 Gayathri Gopakumar
Gayathri Gopakumar Mauricio J. C. Coppo
Mauricio J. C. Coppo Andrés Diaz-Méndez
Andrés Diaz-Méndez Carol A. Hartley
Carol A. Hartley Joanne M. Devlin
Joanne M. Devlin- 1Asia-Pacific Centre for Animal Health, Melbourne Veterinary School, Faculty of Science, The University of Melbourne, Parkville, VIC, Australia
- 2Escuela de Medicina Veterinaria, Universidad Andrés Bello, Concepción, Chile
A glycoprotein-G-deleted live-attenuated vaccine strain of the infectious laryngotracheitis virus (ILTV), ΔgG-ILTV, is safe and efficacious against ILTV challenge. In the current study, the transcriptome of peripheral blood mononuclear cells (PBMCs) of the ΔgG-ILTV-vaccinated group of specific-pathogen-free chickens were compared to those of the nonvaccinated group at 7 days post-vaccination. Tracheal transcriptomes after challenge with virulent ILTV were compared between groups of the non-vaccinated-challenged and the vaccinated-challenged as well as the non-vaccinated-challenged and the uninfected chickens at 4 to 5 days post-challenge. The clinical outcomes after challenge between these groups were also evaluated. Significant differences were observed in the tracheal transcriptome of the non-vaccinated-challenged birds compared to the other two groups. Enriched gene ontologies and pathways that indicated heightened immune responses and impairments to ciliary and neuronal functions, cell junction components, and potential damages to cartilaginous and extracellular components in the trachea of the non-vaccinated-challenged birds were consistent with their severe tracheal pathology compared to the other two groups. On the contrary, the absence of any difference in the tracheal transcriptome between the vaccinated-challenged and the uninfected birds were reflected by the preservation of tracheal mucosal integrity in both groups and mild infiltration of leukocytes in the vaccinated-challenged birds. The results from this study demonstrated that vaccination with ΔgG-ILTV prevented the changes in tracheal transcriptome induced during ILTV challenge, resulting in clinical protection. Additionally, these results also provide insights into the molecular mechanisms underlying the tracheal pathology induced by ILTV infection.
1 Introduction
Infectious laryngotracheitis virus (ILTV) is the etiological agent that causes infectious laryngotracheitis (ILT), a respiratory disease affecting poultry (1). Chickens of all age groups are susceptible to ILTV infection. However, outbreaks are commonly reported in birds older than 3 weeks of age.
ILTV infection induces significant pathological changes in the trachea and conjunctiva of birds and causes morbidity, mortality, and reduced egg production in the affected flocks, leading to significant economic losses to poultry industries worldwide (2). Despite the early onset of humoral immune response, disease protection is directly correlated to cell-mediated immune response. Currently, the disease is controlled by the implementation of biosecurity measures and vaccination practices (3).
Several categories of ILT vaccines have been developed over the years (3). These include a glycoprotein G (gG)-deleted live-attenuated vaccine (ΔgG-ILTV) (4) that has been well characterized using in vitro and in vivo studies (4–11). Initial studies of the ΔgG-ILTV vaccine demonstrated its attenuation (4) and evaluated its levels of immunogenicity and pathogenicity (5). A subsequent study demonstrated the safety and efficacy of this vaccine when delivered through eye-drop and drinking water, which indicated the suitability of the vaccine for mass vaccination of large-scale poultry operations (7). A further study identified the role of ILTV gG as a viral chemokine binding protein (6). The absence of gG during ILTV infection appeared to shift the host immune response away from humoral to the more protective cell-mediated immune response that aligned with the immunomodulatory role of gG (6). The minimum protective dose of the ΔgG-ILTV vaccine for eye-drop delivery has also been determined recently (9). Studies have also assessed the efficacy of drinking water method of administration of the ΔgG-ILTV vaccine in broiler chickens (8) and in ovo administration at different doses (10). A more recent study also investigated the latency establishment capacity of the ΔgG-ILTV vaccine (11).
In addition to these clinical investigations, transcriptome-level studies have also been conducted for the ΔgG-ILTV vaccine strain and the parent wild-type strain to evaluate the role of gG during ILTV infection. These studies have demonstrated the influence of gG on the transcription of a selected set of chicken chemokine- and cytokine-encoding genes (12) and the role of gG in the expression of other ILTV genes using qPCR analysis (13). Nevertheless, the genome-wide changes associated with ΔgG-ILTV vaccination in chickens as well as the genes and pathways associated with ΔgG-ILTV vaccine efficacy remain underexplored.
The aim of the current study was to identify the molecular markers associated with ΔgG-ILTV vaccination in peripheral blood mononuclear cells (PBMCs) of vaccinated chickens at 7 days post-vaccination (dpv) and to evaluate the host-response associated with ILTV challenge in the tracheal mucosae of ΔgG-ILTV vaccinated and non-vaccinated SPF chickens at 4 to 5 days post-challenge with a virulent field strain of ILTV.
2 Materials and methods
2.1 ILTV vaccine and challenge viral strain
Live-attenuated ΔgG-ILTV vaccine (Vaxsafe® ILT, Bioproperties Pty. Ltd.) (4–11) and an Australian virulent strain of ILTV, classified as class 9 ILTV (14, 15), were used in this study. The ΔgG-ILTV vaccine (4) was obtained from the manufacturer and reconstituted as per the manufacturer’s instructions. The class 9 ILTV was first isolated by chorioallantoic membrane inoculation and then propagated on chicken embryo kidney cells and leghorn male hepatoma (LMH) cells (16). Both strains of ILTV were quantitated using plaque-forming unit (PFU) assay on LMH cells prior to inoculation, as described previously (4).
2.2 ILTV vaccination and challenge of chickens
The methodology adopted for ILTV vaccination and infection of chickens has been previously described (7). Briefly, 45 white leghorn (SPF) chickens (Australian SPF Services Pty. Ltd.) were wing-tagged after hatching and randomly assigned into three groups of 15. The groups were housed in separate negative pressure isolator units and provided with irradiated feed and water ad libitum. At 1 week of age, the birds were vaccinated via eye-drop. Groups 1 and 2 were inoculated with 25 µL of sterile diluent. Group 3 was inoculated with 25 µL of ΔgG-ILTV vaccine (Vaxsafe® ILT) containing a dose of 103.8 plaque-forming units (PFU)/bird.
At 3 weeks post-vaccination, the birds were weighed and inoculated with a virulent field strain of ILTV to assess the level of protection provided by the vaccine. Group 1 was inoculated with sterile diluent and designated as the uninfected negative control group. Groups 2 and 3 were inoculated with the class 9 virulent field strain of ILTV at a dose of 500 PFU/bird and designated as the non-vaccinated-challenged positive control group and the vaccinated-challenged group, respectively. Inoculations were performed via eye-drop (125 PFU/40 µL/each eye) and intratracheal (250 PFU/150 µL) routes, delivering half the dose via each route. After challenge, the birds were monitored daily for clinical signs of ILT or mortality. At 3 and 4 days post-challenge (dpc), the birds were clinically assessed and scored as previously described (5) and as summarized in Supplementary Table S1. During clinical assessment, changes in demeanor, dyspnea, and signs of conjunctivitis of the birds were evaluated individually. A score ranging from 0 (normal) to 2 (severely depressed) for demeanor, 0 (normal) to 4 (severe gasping or birds dying due to respiratory distress) for dyspnea, and 0 (normal) to 2 (marked conjunctivitis) for conjunctivitis were used for the assessment.
A total of 10 birds in each group at 4 dpc and five birds in each group at 7 dpc were scheduled for euthanasia for the assessment of protection at the peak of ILTV infection (peak period of ILTV replication and clinical signs) and at a later period during the infection, respectively. Humane killing at the final timepoint or at another timepoint to alleviate suffering was undertaken using intravenous barbiturate (1:1 mixture of pentobarbitone solution and sterile water containing approximately 160 mg of pentobarbitone/mL, resulting in a dose of approximately 160 mg of pentobarbitone/kg body weight) overdose. All birds were weighed after death, prior to post-mortem examination, and percentage change in body weight was calculated based on the difference in weight before challenge and at the time of death. The post-mortem examination included an assessment and scoring of the gross pathology of the tracheal mucosae as described previously (5) and as summarized in Supplementary Table S2. The tracheal pathology scores ranged from 0 (absence of pathology) to 4 (severe mucus and hemorrhagic exudates), depending on the severity of the lesions.
All interventions as well as clinical and pathological scoring were performed by trained researchers. To ensure biosecurity, the uninfected group of birds was clinically scored first. Necropsy and sample collection were also conducted first in this group to minimize the risk of cross-contamination. This approach ensured that the researchers remained blinded during the clinical and pathological scoring of both the vaccinated-challenged and non-vaccinated-challenged birds, except for the uninfected group. Additionally, the researchers were blinded during the histopathological assessments of the trachea. Due to ethical considerations and animal welfare concerns, the animal trial in this study was conducted without any repetitions.
2.3 Sample collection
At 7 dpv, approximately 0.8 mL of blood was drawn from the basilic vein of both uninfected and vaccinated birds. This blood was collected into separate heparin-coated Vacutainer tubes (BD Biosciences) and stored on ice before isolation of PBMCs. Tracheal samples were collected from birds euthanized at 4 dpc (birds scheduled for euthanasia) or 5 dpc (birds euthanized due to the severity of clinical signs) and from the uninfected birds (4 dpc). A transverse section of the upper trachea (2 to 3 mm thick), immediately distal to the larynx, was collected and fixed in 10% v/v neutral buffered formalin prior to standard histological processing and hematoxylin and eosin staining. Tracheal sections were microscopically examined and scored from 0 (normal) to 5 (severe changes), depending upon the severity of the microscopic lesions as previously described (17) and as summarized in Supplementary Table S3. Tracheal mucosal scrapings were collected and stored in 600 µL RLT buffer (RNeasy Mini Kit, Qiagen) containing 1% (v/v) ß-mercaptoethanol and stored at -80°C for nucleic acid extraction.
2.4 Isolation of PBMCs
PBMCs were isolated from whole blood samples using Ficoll-Paque PLUS (Cytiva Lifesciences, Marlborough, MA, USA) gradient centrifugation according to the manufacturer’s instructions. Briefly, heparinized blood samples diluted 1:1 in phosphate-buffered solution (PBS) were gently layered over Ficoll-Paque PLUS in 15-mL tubes and centrifuged for 30 min at 400 × g at 20°C with no brake. The interface containing PBMCs was collected, washed with phosphate-buffered saline (PBS) twice by centrifugation at 1,000 × g for 7 min at room temperature, and the cell pellet was resuspended in 600 µL RLT buffer (RNeasy Mini Kit, Qiagen) containing 1% (v/v) ß-mercaptoethanol prior to storage at -80°C until nucleic acid extraction.
2.5 Nucleic acid extraction
Nucleic acids were extracted from the tracheal scrapings and from the PBMCs as mentioned in a previous study (18). For the extraction of nucleic acid from tracheal scrapings, approximately 30 mg of each of the scrapings was homogenized in 600 μL RLT buffer (RNeasy Mini Kit, Qiagen, Hilden, Germany) with 1% (v/v) ß-mercaptoethanol. Then, 200-μL aliquots of the homogenates were subjected to DNA extraction using MagMAX CORE Nucleic Acid Purification Kit (Thermo Fisher Scientific, Waltham, USA) coupled with the KingFisher Flex Purification System (Thermo Fisher Scientific) for automated extractions in 96-well plates as per the manufacturers’ instructions. The eluates (90 μL) were stored at -80°C until qPCR for the detection of ILTV genome copy numbers. The rest of the tracheal homogenates (approximately 300 μL each) and the leukocytes stored in RLT buffer (600 μL each) were subjected to RNA extraction using RNeasy Mini kit (Qiagen). They were then subjected to DNase treatment using Turbo DNA-free kit (Invitrogen, Carlsbad, CA, USA) and concentrated using Zymo RNA Clean and Concentrator-25 (Zymo research Corporation, Irvine, CA, USA) as per the manufacturers’ instructions. The integrity of the RNA eluates (50 μL) was assessed using 4200 Tape Station System (Agilent Technologies, Santa Clara, CA, USA) before storage at -80°C until RNA-seq analysis.
2.6 Quantification of ILTV in tracheal scrapings
Quantification of ILTV in tracheal scrapings was performed by employing qPCR analysis of the DNA samples in a Rotor-Gene Q Thermocycler (Qiagen, Hilden Germany). Amplification standard curves were generated using 10-fold serial dilutions of pGEM-T (Promega Corporation, Madison, WI, USA) plasmid carrying the ILTV UL15 gene amplicon (113 bp) (19). The UL15 genome copy numbers (GCN) in the samples were calculated based on cycle threshold values (Ct) using the Rotorgene Q software (version 2.1.0, Qiagen) at a cutoff of 100 genome copies per reaction for the detection and quantification of ILTV DNA.
2.7 cDNA library preparation and Illumina sequencing
Total RNA from the PBMCs of four uninfected and eight vaccinated birds and from the tracheal mucosae of eight non-vaccinated-challenged and six each of the uninfected and the most well-protected (based on clinical findings and tracheal pathology) vaccinated-challenged birds, respectively, was subjected to cDNA library preparation and Illumina sequencing at the Australian Genome Research Facility (AGRF, Melbourne, Victoria, Australia). All selected samples had RNA integrity numbers (RIN) of ≥7. Libraries were prepared using the TrueSeq Stranded mRNA library preparation kit (Illumina Inc., San Diego, CA, USA) and sequenced on NovaSeq (Illumina Inc.) using the Illumina DRAGEN BCL Convert 07.021.645.4.0.3 pipeline to generate 150-bp paired-end reads.
2.8 RNA-seq data pre-processing and differential gene expression analysis
Analyses of RNA-seq data were performed using the web-based bioinformatics platform GALAXY at usegalxy.au following the published Galaxy workflow (20–22). The quality of the raw data sets was analyzed using FastQC (Galaxy Version 0.74+galaxy0). Adaptor sequences were removed using Trimmomatic (Galaxy Version 0.36.6) and read pairs with PHRED quality score >20 and length >20 bp were retained. Using the Ensemble release 106 of the annotated chicken (Gallus gallus) genome (FASTA and gene transfer format) as reference, mapping of the reads was performed using RNA STAR (Galaxy Version 2.7.10b+galaxy3). Exon-level read counts of the BAM outputs were performed using featureCounts (Galaxy Version 2.0.3+galaxy1) and paired-end reads that mapped to multiple locations or reads with a minimum mapping quality score (minMOS) <10 were eliminated. Differential gene expression (DGE) analysis was performed using Deseq2 (Galaxy Version 2.11.407 +galaxy2) using the default GALAXY settings. Protein-coding genes of the chicken, differentially expressed in pair-wise comparisons between the different groups at P-adj value (P-value corrected using Benjamini–Hochberg procedure for the Wald statistic) <0.01 and with greater than or equal to twofold change (log2FC1) in expression, were considered significant. To assess the stability of the RNA-seq assay, the transcription of the 10 constitutively transcribed house-keeping genes reported for chicken trachea (23) was examined in the pair-wise comparisons between the different groups.
2.9 Gene ontology analysis
Gene ontology (GO) analysis was conducted using PANTHER (version 17.0) (24, 25). GO terms enriched for biological processes (BP), molecular functions (MF), and cellular components (CC) separately with the upregulated or downregulated genes in the trachea of the non-vaccinated-challenged birds were identified and compared to those of the vaccinated-challenged and the uninfected birds (18). The most enriched GO terms selected based on a false discovery rate (FDR) <0.05 and with fold enrichment values >1 were summarized using REVIGO (26) for the removal of redundant GO terms and reported.
2.10 Pathway and protein class analysis
Pathway and protein class analysis were performed using PANTHER (version 17.0) (24, 25). Reactome pathways and PANTHER protein classes enriched separately with the upregulated and downregulated genes in the trachea of the non-vaccinated-challenged birds compared to those of the other two groups at FDR <0.05 were identified and reported.
2.11 Statistical analysis and data visualization
Mann–Whitney U-test was used for the comparisons of clinical scores, tracheal gross pathology scores, and tracheal histopathology scores. Fisher’s exact test was used for the comparison of proportions between groups. Normality of the data for weigh change and ILTV genome copy numbers (GNCs) were assessed using Shapiro–Wilk’s test. One-way ANOVA corrected for multiple comparisons using Tukey’s test was used for the comparison of the normally distributed percentage weight change data. Kruskal–Wallis test corrected for multiple comparisons using Dunn’s test was used for the comparison of the non-normally distributed ILTV GCN data. Statistical analyses were performed using Prism-GraphPad version 9.4.1. Data visualization was performed using R version 4.0 and SRplot.
3 Results
3.1 Vaccinated birds were protected from ILTV challenge
The vaccinated birds had significantly (P < 0.05) less severe clinical signs than the non-vaccinated-challenged birds at 3 (P = 0.0002) and 4 dpc (P = 0.0087) (Figures 1A, B). Significant differences were also observed in the cumulative mortality between the vaccinated-challenged and the non-vaccinated-challenged birds (P = 0.002). While there were no mortalities among the vaccinated-challenged birds (0% cumulative mortality at 7 days post-challenge), the non-vaccinated-challenged group experienced two sudden deaths—one bird at 3 dpc and another at 4 dpc. Additionally, six birds in the non-vaccinated-challenged group were subjected to humane killing at 3 dpc due to severe clinical signs, resulting in a cumulative mortality rate of 53.33% by 7 days post-challenge, in accordance with the specified interventions outlined in the animal ethics approval. These birds were assigned a summed clinical score of 5, as individual scoring was not feasible due to the immediate need for euthanasia based on ethical considerations.
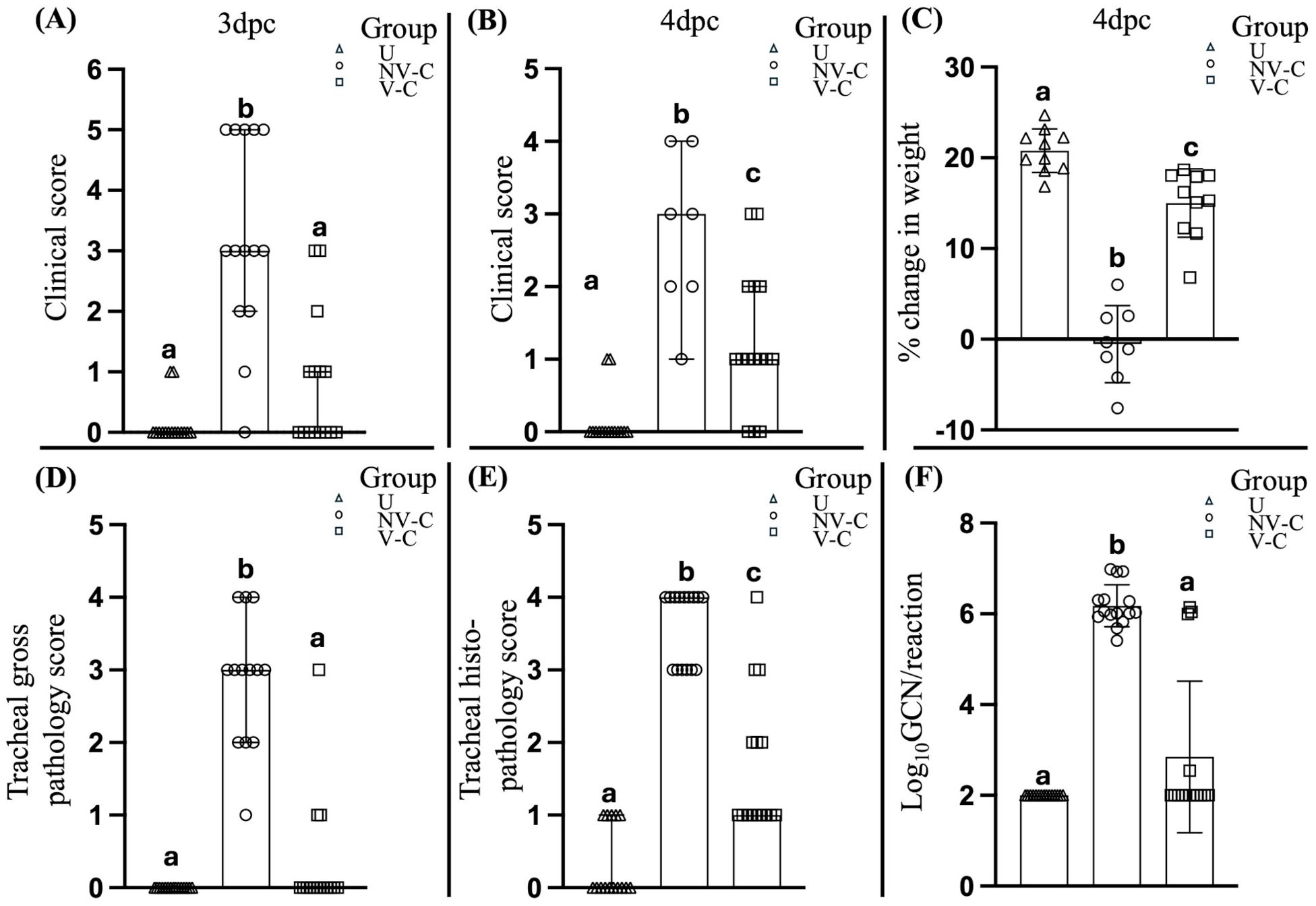
Figure 1. Clinical assessment of the uninfected (negative control) birds, non-vaccinated-challenged birds, and vaccinated-challenged birds after challenge with virulent ILTV. Scatter plot with bars (A, B) represents the clinical scores of the birds at 3 and 4 days post-challenge (dpc) respectively. Scatter plot with bars indicating (C) percentage change in body weight, (D) tracheal gross pathology scores, (E) tracheal histopathology scores, and (G) ILTV genome copy number (GCN)/reaction; lines at median with 95% confidence interval in (A, B, D–F) and line at mean with standard deviation in (C). Log10 GCN/reaction values of the samples that tested negative for ILTV DNA (F) were adjusted to 2, as 100 copies per reaction were used as the cutoff for the detection and quantification of ILTV DNA; values marked with the same superscript letter above each group in each panel were not significantly different (P > 0.05); U, uninfected; NV-C, non-vaccinated-challenged; V-C, vaccinated-challenged; dpc, days post-challenge.
The vaccinated-challenged birds also had a significantly greater weight gain than the non-vaccinated-challenged birds (P < 0.0001) at 4 dpc (Figure 1C).
3.2 Vaccinated birds had reduced tracheal pathology and ILTV GCN after challenge with virulent ILTV
The vaccinated birds had significantly lower (P < 0.05) tracheal gross pathology and histopathology scores after challenge compared to the non-vaccinated-challenged birds (Figures 1D, E). The non-vaccinated-challenged birds had a median gross tracheal pathology score of 3, corresponding to a large amount of bloody mucus with the occasional presence of diphtheritic plaques, while the vaccinated-challenged birds had a score of 0 (Figure 1D). A median tracheal histopathology score of 4 in the non-vaccinated-challenged birds (Figure 1E) reflected severe changes to the trachea. Compared to the uninfected birds (Figures 2A, B), the tracheae of the non-vaccinated-challenged birds exhibited heavy infiltration of leukocytes accompanied by edema or cellular exudate and an absence of normal epithelium (Figures 2C, D). On the other hand, the median tracheal histopathology score of 1 in the vaccinated challenged birds (Figure 1E) reflected minimal changes, with intact ciliated columnar epithelial cells and mild leukocyte infiltration (Figures 2E, F).
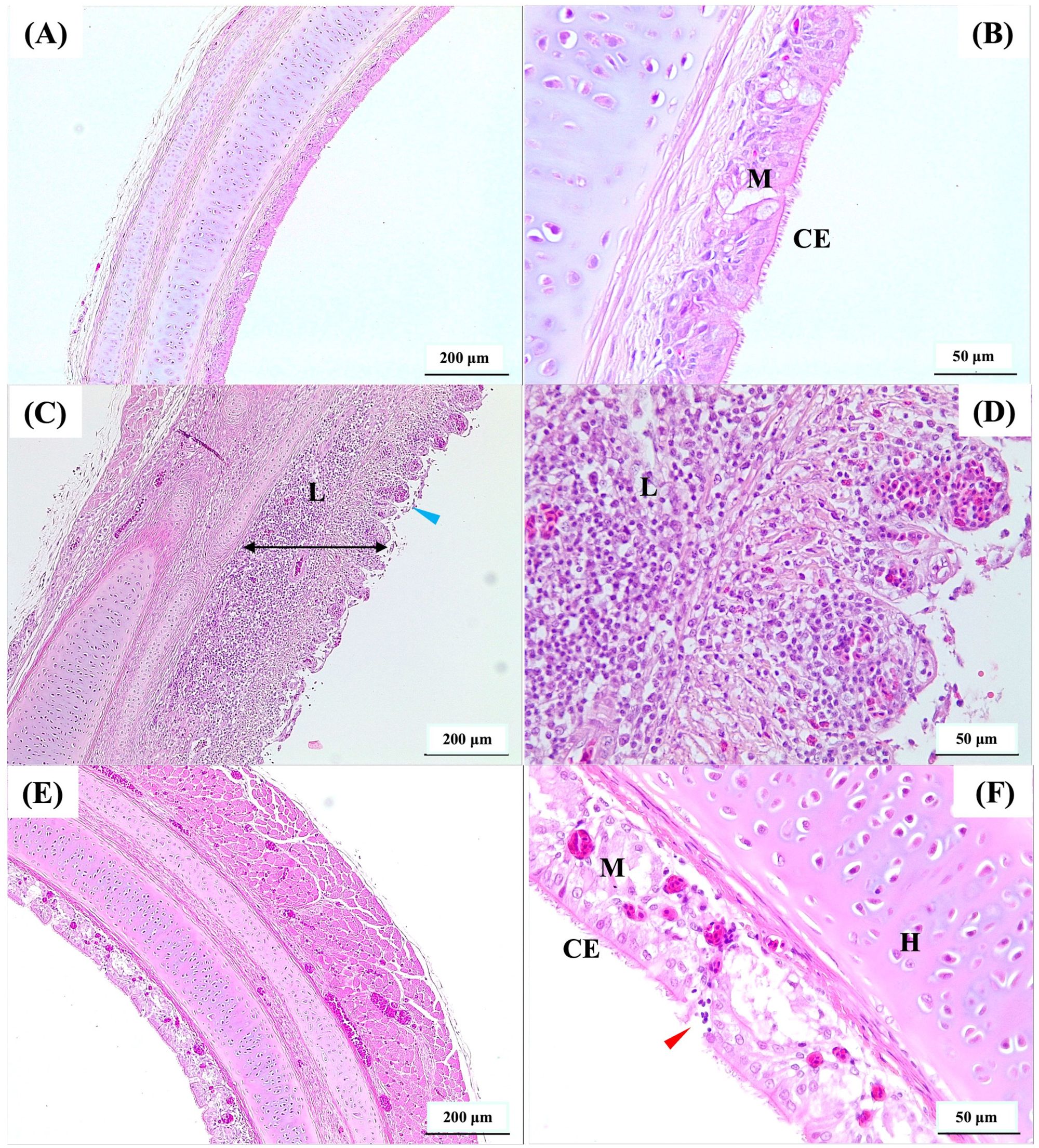
Figure 2. Photomicrographs of hematoxylin and eosin-stained cross-sections of the upper trachea. (A, B) represent uninfected, (C, D) represent non-vaccinated-challenged, and (E, F) represent vaccinated-challenged birds with median histopathology scores of 0, 4, and 1, respectively. The presence of abnormal epithelium without cilia (blue arrowhead) and increased thickness of mucosa (double headed black arrow) due to edema and heavy infiltration of leukocytes (L) was observed in (C, D). Ciliated pseudostratified columnar epithelial cells (CE), mucus or goblet cells (M), and mild infiltration of leukocytes (red arrowhead) were observed in (E, F). Scale bars in (A, C, E) correspond to 200 μm and in (B, D, F) indicate 50 μm. H, hyaline cartilage.
Significant differences (P < 0.05) between the vaccinated-challenged and the non-vaccinated-challenged birds were also observed in the proportion of birds that tested positive for ILTV in the trachea (Figure 1F). ILTV was only detected in 4/15 of vaccinated birds but detected in each of the 15 non-vaccinated-challenged birds. Additionally, the genomic copy numbers (GCNs) of ILTV were significantly lower (P = 0.0002) in the vaccinated birds compared to the non-vaccinated-challenged birds (Figure 1F).
3.3 Differential gene expression analysis
RNA-seq reads after quality assessment and trimming were mapped to the reference chicken genome (Supplementary Tables S4, S5). The global gene expression pattern of the PBMCs, visualized via principal component analysis (PCA), showed no clear distinction between the vaccinated and the non-vaccinated (uninfected) groups (Supplementary Figure S1). No difference was observed in the gene expression of the PBMCs of the vaccinated birds at 7 dpv compared to the uninfected group, at P-adj value <0.01 and greater than or equal to twofold change (log2FC 1) in expression.
The global gene expression pattern of the tracheal mucosae of the vaccinated-challenged and the uninfected groups were similar at 4 or 5 dpc (Supplementary Figure S2). No genes exhibited differential regulation between the groups at P-adj value < 0.01 and a fold change in expression of at least twofold (log2FC). On the other hand, the non-vaccinated-challenged group segregated into a distinct cluster, separated from the other two groups. A greater number of genes was upregulated than downregulated in the non-vaccinated-challenged group compared to the other two group (Figures 3A, B, respectively), and a large proportion of the differentially expressed genes observed in the non-vaccinated-challenged group was shared between the vaccinated-challenged and the uninfected groups (Figures 3C, D, respectively). Of the 10 most stable and reliable chicken tracheal housekeeping genes (23), the majority were found to be constitutively transcribed, with no significant difference in their expression in the pairwise comparisons between the non-vaccinated-challenged and the uninfected birds (8/10 genes) and the non-vaccinated-challenged and the vaccinated-challenged bids (7/10), thus validating the stability of the RNA-seq results (Supplementary Tables S6, S7).
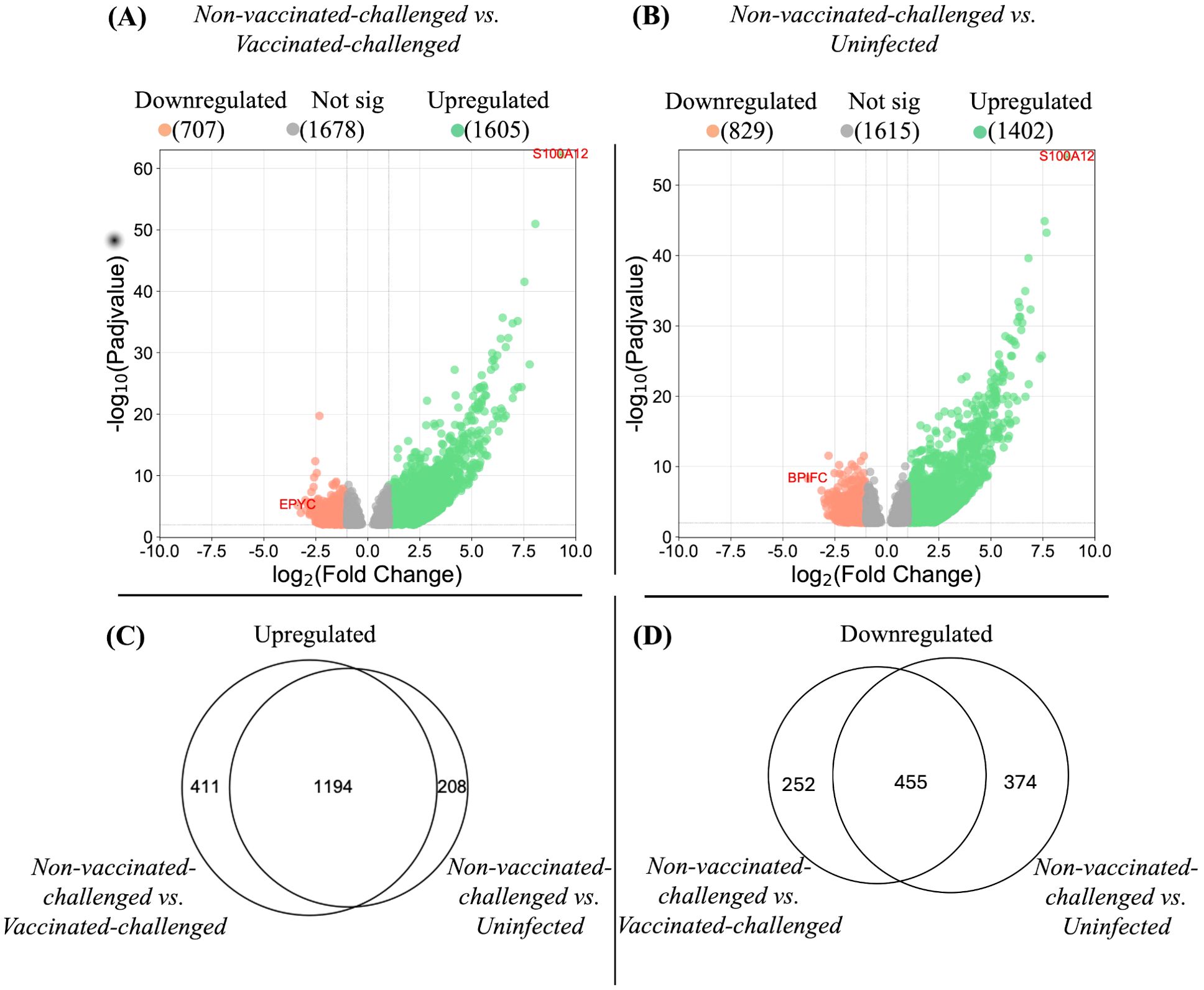
Figure 3. Differentially expressed genes in tracheal mucosae. (A, B) represent the differentially expressed genes in the tracheal mucosae of the non-vaccinated-challenged group compared to the vaccinated-challenged and to the uninfected groups, respectively. Genes differentially regulated at P-adj value <0.01 and log2 fold change ≥1 were considered significantly upregulated, and genes differentially regulated at P-adj value <0.01 and log2 fold change ≤-1 were considered significantly downregulated. The position of the most significantly up- and downregulated gene in each comparison is labeled with gene symbols. Proportional Venn diagrams (C, D) represent the number of differentially expressed genes in the tracheal mucosae of the non-vaccinated-challenged group compared to the vaccinated-challenged and/or the uninfected groups. (C) Upregulated genes. (D) Downregulated genes. The numbers in the intersection of the circles represent the number of genes up- or downregulated in the non-vaccinated-challenged compared to the vaccinated-challenged or the uninfected group, while the numbers outside the intersection of the circles represent the uniquely up- or downregulated genes in the non-vaccinated-challenged group compared to each of the groups.
3.4 Genes most significantly upregulated in the trachea of the non-vaccinated-challenged birds were mostly related to immune response, cell proliferation, and cell activation
The gene encoding S100A12, a chemoattractant protein expressed in granulocytes, exhibited the highest level of upregulation in the tracheal mucosa of the non-vaccinated-challenged birds compared to the vaccinated-challenged (log2FC 9.27) and uninfected birds (log2FC 8.67). This inflammatory marker protein is involved in chemotaxis, production of cytokines, cell proliferation and differentiation, and induction of oxidative stress (27, 28). Several other genes involved in immune and inflammatory responses were also upregulated in the non-vaccinated-challenged birds and constituted the largest category of genes by function, among the top 20 upregulated genes (Table 1). This included the genes encoding several chemokines and cytokines and/or their receptors, immunoglobulin receptors, TLR15, ACOD, a mitochondrial enzyme that regulates immunometabolism, inflammation, and infection in several species (29), and the GTPase, RAB3C, involved in the recycling of phagocytosed MHC class I complex (in humans) (30). The other categories of genes in the list included those that encode proteins involved in the proliferation and activation of cells (granulocytes, macrophages, osteoclasts, and chondrocytes), regulation of apoptosis and necroptosis, extracellular matrix (ECM) remodeling (metallopeptidases), and vascular repair (NOXO1) and fluid homeostasis (SCNNID).
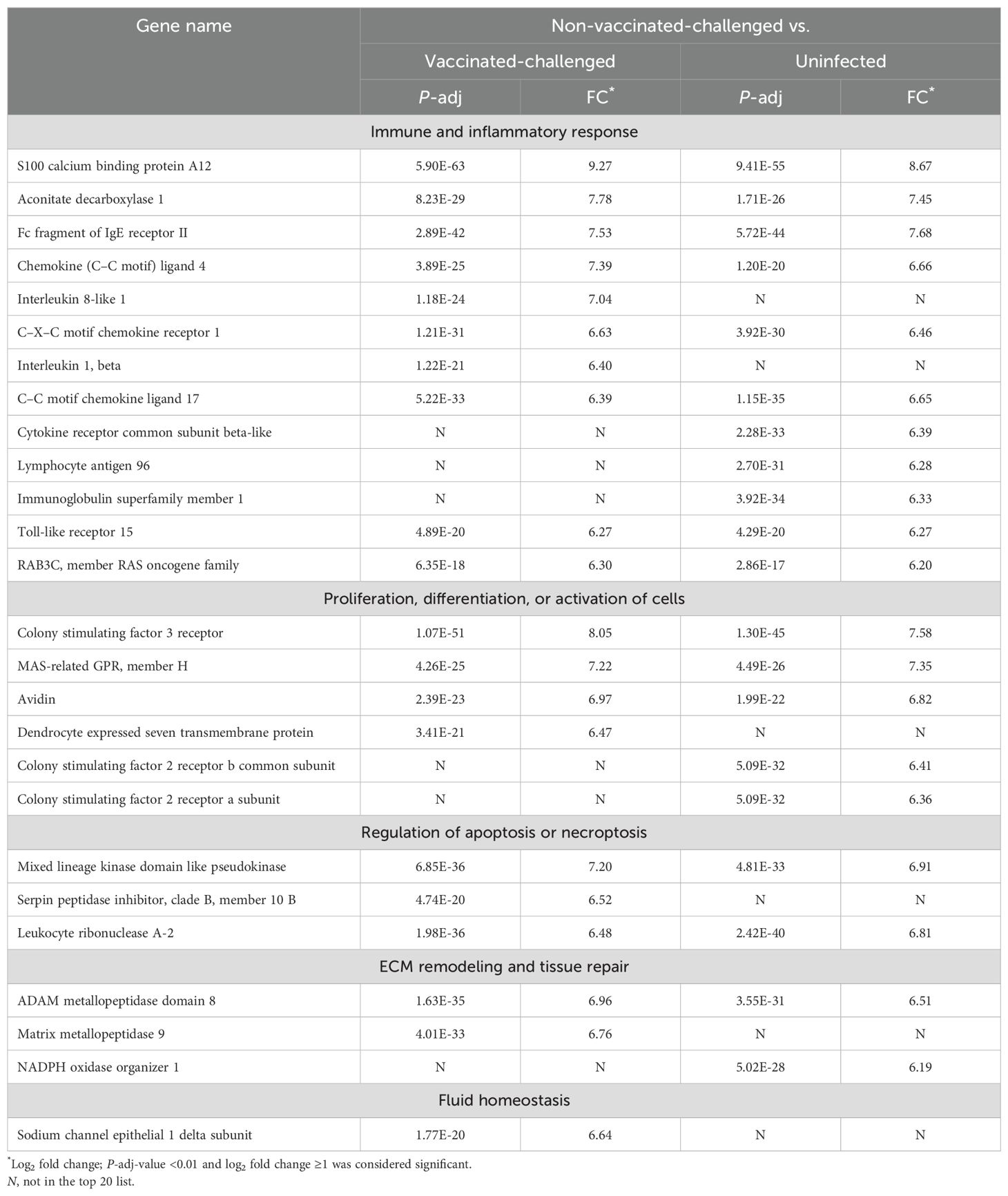
Table 1. Top 20 upregulated genes in trachea of the non-vaccinated-challenged birds compared to the vaccinated-challenged or uninfected birds.
3.5 Genes most significantly downregulated in the trachea of the non-vaccinated-challenged birds were mostly related to extracellular matrix organization and cell adhesion
Genes encoding EPYC and BPIFC were the most downregulated genes in the trachea of the non-vaccinated-challenged birds compared to the vaccinated-challenged (log2FC = -3.37) and the uninfected birds (log2FC = -3.80), respectively. The former is a proteoglycan that interacts with collagen fibers and ECM proteins and aids in cartilage matrix organization (31), while the latter is a palate, lung, and nasal epithelium clone (PLUNC) protein expressed by airway epithelial and/or mucus cells, with innate immune functions (32, 33). Of the top 20 downregulated genes in non-vaccinated-challenged birds, genes that encode proteins involved in tissue remodeling and ECM organization constituted the largest category of genes by function (Table 2). This included the genes encoding ACAN, a proteoglycan involved in cartilage remodeling and ECM organization (34), the bone matrix non-collagenous protein OC3 (35), the cartilage matrix protein CHAD (36), growth factors such as CTGFL and GDF10 and the cell-line-derived transforming sequencing MCF2 with roles in bone morphogenesis and remodeling (37–39), AVPR1A involved in hematopoiesis (40), and the muscle protein MYH1C (41). The second major category of genes included those that encode proteins with roles in cell adhesion. Genes related to other functions in the list included those with neuronal functions, transmembrane transport, immune and inflammatory responses, steroid metabolism, protein production, and protein-to-protein interactions.
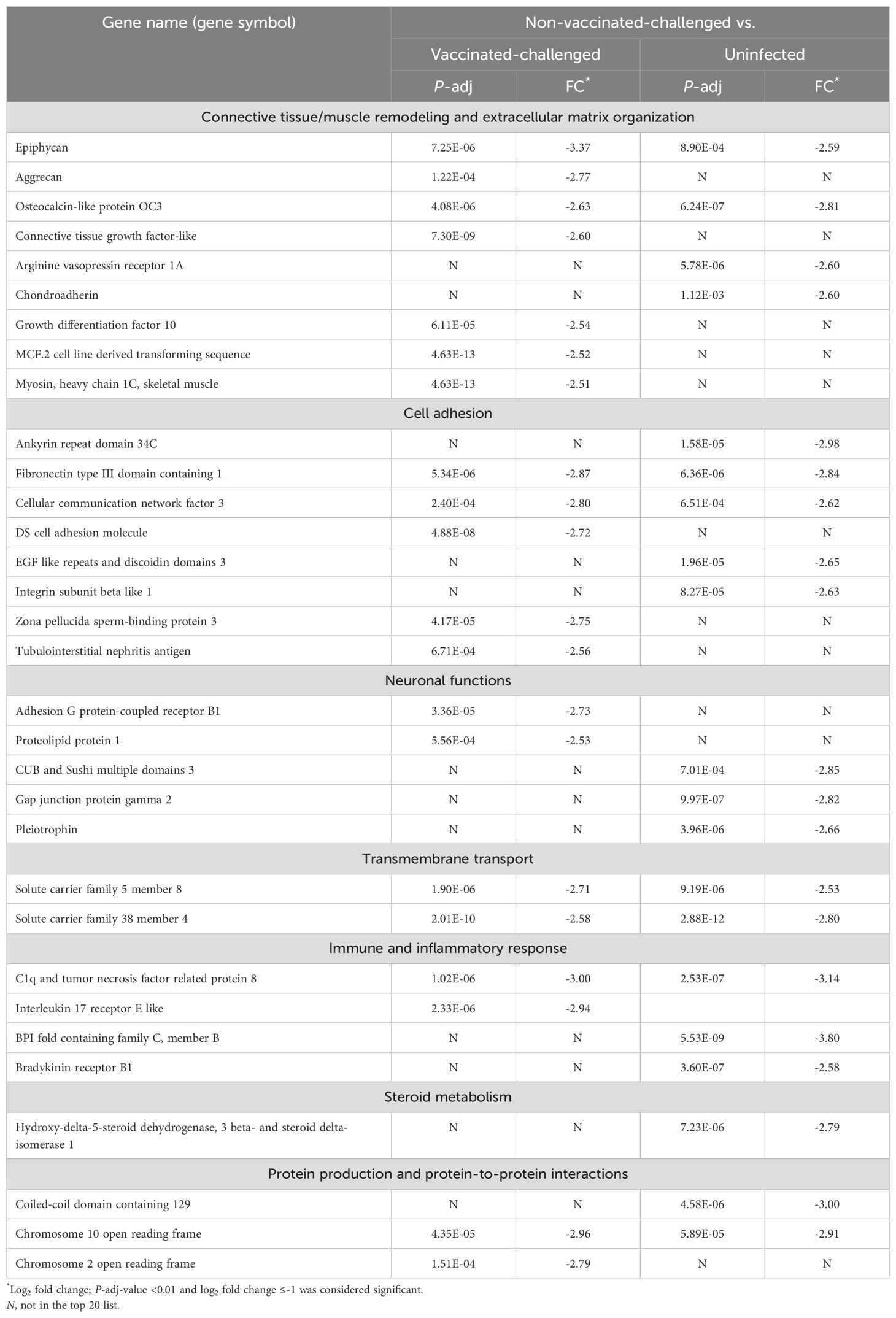
Table 2. Top 20 downregulated genes in the trachea of the non-vaccinated-challenged birds compared to the vaccinated-challenged or uninfected birds.
3.6 Genes differentially regulated in the non-vaccinated-challenged birds included many with immune-related functions
A large number of genes with role in immune response were upregulated in the tracheae of the non-vaccinated-challenged birds compared to the birds in the other two groups. Of these, the largest category was constituted by chemokines, cytokines, and their receptors (Supplementary Table S8). The chemokine-encoding genes most significantly upregulated in the non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups were IL8L1 (log2FC = 7.04) and CCL17 (log2FC = 6.65), respectively. While the expression of genes encoding several other interleukins, interleukin receptors, -CC and -CXC families of chemokines, and chemokine receptors, were upregulated in the non-vaccinated-challenged group compared to either of the other groups, the gene for XCL1 (log2FC = 1.90), one of the two members of the C-family of chemokines and CX3CR1 (log2FC = 2.21), the receptor for the only known CX3C subfamily of chemokines, was upregulated uniquely compared to the uninfected group (42, 43). Genes encoding several toll-like receptor family members (TLR15, TLR4, TLR7, TLR2B, TLR1B, and TLR2A) and MHC-related proteins, including MHC class I, MHC class II, and MHC B-G antigen-related proteins, were also upregulated in the non-vaccinated-challenged group. Upregulated interferon-encoding genes included IFNκL1 and IFN-γ, which were upregulated compared to both the vaccinated-challenged (log2FC = 5.08 and 4.51, respectively) and the uninfected groups (log2FC = 4.97 and 4.14, respectively), while the gene for IFN-ω1 was upregulated uniquely when compared to the former (log2FC = 2.30). Genes encoding several interferon regulatory factors and interferon-induced proteins constituted the other interferon-related genes that were upregulated in the non-vaccinated-challenged group. Among the genes encoding nuclear factors that were upregulated in the non-vaccinated-challenged birds, genes for NFE2, which regulate antioxidant response elements during oxidative airway disorders (44, 45), were upregulated when compared to both the vaccinated-challenged and the uninfected groups (log2FC 1.80 and 1.53, respectively). Additionally, genes encoding NFIL3 and NFκB2 (log2FC = 1.37 and 1.18, respectively) were also upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged birds. The other upregulated immune-related genes included those encoding several TNF superfamily members, TNF-alpha-induced proteins, and CD molecules as well as complement-related proteins.
The largest category of genes with immune-related functions that was downregulated in the non-vaccinated-challenged group was that of “complement components”, which included complement factors and C1q tumor necrosis factor (TNF)-related superfamily of proteins (CTRP) (Supplementary Table S9). Downregulated chemokines and/or chemokine-receptor-encoding genes included CCL20 (log2FC = -1.86, -2.52) and receptors for interleukin 1 (IL1R1, log2FC = -1.07, -1.08) and 17 (IL17REL, log2FC = -2.94, -2.13) and an accessory protein for interleukin 1 receptor (IL1RAPL2, log2FC = -2.10, -1.74). In addition to this, a few immunoglobin-related genes and CD molecules were also downregulated.
3.7 Gene ontology terms related to immune response, cytokine and chemokine activity, and membrane were the most upregulated terms in the non-vaccinated-challenged birds
The gene ontology analysis of the upregulated genes of the non-vaccinated-challenged group compared to the vaccinated-challenged and to the uninfected groups revealed comparable profiles for the most enriched (FDR <0.05 and fold enrichment >1) BP, MF, and CC root categories. A total of 76, 18, and 10 GO terms for BPs, MFs, and CCs, respectively, were enriched with the genes upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged birds, while 59, eight, and nine GO terms for the same, respectively, were enriched with the genes upregulated in the non-vaccinated-challenged birds compared to the uninfected birds. The largest compartments in the REVIGO summarized tree maps for the upregulated GOs were constituted by the representative BP terms “immune response” and “regulation of immune response”, MF terms for “cytokine binding” and “cytokine receptor activity”, and the CC term for “external side of plasma membrane” and “side of membrane” (Supplementary Figures S3-S8). The GO terms enriched most significantly with the upregulated genes in the non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups are shown in Figure 4. The top 10 upregulated BPs were mostly related to immune, defense, and inflammatory responses, signaling, and response to cytokines. The top five MFs were mostly related to cytokine and chemokine activity and binding, while the top five CCs mostly highlighted extracellular components and membranes (Figure 4). The upregulated BP, MF, and CC terms in the non-vaccinated-challenged group with the greatest fold enrichment (FE) compared to those of the vaccinated challenged birds were the terms for “cellular response to biotic stimulus” (16/21 genes, FE = 8.6), “complement binding” (5/5 genes, FE = 11.28), and “intermediate filament” (10/34 genes, FE = 3.32), respectively, while those compared to the uninfected birds were the terms for “positive regulation of adaptive immune response” (4/5 genes, FE = 10.33), “MHC protein binding” (5/6 genes, FE = 10.76), and “external side of plasma membrane” (67/146 genes, FE = 5.93), respectively.
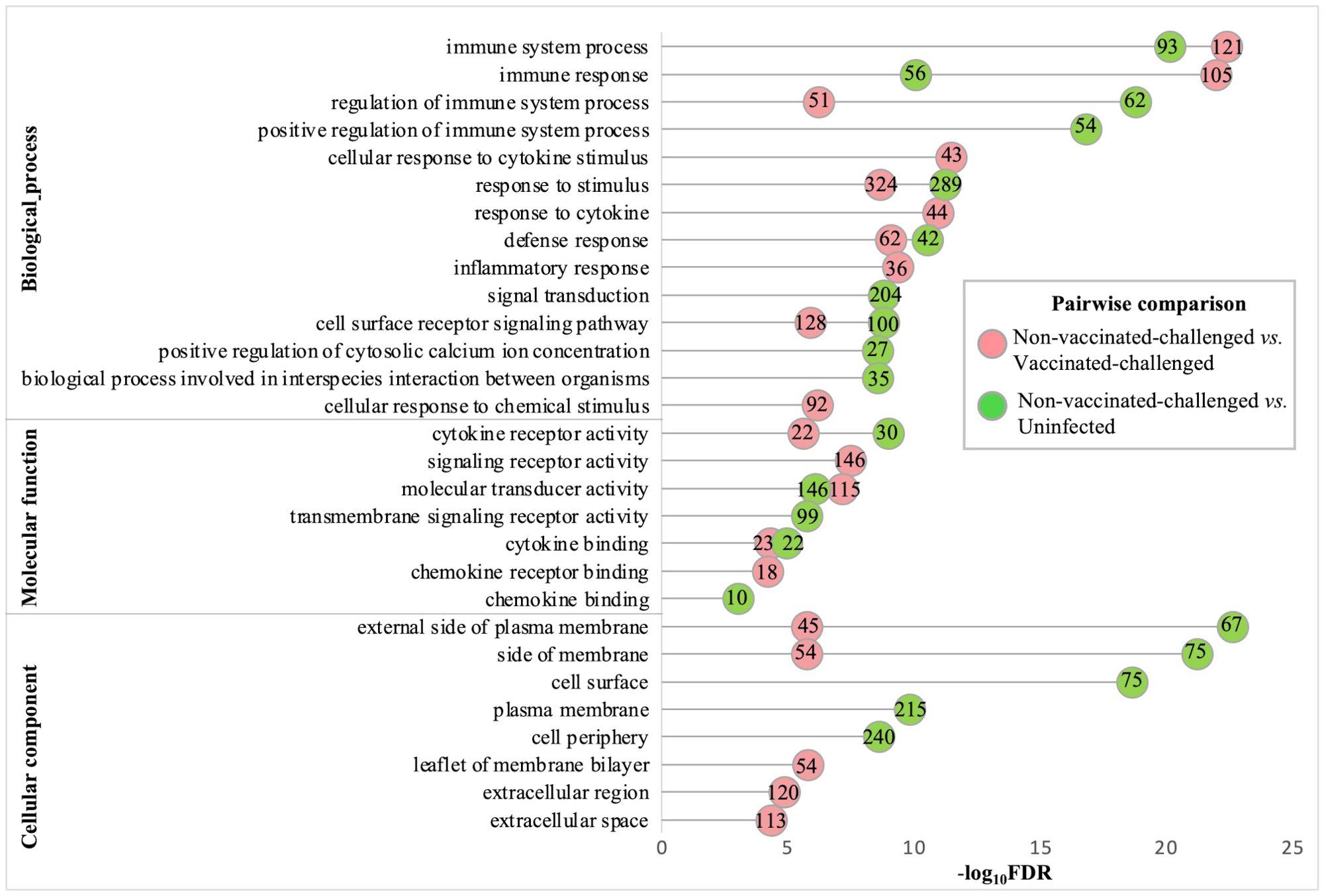
Figure 4. Gene ontologies most significantly enriched (FDR <0.05 and fold enrichment >1) with the upregulated genes of the non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups. The top 10 (FDR < 0.05) biological process (BP), five molecular functions (MF), and five cellular components (CC) terms are listed. The numbers inside the circles denote the number of genes enriched for the GO terms.
3.8 Gene ontology terms related to developmental processes, extracellular structure organization, channel activity, and adhesion were the most downregulated terms in the non-vaccinated-challenged birds
The gene ontologies enriched with the downregulated genes in the non-vaccinated-challenged group compared to the other two groups were similar for the most enriched (FDR <0.05 and fold enrichment >1) BP, MF, and CC categories. A total of 23, six, and 13 GO terms for BPs, MFs, and CCs, respectively, were enriched with the genes downregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged birds, while 40, 12, and nine GO terms for the same, respectively, were enriched with the genes downregulated in the non-vaccinated-challenged birds compared to the uninfected birds. The largest compartments in the REVIGO summarized tree maps for the GOs downregulated in non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups, respectively, were constituted by the representative BP terms “anatomical structure development” and “regulation of multicellular organismal development”, MF terms “voltage-gated monoatomic ion channel activity” and “cell adhesion molecule binding”, and the CC term “extracellular matrix” (Supplementary Figures S9-S14). The GO terms enriched most significantly (FDR < 0.05) with the downregulated genes in the non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups are shown in Figure 5. The top 10 downregulated BPs were mostly related to various developmental processes, extracellular structure organization, cell adhesion, cell communication, signaling, and generation of neurons. The top five MFs were mostly related to channel or transporter activity and protein binding, including receptors and cell adhesion molecules, while the top five CCs mostly highlighted extracellular components and membranes including cell junctions (Figure 5). The downregulated BP, MF, and CC terms in the non-vaccinated-challenged group with the greatest fold enrichment (FE) compared to the vaccinated challenged birds were the terms for “extracellular matrix organization” (11/62 genes, FE = 4.54), “metalloendopeptidase activity” (10/69 genes, FE = 3.71), and “post-synaptic density” (9/57 genes, FE = 4.04), respectively, while those compared to the uninfected birds were the terms for “cognition” (4/9 genes, FE = 9.71), “heparin binding” (6/21 genes, FE = 6.24), and “external encapsulating structure” (43/254 genes, FE = 3.7).
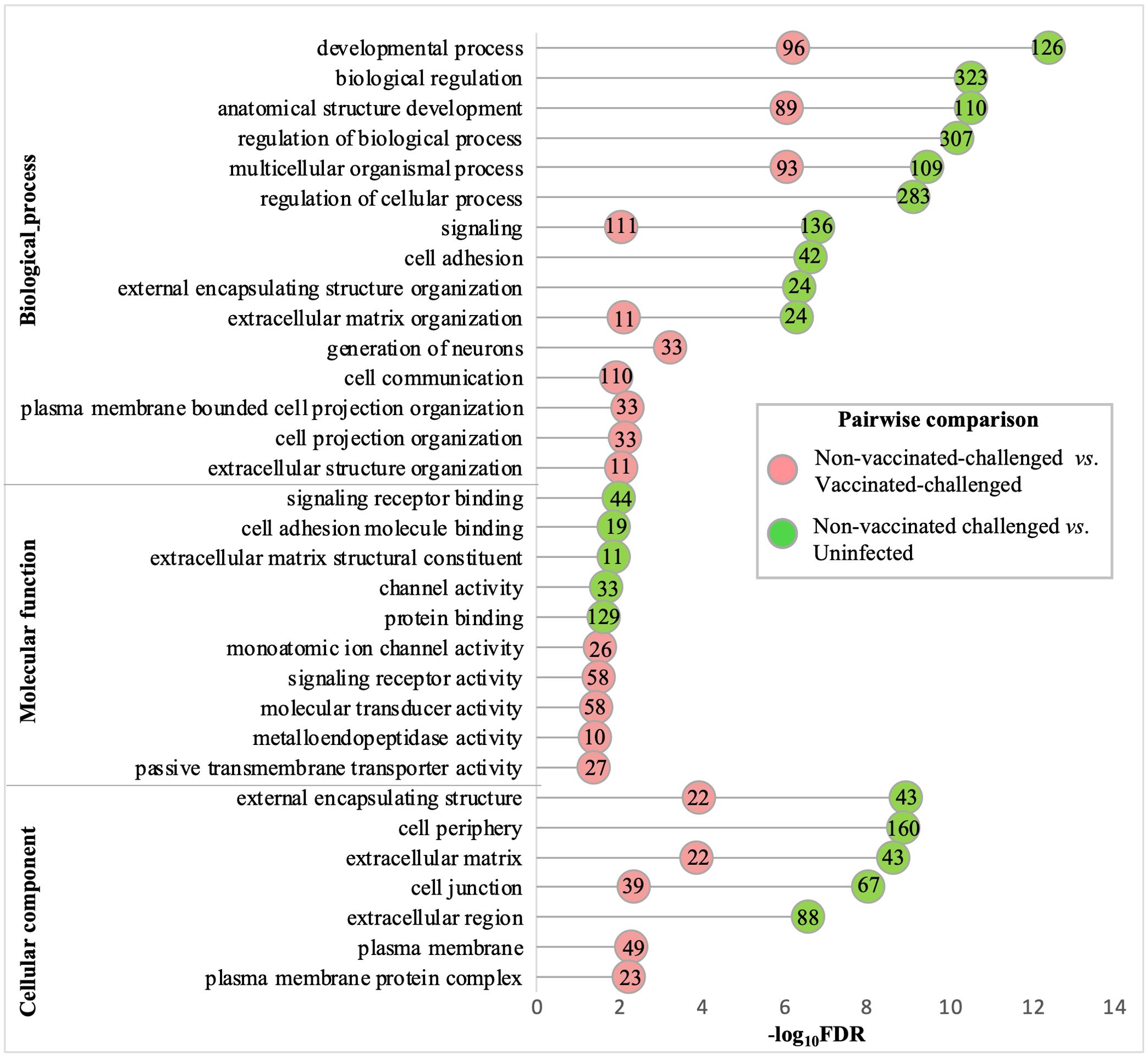
Figure 5. Gene ontologies most significantly enriched (FDR <0.05 and fold enrichment >1) with the downregulated genes of the non-vaccinated-challenged group compared to the vaccinated-challenged and the uninfected groups. The top 10 (FDR < 0.05) biological process (BP), five molecular functions (MF), and five cellular components (CC) terms are listed. The numbers inside the circles denote the number of genes enriched for the GO terms.
3.9 Pathways mainly involved in immune response were upregulated, while ECM and collagen organization were downregulated in the non-vaccinated-challenged birds
A total of 36 and 51 Reactome pathways were enriched with the genes upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged and the uninfected birds, respectively (Figure 6). This included pathways related to immune response, signaling, RNA metabolism, gene expression, and transport, which were upregulated in the non-vaccinated-challenged birds compared to both the vaccinated-challenged and the uninfected birds as well as the pathway for “hemostasis”, upregulated solely in comparison with the uninfected birds. Of this, pathways involved in immune response formed the largest category that constituted 20/36 and 29/51 of the upregulated pathways in the non-vaccinated-challenged birds when compared to the vaccinated-challenged and the uninfected birds, respectively.
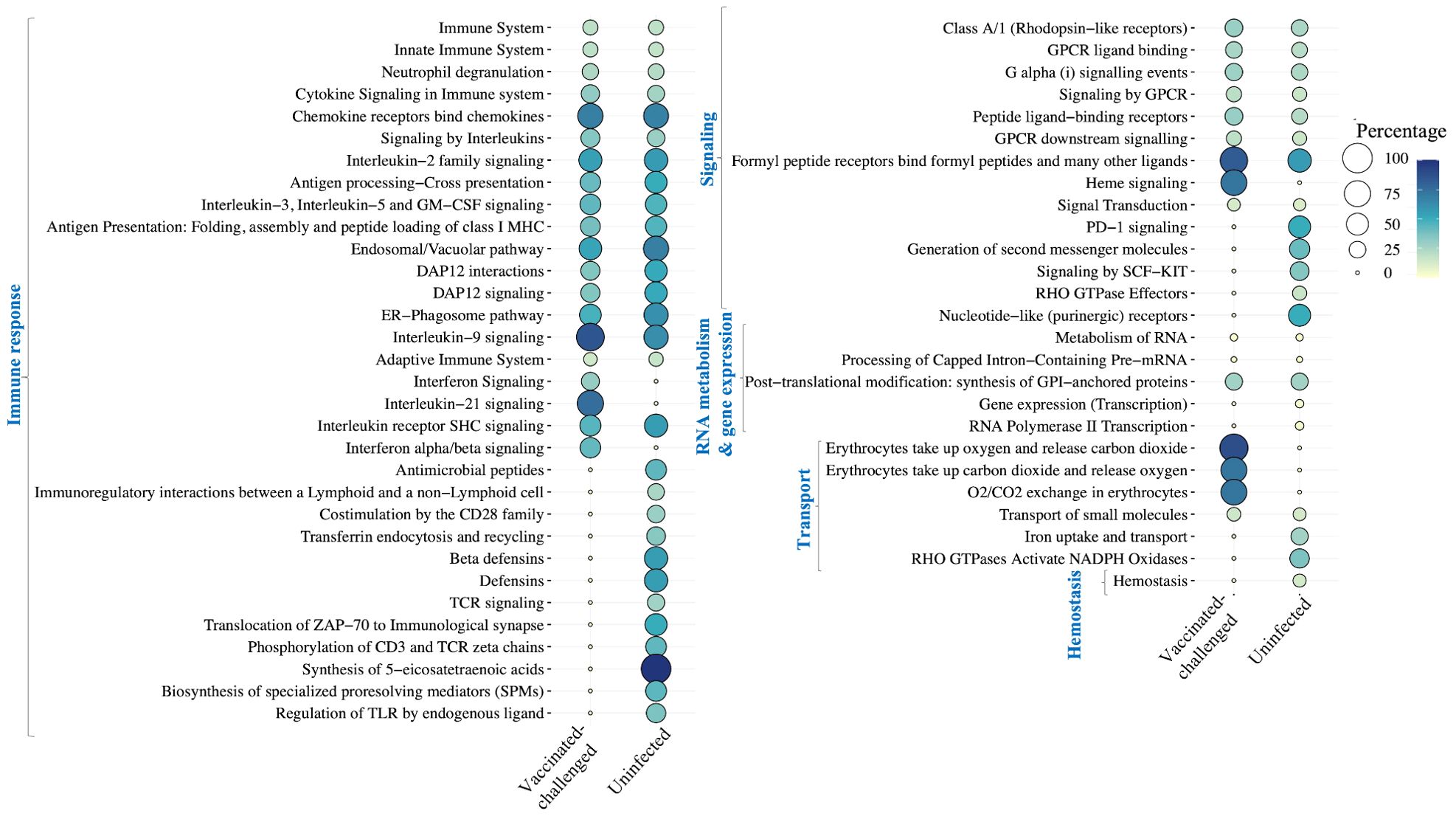
Figure 6. Reactome pathways upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged or the uninfected birds at false discovery rate (FDR) <0.05. The pathways are arranged into categories based on functions, indicated in blue font on the left side. Percentage indicates the proportion of genes enriched in the pathway.
A total of 11 and 22 Reactome pathways were enriched with the genes downregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged and the uninfected birds, respectively (Figure 7). This included pathways related to extracellular matrix and collagen organization, immune response, and growth and development that were upregulated in the non-vaccinated-challenged birds compared to both the vaccinated-challenged and the uninfected birds as well as pathways related to cell adhesion and cell differentiation, translation, and signal transduction, downregulated solely in comparison with the uninfected birds. Of this, pathways involved in extracellular matrix and collagen organization formed the largest category that constituted 8/11 and 11/22 of the downregulated pathways in the non-vaccinated-challenged birds when compared to the vaccinated-challenged and the uninfected birds, respectively.
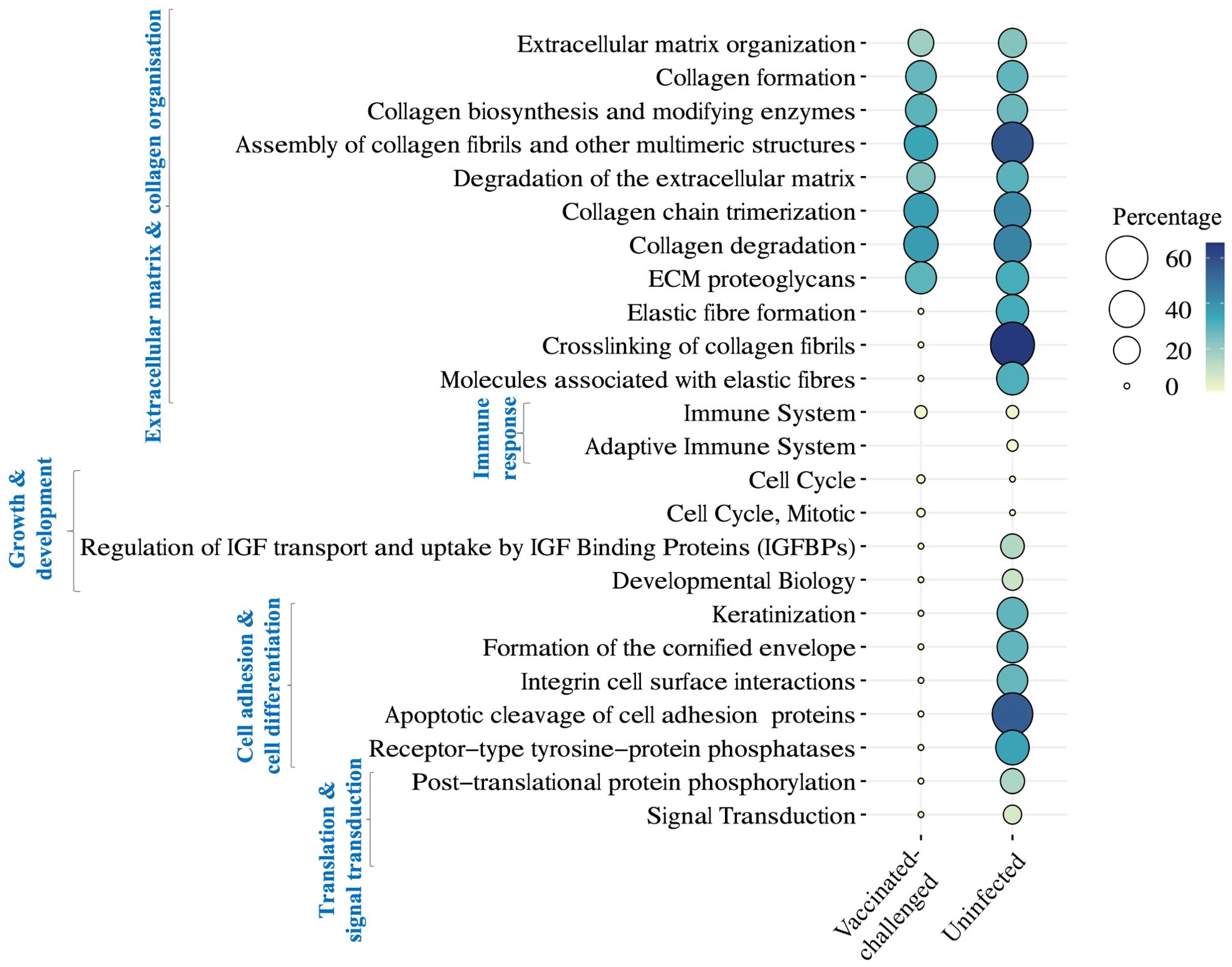
Figure 7. Reactome pathways downregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged or the uninfected birds at false discovery rate (FDR) <0.05. The pathways are arranged into categories based on functions, indicated in blue font on the left side. Percentage indicates the proportion of genes enriched in the pathway.
3.10 Immune response and ECM/structural organization formed the largest categories of the up- and downregulated protein classes, respectively, in the non-vaccinated-challenged birds
A total of 16 and 17 PANTHER protein classes were enriched with the genes upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged and the uninfected birds, respectively (Table 3). These included protein classes with roles in defense and immune response, signal transduction, RNA metabolism, and/or gene expression that were upregulated in the non-vaccinated-challenged birds compared to both the vaccinated-challenged and the uninfected birds. Additionally, a class of enzyme was also upregulated in the non-vaccinated-challenged birds compared to the uninfected birds. The protein class enriched with the greatest number of upregulated genes per class in the non-vaccinated-challenged group was the “major histocompatibility complex protein” with 19/33 (57.58%) and 21/32 (65.63%) of the upregulated genes compared to the vaccinated-challenged and the uninfected birds, respectively.
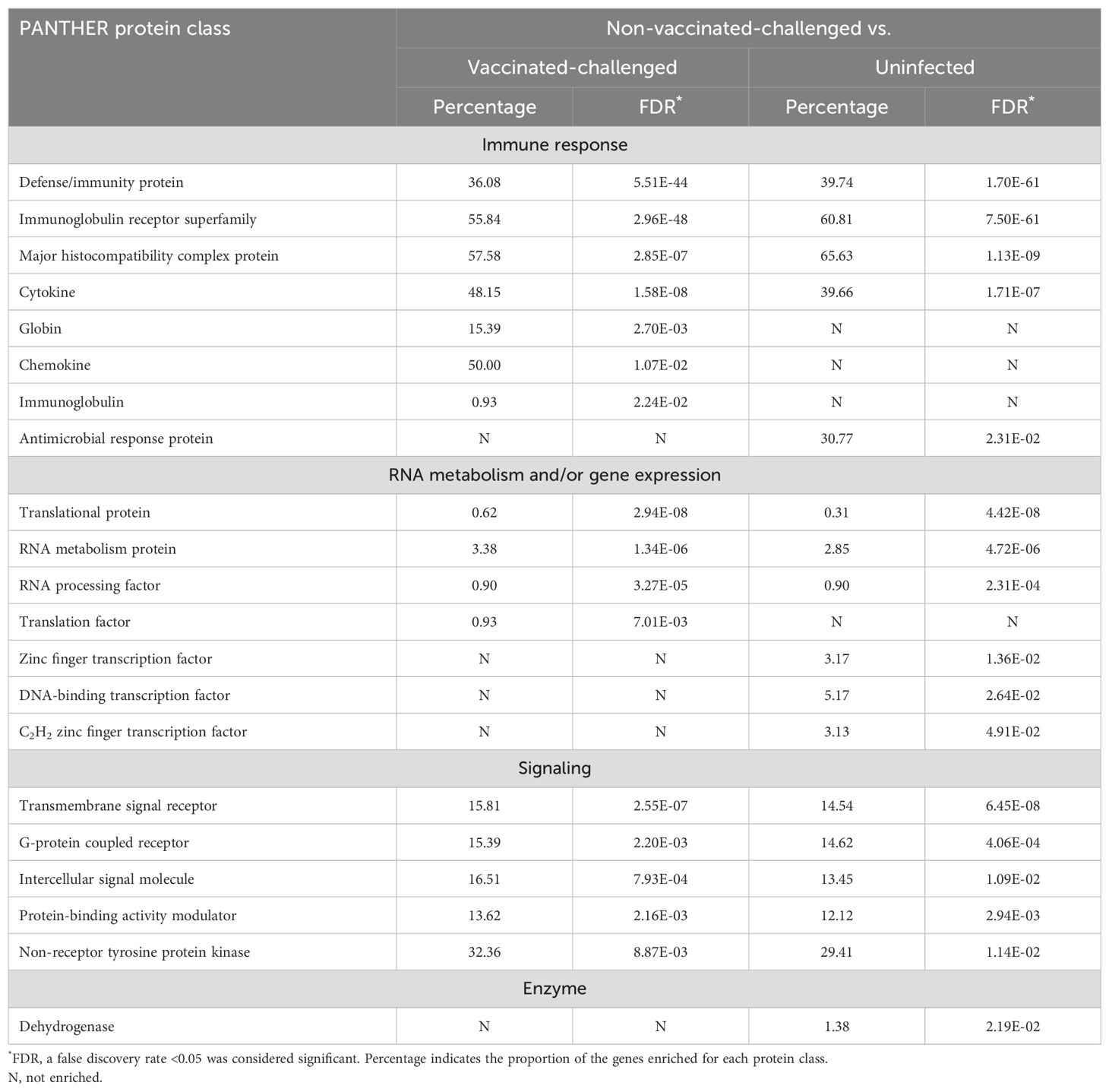
Table 3. Protein classes upregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged or uninfected birds.
Seven and 16 PANTHER protein classes were enriched with the genes downregulated (Table 4) in the non-vaccinated-challenged birds compared to the vaccinated-challenged and the uninfected birds, respectively. These included the protein classes for ECM and/or structural proteins, classes with role in cell adhesion, nucleic acid metabolism, translation, and immune response that were downregulated in the non-vaccinated-challenged birds compared to the other two groups. Additionally, a few classes of enzymes were also downregulated in the non-vaccinated-challenged birds compared to the uninfected birds. The protein classes enriched with the greatest number of downregulated genes per class in the non-vaccinated-challenged group were “extracellular matrix structural protein” with 16/80 (20%) and “intermediate filament binding protein” with 4/10 (40%) of the downregulated genes compared to the vaccinated-challenged and the uninfected birds, respectively.
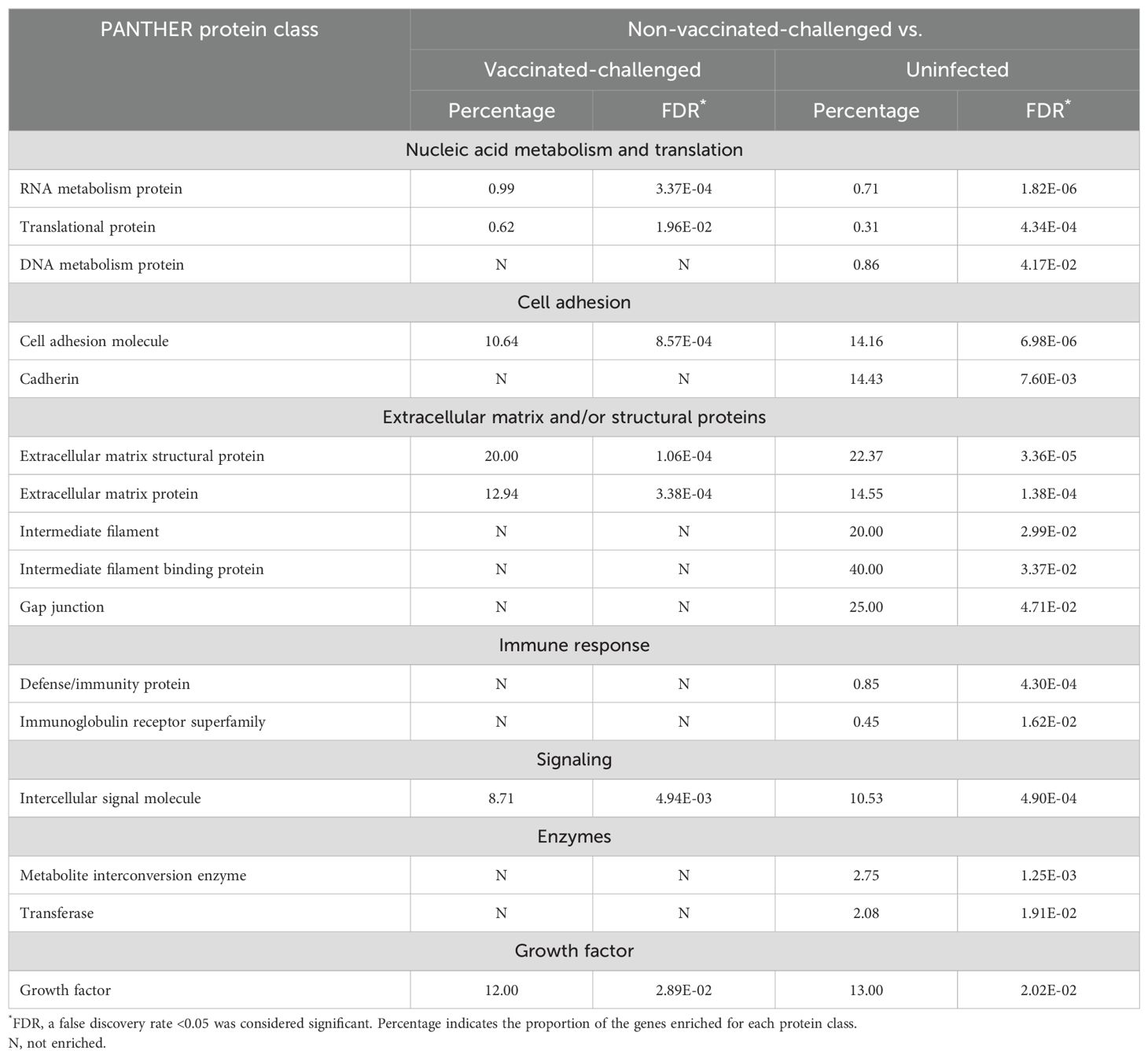
Table 4. Protein classes downregulated in the non-vaccinated-challenged birds compared to the vaccinated-challenged or uninfected birds.
4 Discussion
A comparative analysis of the mortality data, weight gain, clinical signs scores, tracheal gross and histopathology scores, and the qPCR results from tracheal swabs between the non-vaccinated-challenged and the vaccinated-challenged birds indicated that vaccination with ΔgG-ILTV provided effective protection against challenge with virulent ILTV. These results are consistent with those of a previous study that assessed the immunogenicity and pathogenicity of ΔgG-ILTV (5). The absence of any significant difference in tracheal transcriptome between the vaccinated-challenged and the uninfected birds, in contrast to the extensive differences between these groups and the non-vaccinated-challenged group at 4 to 5 dpc, provides further evidence of a high level of vaccine efficacy and helps to correlate differences in tracheal host gene transcription with the outcome of infection.
Tracheal transcriptome data at 4 to 5 dpc (Supplementary Tables S10-S13) was used to collate an understanding of the molecular events pertaining to ILT infection, particularly immune response and the organization of structural components, that distinguished the non-vaccinated-challenged birds from the other two groups (Figure 8). The gene encoding CCL20 was the only upregulated (downregulated in the non-vaccinated-challenged birds) chemokine in the vaccinated-challenged birds (Supplementary Table S9). Upregulation of this gene commonly with the uninfected group and its constitutive expression in mucosal tissues suggested that the chemokine potentially functioned in orchestrating homeostatic trafficking of leukocytes (46). This was consistent with the mild infiltration of inflammatory cells in the trachea of the vaccinated-challenged birds (Figure 2F), which aligned with their significantly lower tracheal pathology (12). Conversely, upregulation of serval TLRs (TLR1B, TLR2A, TLR2B, TLR4, TLR7, and TLR15) in the non-vaccinated-challenged birds (Supplementary Table S8) was consistent with the activation of several downstream signaling pathways that triggered diverse immune and inflammatory responses, thus exacerbating the tracheal pathology in these birds. Ligand-induced stimulation of TLR2 and TLR 4 has been shown to result in antiviral response to ILTV infection in vitro or in ovo (47–50), while the involvement of TLR7 and TLR15 has been demonstrated during Mark’s disease virus infection and TLR1 during avian leukosis virus infections in chickens (51, 52) in previous studies.
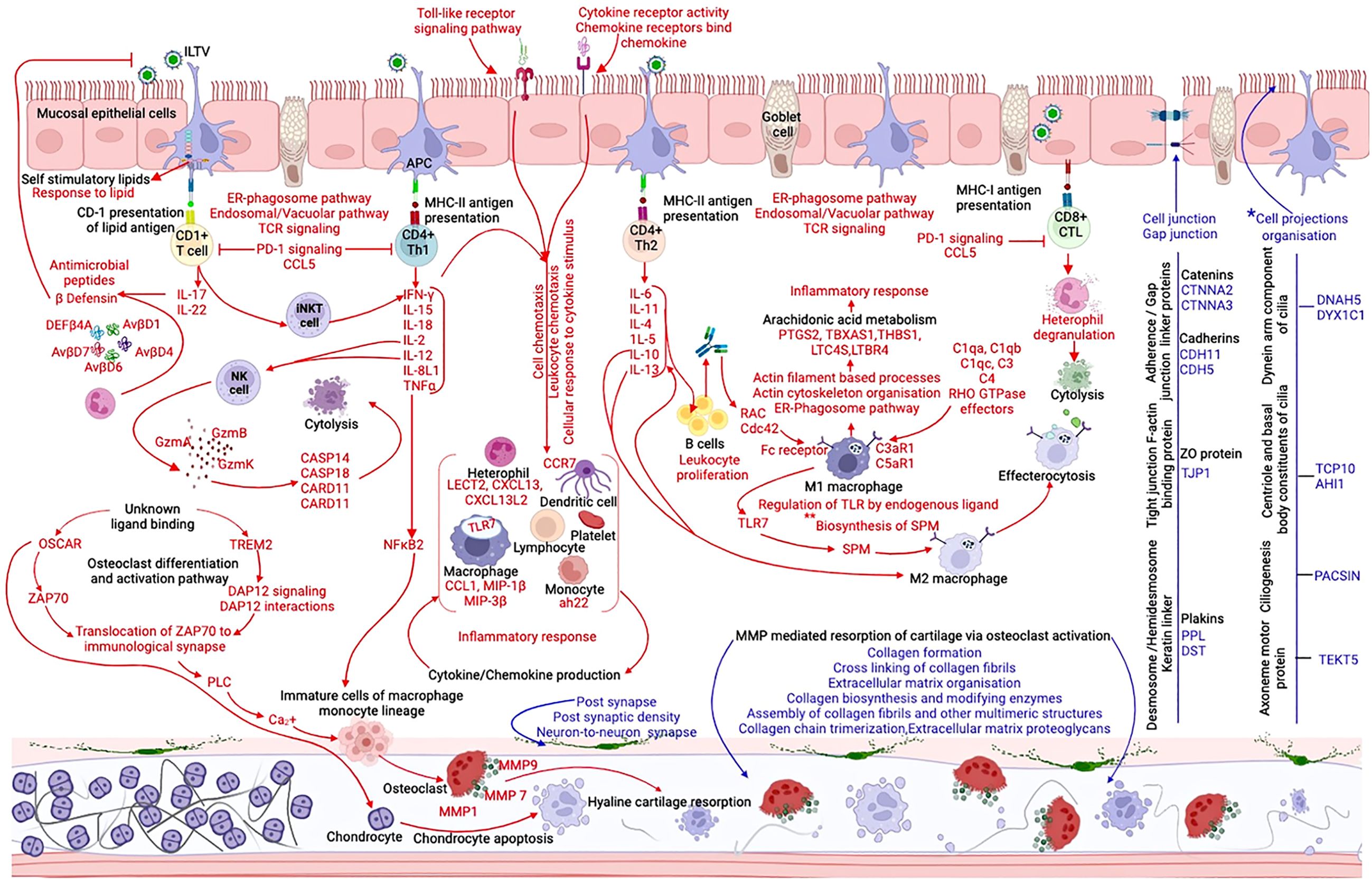
Figure 8. Schematic representation of the immune and inflammatory response in the trachea of non-vaccinated-challenged birds at 4 to 5 days post-challenge, created with BioRender.com. The arrows and texts in red indicate the upregulated and those in blue indicate the downregulated genes, gene ontologies, pathways, or protein classes in the non-vaccinated-challenged birds compared to the vaccinated-challenged and/or uninfected birds. *, differentially regulated only in comparison with the vaccinated-challenged group. **, differentially regulated only in comparison with the uninfected group.
Upregulation of the cathepsin-mediated “vacuolar pathway” as well as the cytosolic “ER-phagosome pathway” in the non-vaccinated-challenged birds (Figure 6) indicated the involvement of both TAP-independent and TAP- dependent mechanisms, respectively, for MHC-peptide antigen cross-presentation during ILTV infection (53, 54). Furthermore, upregulation of the genes encoding the CD1 molecules, CD1b and CD1c (gene synonyms CD1.1, CD1.2, CD1A1, and CD1D) (55, 56), also suggested the activation of the CD1-dependent lipid antigen presentation pathway (Figure 8) (57–59). The involvement of this pathway in the defense response to several bacterial and viral pathogens, including HSV-1, has been shown to be mediated through the activation of Th17 cytokines or through the activation of invariant natural killer T (iNKT) cells that facilitates an antiviral response through the enhanced production of Th1 cytokines like IFN-γ (60, 61). Activation of this pathway in the non-vaccinated-challenged birds is likely mediated via the interaction between virus-induced self-stimulatory lipids or lipopeptides (generated by cellular lipidation of viral proteins) and specific CD-1 restricted T cells (29, 57, 61–63). This predicted interaction was evidenced by the upregulation of BP terms indicating “response to lipid” in these birds (Supplementary Figure S3). In agreement with a previous study that demonstrated the destructive phagocytosis of ILTV-infected macrophages (64), the current study detected evidence of both type I antibody-mediated and type II complement-mediated opsonophagocytosis (65) in the non-vaccinated-challenged birds (Figure 8). This was substantiated by the upregulation of genes encoding several Rho-GTPase-activating/associated proteins, complement receptors, and Fc-fragment receptors and the enrichment of BP terms indicating actin cytoskeleton remodeling in these birds (Supplementary Figure S3) (66).
The upregulated CD molecules in the non-vaccinated-challenged birds (Supplementary Table S8) indicated diverse cell types populating their trachea, consistent with the histopathological findings (Figures 2C, D). This aligned with the upregulation of several cytokine- and chemokine-encoding genes (Supplementary Table S8) and the upregulated gene ontologies indicating immune cell migration (Supplementary Figures S3, S6). The upregulated chemokine- and cytokine-encoding genes in these birds included the -C-X-C family of chemokines and LECT2 that are chemotactic for heterophils/neutrophils (67, 68), several -C-C family of chemokine that are chemotactic for monocytes and T cells (68), and ah221, belonging to the monocyte chemotactic family (69), among the many others. The presence of dendritic cells was indicated by the upregulation of CCR7, a chicken dendritic cell marker (70), while the upregulation of macrophage inflammatory proteins, MIP-1b (CCL4) and MIP-3b (CCL19), was consistent with the recruitment of macrophages (71–73).
It could be hypothesized that the downregulation of IL1R1 promoted the upregulation of IL-1β in the non-vaccinated-challenged birds as IL1R1–IL-1β interactions result in negative feedback, reducing the IL-1β levels (74). Similarly, downregulation of the gene encoding IL17REL, a negative regulator of IL-17 receptors (75), might have enhanced the expression of the IL-17A- and IL-17F-encoding genes in these birds, consequently promoting the production of several proinflammatory cytokines, recruitment of heterophils, and the production of antimicrobial peptides such as β-defensins (76, 77) (Figure 6). Among the 14 avian β-defensins identified to date (78), five were upregulated in the non-vaccinated-challenged birds (Figure 8). Their role in antiviral immunity to ILTV has not been demonstrated yet. Nevertheless, multiple antiviral mechanisms demonstrated for these innate immune components against Marek’s disease virus (MDV), chicken infectious bronchitis virus (IBV), and Newcastle disease virus (NDV) in chickens (79–83) warrant investigation of their role during ILTV infection.
The results of cytokine and chemokine transcription in the non-vaccinated-challenged birds were largely in agreement with Vagnozzi et al. and the in vivo component of the investigation conducted by Coppo et al. (12, 84). Importantly, the absence of IFN-β, but IFN-γ gene upregulation in the nonvaccinated- challenged birds underscores the finding that ILTV infection interferes with IFN-β gene transcription (84). Although there was no upregulation of the gene encoding IFN-α in these birds, upregulation of genes for several interferon α-inducible proteins and the upregulated pathway for “interferon α/β signaling” suggested its activation. There are very limited studies that explored the functions of IFN-κL1 and IFN-ω1 in chickens (85). The upregulation of these genes in the non-vaccinated-challenged birds during the peak of infection indicates that they likely have a role in immune response to ILTV infection in chickens. Genes encoding the anti-inflammatory cytokines IL-10 and IL-13, upregulated in the non-vaccinated-challenged birds, were also in agreement with Vagnozzi et al. (2018), who noticed their peak upregulation at 5 dpi, parallel to the severe pathological changes in the trachea (84). These chemokines were suggested to have a role in dampening the inflammatory responses through M2 macrophage polarization (Figure 8), and hence their upregulation suggests the onset of a transition from proinflammatory phase to tracheal regenerative phase (84). In addition to this, the pathway upregulated for “PD-1 signaling”, which inhibits T cell activation, proliferation, and survival (86), in combination with two other upregulated pathways involved in the production of specialized pro-resolving mediators (SPM) (Figures 6, 8), which promotes M2 macrophage polarization and effecterocytosis, consequently dampening the inflammatory responses (87–89) (Figure 8), and underpins the notion of tissue restoration in these birds.
As observed in a few recent ILTV studies (71, 90), the non-vaccinated-challenged birds in the current study also displayed loss of or damage to the cilia on tracheal epithelial cells (Figures 1D, 2C). Consistent with these observations, the biological process of “cell projections organization” (Supplementary Figure S9) as well as several genes (DNAH5, DYX1C1, TCP10, AHI1, PACSINs, and TEKT5) involved in ciliogenesis, cilium assembly, and ciliary motility (91–96) were downregulated in these birds (Figure 8). This indicated that damage to ciliary components during ILTV infection is not solely a consequence of viral replication in mucosal epithelial cells but, rather, is regulated at the transcriptome level, thus confirming suggestions from an earlier study (12). Due to the significant role that cilia play in the mucociliary clearance of particles including viruses in respiratory epithelium, impairment of their function indicates a compromise to innate immune mechanisms in these birds. There were also indications of damage to the components of the extracellular matrix and cartilaginous structures in the trachea of the non-vaccinated-challenged birds (Figure 8). Several alleles of the gene encoding the immunoglobulin receptor OSCAR, upregulated in the non-vaccinated-challenged birds (Supplementary Table S8), are directly correlated to the destruction of chondrocytes via the induction of chondrocyte apoptosis in cartilages (97). Furthermore, the other upregulated genes encoding TREM-2, ZAP70, and PLC, together with OSCAR, induce the Ca2+ influx necessary for the differentiation and maturation of osteoclasts, which further mediates cartilage resorption/ECM and collagen degradation through the expression of MMPs (also upregulated) (98, 99). These predicted effects also aligned with the upregulation of signaling pathways for DAP12 and ZAP-70 molecules, positive regulation of cytosolic calcium ion concentrations, and the simultaneous downregulation of several pathways for ECM and collagen organization and synthesis (Figures 6–8) in non-vaccinated-challenged birds. Activation of osteoclast by NF-kB is yet another possible pathway that culminated in these potential structural modifications (99) (Figure 8). Previous studies have also reported the upregulation of MMPs following the infection of chicken embryo lung cells with wild-type ILTV (100) and downregulation of several classes of collagen in chickens immunized with a CEO ILT vaccine that associated these changes to a potential compromise to tracheal functions and structure (101). Nevertheless, immunostaining using osteoclast-specific and chondrocyte-specific markers is required to confirm this hypothesis, as these changes were not obvious upon H&E staining of tracheal tissues in the current study (Supplementary Figure S15).
Downregulation of the “gap junction” class of proteins (Table 4) and the CC term “cell junction” (Figure 5) enriched with several barrier molecules (BMs) in the non-vaccinated-challenged birds suggested impairment to junction complexes (Figure 8). Downregulated BMs included catenins (CTNNA2 and CTNNA3), cadherins (CDH13, CDH11, and CDH5), plakins (PPL and DST), and the tight junction ZO protein TJP1 (102–104). These BMs play integral roles in the formation of various junction complexes, including tight junctions (TJs), adherens junctions (AJs), and desmosomes (105, 106) (Figure 8). Disruption of junction complexes in these birds possibly enhanced the trans-endothelial migration of leukocytes (107) and elevated the chemokine milieu at the site of infection, which aggravated the tracheal pathology (108). This may also have facilitated viral spread outside of the tracheal mucosae (109) of these birds as seen in HSV-1 infection (110) and as evidenced by the detection of ILTV in a range of extra-respiratory and neural tissues reported in numerous previous studies (11, 111–115). Aligning with these findings, the current study demonstrated downregulation of several GO terms related to neuron generation and synapse formation (Figures 5, 8; Supplementary Figures S9, S11, S12, S14) in the non-vaccinated-challenged birds. These results, in combination, could reflect ILTV infection of neurons innervating tracheal tissues (116), consequently inducing the transcriptional modifications of these genes. While it remains unclear whether ILTV infects peripheral nervous structures within the trachea, previous research has shown evidence of latent infection in tracheal tissues (115) as well as in sensory neurons of the trigeminal ganglia (11, 115). Infection of the peripheral neural ganglia innervating the trachea would also be consistent with the impairment of cell junction components at the epithelium, while the persistence of virulent ILTV in non-vaccinated-challenged birds may have prolonged the window of opportunity for neuronal infection in these birds compared to the vaccinated-challenged birds (11). Further studies aimed at detecting and quantifying the virus within neuronal tissues and trigeminal ganglia following ILTV infection of both ΔgG-ILTV vaccinated and non-vaccinated birds are required to validate these findings.
Taking these results in combination, the differences in tracheal transcriptome between the ΔgG-ILTV vaccinated and non-vaccinated-challenged birds aligned with the clinical outcomes observed following challenge with virulent ILTV in these groups of birds. A heightened immunological response and a prolonged viral persistence, together with the pathological changes in mucosa, junctional complexes, and cytoskeletal structures, that impaired ciliary and neuronal functions were associated with virulent ILTV infection in the non-vaccinated-challenged birds. In contrast, vaccination appeared to result in protection from damage to the cellular components and significantly reduced the immune and inflammatory responses associated with ILTV pathology in the vaccinated-challenged birds so that the transcriptomic and clinical profiles of these vaccinated birds after challenge were a little different from those of uninfected birds. It appears that the vaccinated birds attained immune homeostasis shortly after ILTV challenge. Future studies aimed at evaluating the immune responses associated with the protection offered by the ΔgG-ILTV vaccine should focus on assessing the early stages of transcriptional changes after ILTV vaccination and again soon after challenge. These studies have the potential to unveil the specific markers that underline the immune protection triggered by ILTV exposure, whether through vaccination or natural infection. Given the critical role of cell-mediated immune response in ILT disease protection and ILTV-induced pathology, studies investigating tracheal cellular dynamics after vaccination and after virulent ILTV challenge are also warranted.
Data availability statement
The datasets presented in this study can be found online at NCBI, under the accession number: PRJNA1100627 (SRA).
Ethics statement
The animal study was approved by The University of Melbourne Animal ethics committee (approval number: 24993). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
GG: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. MC: Conceptualization, Investigation, Methodology, Supervision, Writing – review & editing, Data curation. AD-M: Conceptualization, Investigation, Methodology, Supervision, Writing – review & editing, Data curation. CH: Conceptualization, Investigation, Methodology, Supervision, Writing – review & editing, Data curation. JD: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing, Data curation.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
Illumina sequencing was performed at The Australian Genome Research Facility (AGRF). The authors acknowledge Dr. Greg Underwood, Bioproperties Pty. Ltd. for his thoughtful contributions to the study design and helpful discussions. The authors acknowledge Dr. Pollob Shil, Ms. June Daly, and Ms. Angela Chircop for the maintenance of experimental chickens and for the help during the animal trial. The authors also thank Dr. Alistair Legione for the insightful suggestions on bioinformatics analysis, Mr. Walter Tsang, Dr. Turgut Aktepe, Mr. Sagar Regmi, and Dr. Paola Vaz for their help with interventions during animal trial and necropsies, and Ms. Faye Docherty for the assistance in tissue processing and staining of histology slide.
Conflict of interest
Authors JD and CH are employed by the University of Melbourne. The University of Melbourne licenses the ΔgG ILT vaccine to Bioproperties Pty. Ltd. As employees of the university involved in the development of the ΔgG ILT vaccine strain, these authors are entitled to a share of any royalties generated from this license.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1458218/full#supplementary-material
References
1. Davison AJ, Eberle R, Ehlers B, Hayward GS, McGeoch DJ, Minson AC, et al. The order herpesvirales. Arch Virology. (2009) 154:171–7. doi: 10.1007/s00705-008-0278-4
2. Guy JS, Bagust TJ. Laryngotracheitis. In: Saif YM, Barnes HJ, Glisson JR, Fadly AM, McDougald LR, Swayne D, editors. Diseases of poultry. Iowa State University Press, USA (2003).
3. Coppo MJ, Noormohammadi AH, Browning GF, Devlin JM. Challenges and recent advancements in infectious laryngotracheitis virus vaccines. Avian Pathology. (2013) 42:195–205. doi: 10.1080/03079457.2013.800634
4. Devlin J, Browning G, Hartley C, Kirkpatrick N, Mahmoudian A, Noormohammadi A, et al. Glycoprotein G is a virulence factor in infectious laryngotracheitis virus. J Gen virology. (2006) 87:2839–47. doi: 10.1099/vir.0.82194-0
5. Devlin JM, Browning GF, Hartley CA, Gilkerson JR. Glycoprotein G deficient infectious laryngotracheitis virus is a candidate attenuated vaccine. Vaccine. (2007) 25:3561–6. doi: 10.1016/j.vaccine.2007.01.080
6. Devlin JM, Viejo-Borbolla A, Browning GF, Noormohammadi AH, Gilkerson JR, Alcami A, et al. Evaluation of immunological responses to a glycoprotein G deficient candidate vaccine strain of infectious laryngotracheitis virus. Vaccine. (2010) 28:1325–32. doi: 10.1016/j.vaccine.2009.11.013
7. Devlin JM, Browning GF, Gilkerson JR, Fenton SP, Hartley CA. Comparison of the safety and protective efficacy of vaccination with glycoprotein-G-deficient infectious laryngotracheitis virus delivered via eye-drop, drinking water or aerosol. Avian Pathology. (2008) 37:83–8. doi: 10.1080/03079450701802214
8. Korsa MG, Browning GF, Coppo MJ, Legione AR, Gilkerson JR, Noormohammadi AH, et al. Protection Induced in Broiler Chickens following Drinking-Water Delivery of Live Infectious Laryngotracheitis Vaccines against Subsequent Challenge with Recombinant Field Virus. PloS One. (2015) 10:e0137719. doi: 10.1371/journal.pone.0137719
9. Korsa MG, Devlin JM, Hartley CA, Browning GF, Coppo MJC, Quinteros JA, et al. Determination of the minimum protective dose of a glycoprotein-G-deficient infectious laryngotracheitis virus vaccine delivered via eye-drop to week-old chickens. PloS One. (2018) 13:e0207611–e. doi: 10.1371/journal.pone.0207611
10. Legione AR, Coppo MJC, Lee S-W, Noormohammadi AH, Hartley CA, Browning GF, et al. Safety and vaccine efficacy of a glycoprotein G deficient strain of infectious laryngotracheitis virus delivered in ovo. Vaccine. (2012) 30:7193–8. doi: 10.1016/j.vaccine.2012.10.023
11. Thilakarathne DS, Coppo MJC, Hartley CA, Diaz-Mendez A, Quinteros JA, Fakhri O, et al. Attenuated infectious laryngotracheitis virus vaccines differ in their capacity to establish latency in the trigeminal ganglia of specific pathogen free chickens following eye drop inoculation. PloS One. (2019) 14:e0213866. doi: 10.1371/journal.pone.0213866
12. Coppo MJC, Devlin JM, Legione AR, Vaz PK, Lee SW, Quinteros JA, et al. Infectious laryngotracheitis virus viral chemokine-binding protein glycoprotein G alters transcription of key inflammatory mediators in vitro and in vivo. J Virology. (2018) 92:10–1128. doi: 10.1128/JVI.01534-17
13. Mahmoudian A, Markham PF, Noormohammadi AH, Devlin JM, Browning GF. Differential transcription patterns in wild-type and glycoprotein G-deleted infectious laryngotracheitis viruses. Avian pathology: J WVPA. (2013) 42:253–9. doi: 10.1080/03079457.2013.780649
14. Blacker HP, Kirkpatrick NC, Rubite A, O’Rourke D, Noormohammadi AH. Epidemiology of recent outbreaks of infectious laryngotracheitis in poultry in Australia. Aust Veterinary J. (2011) 89:89–94. doi: 10.1111/j.1751-0813.2010.00665.x
15. Kirkpatrick NC, Mahmoudian A, Colson CA, Devlin JM, Noormohammadi AH. Relationship between mortality, clinical signs and tracheal pathology in infectious laryngotracheitis. Avian Pathology. (2006) 35:449–53. doi: 10.1080/03079450601028803
16. Kawaguchi T, Nomura K, Hirayama Y, Kitagawa T. Establishment and characterization of a chicken hepatocellular carcinoma cell line, LMH. Cancer Res. (1987) 47:4460–4.
17. Guy JS, Barnes HJ, Morgan LM. Virulence of infectious laryngotracheitis viruses: comparison of modified-live vaccine viruses and North Carolina field isolates. Avian Dis. (1990) 34:106–13. doi: 10.2307/1591340
18. Kulappu Arachchige SN, Young ND, Kanci Condello A, Omotainse OS, Noormohammadi AH, Wawegama NK, et al. Transcriptomic Analysis of Long-Term Protective Immunity Induced by Vaccination With Mycoplasma gallisepticum Strain ts-304. Front Immunol. (2020) 11:628804. doi: 10.3389/fimmu.2020.628804
19. Mahmoudian A, Kirkpatrick NC, Coppo M, Lee SW, Devlin JM, Markham PF, et al. Development of a SYBR Green quantitative polymerase chain reaction assay for rapid detection and quantification of infectious laryngotracheitis virus. Avian Pathology. (2011) 40:237–42. doi: 10.1080/03079457.2011.553582
20. Afgan E, Baker D, Batut B, van den Beek M, Bouvier D, Čech M, et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. (2018) 46:W537–W44. doi: 10.1093/nar/gky379
21. Hiltemann S, Rasche H, Gladman S, Hotz H-R, Larivière D, Blankenberg D, et al. Galaxy Training: A powerful framework for teaching! PloS Comput Biol. (2023) 19:e1010752. doi: 10.1371/journal.pcbi.1010752
22. Batut B, Hiltemann S, Bagnacani A, Baker D, Bhardwaj V, Blank C, et al. Community-driven data analysis training for biology. Cell Systems. (2018) 6:752–8.e1. doi: 10.1016/j.cels.2018.05.012
23. Hasanpur K, Hosseinzadeh S, Mirzaaghayi A, Alijani S. Investigation of chicken housekeeping genes using next-generation sequencing data. Front Genet. (2022) 13:827538. doi: 10.3389/fgene.2022.827538
24. Mi H, Thomas P. PANTHER pathway: an ontology-based pathway database coupled with data analysis tools. Methods Mol Biol. (2009) 563:123–40. doi: 10.1007/978-1-60761-175-2_7
25. Mi H, Muruganujan A, Ebert D, Huang X, Thomas PD. PANTHER version 14: more genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. (2019) 47:D419–d26. doi: 10.1093/nar/gky1038
26. Supek F, Bošnjak M, Škunca N, Šmuc T. REVIGO summarizes and visualizes long lists of gene ontology terms. PloS One. (2011) 6:e21800. doi: 10.1371/journal.pone.0021800
27. Xia C, Braunstein Z, Toomey AC, Zhong J, Rao X. S100 proteins as an important regulator of macrophage inflammation. Front Immunol. (2018) 8:1908. doi: 10.3389/fimmu.2017.01908
28. Meijer B, Gearry R, Day A. The role of S100A12 as a systemic marker of inflammation. Int J Inflammation. (2012) 2012:907078. doi: 10.1155/2012/907078
29. Wu R, Chen F, Wang N, Tang D, Kang R. ACOD1 in immunometabolism and disease. Cell Mol Immunol. (2020) 17:822–33. doi: 10.1038/s41423-020-0489-5
30. Zou L, Zhou J, Zhang J, Li J, Liu N, Chai L, et al. The GTPase Rab3b/3c-positive recycling vesicles are involved in cross-presentation in dendritic cells. Proc Natl Acad Sci U S A. (2009) 106:15801–6. doi: 10.1073/pnas.0905684106
31. Johnson HJ, Rosenberg L, Choi HU, Garza S, Höök M, Neame PJ. Characterization of epiphycan, a small proteoglycan with a leucine-rich repeat core protein. J Biol Chem. (1997) 272:18709–17. doi: 10.1074/jbc.272.30.18709
32. Kim C-H, Kim K, Jik Kim H, Kook Kim J, Lee J-G, Yoon J-H. Expression and regulation of PLUNC in human nasal epithelium. Acta Oto-Laryngologica. (2006) 126:1073–8. doi: 10.1080/00016480600606749
33. Bingle L, Bingle Colin D. Distribution of human PLUNC/BPI fold-containing (BPIF) proteins. Biochem Soc Trans. (2011) 39:1023–7. doi: 10.1042/BST0391023
34. Roughley PJ, Mort JS. The role of aggrecan in normal and osteoarthritic cartilage. J Exp Orthopaedics. (2014) 1:8. doi: 10.1186/s40634-014-0008-7
35. Carvalho MS, Cabral JMS, da Silva CL, Vashishth D. Bone matrix non-collagenous proteins in tissue engineering: creating new bone by mimicking the extracellular matrix. Polymers (Basel). (2021) 13:1095. doi: 10.3390/polym13071095
36. Shum L. Chondroadherin binds to type II collagen. Arthritis Res Ther. (2001) 3:72650. doi: 10.1186/ar-2001-72650
37. Arnott JA, Lambi AG, Mundy C, Hendesi H, Pixley RA, Owen TA, et al. The role of connective tissue growth factor (CTGF/CCN2) in skeletogenesis. Crit Rev Eukaryotic Gene Expression. (2011) 21:43–69. doi: 10.1615/critreveukargeneexpr.v21.i1.40
38. Cunningham NS, Jenkins NA, Gilbert DJ, Copeland NG, Reddi AH, Lee SJ. Growth/differentiation factor-10: a new member of the transforming growth factor-beta superfamily related to bone morphogenetic protein-3. Growth Factors. (1995) 12:99–109. doi: 10.3109/08977199509028956
39. Shapiro IM, Layfield R, Lotz M, Settembre C, Whitehouse C. Boning up on autophagy. Autophagy. (2014) 10:7–19. doi: 10.4161/auto.26679
40. Mayer B, Németh K, Krepuska M, Myneni VD, Maric D, Tisdale JF, et al. Vasopressin stimulates the proliferation and differentiation of red blood cell precursors and improves recovery from anemia. Sci Trans Med. (2017) 9:eaao1632. doi: 10.1126/scitranslmed.aao1632
41. Wang H, Shen Z, Zhou X, Yang S, Yan F, He K, et al. Identification of differentially expressed genes in different types of broiler skeletal muscle fibers using the RNA-seq technique. BioMed Res Int. (2020) 2020:9478949. doi: 10.1155/2020/9478949
42. Fox JC, Nakayama T, Tyler RC, Sander TL, Yoshie O, Volkman BF. Structural and agonist properties of XCL2, the other member of the C-chemokine subfamily. Cytokine. (2015) 71:302–11. doi: 10.1016/j.cyto.2014.11.010
43. DeVries ME, Cao H, Wang J, Xu L, Kelvin AA, Ran L, et al. Genomic organization and evolution of the CX3CR1/CCR8 chemokine receptor locus*. J Biol Chem. (2003) 278:11985–94. doi: 10.1074/jbc.M211422200
44. Cho H-Y, Kleeberger SR. Nrf2 protects against airway disorders. Toxicol Appl Pharmacol. (2010) 244:43–56. doi: 10.1016/j.taap.2009.07.024
45. Wyler E, Franke V, Menegatti J, Kocks C, Boltengagen A, Praktiknjo S, et al. Single-cell RNA-sequencing of herpes simplex virus 1-infected cells connects NRF2 activation to an antiviral program. Nat Commun. (2019) 10:4878. doi: 10.1038/s41467-019-12894-z
46. Schutyser E, Struyf S, Van Damme J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev. (2003) 14:409–26. doi: 10.1016/S1359-6101(03)00049-2
47. Haddadi S, Kim DS, Jasmine H, van der Meer F, Czub M, Abdul-Careem MF. Induction of Toll-like receptor 4 signaling in avian macrophages inhibits infectious laryngotracheitis virus replication in a nitric oxide dependent way. Veterinary Immunol Immunopathology. (2013) 155:270–5. doi: 10.1016/j.vetimm.2013.08.005
48. Ahmed-Hassan H, Abdul-Cader MS, Sabry MA, Hamza E, Abdul-Careem MF. Toll-like receptor (TLR)4 signalling induces myeloid differentiation primary response gene (MYD) 88 independent pathway in avian species leading to type I interferon production and antiviral response. Virus Res. (2018) 256:107–16. doi: 10.1016/j.virusres.2018.08.008
49. Thapa S, Nagy E, Abdul-Careem MF. In ovo delivery of Toll-like receptor 2 ligand, lipoteichoic acid induces pro-inflammatory mediators reducing post-hatch infectious laryngotracheitis virus infection. Veterinary Immunol Immunopathology. (2015) 164:170–8. doi: 10.1016/j.vetimm.2015.02.006
50. Abdul-Cader MS, De Silva Senapathi U, Ahmed-Hassan H, Sharif S, Abdul-Careem MF. Single stranded (ss)RNA-mediated antiviral response against infectious laryngotracheitis virus infection. BMC Microbiol. (2019) 19:34. doi: 10.1186/s12866-019-1398-6
51. Jie H, Lian L, Qu LJ, Zheng JX, Hou ZC, Xu GY, et al. Differential expression of Toll-like receptor genes in lymphoid tissues between Marek’s disease virus-infected and noninfected chickens. Poultry Science. (2013) 92:645–54. doi: 10.3382/ps.2012-02747
52. Feng M, Xie T, Li Y, Zhang N, Lu Q, Zhou Y, et al. A balanced game: chicken macrophage response to ALV-J infection. Veterinary Res. (2019) 50:20. doi: 10.1186/s13567-019-0638-y
53. Lecoultre M, Dutoit V, Walker P. Phagocytic function of tumor-associated macrophages as a key determinant of tumor progression control: A review. J ImmunoTherapy Cancer. (2020) 8:1408. doi: 10.1136/jitc-2020-001408
54. Tang-Huau T-L, Gueguen P, Goudot C, Durand M, Bohec M, Baulande S, et al. Human in vivo-generated monocyte-derived dendritic cells and macrophages cross-present antigens through a vacuolar pathway. Nat Commun. (2018) 9:2570. doi: 10.1038/s41467-018-04985-0
55. Miller MM, Wang C, Parisini E, Coletta RD, Goto RM, Lee SY, et al. Characterization of two avian MHC-like genes reveals an ancient origin of the CD1 family. Proc Natl Acad Sci U S A. (2005) 102:8674–9. doi: 10.1073/pnas.0500105102
56. Salomonsen J, Sørensen MR, Marston DA, Rogers SL, Collen T, van Hateren A, et al. indicating that CD1 genes are ancient and likely to have been present in the primordial MHC. Proc Natl Acad Sci. (2005) 102:8668–73. doi: 10.1073/pnas.0409213102
57. Kelly H, Mandraju R, Coelho-dos-Reis JGA, Tsuji M. Effects of HIV-1-induced CD1c and CD1d modulation and endogenous lipid presentation on CD1c-restricted T-cell activation. BMC Immunol. (2013) 14:4. doi: 10.1186/1471-2172-14-4
58. Cohen NR, Garg S, Brenner MB. Antigen presentation by CD1 lipids, T cells, and NKT cells in microbial immunity. Adv Immunol. (2009) 102:1–94. doi: 10.1016/S0065-2776(09)01201-2
59. Fox LM, Miksanek J, May NA, Scharf L, Lockridge JL, Veerapen N, et al. Expression of CD1c enhances human invariant NKT cell activation by α-GalCer. Cancer Immunity. (2013) 13:9.
60. Rao P, Wen X, Lo JH, Kim S, Li X, Chen S, et al. Herpes simplex virus 1 specifically targets human CD1d antigen presentation to enhance its pathogenicity. J Virol. (2018) 92:10–1128. doi: 10.1128/JVI.01490-18
61. Schönrich G, Raftery MJ. CD1-restricted T cells during persistent virus infections: “Sympathy for the devil. Front Immunolgy. (2018) 9:545. doi: 10.3389/fimmu.2018.00545
62. Kasmar A, Van Rhijn I, Moody DB. The evolved functions of CD1 during infection. Curr Opin Immunol. (2009) 21:397–403. doi: 10.1016/j.coi.2009.05.022
63. Van Rhijn I, Young DC, De Jong A, Vazquez J, Cheng T-Y, Talekar R, et al. CD1c bypasses lysosomes to present a lipopeptide antigen with 12 amino acids. J Exp Med. (2009) 206:1409–22. doi: 10.1084/jem.20082480
64. Loudovaris T, Calnek BW, Yoo BH, Fahey KJ. Genetic susceptibility of chicken macrophages to in vitro infection with infectious laryngotracheitis virus. Avian pathology: J WVPA. (1991) 20:291–302. doi: 10.1080/03079459108418765
65. Watts C. Phagocytosis: how the phagosome became the phag-ER-some. Curr Biol. (2002) 12:R666–R8. doi: 10.1016/S0960-9822(02)01163-6
66. Caron E, Hall A. Identification of two distinct mechanisms of phagocytosis controlled by different Rho GTPases. Science. (1998) 282:1717–21. doi: 10.1126/science.282.5394.1717
67. Lee S, Lee RH, Kim SJ, Lee HK, Na CS, Song KD. Transcriptional regulation of chicken leukocyte cell-derived chemotaxin 2 in response to toll-like receptor 3 stimulation. Asian-Australasian J Anim Sci. (2019) 32:1942–9. doi: 10.5713/ajas.19.0192
68. Lam KM. The macrophage inflammatory protein-1β in the supernatants of Mycoplasma gallisepticum-infected chicken leukocytes attracts the migration of chicken heterophils and lymphocytes. Dev Comp Immunol. (2002) 26:85–93. doi: 10.1016/S0145-305X(01)00053-2
69. Kaiser P, Poh TY, Rothwell L, Avery S, Balu S, Pathania US, et al. A genomic analysis of chicken cytokines and chemokines. J Interferon Cytokine Res. (2005) 25:467–84. doi: 10.1089/jir.2005.25.467
70. Wu Z, Hu T, Kaiser P. Chicken CCR6 and CCR7 are markers for immature and mature dendritic cells respectively. Dev Comp Immunol. (2011) 35:563–7. doi: 10.1016/j.dci.2010.12.015
71. Elshafiee EA, Isham IM, Najimudeen SM, Perez-Contreras A, Barboza-Solis C, Ravi M, et al. Host responses following infection with canadian-origin wildtype and vaccine revertant infectious laryngotracheitis virus. Vaccines. (2022) 10:782. doi: 10.3390/vaccines10050782
72. Hayashi S, Odagiri Y, Kotani T, Horiuchi T. Pathological changes of tracheal mucosa in chickens infected with infectious laryngotracheitis virus. Avian Diseases. (1985) 29:943–50.
73. Purcell DA. The ultrastructural changes produced by infectious laryngotracheitis virus in tracheal epithelium of the fowl. Res Veterinary Science. (1971) 12:455–63.
74. Norman JG, Fink GW, Sexton C, Carter G. Transgenic animals demonstrate a role for the IL-1 receptor in regulating IL-1beta gene expression at steady-state and during the systemic stress induced by acute pancreatitis. J Surg Res. (1996) 63:231–6. doi: 10.1006/jsre.1996.0253
75. Wu B, Jin M, Zhang Y, Wei T, Bai Z. Evolution of the IL17 receptor family in chordates: a new subfamily IL17REL. Immunogenetics. (2011) 63:835–45. doi: 10.1007/s00251-011-0554-4
76. Zhang L, Liu R, Song M, Hu Y, Pan B, Cai J, et al. Eimeria tenella: Interleukin 17 contributes to host immunopathology in the gut during experimental infection. Exp Parasitology. (2013) 133:121–30. doi: 10.1016/j.exppara.2012.11.009
77. Kim WH, Jeong J, Park AR, Yim D, Kim Y-H, Kim KD, et al. Chicken IL-17F: Identification and comparative expression analysis in Eimeria-infected chickens. Dev Comp Immunol. (2012) 38:401–9. doi: 10.1016/j.dci.2012.08.002
78. Hong Y, Lee J, Vu TH, Lee S, Lillehoj HS, Hong YH. Chicken avian β-defensin 8 modulates immune response via the mitogen-activated protein kinase signaling pathways in a chicken macrophage cell line. Poultry Science. (2020) 99:4174–82. doi: 10.1016/j.psj.2020.05.027
79. Niu S, Jahejo AR, Jia FJ, Li X, Ning GB, Zhang D, et al. Transcripts of antibacterial peptides in chicken erythrocytes infected with Marek’s disease virus. BMC veterinary Res. (2018) 14:363. doi: 10.1186/s12917-018-1678-7
80. Hong Y, Lee J, Vu TH, Lee S, Lillehoj HS, Hong YH. Immunomodulatory effects of avian β-defensin 5 in chicken macrophage cell line. Res Veterinary Science. (2020) 132:81–7. doi: 10.1016/j.rvsc.2020.06.002
81. Elhamouly M, Terada T, Nii T, Isobe N, Yoshimura Y. Innate antiviral immune response against infectious bronchitis virus and involvement of prostaglandin E2 in the uterine mucosa of laying hens. Theriogenology. (2018) 110:122–9. doi: 10.1016/j.theriogenology.2017.12.047
82. Xu Y, Zhang T, Xu Q, Han Z, Liang S, Shao Y, et al. Differential modulation of avian β-defensin and Toll-like receptor expression in chickens infected with infectious bronchitis virus. Appl Microbiol Biotechnol. (2015) 99:9011–24. doi: 10.1007/s00253-015-6786-8
83. Liu C, Jiang L, Liu L, Sun L, Zhao W, Chen Y, et al. Induction of avian β-defensin 2 is possibly mediated by the p38 MAPK signal pathway in chicken embryo fibroblasts after newcastle disease virus infection. Front Microbiol. (2018) 9:751. doi: 10.3389/fmicb.2018.00751
84. Vagnozzi AE, Beltrán G, Zavala G, Read L, Sharif S, García M. Cytokine gene transcription in the trachea, Harderian gland, and trigeminal ganglia of chickens inoculated with virulent infectious laryngotracheitis virus (ILTV) strain. Avian pathology: J WVPA. (2018) 47:497–508. doi: 10.1080/03079457.2018.1492090
85. Santhakumar D, Iqbal M, Nair V, Munir M. Chicken IFN kappa: A novel cytokine with antiviral activities. Sci Rep. (2017) 7:2719. doi: 10.1038/s41598-017-02951-2
86. Han Y, Liu D, Li L. PD-1/PD-L1 pathway: current researches in cancer. Am J Cancer Res. (2020) 10:727–42.
87. Kumar V, Yasmeen N, Chaudhary AA, Alawam AS, Al-Zharani M, Suliman Basher N, et al. Specialized pro-resolving lipid mediators regulate inflammatory macrophages: A paradigm shift from antibiotics to immunotherapy for mitigating COVID-19 pandemic. Front Mol Biosciences. (2023) 10:1104577. doi: 10.3389/fmolb.2023.1104577
88. de la Rosa X, Norris PC, Chiang N, Rodriguez AR, Spur BW, Serhan CN. Identification and complete stereochemical assignments of the new resolvin conjugates in tissue regeneration in human tissues that stimulate proresolving phagocyte functions and tissue regeneration. Am J Pathology. (2018) 188:950–66. doi: 10.1016/j.ajpath.2018.01.004
89. Koltsida O, Karamnov S, Pyrillou K, Vickery T, Chairakaki AD, Tamvakopoulos C, et al. Toll-like receptor 7 stimulates production of specialized pro-resolving lipid mediators and promotes resolution of airway inflammation. EMBO Mol Med. (2013) 5:762–75. doi: 10.1002/emmm.201201891
90. Barboza-Solis C, Najimudeen SM, Perez-Contreras A, Ali A, Joseph T, King R, et al. Evaluation of recombinant herpesvirus of Turkey laryngotracheitis (rHVT-LT) vaccine against genotype VI canadian wild-type infectious laryngotracheitis virus (ILTV) infection. Vaccines. (2021) 9:1425. doi: 10.3390/vaccines9121425
91. Hornef N, Olbrich H, Horvath J, Zariwala MA, Fliegauf M, Loges NT, et al. DNAH5 mutations are a common cause of primary ciliary dyskinesia with outer dynein arm defects. Am J Respir Crit Care Med. (2006) 174:120–6. doi: 10.1164/rccm.200601-084OC
92. Tarkar A, Loges NT, Slagle CE, Francis R, Dougherty GW, Tamayo JV, et al. DYX1C1 is required for axonemal dynein assembly and ciliary motility. Nat Genet. (2013) 45:995–1003. doi: 10.1038/ng.2707
93. Carvalho-Santos Z, MaChado P, Branco P, Tavares-Cadete F, Rodrigues-Martins A, Pereira-Leal JB, et al. Stepwise evolution of the centriole-assembly pathway. J Cell science. (2010) 123:1414–26. doi: 10.1242/jcs.064931
94. Insinna C, Lu Q, Teixeira I, Harned A, Semler EM, Stauffer J, et al. Investigation of F-BAR domain PACSIN proteins uncovers membrane tubulation function in cilia assembly and transport. Nat Commun. (2019) 10:428. doi: 10.1038/s41467-018-08192-9
95. Cao W, Ijiri TW, Huang AP, Gerton GL. Characterization of a novel tektin member, TEKT5, in mouse sperm. J andrology. (2011) 32:55–69. doi: 10.2164/jandrol.109.009456
96. Muñoz-Estrada J, Ferland RJ. Ahi1 promotes Arl13b ciliary recruitment, regulates Arl13b stability and is required for normal cell migration. J Cell Sci. (2019) 132:jcs230680. doi: 10.1242/jcs.230680
97. Park DR, Kim J, Kim GM, Lee H, Kim M, Hwang D, et al. Osteoclast-associated receptor blockade prevents articular cartilage destruction via chondrocyte apoptosis regulation. Nat Commun. (2020) 11:4343. doi: 10.1038/s41467-020-18208-y
98. Löfvall H, Newbould H, Karsdal MA, Dziegiel MH, Richter J, Henriksen K, et al. Osteoclasts degrade bone and cartilage knee joint compartments through different resorption processes. Arthritis Res Ther. (2018) 20:67. doi: 10.1186/s13075-018-1564-5
100. Lee JY, Song JJ, Wooming A, Li X, Zhou H, Bottje WG, et al. Transcriptional profiling of host gene expression in chicken embryo lung cells infected with laryngotracheitis virus. BMC Genomics. (2010) 11:1–15. doi: 10.1186/1471-2164-11-445
101. Luo J, Carrillo JA, Menendez KR, Tablante NL, Song J. Transcriptome analysis reveals an activation of major histocompatibility complex 1 and 2 pathways in chicken trachea immunized with infectious laryngotracheitis virus vaccine. Poultry Science. (2014) 93:848–55. doi: 10.3382/ps.2013-03624
102. Künzli K, Favre B, Chofflon M, Borradori L. One gene but different proteins and diseases: the complexity of dystonin and bullous pemphigoid antigen 1. Exp Dermatol. (2016) 25:10–6. doi: 10.1111/exd.2016.25.issue-1
103. Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochim Biophys Acta (BBA) - Rev Cancer. (2008) 1778:660–9. doi: 10.1016/j.bbamem.2007.07.012
104. Lee Y-C, Tsai K-W, Liao J-B, Kuo W-T, Chang Y-C, Yang Y-F. High expression of tight junction protein 1 as a predictive biomarker for bladder cancer grade and staging. Sci Rep. (2022) 12:1496. doi: 10.1038/s41598-022-05631-y
105. Leick KM, Rodriguez AB, Melssen MM, Benamar M, Lindsay RS, Eki R, et al. The barrier molecules junction plakoglobin, filaggrin, and dystonin play roles in melanoma growth and angiogenesis. Ann surgery. (2019) 270:712. doi: 10.1097/SLA.0000000000003522
106. Marchiando AM, Graham WV, Turner JR. Epithelial barriers in homeostasis and disease. Annu Rev Pathology: Mech Disease. (2010) 5:119–44. doi: 10.1146/annurev.pathol.4.110807.092135
107. Muller WA. Mechanisms of leukocyte transendothelial migration. Annu Rev Pathology. (2011) 6:323–44. doi: 10.1146/annurev-pathol-011110-130224
108. Chen ML, Ge Z, Fox JG, Schauer DB. Disruption of tight junctions and induction of proinflammatory cytokine responses in colonic epithelial cells by Campylobacter jejuni. Infection immunity. (2006) 74:6581–9. doi: 10.1128/IAI.00958-06
109. Tugizov S. Virus-associated disruption of mucosal epithelial tight junctions and its role in viral transmission and spread. Tissue Barriers. (2021) 9:1943274. doi: 10.1080/21688370.2021.1943274
110. Sufiawati I, Tugizov SM. HIV-associated disruption of tight and adherens junctions of oral epithelial cells facilitates HSV-1 infection and spread. PloS One. (2014) 9:e88803. doi: 10.1371/journal.pone.0088803
111. Oldoni I, Rodriguez-Avila A, Riblet SM, Zavala G, Garcia M. Pathogenicity and growth characteristics of selected infectious laryngotracheitis virus strains from the United States. Avian Pathology. (2009) 38:47–53. doi: 10.1080/03079450802632031
112. Rodriguez-Avila A, Oldoni I, Riblet S, Garcia M. Replication and transmission of live attenuated infectious laryngotracheitis virus (ILTV) vaccines. Avian Diseases. (2007) 51:905–11 . doi: 10.1637/8011-041907-REGR.1
113. Zhao Y, Kong C, Cui X, Cui H, Shi X, Zhang X, et al. Detection of infectious laryngotracheitis virus by real-time PCR in naturally and experimentally infected chickens. PloS One. (2013) 8:e67598. doi: 10.1371/journal.pone.0067598
114. Wang L-G, Ma J, Xue C-Y, Wang W, Guo C, Chen F, et al. Dynamic distribution and tissue tropism of infectious laryngotracheitis virus in experimentally infected chickens. Arch virology. (2013) 158:659–66. doi: 10.1007/s00705-012-1414-8
115. Bagust TJ, Calnek BW, Fahey KJ. Gallid-1 herpesvirus infection in the chicken. 3. Reinvestigation of the pathogenesis of infectious laryngotracheitis in acute and early post-acute respiratory disease. Avian Diseases. (1986) 30:179–90. doi: 10.2307/1590631
Keywords: infectious laryngotracheitis virus, vaccination-challenge, RNA-sequencing, transcriptome, immune response, gene enrichment analysis, tracheal mucosae
Citation: Gopakumar G, Coppo MJC, Diaz-Méndez A, Hartley CA and Devlin JM (2025) Clinical assessment and transcriptome analysis of host immune responses in a vaccination-challenge study using a glycoprotein G deletion mutant vaccine strain of infectious laryngotracheitis virus. Front. Immunol. 15:1458218. doi: 10.3389/fimmu.2024.1458218
Received: 02 July 2024; Accepted: 17 December 2024;
Published: 24 January 2025.
Edited by:
Amol Suryawanshi, North Carolina State University, United StatesReviewed by:
Jamal Hussen, King Faisal University, Saudi ArabiaChetan Pundkar, Walter Reed Army Institute of Research, United States
Copyright © 2025 Gopakumar, Coppo, Diaz-Méndez, Hartley and Devlin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gayathri Gopakumar, Z2F5YXRocmkuZ29wYWt1bWFyQHVuaW1lbGIuZWR1LmF1