
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 05 September 2024
Sec. Microbial Immunology
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1444469
This article is part of the Research TopicGenetic and Molecular Determinants in Bone Health and Diseases - Volume IIView all 4 articles
 Jia-Qi Zhou1,2,3†
Jia-Qi Zhou1,2,3† Zi-Xian Liu4,5†
Zi-Xian Liu4,5† Hong-Fa Zhong6†
Hong-Fa Zhong6† Guan-Qiao Liu1Ming-Cong Ding4,5
Guan-Qiao Liu1Ming-Cong Ding4,5 Yu Zhang2,3*
Yu Zhang2,3* Bin Yu1*
Bin Yu1* Nan Jiang1,6*
Nan Jiang1,6*Currently, despite advancements in diagnostic and therapeutic modalities, osteomyelitis and prosthetic joint infection (PJI) continue to pose significant challenges for orthopaedic surgeons. These challenges are primarily attributed to the high degree of heterogeneity exhibited by these disorders, which are influenced by a combination of environmental and host factors. Recent research efforts have delved into the pathogenesis of osteomyelitis and PJI by investigating single nucleotide polymorphisms (SNPs). This review comprehensively summarizes the current evidence regarding the associations between SNPs and the predisposition to osteomyelitis and PJI across diverse populations. The findings suggest potential linkages between SNPs in genes such as IL-1, IL-6, IFN-γ, TNF-α, VDR, tPA, CTSG, COX-2, MMP1, SLC11A1, Bax, NOS2, and NLRP3 with the development of osteomyelitis. Furthermore, SNPs in genes like IL-1, IL-6, TNF-α, MBL, OPG, RANK, and GCSFR are implicated in susceptibility to PJI. However, it is noted that most of these studies are single-center reports, lacking in-depth mechanistic research. To gain a more profound understanding of the roles played by various SNPs in the development of osteomyelitis and PJI, future multi-center studies and fundamental investigations are deemed necessary.
Bone and joint infections, such as osteomyelitis and orthopedic implant-associated infections (1), have posed a persistent challenge in the field of orthopedics. Osteomyelitis, characterized by irregular bone neoformation and progressive bone deterioration (2, 3), remains a complex condition, despite the combined efforts of medical and surgical interventions. Alarmingly, even with adequate treatment, up to 30% of osteomyelitis cases may develop into chronic stage, resulting in substantial financial burden, morbidity, and mortality (4).
Total joint replacement stands as a highly effective treatment option for patients suffering from advanced osteoarthritis (5). Nevertheless, certain factors, including aseptic prosthesis loosening and prosthetic joint infection (PJI), can significantly influence the durability of artificial joints (6). Despite the routine employment of contemporary surgical techniques and antibacterial prophylaxis, some surgical patients continue to experience PJI (7). Therefore, a thorough comprehension of the underlying pathogenesis of bone and joint infections is paramount.
The pathogenesis of these infections is intricately linked to both environmental and host components. Notably, there is a growing body of evidence indicating that single nucleotide polymorphisms (SNPs) play a crucial role in the development of bone and joint infections (8, 9). SNPs refer to variations in a single nucleotide that arise at precise locations within the genome, possessing the potential to modulate an individual’s response to disease development (10). The comprehension of the significance of SNPs in the context of osteomyelitis and PJI is paramount, as these genetic alterations can influence the immune response, predisposition to infections, and disease progression (11, 12). The identification of specific SNPs correlated with these infections holds the potential to aid in the development of targeted preventive measures and individualized therapeutic approaches, ultimately enhancing patient outcomes (13). Current research has amassed increasing evidence suggesting that SNPs in various genes, including IL-1, IL-6, IFN-γ, TNF-α, VDR, tPA, CTSG, COX-2, MMP1, SLC11A1, Bax, NOS2, and NLRP3, may contribute to the development of osteomyelitis. Analogously, SNPs in the genes, such as IL-1, IL-6, TNF-α, MBL, OPG, RANK, and GCSFR, have been associated with susceptibility to PJI (Table 1). These genetic insights are fundamental in understanding the diversity of host responses to infections and in identifying patients who may be at a heightened risk. However, it is noteworthy that most studies conducted hitherto have been single-center reports, lacking in-depth mechanistic investigations, thus emphasizing the necessity for future multi-center studies and foundational research.
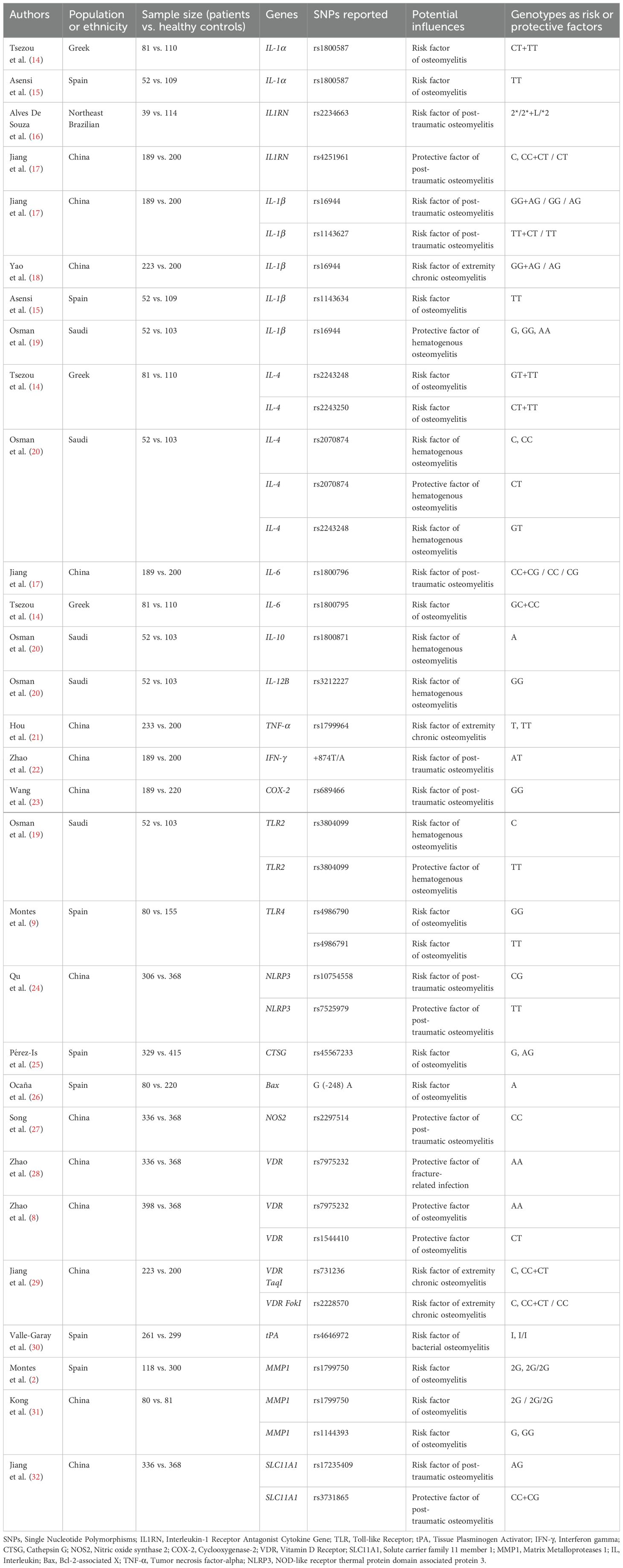
Table 1. SNPs involving in the development of bone and joint infections.
This comprehensive review encapsulates the current evidence pertaining to the associations between SNPs of diverse genes and the predisposition to osteomyelitis and PJI across various populations. It underscores the significant potential of genetic research in augmenting our comprehension and management strategies for bone and joint infections.
Interleukin-1 (IL-1), serving as a pivotal regulator of innate immunity and inflammation, manifests in two distinct forms: IL-1α and IL-1β (16, 18). Notably, IL-1β, the gene encoding the IL-1β cytokine, exhibits a high degree of polymorphism. Furthermore, IL-1 receptor antagonist (IL-1Ra), a naturally occurring competitive inhibitor, binds to the same receptor as IL-1α and IL-1β, yet it does not elicit an inflammatory response (33). Recent investigations have revealed a close association between IL-1 and bone homeostasis as well as bone-related disorders (34).
Extensive research has focused on elucidating the potential associations between the SNPs of IL-1 and the development of osteomyelitis. In a case-control study conducted in 2008 by Tsezou et al. (14), an examination was made of the relationship between IL-1α SNPs and chronic osteomyelitis in a Greek cohort. The study encompassed 81 patients diagnosed with osteomyelitis and 110 healthy controls. The results revealed a nearly sevenfold increase in the risk of osteomyelitis among the TT individuals carrying the IL-1 -889-C/T polymorphism. Comparable findings were observed in a study conducted by Asensi et al. (15) on a Spanish population. Additionally, studies have shown an elevation in IL-1 levels in the serum of rodents suffering from experimental post-traumatic osteomyelitis (PTOM) (35). Consequently, it is plausible to posit that the IL-1 -889-C/T mutation represents a significant risk factor for the development of osteomyelitis. However, given that both aforementioned studies were conducted at a single center and involved a relatively small number of participants, it is imperative to conduct further multicenter studies with a larger sample size to validate these findings.
In 2003, Asensi et al. (15) documented a correlation between the IL-1β gene polymorphism (rs1143634) and the likelihood of developing osteomyelitis in a Spanish cohort. Their findings indicated that individuals with TT genotype exhibited a heightened risk of osteomyelitis compared to those with CT and CC genotypes. A subsequent case-control study conducted by Yao et al. (18) in 2015 revealed a potential association between the IL-1β gene rs16944 and an increased predisposition for chronic osteomyelitis among an Chinese Han population, specifically those with AG genotype. Furthermore, they observed a potential link between the IL-1 gene polymorphism (rs1143627) and an elevated risk of chronic osteomyelitis across dominant, homozygote, and heterozygous models, albeit statistical significance in the comparison between patients and healthy controls was not achieved. Of particular note, Osman et al. (19) conducted a case-control study encompassing 52 patients with hematogenous osteomyelitis and 103 healthy controls, and discovered that the G allele of rs16944 was significantly associated with a diminished risk of hematogenous osteomyelitis in a Saudi population. A recent case-control study (17) reported a heightened propensity for PTOM among individuals with the rs16944 genotypes AG and GG, rs1143627 genotype TT, and rs1800796 genotypes CG and GG. However, their findings regarding the influence of IL-1 gene polymorphism (rs16944) on osteomyelitis susceptibility are inconsistent. This suggests the need for a more nuanced examination of different osteomyelitis subtypes and an independent exploration of the relationships between IL-1β gene polymorphisms and these varying types of osteomyelitis. Additionally, population heterogeneity may serve as a contributing factor.
The presence of variable numbers of tandem repeats (VNTR, 86-bp repeats) within intron 2 of the IL-1 receptor antagonist cytokine (IL-1 RN) gene constitutes a significant genetic polymorphism. Alves De Souza et al. (16) reported a more than twofold increased risk of osteomyelitis in a model dominated by the short allele (IL1RN*2) with two repetitive sequences. Furthermore, their study revealed that individuals harboring the IL1RN*2/*2 + IL1B-511T/T polymorphism genotype were all afflicted with osteomyelitis. The infrequent association and separation of the two polymorphism alleles (IL1RN VNTR and IL1B-511C > T) by 280kb (36) suggests a causal relationship rather than a mere linkage disequilibrium. A study conducted by Jiang et al. (17) in 2020 examined the potential correlation between the IL-1RN SNP rs4251961 and susceptibility to PTOM in a Chinese Han population. The C allele of rs4251961 emerged as an anti-osteomyelitis factor among the 389 participants.
The polymorphisms in the IL-1 gene, by modulating the levels of IL-1 receptor antagonist (IL-1 Ra), IL-1α, and IL-1β at inflammatory sites, significantly influence the inflammatory response, including its initiation, persistence, and resolution (33). However, the precise impact of these polymorphisms on the cytokine levels and the underlying mechanisms contributing to osteomyelitis development remain unclear and warrant further exploration.
In the evolution of chronic osteomyelitis, a crucial aspect is the appearance of necrotic bone fragments (37). It has been definitively established that IL-4 and IL-6 play a direct role in bone resorption and the regulation of osteoclast activity in chronic osteomyelitis (14). Moreover, patients afflicted with osteomyelitis have exhibited elevated levels of IL-4 and IL-6 (38). Recent studies have uncovered a correlation between the development of osteomyelitis and specific SNPs in IL-4 (rs2243248, rs2243250, and rs2070874) as well as SNPs in IL-6 (rs1800795, rs1800796).
In a case-control study conducted in 2008, Tsezou et al. (14) observed a significant association between two polymorphisms of the IL-4 gene (rs2243248 and rs2243250) and elevated risks of osteomyelitis. Specifically, individuals carrying the T allele exhibited a heightened risk of developing osteomyelitis. Additionally, the study found that individuals with the homozygote for the C allele of IL-6 rs1800795 C allele were at an increased risk of developing osteomyelitis. In 2015, Osman et al. (20) reported that in an Saudi population, the IL-4 rs2243248 genotype GT was associated with a greater likelihood to develop hematogenous osteomyelitis. They further discovered that the heterozygous genotype (CT) of IL-4 rs2070874 exhibited protective effects compared to the CC and TT genotypes, while the C allele was identified as a risk factor for hematogenous osteomyelitis. Jiang et al. (17) observed a correlation between the IL-6 gene rs1800796 and an augmented vulnerability to PTOM, where individuals with CG and CC genotypes had a higher propensity for developing this condition. However, it is noteworthy that these findings are derived from single-center studies with limited sample sizes, thus necessitating further confirmation through multicenter studies encompassing larger sample sizes in the future. While numerous studies have uncovered associations between IL-4 and IL-6 SNPs and various inflammatory diseases, the underlying mechanisms remain unclear.
IL-10 was initially identified for its capability to restrain the activation and effector functions of immune cells (e.g. T cells, monocytes, and macrophages) (39). Beyond its traditional function of limiting and ultimately terminating inflammatory responses, IL-10 has been identified as a pivotal player in the differentiation and functioning of T regulatory cells (39). Recent investigations have uncovered a correlation between IL-10 SNPs and a spectrum of inflammatory disorders, such as sepsis (40), osteomyelitis (20), and pancreatitis (41).
Osman et al. (20) conducted a rigorous case-control study in 2015 to investigate potential association between IL-10 SNP rs1800871 polymorphism and the predisposition to hematogenous osteomyelitis among a Saudi population. The analysis of 155 subjects revealed that the presence of the A allele of the IL-10 rs1800871 variant constituted a significant risk factor for the development of hematogenous osteomyelitis. Furthermore, a study by Hofmann et al. (42) demonstrated that individuals afflicted with chronic recurrent multifocal osteomyelitis exhibited diminished IL-10 expression, which subsequently triggered the NLRP3 inflammasome and contributed to inflammatory bone loss. These cumulative findings suggest that IL-10 SNP rs1800871, through its ability to modulate IL-10 levels and further influence the activation of the NLRP3 inflammasome, may exert an influence on the progression of osteomyelitis. However, further in-depth mechanistic studies are imperative to elucidate these complex interactions.
Interleukin-12 (IL-12) is a cytokine that promotes inflammation, produced primarily by non-T cells, especially antigen-presenting cells integral to the T-helper 1 system (43). IL-12 exerts its effects primarily on T cells and natural killer cells, exhibiting numerous biological effects. The IL-12B gene is responsible for encoding a component of IL-12. Recent investigations have identified a linkage between variations in the IL-12B gene and various infectious diseases, including chronic hepatitis B (43), Q fever caused by Coxsackie burnetii (44), and chronic osteomyelitis (20). Among these variations, the SNP locus rs3212227 has garnered significant attention in research.
In a case-control study conducted in 2015, Osman et al. (20) conducted an analysis to examine the potential association between SNP rs1800871 in the IL-12B gene and susceptibility to hematogenous osteomyelitis among a Saudi population. The findings revealed that individuals possessing GG genotype exhibited a heightened risk of developing osteomyelitis in comparison to those with GT and TT genotypes. While Osman et al. did not delve deeper into the underlying mechanisms, research has demonstrated that endogenous IL-12 production or the exogenous administration of recombinant IL-12 enhances the body’s immunity against a diverse array of cellular pathogens (45). Furthermore, a substantial component of this defense mechanism appears to be influenced by IL-12, which prompts T lymphocytes and natural killer cells to produce interferon (46). This increased interferon stimulates macrophages and T lymphocytes, thereby enabling them to more effectively eliminate pathogens from invaded cells (47). S. aureus, the most prevalent pathogen in all forms of osteomyelitis, has the ability to colonize the bone matrix and be internalized and retained by osteoblasts, potentially contributing to the development of chronic infection (45). Consequently, polymorphisms in the IL-12B gene may prevent the development of osteomyelitis by modulating serum IL-12 levels and the IL-12/interferon pathway. However, further research is imperative as the current understanding of the relationship between osteomyelitis and IL-12B polymorphisms remains limited.
Tumor necrosis factor (TNF), a crucial proinflammatory cytokine primarily secreted by macrophages, plays a pivotal role in the progression of numerous infectious disorders. Alterations in the DNA sequence within regulatory regions can potentially alter transcriptional regulation, thereby affecting TNF levels in the bloodstream and augmenting vulnerability to various infectious diseases (48). Recent investigations have indicated that genetic variants in the promoter region of the TNF-α gene, specifically SNPs, may modulate the secretion of TNF-α (49). Among the most extensively studied TNF-α gene polymorphism is rs1799964.
In a case-control study of 433 subjects (233 patients with extremity chronic osteomyelitis and 200 healthy controls), Hou et al. (21) reported that the frequency of the T allele in rs1799964 was significantly higher in case group subjects than that in the control subjects, indicating that having the rs1799964 T allele might be a potential risk factor. Additionally, they determined that there was a statistical connection between the quantity of leukocytes in patients and the TNF-α gene polymorphism rs1799964. The potential link between TNF-α SNP and leukocyte counts needs to be further investigated. Ma et al. (50) found that during S. aureus infection, elevated TNF-α enhanced the expression of endogenous miR-129-5p, inhibiting endothelial nitric oxide synthase (eNOS), proving the role of TNF-α/miR-129-5p/eNOS in the pathogenesis of osteomyelitis. There was no evidence to support either the leukocyte/inflammatory factor route or the TNF-α/miR-129-5p/eNOS pathway as the mechanism by which TNF-α gene polymorphisms caused osteomyelitis. Moreover, considering that the study by Hou et al. was a single-center study with a limited number of participants, there is a need for future multicenter studies with larger sample sizes.
Interferon gamma (IFN-γ) serves as a pivotal immunomodulatory factor, derived from T cells and natural killer cells (NK cells), in the context of immune responses (51). IFN-γ plays a crucial role in the anti-infective immune response to infection, primarily through augmenting antigen presentation and phagocytosis in macrophages (52). Additionally, IFN-γ has been employed in the therapeutic management of certain chronic and inflammatory disorders (53, 54), thereby indicating its potential significance in the immune regulation of osteomyelitis (55). However, it is noteworthy that genetic polymorphisms can lead to variations in IFN-γ production levels and immune responsiveness, ultimately influencing an individual’s resilience to pathogens.
Zhao et al. (22) conducted a study which revealed that the A allele in the IFN-γ +874T/A polymorphism appears to heighten an individual’s predisposition towards PTOM. Specifically, the presence of the A allele, particularly in AT heterozygotes, was observed to significantly increase the likelihood of osteomyelitis development compared to the TT heterozygote. Furthermore, the research suggests that genetic variations in IFN-γ +874T/A may modulate an individual’s vulnerability to PTOM by influencing serum IFN-γ concentrations. Specifically, the A allele is postulated to be associated with reduced IFN-γ levels, potentially impairing the ability to mount an effective antimicrobial response and subsequently elevating the risk of PTOM. In essence, the IFN-γ +874T/A gene polymorphisms may serve as a potential marker for future genetic screening and personalized therapeutic approaches in the context of PTOM.
Toll-like receptors (TLRs) function as pattern recognition receptors that identify pathogen-associated molecular patterns and subsequently trigger the immune system and host defense-stimulating activities (56). TLRs influenced the release of pro-inflammatory cytokines by interacting with pathogen-associated molecular patterns (19). However, excessive release of pro-inflammatory factors inhibited tissue regeneration. Recent research has demonstrated that TLR SNPs play a role in the development of osteomyelitis.
In a case-control study published in 2016, Osman et al. (19) examined the relationship between the TLR2 gene polymorphism rs3804099 and hematogenous osteomyelitis susceptibility in a Saudi population. They noted that people with C allele had a higher risk to develop osteomyelitis. It was reported by Montes et al. (9) that osteomyelitis was linked to the TLR4 polymorphism rs4986790’s GG genotype, regardless of the frequency of the G allele. This finding would suggest that the mutation has a dosage effect and needs both alleles to have the full pathogenic effect. Similar results were found in the TLR4 polymorphism rs4986791. It has been reported that aureus peptidoglycan promotes osteoclast formation through TLR2-mediated activation of the nuclear factor kappa-B (NF-κB)/nuclear factor of activated T cells 1 (NFATc1) signaling pathway (57), causing bone destruction and the formation of osteomyelitis. The association between TLR2 polymorphisms and vulnerability to osteomyelitis and this signaling pathway, however, has not been proven. Neutrophils typically have a short half-life and pass away by apoptosis. Further research by Montes et al. (9) revealed that individuals with the G allele of the TLR4 (rs4986790) polymorphism have considerably lower neutrophil apoptosis and reactivity to lipopolysaccharide, which may help explain the persistent nature of bone infections. The specific mechanism influencing the osteomyelitis pathogenesis requires to be identified to fully understand the association between TLR polymorphisms and osteomyelitis susceptibility.
Vitamin D receptor (VDR) is encoded by the VDR gene located on chromosome 12. VDR participates in several biological processes, including the modulation of immune response and bone metabolism (28, 29). High polymorphism existed in the VDR gene. Studies already conducted have shown that VDR gene variations are linked to an increased risk of contracting several inflammatory disorders, including tuberculosis (58), chronic periodontitis (59), and leprosy (60). The VDR gene variations TaqI (rs731236), BsmI (rs1544410), ApaI (rs7975232), and FokI (rs2228570) have been the subjects of the most studies.
In a 2016 case-control study, Jiang et al. (29) reported the association between the VDR gene polymorphisms rs731236 and rs2228570 and susceptibilities to chronic osteomyelitis in a Chinese population and found that the frequencies of the C allele in both rs731236 and rs2228570 were significantly higher in subjects in the case group than in those in the control group. Furthermore, the homozygote model revealed a substantial association between the likelihood of acquiring chronic osteomyelitis and the gene polymorphisms rs731236 and rs2228570. Therefore, people having the C allele in the VDR gene polymorphisms rs731236 and rs2228570 were more likely to develop osteomyelitis. By using recessive and homozygote models, Zhao et al. (28) observed a significant connection between the VDR gene SNP rs7975232 and a risk of developing fracture-related infections, suggesting that individuals with the AA genotype at this location are less vulnerable to these infection. Zhao et al. (8) similarly found that rs7975232 was associated with susceptibility to osteomyelitis, with the AA genotype as a protective factor. In addition to this, the heterozygote model showed a significant correlation between rs1544410 and susceptibility to osteomyelitis, suggesting that the CT genotype may have a protective effect against osteomyelitis. Through in vivo and in vitro experiments, Zhao et al. (8) also showed that the protective effect of vitamin D against osteomyelitis may be attained in part by preventing macrophage death by inhibiting excessive reactive oxygen species (ROS) production by the VDR-Bmi1 signaling pathway.
Tissue plasminogen activator (tPA), encoded by the human gene t-PA located on chromosome 8p11.21, is a key protease of the fibrinolytic system (61). The tPA Alu insertion/deletion [I/D] is a 311 bp sequence inserted/deleted in the 8th intron of the tPA gene (30). Studies that are now available have demonstrated definite associations between tPA polymorphism and numerous disorders, such as stroke (61) and bacterial infections (62).
In a 2013 case-control study, Valle-Garay et al. (30) reported that the tPA polymorphism (rs4646972) was associated with an increased susceptibility to bacterial osteomyelitis in a Spanish population, with the I allele as a risk factor. They also noted that patients with osteomyelitis had a considerably higher prevalence of the I/I genotype compared to controls. The release rate of tPA was considerably greater in people with the I/I genotype than in those with other genotypes, according to Jern et al.’s (63) study of subjects with various genotypes of the tPA polymorphism (rs4646972). This accelerated rate of tPA release may be involved in the development of osteomyelitis through an in vivo process. However, further studies are needed.
Cathepsin G (CTSG), encoded by the human gene CTSG gene located on chromosome 14q11.2, is a 26-kDa serine protease (25). CTSG was hypothesized to be connected to tissue remodeling at locations of tissue damage, neutrophil response to different pathogens, and blood coagulation (64). In a 2019 study, Pérez-Is et al. (25) investigated the relationship between the N125S polymorphism of the CTSG gene and the risk of developing osteomyelitis, and the results suggested that the G allele and the AG genotype are risk factors for the development of osteomyelitis. In individuals with the G allele, they also discovered a substantial rise in serum CTSG activity and lactoferrin levels. The CTSG gene N125S polymorphism may therefore increase osteomyelitis susceptibility by increasing lactoferrin and CTSG activity. The precise molecular mechanism is still unclear, though. Pyroptosis, a process of inflammatory cell death, can trigger a potent inflammatory response to protect the host from microbial infection (65). A protein produced by Mycobacterium tuberculosis known as Rv3364c has been discovered to bind to host CTSG and prevent pyroptosis in infected macrophages, allowing bound mycobacteria to survive in macrophages (66). It is unclear if osteomyelitis development is governed by analogous mechanisms. To prove this, more research is required.
Cyclooxygenase-2 (COX-2) is encoded by the COX-2 gene located on chromosome 1q25.2-25.3 with a transcript of 4.5 kb (67). In the presence of COX, free arachidonic acid is converted to prostaglandins, regulating the inflammatory response process (68). COX-2 polymorphisms have been reported to be associated with a variety of inflammatory diseases, such as inflammatory bowel disease (69), periodontitis (70), and asthma (71). Wang et al. (23) investigated potential associations of COX-2 gene polymorphisms with susceptibility to osteomyelitis in a Chinese population. Results from 409 subjects (189 patients and 220 controls) showed a significant association between rs689466 and PTOM, with GG genotype as a risk factor. They also compared whether there were statistical differences in the levels of the six cytokines among the people with different genotypes to clarify the potential mechanisms of this polymorphism in the pathogenesis of osteomyelitis. But there were no statistically significant differences. Through an in vivo research, Johansen et al. (72) reported that COX-2-mediated prostaglandins play a role in early osteomyelitis bone resorption. However, the underlying mechanisms of COX-2 polymorphisms in the pathogenesis of osteomyelitis remain unclear.
Nitric oxide synthase 2 (NOS2), also known as inducible nitric oxide synthase (iNOS), is one of the three key enzymes in the production of nitric oxide (NO) in the human body (73). It has been shown that in chronic inflammatory diseases such as rheumatoid arthritis (74), asthma (75), and inflammatory bowel disease (76), polymorphisms in the NOS2 gene may affect its expressions and activities, thereby influencing an individual’s susceptibility to inflammatory diseases (77). The study by Song et al. (27) included 336 patients with PTOM and 368 healthy controls. The rs2297514 and rs2248814 loci in the NOS2 gene of the participants were genotyped by SNaPshot genotyping methods. It was found that the frequency of the C allele at the rs2297514 locus was significantly lower in the group of PTOM patients than that in the group of healthy controls (48.7% vs. 54.5%), suggesting that the C allele may have a protective role in reducing the susceptibility to PTOM. In addition, the median level of CRP was significantly lower in PTOM patients with the CC genotype than in those with the TT genotype (4.1 mg/L vs. 8.9 mg/L, P = 0.027), suggesting that the CC genotype may exert its protective effect by influencing the severity of the inflammatory response. However, no significant association was found between rs2248814 and PTOM susceptibility. This finding provides further evidence of potential links between polymorphisms in the NOS2 gene and the pathogenesis of PTOM and may contribute to the development of future preventive and therapeutic strategies for such diseases.
Matrix metalloproteases (MMPs) are zinc-dependent protein hydrolases that break down different extracellular matrix protein components and are crucial for both healthy physiological processes and pathological conditions (31, 78). One of the crucial MMPs, MMP1, is capable of dissolving type I collagen fibers (79). The emergence of chronic inflammation is strongly correlated with the MMP1 gene SNP (80). In a case-control study in 2010, Montes et al. (2) reported that the MMP1 polymorphism (rs1799750) was associated with an increased susceptibility to osteomyelitis in a Spanish population, with the 2G allele as a risk factor. The study by Kong et al. (31) produced similar outcomes. Along with rs1799750, Kong et al. (31) additionally observed that the G allele of the MMP1 polymorphism rs1144393 increased the incidence of osteomyelitis. The serum of osteomyelitis patients included high levels of inflammatory mediators such TNF-α and IL-1, which mediated enhanced MMP expression (2). In cells with the 2G allele (rs1799750), Cao et al. (80) discovered changes in MMP1 protein and transcript levels. However, no additional research into the underlying molecular pathways has been done. Furthermore, research on the MMP1 rs1144393 polymorphism’s pathogenic mechanism is scarce.
Bcl-2-associated X protein (Bax), encoded by the BAX gene of the BCL-2 gene family, is the predominant apoptotic gene in humans and functions by forming a heterodimer with Bcl-2 protein (81). A critical element in regulating the potency of the inhibitory impact on apoptosis is the ratio between the Bax/Bcl-2 proteins (82). The expression and function of the Bax gene are impacted by mutations in its coding area and promoter (26). In a case-control study in 2007, Ocaña et al. (26) investigated the association between the Bax polymorphism rs4645878 and chronic osteomyelitis in a Spanish population. A total of 80 patients with osteomyelitis as well as 220 healthy controls were included. The findings demonstrated that individuals with the A allele had a greater incidence of osteomyelitis occurence, lower expression of the Bax protein level, and a longer median survival time for peripheral neutrophils. More research is required to better understand how this polymorphism’s anti-apoptotic impact influences susceptibility to osteomyelitis.
NOD-like receptor thermal protein domain associated protein 3 (NLRP3) is an important pattern recognition receptor in the cytoplasm that, together with apoptosis-associated speck-like protein containing a CARD (ASC) and pro-cysteinyl aspartate specific proteinase-1 (pro-caspase-1), forms the NLRP3 inflammatory vesicle (83). The NLRP3 protein is encoded by a gene with nine exons that is found on chromosome 1q44 (24). Pro-inflammatory cytokines mature and pyroptosis was induced when NLRP3 inflammatory vesicles were activated (83). However, excessive activation of NLRP3 activation also promoted the development of a number of inflammatory disorders, including type 2 diabetes, inflammatory bowel disease, and gouty arthritis (84, 85). In a case-control study conducted in 2023, Qu et al. (24) discovered that the NLRP3 gene polymorphisms rs10754558 and rs7525979 may be linked to PTOM susceptibility in an Chinese Han population, with the CG genotype (rs10754558) population possibly being at high risk and the TT genotype (rs7525979) population being protective. Although the pathogenic mechanism of the NLRP3 polymorphism (rs10754558) has not been further explored by Qu et al., Zhang et al. (86) found that the polymorphism rs10754558 may be involved in regulating the immune and inflammatory response in patients with primary gouty arthritis by affecting the expression of NLRP3 inflammasome components. Whether the NLRP3/IL-1β signaling pathway is involved in susceptibility to osteomyelitis due to NLRP3 polymorphism (rs10754558) is unclear. In a study by von Herrmann et al. (87), it was reported that the NLRP3 polymorphism rs7525979 was connected to a considerably decreased risk of Parkinson’s disease development. They also showed that the polymorphism rs7525979 influences the translation efficiency of the NLRP3 protein, which in turn affects how stable, ubiquitinated, and soluble it is. Further research is necessary to determine whether comparable pathways are present in models of osteomyelitis.
Solute Carrier Family 11 Member 1 (SLC11A1), is a gene that encodes a transmembrane protein that is expressed in macroscopic immune cells such as macrophages and plays an important role in the transport of iron and other metal ions (88). Polymorphisms in the SLC11A1 gene may affect the function of this protein and thus the susceptibility to some diseases (89–92). The study by Jiang et al. (32) included 336 PTOM patients and 368 healthy controls. Participants were genotyped to assess the prevalence of the rs17235409 and rs3731865. Results indicated that rs17235409 was associated with an increased risk of developing PTOM. Specifically, the AG genotype of rs17235409 was found to increase risk, and patients with the AG genotype had relatively higher levels of inflammatory biomarkers than those with the AA and GG genotypes, particularly white blood cell counts and C-reactive protein levels. This suggests that this genotype may be a risk factor of PTOM development. Whereas rs3731865 was found to possibly reduce the risk of PTOM, although the results were not statistically significant. In conclusion, this study demonstrates a potential link between specific genetic variants in the SLC11A1 gene and susceptibilities to PTOM, emphasizing the importance of genetic factors in the development of this disease. However, more research is needed to fully understand these mechanisms and confirm these findings, especially for rs3731865.
IL and TNF-α are pivotal inflammatory mediators that play a fundamental role in the underlying mechanisms of aseptic prosthesis loosening (APL)/aseptic failure of arthroplasty, as well as in the pathogenesis of PJI (93). IL-1, IL-6 and TNF-α are closely related to osteoclastogenesis and thus play an important role in bone remodeling (94, 95). Levels of IL-1, IL-6, and TNF-α were observed to be elevated in individuals who have experienced arthroplasty failure (96). Furthermore, cytokines associated with the Th-17 immune response, which are instrumental in providing immune protection against microorganisms, are also potential genetic candidates that may confer an increased susceptibility to PJI (97).
In a 2012 case-control study, Stahelova et al. (98) investigated the relationships between genetic polymorphisms of IL-1β, IL-6, and TNF-α and susceptibilities to PJI, and enrolled 471 subjects (89 PJI vs. 214 APL vs. 168 healthy controls). They found that the IL-1β gene T allele of polymorphism rs16944 was a risk factor for PJI development, while there were no significant associations between IL-6 (-174 G/C, A/G nt565) or TNF-α (rs1800629, rs361525) SNPs and PJI development. Similarly, Malik et al. (99) failed to find definite link between IL-6-174 SNP and PJI occurrence. However, in a prospective study, Erdemli et al. (100) observed that the frequency of IL-6-174 C allele among the PJI patients was significantly higher than that among the APL patients.Meanwhile, they found that the G allele of the TNF-α polymorphism rs361525, IL1RN*2/*2 and IL1RN*1/*2 genotypes were associated with increased susceptibilities to PJI. Racial heterogeneity could potentially be a contributing factor to this discrepancy. Furthermore, the research conducted by Erdemli et al. was structured as a prospective investigation and did not incorporate a comparison group of healthy individuals. Furthermore, López-Anglada et al. (93) investigated potential associations between IL-1β SNP rs1143634 and susceptibilities to PJI in a Spanish population. Based on an analysis of 117 PJI patients, 77 APL patients, and 145 healthy controls, the findings revealed that, in comparison to the healthy controls, the presence of the IL-1β SNP rs1143634 T allele and the TT genotype were identified as risk factors for APL. Conversely, no such associations were observed among the PJI patients. Additionally, in line with the findings of Stahelova et al. (98), López-Anglada et al. (93) reported that no positive correlation was established between rs1800629 (TNF-α gene) and the development of PJI. Navratilova et al. (101) indicated that, despite observing an increased Th-17 immune response among individuals with PJI, no significant positive correlations were established between cytokine polymorphisms associated with Th-17 immunological reactivity and the susceptibility to PJI.
Mannose-binding lectin (MBL), a serum protein derived from the liver, is intricately involved in the activation process of the complement pathway. Additionally, it elicits the activation of macrophages through the mediation of the C1q receptor. Consequently, MBL occupies a pivotal position in innate immunity, underscoring its critical role in the body’s natural defense mechanisms (102). Siassi et al. (103) observed that diminished levels of MBL may elevate the potential for postoperative infection, irrespective of whether they display an immediate reaction to surgical trauma. Furthermore, there exists a strong correlation between reduced MBL levels and SNPs within the MBL2 gene, which is located on chromosome 10 (104). Clinical research has demonstrated that these SNPs are linked to increased risks of developing inflammatory conditions, such systemic lupus erythematosus (105) and arthritis (106). Malik et al. (102) conducted a rigorous investigation into the correlations between MBL SNPs located at the promoter position -500 G/C (MBL-550 G/C) and codon 54 G/A (MBL-54 G/A) and their potential impacts on the susceptibilities to PJI in a cohort from the UK. Their findings indicated that the C allele and the CC genotype of the -550 SNP, as well as the -54 GG genotype, serve as risk factors for the development of PJI. Similarly, Navratilova et al. (6) indicated that there were significant increases in the frequencies of the L allele and LL genotype of the MBL-550 SNP among the PJI patients, as compared to the controls and the APL patients. Furthermore, it was observed that individuals carrying the L allele exhibited lower serum MBL concentrations than those who did not carry the allele. This finding suggests that such a genetic variant may contribute to an elevated risk of PJI, potentially through the regulation of serological MBL levels. However, it is unfortunate to note that no subsequent studies were undertaken to elucidate the underlying molecular mechanisms linking low MBL levels to the occurrence of PJI.
Aside from IL, TNF-α, and MBL, polymorphisms in certain genes, including TLRs, osteoprotegerin (OPG), receptor activator of NF-kappaB (RANK), VDR, and granulocyte colony stimulating factor receptor (GCSFR), have been examined to determine their potential correlations with PJI development. Given the scarcity of research in this area and the fact that some of these genes have already been previously investigated, we shall delve into their discussions collectively in this context.
Despite TLRs’ pivotal role in innate immunity, the research conducted by Mrazek et al. (107) indicated that the presence of multiple genetic polymorphisms within TLRs did not amplify the predisposition to PJI. Interestingly, El-Helou and colleagues (108), through rigorous in vitro experimentation, observed that the TLR2 R753Q SNP effectively suppressed the cellular response triggered by S. aureus peptidoglycan. Furthermore, osteoclastogenesis exhibits a profound link with OPG and RANK, and it is noteworthy that bacterial infections may contribute to bone loss in the context of periprosthetic prostheses for total joint arthroplasty by initiating a series of reactions mediated by the OPG/RANK/RANKL pathway (109). In a case-control study conducted by Malik et al. (5), a significant association was observed between the A allele and AA genotype of the OPG SNP rs3102735, as well as the T allele and TT genotype of the RANK SNP at position 575 in exon 6 (RANK + 575 C/T), with APL. Additionally, the study revealed that, when compared to the control group, the A allele and AA genotype of the OPG SNP rs3102735 served as risk factors for the development of PJI.
The SNPs in the promoter region of the OPG gene can affect the expression levels of OPG proteins by modulating transcription, whereas the SNP in the RANK gene may alter the functionality of the resulting protein product. Malik et al. (99) observed that, when compared to controls, the presence of the T allele and the TT genotype of VDR Taqman Test (VDR-T) assay reagents intensified osteolysis caused by deep infections. Granulocyte Colony Stimulating Factor (GCSF), an inflammatory cytokine, binds to the GCSF receptor (GCSFR), thereby modulating inflammatory processes. Erdemli et al. (100) noted a significant association between the CT genotype of GCSFR and PJI. Nevertheless, research on these genetic polymorphisms remains limited in elucidating the precise relationship between them and the development of PJI. Consequently, there is a need for additional multicenter studies involving larger sample sizes.
Detecting SNPs associated with osteomyelitis and PJI allows for an accurate assessment of risk for individuals, enabling earlier intervention and the implementation of preventive measures for high-risk populations (24). In the case of osteomyelitis, key SNPs within genes such as IL-1β (including rs1143634 and rs16944), IL-6 (e.g., rs1800795), VDR, and TNF-α are instrumental in evaluating susceptibility. Similarly, for PJI, significant SNPs encompass those located in the MBL gene (-550 G/C, -54 G/A), the IL-1β gene (rs16944), and specific polymorphisms of the IL-6 gene. By integrating genetic screening into clinical protocols, healthcare professionals can more effectively identify individuals with an elevated risk, customize personalized treatment plans, and administer targeted preventive strategies, thereby enhancing patient outcomes and mitigating the occurrence of these infections.
The comprehension of the functional role of specific SNPs in osteomyelitis and PJI serves as a valuable tool in the development of tailored treatment strategies, which ultimately enhances therapeutic efficacy and mitigates undesired side effects. In the case of high-risk patients harboring IL-1 gene polymorphisms, such as rs16944, the utilization of IL-1 receptor antagonists, such as Anakinra, can significantly attenuate inflammatory responses (110). Furthermore, the identification of individuals with TLR gene polymorphisms may suggest an increased susceptibility to certain antibiotics, enabling the customization of antibiotic regimens to bolster effectiveness and diminish the potential for antibiotic resistance (111).
In cases where patients are identified as possessing high-risk gene polymorphisms, it is advisable to adopt more stringent monitoring and care protocols both before and after surgical procedures to mitigate the potential risk of postoperative infection (112). Prior to surgery, preparatory measures for patients with high-risk PJI profiles may entail prophylactic antibiotic therapy tailored to their specific genetic makeup, thus substantially diminishing the likelihood of infection (113). Following surgical procedures, individuals with specific SNP should undergo rigorous management strategies that incorporate close monitoring to promptly identify and address any signs of infection. This entails conducting regular blood tests, imaging assessments, and swift intervention upon the earliest manifestation of infection symptoms. The execution of these genotype-specific strategies is aimed at enhancing the quality of patient care, while also fostering a more efficient allocation of medical resources. Consequently, this approach contributes to the reduction of overall healthcare costs and the optimization of long-term outcomes for patients afflicted with osteomyelitis and PJI.
Investigating the precise mechanisms of SNPs possesses considerable potential to propel the progression of novel targeted therapies, thereby providing patients with more efficacious and individualized treatment approaches. For instance, the investigation of genetic variations within the TNF-α gene, such as the rs1799964 polymorphism, has the capability to enable the development of therapeutic strategies that specifically modulate TNF-α activity. Additionally, medications like infliximab and adalimumab, which serve as TNF-α inhibitors, have demonstrated efficacy in managing osteomyelitis and mitigating inflammatory responses (114, 115). Furthermore, for individuals who possess MBL gene polymorphisms that lead to decreased MBL levels, the development of MBL replacement therapies holds the potential to fortify immune responses, consequently mitigating the risk of infection and optimizing overall patient wellbeing (116, 117).
Similarly, the application of precise inhibitors that specifically target genetic variations in the IL-6 gene, notably the rs1800795 polymorphism, effectively reduces inflammation and arrests the progression of osteomyelitis and PJI. Medications, such as tocilizumab and sarilumab, which function as antagonists to the IL-6 receptor, have exhibited substantial therapeutic efficacy in these scenarios (118, 119). By leveraging these genetic insights, healthcare providers are capable of developing highly personalized therapeutic strategies that target the underlying genetic factors of these conditions, thereby enhancing the effectiveness and individualization of patient care.
Genetic polymorphism data can be leveraged to precisely calibrate drug dosages, thereby optimizing therapeutic efficacy while minimizing the occurrence of undesirable side effects (120, 121). Specifically, the adjustment of anti-inflammatory medication dosages in accordance with the genetic variations of TNF-α and IL-6 genes ensures that patients receive individualized, optimal, and effective treatments that are specifically tailored to their unique genetic profiles (122, 123). This tailored strategy significantly improves the efficacy of the treatment while markedly reducing the potential for adverse reactions and complications.
The identification of individuals harboring high-risk gene polymorphisms enables the implementation of comprehensive preventative measures in both lifestyle adjustments and medical oversight, thereby minimizing the likelihood of disease occurrence (124). It is imperative to provide focused health education to these individuals, highlighting the significance of maintaining optimal hygiene, avoiding traumatic injuries, and adhering to recommended wound care practices (125). Furthermore, the introduction of regular health screenings for high-risk populations serves to facilitate the timely detection and prompt management of potential infections (126). These screenings may encompass routine blood testing, imaging studies, and physical examinations, aimed at detecting early indicators of osteomyelitis or PJI (127, 128). Genetic risk factors-guided early intervention strategies can significantly enhance health outcomes by halting the progression of these conditions and minimizing the occurrence of severe complications.
SNPs occupy a pivotal role in the diagnosis, therapeutic intervention, and preventative measures pertaining to osteomyelitis and PJI. Genetic testing presents an opportunity for the implementation of tailored risk assessment and individualized treatment approaches, which subsequently enhance treatment effectiveness and minimize the incidence of complications. Further advancements in research endeavors and clinical applications possess the potential to refine these strategies and improve patient prognosis.
In summary, SNPs located in the genes of IL-1α (rs1800587), IL-1β (rs16944, rs1143627, rs1143634), IL1RN (rs2234663, rs4251961), IL-4 (rs2243248, rs2243250, rs2070874), IL-6 (rs1800795, rs1800796), IL-10 (rs1800871), IL-12B (rs3212227), TLR2 (rs3804099), TNF-α (rs1799964), tPA (rs4646972), CTSG (rs45567233), COX-2 (rs689466), VDR (rs731236, rs2228570, rs7975232, rs1544410), MMP1 (rs1799750, rs1144393), NLRP3 (rs10754558, rs7525979) and Bax (G (-248) A) may play an important role in the development of osteomyelitis. SNPs located in the genes of IL-1β (rs16944, rs1143634), IL1RN (rs2234663), IL-6 (-174 G/C, A/G nt565), TNF-α (rs361525), MBL-550 G/C, MBL-54 G/A, MBL2 (rs11003125), OPG (rs3102735), RANK + 575 C/T, VDR-T, GCSFR may be involved in the PJI (Figure 1, Table 1).
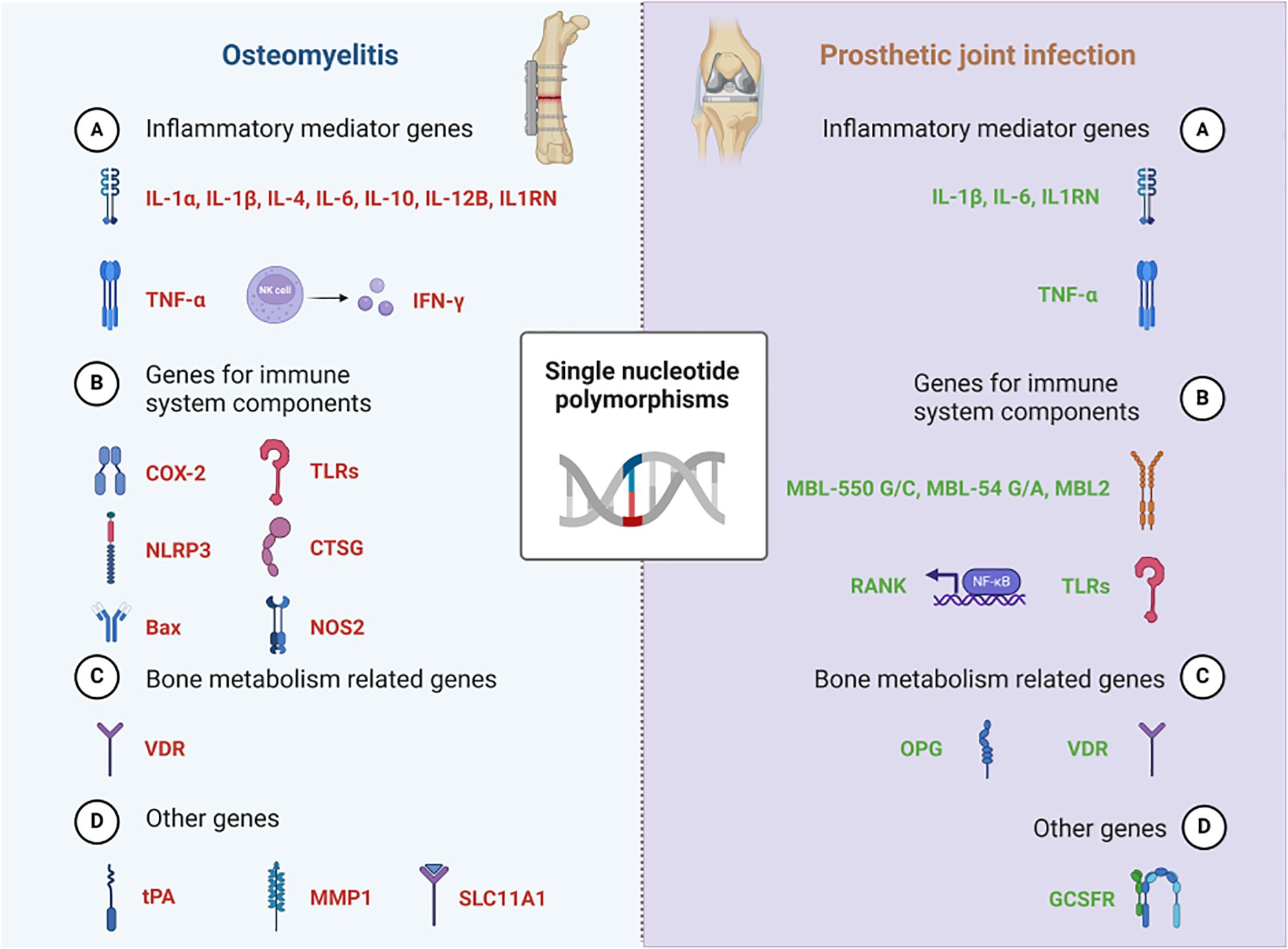
Figure 1. SNPs involved in Osteomyelitis & Prosthetic joint infection.
In the realm of research pertaining to osteomyelitis and PJI, SNPs have yielded pivotal insights into the intricate interplay between genetic predisposition and the susceptibility to, as well as the progression of, these diseases. Despite remarkable strides made in this field, the investigation of SNPs remains confronted with a multitude of constraints and hurdles within the clinical arena. The subsequent paragraphs delve into these limitations in a comprehensive manner, and further propose avenues for future exploration (Figure 2).
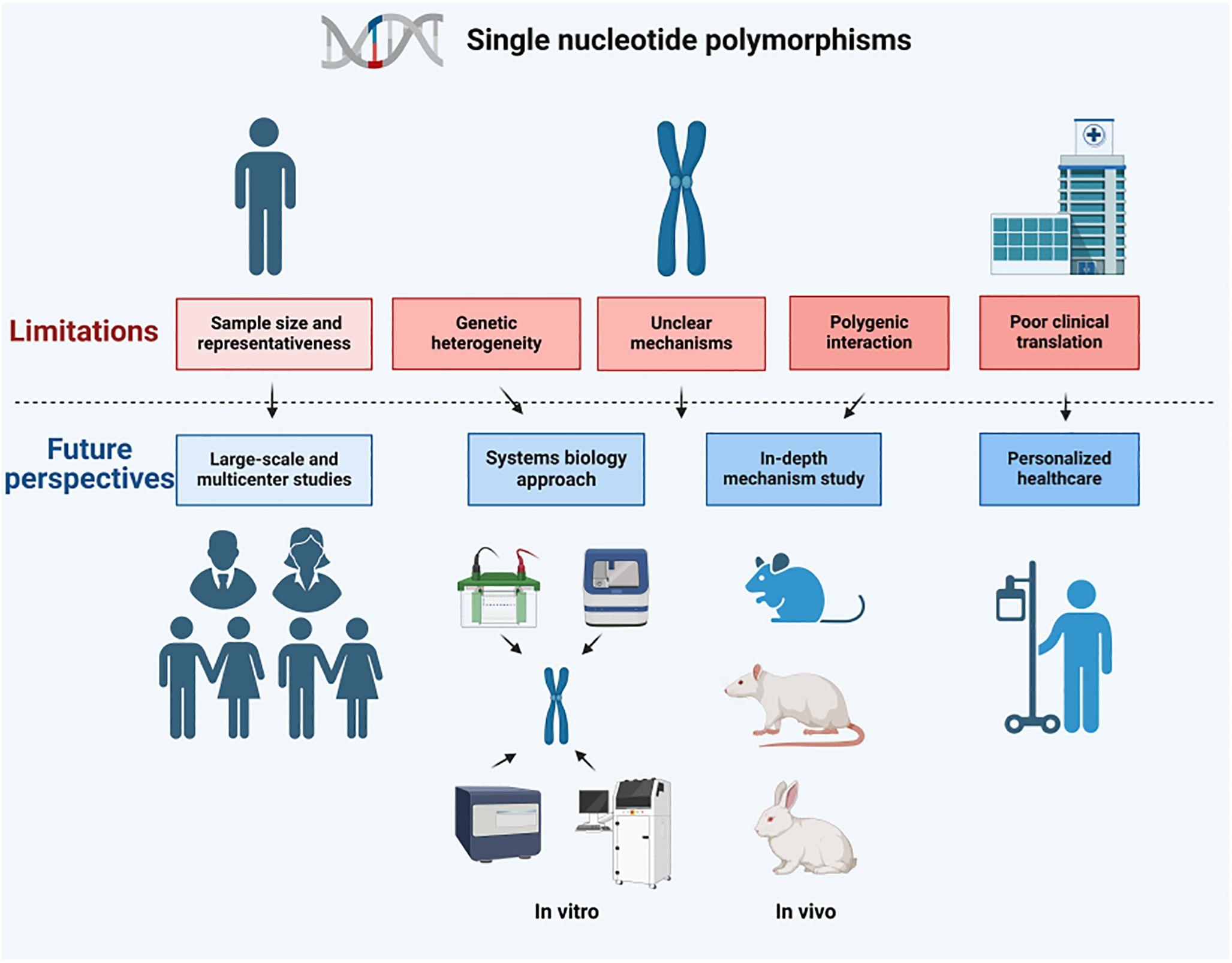
Figure 2. Limitations and future perspectives for SNPs.
(1) Numerous investigations into the relationship between SNPs and osteomyelitis or PJI have been undertaken, relying predominantly on small sample sizes. This approach, however, carries the risk of introducing statistical bias and confines the applicability of the findings. Furthermore, these studies tend to concentrate on specific racial or geographical cohorts, neglecting a holistic, multiracial, and multiregional perspective. The majority of rigorous and methodical research endeavors exploring the correlation between SNPs and both osteomyelitis and PJI have primarily aimed to illuminate the intricate interplay between genetic variations and the predispositions to these diseases. These studies generally explore the potential connections between various SNP genotypes and the probability of disease development, while limited efforts have been directed towards an exhaustive evaluation of sample characteristics, considering the differing prevalence rates among distinct SNP genotypes. (2) A discernible gap is evident in the realm of subgroup analyses, where only a limited number of studies have briefly acknowledged patient age and other relevant susceptibility factors. However, these variables have rarely been subjected to rigorous examination. In particular, the systematic investigation of the characteristic patterns of age groups under varying SNP genotypes and their subsequent impacts on disease progression remains scarce. Consequently, this limitation restricts the breadth and depth of research endeavors aimed at elucidating the influence of diverse genotypes on disease predisposition. (3) Another significant constraint lies in the inherent limitations of the primary data. Studies that have undertaken direct comparisons of the occurrences of osteomyelitis and PJI across various SNP genotypes have frequently refrained from divulging comprehensive patient information. This lack of detailed data has, as a result, obstructed further statistical investigations, thereby restricting our capacity to delve deeply into the specific pathological characteristics and their contributory factors within the framework of distinct SNP genotypes. (4) Even for individuals with the same disease, their genetic backgrounds exhibit significant variations (129–131). Given the genetic heterogeneity, it becomes challenging to discern the distinct impacts of individual SNPs amidst other confounding genetic and environmental factors (132). (5) Despite the identification of numerous SNPs that are linked to osteomyelitis and PJI, the precise biological pathways through which these SNPs influence disease progression remain elusive. The absence of thorough mechanistic investigations hinders the effective translation of this genetic knowledge into clinical practice. (6) Multi-gene interactions play a pivotal role in the development of osteomyelitis and PJI, which are typically not attributed to a solitary genetic variant, but rather to a complex interplay of multiple genes (133, 134). Current research often ignores the complexity of this genetic network. (7) Despite the identification of correlations between genes and diseases, there persist formidable challenges in converting this knowledge into viable diagnostic and therapeutic methodologies (135, 136), especially in genetic counseling, risk assessment, and treatment decisions (137).
(1) Large-scale and multi-center studies: The representativeness of the study and the reliability of the results can be enhanced by expanding the sample size and covering more races and regions. Such studies can help define more precisely the impacts of SNPs on disease susceptibilities. Furthermore, researchers are strongly encouraged to provide comprehensive raw data, thereby enabling the scientific community to conduct further analysis and validation, ultimately leading to a deeper understanding of the intricate relationships between SNPs and disease susceptibilities.
(2) Systems biology approach: Adopting a rigorous systems biology methodology to investigate the intricate interplay between multiple SNPs and other genetic elements within a network framework can potentially uncover the complex underlying genetic architecture of diseases. Furthermore, the implementation of sophisticated subgroup analysis methodologies is intended to clarify the impacts of age, pathological conditions, and other pertinent variables on disease predisposition among individuals with diverse SNP genotypes.
(3) In-depth mechanistic studies: To investigate the impact of particular SNPs on the progression of diseases, research endeavors should delve into their mechanisms of modulating cell signaling, governing gene expression regulation, and influencing other biological pathways. This can be achieved through rigorous experimentation utilizing in vitro systems and animal models, ensuring a comprehensive and scientific approach to elucidating their role in disease development.
(4) Preclinical and translational research: To advance the field of preclinical and translational research, it is essential to intensify efforts in exploring individualized preventive and therapeutic strategies that are grounded in SNPs. This includes the development of tailored drug therapies and the construction of predictive models, both of which hold the potential to significantly enhance the precision and effectiveness of medical interventions.
(5) Interdisciplinary collaboration: It is advocated for the advancement of genetic research pertaining to osteomyelitis and PJI through the concerted efforts and integration of expertise from various disciplines, such as genetics, immunology, microbiology, and clinical medicine.
JZ: Validation, Writing – original draft. ZL: Validation, Writing – original draft. HZ: Funding acquisition, Validation, Writing – review & editing. GL: Funding acquisition, Validation, Writing – review & editing. MD: Validation, Writing – review & editing. YZ: Conceptualization, Writing – review & editing. BY: Conceptualization, Funding acquisition, Writing – review & editing. NJ: Conceptualization, Funding acquisition, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the National Natural Science Foundation of China (grant nos: 82172197, 82272517), Guangdong Basic and Applied Basic Research Foundation (grant nos: 2022A1515012385, 2023A1515140162) and the Ganzhou City Science and Technology Plan Project (grant no. 2023LNS17537).
We would like to thank the BioRender.com for drawing the figures of our study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
SNPs, Single Nucleotide Polymorphisms; PJI, Prosthetic joint infection; PTOM, Post-traumatic osteomyelitis; APL, Aseptic prosthesis loosening; IL1RN, Interleukin-1 Receptor Antagonist Cytokine Gene; TLR, Toll-like Receptor; tPA, Tissue Plasminogen Activator; CTSG, Cathepsin G; NOS2, Nitric oxide synthase 2; iNOS, inducible nitric oxide synthase; eNOS, endothelial nitric oxide synthase; NO, Nitric oxide; ROS, Reactive oxygen species; COX-2, Cyclooxygenase-2; VDR, Vitamin D Receptor; SLC11A1, Solute carrier family 11 member 1; MMP1, Matrix Metalloproteases 1; IL, Interleukin; IFN-γ, Interferon gamma; Bax, Bcl-2-associated X; TNF-α, Tumor necrosis factor-alpha; NLRP3, NOD-like receptor thermal protein domain associated protein 3; RANK, NF-kappaB; GCSF, Granulocyte colony stimulating factor; GCSFR, Granulocyte colony stimulating factor receptor; OPG, Osteoprotegerin.
1. Morata L, Cobo J, Fernández-Sampedro M, Guisado Vasco P, Ruano E, Lora-Tamayo J, et al. Safety and efficacy of prolonged use of dalbavancin in bone and joint infections. Antimicrob Agents Chemother. (2019) 63. doi: 10.1128/AAC.02280-18
2. Montes AH, Valle-Garay E, Alvarez V, Pevida M, García Pérez E, Paz J, et al. A functional polymorphism in MMP1 could influence osteomyelitis development. J Bone Mineral Res. (2010) 25:912–9. doi: 10.1359/jbmr.091013
3. Xie X, Li J, Gu F, Zhang K, Su Z, Wen Q, et al. Genetic determinants for bacterial osteomyelitis: A focused systematic review of published literature. Front Genet. (2021) 12:654792. doi: 10.3389/fgene.2021.654792
4. Lew DP, Waldvogel FA. Osteomyelitis. Lancet. (2004) 364:369–79. doi: 10.1016/S0140-6736(04)16727-5
5. Malik MH, Bayat A, Jury F, Ollier WE, Kay PR. Genetic susceptibility to hip arthroplasty failure–association with the RANK/OPG pathway. Int Orthop. (2006) 30:177–81. doi: 10.1007/s00264-006-0074-2
6. Navratilova Z, Gallo J, Mrazek F, Lostak J, Petrek M. MBL2 gene variation affecting serum MBL is associated with prosthetic joint infection in czech patients after total joint arthroplasty. Tissue Antigens. (2012) 80:444–51. doi: 10.1111/tan.12001
7. Jämsen E, Furnes O, Engesaeter LB, Konttinen YT, Odgaard A, Stefánsdóttir A, et al. Prevention of deep infection in joint replacement surgery. Acta Orthop. (2010) 81:660–6. doi: 10.3109/17453674.2010.537805
8. Zhao XQ, Wan HY, He SY, Qin HJ, Yu B, Jiang N. Vitamin d receptor genetic polymorphisms associate with a decreased susceptibility to extremity osteomyelitis partly by inhibiting macrophage apoptosis through inhibition of excessive ROS production via VDR-bmi1 signaling. Front Physiol. (2022) 13:808272. doi: 10.3389/fphys.2022.808272
9. Montes AH, Asensi V, Alvarez V, Valle E, Ocaña MG, Meana A, et al. The toll-like receptor 4 (Asp299Gly) polymorphism is a risk factor for gram-negative and haematogenous osteomyelitis. Clin Exp Immunol. (2006) 143:404–13. doi: 10.1111/j.1365-2249.2005.03002.x
10. Bahreini F, Rayzan E, Rezaei N. microRNA-related single-nucleotide polymorphisms and breast cancer. J Cell Physiol. (2021) 236:1593–605. doi: 10.1002/jcp.29966
11. Gowin E, Januszkiewicz-Lewandowska D. Genes and their single nucleotide polymorphism involved in innate immune response in central nervous system in bacterial meningitis: review of literature data. Inflammation Res. (2018) 67:655–61. doi: 10.1007/s00011-018-1158-3
12. Skevaki C, Pararas M, Kostelidou K, Tsakris A, Routsias JG. Single nucleotide polymorphisms of toll-like receptors and susceptibility to infectious diseases. Clin Exp Immunol. (2015) 180:165–77. doi: 10.1111/cei.12578
13. Grezzana GB, Vieira JL, Portal VL. Single-nucleotide polymorphisms: a perspective of cardiovascular prevention. Rev Assoc Med Bras (1992). (2015) 61:458–68. doi: 10.1590/1806-9282.61.05.458
14. Tsezou A, Poultsides L, Kostopoulou F, Zintzaras E, Satra M, Kitsiou-Tzeli S, et al. Influence of interleukin 1alpha (IL-1alpha), IL-4, and IL-6 polymorphisms on genetic susceptibility to chronic osteomyelitis. Clin Vaccine Immunol. (2008) 15:1888–90. doi: 10.1128/CVI.00209-08
15. Asensi V, Alvarez V, Valle E, Meana A, Fierer J, Coto E, et al. IL-1 alpha (-889) promoter polymorphism is a risk factor for osteomyelitis. Am J Med Genet A. (2003) 119A:132–6. doi: 10.1002/ajmg.a.20137
16. Alves De Souza C, Queiroz Alves De Souza A, Queiroz Alves De Souza MDS, Dias Leite JA, Silva De Morais M, Barem Rabenhorst SH. A link between osteomyelitis and IL1RN and IL1B polymorphisms-a study in patients from northeast brazil. Acta Orthop. (2017) 88:556–61. doi: 10.1080/17453674.2017.1348439
17. Jiang N, Li SY, Ma YF, Hu YJ, Lin QR, Yu B. Associations between interleukin gene polymorphisms and risks of developing extremity posttraumatic osteomyelitis in chinese han population. Mediators Inflamm. (2020) 2020:3278081. doi: 10.1155/2020/3278081
18. Yao ZL, Lin QR, Hu YJ, Hou YL, Ma YF, Yu B, et al. Interleukin-1 beta gene polymorphism rs16944 may associate with increased susceptibility to extremity chronic osteomyelitis in chinese han population. BioMed Res Int. (2019) 2019:7483537. doi: 10.1155/2019/7483537
19. Osman AE, Mubasher M, ElSheikh NE, AlHarthi H, AlZahrani MS, Ahmed N, et al. Association of single nucleotide polymorphisms in proinflammatory cytokine and toll-like receptor genes with pediatric hematogenous osteomyelitis. Genet Mol Res. (2016) 15. doi: 10.4238/gmr.15027718
20. Osman AE, Mubasher M, ElSheikh NE, AlHarthi H, AlAlallah IA, Elbeshir AA, et al. Investigation of polymorphisms in anti-inflammatory cytokine genes in hematogenous osteomyelitis. Genet Mol Res. (2015) 14:16981–6. doi: 10.4238/2015.December.15.4
21. Hou Y, Bai L, Jiang N, Yao Z, Xue L, Yu B. Screening of TNF-a gene polymorphisms in patients with extremity chronic osteomyelitis in china. Per Med. (2018) 15:395–401. doi: 10.2217/pme-2018-0017
22. Zhao XQ, Jiang N, Hu YJ, Yu B. IFN-γ +874T/A polymorphism increases susceptibility to posttraumatic osteomyelitis. Int J Immunogenet. (2020) 47:163–8. doi: 10.1111/iji.12462
23. Wang L, Jiang N, Lin QR, Qin CH, Hu YJ, Yu B. Cyclooxygenase-2 (COX-2) polymorphism rs689466 may contribute to the increased susceptibility to post-traumatic osteomyelitis in chinese population. Infect Dis (Lond). (2017) 49:817–23. doi: 10.1080/23744235.2017.1347816
24. Qu Y, Li J, Zhang W, Xia C, Ou S, Yang Y, et al. Posttraumatic osteomyelitis risks associated with NLRP3 gene polymorphisms in the chinese population. J Pers Med. (2023) 13:253. doi: 10.3390/jpm13020253
25. Pérez-Is L, Ocaña MG, Montes AH, Carton JA, Álvarez V, Meana Á, et al. The N125S polymorphism in the cathepsin g gene (rs45567233) is associated with susceptibility to osteomyelitis in a spanish population. PloS One. (2019) 14:e0220022. doi: 10.1371/journal.pone.0220022
26. Ocaña MG, Valle-Garay E, Montes AH, Meana A, Cartón JA, Fierer J, et al. Bax gene g(-248)A promoter polymorphism is associated with increased lifespan of the neutrophils of patients with osteomyelitis. Genet Med. (2007) 9:249–55. doi: 10.1097/GIM.0b013e318039b23d
27. Song CS, Zhang P, Lin QR, Hu YY, Pan CQ, Jiang N, et al. Nitric oxide synthase 2 genetic variation rs2297514 associates with a decreased susceptibility to extremity post-traumatic osteomyelitis in a chinese han population. Front Cell Infect Microbiol. (2023) 13:1177830. doi: 10.3389/fcimb.2023.1177830
28. Zhao XQ, Chen K, Wan HY, He SY, Qin HJ, Yu B, et al. Vitamin d receptor genetic variations may associate with the risk of developing late fracture-related infection in the chinese han population. J Immunol Res. (2022) 2022:9025354. doi: 10.1155/2022/9025354
29. Jiang N, Zhao XQ, Qin CH, Hu YJ, Wang L, Xie GP, et al. Association of vitamin d receptor gene TaqI, BsmI, FokI and ApaI polymorphisms and susceptibility to extremity chronic osteomyelitis in chinese population. Injury. (2016) 47:1655–60. doi: 10.1016/j.injury.2016.06.005
30. Valle-Garay E, Montes AH, Corte JR, Meana A, Fierer J, Asensi V. tPA alu (I/D) polymorphism associates with bacterial osteomyelitis. J Infect Dis. (2013) 208:218–23. doi: 10.1093/infdis/jit158
31. Kong Q, Jin Y, Yan S, Wang Y, Zhao J, Feng Z, et al. Examining the association of MMP-1 gene -1607 (2G/1G) and -519 (A/G) polymorphisms with the risk of osteomyelitis: A case-control study. Med (Baltimore). (2017) 96:e4969. doi: 10.1097/MD.0000000000004969
32. Jiang N, Zhong YC, Lin QR, Song CS, Yu B, Hu YJ. Solute carrier family 11 member 1 genetic polymorphisms rs17235409 and rs3731865 associate with susceptibility to extremity post-traumatic osteomyelitis in a chinese han population. Int J Immunogenet. (2023) 50:127–33. doi: 10.1111/iji.12620
33. Witkin SS, Gerber S, Ledger WJ. Influence of interleukin-1 receptor antagonist gene polymorphism on disease. Clin Infect Dis. (2002) 34:204–9. doi: 10.1086/338261
34. Lukens JR, Gross JM, Calabrese C, Iwakura Y, Lamkanfi M, Vogel P, et al. Critical role for inflammasome-independent IL-1b production in osteomyelitis. Proc Natl Acad Sci USA. (2014) 111:1066–71. doi: 10.1073/pnas.1318688111
35. Kobayashi N, Kadono Y, Naito A, Matsumoto K, Yamamoto T, Tanaka S, et al. Segregation of TRAF6-mediated signaling pathways clarifies its role in osteoclastogenesis. EMBO J. (2001) 20:1271–80. doi: 10.1093/emboj/20.6.1271
36. Wang Y, Shumansky K, Sin DD, Man SF, Akhabir L, Connett JE, et al. Associations of interleukin-1 gene cluster polymorphisms with c-reactive protein concentration and lung function decline in smoking-induced chronic obstructive pulmonary disease. Int J Clin Exp Pathol. (2015) 8:13125–35.
37. Pesanti EL, Lorenzo JA. Osteoclasts and effects of interleukin 4 in development of chronic osteomyelitis. Clin Orthop Relat Res. (1998) 1998:290–9. doi: 10.1097/00003086-199810000-00031
38. Yoshii T, Magara S, Miyai D, Nishimura H, Kuroki E, Furudoi S, et al. Local levels of interleukin-1beta, -4, -6 and tumor necrosis factor alpha in an experimental model of murine osteomyelitis due to staphylococcus aureus. Cytokine. (2002) 19:59–65. doi: 10.1006/cyto.2002.1039
39. Moore KW, de Waal Malefyt R, Coffman RL, O'Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol. (2001) 19:683–765. doi: 10.1146/annurev.immunol.19.1.683
40. Zhang N, Wang S, Fan Y, Sheng C, Ge W. Association between IL10 polymorphisms and the susceptibility to sepsis: A meta-analysis. Biochem Genet. (2023) 61:847–60. doi: 10.1007/s10528-022-10310-8
41. Dong L, Zhu Y. Association between IL-10 polymorphisms rs1800896 and rs1800871 and risk of acute pancreatitis in chinese han population: An update systemic review and meta-analysis. Niger J Clin Pract. (2019) 22:149–53. doi: 10.4103/njcp.njcp_57_18
42. Hofmann SR, Kubasch AS, Ioannidis C, Rösen-Wolff A, Girschick HJ, Morbach H, et al. Altered expression of IL-10 family cytokines in monocytes from CRMO patients result in enhanced IL-1b expression and release. Clin Immunol. (2015) 161:300–7. doi: 10.1016/j.clim.2015.09.013
43. Ben Dhifallah I, Ayouni K, Jmel H, Kammoun W, Hamzaoui K, Sadraoui A, et al. Strong association of functional polymorphism in IL-12B with susceptibility to chronic hepatitis B in tunisia. J Med Virol. (2021) 93:4949–56. doi: 10.1002/jmv.26946
44. Schoffelen T, Textoris J, Bleeker-Rovers CP, Ben Amara A, van der Meer JW, Netea MG, et al. Intact interferon-g response against coxiella burnetii by peripheral blood mononuclear cells in chronic Q fever. Clin Microbiol Infect. (2017) 23:209.e9–209.e15. doi: 10.1016/j.cmi.2016.11.008
45. Bost KL, Ramp WK, Nicholson NC, Bento JL, Marriott I, Hudson MC. Staphylococcus aureus infection of mouse or human osteoblasts induces high levels of interleukin-6 and interleukin-12 production. J Infect Dis. (1999) 180:1912–20. doi: 10.1086/315138
46. Manetti R, Gerosa F, Giudizi MG, Biagiotti R, Parronchi P, Piccinni MP, et al. Interleukin 12 induces stable priming for interferon gamma (IFN-gamma) production during differentiation of human T helper (Th) cells and transient IFN-gamma production in established Th2 cell clones. J Exp Med. (1994) 179:1273–83. doi: 10.1084/jem.179.4.1273
47. Trinchieri G. Interleukin-12: a cytokine at the interface of inflammation and immunity. Adv Immunol. (1998) 70:83–243. doi: 10.1016/S0065-2776(08)60387-9
48. Qidwai T, Khan F. Tumour necrosis factor gene polymorphism and disease prevalence. Scand J Immunol. (2011) 74:522–47. doi: 10.1111/j.1365-3083.2011.02602.x
49. Sezgin M, Barlas IO, Ankarali HC, Altintaş ZM, Türkmen E, Gökdoğan T, et al. Tumour necrosis factor alpha -308G/A gene polymorphism: lack of association with knee osteoarthritis in a turkish population. Clin Exp Rheumatol. (2008) 26:763–8.
50. Ma X, Xia W, Zong Y, Jiang C, Shan H, Lin Y, et al. Tumor necrosis factor-a promotes staphylococcus aureus-induced osteomyelitis through downregulating endothelial nitric oxide synthase. J Microbiol Immunol Infect. (2021) 54:1018–27. doi: 10.1016/j.jmii.2020.08.002
51. Jorgovanovic D, Song M, Wang L, Zhang Y. Roles of IFN-γ in tumor progression and regression: a review. biomark Res. (2020) 8:49. doi: 10.1186/s40364-020-00228-x
52. Clark DN, Begg LR, Filiano AJ. Unique aspects of IFN-γ/STAT1 signaling in neurons. Immunol Rev. (2022) 311:187–204. doi: 10.1111/imr.13092
53. Kursunel MA, Esendagli G. The untold story of IFN-γ in cancer biology. Cytokine Growth Factor Rev. (2016) 31:73–81. doi: 10.1016/j.cytogfr.2016.07.005
54. Tang M, Tian L, Luo G, Yu X. Interferon-Gamma-Mediated osteoimmunology. Front Immunol. (2018) 9:1508. doi: 10.3389/fimmu.2018.01508
55. Tsumura M, Miki M, Mizoguchi Y, Hirata O, Nishimura S, Tamaura M, et al. Enhanced osteoclastogenesis in patients with MSMD due to impaired response to IFN-γ. J Allergy Clin Immunol. (2022) 149:252–61.e6. doi: 10.1016/j.jaci.2021.05.018
56. Fitzgerald KA, Kagan JC. Toll-like receptors and the control of immunity. Cell. (2020) 180:1044–66. doi: 10.1016/j.cell.2020.02.041
57. Cao F, Zhou W, Liu G, Xia T, Liu M, Mi B, et al. Staphylococcus aureus peptidoglycan promotes osteoclastogenesis via TLR2-mediated activation of the NF-κB/NFATc1 signaling pathway. Am J Transl Res. (2017) 9:5022–30.
58. Mohammadi A, Khanbabaei H, Nasiri-Kalmarzi R, Khademi F, Jafari M, Tajik N. Vitamin d receptor ApaI (rs7975232), BsmI (rs1544410), Fok1 (rs2228570), and TaqI (rs731236) gene polymorphisms and susceptibility to pulmonary tuberculosis in an iranian population: A systematic review and meta-analysis. J Microbiol Immunol Infect. (2020) 53:827–35. doi: 10.1016/j.jmii.2019.08.011
59. Deng H, Liu F, Pan Y, Jin X, Wang H, Cao J. BsmI, TaqI, ApaI, and FokI polymorphisms in the vitamin d receptor gene and periodontitis: a meta-analysis of 15 studies including 1338 cases and 1302 controls. J Clin Periodontol. (2011) 38:199–207. doi: 10.1111/j.1600-051X.2010.01685.x
60. Pepineli AC, Alves HV, Tiyo BT, Macedo LC, Visentainer L, de Lima Neto QA, et al. Vitamin d receptor gene polymorphisms are associated with leprosy in southern brazil. Front Immunol. (2019) 10:2157. doi: 10.3389/fimmu.2019.02157
61. Babu MS, Prabha TS, Kaul S, Al-Hazzani A, Shafi G, Roy S, et al. Association of genetic variants of fibrinolytic system with stroke and stroke subtypes. Gene. (2012) 495:76–80. doi: 10.1016/j.gene.2011.12.046
62. Brouwer MC, Meijers JC, Baas F, van der Ende A, Pfister HW, Giese A, et al. Plasminogen activator inhibitor-1 influences cerebrovascular complications and death in pneumococcal meningitis. Acta Neuropathol. (2014) 127:553–64. doi: 10.1007/s00401-013-1216-4
63. Jern C, Ladenvall P, Wall U, Jern S. Gene polymorphism of t-PA is associated with forearm vascular release rate of t-PA. Arterioscler Thromb Vasc Biol. (1999) 19:454–9. doi: 10.1161/01.atv.19.2.454
64. MacIvor DM, Shapiro SD, Pham CT, Belaaouaj A, Abraham SN, Ley TJ. Normal neutrophil function in cathepsin g-deficient mice. Blood. (1999) 94:4282–93. doi: 10.1182/blood.V94.12.4282
65. Burdette BE, Esparza AN, Zhu H, Wang S. Gasdermin d in pyroptosis. Acta Pharm Sin B. (2021) 11:2768–82. doi: 10.1016/j.apsb.2021.02.006
66. Danelishvili L, Everman JL, McNamara MJ, Bermudez LE. Inhibition of the plasma-Membrane-Associated serine protease cathepsin g by mycobacterium tuberculosis Rv3364c suppresses caspase-1 and pyroptosis in macrophages. Front Microbiol. (2012) 2:281. doi: 10.3389/fmicb.2011.00281
67. Tay A, Squire JA, Goldberg H, Skorecki K. Assignment of the human prostaglandin-endoperoxide synthase 2 (PTGS2) gene to 1q25 by fluorescence in situ hybridization. Genomics. (1994) 23:718–9. doi: 10.1006/geno.1994.1569
68. Warner TD, Mitchell JA. Cyclooxygenases: new forms, new inhibitors, and lessons from the clinic. FASEB J. (2004) 18:790–804. doi: 10.1096/fj.03-0645rev
69. Andersen V, Nimmo E, Krarup HB, Drummond H, Christensen J, Ho GT, et al. Cyclooxygenase-2 (COX-2) polymorphisms and risk of inflammatory bowel disease in a scottish and danish case-control study. Inflammation Bowel Dis. (2011) 17:937–46. doi: 10.1002/ibd.21440
70. Schaefer AS, Richter GM, Nothnagel M, Laine ML, Noack B, Glas J, et al. COX-2 is associated with periodontitis in europeans. J Dent Res. (2010) 89:384–8. doi: 10.1177/0022034509359575
71. Shi J, Misso NL, Kedda MA, Horn J, Welch MD, Duffy DL, et al. Cyclooxygenase-2 gene polymorphisms in an australian population: association of the -1195G > a promoter polymorphism with mild asthma. Clin Exp Allergy. (2008) 38:913–20. doi: 10.1111/j.1365-2222.2008.02986.x
72. Johansen LK, Iburg TM, Nielsen OL, Leifsson PS, Dahl-Petersen K, Koch J, et al. Local osteogenic expression of cyclooxygenase-2 and systemic response in porcine models of osteomyelitis. Prostaglandins Other Lipid Mediat. (2012) 97:103–8. doi: 10.1016/j.prostaglandins.2012.01.002
73. Basudhar D, Bharadwaj G, Somasundaram V, Cheng RYS, Ridnour LA, Fujita M, et al. Understanding the tumour micro-environment communication network from an NOS2/COX2 perspective. Br J Pharmacol. (2019) 176:155–76. doi: 10.1111/bph.14488
74. Pascual M, López-Nevot MA, Cáliz R, Koeleman BP, Balsa A, Pascual-Salcedo D, et al. Genetic determinants of rheumatoid arthritis: the inducible nitric oxide synthase (NOS2) gene promoter polymorphism. Genes Immun. (2002) 3:299–301. doi: 10.1038/sj.gene.6363856
75. Hirai K, Shirai T, Suzuki M, Shimomura T, Itoh K. Association between (CCTTT)n repeat polymorphism in NOS2 promoter and asthma exacerbations. J Allergy Clin Immunol. (2018) 142:663–5.e3. doi: 10.1016/j.jaci.2018.02.023
76. Sardoiwala MN, Mohanbhai SJ, Kushwaha AC, Dev A, Biswal L, Sharma SS, et al. Melatonin mediated inhibition of EZH2-NOS2 crosstalk attenuates inflammatory bowel disease in preclinical in vitro and in vivo models. Life Sci. (2022) 302:120655. doi: 10.1016/j.lfs.2022.120655
77. Senhaji N, Nadifi S, Zaid Y, Serrano A, Rodriguez DAL, Serbati N, et al. Polymorphisms in oxidative pathway related genes and susceptibility to inflammatory bowel disease. World J Gastroenterol. (2017) 23:8300– 7. doi: 10.3748/wjg.v23.i47.8300
78. Assunção JH, Godoy-Santos AL, Dos Santos MCLG, Malavolta EA, Gracitelli MEC, Ferreira Neto AA. Matrix metalloproteases 1 and 3 promoter gene polymorphism is associated with rotator cuff tear. Clin Orthop Relat Res. (2017) 475:1904–10. doi: 10.1007/s11999-017-5271-3
79. Pardo A, Selman M. MMP-1: the elder of the family. Int J Biochem Cell Biol. (2005) 37:283–8. doi: 10.1016/j.biocel.2004.06.017
80. Cao Z, Li C, Xiang J. Effect of matrix metalloproteinase-1 promoter genotype on interleukin-1beta-induced matrix metalloproteinase-1 production in human periodontal ligament cells. J Periodontal Res. (2010) 45:109–15. doi: 10.1111/j.1600-0765.2009.01208.x
81. Droin NM, Green DR. Role of bcl-2 family members in immunity and disease. Biochim Biophys Acta. (2004) 1644:179–88. doi: 10.1016/j.bbamcr.2003.10.011
82. Liu JJ, Huang RW, Lin DJ, Peng J, Wu XY, Lin Q, et al. Expression of survivin and bax/bcl-2 in peroxisome proliferator activated receptor-gamma ligands induces apoptosis on human myeloid leukemia cells in vitro. Ann Oncol. (2005) 16:455–9. doi: 10.1093/annonc/mdi077
83. Huang Y, Xu W, Zhou R. NLRP3 inflammasome activation and cell death. Cell Mol Immunol. (2021) 18:2114–27. doi: 10.1038/s41423-021-00740-6
84. Lin Y, Luo T, Weng A, Huang X, Yao Y, Fu Z, et al. Gallic acid alleviates gouty arthritis by inhibiting NLRP3 inflammasome activation and pyroptosis through enhancing Nrf2 signaling. Front Immunol. (2020) 11:580593. doi: 10.3389/fimmu.2020.580593
85. Wang Z, Zhang S, Xiao Y, Zhang W, Wu S, Qin T, et al. NLRP3 inflammasome and inflammatory diseases. Oxid Med Cell Longev. (2020) 2020:4063562. doi: 10.1155/2020/4063562
86. Zhang QB, Qing YF, He YL, Xie WG, Zhou JG. Association of NLRP3 polymorphisms with susceptibility to primary gouty arthritis in a chinese han population. Clin Rheumatol. (2018) 37:235–44. doi: 10.1007/s10067-017-3900-6
87. von Herrmann KM, Salas LA, Martinez EM, Young AL, Howard JM, Feldman MS, et al. NLRP3 expression in mesencephalic neurons and characterization of a rare NLRP3 polymorphism associated with decreased risk of parkinson's disease. NPJ Parkinsons Dis. (2018) 4:24. doi: 10.1038/s41531-018-0061-5
88. Xu H, Zhang A, Fang C, Zhu Q, Wang W, Liu Y, et al. SLC11A1 as a stratification indicator for immunotherapy or chemotherapy in patients with glioma. Front Immunol. (2022) 13:980378. doi: 10.3389/fimmu.2022.980378
89. Kavian Z, Sargazi S, Majidpour M, Sarhadi M, Saravani R, Shahraki M, et al. Association of SLC11A1 polymorphisms with anthropometric and biochemical parameters describing type 2 diabetes mellitus. Sci Rep. (2023) 13:6195. doi: 10.1038/s41598-023-33239-3
90. Stewart LC, Day AS, Pearson J, Barclay ML, Gearry RB, Roberts RL, et al. SLC11A1 polymorphisms in inflammatory bowel disease and mycobacterium avium subspecies paratuberculosis status. World J Gastroenterol. (2010) 16:5727–31. doi: 10.3748/wjg.v16.i45.5727
91. Sophie M, Hameed A, Muneer A, Samdani AJ, Saleem S, Azhar A. SLC11A1 polymorphisms and host susceptibility to cutaneous leishmaniasis in pakistan. Parasit Vectors. (2017) 10:12. doi: 10.1186/s13071-016-1934-2
92. Kim EJ, Kim KM, Park SH, Kim JS, Lee WK, Cha SI, et al. SLC11A1 polymorphisms are associated with the risk of chronic obstructive pulmonary disease in a korean population. Biochem Genet. (2008) 46:506–19. doi: 10.1007/s10528-008-9166-6
93. López-Anglada E, Collazos J, Montes AH, Pérez-Is L, Pérez-Hevia I, Jiménez-Tostado S, et al. IL-1 β gene (+3954 C/T, exon 5, rs1143634) and NOS2 (exon 22) polymorphisms associate with early aseptic loosening of arthroplasties. Sci Rep. (2022) 12:18382. doi: 10.1038/s41598-022-22693-0
94. Levescot A, Chang MH, Schnell J, Nelson-Maney N, Yan J, Martínez-Bonet M, et al. IL-1β-driven osteoclastogenic tregs accelerate bone erosion in arthritis. J Clin Invest. (2021) 131:e141008. doi: 10.1172/JCI141008
95. Yokota K, Sato K, Miyazaki T, Aizaki Y, Tanaka S, Sekikawa M, et al. Characterization and function of tumor necrosis factor and interleukin-6-Induced osteoclasts in rheumatoid arthritis. Arthritis Rheumatol. (2021) 73:1145–54. doi: 10.1002/art.41666
96. Chiba J, Rubash HE, Kim KJ, Iwaki Y. The characterization of cytokines in the interface tissue obtained from failed cementless total hip arthroplasty with and without femoral osteolysis. Clin Orthop Relat Res. (1994) 1994:304–12.
97. Hirota K, Martin B, Veldhoen M. Development, regulation and functional capacities of Th17 cells. Semin Immunopathol. (2010) 32:3–16. doi: 10.1007/s00281-009-0187-y
98. Stahelova A, Mrazek F, Smizansky M, Petrek M, Gallo J. Variation in the IL1B, TNF and IL6 genes and individual susceptibility to prosthetic joint infection. BMC Immunol. (2012) 13:25. doi: 10.1186/1471-2172-13-25
99. Malik MH, Jury F, Bayat A, Ollier WE, Kay PR. Genetic susceptibility to total hip arthroplasty failure: a preliminary study on the influence of matrix metalloproteinase 1, interleukin 6 polymorphisms and vitamin d receptor. Ann Rheum Dis. (2007) 66:1116–20. doi: 10.1136/ard.2006.062018
100. Erdemli B, Özbek EA, Başarir K, Karahan ZC, Öcal D, Biriken D. Proinflammatory biomarkers' level and functional genetic polymorphisms in periprosthetic joint infection. Acta Orthop Traumatol Turc. (2018) 52:143–7. doi: 10.1016/j.aott.2017.11.002
101. Navratilova Z, Gallo J, Mrazek F, Petrek M. Genetic variation in key molecules of the th-17 immune response is not associated with risk for prosthetic joint infection in a czech population. BioMed Pap Med Fac Univ Palacky Olomouc Czech Repub. (2012) 156:248–52. doi: 10.5507/bp.2012.023
102. Malik MH, Bayat A, Jury F, Kay PR, Ollier WE. Genetic susceptibility to total hip arthroplasty failure–positive association with mannose-binding lectin. J Arthroplasty. (2007) 22:265–70. doi: 10.1016/j.arth.2006.02.163
103. Siassi M, Hohenberger W, Riese J. Mannan-binding lectin (MBL) serum levels and post-operative infections. Biochem Soc Trans. (2003) 31:774–5. doi: 10.1042/bst0310774
104. Lipscombe RJ, Sumiya M, Hill AV, Lau YL, Levinsky RJ, Summerfield JA, et al. High frequencies in african and non-african populations of independent mutations in the mannose binding protein gene. Hum Mol Genet. (1992) 1:709–15. doi: 10.1093/hmg/1.9.709
105. Garred P, Madsen HO, Halberg P, Petersen J, Kronborg G, Svejgaard A, et al. Mannose-binding lectin polymorphisms and susceptibility to infection in systemic lupus erythematosus. Arthritis Rheumatol. (1999) 42:2145–52. doi: 10.1002/1529-0131(199910)42:10<2145::AID-ANR15>3.0.CO;2-#
106. Jacobsen S, Madsen HO, Klarlund M, Jensen T, Skjødt H, Jensen KE, et al. The influence of mannose binding lectin polymorphisms on disease outcome in early polyarthritis. TIRA Group J Rheumatol. (2001) 28:935–42.
107. Mrazek F, Gallo J, Stahelova A, Petrek M. Coding variants of TLR2 and TLR4 genes do not substantially contribute to prosthetic joint infection. Inflammation Res. (2013) 62:483–7. doi: 10.1007/s00011-013-0601-8
108. El-Helou O, Berbari EF, Brown RA, Gralewski JH, Osmon DR, Razonable RR. Functional assessment of toll-like receptor 2 and its relevance in patients with staphylococcus aureus infection of joint prosthesis. Hum Immunol. (2011) 72:47–53. doi: 10.1016/j.humimm.2010.10.001
109. Haynes DR, Crotti TN, Potter AE, Loric M, Atkins GJ, Howie DW, et al. The osteoclastogenic molecules RANKL and RANK are associated with periprosthetic osteolysis. J Bone Joint Surg Br. (2001) 83:902–11. doi: 10.1302/0301-620x.83b6.10905
110. Alehashemi S, Dasari S, Metpally A, Uss K, Castelo-Soccio LA, Heller T, et al. Anakinra-associated systemic amyloidosis. Arthritis Rheumatol. (2024) 76:100–6. doi: 10.1002/art.42664
111. Strle K, Shin JJ, Glickstein LJ, Steere AC. Association of a toll-like receptor 1 polymorphism with heightened Th1 inflammatory responses and antibiotic-refractory lyme arthritis. Arthritis Rheumatol. (2012) 64:1497–507. doi: 10.1002/art.34383
112. Azim K, McManus R, Brophy K, Ryan A, Kelleher D, Reynolds JV. Genetic polymorphisms and the risk of infection following esophagectomy. positive association with TNF-alpha gene -308 genotype. Ann Surg. (2007) 246:122–8. doi: 10.1097/01.sla.0000259389.09161.49
113. de Rooij BJ, van Hoek B, ten Hove WR, Roos A, Bouwman LH, Schaapherder AF, et al. Lectin complement pathway gene profile of donor and recipient determine the risk of bacterial infections after orthotopic liver transplantation. Hepatology. (2010) 52:1100–10. doi: 10.1002/hep.23782
114. Gordon M, Sinopoulou V, Akobeng AK, Sarian A, Moran GW. Infliximab for maintenance of medically-induced remission in crohn's disease. Cochrane Database Syst Rev. (2024) 2:CD012609. doi: 10.1002/14651858.CD012609.pub2
115. Lu X, Hu R, Peng L, Liu M, Sun Z. Efficacy and safety of adalimumab biosimilars: Current critical clinical data in rheumatoid arthritis. Front Immunol. (2021) 12:638444. doi: 10.3389/fimmu.2021.638444
116. Matusiak AE, Stępniak J, Lewiński A, Karbownik-Lewińska M. Decreased mannan-binding lectin level in adults with hypopituitarism; dependence on appropriate hormone replacement therapies. Front Immunol. (2023) 14:1107334. doi: 10.3389/fimmu.2023.1107334
117. Keizer MP, Wouters D, Schlapbach LJ, Kuijpers TW. Restoration of MBL-deficiency: redefining the safety, efficacy and viability of MBL-substitution therapy. Mol Immunol. (2014) 61:174–84. doi: 10.1016/j.molimm.2014.06.005
118. Scott LJ. Tocilizumab: A review in rheumatoid arthritis. Drugs. (2017) 77:1865–79. doi: 10.1007/s40265-017-0829-7
119. Raimondo MG, Biggioggero M, Crotti C, Becciolini A, Favalli EG. Profile of sarilumab and its potential in the treatment of rheumatoid arthritis. Drug Des Devel Ther. (2017) 11:1593–603. doi: 10.2147/DDDT.S100302
120. Zhao W, Meng H. Effects of genetic polymorphism of drug-metabolizing enzymes on the plasma concentrations of antiepileptic drugs in chinese population. Bioengineered. (2022) 13:7709–45. doi: 10.1080/21655979.2022.2036916
121. Roots I, Gerloff T, Meisel C, Kirchheiner J, Goldammer M, Kaiser R, et al. Pharmacogenetics-based new therapeutic concepts. Drug Metab Rev. (2004) 36:617–38. doi: 10.1081/dmr-200033458
122. Liu J, Dong Z, Zhu Q, He D, Ma Y, Du A, et al. TNF-α promoter polymorphisms predict the response to etanercept more powerfully than that to Infliximab/Adalimumab in spondyloarthritis. Sci Rep. (2016) 6:32202. doi: 10.1038/srep32202
123. Sawczenko A, Azooz O, Paraszczuk J, Idestrom M, Croft NM, Savage MO, et al. Intestinal inflammation-induced growth retardation acts through IL-6 in rats and depends on the -174 IL-6 G/C polymorphism in children. Proc Natl Acad Sci USA. (2005) 102:13260–5. doi: 10.1073/pnas.0503589102
124. Grotenfelt NE, Wasenius NS, Rönö K, Laivuori H, Stach-Lempinen B, Orho-Melander M, et al. Interaction between rs10830963 polymorphism in MTNR1B and lifestyle intervention on occurrence of gestational diabetes. Diabetologia. (2016) 59:1655–8. doi: 10.1007/s00125-016-3989-1
125. Yanes T, McInerney-Leo AM, Law MH, Cummings S. The emerging field of polygenic risk scores and perspective for use in clinical care. Hum Mol Genet. (2020) 29:R165–76. doi: 10.1093/hmg/ddaa136
126. Lu C, Xu CF, Wan XY, Zhu HT, Yu CH, Li YM. Screening for pancreatic cancer in familial high-risk individuals: A systematic review. World J Gastroenterol. (2015) 21:8678–86. doi: 10.3748/wjg.v21.i28.8678
127. Dinh T, Snyder G, Veves A. Current techniques to detect foot infection in the diabetic patient. Int J Low Extrem Wounds. (2010) 9:24–30. doi: 10.1177/1534734610363004
128. Stephan AM, Faino A, Caglar D, Klein EJ. Clinical presentation of acute osteomyelitis in the pediatric emergency department. Pediatr Emerg Care. (2022) 38:e209–13. doi: 10.1097/PEC.0000000000002217
129. Bufi R, Korstanje R. The impact of genetic background on mouse models of kidney disease. Kidney Int. (2022) 102:38–44. doi: 10.1016/j.kint.2022.03.020
130. Uniken Venema WT, Voskuil MD, Dijkstra G, Weersma RK, Festen EA. The genetic background of inflammatory bowel disease: from correlation to causality. J Pathol. (2017) 241:146–58. doi: 10.1002/path.4817
131. Gadelha MR, Kasuki L, Korbonits M. The genetic background of acromegaly. Pituitary. (2017) 20:10–21. doi: 10.1007/s11102-017-0789-7
132. Kasher M, Williams FMK, Freidin MB, Malkin I, Cherny SS, CHARGE Inflammation Working Group, et al. Understanding the complex genetic architecture connecting rheumatoid arthritis, osteoporosis and inflammation: discovering causal pathways. Hum Mol Genet. (2022) 31:2810–9. doi: 10.1093/hmg/ddac061
133. Zhou Z, Chen Y, Min HS, Wan Y, Shan H, Lin Y, et al. Merlin functions as a critical regulator in staphylococcus aureus-induced osteomyelitis. J Cell Physiol. (2022) 237:815–23. doi: 10.1002/jcp.30550
134. Butrico CE, Cassat JE. Quorum sensing and toxin production in staphylococcus aureus osteomyelitis: Pathogenesis and paradox. Toxins (Basel). (2020) 12:516. doi: 10.3390/toxins12080516
135. Srinivasan S, Clements JA, Batra J. Single nucleotide polymorphisms in clinics: Fantasy or reality for cancer? Crit Rev Clin Lab Sci. (2016) 53:29–39. doi: 10.3109/10408363.2015.1075469
136. Wu K, Kong F, Zhang J, Tang Y, Chen Y, Chao L, et al. Recent progress in single-nucleotide polymorphism biosensors. Biosensors (Basel). (2023) 13:864. doi: 10.3390/bios13090864
Keywords: osteomyelitis, prosthetic joint infection, single nucleotide polymorphisms, single nucle otide variations, narrative review
Citation: Zhou J-Q, Liu Z-X, Zhong H-F, Liu G-Q, Ding M-C, Zhang Y, Yu B and Jiang N (2024) Single nucleotide polymorphisms in the development of osteomyelitis and prosthetic joint infection: a narrative review. Front. Immunol. 15:1444469. doi: 10.3389/fimmu.2024.1444469
Received: 05 June 2024; Accepted: 08 August 2024;
Published: 05 September 2024.
Edited by:
Michela Rossi, Bambino Gesù Children’s Hospital (IRCCS), ItalyReviewed by:
Madhavi Annamanedi, West Virginia University, United StatesCopyright © 2024 Zhou, Liu, Zhong, Liu, Ding, Zhang, Yu and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nan Jiang, aG54eWpuQHNtdS5lZHUuY24=; Bin Yu, eXViaW5Ac211LmVkdS5jbg==; Yu Zhang, bHVja18yMDAxQDEyNi5jb20=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.