 Inga Tometten1*
Inga Tometten1* Tobias Brandt2
Tobias Brandt2 Maike Schlotz3Ricarda Stumpf3Sinje Landmann2
Maike Schlotz3Ricarda Stumpf3Sinje Landmann2 Marta Kantauskaite2Joshua Lamberti2Jonas Hillebrandt2
Marta Kantauskaite2Joshua Lamberti2Jonas Hillebrandt2 Lisa Müller1Margarethe Kittel2Katrin Ivens2,4
Lisa Müller1Margarethe Kittel2Katrin Ivens2,4 Henning Gruell3Anja Voges1Heiner Schaal1Nadine Lübke1
Henning Gruell3Anja Voges1Heiner Schaal1Nadine Lübke1 Eva Königshausen2,4
Eva Königshausen2,4 Lars Christian Rump2,4Florian Klein3,5,6
Lars Christian Rump2,4Florian Klein3,5,6 Johannes Stegbauer2,4†
Johannes Stegbauer2,4† Jörg Timm1†
Jörg Timm1†- 1Institute of Virology, Medical Faculty, University Hospital Düsseldorf, Heinrich-Heine-University Düsseldorf, Düsseldorf, Germany
- 2Department of Nephrology, Medical Faculty, University Hospital Düsseldorf, Heinrich-Heine-University Düsseldorf, Düsseldorf, Germany
- 3Laboratory of Experimental Immunology, Institute of Virology, Faculty of Medicine and University Hospital Cologne, University of Cologne, Cologne, Germany
- 4KfH Kuratorium für Dialyse und Nierentransplantation e.V., Düsseldorf, Germany
- 5Center for Molecular Medicine Cologne (CMMC), University of Cologne, Cologne, Germany
- 6German Center for Infection Research (DZIF), partner site Bonn-Cologne, Cologne, Germany
Background: The emergence of novel SARS-CoV-2 variants challenges immunity, particularly among immunocompromised kidney transplant recipients (KTRs). To address this, vaccines have been adjusted to circulating variants. Despite intensive vaccination efforts, SARS-CoV-2 infections surged among KTRs during the Omicron wave, enabling a direct comparison of variant-specific immunity following-vaccination against Omicron BA.4/5 or Omicron infection in KTRs.
Methods: 98 SARS-CoV-2 naïve KTRs who had received four vaccine doses were studied. Before and after a 5th antigen exposure, either via the bivalent vaccine composed of ancestral SARS-CoV-2 and Omicron BA.4/5 (29 KTRs) or via natural infection with Omicron (38 BA.4/5, 31 BA.1/2), spike-specific T cells were quantified using Elispot and serum pseudovirus neutralizing activity was assessed against the ancestral Wuhan strain, BA.5 and XBB.1.5.
Results: Compared to BA.4/5 vaccination, spike-specific T-cell responses and neutralization activity were higher up to six months post-Omicron infection and reached levels similar to healthy controls. Vaccinated KTRs showed modestly boosted neutralization activity against the Wuhan strain and BA.5, but not XBB.1.5. Baseline immunity correlated with immune responses three months post-vaccination and post-infection, indicating a predictive value for peak immune responses. Tixagevimab/Cilgavimab treatment was associated with robust neutralization of the Wuhan strain, but ineffective against XBB.1.5.
Conclusion: The BA.4/5 vaccine improved neutralizing activity against the BA.4/5 variant, but not against the subsequently circulating XBB.1.5 variant in KTRs. Conversely, omicron infection boosted T cells and humoral responses more effectively, showing efficacy against XBB.1.5. These findings suggest that infection-induced immunity associates with greater protection than vaccination against future variants in KTRs.
1 Introduction
The SARS-CoV-2 pandemic posed a challenge to global health with the constant emergence of new variants. In 2022, the omicron variant emerged and became predominant worldwide, associated with a massive increase in infection rates (1, 2). Because high population immunity was achieved and the overall medical burden declined, measures to contain the spread of SARS-CoV-2 were reduced in many countries. Since then, multiple distinct Omicron subvariants have emerged with distinct antigenic properties (3). In June 2022 the BA.5 variant became predominant in Germany until January 2023, when it was replaced by new variants such as XBB.1.5. mRNA vaccines offer the possibility of rapid adaptation to new variants. Shortly after the approval of vaccines adapted to the Omicron variant BA.1 (Comirnaty Original/Omicron BA.1 and Spikevax Bivalent Original/Omicron BA.1), Comirnaty Original/Omicron BA.4-5 (adapted to Omicron BA.4/BA.5) was approved by the EMA in September 2022. These adapted bivalent vaccines were then recommended as booster vaccinations.
Kidney transplant recipients (KTRs) are among the most vulnerable groups regarding severe COVID-19, even after vaccination (4). This is mainly due to an abrogated B- and T-cell response after SARS-CoV-2 vaccination compared to healthy individuals (5–10). Data from the vaccine trials with adapted mRNA vaccines have demonstrated superior variant-specific immunity with adapted vaccines compared to the original vaccine (11, 12). Accordingly, booster vaccination with the BA.4/5 bivalent vaccines was recommended for KTRs by the German Standing Vaccination Commission in September 2022. Because a substantial subgroup of KTRs had not responded to prior booster vaccinations with detectable and sustained immune responses, we sought to determine how KTRs respond to an adapted vaccine and if variant-specific immunity against the currently circulating variant BA.5 as well as one of the subsequently circulating variants (XBB.1.5) was induced. Interestingly, coinciding with the start of the vaccination campaign, the incidence of SARS-CoV-2 Omicron infections increased, opening up the opportunity to directly compare SARS-CoV-2 specific immunity after BA.4/5 vaccination with immunity after SARS-CoV-2 BA.5 infections in KTRs. Because it is expected that KTRs will be exposed to SARS-CoV-2 at some point, the impact of the infection on SARS-CoV-2 immunity and specifically the immunity against future variants is of great interest.
Here, we studied a cohort of KTRs exposed to a 5th SARS-CoV-2 antigen by either BA.4/5 vaccination or natural infection. SARS-CoV-2-specific T-cell and IgG levels were quantified and neutralizing activity against the original Wuhan strain, the BA.5 variant circulating at the time of the vaccination campaign and the XBB.1.5 variant following BA.5 was determined.
2 Methods
2.1 Study cohort
In this observational cohort study, KTRs who previously received four vaccine doses were included. The cohort is part of a previously described observational study (NCT04743947) of the Department of Nephrology, University Hospital Düsseldorf (7, 10, 13). We determined the immune response against SARS-CoV-2 before as well as 3 months and 6 months after their 5th antigen contact. Inclusion criteria were: age ≥ 18 years, kidney transplantation before the first SARS-CoV-2 vaccine dose, four prior vaccinations with any licensed COVID-19 vaccine (from AstraZeneca, Johnson & Johnson, Moderna or BioNTech), being SARS-CoV-2 naïve before the 5th antigen contact. KTRs were considered SARS-CoV-2 naïve when all of the following criteria applied: no documented positive SARS-CoV-2 test result, no self-reported positive SARS-CoV-2 antigen test, undetectable anti-nucleocapsid IgG and, when available, undetectable anti-nucleocapsid T cells before 5th antigen contact. The study cohort was divided into 2 groups: KTRs vaccinated with the bivalent BA.4/5-adapted mRNA vaccine as the 5th antigen contact (Vac); KTRs infected with SARS-CoV-2 as their 5th antigen contact (Inf). The group with infection was further subdivided into KTRs who were likely infected with BA.1 or BA.2 (Inf BA.1/2) and KTRs likely infected with BA.4 or BA.5 (Inf BA.4/5). When no viral sequence data were available, the most prevalent variant at the time of infection was considered. According to public health reports of SARS-CoV-2 variants, from January to May 2022 the variants BA.1 and BA.2 and from June to December 2022 the variant BA.5 were predominant (Supplementary Figure 1). To evaluate the humoral immune response, infected KTRs were further divided into patients receiving or not receiving monoclonal antibodies. As a reference group, vaccinated, immunocompetent, non-transplanted individuals (age ≥ 18 years) were included who were infected with SARS-CoV-2 between June and December 2022. Due to the complexity of comorbidities in the KTR cohort, matching for pre-existing conditions was not performed. B- and T- cell immune responses were examined before, 3 and 6 months after the 5th antigen contact when possible (Supplementary Table 1). All KTRs and healthy controls gave written informed consent and the study was approved by the ethics committee of the Medical Faculty of the Heinrich-Heine-University Düsseldorf, Germany (ID 2020-1237 and 2021-1316).
2.2 Pseudovirus neutralization assay
Neutralization was determined using an in-house pseudovirus neutralization assay as previously described with minor modifications (14). This assay is based on a protocol for pseudotyping lentiviral particles (15). In brief, pseudoviruses were produced in HEK293T cells cultured at 37°C/5% CO2 by cotransfection of plasmids encoding the SARS-CoV-2 spike protein, HIV-1 Gag/Pol, HIV-1 Tat, HIV-1 Rev, and Luciferase-IRES-ZsGreen, respectively. Following a medium exchange, pseudovirus supernatants in cell culture medium (high glucose DMEM supplemented with 100 IU/ml penicillin, 2 mM L-glutamine, 1 mM sodium pyruvate, 100 µg/ml streptomycin (all Thermo Fisher) and 10% FBS (Sigma-Aldrich)) were harvested at 48 h and/or 72 h post-transfection, centrifuged, purified (0.45 mm filter) and stored at -80°C.
Heat-inactivated serum samples (45 min at 56°C) were tested in three-fold serial dilutions starting at 1:10. Following a 60 min co-incubation of serum samples and pseudoviruses at 37°C and 5% CO2, 293T-ACE2 cells were added. After two days, cells were lysed using luciferin/lysis buffer (0.3 mM ATP, 10 mM MgCl2, 17 mM IGEPAL CA-630, 0.5 mM coenzyme A (all Sigma-Aldrich), and 1 mM D-Luciferin (GoldBio) in Tris-HCL) and bioluminescence was determined using a microplate reader (Berthold). Average background relative light units (RLUs) of non-infected cells were subtracted, and serum 50% inhibitory dilutions (ID50s) were determined as the serum dilutions that resulted in a 50% reduction of RLUs compared to the average of virus-infected untreated controls cells using a non-linear fit model plotting an agonist versus normalized dose response curve with variable slope using the least squares fitting method in Prism (GraphPad).
Serum samples were analyzed at a starting dilution of 1:10 and in seven 3-fold serial dilutions (i.e., 1:10, 1:30, 1:90, 1:270, 1:810, 1:2430, 1:7290, 1:21870). Dilution series were performed in duplicate or triplicate and the geometric mean ID50 for each sample was calculated using the two individually determined ID50s closest to each other. The limits of quantification were defined by the lowest and highest serum dilutions tested. In case of neutralization above the upper limit of quantification (ULOQ) in one of the two ID50 values (ID50 >21.870), the sample was assigned an ID50 value of 21.870. If both ID50 values were above the ULOQ, the sample was assigned an ID50 of 43.740 (2x ULOQ). In case of neutralization below the lower limit of quantification (LLOQ) in one of the two values (ID50 <10), the sample was assigned an ID50 of 10. If both ID50 values were below the LLOQ, the sample was assigned an ID50 of 5 (1/2 x LLOQ).
2.3 Quantification of SARS-CoV-2 specific T cells
The SARS-CoV-2-specific T-cell response was quantified by a commercially available standardized Interferon gamma (IFNγ) release assay (T.SPOT COVID Test from Oxford Immunotec, UK) according to the manufacturer’s instructions. In brief, whole blood was collected and stored at room temperature until further processing within 24 hours. T-cell Xtend reagent (Oxford Immunotec, UK) was added and mononuclear cells were separated through density gradient centrifugation. Cells were counted and 2.5 x 105 PBMCs were incubated for 20 hours with overlapping peptide pools covering the SARS-CoV-2 spike protein in the ELISPOT plate coated with anti-IFNγ. For each patient, analyses of negative (medium) and positive controls (phytohemagglutinine) were performed. On the next day, the plate was washed with PBS before IFNγ was stained. After the plate was dried, spots were counted using the BioReader 7000-E (Bio Sys, Germany). According to the manufacturer’s instructions <5 IFNγ spots (= spot forming units, SFU)/2.5 x 105 PBMCs were considered negative. 5-7 SFU/2.5 x 105 PBMCs were considered borderline and > 7 SFU/2.5 x 105 PBMCs were considered positive.
2.4 Quantification of SARS-CoV-2-specific antibodies
Antibody levels against SARS-CoV-2 spike protein (anti-S antibodies) were quantified in serum using the SARS-CoV-2 QuantiVac ELISA (Euroimmun, Germany). According to the manufacturer’s instructions, results <25.6 BAU/ml were considered negative, values between 25.6-35.2 BAU/ml as borderline and values >35.2 BAU/ml as positive.
2.5 Statistics
Statistical analyses were performed with GraphPad Prism 9. Non-parametric tests were used for comparison of quantitative results between groups as indicated in the figure legends. P values <0.05 were considered as statistically significant.
3 Results
3.1 Study cohort
A total of 102 KTRs with four documented vaccinations and no evidence of previous SARS-CoV-2 infection participated. Of these, 29 received the bivalent BA.4/5 vaccine as the 5th antigen contact and were included in the vaccination cohort (Vac). 73 patients got infected with SARS-CoV-2 as their 5th antigen contact. Samples from 35 patients were sequenced, resulting in 20 patients being assigned to the BA.1/2 cohort and 11 patients to the BA.4/5 cohort. 4 patients with other variants were excluded. Patients without SARS-CoV-2 sequencing data were assigned to the temporally predominant variant based on the period of their infection: 11 patients infected between January and May 2022 were assigned to the Omicron BA.1/BA.2 cohort (Inf BA.1/2), 27 patients infected between June and December 2022 were assigned to the Omicron BA.4/BA.5 cohort (Inf BA.4/5) (Figure 1). 24 of the 31 BA.1/2 infected patients and 22 of the 38 BA.4/5 infected patients received therapy with monoclonal antibodies (Table 1). None of the KTRs received Tixagevimab/Cilgavimab as pre-exposure prophylaxis. Regarding immunosuppression, 79% of the vaccinated cohort, 87% of the BA.1/2 infected cohort, and 68% of the BA.4/5 infected cohort received triple immunosuppression with mycophenolate mofetil (MMF), calcineurin inhibitors, and steroids. The remaining patients received other combinations. The MMF dosage and tacrolimus trough levels between the fourth and the fifth antigen exposure as a degree for the level of immunosuppression did not differ significantly between the three cohorts (Supplementary Figure 2). As infected patients presented with mild to moderate symptoms, the immunosuppressive treatment was not reduced during infection. In terms of vaccination history prior to the 5th antigen contact, KTRs were also comparable: Only 14%, 10%, and 18% of patients in the Vac, Inf BA.4/5, and Inf BA.1/2 cohorts had received a homologous vaccination scheme with mRNA-based vaccines only. The others had received at least one dose of a vector-based vaccine dose, either AstraZeneca or Johnson& Johnson (Table 1). 25 immunocompetent and non-transplanted individuals who had not yet received a bivalent vaccine but had a SARS-CoV-2 infection between June and December 2022 as their most recent antigen contact were included as controls, with 16 having received 3 vaccine doses before infection and 9 having been vaccinated 4 times.
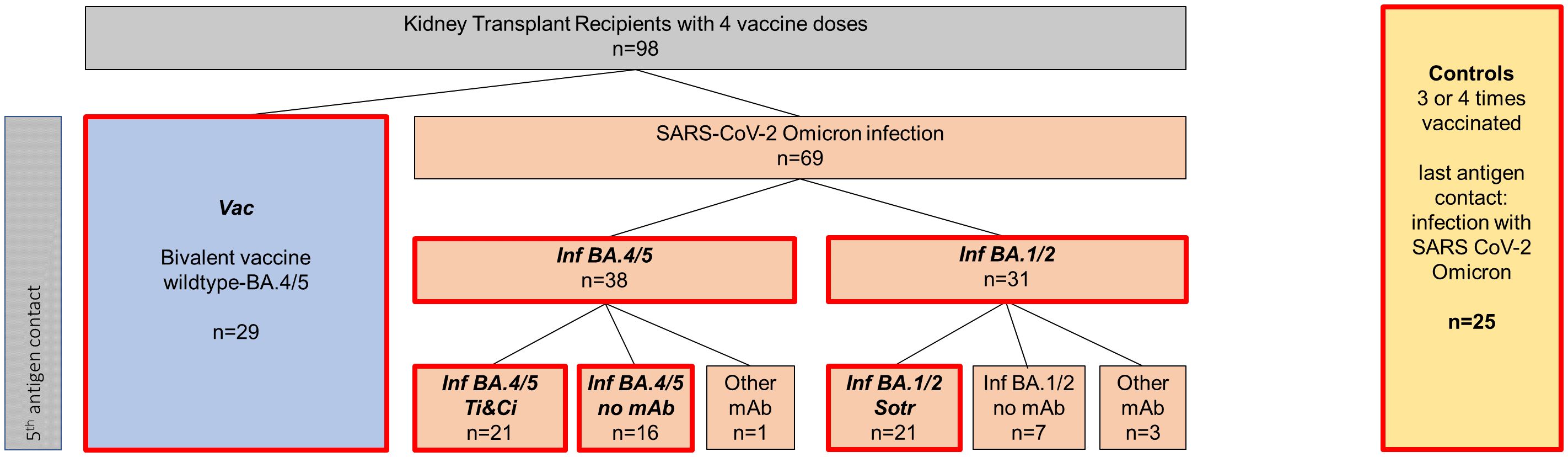
Figure 1. Classification of included patients into different cohorts. A total of 98 SARS-CoV-2 naive patients with four prior vaccinations were included. Of those, 29 patients received the wildtype/Omicron BA.4/5 bivalent vaccine as their fifth antigen exposure (highlighted in blue), referred to as “Vac” for “vaccinated kidney transplant recipients” throughout the manuscript. 69 patients were infected with SARS-CoV-2 Omicron as their fifth antigen exposure (represented in orange), with 38 putatively infected with Omicron BA.4/5 (Inf BA.4/5) and 31 putatively infected with Omicron BA.1/2 (Inf BA.1/2). 21 patients in the Inf BA.4/5 cohort received Tixagevimab/Calgevimab therapy, labeled “Inf BA.4/5 Ti&Ci” throughout the manuscript while 16 received no monoclonal antibody treatment and are classified as “ Inf BA.4/5 no mAb”. 21 BA.1/2 patients received Sotrovimab and are designated “ Inf BA.1/2 Sotr” Untreated BA.1/2 patients, as well as infected patients with other mAb than Tixagevimab/Cilgavimab or Sotrovimab were not analyzed separately due to small patient numbers. 25 healthy controls, who received 3-4 vaccine doses and who had a SARS-CoV-2 Omicron infection as their last antigen exposure, were included as a reference group.
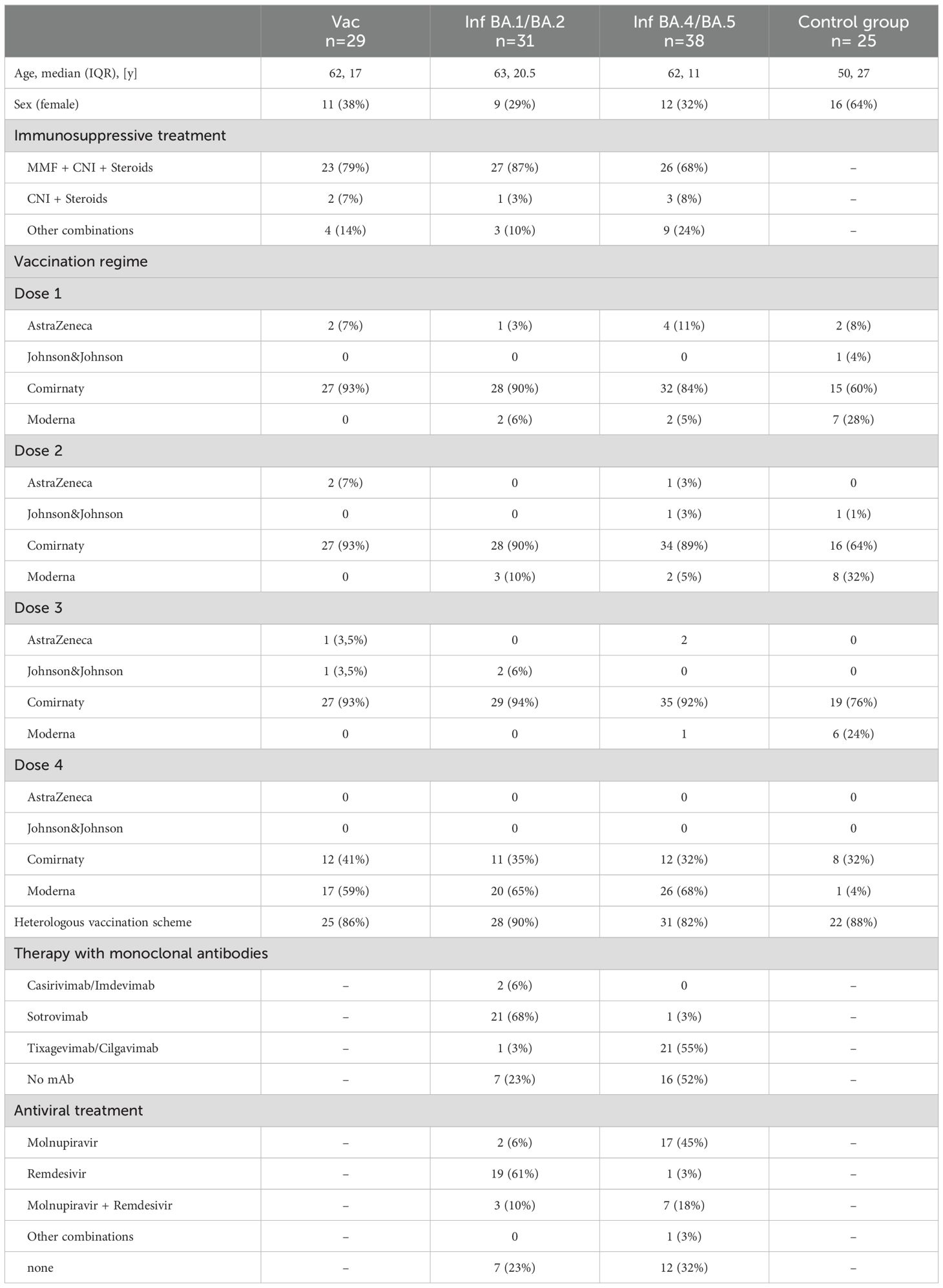
Table 1. Base line characteristics.
3.2 SARS-CoV-2 Omicron infection but not bivalent SARS-CoV-2 BA.4/5 vaccination boosts the T cell immune response in KTRs
The study compared the T-cell response to the SARS-CoV-2 spike protein in individuals who received the bivalent BA.4/5 vaccine with those who had contracted SARS-CoV-2 Omicron infection. Notably, KTRs with SARS-CoV-2 infection often received monoclonal antibody treatment (primarily Sotrovimab during the BA.1/2 wave and Tixagevimab/Cilgavimab during the BA.4/5 wave). The impact of monoclonal antibodies on SARS-CoV-2-specific T-cell immunity was therefore determined, revealing no significant difference in T-cell response between untreated infected KTRs and those treated with monoclonal antibodies (Supplementary Figure 3). Consequently, untreated and treated KTRs with SARS-CoV-2 infection were combined for T-cell response comparison. Following vaccination with the bivalent BA.4/5 vaccine, KTRs did not show improved T-cell responses at 3 and 6 months compared to pre-vaccination levels (Figure 2). Moreover, on a qualitative level, the proportion of KTRs with undetectable T-cell responses remained unchanged post-vaccination, with approximately half of KTRs still lacking detectable T-cell responses after the 5th vaccine dose. This was in stark contrast to KTRs infected with SARS-CoV-2 BA.1/2 or Omicron BA.4/5, where the T-cell response significantly increased at 3 and 6 months post-infection compared to pre-infection levels. In contrast to vaccinated KTRs, the proportion of KTRs with undetectable T-cell responses decreased to 20% after BA.4/5 infection. Although the number of KTRs with undetectable T-cell responses was slightly higher 6 months after infection compared to healthy controls, the frequency of spike-specific T cells reached similar levels. Taken together, the induction of SARS-CoV-2 spike-specific T cells was substantially more robust after SARS-CoV-2 Omicron infection compared to SARS-CoV-2 BA.4/5 vaccination.
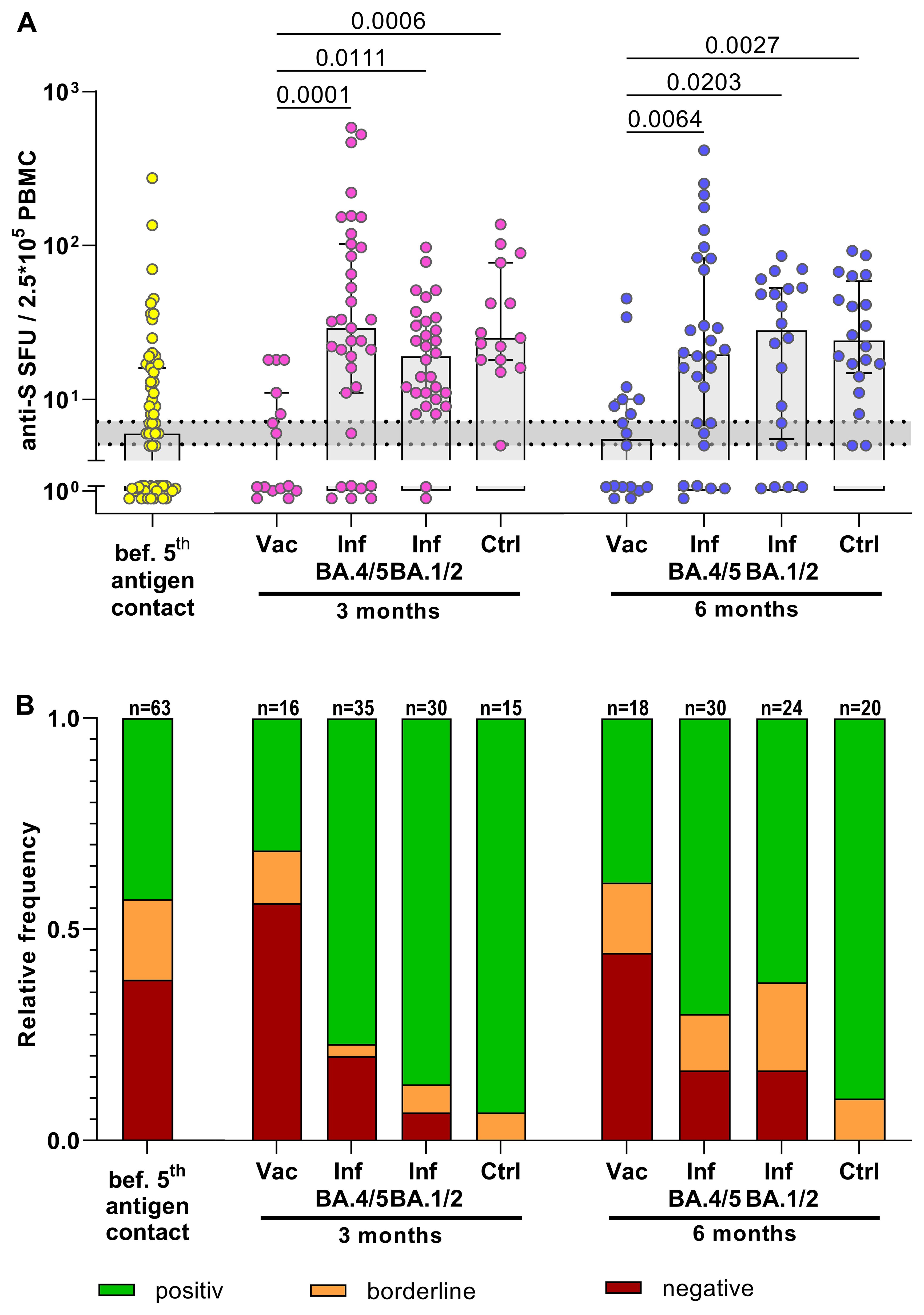
Figure 2. Boosted T-cell immune response in KTRs after SARS-CoV-2 Omicron infection but not after bivalent SARS-CoV-2 BA.4/5 vaccination. (A) Frequencies of anti-S specific T cells. Yellow: T-cell response in quadruple vaccinated KTRs before their 5th antigen contact. Rose/Blue: T-cell response in KTRs 3 months/6 months after their 5th antigen contact (BA.4/5 bivalent vaccine or BA.4/5 infection or BA.1/2 infection) compared to BA.4/5-infected healthy controls (Ctrl). An anti-S T-cell response >7 SFUs/2.5*105 PBMCs was considered as positive, <5 SFUs/2.5*105 PBMCs as negative. The range of borderline responses (5-7 SFUs/2.5*105 PBMCs) is marked in grey. (B) Corresponding relative frequencies of a qualitative positive, borderline or negative anti-S specific T-cell response. For statistical analysis, Mann-Whitney test or Kruskal-Wallis-Test followed by Dunn’s post-hoc test was applied. Boxes and bars represent median and interquartile range.
3.3 SARS-CoV-2 Omicron BA.4/5 infection induces stronger neutralization activity against BA.5 and XBB.1.5 than BA.4/5 bivalent vaccination in KTRs
To compare SARS-CoV-2-specific antibodies and neutralization activity, SARS-CoV-2-infected KTRs treated with monoclonal antibodies were analyzed separately from untreated infected KTRs. This distinction was necessary because the assays cannot differentiate between the patient’s own humoral response and therapeutically applied antibodies as long as the viral strain in use is not completely resistant to the administered antibody. Three months after the 5th vaccination, KTRs exhibited a 9.3-fold increase in the median anti-S IgG titer compared to pre-vaccination levels (Figure 3). However, this anti-S IgG titer (591 BAU/ml, IQR 2227 BAU/ml) remained lower than that in BA.4/5-infected KTRs (1143 BAU/ml, IQR 2478.7 BAU/ml) who had not received mAb therapy. As anticipated, SARS-CoV-2-infected KTRs treated with Tixagevimab/Cilgavimab or Sotrovimab showed higher levels of anti-S antibodies, similar to those observed after infection in healthy controls.
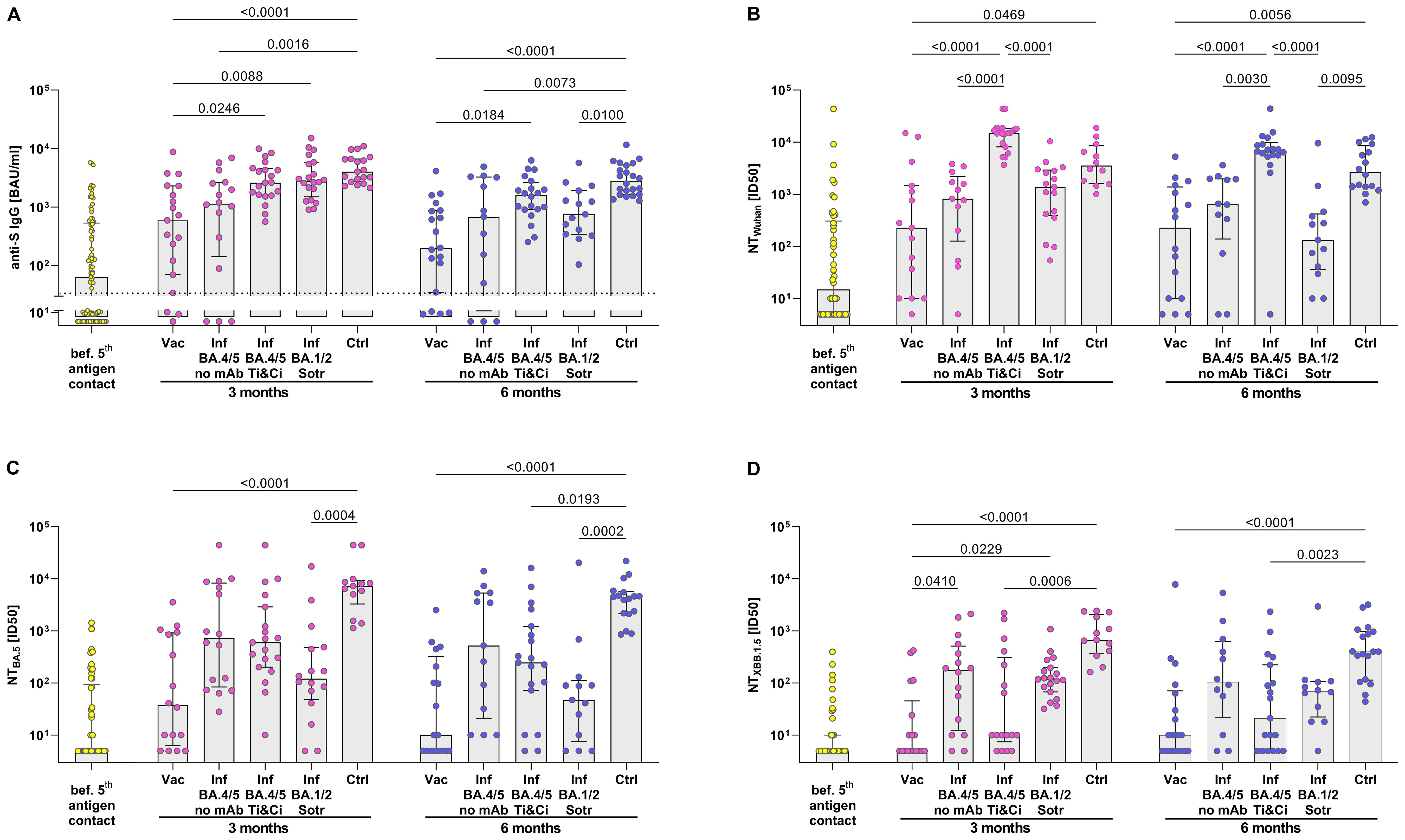
Figure 3. Stronger neutralization activity against BA.5 and XBB.1.5 in KTRs after SARS-CoV-2 Omicron BA.4/5 infection than after BA.4/5 bivalent vaccination. (A) Anti-S IgG Titers and (B–D) serum neutralization activity against Wuhan, Omicron BA.4/5 and Omicron XBB.1.5 pseudoviruses were determined. Yellow: B cell response in quadruple vaccinated KTRs before their 5th antigen contact. Rose/Blue: B cell response in KTRs 3 months/6 months after their 5th antigen: BA.4/5 bivalent vaccine (Vac) or BA.4/5 infection without treatment with monoclonal antibodies (Inf BA.4/5) or BA.4/5 infection receiving Tixagevimab/Cilgavimab (Inf BA.4/5 Ti&Ci) or BA.1/2 infection receiving Sotrovimab (Inf BA.1/2 Sotr) compared to BA.4/5 infected healthy controls (Ctrl). For statistical analysis, Kruskal-Wallis-Test followed by Dunn’s post-hoc test was applied. Boxes and bars represent median and interquartile range.
Neutralization activity post-vaccination and infection was assessed using pseudoviruses for the original Wuhan strain, the Omicron BA.5 variant, and the Omicron XBB.1.5 variant. At 3 months post-vaccination, there was a modest increase in neutralization activity against the Wuhan strain and the BA.5 variant, but no enhancement in neutralization of the XBB.1.5 variant. This suggests that the bivalent BA.4/5 vaccine boosted the antibody response against the original prototype and the BA.5 variant, but not against the subsequently circulating XBB.1.5 variant. This stands in contrast to KTRs with SARS-CoV-2 BA.4/5 infection, where neutralization was augmented against the Wuhan strain as well as both Omicron variants, including the XBB.1.5 variant. Consequently, at 3 and 6 months post-infection, neutralization activity against the three tested SARS-CoV-2 variants was higher in infected KTRs compared to vaccinated KTRs. Notably, the neutralization activity in infected KTRs almost reached levels comparable to healthy controls.
Treating infected KTRs with Tixagevimab/Cilgavimab or Sotrovimab also provided an opportunity to examine the long-term neutralization levels achieved with these monoclonal antibodies in vivo. At 3 and 6 months post-infection, KTRs treated with Tixagevimab/Cilgavimab exhibited significantly greater neutralization activity against the Wuhan strain compared to those treated with Sotrovimab. However, when it came to neutralizing the BA.5 and XBB.1.5 variants, there was no significant difference in activity between KTRs treated with Tixagevimab/Cilgavimab or Sotrovimab and those not treated with monoclonal antibodies. Interestingly, although not statistically significant, neutralization of the XBB.1.5 variant even tended to be lower in KTRs treated with Tixagevimab/Cilgavimab compared to untreated KTRs.
In summary, analyzing antibody levels in KTRs post-exposure to a 5th antigen suggests overall higher neutralization activity after SARS-CoV-2 Omicron infection compared to bivalent BA.4/5 vaccination, including improved neutralization of the subsequently circulating XBB.1.5 variant.
3.4 Immunity prior to the 5th antigen contact correlates with the level of immunity after vaccination or infection in KTRs
We previously noted that among various factors, pre-existing immunity prior to vaccination predicts the immune response post-vaccination in KTRs (10). However, it remains uncertain whether this holds true in the context of infection. To explore this, we correlated neutralization activities against the Wuhan strain, the BA.5 variant, and the XBB.1.5 variant before and 3 months after the 5th antigen exposure (Figure 4). Neutralization of the Wuhan strain and BA.5 variant before vaccination strongly correlated with neutralization 3 months after the bivalent BA.4/5 vaccine (r=0.9785; p<0.0001 and r=0.9289; p<0.0001). However, the correlation for neutralization of the XBB.1.5 variant was less robust and not significant (r=0.4966; p<0.0615), likely due to overall low levels of neutralization. Moreover, there was a significant correlation between neutralization of the Wuhan strain and the BA.5 variant before and 3 months after infection (r=0.7379; p=0.0197 and r=0.7556; p=0.0027), although the correlation coefficient was weaker compared to that after vaccination. Notably, in the context of KTR infection, neutralization of the XBB.1.5 variant before the 5th antigen exposure strongly correlated with levels 3 months afterward. Taken together, these findings suggest that similar to immunity induced by vaccination, the level of humoral immune response prior to infection can serve as a predictor for the response level after infection in KTRs.
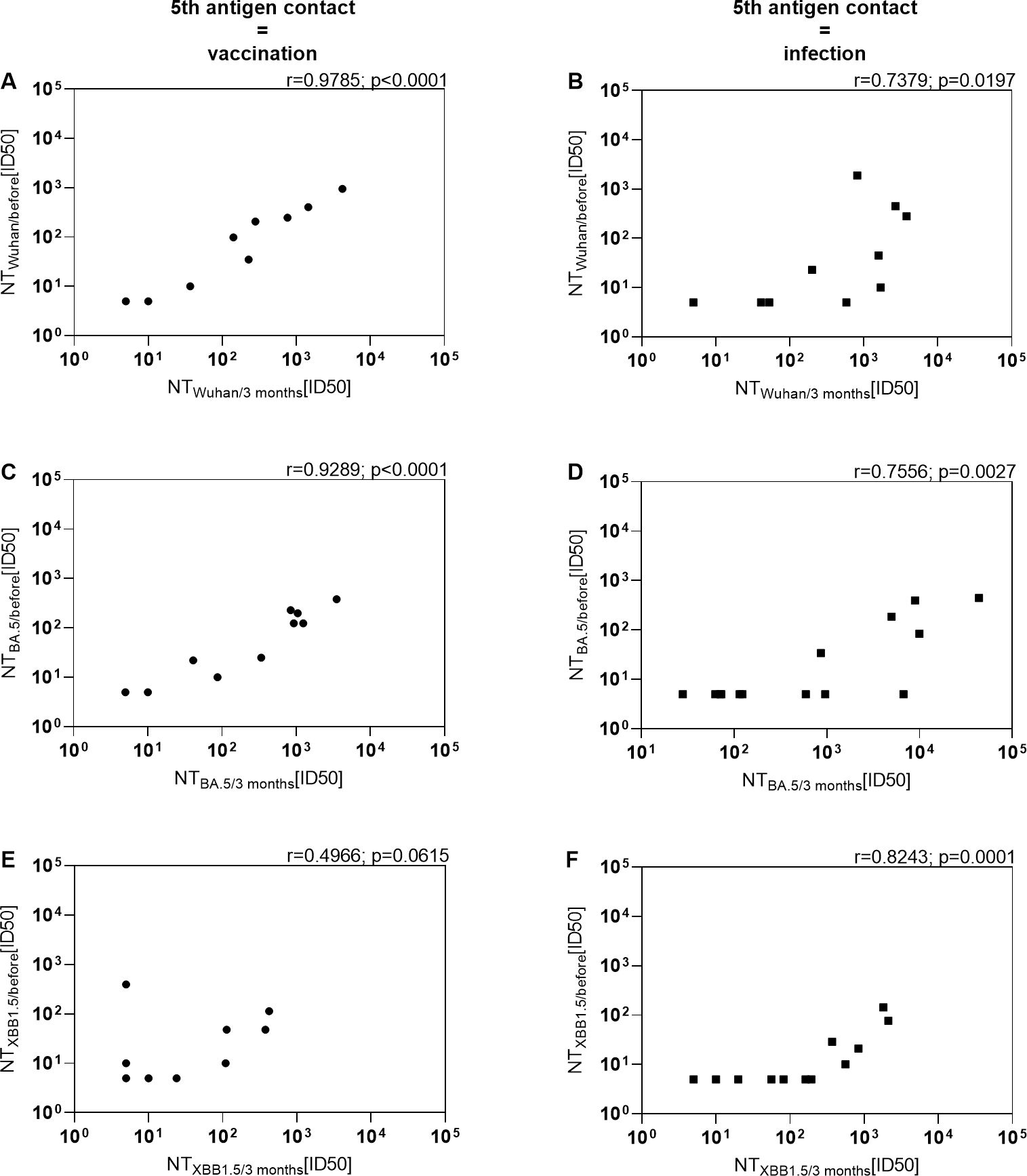
Figure 4. Positive correlation between level of immunity prior to the 5th antigen contact and level of immunity after vaccination or infection in KTRs. Serum neutralization activity against (A, B) Wuhan, (C, D) Omicron BA.5 and (E, F) Omicron XBB.1.5 before and 3 months after BA.4/5 bivalent vaccination or BA.4/5 infection were correlated. Left: KTRs with the bivalent BA.4/5 vaccination as their 5th antigen contact. Right: KTRs with an Omicron BA.4/5 infection as their 5th antigen contact without treatment with monoclonal antibodies.
4 Discussion
Population immunity has substantially increased after vaccination campaigns have been implemented and was further promoted by natural SARS-CoV-2 infections, leading to a lower incidence of severe COVID-19 and significantly declined overall mortality (16–18). Protective measures such as contact bans or mandatory use of protective masks, predominantly implemented for protection of patient groups at high risk for severe COVID-19, were continuously lifted. At the same time, the immune evasive Omicron variant emerged and rapidly spread globally. Although the number of severe cases remained low, this resulted in unprecedented high SARS-CoV-2 infection rates as demonstrated in the highest peak concentrations of SARS-CoV-2 RNA in late 2023 since the beginning of waste water surveillance in Germany (19). It is expected that also vulnerable patient groups such as KTRs will be exposed to SARS-CoV-2 at some point. Accordingly, these groups continue to be a prime target of the vaccination strategy, in which vaccines are used that have been adapted to circulating SARS-CoV-2 variants.
In this study, the immune response to SARS-CoV-2 following vaccination with bivalent vaccines adapted to the BA.4/5 variants was directly compared with immunity post-SARS-CoV-2 Omicron infection in KTRs. The objective was to characterize immunity against presently circulating variants and potential future variants in immunocompromised KTRs. To facilitate a direct comparison, only KTRs who experienced their 5th antigen encounter either through vaccination or SARS-CoV-2 infection were included. One notable strength of the study lies in its prospective nature, with analyses conducted on samples taken before vaccination and infection, respectively. This was made possible by utilizing an observational cohort study of KTRs that was initiated at the beginning of vaccination campaigns (7, 10, 13, 20), thereby ensuring comprehensive characterization of the immune response in all participants before the 5th antigen contact.
A central finding was lack or only weak induction of T-cell responses by vaccination. This was in line with previous data on booster immunization with the standard vaccines in this cohort (10) and can now be extended to an adapted vaccine (21, 22). Of note, the peptides used for the ELISpot assay covered the SARS-CoV-2-spike protein of the Wuhan strain and were not adapted to the Omicron variant, however, it has been described that the differences in peptide sequences have no or only a minor effect on quantification of SARS-CoV-2 specific T cells (23–25). The data suggest that in KTRs activation and expansion of SARS-CoV-2 spike-specific T cells continues to be challenging even with mRNA vaccines, especially in those KTRs who also did not respond well to previous vaccines (26, 27). This is in contrast to SARS-CoV-2 Omicron infection, which led to a robust expansion of SARS-CoV-2 spike-specific T cells at 3 and 6 months post-infection. Similar results have also been observed in non-immunocompromised individuals, with stronger T-cell responses after breakthrough infection compared to fully vaccinated individuals (25).
Consistent with the literature, the 5th vaccination with the bivalent BA.4/5 specific vaccine led to an increase in the anti-Spike IgG titers (28, 29). 3 months post-vaccination, anti-S IgG titers did not significantly differ between vaccinated and infected KTRs, underlying a quantitative robust B cell immune response after vaccination. In addition, 93% of the vaccinated cohort showed detectable neutralization activity against the Wuhan strain. In contrast, the neutralization activity was significantly reduced against the Omicron variant BA.5. which is in line with the previously reported immune evasive properties of this variant. After the first appearance of Omicron, continuous selection of immune evasive subvariants lead to the emergence and spread of XBB.1.5 in January 2023 after the vaccination campaigns had started with the adapted vaccines containing Omicron BA.4/5. The XBB.1.5 associated with a further decrease in neutralization activity (28, 30–32), which was confirmed here in our cohort of KTRs after the 5th vaccination. Collectively, our results suggest that a booster effect for virus neutralization was limited to the original Wuhan strain and to some extent to the BA.5 strain justifying adaption of the vaccine. However, in our cohort there was no significant booster effect on neutralizing activity on the serum level against the XBB.1.5 variant as a following emerging variant.
In turn, after BA.4/5 infection, peak neutralization activity was high and achieved levels 21.5-fold higher against BA.5 and 53-fold higher against XBB.1.5. compared to KTRs pos-vaccination. This strongly supports previous observations, that hybrid immunity induced by a combination of vaccination and infection is more robust and associates with broader neutralization capacity than vaccine-induced immunity (33–35). In KTRs this seems to be true for the humoral response as well as the T-cell response. Importantly, in this setting, hybrid immunity by vaccination and infection with the BA.5 variant also showed higher neutralization of the XBB.1.5 as the next following variant circulating in the population. As MMF may dose-dependently impair humoral immunity (20), lower immunosuppression in the infected cohort might theoretically account for their enhanced immune response. However, immunosuppression was not reduced in the infected cohort because infections were mild-to-moderate, and dosage of MMF and trough levels of Tacrolimus were comparable between vaccinated and infected cohorts. Additionally, absolute anti-Spike IgG titers did not differ significantly between vaccinated and mAb-untreated infected KTRs. Thus, different levels of immunosuppression is unlikely to account for the observed differences in humoral immune responses between KTR groups. Although SARS-CoV-2 infections in vaccinated KTRs unfortunately still come with the risk of severe disease, they seem inevitable when the infection pressure in the population is high as has been observed during the season 2022/23. In these cases, infection associated immunity may strongly contribute to protection and also promotes immunity against future variants, which seems difficult to achieve in immunosuppressed KTRs by vaccination only.
Finally, the application of monoclonal antibodies as SARS-CoV-2 treatment in many KTRs opened up the opportunity to study the neutralizing levels achieved in vivo against the different variants. Although the mAb combination Tixagevimab/Cilgavimab was clearly associated with a high neutralizing activity against the Wuhan strain in serum over a period of at least 6 months, there was only limited activity against the BA.5 variant and no activity against the XBB.1.5 variant. This was expected as Tixagevimab/Cilgavimab has been described to be not effective against XBB.1.5 variant in healthy patients treated with these mAbs as pre-exposure prophylaxis and because XBB.1.5 is resistant to Tixagevimab/Cilgavimab in vitro (36, 37). Interestingly, 10 of 17 Tixagevimab/Cilgavimab-treated KTRs (59%) showed no relevant neutralizing activity (<=10) against XBB.1.5 compared to 24% in the mAb-untreated cohort (4 of 17). Although not conclusive, a negative effect of Tixagevimab/Cilgavimab on the induction of novel variant-specific antibodies by the host should be considered. Recent data suggest, that the administration of monoclonal antibodies has an effect on memory B cell selection (38, 39). Nevertheless, the possible negative consequences of therapeutically applied antibodies lacking activity against the relevant variant on the development of an autologous humoral response need further investigation. An alternative explanation for weaker neutralization of XBB.1.5 in the Tixagevimab/Cilgavimab-treated cohort compared to mAb-untreated KTRs may be a potential selection bias, as only those KTRs who sought medical attention at the hospital received treatment with Tixagevimab/Cilgavimab. A possible negative impact on neutralization activity was not observed for Sotrovimab-treated BA.1/2 infected KTRs. This is in line with existing data, reporting some residual neutralization activity against XBB.1.5 in Sotrovimab-treated patients in contrast to Tixagevimab/Cilgavimab treated patients (40, 41). Nevertheless, in the context of continuous evolution of subsequent variants with even higher immune evasive properties, both Tixagevimab/Cilgavimab and Sotrovimab are not generally recommended any longer (42).
There are few limitations to the study. Data were analysed pseudo longitudinally instead of longitudinally, due to the observational nature of this study, which integrated patients from routine clinical visits without consistent sampling at all time points. This led to incomplete data across cohorts, especially in the BA.1/2-infected group, where 25 of 31 KTRs missed evaluations between the fourth and fifth antigen exposures. At that stage, all KTRs had received four vaccinations and were SARS-CoV-2 naive, ensuring equivalent antigen exposure, with cohort differentiation only occurring at the fifth exposure. As we have almost complete data for vaccinated and BA.4/5-infected cohorts at this time, we believe the missing BA.1/2 data did not significantly impact our findings. Considering the comparability of the cohorts, it is notable that the control group was slightly younger than the KTRs (p = 0.011). However, the control group was included as a reference to contextualize the immune response of the KTRs. No conclusions were drawn based on direct comparisons between healthy controls and KTRs. Hence, the age difference of the control group was not considered to be a problem when immune responses of vaccinated KTRs and infected KTRs were compared. Furthermore, the higher proportion of women in the control group should be noted, as sex-specific differences in the immune response exist, which are particularly complex and thus are not further discussed here (43). The exact vaccination histories regarding the specific vaccines of the included patients are heterogeneous. However, this represents the real-world scenario, as the choice of vaccine was influenced by external factors such as vaccine availability, which changed over time with the approval of new vaccines. Here, 86% of the vaccinated KTRs, 90% of the BA.1/2 infected KTRs, and 82% of the BA.4/5 infected KTRs had a prior vaccination history with at least one dose of a vector-based vaccine. Thus, the patient groups appear roughly comparable in this regard. Furthermore, the exact SARS-CoV-2 genotype was determined only for the minority of the infected patients. Therefore, the most likely genotype was inferred from data on the circulating variants in Germany published by the public health authorities. The inclusion of infected KTRs as BA.4/5 or BA.1/2 infected KTRs was based on the time of infection and the predominant variant at that time (BA.4/5 between June and December 2022 and BA.1/2 between January to May 2022). This represents the real-life scenario when therapeutic decisions are based on the predominant variants. Because the distribution of variants was overall rather homogenous during that time, we believe that inference of the genotype base on epidemiological data is a reasonable proxy. Importantly immunity induced by infections with the XBB.1.5 variant can be excluded, because this variant was not circulating in Germany during that time. In the infected cohort, variations in treatment approaches, such as the use of different mAbs, present an additional limitation to the study, as they may have influenced the humoral immune response differently. Treatment data with mAbs was available for all infected patients, allowing us to compare groups according to the monoclonal antibody therapy. Importantly, we were also able to include an untreated BA.4/5 infected cohort. Therefore, we believe this limitation does not compromise the overall validity of our data.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Ethics statement
The studies involving humans were approved by ethics committee of the Medical Faculty of the Heinrich-Heine-University Düsseldorf, Germany ID 2020-1237 and 2021-1316. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
IT: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing – original draft. TB: Formal analysis, Writing – review & editing, Investigation. MS: Formal analysis, Writing – review & editing, Investigation. RS: Formal analysis, Writing – review & editing, Investigation. SL: Formal analysis, Writing – review & editing, Investigation. MKi: Formal analysis, Writing – review & editing, Investigation. JL: Writing – review & editing, Investigation. JH: Writing – review & editing, Investigation. LM: Data curation, Formal analysis, Investigation, Writing – review & editing. MKa: Writing – review & editing, Investigation. KI: Investigation, Writing – review & editing. HG: Data curation, Formal analysis, Investigation, Writing – review & editing. AV: Investigation, Writing – review & editing. HS: Investigation, Writing – review & editing. NL: Investigation, Writing – review & editing. EK: Investigation, Writing – review & editing. LR: Investigation, Writing – review & editing. FK: Data curation, Formal analysis, Investigation, Supervision, Writing – review & editing. JS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Supervision, Writing – review & editing, Writing – original draft. JT: Conceptualization, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing, Data curation, Formal analysis, Investigation.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The study was supported by funds of the Ministry of Labour, Health and Social Affairs of North-Rhine Westphalia (CPS-1-1A), the Federal Ministry of Education and Research (01KX2021) and by the VIRus Alliance NRW (VIRAL) of the Ministry of Culture and Science of the state of North Rhine-Westphalia (323-8.03-151826).
Acknowledgments
We would like to thank all patients for participating in this observational cohort study. We thank Finn Teipel for providing the XBB.1.5 pseudovirus plasmid. The data were partially presented as a poster presentation at the 2024 Annual Meeting of the ESCV.
Conflict of interest
HG and FK are inventors on patent applications on SARS-CoV-2 neutralizing antibodies filed by the University of Cologne and have received payments from the University of Cologne for licensed antibodies. LK received consulting fees or payment/honoraria from ViiV, Theratechnologies, Gilead and MSD.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1476294/full#supplementary-material
References
1. Cao Y, Wang J, Jian F, Xiao T, Song W, Yisimayi A, et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature. (2022) 602:657–63. doi: 10.1038/s41586-021-04385-3
2. Viana R, Moyo S, Amoako DG, Tegally H, Scheepers C, Althaus CL, et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature. (2022) 603:679–86. doi: 10.1038/s41586-022-04411-y
3. Chatterjee S, Bhattacharya M, Nag S, Dhama K, Chakraborty C. A detailed overview of SARS-CoV-2 omicron: its sub-variants, mutations and pathophysiology, clinical characteristics, immunological landscape, immune escape, and therapies. Viruses. (2023) 15(1):167. doi: 10.3390/v15010167
4. Caillard S, Thaunat O. COVID-19 vaccination in kidney transplant recipients. Nat Rev Nephrol. (2021) 17:785–7. doi: 10.1038/s41581-021-00491-7
5. Zhang R, Shin BH, Gadsden TM, Petrosyan A, Vo A, Ammerman N, et al. Assessment of humoral and cellular immune responses to SARS CoV-2 vaccination (BNT162b2) in immunocompromised renal allograft recipients. Transpl Infect Dis. (2022) 24:e13813. doi: 10.1111/tid.13813
6. Udomkarnjananun S, Gatechompol S, Leelahavanichkul A, Kerr SJ. Cellular immune response of SARS-CoV-2 vaccination in kidney transplant recipients: a systematic review and meta-analysis. Front Immunol. (2023) 14:1220148. doi: 10.3389/fimmu.2023.1220148
7. Kolb T, Fischer S, Muller L, Lubke N, Hillebrandt J, Andree M, et al. Impaired immune response to SARS-CoV-2 vaccination in dialysis patients and in kidney transplant recipients. Kidney360. (2021) 2:1491–8. doi: 10.34067/KID.0003512021
8. Werbel WA, Karaba AH, Chiang TP, Massie AB, Brown DM, Watson N, et al. Persistent SARS-CoV-2-specific immune defects in kidney transplant recipients following third mRNA vaccine dose. Am J Transpl. (2023) 23:744–58. doi: 10.1016/j.ajt.2023.03.014
9. Hamaya T, Hatakeyama S, Yoneyama T, Tobisawa Y, Kodama H, Fujita T, et al. Seroprevalence of SARS-CoV-2 spike IgG antibodies after the second BNT162b2 mRNA vaccine in Japanese kidney transplant recipients. Sci Rep. (2022) 12:5876. doi: 10.1038/s41598-022-09897-0
10. Tometten I, Landmann S, Kantauskaite M, Lamberti J, Hillebrandt J, Muller L, et al. Factors associated with vaccine-induced T-cell immune responses against severe acute respiratory syndrome Coronavirus 2 in kidney transplant recipients. J Infect Dis. (2023) 227:641–50. doi: 10.1093/infdis/jiac449
11. Usdan L, Patel S, Rodriguez H, Xu X, Lee DY, Finn D, et al. A Bivalent Omicron-BA.4/BA.5-Adapted BNT162b2 Booster in >/=12-Year-Olds. Clin Infect Dis. (2023) 78(5):1194,203. doi: 10.1093/cid/ciad718
12. Chalkias S, Harper C, Vrbicky K, Walsh SR, Essink B, Brosz A, et al. A bivalent omicron-containing booster vaccine against Covid-19. N Engl J Med. (2022) 387:1279–91. doi: 10.1056/NEJMoa2208343
13. Kantauskaite M, Muller L, Hillebrandt J, Lamberti J, Fischer S, Kolb T, et al. Immune response to third SARS-CoV-2 vaccination in seronegative kidney transplant recipients: Possible improvement by mycophenolate mofetil reduction. Clin Transpl. (2022) 36:e14790. doi: 10.1111/ctr.v36.11
14. Vanshylla K, Di Cristanziano V, Kleipass F, Dewald F, Schommers P, Gieselmann L, et al. Kinetics and correlates of the neutralizing antibody response to SARS-CoV-2 infection in humans. Cell Host Microbe. (2021) 29:917–29.e4. doi: 10.1016/j.chom.2021.04.015
15. Crawford KHD, Eguia R, Dingens AS, Loes AN, Malone KD, Wolf CR, et al. Protocol and reagents for pseudotyping lentiviral particles with SARS-CoV-2 spike protein for neutralization assays. Viruses. (2020) 12(5):513. doi: 10.3390/v12050513
16. Huang C, Yang L, Pan J, Xu X, Peng R. Correlation between vaccine coverage and the COVID-19 pandemic throughout the world: Based on real-world data. J Med Virol. (2022) 94:2181–7. doi: 10.1002/jmv.27609
17. Mohammed I, Nauman A, Paul P, Ganesan S, Chen KH, Jalil SMS, et al. The efficacy and effectiveness of the COVID-19 vaccines in reducing infection, severity, hospitalization, and mortality: a systematic review. Hum Vaccin Immunother. (2022) 18:2027160. doi: 10.1080/21645515.2022.2027160
18. Hovd M, Asberg A, Munthe LA, Heldal K, Reisaeter AV, Vaage JT, et al. Humoral vaccine response and breakthrough infections in kidney transplant recipients during the COVID-19 pandemic: a nationwide cohort study. EClinicalMedicine. (2023) 60:102035. doi: 10.1016/j.eclinm.2023.102035
19. Umweltbundesamt, Robert Koch-Institut. Abwassersurveillance zu SARS-CoV-2(2024). Available online at: https://www.rki.de/DE/Content/Institut/OrgEinheiten/Abt3/FG32/Abwassersurveillance/Bericht_Abwassersurveillance.html?:blob=publicationFile (accessed April 24, 2024).
20. Kantauskaite M, Muller L, Kolb T, Fischer S, Hillebrandt J, Ivens K, et al. Intensity of mycophenolate mofetil treatment is associated with an impaired immune response to SARS-CoV-2 vaccination in kidney transplant recipients. Am J Transpl. (2022) 22:634–9. doi: 10.1111/ajt.16851
21. Solera JT, Ierullo M, Arbol BG, Mavandadnejad F, Kurtesi A, Qi F, et al. Bivalent COVID-19 mRNA vaccine against omicron subvariants in immunocompromised patients. Lancet Infect Dis. (2023) 23:e266–e7. doi: 10.1016/S1473-3099(23)00357-2
22. Fernandez-Ruiz M, Almendro-Vazquez P, Redondo N, Ruiz-Merlo T, Abella S, Somoza A, et al. Cell-mediated and neutralizing antibody responses to the SARS-CoV-2 omicron BA.4/BA.5-adapted bivalent vaccine booster in kidney and liver transplant recipients. Transplant Direct. (2023) 9(10):e1536. doi: 10.1097/TXD.0000000000001536
23. Tarke A, Coelho CH, Zhang Z, Dan JM, Yu ED, Methot N, et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell. (2022) 185:847–59.e11. doi: 10.1016/j.cell.2022.01.015
24. Keeton R, Tincho MB, Ngomti A, Baguma R, Benede N, Suzuki A, et al. T cell responses to SARS-CoV-2 spike cross-recognize Omicron. Nature. (2022) 603:488–92. doi: 10.1038/s41586-022-04460-3
25. Collier AY, Brown CM, McMahan KA, Yu J, Liu J, Jacob-Dolan C, et al. Characterization of immune responses in fully vaccinated individuals after breakthrough infection with the SARS-CoV-2 delta variant. Sci Transl Med. (2022) 14:eabn6150. doi: 10.1126/scitranslmed.abn6150
26. Yang J, Lee KW, Baek JY, Bae S, Lee YH, Kim H, et al. Augmented humoral and cellular immunity against severe acute respiratory syndrome coronavirus 2 after breakthrough infection in kidney transplant recipients who received 3 doses of coronavirus disease 2019 vaccine. Am J Transpl. (2023) 23:565–72. doi: 10.1016/j.ajt.2022.12.022
27. Rezahosseini O, Hamm SR, Heftdal LD, Perez-Alos L, Moller DL, Perch M, et al. Humoral and T-cell response 12 months after the first BNT162b2 vaccination in solid organ transplant recipients and controls: Kinetics, associated factors, and role of SARS-CoV-2 infection. Front Immunol. (2022) 13:1075423. doi: 10.3389/fimmu.2022.1075423
28. Pedersen RM, Bang LL, Holm DK, Madsen LW, Johansen IS, Jensen TG, et al. Serum neutralization of omicron XBB.1.5 in kidney transplant recipients after bivalent mRNA booster vaccination. Kidney Int Rep. (2023) 8:1665–8. doi: 10.1016/j.ekir.2023.05.020
29. Hirotsu Y, Sugiura H, Takatori M, Mochizuki H, Omata M. Antibody Response to the BA.5 Bivalent Vaccine Shot: a Two-Year Follow-Up Study following Initial COVID-19 mRNA Vaccination. Microbiol Spectr. (2023) 11(3):e0131623. doi: 10.1128/spectrum.01316-23
30. Amano M, Otsu S, Uemura Y, Ichikawa Y, Matsumoto S, Higashi-Kuwata N, et al. Neutralization against Omicron sublineages (BA.2/BA.5/BQ.1.1/XBB/XBB.1.5) in bivalent BNT162b2-vaccinated HCWs with or without risk factors, or following BT infection with Omicron. Sci Rep. (2023) 13:17404. doi: 10.1038/s41598-023-44484-x
31. Qu P, Faraone JN, Evans JP, Zheng YM, Carlin C, Anghelina M, et al. Extraordinary evasion of neutralizing antibody response by omicron XBB.1.5, CH.1.1 and CA.3.1 variants. bioRxiv. (2023) 42(5):112443. doi: 10.1101/2023.01.16.524244
32. Yau K, Kurtesi A, Qi F, Delgado-Brand M, Tursun TR, Hu Q, et al. Omicron variant neutralizing antibodies following BNT162b2 BA.4/5 versus mRNA-1273 BA.1 bivalent vaccination in patients with end-stage kidney disease. Nat Commun. (2023) 14:6041. doi: 10.1038/s41467-023-41678-9
33. Bobrovitz N, Ware H, Ma X, Li Z, Hosseini R, Cao C, et al. Protective effectiveness of previous SARS-CoV-2 infection and hybrid immunity against the omicron variant and severe disease: a systematic review and meta-regression. Lancet Infect Dis. (2023) 23:556–67. doi: 10.1016/S1473-3099(22)00801-5
34. Andreano E, Paciello I, Piccini G, Manganaro N, Pileri P, Hyseni I, et al. Hybrid immunity improves B cells and antibodies against SARS-CoV-2 variants. Nature. (2021) 600:530–5. doi: 10.1038/s41586-021-04117-7
35. Lasrado N, Barouch DH. SARS-CoV-2 hybrid immunity: the best of both worlds. J Infect Dis. (2023) 228:1311–3. doi: 10.1093/infdis/jiad353
36. Zhao Q, Wang X, Zhang Z, Liu X, Wang P, Cao J, et al. Serum neutralization of SARS-CoV-2 Omicron BA.2, BA.2.75, BA.2.76, BA.5, BF.7, BQ.1.1 and XBB.1.5 in individuals receiving Evusheld. J Med Virol. (2023) 95(7):e28932. doi: 10.1002/jmv.28932
37. Hoffmann M, Arora P, Nehlmeier I, Kempf A, Cossmann A, Schulz SR, et al. Profound neutralization evasion and augmented host cell entry are hallmarks of the fast-spreading SARS-CoV-2 lineage XBB.1.5. Cell Mol Immunol. (2023) 20:419–22. doi: 10.1038/s41423-023-00988-0
38. Schaefer-Babajew D, Wang Z, Muecksch F, Cho A, Loewe M, Cipolla M, et al. Antibody feedback regulates immune memory after SARS-CoV-2 mRNA vaccination. Nature. (2023) 613:735–42. doi: 10.1038/s41586-022-05609-w
39. Bloom N, Ramirez SI, Coelho C, Parikh UM, Heaps A, Sieg SF, et al. SARS-CoV-2 monoclonal antibody treatment followed by vaccination shifts human memory B cell epitope recognition suggesting antibody feedback. bioRxiv. (2023) 230(5):1187–96. doi: 10.1101/2023.11.21.567575
40. Bruel T, Vrignaud LL, Porrot F, Staropoli I, Planas D, Guivel-Benhassine F, et al. Antiviral activities of sotrovimab against BQ.1.1 and XBB.1.5 in sera of treated patients. medRxiv. (2023) 05(25):23290512. doi: 10.1101/2023.05.25.23290512
41. Pochtovyi AA, Kustova DD, Siniavin AE, Dolzhikova IV, Shidlovskaya EV, Shpakova OG, et al. In vitro efficacy of antivirals and monoclonal antibodies against SARS-CoV-2 omicron lineages XBB.1.9.1, XBB.1.9.3, XBB.1.5, XBB.1.16, XBB.2.4, BQ.1.1.45, CH.1.1, and CL.1. Vaccines (Basel). (2023) 11(10). doi: 10.3390/vaccines11101533
42. COVID-19 Treatment Guidelines Panel. Coronavirus Disease 2019 (COVID-19) treatment Guidelines. National Institutes of Health (2024).
Keywords: SARS-CoV-2, Omicron BA.4/5, Omicron XBB.1.5, bivalent vaccination, kidney transplant recipients
Citation: Tometten I, Brandt T, Schlotz M, Stumpf R, Landmann S, Kantauskaite M, Lamberti J, Hillebrandt J, Müller L, Kittel M, Ivens K, Gruell H, Voges A, Schaal H, Lübke N, Königshausen E, Rump LC, Klein F, Stegbauer J and Timm J (2025) Comparison of immune responses to SARS-CoV-2 spike following Omicron infection or Omicron BA.4/5 vaccination in kidney transplant recipients. Front. Immunol. 15:1476294. doi: 10.3389/fimmu.2024.1476294
Received: 05 August 2024; Accepted: 06 December 2024;
Published: 14 January 2025.
Edited by:
Mrinmoy Sanyal, Stanford University, United StatesReviewed by:
Sivaram Gunisetty, Emory University, United StatesLeonardo Sorrentino, Emory University, United States
Louise Benning, University of Heidelberg, Germany
Copyright © 2025 Tometten, Brandt, Schlotz, Stumpf, Landmann, Kantauskaite, Lamberti, Hillebrandt, Müller, Kittel, Ivens, Gruell, Voges, Schaal, Lübke, Königshausen, Rump, Klein, Stegbauer and Timm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Inga Tometten, aW5nYS50b21ldHRlbkBtZWQudW5pLWR1ZXNzZWxkb3JmLmRl
†These authors have contributed equally to this work and share last authorship