 Per Wågström1,2*‡
Per Wågström1,2*‡ Maria Hjorth2†
Maria Hjorth2† Daniel Appelgren3Janne Björkander4,5
Daniel Appelgren3Janne Björkander4,5 Charlotte Dahle2
Charlotte Dahle2 Mats Nilsson5,6Åsa Nilsdotter-Augustinsson2
Mats Nilsson5,6Åsa Nilsdotter-Augustinsson2 Jan Ernerudh2†
Jan Ernerudh2† Sofia Nyström2
Sofia Nyström2- 1Department of Infectious Diseases, Ryhov County Hospital, Jönköping, Sweden
- 2Division of Inflammation and Infection, Department of Biomedical and Clinical Sciences, Linköping University, Linköping, Sweden
- 3Division of Diagnostics and Specialist Medicine, Department of Health, Medicine and Caring Sciences, Linköping University, Linköping, Sweden
- 4Wetterhälsan Primary Health Care Centre, Region Jönköping County Jönköping, Sweden
- 5Futurum – the Academy for Health and Care, Region Jönköping County, Sweden
- 6Department of Health, Medicine and Caring Sciences, Linköping University, Linköping, Sweden
Background: Immunoglobulin G subclass deficiencies (IgGsd) comprise a wide clinical spectrum from no symptoms to repeated respiratory infections and risk for the development of lung damage. Our aims were to investigate whether the immunological phenotype of IgGsd patients on and off immunoglobulin replacement therapy (IgRT) was reflected in the clinical features of IgGsd.
Method: Thirty patients with IgGsd were included in this prospective study of 18 months of IgRT, followed by 7-18 months of IgRT discontinuation. Blood samples were collected when patients were on and off IgRT and compared with samples from 34 cross-sectional healthy controls. An in-depth lymphocyte phenotyping was performed by flow cytometry and plasma levels of immune checkpoints were assessed.
Results: IgG3 subclass deficiency was most common. Patients with IgGsd had decreased levels of activated T cells and B cells and plasma levels of negative immune checkpoint molecules correlated negatively with T cell and B cell activation. The decreased T cell activation level was unaffected by IgRT, while the B cell activation was partly restored. Of note, decreased levels of activated regulatory T cells (Tregs) were found in IgGsd patients and was partly restored during IgRT. The profile of comorbidities did not associate with Treg levels.
Discussion: IgGsd is associated with decreased B cell and T cell activation including Tregs, and increased plasma levels of negative immune checkpoint molecules. The consequence of reduced activated Tregs in IgGsd remains unclear. Decreased immune cell activation was partly restored during IgRT, demonstrating that IgRT may contribute to improved immune function in patients with IgGsd.
Introduction
Predominantly antibody deficiencies (PAD) comprise a group of inborn errors of immunity with poor antibody responses (1). PAD is associated with increased susceptibility to infections and with chronic inflammatory disorders (2). Immunoglobulin G subclass deficiency (IgGsd) is a mild form of PAD characterized by increased frequencies of infections, and reduced levels of at least one IgG subclass. According to the international classification, which aligns with Swedish classification, IgGsd characterized by reduced levels of least one of IgG subclass and is mostly an asymptomatic PAD, but IgGsd can also be associated with increased frequencies of infections. IgGsd can be associated with subnormal levels of IgA or moderately reduced IgG (1). According to the European Society of immunodeficiency, IgGsd should be accompanied by normal levels of IgG, IgM and IgA otherwise it is considered an unclassified antibody deficiency (3). The clinical presentation of IgGsd varies from asymptomatic to recurrent infections and symptomatic IgGsd is often associated with atopic disease and chronic lung disease (4–6). Bronchiectasis, affect 38%-48% of patients with IgGsd. Recurrent airway infections may contribute to airway remodeling and deteriorated lung function in patients with IgGsd (7–9). Autoimmune conditions are also frequent among patients with IgGsd. The prevalence of autoimmune conditions, such as rheumatoid arthritis, thyroiditis and Sjögren syndrome, was over 40% in a large cohort of patients with IgGsd (10). Furthermore, in a study on inborn errors of immunity, rheumatologic complications were most common in patients with IgGsd (11%) when compared to other groups of patients with predominantly antibody deficiencies (11). Hence, low IgG subclass levels may reflect an underlying immune dysregulation, but the understanding of factors contributing to autoimmune complications in IgGsd is scarce.
To date, there is no pathogenic gene variant associated with IgGsd, or with any other PAD characterized by reduced IgG levels not fulfilling the criteria of common variable immunodeficiency (CVID). Common variable immunodeficiency (CVID) refers to a group of heterogenic severe PAD characterized by significant hypogammaglobulinemia, in combination with perturbations of circulating B cell and T cell subsets (12). In CVID an increasing number of pathogenic gene variants have been reported (1). As in unspecified PAD and IgGsd, autoimmunity and lung disease affect many patients with CVID (13). In contrast to IgGsd, lymphoproliferation (i. e polyclonal lymphocytic infiltration of non-lymphoid tissues) may complicate CVID, and CVID is associated with markers of more severe immune defects than IgGsd (14).
Patients with CVID are characterized by decreased class-switched memory B cells (15) and low regulatory T cells (Tregs) associates with autoimmune manifestations in CVID (16). There is, to our knowledge, there is only one previous report on circulating lymphocyte subsets, e.g., B cells, CD4+ helper (h) T cells, CD8+ cytotoxic T cells and natural killer (NK) cells, in patients with IgGsd. In a study of 16 patients with IgG2 deficiency, the only anomaly found among lymphocytes, was subnormal levels of Th cells in one patient (10). In the vast majority of the patients in a cohort of 17 patients with IgG3 deficiency, there were normal numbers of, or close to normal numbers, of B cells, Th cells, cytotoxic T cells and NK cells (17). Notably, altered functional T cell responses were reported in occasional patients in this restricted cohort, which may reflect T cell subpopulation perturbations. Decreased switched memory B cells have been reported in unspecified IgG deficiency (14) and it can be hypothesized that the numbers of class-switched memory cells are informative about the underlying immune response also in IgGsd. Impaired infection control may also result in perturbation of the T cell compartment. Accumulating data support a role for Tregs in various autoimmune diseases (18). Given the high prevalence of autoimmunity in IgGsd, data on the frequencies of Tregs could increase the understanding of factors contributing to autoimmunity in patients with IgGsd.
Immunoglobulin replacement therapy (IgRT) consists of monomeric IgG pooled from healthy donors which is administrated subcutaneously or intravenously and aims to improve the immune response to pathogens in patients with PAD. High dose intravenous immunoglobulin therapy, on the other hand, is used to alleviate inflammatory conditions by immune modulatory mechanisms. In patients with IgGsd, IgRT reduces bacterial infections and improve the quality of life (6, 17, 19) and IgRT should be considered in patients with clinically significant infections (20). Consensus on the use of IgRT in IgGsd and other minor PAD is lacking, and it is essential to define reliable predictors identifying the subgroup of patients with IgGsd that benefit from IgRT. In Sweden, the decision to introduce IgRT is based on the frequency and severity of bacterial infections and the presence of any lung damage. According to present Swedish national guidelines (21), the effect of IgRT should be evaluated after 12-18 months and followed by a discontinuation trial, during which IgRT is reintroduced if clinically significant infections reoccur or if there are signs of lung function deterioration when IgRT is discontinued. It has been suggested that immunoglobulins used in replacement therapies may modulate the immune response (22, 23), however to our knowledge there is only one in vivo study on the long term immune-modulatory effects of IgRT in CVID (24). Longitudinal prospective studies of the immune modulatory effects of IgRT in hypogammaglobulinemia are important for the understanding of the pathological processes.
In this study, we performed in-depth characterization of the B cell and T cell compartments of a clinically well-characterized cohort of patients with IgGsd. Plasma levels of costimulatory and regulatory immune checkpoint molecules were also evaluated. In addition, any changes in subsets of B cells, T cells and the levels of costimulatory and regulatory immune checkpoints in patients with IgGsd during IgRT were investigated. We found that IgGsd was associated with decreased levels of activated T cells and B cells and that plasma levels of negative immune checkpoint molecules correlated negatively with T cell and B cell activation. IgGsd was also characterized by decreased levels of activated regulatory T cells (Tregs), which were partly restored during IgRT.
Material and methods
Ethics
The study protocol was approved by Regional Ethical Review Board in Linköping, Sweden (Dnr 2011/506-31). Written informed consent was obtained from all study participants.
Study population and design of the study
This prospective study was conducted at two Swedish regional centers of infectious disease during 2012-2015. Of 85 adult patients diagnosed with IgGsd, 35 (22 women and 13 men) fulfilled the inclusion criteria (no severe lung disease and no previous IgRT discontinuation trial). The cohort has previously been described (25). Patients kept track of their infections during the study period and blood samples were collected after 18 months on IgRT, and at least seven months after discontinuation of IgRT. Samples both when on and off IgRT were available for thirty patients (19 women and 11 men). Thirty-four healthy blood donors (15 men and 19 women, median aged 54, range 28-68 years), served as cross-sectional controls. The control population was in good physical health. Three controls (all 62 years or older) reported regular use of antihypertensive drugs and another control was on amitriptyline for neuropathic pain.
Handling of blood samples
Blood samples were drawn in vacutainers, and immune phenotyping were performed on whole blood. Levels of IgG, IgA, IgM, and IgG subclasses IgG1, IgG2 and IgG3 were measured according to standard operating procedures at the Laboratories of Clinical Chemistry in the Counties of Jönköping and Östergötland. Plasma samples were stored at -80°C until plasma protein profiling.
Lymphocyte phenotyping with flow cytometric analysis
Absolute T-, B- and NK-cell numbers were determined by flow cytometry with the use of BD Multitest™ reagent (BD Biosciences; 340499 and 340500). Briefly, antibodies were added to fresh EDTA whole blood and erythrocytes were lysed with BD FACS™ lysing solution (BD Biosciences). Data were acquired with BD FACSCanto™ II (BD Biosciences) flow cytometer. Phenotyping of T- and B-cell subsets was performed in 7- or 8-color combinations. The mAbs used are listed in (Supplementary Table S1). Kaluza flow cytometry software version 1.5 (Beckman Coulter, Miami, USA) was used for data analysis. Identification of lymphocytes was performed by CD45 and side scatter (SSC) or forward scatter (FSC) and SSC. HLA-DR was used as a marker for activation of CD4+ helper and CD8+ cytotoxic T cells. Differentiation status of CD4+ T helper and CD8+ cytotoxic T cells was determined with CD45RA and CCR7. Regulatory T cells (Tregs) were defined as resting (CD45RA+CD25dim) or activated (CD45RA-CD25bright) T helper cells. Subtyping of naïve and memory B cells was determined in CD19+ cells using CD27 and IgD. Further subtyping of B cells was performed using CD21, CD24, CD25, CD38 and IgM.
Plasma protein profiling
Frozen plasma samples were sent to the Clinical Biomarkers facility, Science for Life Laboratory, Uppsala University (Uppsala, Sweden) and analyzed there according to their standard operating procedures. Proseek multiplex assay inflammation panel (95302) was used for detection of 92 different inflammatory associated proteins (Olink Proteomics, Uppsala, Sweden). Of 81 proteins detected in at least 40% of samples, 19 had a predominantly plasma membrane location and were selected for further investigation (Supplementary Table S2).
Statistical analyses
For comparisons between two groups, Mann-Whitney U-test was used, or the Wilcoxon test for paired samples, and ANOVA with Tukey post-test if parametric data. Statistical significance was defined as p<0.05. SAS/STAT® ver. 13.1 software (Copyright © 2002-2012 by SAS Institute Inc., Cary, NC, USA), Statistica Ver.13, (Copyright © 1984-2017, TIBCO Software Inc., CA, USA), IBM SPSS Statistics 28 (SPSS inc., IL, USA) and GraphPad Prism 9.3.1 (GraphPad software, CA, USA) were used for the calculations. GraphPad Prism was used for graphics. Bioinformatics & Evolutionary Genomics was used for the construction of Venn-diagrams (26).
Results
Characteristics of the study cohort
The group of IgGsd patients was heterogeneous, with diverse types of comorbidities (Table 1), and has previously been described in detail (25). The most common associated comorbidities were autoimmunity and lung disease. Seven patients were affected by both autoimmunity and lung disease. Ten patients were considered having a profession associated with an increased risk of exposure to airway pathogens, e. g preschool teacher and healthcare worker. Sixteen patients had subnormal IgG levels and seven patient subnormal IgA levels (Figures 1A, B). Subnormal IgA or IgG was any concentration below the lower reference limits referring to the 2.5 percentile of a healthy Swedish population (27). IgG3 deficiency was present in ten of the 14 patients with normal levels of total IgG and in 16 of 23 patients with normal levels of IgA. IgG1 deficiency and mixed IgGsd were most common in the group of patients with subnormal IgG levels (Figure 1A). Six patients of the seven patients with subnormal IgA had IgG2 deficiency (Figure 1B). Subnormal IgM levels were found in five patients (results not shown).
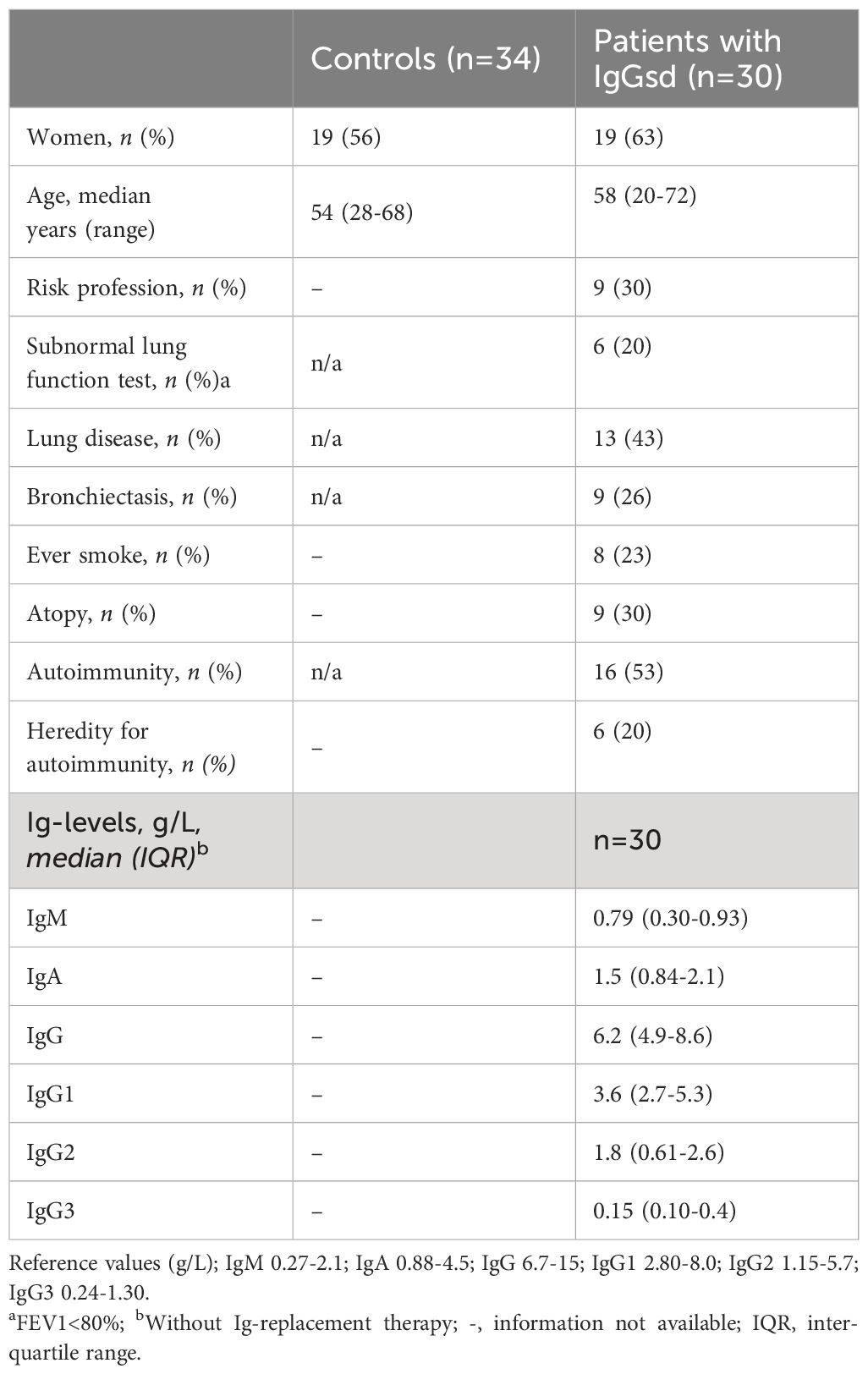
Table 1. Demographics, comorbidities, and Ig levels in patients with IgG subclass deficiency.
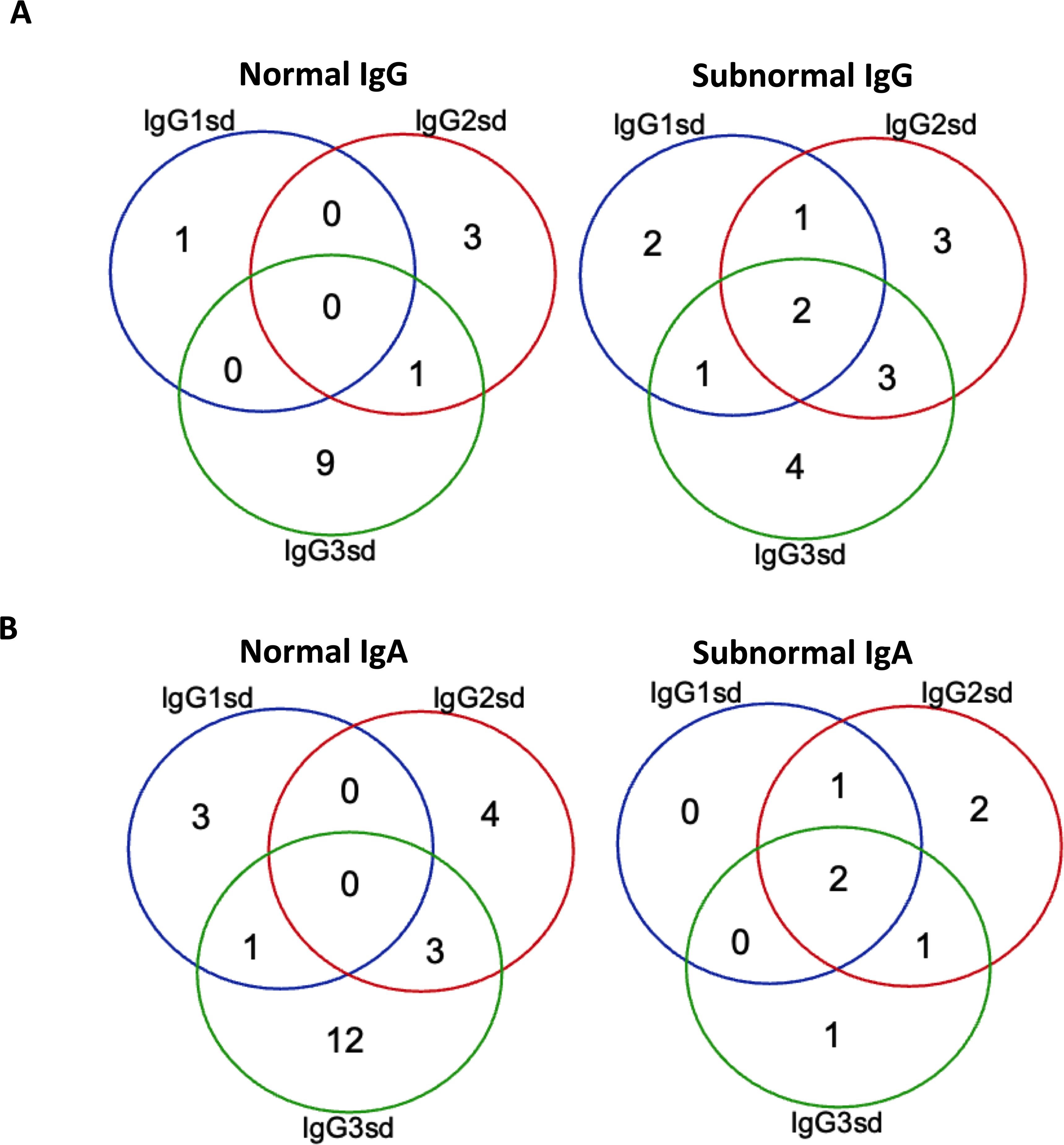
Figure 1. Isolated IgG3 subclass deficiency most common among patients with normal IgG. Venn-diagram showing the distribution of single and combined IgG-subclass deficiencies related to plasma levels of IgG (A) and IgA (B) in patients with IgGsd (n=30). Quantitative detection of serum immunoglobulins was performed by routine analyses using validated techniques.
Circulating lymphocyte populations are unaffected by IGRT in IgGsd
To evaluate if patients with IgGsd had phenotypic abnormalities in their lymphocyte populations, a broad flow cytometry assessment was performed with paired samples collected when on and when off IgRT. Samples collected from patients were compared to 34 cross-sectional healthy controls. The absolute numbers and percentages of major lymphocyte populations (CD4+ and CD8+ T cells, B cells and NK cells) were similar in patients with IgGsd and in the controls (Table 2). The major lymphocytes populations, e.g., Th cells, cytotoxic T cells, B cells and NK cells, were not affected by IgRT since levels were similar when on and off treatment.
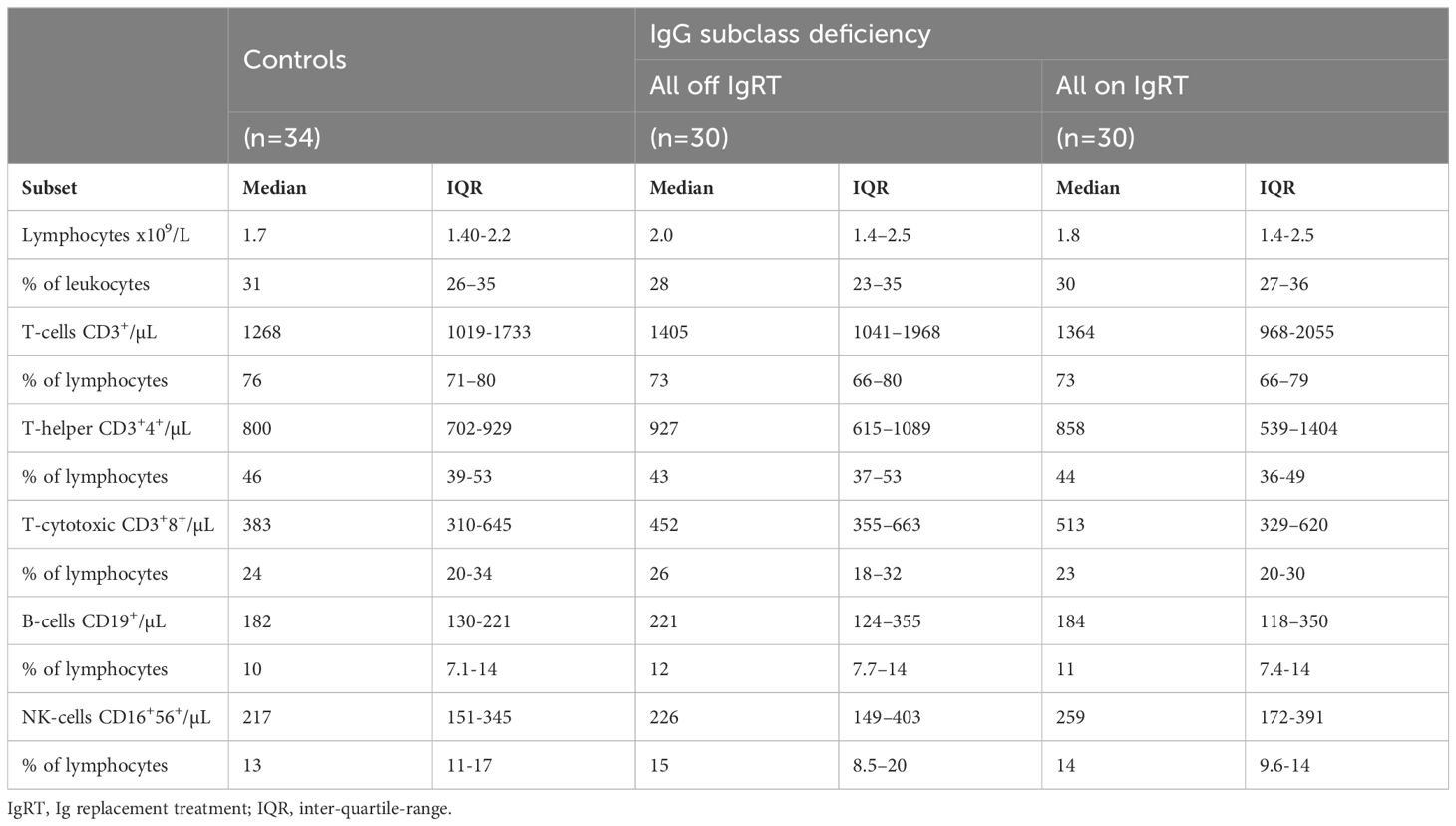
Table 2. Lymphocyte populations in patients with IgG subclass deficiency and controls.
Alterations in IgGsd patients’ B cell subsets normalized during IgRT
A deeper characterization of the B cells revealed aberrations restricted to the CD25+ B cell subsets in patients off IgRT compared to controls (Table 3). The proportions of activated CD25+ B cells and activated memory CD25+ CD27+ B cells were lower in IgGsd when off IgRT, while levels on treatment were normalized and did not differ from controls (Figure 2). Activated CD21Low and switched memory IgD- CD27+ B cells were similar across groups. In summary, patients with IgGsd had aberrant activation of B cell subsets that were normalized during IgRT.
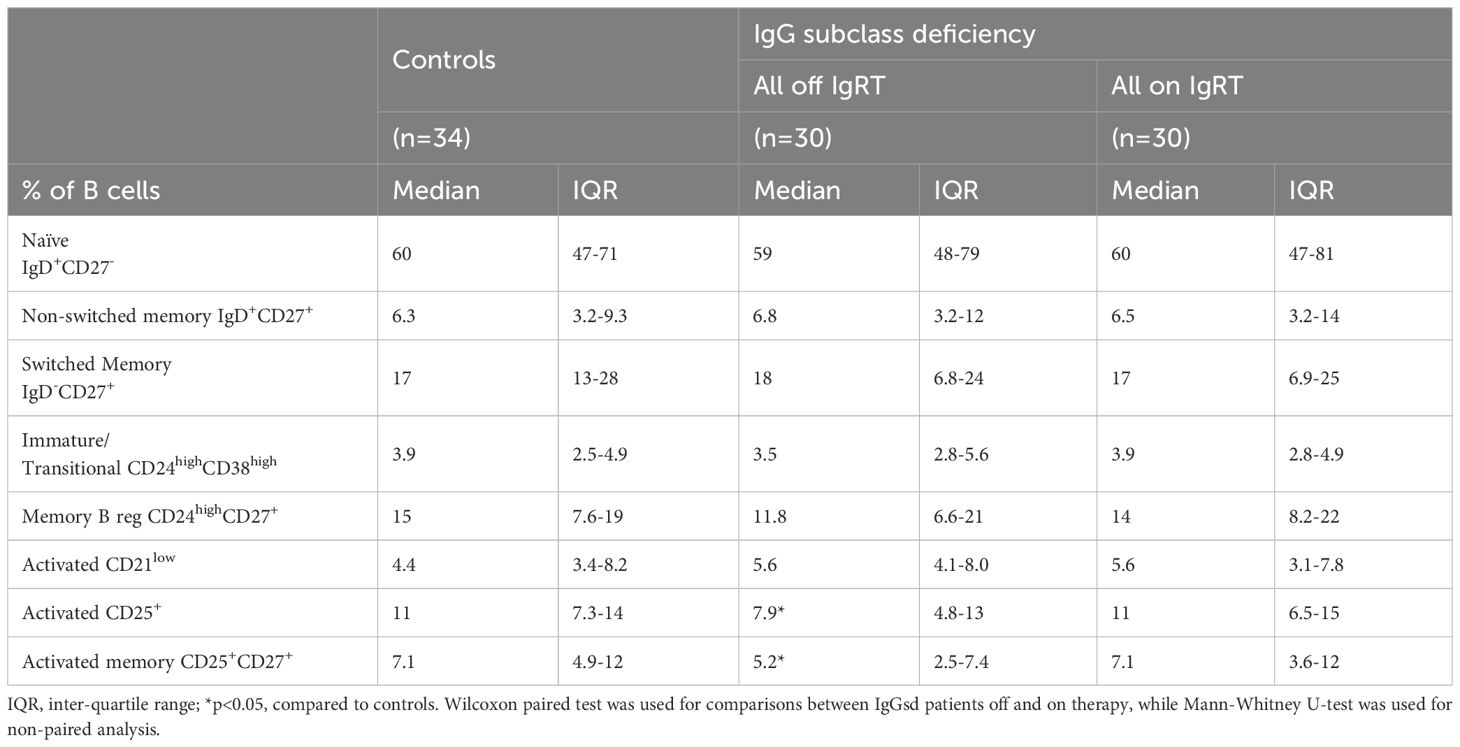
Table 3. B cell subpopulations in patients with IgG subclass deficiency.
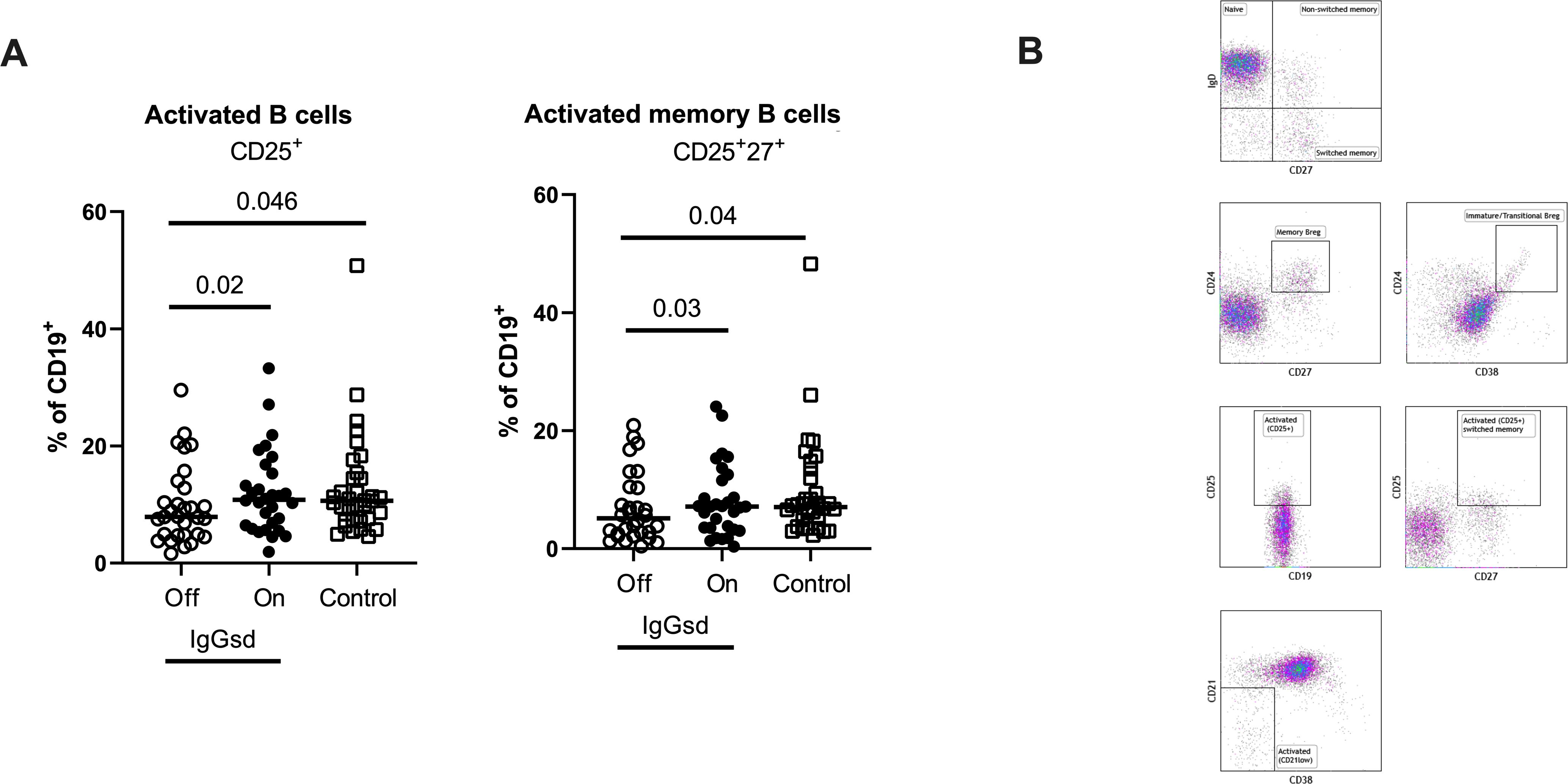
Figure 2. Ig-subclass deficiency was associated with decreased activation of B cells. Frequencies of activated and activated memory B-cells in Ig-subclass deficiency (IgGsd) patients when off (open circle) and on (filled circle) Ig-replacement therapy (IgRT) in relation to healthy controls (open squares) were determined by flow cytometry (A). Definition and gating strategies of CD19+ B cell subsets (B). Wilcoxon paired test was used for comparisons between IgGsd patients off and on IgRT, while Mann-Whitney U-test was used for non-paired analysis. Breg, regulatory B cells.
Decreased T cell activation in IgGsd and increased CD8+ T cell differentiation during IgRT
Within the T cell compartment, we found decreased proportions of activated (HLA-DR+) CD4+ and CD8+ T cells in IgGsd patients off IgRT compared to controls (Figure 3A). The differences in T cell activation remained unaffected by IgRT. The proportions of terminally differentiated memory (TEMRA) CD4+ and CD8+ T cells were similar across groups, as well as the proportions of senescent (CD28-) CD4+ and CD8+ T cells, both when on and off IgRT (Table 4). On the other hand, IgRT induced a redistribution of memory cells within the CD8+ T cell population. When on IgRT, the proportions of CD45RA-CCR7+ central memory CD8+ cells were lower, while the proportions of CD45RA- CCR7- effector memory CD8+ T cells were higher compared with off-treatment samples (Figure 3B). Gating strategies for T cell subsets are shown in Figure 3C. Taken together, IgGsd was characterized by a decrease in activated T cells which was unaffected by IgRT.
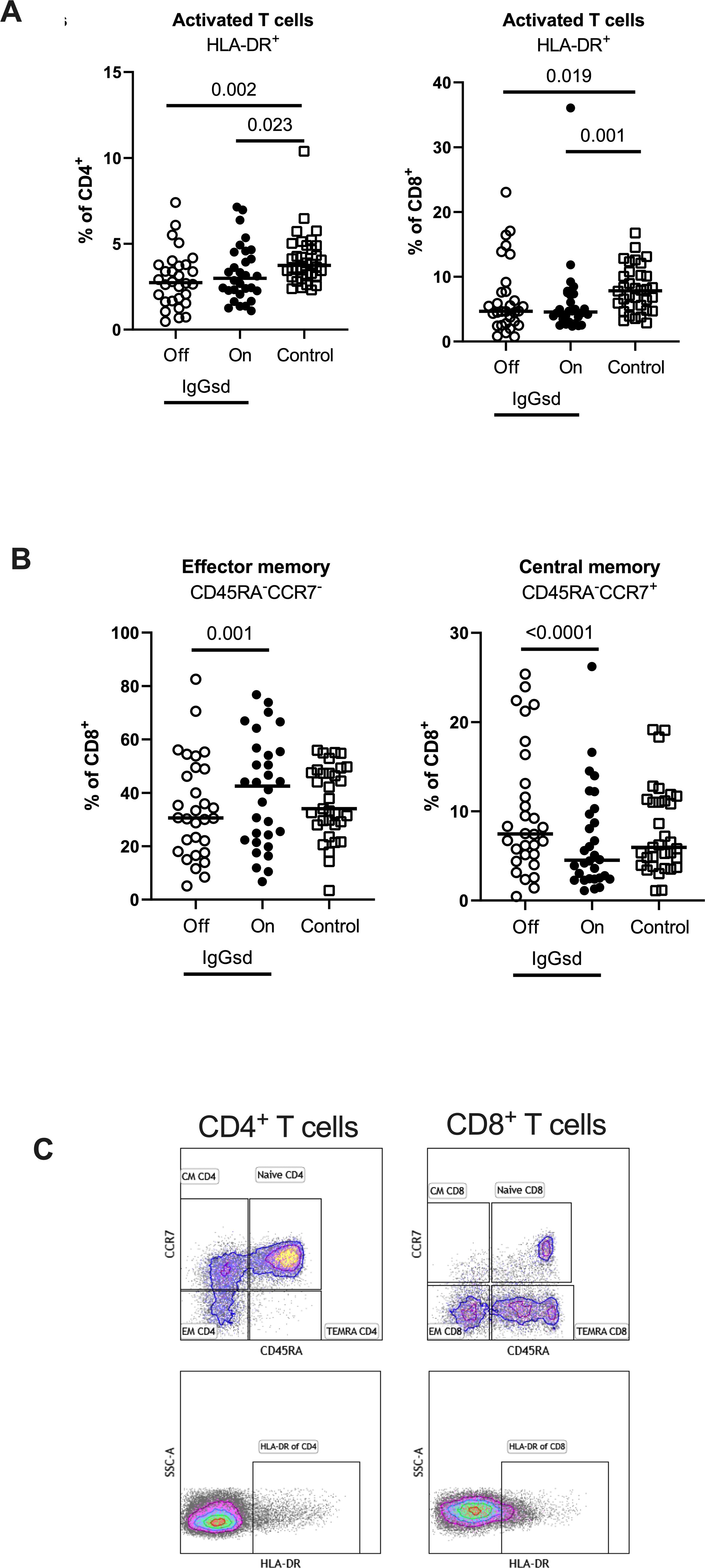
Figure 3. IgG replacement therapy is associated with increased CD8+ T cell differentiation in patients with IgG-subclass deficiency. Frequencies of activated CD4+ T-helper and CD8+ cytotoxic T cells (A), effector memory and central memory CD8+ cytotoxic T cells (B) were assessed by flow cytometry in patients with Ig-subclass deficiency (IgGsd) when off (open circle) and on (filled circle) immunoglobulin replacement therapy (IgRT) and compared to healthy controls (open squares). Definition and gating strategies of naïve- memory- and activated subsets of CD4+ and CD8+ T cells (C). Wilcoxon paired test was used for comparisons between IgGsd patients off and on immunoglobulin replacement therapy, while Mann-Whitney U-test was used for non-paired analysis. CM, central memory; EM, effector memory; TEMRA, terminally differentiated effector memory RA+.
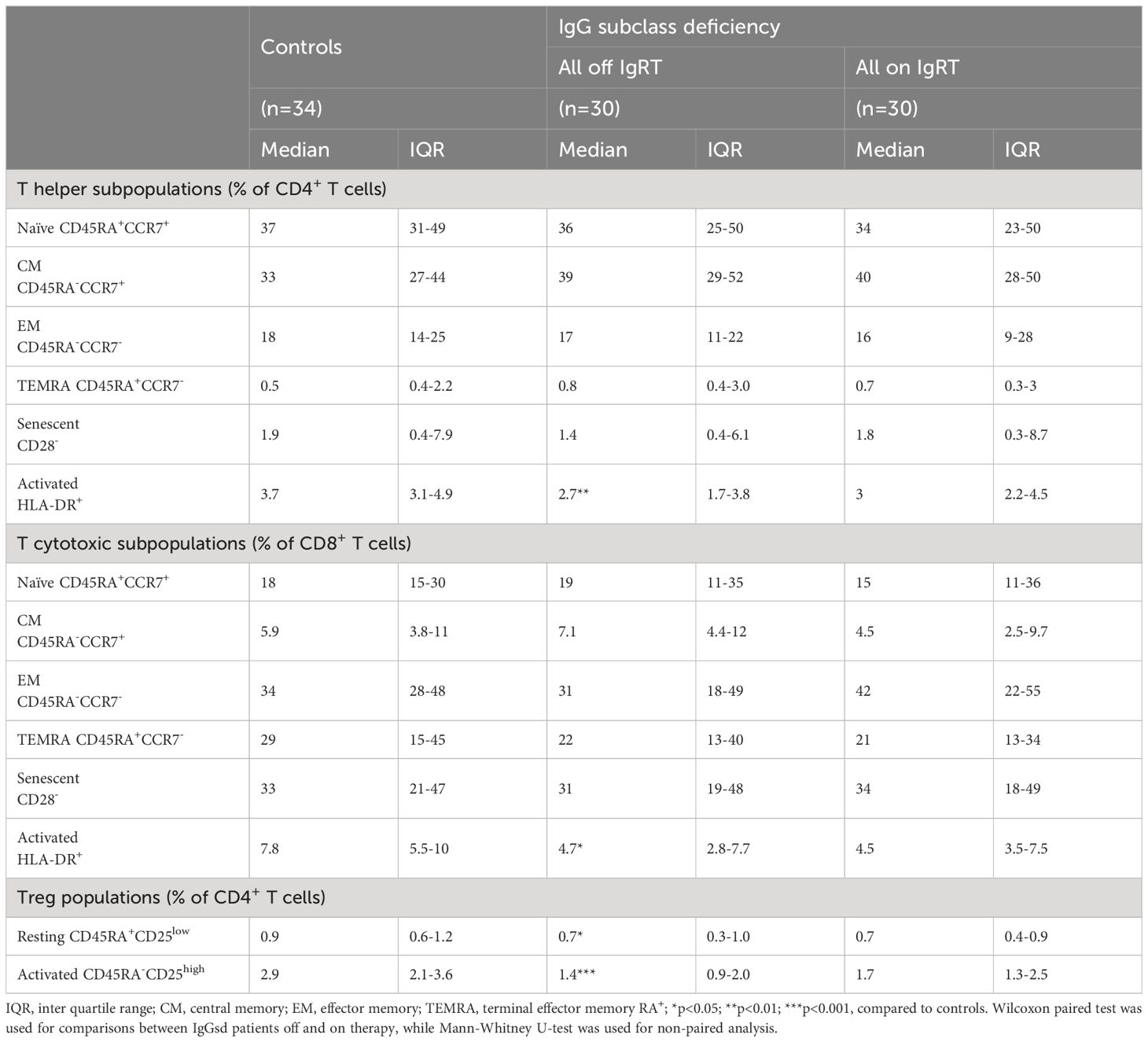
Table 4. T cell subpopulations in patients with IgG subclass deficiency.
Reduced regulatory T cells in patients with IgG subclass deficiency
Further, we found reduced levels of CD4+ Tregs in patients with IgGsd compared to controls (Table 4) and this finding was mainly related to the activated CD45RA-CD25bright Treg subset (Figures 4A, B). There was a high compliance between this gating strategy for Tregs and an alternate gating strategy that included the transcription factor Foxp3 with Spearman correlation (rs) 0.91 for activated Treg cells (Figures 4C, D). When patients were on IgRT, the Treg compartment expanded within the Th cell population, mainly due to increased proportions of activated Treg cells, but the levels were still lower compared with healthy controls (Figure 4A). We found no association between activated Tregs and the presence of autoimmune conditions in patients with IgGsd. In summary, IgGsd was characterized by decreased activated and total Tregs, which were partly restored during IgRT.
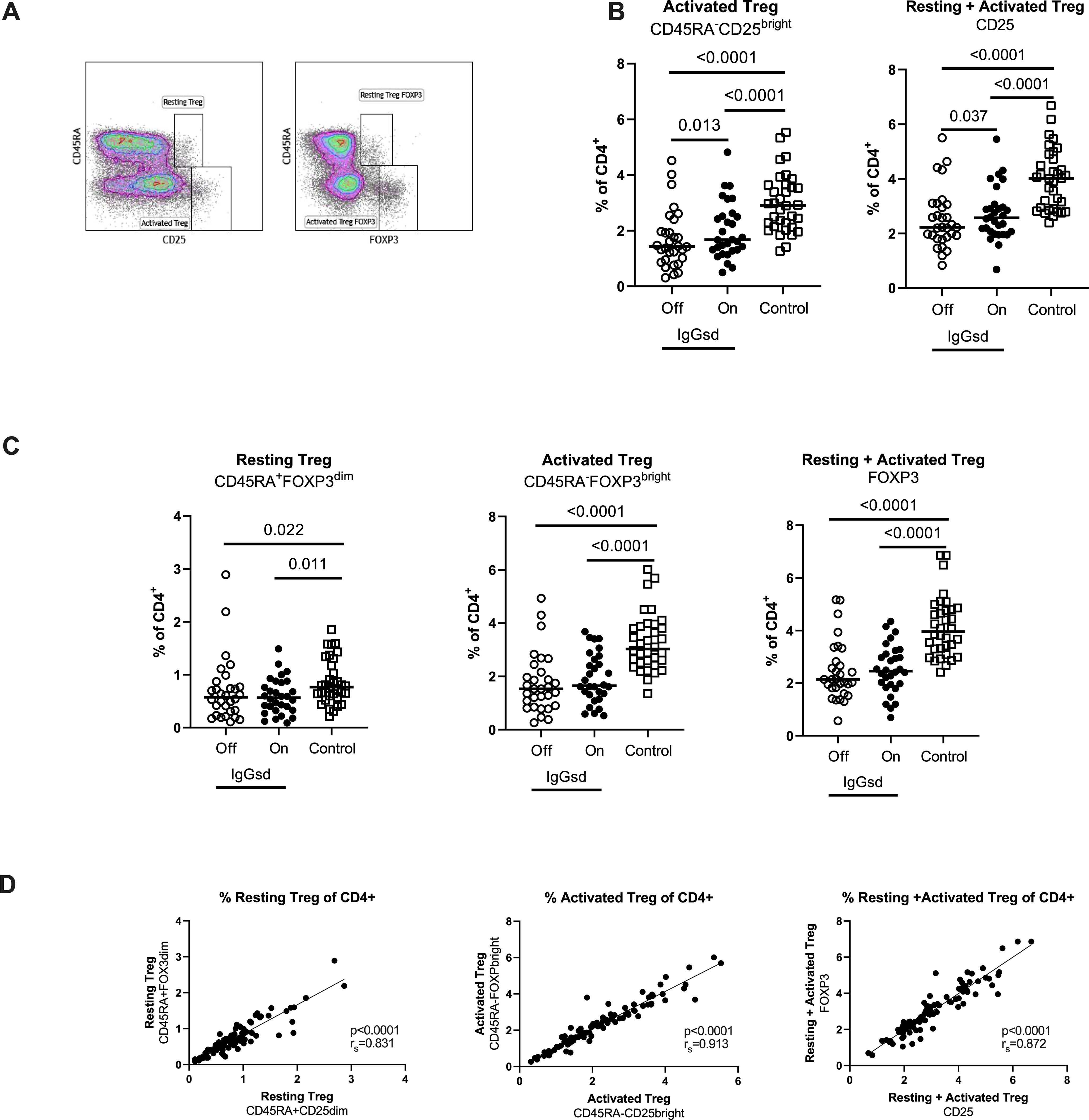
Figure 4. Decreased activated regulatory T cells (Tregs) in IgG-subclass deficiency patients is partly restored during Ig replacement therapy. Frequencies of Tregs were assessed by flow cytometry using two different strategies: cell surface expression of CD25 or intra cellular expression of FoxP3, and the activation status of Tregs was defined by the expression of CD45RA (A). Lower levels of activated Tregs in IgG subclass deficiency (IgGsd) patients (open circle) were partly restored during (filled circle) immunoglobulin replacement therapy (IgGRT) when compared to healthy controls (B). Frequencies of Resting, activated, and resting + activated Tregs defined by FoxP3 and CD45RA expression showed a pattern similar to Tregs defined by CD25 and CD45RA (C). There was a strong correlation between frequencies of Treg populations defined by CD25 and FoxP3, respectively (D). Wilcoxon paired test was performed between IgGsd patients Off and On immunoglobulin replacement therapy, while Mann-Whitney U-test was used for non-paired analysis, and Spearman (rs) for correlation analysis.
Increased plasma levels of costimulatory and regulatory immune checkpoint molecules in patients with IgG subclass deficiency
Shedded transmembrane proteins can be detected in plasma and provide additional aspects of the immune function in patients with IgGsd. Circulating levels of transmembrane receptors were analyzed in samples collected from 28 patients when on and off IgRT and compared to cross-sectional healthy control samples. In our dataset, there were 18 proteins with predominant location in the plasma membrane. Sixteen of them were detected in more than 90% of samples and selected for further analysis (Supplementary Table S2). Whether the patients had ongoing IgRT or had no therapy, did not affect the plasma levels of any factor. Plasma levels of CD40, CD137 and CD244 were elevated in patients with IgGsd, both when on and when off IgRT, compared to controls (Figure 5A). These proteins regulate immune responses, and soluble isoforms of CD40 and CD137 are secreted by various cell types including immune cells (28, 29). Circulating levels of soluble CD40, CD137 and CD244 all showed a moderate correlation with activated HLA-DR+ CD8+ T cells (Figure 5B). Moreover, soluble CD137 correlated moderately and soluble CD40 correlated weakly with activated CD25+ memory B cells (Figure 5C). In addition, levels of CD6, CD215 (interleukin, IL-15 receptor A), CD218 (IL-18 receptor 1) and CD318, a ligand for CD6 also known as CDCP1 (for CUB domain-containing protein 1) were elevated in IgGsd patients in samples collected when on IgRT (CD218) or when off IgRT (CD6, CD215 and CD318) (Table 5). All these proteins are membrane bound, but CD6, CD215 and CD318 are also produced as soluble molecules (30–32). Taken together, the elevated levels of proteins involved in immune responses such as immune checkpoints associated with T and B cell phenotypes in IgGsd.
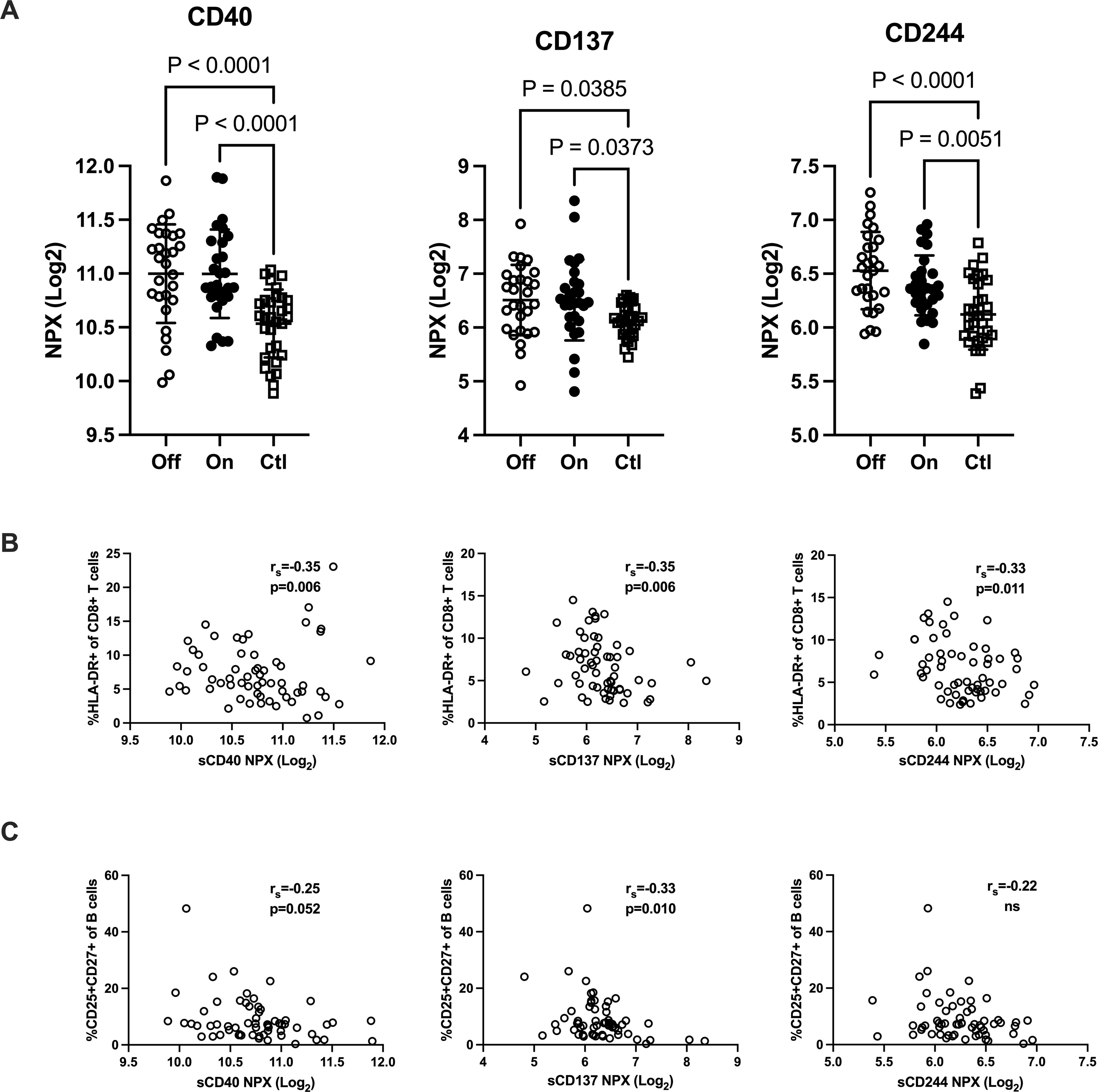
Figure 5. IgG-subclass deficiency is associated with increased plasma levels of checkpoint molecules. Plasma levels of T cell response regulating factors CD40, CD137 (TNFRSF9) and CD244 were analyzed with proximity extension assay in patients with IgG-subclass deficiency (IgGsd) when off and on Ig-replacement therapy (IgRT) compared to healthy controls (A). Correlation of T cell response regulating factors with activated CD8+ T cells (B) and activated memory B cells (C). Data is presented as normalized protein expression (NPX) values in Log2 scale. Error-bars indicate 1SD, p represent ANOVA with Tukey posttest and Spearman correlation (rs), respectively.
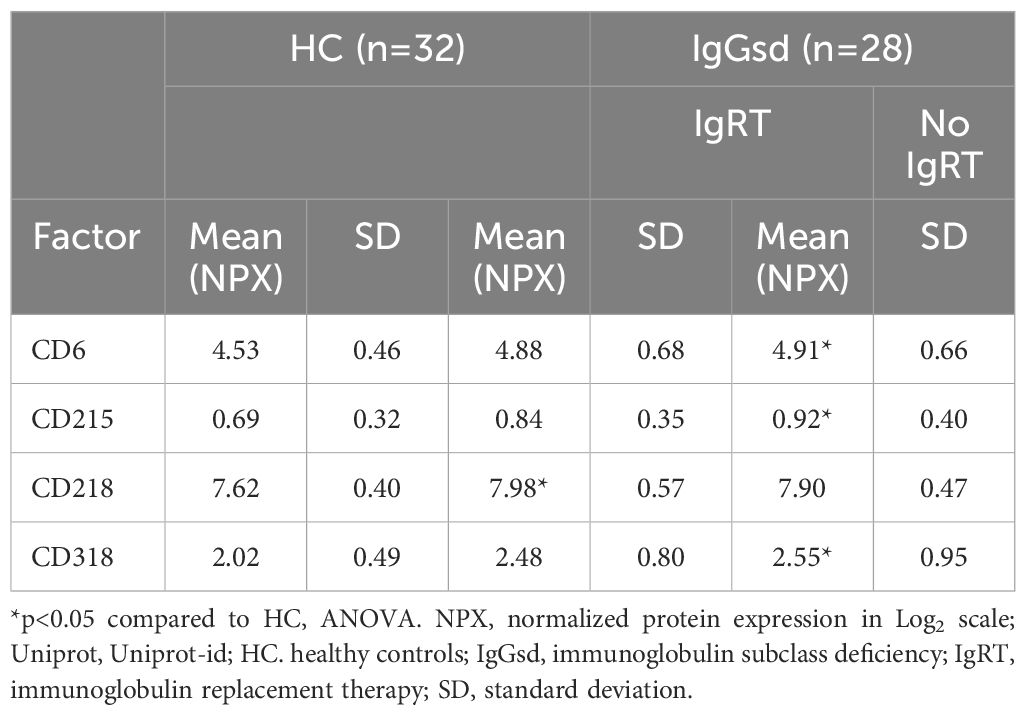
Table 5. Elevated immune checkpoints in plasma of patients with IgGsd.
Discussion
In this prospective study we have compared the adaptive arm of the immune system in patients with IgGsd, on and off subcutaneous IgRT, with a group of cross-sectional healthy controls. The study protocol required at least 6 months of IgRT discontinuation before considering patients as being off IgRT to ensure that residual IgRT not was captured in laboratory testing. IgG3 deficiency was the most common finding in our cohort, either alone or in combination with other IgG-subclass deficiencies. Only 23% and 43% of patients were diagnosed with IgG1 and IgG2 subclass deficiency, respectively, which may explain why IgG1- and IgG2 median values were within the reference intervals based on the 2.5 – 95 percentiles of a healthy population (33). Serum levels of IgG4 were not tested since IgG4 constitute less than 5% of total serum IgG and is of minor importance in infection defense (34). In addition, the lower reference limit (2.5 percentile) of IgG4 is 0.05g/L which makes the clinical relevance of an isolated IgG4sd uncertain (33).
Most patients with IgGsd had one or several comorbidities such as autoimmunity, atopy, or reduced lung function. We found no connection with any of the comorbidities and more complex IgG subclass deficiencies or subnormal levels of IgG and IgA. Overall, the IgGsd patients in this cohort shared clinical characteristics with other cohorts of patients with different forms of IgGsd (6, 7, 10). Absolute numbers of B cells, NK cells, Th cells and cytotoxic T cells did not differ between IgGsd patients and healthy controls and were not affected by IgRT, which is in line with previous reports of minor PAD (10, 17, 35). Deeper analysis of the B cell and T cell subsets in our study showed that activated (CD25+) memory B cells were lower in IgGsd and normalized during IgRT, which might reflect the important role of antibodies in B-cell activation. Low frequencies of switched memory B cells are common in CVID and correlate with lymphoproliferation (15, 36). Only two of the patients had subnormal switched memory B cells, suggesting that the underlying pathophysiology of IgGsd may differ from other more severe PAD in which low switched memory B cells is a common finding such as CVID and CVID-like disorders.
The activation status of Th cells and cytotoxic T cells was lower in IgGsd compared to controls. While on IgRT, the effector memory subset of CD8+ T cells increased, which might reflect an improvement of T cell function. This is in line with the reduced infection burden in patients with IgGsd treated with IgRT (6).
Compared to controls, decreased frequencies of Treg subsets were a consistent finding in IgGsd both while on and off IgRT. This is a novel finding in IgGsd, and its importance remains to be determined. Tregs suppress immune responses by cellular interactions with antigen presenting cells, and modulation of the local environment by the secretion of immune inhibitory cytokines and deprivation of immune stimulatory cytokines (37). Most autoimmune diseases display defects in either the number or function of Tregs (18). Lower Tregs could contribute to the development of autoimmunity and atopic disease in patients with IgGsd. Also, bronchiectasis can be considered a chronic inflammatory disease (38), which may worsen by impaired Treg function. When phenotyping Treg cells we used an appropriate strategy that avoids inclusion of false positive Foxp3 cells (37), and we found a good correlation between alternative gating strategies. Tregs have been extensively studied in CVID, where decreased proportions of Treg cells are considered to be associated with an increased risk of autoimmune manifestations (39), even if some conflicting data exist (40). In our cohort, we did not find a clear association between Treg frequency and autoimmunity, but this needs to be further explored in future studies of IgGsd. In CVID it has been demonstrated that IgRT transiently increases the number of Tregs immediately after infusion (41), but a long-term follow-up trial did not show any effect of IgRT on the Foxp3+ regulatory T cell compartment in CVID (24). Our observation, that the Treg compartment was only partially restored during IgRT may relate to the ability of IgG to act as an effector molecule as well as a regulatory molecule. There are accumulating data that high-dose intravenous immunoglobulin therapy increases the number of circulating Tregs and their immunosuppressive capacity in many inflammatory and autoimmune conditions (42). Based on these findings, a more significant effect on the Treg compartment in IgGsd during IgRT would have been expected. However, the use of lower doses and that subcutaneous administration instead of intravenous, as well as chronic longstanding inflammation rather than an acute inflammatory condition in IgGsd and other PAD, make any comparison difficult.
Increased levels of circulating costimulatory and regulatory immune checkpoints molecules, may contribute to the aberrant B- and T cell subpopulations found in patients with IgGsd. Both CD40 and CD137 belong to the TNF receptor superfamily and are key regulators of adaptive immune responses. CD40 is expressed not only by immune cells but also by epithelial and endothelial cells (29). Soluble (s) CD40 results mainly from shedding (43) and can act as competitor of the interaction of membrane CD40 with its ligand CD154 and negatively affect B cell activation and T cell dependent antibody responses (29). Soluble CD137 is secreted by activated T cells and encompasses the whole extracellular domain (44). Hence, increased sCD137 may counteract CD137 and CD137 ligand interactions, hampering T cell activation (28) and T cell dependent antibody responses (45). Negative correlations between sCD40 as well as between sCD137 and the proportions of activated memory B cell and activated cytotoxic T cells in this study may reflect inadequate immune suppression in IgGsd, which may confer impaired immune responses and increased susceptibility to infections. CD244 is a transmembrane receptor present mainly in NK cells and T cells, which provides stimulatory and inhibitory signals, regulate cytotoxic cellular immune responses. To date, there are no evidence for soluble forms of CD244. However, increased plasma levels of CD244 have been reported in hepatitis C infection (46). The elevated levels of sCD244 in the present study could reflect increased shedding and/or turnover of T cells and NK cells.
CD6, CD215, CD218 and CD318 were other dysregulated transmembrane receptors in patients with IgGsd. CD6 is a pattern recognition receptor expressed by most T cells, which is released into circulation by proteolytic cleavage (46). CD318 is widely expressed by epithelial cells and is a ligand for CD6 (32). CD6 has an inhibitory effect on T cell responses (47). Increased levels of sCD215 (IL-15RA) have been reported in synovial fluid of patients with rheumatoid arthritis (48). Membrane CD218 (IL-18R1) is involved in activating T helper 1 responses (49) but knowledge about how sCD218 may affect immune responses is missing. CD218 was the single factor only elevated in samples collected when on IgRT. In summary, increased levels of sCD40 and sCD137 may contribute to the decreased activation of B cells and T cells found in patients with IgGsd and it can be hypothesized that sCD40 and sCD137 impair immune responses and the secretion of IgG in IgGsd.
In a previous study, where we investigated a large number of secreted cytokines in plasma of patients with IgGsd, levels were unaffected by IgRT (25). Together these findings indicate no anti-inflammatory effects of IgRT in IgGsd. Instead, we found that CD4+ T cell activation and B cell activation were increased, and partly restored, as well as increased effector memory CD8+ T cells in patients when on IgRT. These findings imply that IgRT may augment the adaptive arm of the immune response in IgGsd. Increased soluble checkpoints both when on and off IgRT, may be driven by underlying inflammatory conditions or tissue damage not affected by IgRT. However, increased soluble immune checkpoints and aberrations in B- and T cell subpopulations clearly are indicatives of an underlying immune dysfunction in patients with IgGsd.
The prospective design and the paired samples for comparing immunological phenotypes when the patients were on and off IgRT are strengths of this study. The low number of patients enrolled in the study is a limitation, and the findings need to be verified in other larger cohorts. The heterogeneity of patients with deficiencies of different IgG subclasses and their different combinations of comorbidities makes it difficult to draw any general conclusions from this cohort. Using severe lung disease as an exclusion criterion may also have impact on the results, since patients with manifest lung disease are the ones that benefits most from IgRT.
In summary, numbers of B cells, T cells and NK cells were similar in patients with IgGsd and healthy controls. Deeper analysis revealed decreased activation of CD4+ and CD8+ T cells, and memory B-cells in patients with IgGsd. The findings were consistent with negative correlation between plasma levels of immune checkpoint molecules and the levels of CD8+ T cell and memory B cell activation. The activation of T cells and immune checkpoint levels were not affected by IgRT, but the activation of B memory B cells was partly restored. Activated Tregs were reduced in patients with IgGsd, and partly restored during IgRT. Hence, IgRT may contribute to improved immunity in patients with IgGsd. The significance of the association between low activated Tregs and IgGsd remains unclear and should be further investigated in future studies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Regional Ethical Review Board in Linköping, Sweden (Dnr 2011/506-31). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
PW: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing. MH: Data curation, Formal analysis, Investigation, Writing – original draft, Writing – review & editing. DA: Conceptualization, Data curation, Formal analysis, Writing – original draft, Writing – review & editing. JB: Conceptualization, Formal analysis, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing. CD: Conceptualization, Formal analysis, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. MN: Data curation, Formal analysis, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. ÅN-A: Conceptualization, Data curation, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JE: Conceptualization, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing. SN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by ALF Region Östergötland (RÖ-969621) (SN), Medical Research Council of Southeast Sweden (FORSS-964617) (PW) and Region Jönköping County (Futurum-64617) (PW).
Acknowledgments
The authors would like to dedicate this paper to the memory of our dear colleagues and friends, MH (1978–2023) and JE (1952-2024). We would also especially like to acknowledge Ramona Fust (Department of Infectious Medicine Linköping University Hospital) and Carina Hagstedt (Department of Infectious Medicine, Ryhov County Hospital, Jönköping) for help with patient sample collection. We further acknowledge the help of the flow cytometry unit at the Department of Clinical Immunology and Transfusion Medicine, Linköping University Hospital for help with sample collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1442749/full#supplementary-material
References
1. Bousfiha A, Moundir A, Tangye SG, Picard C, Jeddane L, Al-Herz W, et al. The 2022 update of IUIS phenotypical classification for human inborn errors of immunity. J Clin Immunol. (2022) 42:1508–20. doi: 10.1007/s10875-022-01352-z
2. Bonilla FA, Khan DA, Ballas ZK, Chinen J, Frank MM, Hsu JT, et al. Practice parameter for the diagnosis and management of primary immunodeficiency. J Allergy Clin Immunol. (2015) 136:1186–205.e1-78. doi: 10.1016/j.jaci.2015.04.049
3. Seidel MG, Kindle G, Gathmann B, Quinti I, Buckland M, van Montfrans J, et al. The european society for immunodeficiencies (ESID) registry working definitions for the clinical diagnosis of inborn errors of immunity. J Allergy Clin Immunol Pract. (2019) 7:1763–70. doi: 10.1016/j.jaip.2019.02.004
4. Barton JC, Bertoli LF, Barton JC, Acton RT. Selective subnormal igG1 in 54 adult index patients with frequent or severe bacterial respiratory tract infections. J Immunol Res. (2016) 2016:1405950. doi: 10.1155/2016/1405950
5. Berger M, Geng B, Cameron DW, Murphy LM, Schulman ES. Primary immune deficiency diseases as unrecognized causes of chronic respiratory disease. Respir Med. (2017) 132:181–8. doi: 10.1016/j.rmed.2017.10.016
6. Olinder-Nielsen AM, Granert C, Forsberg P, Friman V, Vietorisz A, Björkander J. Immunoglobulin prophylaxis in 350 adults with IgG subclass deficiency and recurrent respiratory tract infections: a long-term follow-up. Scand J Infect Dis. (2007) 39:44–50. doi: 10.1080/00365540600951192
7. Kim JH, Park S, Hwang YI, Jang SH, Jung KS, Sim YS, et al. Immunoglobulin G subclass deficiencies in adult patients with chronic airway diseases. J Korean Med Sci. (2016) 31:1560–5. doi: 10.3346/jkms.2016.31.10.1560
8. De Gracia J, Rodrigo MJ, Morell F, Vendrell M, Miravitlles M, Cruz MJ, et al. IgG subclass deficiencies associated with bronchiectasis. Am J Respir Crit Care Med. (1996) 153:650–5. doi: 10.1164/ajrccm.153.2.8564113
9. Flume PA, Chalmers JD, Olivier KN. Advances in bronchiectasis: endotyping, genetics, microbiome, and disease heterogeneity. Lancet. (2018) 392:880–90. doi: 10.1016/S0140-6736(18)31767-7
10. Barton JC, Barton JC, Bertoli LF, Acton RT. Characterization of adult patients with IgG subclass deficiency and subnormal IgG2. PloS One. (2020) 15:e0240522. doi: 10.1371/journal.pone.0240522
11. Padem N, Wright H, Fuleihan R, Garabedian E, Suez D, Cunningham-Rundles C, et al. Rheumatologic diseases in patients with inborn errors of immunity in the USIDNET registry. Clin Rheumatol. (2022) 41:2197–203. doi: 10.1007/s10067-021-06044-4
12. Hultberg J, Blixt E, Göransson R, Adolfsson J, Govender M, Larsson M, et al. In-depth immune profiling reveals advanced B- and T-cell differentiation to be associated with Th1-driven immune dysregulation in common variable immunodeficiency. Clin Immunol. (2023) 257:109816. doi: 10.1016/j.clim.2023.109816
13. Cunningham-Rundles C, Bodian C. Common variable immunodeficiency: clinical and immunological features of 248 patients. Clin Immunol. (1999) 92:34–48. doi: 10.1006/clim.1999.4725
14. Filion CA, Taylor-Black S, Maglione PJ, Radigan L, Cunningham-Rundles C. Differentiation of common variable immunodeficiency from igG deficiency. J Allergy Clin Immunol Pract. (2019) 7:1277–84. doi: 10.1016/j.jaip.2018.12.004
15. Wehr C, Kivioja T, Schmitt C, Ferry B, Witte T, Eren E, et al. The EUROclass trial: defining subgroups in common variable immunodeficiency. Blood. (2008) 111:77–85. doi: 10.1182/blood-2007-06-091744
16. Kofod-Olsen E, Jørgensen SE, Nissen SK, Westh L, Møller BK, Østergaard L, et al. Altered fraction of regulatory B and T cells is correlated with autoimmune phenomena and splenomegaly in patients with CVID. Clin Immunol. (2016) 162:49–57. doi: 10.1016/j.clim.2015.11.003
17. Abrahamian F, Agrawal S, Gupta S. Immunological and clinical profile of adult patients with selective immunoglobulin subclass deficiency: response to intravenous immunoglobulin therapy. Clin Exp Immunol. (2010) 159:344–50. doi: 10.1111/j.1365-2249.2009.04062.x
18. Dominguez-Villar M, Hafler DA. Regulatory T cells in autoimmune disease. Nat Immunol. (2018) 19:665–73. doi: 10.1038/s41590-018-0120-4
19. Abdou NI, Greenwell CA, Mehta R, Narra M, Hester JD, Halsey JF. Efficacy of intravenous gammaglobulin for immunoglobulin G subclass and/or antibody deficiency in adults. Int Arch Allergy Immunol. (2009) 149:267–74. doi: 10.1159/000199723
20. Albin S, Cunningham-Rundles C. An update on the use of immunoglobulin for the treatment of immunodeficiency disorders. Immunotherapy. (2014) 6:1113–26. doi: 10.2217/imt.14.67
21. SlLIPI. Riktlinjer för utredning, diagnostik och behandling av immunbrster. (2021) Sweden: Sveriges läkares intresseförening för primära immunbrister.
22. Quinti I, Mitrevski M. Modulatory effects of antibody replacement therapy to innate and adaptive immune cells. Front Immunol. (2017) 8:697. doi: 10.3389/fimmu.2017.00697
23. Simón-Fuentes M, Sánchez-Ramón S, Fernández-Paredes L, Alonso B, Guevara-Hoyer K, Vega MA, et al. Intravenous immunoglobulins promote an expansion of monocytic myeloid-derived suppressor cells (MDSC) in CVID patients. J Clin Immunol. (2022) 42:1–13. doi: 10.1007/s10875-022-01277-7
24. Paquin-Proulx D, Santos BA, Carvalho KI, Toledo-Barros M, Barreto de Oliveira AK, Kokron CM, et al. IVIg immune reconstitution treatment alleviates the state of persistent immune activation and suppressed CD4 T cell counts in CVID. PloS One. (2013) 8:e75199. doi: 10.1371/journal.pone.0075199
25. Wågström P, Nilsdotter-Augustinsson Å, Nilsson M, Björkander J, Dahle C, Nyström S. Fatigue is common in immunoglobulin G subclass deficiency and correlates with inflammatory response and need for immunoglobulin replacement therapy. Front Immunol. (2021) 12:797336. doi: 10.3389/fimmu.2021.797336
26. Bioinformatics & Evolutionary Genomics Online. A tool you can calculate the intersection(s) of list of elements . Available online at: https://bioinformatics.psb.ugent.be/webtools/Venn/.
27. Bäck SE, Nilsson JE, Fex G, Jeppson JO, Rosén U, Tryding N, et al. Towards common reference intervals in clinical chemistry. An attempt at harmonization between three hospital laboratories in Skåne, Sweden. Clin Chem Lab Med. (1999) 37:573–92. doi: 10.1515/CCLM.1999.091
28. Luu K, Shao Z, Schwarz H. The relevance of soluble CD137 in the regulation of immune responses and for immunotherapeutic intervention. J Leukoc Biol. (2020) 107:731–8. doi: 10.1002/JLB.2MR1119-224R
29. Wagner AH, Klersy A, Sultan CS, Hecker M. Potential role of soluble CD40 receptor in chronic inflammatory diseases. Biochem Pharmacol. (2023) 217:115858. doi: 10.1016/j.bcp.2023.115858
30. Consuegra-Fernández M, Lin F, Fox DA, Lozano F. Clinical and experimental evidence for targeting CD6 in immune-based disorders. Autoimmun Rev. (2018) 17:493–503. doi: 10.1016/j.autrev.2017.12.004
31. Mortier E, Bernard J, Plet A, Jacques Y. Natural, proteolytic release of a soluble form of human IL-15 receptor alpha-chain that behaves as a specific, high affinity IL-15 antagonist. J Immunol. (2004) 173:1681–8. doi: 10.4049/jimmunol.173.3.1681
32. Enyindah-Asonye G, Li Y, Ruth JH, Spassov DS, Hebron KE, Zijlstra A, et al. CD318 is a ligand for CD6. Proc Natl Acad Sci U S A. (2017) 114:E6912–e21. doi: 10.1073/pnas.1704008114
33. Schauer U, Stemberg F, Rieger CH, Borte M, Schubert S, Riedel F, et al. IgG subclass concentrations in certified reference material 470 and reference values for children and adults determined with the binding site reagents. Clin Chem. (2003) 49:1924–9. doi: 10.1373/clinchem.2003.022350
34. Vidarsson G, Dekkers G, Rispens T. IgG subclasses and allotypes: from structure to effector functions. Front Immunol. (2014) 5:520. doi: 10.3389/fimmu.2014.00520
35. Dinh T, Oh J, Cameron DW, Lee SH, Cowan J. Differential immunomodulation of T-cells by immunoglobulin replacement therapy in primary and secondary antibody deficiency. PloS One. (2019) 14:e0223861. doi: 10.1371/journal.pone.0223861
36. Hultberg J, Ernerudh J, Larsson M, Nilsdotter-Augustinsson A, Nystrom S. Plasma protein profiling reflects TH1-driven immune dysregulation in common variable immunodeficiency. J Allergy Clin Immunol. (2020) 146(2):417–28. doi: 10.1016/j.jaci.2020.01.046
37. Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity. (2009) 30:899–911. doi: 10.1016/j.immuni.2009.03.019
38. Long MB, Chotirmall SH, Shteinberg M, Chalmers JD. Rethinking bronchiectasis as an inflammatory disease. Lancet Respir Med. (2024) 3:S2213–2600. doi: 10.1016/S2213-2600(24)00176-0
39. Azizi G, Hafezi N, Mohammadi H, Yazdani R, Alinia T, Tavakol M, et al. Abnormality of regulatory T cells in common variable immunodeficiency. Cell Immunol. (2017) 315:11–7. doi: 10.1016/j.cellimm.2016.12.007
40. Gupta S, Demirdag Y, Gupta AA. Members of the regulatory lymphocyte club in common variable immunodeficiency. Front Immunol. (2022) 13:864307. doi: 10.3389/fimmu.2022.864307
41. Kasztalska K, Ciebiada M, Cebula-Obrzut B, Górski P. Intravenous immunoglobulin replacement therapy in the treatment of patients with common variable immunodeficiency disease: an open-label prospective study. Clin Drug Investig. (2011) 31:299–307. doi: 10.1007/BF03256928
42. Bayry J, Ahmed EA, Toscano-Rivero D, Vonniessen N, Genest G, Cohen CG, et al. Intravenous immunoglobulin: mechanism of action in autoimmune and inflammatory conditions. J Allergy Clin Immunol Pract. (2023) 11:1688–97. doi: 10.1016/j.jaip.2023.04.002
43. Contin C, Pitard V, Itai T, Nagata S, Moreau JF, Déchanet-Merville J. Membrane-anchored CD40 is processed by the tumor necrosis factor-alpha-converting enzyme. Implications for CD40 signaling. J Biol Chem. (2003) 278:32801–9. doi: 10.1074/jbc.M209993200
44. Shao Z, Sun F, Koh DR, Schwarz H. Characterisation of soluble murine CD137 and its association with systemic lupus. Mol Immunol. (2008) 45:3990–9. doi: 10.1016/j.molimm.2008.05.028
45. Alfaro C, Echeveste JI, Rodriguez-Ruiz ME, Solorzano JL, Perez-Gracia JL, Idoate MA, et al. Functional expression of CD137 (4-1BB) on T helper follicular cells. Oncoimmunology. (2015) 4:e1054597. doi: 10.1080/2162402X.2015.1054597
46. Khera T, Du Y, Todt D, Deterding K, Strunz B, Hardtke S, et al. Long-lasting imprint in the soluble inflammatory milieu despite early treatment of acute symptomatic hepatitis C. J Infect Dis. (2022) 226:441–52. doi: 10.1093/infdis/jiab048
47. Oliveira MI, Gonçalves CM, Pinto M, Fabre S, Santos AM, Lee SF, et al. CD6 attenuates early and late signaling events, setting thresholds for T-cell activation. Eur J Immunol. (2012) 42:195–205. doi: 10.1002/eji.201040528
48. MaChado Diaz AC, Chico Capote A, Arrieta Aguero CA, Rodríguez Alvarez Y, García Del Barco Herrera D, Estévez Del Toro M, et al. Proinflammatory soluble interleukin-15 receptor alpha is increased in rheumatoid arthritis. Arthritis. (2012) 2012:943156. doi: 10.1155/2012/943156
Keywords: predominantly antibody deficiency, IgG subclass deficiency, Ig replacement therapy, T cells, B cells, Tregs, immune checkpoints
Citation: Wågström P, Hjorth M, Appelgren D, Björkander J, Dahle C, Nilsson M, Nilsdotter-Augustinsson Å, Ernerudh J and Nyström S (2024) Immunological characterization of IgG subclass deficiency reveals decreased Tregs and increased circulating costimulatory and regulatory immune checkpoints. Front. Immunol. 15:1442749. doi: 10.3389/fimmu.2024.1442749
Received: 02 June 2024; Accepted: 24 July 2024;
Published: 14 August 2024.
Edited by:
Georgios Sogkas, Hannover Medical School, GermanyReviewed by:
Beatrice Omusiro Ondondo, University Hospitals of Leicester NHS Trust, United KingdomRoberta Bulla, University of Trieste, Italy
Copyright © 2024 Wågström, Hjorth, Appelgren, Björkander, Dahle, Nilsson, Nilsdotter-Augustinsson, Ernerudh and Nyström. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Per Wågström, UGVyLldhZ3N0cm9tQGxpdS5zZQ==
†deceased
‡ORCID: Per Wagström, orcid.org/0000-0001-8549-473X