitness effects of essentially every non-lethal gene deletion in bacterial genomes. It has been applied across a whole range of organisms from different phyla, including E. coli (114), Klebsiella pneumoniae (115), P. aeruginosa (116), Bacteroides thetaiotaomicron (117), as well as Streptococcus and Staphylococcus species (118, 119). While the molecular protocols required to transform the plasmid into bacteria from different taxonomic groups may differ substantially, the overall approach has shown remarkable versatility. It can also be used both in vitro and in vivo (114), again highlighting its flexibility as a large-scale tool for assessing fitness costs associated with specific conditions and genes.
In the specific context of host-bacterial interactions, transposon mutagenesis can be used to specifically determine which genes are involved in surface colonization (120), cell communication between bacteria and host (121), and overall fitness in the host environment (114, 117). In addition, these techniques have also been used to study specific genes involved in tolerance and resistance to antibiotics (122–124), as well as those involved in competition and virulence (125). To study interactions between bacteria and host cells at a large scale, one could design experiments in which transposon mutant libraries containing a mixed pool of tens of thousands of mutants defective in specific genes are added to pre-grown organoids under a variety of conditions. This allows the study of specific genes important for bacterial adhesion to tissues, growth in the presence of host cells, as well as colonization and invasion of organoid representatives of human organs. In addition, since these analyses can be performed with high throughput, these fitness effects can be probed across a range of conditions and concentrations, enabling the resolution of complex environment-host-microbe dependencies.
Notably, as with every other high-throughput sequencing-based technique, transposon mutagenesis will generate a high degree of false positive observations. Therefore, caution has to be taken to make sure that appropriate multiple testing correction is applied to the statistical data. This is usually possible in all statistical packages for this type of analysis (113, 126–128), but is sometimes not the default option and it is important to make sure to use these correction techniques as appropriate when dealing with this type of data. Furthermore, any potentially important gene should be retested in independent experiments with individual single-gene mutants. This practice ensures that any noise introduced from the other mutants in the pooled libraries can be disregarded. Finally, the transposon mutant studies are somewhat limited in specific types of mutant phenotypes for which they can detect significant effects. For example, a mutant deficient in some secreted ‘common goods’ molecule (such as a signaling molecule) would be able to hitchhike on the other mutants in the pool that are deficient in other genes but can produce the common goods molecule. Thus, transposon mutagenesis techniques are restricted to finding genes involved in individual cell fitness and are more likely to identify genes as important if they are, e.g., involved in direct cell-to-cell contact via the membrane, transport of certain molecules from the cell via efflux pumps or intracellular enzymatic processes involving molecules that cannot be easily taken up from the local environment.
4 Microbial model communities
Similar to how organoid systems can be cost-efficient, relatively easy-to-use models that can function as alternatives to animal experiments, but can yet provide a more realistic picture of the complexity of human organs than 2D cell cultures, microbial model communities are model systems aiming to capture interspecies interactions without the full complexity of real-world microbial communities encompassing hundreds or thousands of species (16). Microbial model communities – also sometimes referred to as synthetic communities (SynComs) – consist of a small number of microbial species and strains, chosen in a way that allows the study of interaction phenomena in a controlled manner. A range of microbial model communities exist, targeting different research questions, species combinations, and environments (16). In the context of 3D cell culturing approaches, it is of particular interest to mention those aiming to capture interaction phenomena among bacteria inhabiting humans. For example, the complexity of the human gut microbiota has been targeted in both 14-species (129) and 12-species (130) models, as well as in models aiming to capture a larger swath of the intestinal microbiota diversity (131, 132). These model communities contain different species, represent different degrees of complexity, and cater to somewhat different research needs. Notably, only three species overlap between the 14- and 12-species models, with the species in the model developed by Venturelli et al. (130) being selected based on how well they mirror the functional and phylogenetic diversity of the human gut, while Gutiérrez and Garrido (129) chose to include species based on their involvement in inulin metabolism. Both these communities are aiming to recapture different aspects of interspecies interactions in the human microbiota. In contrast, the more complex communities used by Spragge et al. (131) and Wuyts et al. (132) rather focus on being comprehensive and then systematically remove species from a large pool to better understand ecological interactions (131) or drug metabolism in the human microbiota (132).
Furthermore, there are also a few different microbial model communities developed as wound or tissue infection models (133–135). These contain just two to three species, and they typically involve the opportunistic pathogen P. aeruginosa as a prominent member. This bacterium is among the most common ones included in microbial model communities (16) and is also relatively easy to do transposon mutagenesis in (136), making it – together with E. coli – an excellently well-suited pathogen model to use in organoid infection models aiming to understand pathogenicity. There are also microbial model communities for the human lung (135) and stomach (Mirjam Dannborg et al., personal communication) environments, which would also be highly suitable to combine with organoid studies.
The benefit of introducing microbial model communities as a tool to better understand host-bacteria interactions is that they allow a degree of bacterial interplay to take place in addition to the direct host-cell and bacterial-cell interactions. This adds an additional layer of ecological complexity to these systems; for example, microbial communities may be protecting the human cells, providing colonization resistance against pathogens, as observed in the human gut (137, 138). There might also be situations where an opportunistic pathogen alone will not be able to significantly disturb the host organs, but where substances produced by other bacteria can induce a more virulent phenotype. Finally, competition between bacteria for, e.g., nutrients can introduce highly unpredictable behaviors towards host cells, which would be impossible to capture in single-microbe systems.
5 Combining large-scale -omics with 3D cell culturing approaches
A variety of pathogens have been studied using 3D models of enteroids, colloids and organoids, including Salmonella spp., C. difficile, enteropathogenic E. coli (EPEC), EHEC, ETEC, norovirus, rotavirus, enterovirus, Toxoplasma gondii, and coronavirus (15). By combining these infection model systems with large-scale -omics approaches, including transcriptomics, metagenomics, and transposon sequencing, we can gain both a deep and broad insight into how the pathogen and the host interact, and how the host tissue responds to pathogen invasion and vice versa. Notably, the combination of these approaches can also be used to study interactions between host cells and the beneficial microbiome in a semi-realistic setting. Here, the addition of microbial model communities makes it possible to also decipher the molecular and genetic mechanisms behind host-microbiome interactions. Below, we will highlight a few prominent examples of studies combining omics and 3D culture system approaches.
3D organoid models have been used to mimic human gastroenteritis shortly after Salmonella infection. The pathogen is here restricted to the epithelial compartment, which is an advantage over existing mouse models. Based on immunohistochemical and microscopic analysis, Nickerson et al. (57) reported that 3D cultured intestinal cells (Int-407) more accurately replicated the complex environment that Salmonella enterica serovar Thyphimurium faces during the natural course of human infection. The authors showed that there were differences in adherence, invasion, and tissue pathology between non-infected monolayers and 3D cultures. It is interesting to note that S. Thyphimurium presented low ability to adhere and invade 3D intestinal cells, which presented tissue organization and differentiation similar to in vivo conditions, for example, with regard to the presence of basement membrane proteins, microvilli, tight junctions, mucin and sialyl Lewis A (indicative of M-cell glycosylation). In contrast, intestinal Int-407 cells in 2D culture were poorly organized and presented reduced expression of cell differentiation markers. Although Nickerson et al. (57) did not employ nucleotide sequencing, it is one of the earliest examples of the use of large-scale techniques to investigate host-pathogen interactions in 3D models. The authors obtained total RNA from cultures to investigate the pattern of expression of cytokines by the intestinal cells following infection by Salmonella, based on a multiprobe commercial assay. It was shown that infection of Int-407 monolayers and 3D aggregates by S. Thyphimurium (1 to 2 hours) resulted in significantly higher expression of proinflammatory and immunomodulatory cytokines, including Tumor Necrosis Factor alpha (TNF-α) and several interleukins (IL-6, IL-1α, IL-1β), compared to uninfected monolayers and 3D culture. More specifically, TNF-α expression increased in monolayers after 1h and continued to rise after 2h, in contrast with 3D cells that presented TNF-α mRNA levels significantly elevated at 1h, but did not increase from 1h to 2h of infection. In fact, at 2 hours the TNF-α expression was more than fivefold higher in 2D cultures in comparison with 3D aggregates. Taken altogether, the results from Nickerson et al. (57) indicate 3D tissue aggregates from RWVs may overcome many limitations of monolayer assays for the investigation of bacterial infections.
Using dual host-bacteria transcriptome sequencing of a scaffold-based 3D model infected with Salmonella, Schulte et al. (53) revealed the communication between epithelial, endothelial, monocytic, and natural killer cells, but also with the pathogen. This study showed that downstream effects of infection on endothelial cells and immune cells not detected in monocultures could be captured in the 3D organoid model. In addition, the study showed that dual RNA-seq can identify bacterial virulence strategies, along with the responses of the infected host cells, highlighting complex time-dependent interactions of these systems that would be hard to capture in, e.g., a mouse model.
Bartfeld et al. (139) used organoids differentiated into specific lines of the stomach, presenting repetitive architecture with regular invaginations called gastric glands, to study pathogen-host interactions. These stomach organoids were used to investigate the response to Helicobacter pylori infection, measured as induction of cytokine mRNA (139, 140). The study combined image analysis with fluorescence-activated cell sorting (FACS), PCR and microarrays, allowing a multifaceted view on host-pathogen interactions. Organoids derived from different stomach tissues expressed their expected markers and assembled into gland and pit domains by self-organization. By adding nicotinamide and withdrawing WNT, the cell type of the organoids could be controlled, and the gastric gland lineages showed a strong inflammatory response to H. pylori (139).
Belanger et al. (141) used a combination of human skin organoid models and transposon sequencing in P. aeruginosa to identify genes important for survival during nosocomial infection. They found that genes typically upregulated during infection, such as those involved in transport of divalent cations and metallic cations, genes encoding the pyochelin synthesis proteins, as well as pchG, were indeed important in the organoid system, but not in typical lab media conditions. These genes also included those involved in nucleotide metabolism, iron uptake and vitamin B12 biosynthesis. This is consistent with previous studies of infection in P. aeruginosa, and highlights the usability of organoids as relevant host-infection models (141).
6 Future perspectives
The above examples outline some of the possibilities on the horizon with better model systems. With both organoid techniques, sequencing technologies – particularly more innovative applications of nucleotide sequencing – and microbial model communities rapidly developing into robust, large-scale, generally applicable tools, there are many reasons to expect a breakthrough at the intersection of these technologies. Particularly, there is a scarcity of research published using microbial model communities in combination with organoid models, utilizing large-scale -omics methods to understand the multiway interactions in these systems. While setting up such combination systems is complicated, the fact that there are several examples combining pathogens and organoids indicates that the time is ripe to bring together model communities and 3D culture systems. From there, using large-scale transcriptomics, metagenomics and transposon sequencing as read-outs for both the organoid tissue and the bacterial community responses seems like a natural step with few hurdles, aside from perhaps the additional costs.
One of the greatest difficulties currently encountered in this type of studies is the visualization of images, as the 3D structures are larger than monolayers due to the presence of several cells in different visual planes. Anchorage-dependent cultures and spheroids are also difficult to image due to plate incompatibility with microscopes (142). The most common way to analyze cellular phenotypes is using conventional wide-field fluorescence or confocal microscopy (142). In this area, new, faster methodologies must be developed, to overcome the current limitations faced by 3D cell culture.
Despite the need for further methods development in the imaging area, and the currently limited use of combinations of high-throughput molecular techniques, 3D culturing systems, and microbial model systems, this intersect provides a lot of promise towards reproducibility, consistency and scalability, which is simply not offered by animal models or ex vivo culturing techniques. Uniting these methods into a common framework will allow models that account for interspecies microbial interactions, host-microbe interactions, and tissue responses in a highly reproducible fashion, which also allows data to be extracted from the models at very large scales. This, in turn, will allow utilization of machine learning approaches to generate new hypotheses and to improve the design of future experiments. Ultimately, this will open up new avenues of research into microbial communities and their interactions with the human body, while at the same time reducing the need for animal experiments. Consequently, animal studies can then be targeted towards the most relevant and promising leads and phenomena identified in the model systems. Ultimately, the human body and the niches it offers microbial inhabitants are immensely complex biological systems. To understand this complexity, we need to reduce it into components where we can more easily control the parameter space, akin to how E. coli grown in the lab is used as a model system to understand bacteria and living cells in general. Based on these components, we can then figure out the larger picture in smaller steps, by extrapolating from simpler to increasingly more complex models.
Author contributions
EM: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. VA: Conceptualization, Writing – original draft, Writing – review & editing. MP: Conceptualization, Writing – original draft. LA: Conceptualization, Funding acquisition, Supervision, Writing – review & editing. NA: Writing – review & editing, Data curation. AA: Data curation, Writing – review & editing, Investigation. MD: Investigation, Visualization, Writing – review & editing. JB-P: Writing – review & editing, Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001. EM is a fellow of the Brazilian National Council for Scientific and Technological Development - CNPq (Process #306998/2022-0) and she is grateful for Research Grants from São Paulo Research Foundation - Brazil (FAPESP #2017/18928-0 and #2020/00898-0) and CNPq (grant number 407025/2021-9). MP received a fellowship from the National Postdoctoral Program (PNPD) of the Coordination for the Improvement of Higher Education Personnel (CAPES) - Graduate Program on Biosciences and Biotechnology - Ribeirão Preto School of Pharmaceutical Sciences - University of São Paulo, Brazil. NA was supported by PhD fellowships CAPES (grant number 88887.670254/2022-0 (PROEX) and 8887.716776/2022-00 (PRINT)) and CNPq (grant number 140875/2024-7). JB-P is supported by funding from the Swedish Research Council (VR; grants 2020-00120 and 2020-03629), the Data-Driven Life Science (DDLS) program supported by the Knut and Alice Wallenberg Foundation (KAW 2020.0239), the Swedish Foundation for Strategic Research (FFL21-0174), the Centre for Antibiotic Resistance Research at the University of Gothenburg, and the Sahlgrenska Academy at the University of Gothenburg.
Acknowledgments
The authors would like to thank Emil Burman for valuable feedback on the conceptualization of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Egan S, Fukatsu T, Francino MP. Opportunities and challenges to microbial symbiosis research in the microbiome era. Front Microbiol. (2020) 11:1150. doi: 10.3389/fmicb.2020.01150
2. Bédard P, Gauvin S, Ferland K, Caneparo C, Pellerin È, Chabaud S, et al. Innovative human three-dimensional tissue-engineered models as an alternative to animal testing. Bioengineering. (2020) 7:115. doi: 10.3390/bioengineering7030115
3. Barroca NCB, Della Santa G, Suchecki D, García-Cairasco N, Umeoka EHdL. Challenges in the use of animal models and perspectives for a translational view of stress and psychopathologies. Neurosci Biobehav Rev. (2022) 140:104771. doi: 10.1016/j.neubiorev.2022.104771
4. European Food Safety Authority. Alternatives to Animal Testing (2024). Available online at: https://www.efsa.europa.eu/en/topics/topic/alternatives-animal-testing (Accessed April 16, 2024).
5. Carvalho HM, Teel LD, Goping G, O’Brien AD. A three-dimensional tissue culture model for the study of attach and efface lesion formation by enteropathogenic and enterohaemorrhagic Escherichia coli. Cell Microbiol. (2005) 7:1771–81. doi: 10.1111/j.1462-5822.2004.00594.x
6. Co JY, Margalef-Català M, Li X, Mah AT, Kuo CJ, Monack DM, et al. Controlling epithelial polarity: A human enteroid model for host-pathogen interactions. Cell Rep. (2019) 26:2509–2520.e4. doi: 10.1016/j.celrep.2019.01.108
7. Walsh AJ, Cook R, Sanders M, Arteaga CL, Skala MC. Drug response in organoids generated from frozen primary tumor tissues. Sci Rep. (2016) 6:18889. doi: 10.1038/srep18889
8. Aguilar C, Alves da Silva M, Saraiva M, Neyazi M, Olsson IAS, Bartfeld S. Organoids as host models for infection biology – a review of methods. Exp Mol Med. (2021) 53:1471–82. doi: 10.1038/s12276-021-00629-4
9. Voloshin N, Tyurin-Kuzmin P, Karagyaur M, Akopyan Z, Kulebyakin K. Practical use of immortalized cells in medicine: current advances and future perspectives. Int J Mol Sci. (2023) 24:12716. doi: 10.3390/ijms241612716
10. Fang Y, Eglen RM. Three-dimensional cell cultures in drug discovery and development. SLAS Discovery. (2017) 22:456–72. doi: 10.1177/1087057117696795
11. Koledova Z. 3D cell culture: An introduction. Methods Mol Biol. (2017) 1612:1–11. doi: 10.1007/978-1-4939-7021-6_1
12. Alzheimer M, Svensson SL, König F, Schweinlin M, Metzger M, Walles H, et al. A three-dimensional intestinal tissue model reveals factors and small regulatory RNAs important for colonization with Campylobacter jejuni. PloS Pathog. (2020) 16:e1008304. doi: 10.1371/journal.ppat.1008304
13. Parente R, Possetti V, Schiavone ML, Campodoni E, Menale C, Loppini M, et al. 3D cocultures of osteoblasts and Staphylococcus aureus on biomimetic bone scaffolds as a tool to investigate the host–pathogen interface in osteomyelitis. Pathogens. (2021) 10:837. doi: 10.3390/pathogens10070837
14. Pellegrino E, Gutierrez MG. Human stem cell-based models for studying host-pathogen interactions. Cell Microbiol. (2021) 23:e13335. doi: 10.1111/cmi.13335
15. Barrila J, Crabbé A, Yang J, Franco K, Nydam SD, Forsyth RJ, et al. Modeling host-pathogen interactions in the context of the microenvironment: three-dimensional cell culture comes of age. Infect Immun. (2018) 86(11):e00282-18. doi: 10.1128/IAI.00282-18
16. Bengtsson-Palme J. Microbial model communities: To understand complexity, harness the power of simplicity. Comput Struct Biotechnol J. (2020) 18:3987–4001. doi: 10.1016/j.csbj.2020.11.043
17. Potapova IA, Gaudette GR, Brink PR, Robinson RB, Rosen MR, Cohen IS, et al. Mesenchymal stem cells support migration, extracellular matrix invasion, proliferation, and survival of endothelial cells in vitro. Stem Cells. (2007) 25:1761–8. doi: 10.1634/stemcells.2007-0022
18. Jensen C, Teng Y. Is it time to start transitioning from 2D to 3D cell culture? Front Mol Biosci. (2020) 7:33. doi: 10.3389/fmolb.2020.00033
19. Sebrell TA, Sidar B, Bruns R, Wilkinson RA, Wiedenheft B, Taylor PJ, et al. Live imaging analysis of human gastric epithelial spheroids reveals spontaneous rupture, rotation and fusion events. Cell Tissue Res. (2018) 371:293–307. doi: 10.1007/s00441-017-2726-5
20. Ryu N-E, Lee S-H, Park H. Spheroid culture system methods and applications for mesenchymal stem cells. Cells. (2019) 8:1620. doi: 10.3390/cells8121620
21. Dahl-Jensen S, Grapin-Botton A. The physics of organoids: a biophysical approach to understanding organogenesis. Development. (2017) 144:946–51. doi: 10.1242/dev.143693
22. Han Y, Yang L, Lacko LA, Chen S. Human organoid models to study SARS-CoV-2 infection. Nat Methods. (2022) 19:418–28. doi: 10.1038/s41592-022-01453-y
23. Kim J, Koo BK, Knoblich JA. Human organoids: model systems for human biology and medicine. Nat. Rev. Mol. Cell. Biol. (2020) 21(10):571–84. doi: 10.1038/s41580-020-0259-3
24. Pereira MG, de Almeida OGG, da Silva HRA, Ishizawa MH, De Martinis ECP. Studies on host-foodborne bacteria in intestinal three-dimensional cell culture model indicate possible mechanisms of interaction. World J Microbiol Biotechnol. (2021) 37:31. doi: 10.1007/s11274-021-02996-6
25. Gardner J, Herbst-Kralovetz M. Three-dimensional rotating wall vessel-derived cell culture models for studying virus-host interactions. Viruses. (2016) 8:304. doi: 10.3390/v8110304
26. Xu F, Dawson C, Lamb M, Mueller E, Stefanek E, Akbari M, et al. Hydrogels for tissue engineering: Addressing key design needs toward clinical translation. Front Bioeng Biotechnol. (2022) 10:849831. doi: 10.3389/fbioe.2022.849831
27. Hölzl K, Lin S, Tytgat L, Van Vlierberghe S, Gu L, Ovsianikov A. Bioink properties before, during and after 3D bioprinting. Biofabrication. (2016) 8:32002. doi: 10.1088/1758-5090/8/3/032002
28. Radulescu DM, Neacsu IA, Grumezescu AM, Andronescu E. New insights of scaffolds based on hydrogels in tissue engineering. Polymers. (2022) 14:799. doi: 10.3390/polym14040799
29. Feaugas T, Sauvonnet N. Organ-on-chip to investigate host-pathogens interactions. Cell Microbiol. (2021) 23(7):e13336. doi: 10.1111/cmi.13336
30. Leung CM, de Haan P, Ronaldson-Bouchard K, Kim G-A, Ko J, Rho HS, et al. A guide to the organ-on-a-chip. Nat Rev Methods Primers. (2022) 2:33. doi: 10.1038/s43586-022-00118-6
31. Koyilot MC, Natarajan P, Hunt CR, Sivarajkumar S, Roy R, Joglekar S, et al. Breakthroughs and applications of organ-on-a-chip technology. Cells. (2022) 11:1828. doi: 10.3390/cells11111828
32. Laschke MW, Menger MD. Life is 3D: boosting spheroid function for tissue engineering. Trends Biotechnol. (2017) 35:133–44. doi: 10.1016/j.tibtech.2016.08.004
33. Tosca EM, Ronchi D, Facciolo D, Magni P. Replacement, reduction, and refinement of animal experiments in anticancer drug development: the contribution of 3D in vitro cancer models in the drug efficacy assessment. Biomedicines. (2023) 11:1058. doi: 10.3390/biomedicines11041058
34. Osswald A, Sun Z, Grimm V, Ampem G, Riegel K, Westendorf AM, et al. Three-dimensional tumor spheroids for in vitro analysis of bacteria as gene delivery vectors in tumor therapy. Microb Cell Fact. (2015) 14:199. doi: 10.1186/s12934-015-0383-5
35. Mukundan S, Singh P, Shah A, Kumar R, O’Neill KC, Carter CL, et al. In vitro miniaturized tuberculosis spheroid model. Biomedicines. (2021) 9:1209. doi: 10.3390/biomedicines9091209
36. Ramírez-Flores CJ, Knoll LJ. Breakthroughs in microbiology made possible with organoids. PloS Pathog. (2021) 17:e1010080. doi: 10.1371/journal.ppat.1010080
37. Suarez-Martinez E, Suazo-Sanchez I, Celis-Romero M, Carnero A. 3D and organoid culture in research: physiology, hereditary genetic diseases and cancer. Cell Biosci. (2022) 12:39. doi: 10.1186/s13578-022-00775-w
38. Forbester JL, Hannan N, Vallier L, Dougan G. Derivation of intestinal organoids from human induced pluripotent stem cells for use as an infection system. Methods Mol Biol. (2016) 1576:157–69. doi: 10.1007/7651_2016_7
39. Dutta D, Heo I, Clevers H. Disease modeling in stem cell-derived 3D organoid systems. Trends Mol Med. (2017) 23:393–410. doi: 10.1016/j.molmed.2017.02.007
40. Verma S, Senger S, Cherayil BJ, Faherty CS. Spheres of influence: insights into Salmonella pathogenesis from intestinal organoids. Microorganisms. (2020) 8:504. doi: 10.3390/microorganisms8040504
41. Hohwieler M, Illing A, Hermann PC, Mayer T, Stockmann M, Perkhofer L, et al. Human pluripotent stem cell-derived acinar/ductal organoids generate human pancreas upon orthotopic transplantation and allow disease modelling. Gut. (2017) 66:473–86. doi: 10.1136/gutjnl-2016-312423
42. Kim W, Gwon Y, Park S, Kim H, Kim J. Therapeutic strategies of three-dimensional stem cell spheroids and organoids for tissue repair and regeneration. Bioact Mater. (2023) 19:50–74. doi: 10.1016/j.bioactmat.2022.03.039
43. Yissachar N, Zhou Y, Ung L, Lai NY, Mohan JF, Ehrlicher A, et al. An intestinal organ culture system uncovers a role for the nervous system in microbe-immune crosstalk. Cell. (2017) 168:1135–1148.e12. doi: 10.1016/j.cell.2017.02.009
44. Cohen LJ, Cho JH, Gevers D, Chu H. Genetic factors and the intestinal microbiome guide development of microbe-based therapies for inflammatory bowel diseases. Gastroenterology. (2019) 156:2174–89. doi: 10.1053/j.gastro.2019.03.017
45. Paolicelli G, Luca A, Jose SS, Antonini M, Teloni I, Fric J, et al. Using lung organoids to investigate epithelial barrier complexity and IL-17 signaling during respiratory infection. Front Immunol. (2019) 10:323. doi: 10.3389/fimmu.2019.00323
46. Peng L, Gao L, Wu X, Fan Y, Liu M, Chen J, et al. Lung organoids as model to study SARS-CoV-2 Infection. Cells. (2022) 11:2758. doi: 10.3390/cells11172758
47. Bosáková V, De Zuani M, Sládková L, Garlíková Z, Jose SS, Zelante T, et al. Lung organoids—The ultimate tool to dissect pulmonary diseases? Front Cell Dev Biol. (2022) 10:899368. doi: 10.3389/fcell.2022.899368
48. Krenn V, Bosone C, Burkard TR, Spanier J, Kalinke U, Calistri A, et al. Organoid modeling of Zika and herpes simplex virus 1 infections reveals virus-specific responses leading to microcephaly. Cell Stem Cell. (2021) 28:1362–1379.e7. doi: 10.1016/j.stem.2021.03.004
49. Watanabe M, Buth JE, Vishlaghi N, de-la-Torre-Ubieta L, Taxidis J, Khakh BS, et al. Self-organized cerebral organoids with human-specific features predict effective drugs to combat Zika virus infection. Cell Rep. (2017) 21:517–32. doi: 10.1016/j.celrep.2017.09.047
50. Adeniyi-Ipadeola G, Nwanosike H, Ramani S. Human intestinal organoids as models to study enteric bacteria and viruses. Curr Opin Microbiol. (2023) 75:102362. doi: 10.1016/j.mib.2023.102362
51. Yin Y, Zhou D. Organoid and enteroid modeling of Salmonella infection. Front Cell Infect Microbiol. (2018) 8:102. doi: 10.3389/fcimb.2018.00102
52. Engevik MA, Yacyshyn MB, Engevik KA, Wang J, Darien B, Hassett DJ, et al. Human Clostridium difficile infection: altered mucus production and composition. Am J Physiol Gastrointest Liver Physiol. (2015) 308:G510–24. doi: 10.1152/ajpgi.00091.2014
53. Schulte LN, Schweinlin M, Westermann AJ, Janga H, Santos SC, Appenzeller S, et al. An advanced human intestinal coculture model reveals compartmentalized host and pathogen strategies during Salmonella Infection. mBio. (2020) 11:e03348–19. doi: 10.1128/mBio.03348-19
54. Ranganathan S, Smith EM, Foulke-Abel JD, Barry EM. Research in a time of enteroids and organoids: how the human gut model has transformed the study of enteric bacterial pathogens. Gut Microbes. (2020) 12:1795389. doi: 10.1080/19490976.2020.1795389
55. Sheikh A, Wangdi T, Vickers TJ, Aaron B, Palmer M, Miller MJ, et al. Enterotoxigenic Escherichia coli degrades the host MUC2 mucin barrier to facilitate critical pathogen-enterocyte interactions in human small intestine. Infect Immun. (2022) 90:e0057221. doi: 10.1128/iai.00572-21
56. Mirabelli C, Santos-Ferreira N, Gillilland MG, Cieza RJ, Colacino JA, Sexton JZ, et al. Human norovirus efficiently replicates in differentiated 3D-human intestinal enteroids. J Virol. (2022) 96:e0085522. doi: 10.1128/jvi.00855-22
57. Nickerson CA, Goodwin TJ, Terlonge J, Ott CM, Buchanan KL, Uicker WC, et al. Three-dimensional tissue assemblies: novel models for the study of Salmonella enterica serovar Typhimurium pathogenesis. Infect Immun. (2001) 69:7106–20. doi: 10.1128/IAI.69.11.7106-7120.2001
58. Höner zu Bentrup K, Ramamurthy R, Ott CM, Emami K, Nelman-Gonzalez M, Wilson JW, et al. Three-dimensional organotypic models of human colonic epithelium to study the early stages of enteric salmonellosis. Microbes Infect. (2006) 8:1813–25. doi: 10.1016/j.micinf.2006.02.020
59. Warren AC, Destura RV, Sevilleja JEAD, Barroso LF, Carvalho H, Barrett LJ, et al. Detection of epithelial-cell injury, and quantification of infection, in the HCT-8 organoid model of cryptosporidiosis. J Infect Dis. (2008) 198:143–9. doi: 10.1086/588819
60. Radtke AL, Wilson JW, Sarker S, Nickerson CA. Analysis of interactions of Salmonella type three secretion mutants with 3-D intestinal epithelial cells. PloS One. (2010) 5:e15750. doi: 10.1371/journal.pone.0015750
61. Salerno-Goncalves R, Fasano A, Sztein MB. Engineering of a multicellular organotypic model of the human intestinal mucosa. Gastroenterol. (2011) 141:e18–20. doi: 10.1053/j.gastro.2011.04.062
62. De Weirdt R, Crabbé A, Roos S, Vollenweider S, Lacroix C, van Pijkeren JP, et al. Glycerol supplementation enhances L. reuteri’s protective effect against S. Typhimurium colonization in a 3-D model of colonic epithelium. PloS One. (2012) 7:e37116. doi: 10.1371/journal.pone.0037116
63. Drummond CG, Nickerson CA, Coyne CB. A three-dimensional cell culture model to study enterovirus infection of polarized intestinal epithelial cells. mSphere. (2016) 1(1):e00030-15. doi: 10.1128/mSphere.00030-15
64. Barrila J, Yang J, Crabbé A, Sarker SF, Liu Y, Ott CM, et al. Three-dimensional organotypic co-culture model of intestinal epithelial cells and macrophages to study Salmonella enterica colonization patterns. NPJ Microgravity. (2017) 3:10. doi: 10.1038/s41526-017-0011-2
65. Nikolova MP, Chavali MS. Recent advances in biomaterials for 3D scaffolds: A review. Bioact Mater. (2019) 4:271–92. doi: 10.1016/j.bioactmat.2019.10.005
66. Nagaich U. Nanofibrous scaffold in tissue engineering. J Adv Pharm Technol Res. (2017) 8:85. doi: 10.4103/japtr.JAPTR_117_17
67. Zulkifli MZA, Nordin D, Shaari N, Kamarudin SK. Overview of electrospinning for tissue engineering applications. Polymers. (2023) 15:2418. doi: 10.3390/polym15112418
68. Huang AJ, O’Brien CL, Dawe N, Tahir A, Scott AJ, Leung BM. Characterization of an engineered mucus microenvironment for in vitro modeling of host–microbe interactions. Sci Rep. (2022) 12:5515. doi: 10.1038/s41598-022-09198-6
69. Baddal B, Marrazzo P. Refining host-pathogen interactions: organ-on-chip side of the coin. Pathogens. (2021) 10:203. doi: 10.3390/pathogens10020203
70. Yum K, Hong SG, Healy KE, Lee LP. Physiologically relevant organs on chips. Biotechnol J. (2014) 9:16–27. doi: 10.1002/biot.201300187
71. Xian C, Zhang J, Zhao S, Li X-G. Gut-on-a-chip for disease models. J Tissue Eng. (2023) 14:204173142211498. doi: 10.1177/20417314221149882
72. Kim HJ, Huh D, Hamilton G, Ingber DE. Human gut-on-a-chip inhabited by microbial flora that experiences intestinal peristalsis-like motions and flow. Lab Chip. (2012) 12:2165. doi: 10.1039/c2lc40074j
73. Villenave R, Wales SQ, Hamkins-Indik T, Papafragkou E, Weaver JC, Ferrante TC, et al. Human gut-on-a-chip supports polarized infection of Coxsackie B1 virus in vitro. PloS One. (2017) 12:e0169412. doi: 10.1371/journal.pone.0169412
74. Grassart A, Malardé V, Gobaa S, Sartori-Rupp A, Kerns J, Karalis K, et al. Bioengineered human organ-on-chip reveals intestinal microenvironment and mechanical forces impacting Shigella infection. Cell Host Microbe. (2019) 26:435–444.e4. doi: 10.1016/j.chom.2019.08.007
75. Thacker VV, Dhar N, Sharma K, Barrile R, Karalis K, McKinney JD. A lung-on-chip model of early Mycobacterium tuberculosis infection reveals an essential role for alveolar epithelial cells in controlling bacterial growth. Elife. (2020) 9:e59961. doi: 10.7554/eLife.59961
76. Kang Y, Rawat S, Duchemin N, Bouchard M, Noh M. Human liver sinusoid on a chip for hepatitis B virus replication study. Micromachines. (2017) 8:27. doi: 10.3390/mi8010027
77. Introini V, Govendir MA, Rayner JC, Cicuta P, Bernabeu M. Biophysical tools and concepts enable understanding of asexual blood stage malaria. Front Cell Infect Microbiol. (2022) 12:908241. doi: 10.3389/fcimb.2022.908241
78. Deinhardt-Emmer S, Rennert K, Schicke E, Cseresnyés Z, Windolph M, Nietzsche S, et al. Co-infection with Staphylococcus aureus after primary influenza virus infection leads to damage of the endothelium in a human alveolus-on-a-chip model. Biofabrication. (2020) 12:025012. doi: 10.1088/1758-5090/ab7073
79. Sunuwar L, Yin J, Kasendra M, Karalis K, Kaper J, Fleckenstein J, et al. Mechanical stimuli affect Escherichia coli heat-stable enterotoxin-cyclic GMP signaling in a human enteroid intestine-chip model. Infect Immun. (2020) 88:e00866–19. doi: 10.1128/IAI.00866-19
80. Salmén F, Ståhl PL, Mollbrink A, Navarro JF, Vickovic S, Frisén J, et al. Barcoded solid-phase RNA capture for spatial transcriptomics profiling in mammalian tissue sections. Nat Protoc. (2018) 13:2501–34. doi: 10.1038/s41596-018-0045-2
81. Strell C, Hilscher MM, Laxman N, Svedlund J, Wu C, Yokota C, et al. Placing RNA in context and space – methods for spatially resolved transcriptomics. FEBS J. (2019) 286:1468–81. doi: 10.1111/febs.14435
82. Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. (2010) 464:59–65. doi: 10.1038/nature08821
83. Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. (2011) 473:174–80. doi: 10.1038/nature09944
84. Bäckhed F, Roswall J, Peng Y, Feng Q, Jia H, Kovatcheva-Datchary P, et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe. (2015) 17:690–703. doi: 10.1016/j.chom.2015.04.004
85. Bengtsson-Palme J, Angelin M, Huss M, Kjellqvist S, Kristiansson E, Palmgren H, et al. The human gut microbiome as a transporter of antibiotic resistance genes between continents. Antimicrob Agents Chemother. (2015) 59:6551–60. doi: 10.1128/AAC.00933-15
86. Bonder MJ, Kurilshikov A, Tigchelaar EF, Mujagic Z, Imhann F, Vila AV, et al. The effect of host genetics on the gut microbiome. Nat Genet. (2016) 48:1407–12. doi: 10.1038/ng.3663
87. Alavi S, Mitchell JD, Cho JY, Liu R, Macbeth JC, Hsiao A. Interpersonal gut microbiome variation drives susceptibility and resistance to cholera infection. Cell. (2020) 181:1533–1546.e13. doi: 10.1016/j.cell.2020.05.036
88. Oh J, Byrd AL, Park M, Kong HH, Segre JA. Temporal stability of the human skin microbiome. Cell. (2016) 165:854–66. doi: 10.1016/j.cell.2016.04.008
89. Hu Y-L, Pang W, Huang Y, Zhang Y, Zhang C-J. The gastric microbiome is perturbed in advanced gastric adenocarcinoma identified through shotgun metagenomics. Front Cell Infect Microbiol. (2018) 8:433. doi: 10.3389/fcimb.2018.00433
90. Hugerth LW, Krog MC, Vomstein K, Du J, Bashir Z, Kaldhusdal V, et al. Defining vaginal community dynamics: daily microbiome transitions, the role of menstruation, bacteriophages, and bacterial genes. Microbiome. (2024) 12:153. doi: 10.1186/s40168-024-01870-5
91. Manenzhe RI, Dube FS, Wright M, Lennard K, Zar HJ, Mounaud S, et al. Longitudinal changes in the nasopharyngeal resistome of South African infants using shotgun metagenomic sequencing. PloS One. (2020) 15:e0231887. doi: 10.1371/journal.pone.0231887
92. Hugerth LW, Larsson J, Alneberg J, Lindh MV, Legrand C, Pinhassi J, et al. Metagenome-assembled genomes uncover a global brackish microbiome. Genome Biol. (2015) 16:279. doi: 10.1186/s13059-015-0834-7
93. Bahram M, Hildebrand F, Forslund SK, Anderson JL, Soudzilovskaia NA, Bodegom PM, et al. Structure and function of the global topsoil microbiome. Nature. (2018) 560:233–7. doi: 10.1038/s41586-018-0386-6
94. Fresia P, Antelo V, Salazar C, Giménez M, D’Alessandro B, Afshinnekoo E, et al. Urban metagenomics uncover antibiotic resistance reservoirs in coastal beach and sewage waters. Microbiome. (2019) 7:35. doi: 10.1186/s40168-019-0648-z
95. Cao J, Hu Y, Liu F, Wang Y, Bi Y, Lv N, et al. Metagenomic analysis reveals the microbiome and resistome in migratory birds. Microbiome. (2020) 8:26. doi: 10.1186/s40168-019-0781-8
96. Cao S, Zhang W, Ding W, Wang M, Fan S, Yang B, et al. Structure and function of the Arctic and Antarctic marine microbiota as revealed by metagenomics. Microbiome. (2020) 8:47. doi: 10.1186/s40168-020-00826-9
97. Solden L, Lloyd K, Wrighton K. The bright side of microbial dark matter: lessons learned from the uncultivated majority. Curr Opin Microbiol. (2016) 31:217–26. doi: 10.1016/j.mib.2016.04.020
98. Pavlopoulos GA, Baltoumas FA, Liu S, Selvitopi O, Camargo AP, Nayfach S, et al. Unraveling the functional dark matter through global metagenomics. Nature. (2023) 622:594–602. doi: 10.1038/s41586-023-06583-7
99. Hurley A, Chevrette MG, Rosario-Meléndez N, Handelsman J. THOR’s Hammer: the antibiotic Koreenceine drives gene expression in a model microbial community. mBio. (2022) 13:e02486–21. doi: 10.1128/mbio.02486-21
100. Thorell K, Bengtsson-Palme J, Liu OH-F, Palacios Gonzales RV, Nookaew I, Rabeneck L, et al. In vivo analysis of the viable microbiota and Helicobacter pylori transcriptome in gastric infection and early stages of carcinogenesis. Infect Immun. (2017) 85:e00031–17. doi: 10.1128/IAI.00031-17
101. Wikström T, Abrahamsson S, Bengtsson-Palme J, Ek J, Kuusela P, Rekabdar E, et al. Microbial and human transcriptome in vaginal fluid at midgestation: Association with spontaneous preterm delivery. Clin Transl Med. (2022) 12:e1023. doi: 10.1002/ctm2.1023
102. Metsky HC, Siddle KJ, Gladden-Young A, Qu J, Yang DK, Brehio P, et al. Capturing sequence diversity in metagenomes with comprehensive and scalable probe design. Nat Biotechnol. (2019) 37:160–8. doi: 10.1038/s41587-018-0006-x
103. Sundararaman B, Sylvester MD, Kozyreva VK, Berrada ZL, Corbett-Detig RB, Green RE. A hybridization target enrichment approach for pathogen genomics. mBio. (2023) 14(5):e01889-23. doi: 10.1128/mbio.01889-23
104. Lloréns-Rico V, Simcock JA, Huys GRB, Raes J. Single-cell approaches in human microbiome research. Cell. (2022) 185:2725–38. doi: 10.1016/j.cell.2022.06.040
105. Terekhov SS, Smirnov IV, Stepanova AV, Bobik TV, Mokrushina YA, Ponomarenko NA, et al. Microfluidic droplet platform for ultrahigh-throughput single-cell screening of biodiversity. PNAS Proc Natl Acad Sci. (2017) 114:2550–5. doi: 10.1073/pnas.1621226114
106. Heravi FS, Zakrzewski M, Vickery K, Hu H. Host DNA depletion efficiency of microbiome DNA enrichment methods in infected tissue samples. J Microbiol Methods. (2020) 170:105856. doi: 10.1016/j.mimet.2020.105856
107. Wangsanuwat C, Heom KA, Liu E, O’Malley MA, Dey SS. Efficient and cost-effective bacterial mRNA sequencing from low input samples through ribosomal RNA depletion. BMC Genomics. (2020) 21:717. doi: 10.1186/s12864-020-07134-4
108. Bikel S, Valdez-Lara A, Cornejo-Granados F, Rico K, Canizales-Quinteros S, Soberón X, et al. Combining metagenomics, metatranscriptomics and viromics to explore novel microbial interactions: towards a systems-level understanding of human microbiome. Comput Struct Biotechnol J. (2015) 13:390–401. doi: 10.1016/j.csbj.2015.06.001
109. Cain AK, Barquist L, Goodman AL, Paulsen IT, Parkhill J, van Opijnen T. A decade of advances in transposon-insertion sequencing. Nat Rev Genet. (2020) 21:526–40. doi: 10.1038/s41576-020-0244-x
110. Langridge GC, Phan M-D, Turner DJ, Perkins TT, Parts L, Haase J, et al. Simultaneous assay of every Salmonella Typhi gene using one million transposon mutants. Genome Res. (2009) 19:2308–16. doi: 10.1101/gr.097097.109
111. Goodman AL, Wu M, Gordon JI. Identifying microbial fitness determinants by insertion sequencing using genome-wide transposon mutant libraries. Nat Protoc. (2011) 6:1969–80. doi: 10.1038/nprot.2011.417
112. Klein BA, Duncan MJ, Hu LT. Defining essential genes and identifying virulence factors of porphyromonas gingivalis by massively parallel sequencing of transposon libraries (Tn-seq). Methods Mol Biol. (2015) 1279:25–43. doi: 10.1007/978-1-4939-2398-4_3
113. Abramova A, Osińska A, Kunche H, Burman E, Bengtsson-Palme J. CAFE: a software suite for analysis of paired-sample transposon insertion sequencing data. Bioinformatics. (2021) 37:121–2. doi: 10.1093/bioinformatics/btaa1086
114. Warr AR, Hubbard TP, Munera D, Blondel CJ, Abel zur Wiesch P, Abel S, et al. Transposon-insertion sequencing screens unveil requirements for EHEC growth and intestinal colonization. PloS Pathog. (2019) 15:e1007652. doi: 10.1371/journal.ppat.1007652
115. Paczosa MK, Silver RJ, McCabe AL, Tai AK, McLeish CH, Lazinski DW, et al. Transposon mutagenesis screen of Klebsiella pneumoniae identifies multiple genes important for resisting antimicrobial activities of neutrophils in mice. Infect Immun. (2020) 88:e00034–20. doi: 10.1128/IAI.00034-20
116. Ranjani J, Sivakumar R, Gunasekaran P, Velmurugan G, Ramasamy S, Rajendhran J. Genome-wide identification of genetic requirements of Pseudomonas aeruginosa PAO1 for rat cardiomyocyte (H9C2) infection by insertion sequencing. Infect Genet Evol. (2022) 98:105231. doi: 10.1016/j.meegid.2022.105231
117. Goodman AL, McNulty NP, Zhao Y, Leip D, Mitra RD, Lozupone CA, et al. Identifying genetic determinants needed to establish a human gut symbiont in its habitat. Cell Host Microbe. (2009) 6:279–89. doi: 10.1016/j.chom.2009.08.003
118. Le Breton Y, Belew AT, McIver KS. Protocols for Tn-seq analyses in the group A Streptococcus. Methods Mol Biol. (2020) 2136:33–57. doi: 10.1007/978-1-0716-0467-0_4
119. Sargison FA, Fitzgerald JR. Advances in transposon mutagenesis of Staphylococcus aureus: insights into pathogenesis and antimicrobial resistance. Trends Microbiol. (2021) 29:282–5. doi: 10.1016/j.tim.2020.11.003
120. Holden ER, Yasir M, Turner AK, Wain J, Charles IG, Webber MA. Massively parallel transposon mutagenesis identifies temporally essential genes for biofilm formation in Escherichia coli. Microb Genom. (2021) 7:673. doi: 10.1099/mgen.0.000673
121. Spencer H, Karavolos MH, Bulmer DM, Aldridge P, Chhabra SR, Winzer K, et al. Genome-wide transposon mutagenesis identifies a role for host neuroendocrine stress hormones in regulating the expression of virulence genes in Salmonella. J Bacteriol. (2010) 192:714–24. doi: 10.1128/JB.01329-09
122. Hu Y, Coates ARM. Transposon mutagenesis identifies genes which control antimicrobial drug tolerance in stationary-phase Escherichia coli. FEMS Microbiol Lett. (2005) 243:117–24. doi: 10.1016/j.femsle.2004.11.049
123. Coward C, Dharmalingham G, Abdulle O, Avis T, Beisken S, Breidenstein E, et al. High-density transposon libraries utilising outward-oriented promoters identify mechanisms of action and resistance to antimicrobials. FEMS Microbiol Lett. (2020) 367:fnaa185. doi: 10.1093/femsle/fnaa185
124. Zhu L, Eraso JM, Mangham RE, Ojeda Saavedra M, Olsen RJ, Beres SB, et al. Genome-wide transposon mutagenesis screens identify group A Streptococcus genes affecting susceptibility to β-lactam antibiotics. J Bacteriol. (2022) 204:e0028722. doi: 10.1128/jb.00287-22
125. Yin F, Hu Y, Bu Z, Liu Y, Zhang H, Hu Y, et al. Genome-wide identification of genes critical for in vivo fitness of multi-drug resistant porcine extraintestinal pathogenic Escherichia coli by transposon-directed insertion site sequencing using a mouse infection model. Virulence. (2023) 14:2158708. doi: 10.1080/21505594.2022.2158708
126. Zomer A, Burghout P, Bootsma HJ, Hermans PWM, van Hijum SAFT. ESSENTIALS: Software for rapid analysis of high throughput transposon insertion sequencing Data. PloS One. (2012) 7:e43012. doi: 10.1371/journal.pone.0043012
127. Blanchard AM, Leigh JA, Egan SA, Emes RD. Transposon insertion mapping with PIMMS – Pragmatic Insertional Mutation Mapping System. Front Genet. (2015) 6:139. doi: 10.3389/fgene.2015.00139
128. Zhao L, Anderson MT, Wu W, Mobley HLT, Bachman MA. TnseqDiff: Identification of conditionally essential genes in transposon sequencing studies. BMC Bioinf. (2017) 18:326. doi: 10.1186/s12859-017-1745-2
129. Gutiérrez N, Garrido D. Species deletions from microbiome consortia reveal key metabolic interactions between gut microbes. mSystems. (2019) 4:e00185–19. doi: 10.1128/mSystems.00185-19
130. Venturelli OS, Carr AV, Fisher G, Hsu RH, Lau R, Bowen BP, et al. Deciphering microbial interactions in synthetic human gut microbiome communities. Mol Syst Biol. (2018) 14:e8157. doi: 10.15252/msb.20178157
131. Spragge F, Bakkeren E, Jahn MT, Araujo BN, Pearson CF, Wang X, et al. Microbiome diversity protects against pathogens by nutrient blocking. Science. (2023) 382:eadj3502. doi: 10.1126/science.adj3502
132. Wuyts S, Alves R, Zimmermann-Kogadeeva M, Nishijima S, Blasche S, Driessen M, et al. Consistency across multi-omics layers in a drug-perturbed gut microbial community. Mol Syst Biol. (2023) 19:e11525. doi: 10.15252/msb.202311525
133. Sun Y, Dowd SE, Smith E, Rhoads DD, Wolcott RD. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regener. (2008) 16:805–13. doi: 10.1111/j.1524-475X.2008.00434.x
134. DeLeon S, Clinton A, Fowler H, Everett J, Horswill AR, Rumbaugh KP. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect Immun. (2014) 82:4718–28. doi: 10.1128/IAI.02198-14
135. Filkins LM, Graber JA, Olson DG, Dolben EL, Lynd LR, Bhuju S, et al. Coculture of Staphylococcus aureus with Pseudomonas aeruginosa drives S. aureus towards fermentative metabolism and reduced viability in a cystic fibrosis model. J Bacteriol. (2015) 197:2252–64. doi: 10.1128/JB.00059-15
136. Sivakumar R, Ranjani J, Vishnu US, Jayashree S, Lozano GL, Miles J, et al. Evaluation of INSeq to identify genes essential for Pseudomonas aeruginosa PGPR2 corn root colonization. G3. (2019) 9:651–61. doi: 10.1534/g3.118.200928
137. Buffie CG, Pamer EG. Microbiota-mediated colonization resistance against intestinal pathogens. Nat Rev Immunol. (2013) 13:790–801. doi: 10.1038/nri3535
138. Jacobson A, Lam L, Rajendram M, Tamburini F, Honeycutt J, Pham T, et al. A gut commensal-produced metabolite mediates colonization resistance to Salmonella infection. Cell Host Microbe. (2018) 24:296–307.e7. doi: 10.1016/j.chom.2018.07.002
139. Bartfeld S, Bayram T, van de Wetering M, Huch M, Begthel H, Kujala P, et al. In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology. (2015) 148:126–136.e6. doi: 10.1053/j.gastro.2014.09.042
140. Bartfeld S. Modeling infectious diseases and host-microbe interactions in gastrointestinal organoids. Dev Biol. (2016) 420:262–70. doi: 10.1016/j.ydbio.2016.09.014
141. Belanger CR, Dostert M, Blimkie TM, Lee AH-Y, Dhillon BK, Wu BC, et al. Surviving the host: microbial metabolic genes required for growth of Pseudomonas aeruginosa in physiologically-relevant conditions. Front Microbiol. (2022) 13:1055512. doi: 10.3389/fmicb.2022.1055512
Keywords: tridimensional cell culture, organoids, organ-on-a-chip, rotating wall vessel (RWV), high-throughput sequencing, transposon sequencing (TnSeq), microbial model communities
Citation: De Martinis ECP, Alves VF, Pereira MG, Andrade LN, Abichabki N, Abramova A, Dannborg M and Bengtsson-Palme J (2025) Applying 3D cultures and high-throughput technologies to study host-pathogen interactions. Front. Immunol. 16:1488699. doi: 10.3389/fimmu.2025.1488699
Received: 30 August 2024; Accepted: 04 February 2025;
Published: 20 February 2025.
Edited by:
Laurel J. Gershwin, University of California, Davis, United StatesReviewed by:
Qing Kang Lyu, La Jolla Institute for Immunology (LJI), United StatesGuan-Jun Yang, Ningbo University, China
Copyright © 2025 De Martinis, Alves, Pereira, Andrade, Abichabki, Abramova, Dannborg and Bengtsson-Palme. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Johan Bengtsson-Palme, am9oYW4uYmVuZ3Rzc29uLnBhbG1lQGNoYWxtZXJzLnNl
†These authors share first authorship