 Abdullah Al Mamun
Abdullah Al Mamun Chuxiao Shao
Chuxiao Shao Peiwu Geng
Peiwu Geng Shuanghu Wang
Shuanghu Wang Jian Xiao
Jian Xiao
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 21 May 2024
Sec. Inflammation
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1395479
This article is part of the Research TopicThe inflammatory response in cutaneous wound healing processView all 6 articles
The skin, being a multifaceted organ, performs a pivotal function in the complicated wound-healing procedure, which encompasses the triggering of several cellular entities and signaling cascades. Aberrations in the typical healing process of wounds may result in atypical scar development and the establishment of a persistent condition, rendering patients more vulnerable to infections. Chronic burns and wounds have a detrimental effect on the overall quality of life of patients, resulting in higher levels of physical discomfort and socio-economic complexities. The occurrence and frequency of prolonged wounds are on the rise as a result of aging people, hence contributing to escalated expenditures within the healthcare system. The clinical evaluation and treatment of chronic wounds continue to pose challenges despite the advancement of different therapeutic approaches. This is mainly owing to the prolonged treatment duration and intricate processes involved in wound healing. Many conventional methods, such as the administration of growth factors, the use of wound dressings, and the application of skin grafts, are used to ease the process of wound healing across diverse wound types. Nevertheless, these therapeutic approaches may only be practical for some wounds, highlighting the need to advance alternative treatment modalities. Novel wound care technologies, such as nanotherapeutics, stem cell treatment, and 3D bioprinting, aim to improve therapeutic efficacy, prioritize skin regeneration, and minimize adverse effects. This review provides an updated overview of recent advancements in chronic wound healing and therapeutic management using innovative approaches.
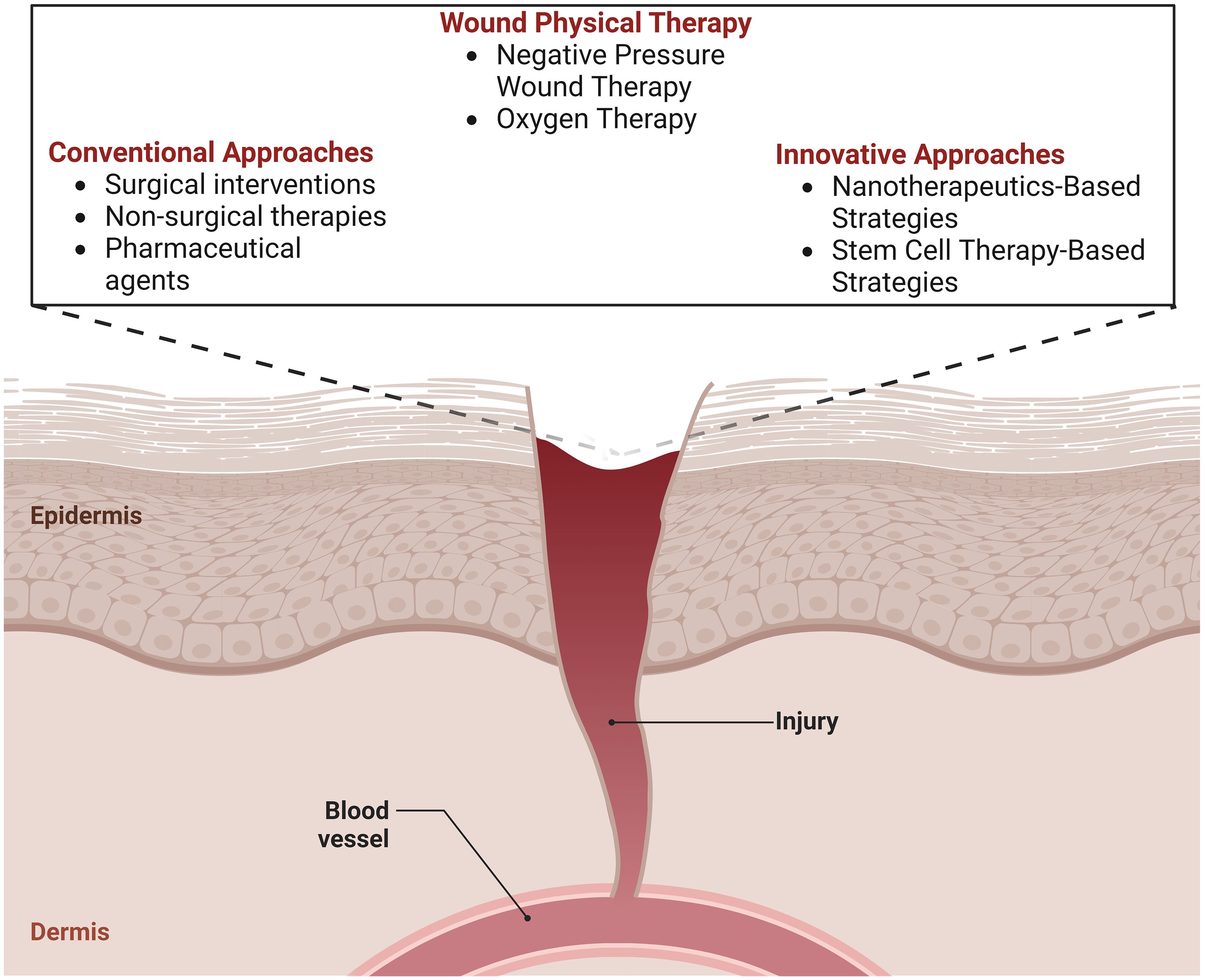
Graphical Abstract
Skin is the largest organ of our body, which accounts for approximately 16% of the total body weight. It is essential for both the maintenance of homeostasis and the functioning of the barrier it provides against the impact of external stimuli (1). Skin integrity is of utmost importance in the preservation of overall health, given that injuries resulting from chronic disorders, burns, trauma, and surgical procedures may lead to impairment and emotional anguish. These consequences provide a significant roadblock to healthcare systems worldwide (2). Multiple cell types must be synchronized sequentially for skin healing. The epidermis is impermeable in healthy skin and protects against external factors (3). The epidermis, which encompasses hair follicles, sebaceous, and sweat glands, provides structural integrity, immunity, and nutrition to the integumentary system. The dermis is characterized by higher levels of extracellular matrix (ECM), vascular, and mechanoreceptors, while subcutaneous adipose tissue functions as an energy reservoir and a consistent supply of growth factors (4). Each skin layer has immune system cells that monitor damage to the skin. Several kinds of cells in various layers must cooperate at crucial stages to repair skin wounds (5, 6). Based on causes and consequences, wounds may be healing (acute) and non-healing (chronic). The healing process of minor acute wounds is mainly facilitated by the inherent regenerative capacity of the skin, which involves cellular mechanisms, remodeling of the ECM, and the presence of growth factors (5, 7). Injuries to the skin and chronic wounds heal slowly owing to infection and fluid loss. These injuries damage the skin’s structure, harming millions and presenting economic and societal problems to global healthcare systems (8, 9). Despite the considerable amount of research conducted, there are still unmet needs in the field of skin wound healing techniques and the treatment of extensive and chronic wounds. These issues primarily arise from difficulties in accurately assessing wounds and effectively managing their care. Hence, the development of upgraded and novel approaches for healing skin wounds has substantial medical significance worldwide (3, 8, 10).
Skin healing from wounds is paramount for sustaining the quality of life and reaching the goal of wound closure. It encompasses a multitude of cell types and mediators operating in a highly intricate sequence (11). Despite the existence of a substantial body of research on the processes behind wound healing, there are several unresolved issues regarding its physiologic regulation (12, 13). Chronic wounds are now a growing problem, being identified at a concerning rate and imposing a substantial economic strain on the medical field. The increasing prevalence of chronic wounds may be attributed to aging populations and the rising rates of obesity and diabetes on a global scale. Consequently, this has resulted in a substantial escalation in the economic costs associated with the management and treatment of these persistent wounds (9). The yearly expenditure to treat chronic wounds exceeds $25 billion, a substantial amount that becomes even more significant when accounting for the additional expenses associated with decreased productivity among afflicted persons and the provision of long-term care in facilities and nursing homes (14). Diabetic foot ulcers and pressure ulcers are significant contributors to morbidity rates and impose a substantial economic burden (15). The investigation of tissue regeneration in chronic wound healing is of utmost importance due to the rising occurrence of diabetes and obesity and the demand for wound care among veterans. The investigation of tissue regeneration in the context of chronic wound healing is crucial in order to effectively tackle these challenges and enhance the overall well-being of patients (9).
Skin wound therapy may be categorized into two main groups: conventional procedures and regenerative approaches. Traditional methods of treating wounds often include measures to limit infection, regular changes of dressings, and the removal of wrecked tissues by debridement (16). Split-thickness skin autografts are a vital medical intervention with limitations like repeated surgeries, limited donor sites, hypertrophic damage, and functional changes. Regenerative wound healing encompasses a range of developing biomedical technologies, such as bioactive biomaterials, innovative dressings for wounds, treatment with stem cells, growth factor administration, gene therapies, and bioengineered skin grafts. These innovative approaches attempt to restore the skin’s original function and repair damaged tissues. This technique enables faster and higher-quality wound healing while minimizing the occurrence of scarring (17, 18). A previous wound healing method centered on layer-by-layer skin regeneration utilizing bioengineered scaffolds or cell-encapsulated hydrogels. Rapid wound healing with little scarring has been achieved using cell-laden matrices, including fibroblasts, keratinocytes, or stem cells (19). Nevertheless, there is potential for enhancement in the spatial distribution and cellular composition to replicate the intricate microarchitecture seen in natural skin tissues (20). Hence, the current emphasis is primarily on the advancement of diverse, developing, and creative approaches to expedite the wound healing process while ensuring the preservation of functional attributes. Within emerging treatment modalities, there has been significant interest in using stem cell-based therapies, specifically those employing various types of stem cells, such as induced pluripotent stem cells and mesenchymal stem cells. These therapies are being explored extensively in clinical and preclinical studies as a component of stem cell-based regenerative medicine (21). The efficacy of stem cell-based therapy is limited by low wound stem cell survival after implantation/grafting owing to a hostile inflammatory environment (22). Stem cell viability enhancement, genome editing, and genetic alterations offer promising methods for personalized wound care for patients with enduring wounds (23).
The main objective of wound treatment is to enhance the accuracy of diagnosis and prognosis to develop individualized treatment strategies. Over the past decade, there has been a significant emphasis on exploring innovative therapeutic methods, such as nanotherapeutics, stem cell therapy, and 3D bioprinting (24, 25). The present article presents an in-depth review of recent breakthroughs in treatments for wound healing. It delves into the molecular mechanisms behind this process, explores traditional treatment modalities, and highlights newer tactics that show promise in this field. The review encompasses an examination of nanotherapeutic methodologies, including the use of nanomaterials, and an exploration of the application of stem cell treatment. Additionally, the article addresses the many difficulties encountered in contemporary wound healing therapy and explores potential advancements in this field.
Skin healing from wounds is a complex phenomenon that incorporates several cellular, humoral, and molecular pathways. It starts promptly upon the appearance of a lesion and may persist for an extended duration, perhaps spanning several years (21). The condition under consideration comprises instances of tissue disruptions resulting in functional impairment. Intrinsic injuries may manifest as either open wounds, which occur on the surface of the body or closed wounds, which entail ruptures to internal organs while the skin remains intact. Either regeneration or repair mechanisms may attain closure. Regeneration entails the replacement of damaged tissue, while skin healing includes the process of fibrosis and subsequent scar formation (26). The phenomenon is sometimes characterized as a symphony or theatrical performance, whereby the interaction among cells, growth factors, and cytokines culminates in the closing of the skin (27). Nevertheless, current research indicates that the insufficiency of a particular cell type or the lack of a mediator may be compensated for by other entities participating in the wound healing process (Figure 1), thus enabling the progression of repair (Table 1) (30, 34).
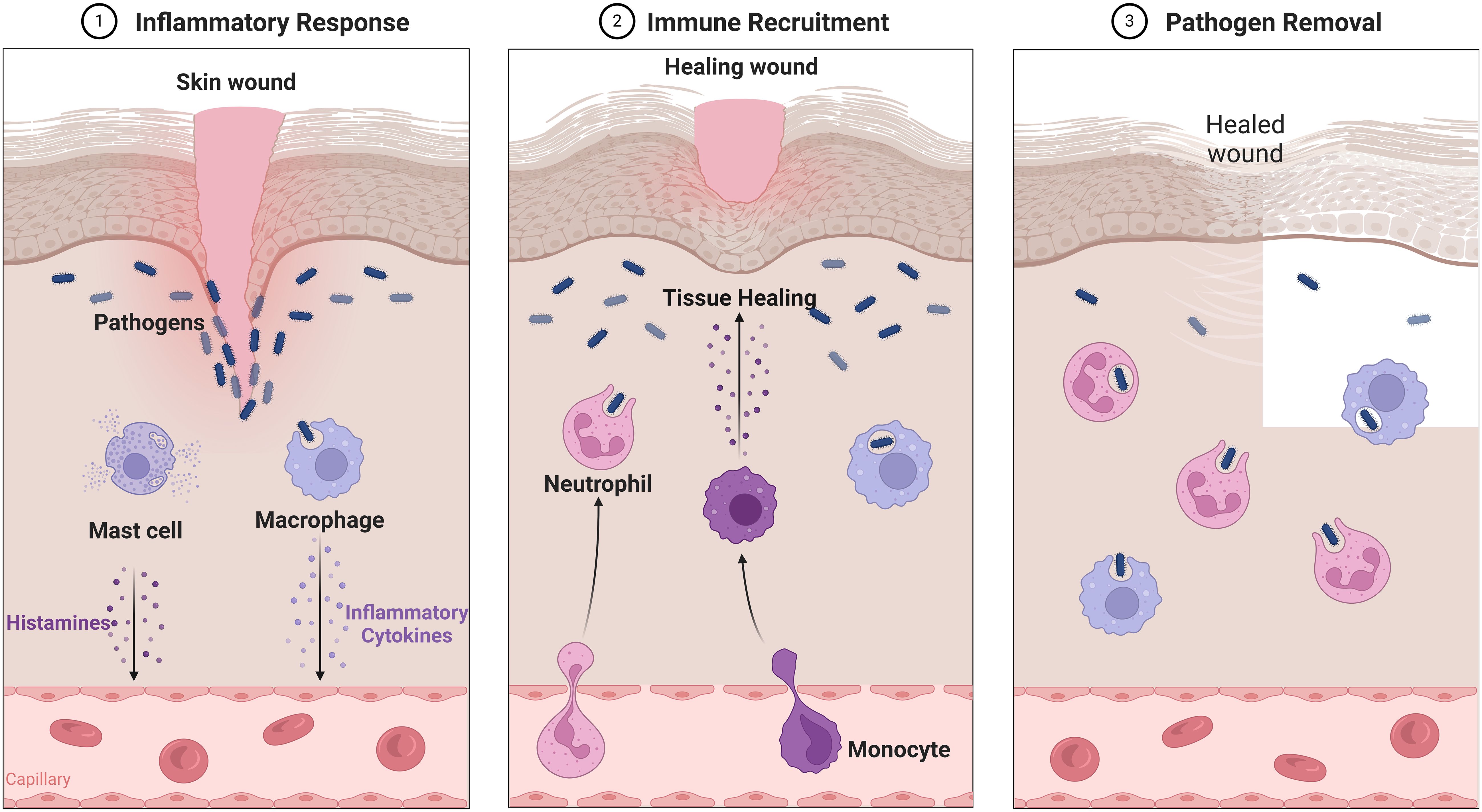
Figure 1 Overview of Wound healing.
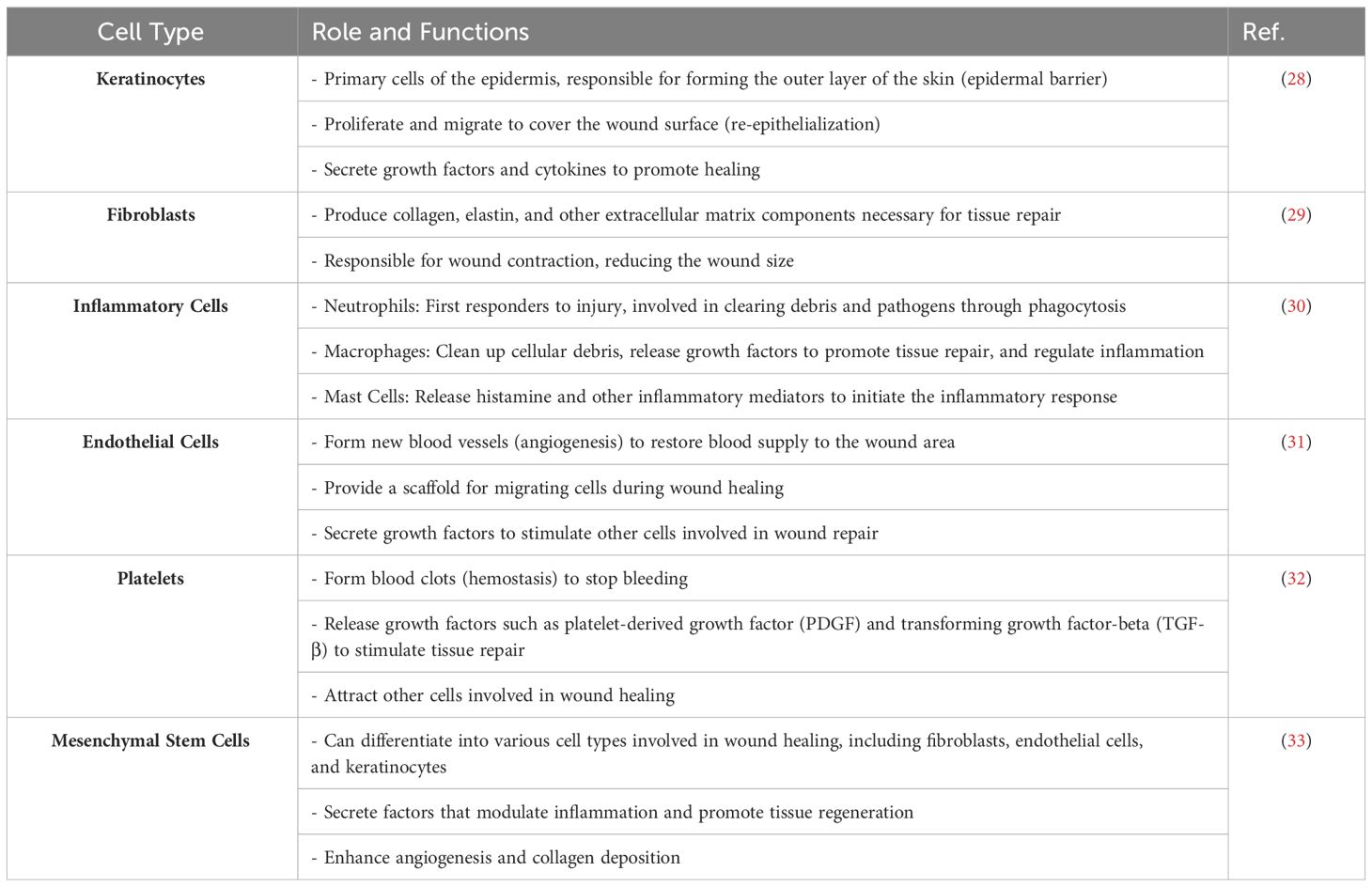
Table 1 The role and functions of different types of cells in skin wound healing.
The efficient process of healing a wound, free from any adverse outcomes, is of utmost importance for the life of an individual. It reinstates the structural integrity of the skin and serves as a protective barrier against potential risks such as dehydration and infection (35). The process of wound healing in adults encompasses a sequence of processes that culminate in the restoration of damaged tissues and the development of scar tissue. The healing process is comprised of several stages of proliferation, inflammation, hemostasis, and remodeling, which occur in a coordinated manner (36). The number of inflammatory macrophages and monocytes infiltrates the tissue more deeply as time goes on, eventually reaching a peak during the proliferative period before gradually decreasing. Early in the process, lymphocytes in the blood migrate to the skin, achieving a point of saturation by day four and continuing to do so for the next two weeks before beginning to decrease. The third step, which involves redesigning the tissue and forming a scar to reinstate the skin’s integrity, occurs in the second week following the damage. This procedure can take many months to complete. This review offers up-to-date information on the critical role that the microenvironment, immune cells, and the connections between these factors play in the process of wound healing (37–39).
Acute wounds are characterized by vascular damage and hemorrhaging, and the first stage of wound healing involves the implementation of measures to mitigate blood loss by means of vasoconstriction and the production of blood clots. The process of hemostasis is triggered when blood components come into contact with the inner lining of blood vessels, leading to the adhesion, aggregation, and formation of an initial hemostatic plug by platelets (35). The procedure mentioned above unfolds via three distinct stages: vasoconstriction, primary hemostasis, and secondary hemostasis. The platelet, which serves as the crucial cell, interacts with fibrinogen, a vital matrix component. The healthy endothelilal cells (ECs) monolayer protects platelets in undamaged skin, preventing premature activation (26). Fibrinogen, which is synthesized by hepatocytes, is distributed throughout the bloodstream and is found inside platelets; nevertheless, it remains unprocessed into fibrin fibers, which are crucial constituents of blood clot formation (40, 41).
In the case of skin injury, the blood arteries inside the skin undergo constriction to impede hemorrhaging. As mentioned above, the process is characterized by the sequential occurrence of primary and secondary hemostasis, which are mediated by two interrelated routes (42). Primary hemostasis encompasses the processes of aggregation of platelets and the development of a platelet plug, which are initiated by the exposure of collagen in the sub-endothelial matrix. Secondary hemostasis is the process by which the coagulation cascade is triggered, leading to the conversion of soluble fibrinogen into insoluble strands that ultimately form a fibrin mesh. The formation of a thrombus serves to halt hemorrhaging while also facilitating the release of complements and growth factors, promoting wound healing via the provision of a supportive scaffold (43).
Following an injury, blood arteries undergo a fast constriction process to minimize bleeding from damaged microvasculature. The method of achieving this phenomenon involves the reflexive contraction of vascular smooth muscle, which is initiated by vasoconstrictors such as endothelin (44). The regulation of vasoconstriction is also influenced by circulating catecholamines, such as epinephrine, norepinephrine, and prostaglandins. Platelets are responsible for the production of platelet-derived growth factor (PDGF). This signaling molecule stimulates the activation of mesenchymal cells, namely smooth muscle cells located inside the walls of blood vessels (45).
Nevertheless, the first reflexive contraction of muscles only temporarily reduces bleeding. This is due to the fact that the wound experiences a rise in hypoxia and acidity, leading to passive muscle relaxation and subsequent resumption of bleeding. The subsequent activation of the coagulation cascade is necessary to control the process of vasoconstriction further and facilitate the resolution of long-term bleeding. Vasoconstriction, accompanied by clot formation, is crucial in preserving life by preventing excessive blood loss. However, this physiological response may also result in impaired local perfusion, heightened glycolytic activity, and alterations in pH levels (43, 46).
After an injury and the subsequent rupture of blood vessels, the thrombogenic subendothelial matrix becomes exposed, which facilitates the binding of platelets and initiates the inside-out signaling route for platelet activation. Consequently, this phenomenon results in the activation of integrins, promoting enhanced platelet adhesion to both neighboring platelets and the adjacent ECM. The external-to-internal signaling route enhances platelet activation and regulates the actin cytoskeleton, resulting in the transformation of the platelet into a cell with a fried-egg-like morphology that exhibits robust adhesion to the ECM, undergoes contraction, and effectively occludes the blood a blood vessel (39, 47, 48). The surface area of the activated platelet is increased as a result of the fusion between intracellular granules and the plasma membrane or surface-connected membranes of the open canalicular system (OCS). The granules mentioned above are responsible for the secretion of more than 300 bioactive compounds, including ADP, serotonin, calcium, histamine, as well as vWF and integrins, which play crucial roles in both primary and secondary hemostasis (42, 49). The surface area of the activated platelet is increased as a result of the fusion between intracellular granules and the plasma membrane or surface-connected membranes of the OCS. The granules, as mentioned above, are responsible for the secretion of more than 300 bioactive compounds, including ADP, serotonin, calcium, histamine, as well as vWF and integrins, which play crucial roles in both primary and secondary hemostasis (50). Activated platelets also secrete molecules such as thromboxane A2, which enhance platelet aggregation, resulting in the formation of the “platelet plug.” The release of cytokines and growth factors by platelets inside the plug plays a crucial role in cellular mediation for the process of healing. The intensity of platelet factor release is highest during the first hour after activation. However, the release of these factors by activated platelets persists for seven days, hence imposing paracrine impacts on many cell types present in the wound (47).
Platelets are crucial in assembling and activating coagulation complexes, triggered by contact with the subendothelial matrix, with traditional coagulation routes recognized in the field. These pathways are triggered upon contact with the subendothelial matrix. The activation of Factor X initiates the process of fibrin synthesis, ultimately resulting in the development of a blood clot. The blood clot consists of cross-linked fibrin, along with cellular components such as erythrocytes and platelets. Additionally, various ECM proteins, including fibronectin, vitronectin, and thrombospondin, are present in the clot (6, 51). Factor XIII is responsible for the covalent crosslinking of fibrin, which results in the binding of the aggregated platelet plug. This process leads to the formation of a final secondary hemostasis plug, also known as the thrombus. The thrombus functions as the temporary ECM for the invasion of various cells throughout the following phases of the healing process. The adhesion of platelets is facilitated by activated integrin receptors located on their surface (52, 53). The adhesion of platelets is facilitated by activated integrin receptors located on their surface. Platelets in the clot degranulated, producing powerful inflammatory cell chemoattractants, local fibroblast and EC activation factors, and vasoconstrictors. CCL5, thrombin, TGF β, PDGF, and VEGF are important chemokines that regulate coagulation and restrict blood vessel formation (54). The process of fibrin breakdown and the subsequent activation of the complement system are essential components in initiating the inflammatory response, promoting the formation of new blood vessels in wounds, and enabling the proliferation of stromal cells. Fibrin interacts with integrin CD11b/CD18 on invading monocytes and neutrophils, as well as fibroblast growth factor 2 (FGF 2) and VEGF, hence facilitating the process of wound tissue vascularization. In cases of thrombocytopenia, macrophages and T lymphocytes present at the site of injury serve to compensate for the deficiency of platelet-derived growth factors (PDGFs) and commence the inflammatory phase (35, 38, 55–57).
Inflammation in wound healing involves a complex coordination of immune system cells and molecular interactions, including macrophages, neutrophils, lymphocytes, and signaling molecules. It begins after hemostasis, where blood arteries dilate due to coagulation and complement cascades, with Bradykinin and anaphylatoxins playing crucial roles (58). Anaphylatoxins increase blood vessel permeability, allowing monocytes and neutrophils to reach injury sites. They also stimulate mast cell production of histamine and leukotrienes, amplifying inflammation response. This enhances EC permeability, disrupting cell-to-cell connections and facilitating inflammatory cell movement at wound sites (59). Neutrophils, which are the primary leukocytes present in wounds, play a crucial role in combating infection by using several mechanisms, such as antimicrobial peptides, proteases, and reactive oxygen species (ROS). These entities possess a limited duration of existence and experience programmed cell death through mechanisms involving caspases and cathepsin D (59, 60). Neutrophils do not hinder wound healing, but their prolonged absence may lead to chronic nonhealing wounds. Monocytes, prevailing inflammatory cells at wound sites, migrate towards the injury site through the chemotactic process facilitated by other immune cells like neutrophils and keratinocytes. CCL2 chemokines direct monocyte migration, while other immune cells like neutrophils and keratinocytes also play a role (61). Monocytes migrate to wound sites and differentiate into macrophages, which is crucial for wound healing. Macrophages eliminate apoptotic neutrophils, present antigens, and produce cytokines and growth hormones. They also eliminate diseased cells and present antigens (62). Growth factors like TGF-α, TGF-β, bFGF, PDGF, and VEGF activate and recruit cells, promoting wound healing through proliferation, ECM formation, and angiogenesis. VEGF also promotes angiogenesis by eliciting the expression of LIGHT, a cytokine within the TNF-α family, in macrophages, potentially causing apoptosis (63). The coordinated interaction between immune system cells and signaling molecules is crucial for the systematic advancement of wound healing, including the initial inflammatory response as well as later stages of repair of tissues and remodeling. A comprehensive comprehension of these processes is of utmost importance in the development of efficacious treatment approaches for wound healing and the management of chronic wound diseases (54).
Macrophages play a crucial role in the process of wound healing since the lack or dysfunction of these cells dramatically impedes the closure of wounds. Research indicates that the depletion of macrophages during the process of skin wound healing might result in poor tissue disposal, a decrease in the number of fibroblasts, and a delay in the healing process. Nevertheless, it is important to note that inflammation is not an essential prerequisite for the process of wound healing of skin (30, 64). The findings of a study using PU.1 null mice have shown that these mice exhibited comparable wound repair rates to their wild-type littermates while notably lacking scar formation. These observations imply that the presence of inflammation may not be a need for the process of wound healing (61, 62). Studies show that modulating the inflammatory response can help reduce scar formation in wound healing. However, impaired macrophage activity at the wound site can hinder inflammation resolution, especially in diabetic wounds. This continuous inflammatory state can compromise wound closure and worsen scar formation, especially in diabetic wounds where the continuous inflammatory state prevents a resolving phenotype (54, 65, 66). Lipid mediators like protectins, resolvins, lipoxins, and maresins regulate inflammation and control wound healing. They elicit actions to resolve and counteract inflammation. B and T lymphocytes are crucial for wound healing as they initiate targeted immune responses against pathogens and foreign substances (62). Mast cells play a vital role in wound healing by triggering inflammation and resolving it through apoptosis, facilitated by interferon-c and TNF α. They also influence wound healing and fibrosis processes. However, their compromised healing in mice and their association with keloids and hypertrophic scars suggest the need for further research into their specific function and potential therapeutic applications (66). Understanding immune cell function in wound healing is crucial for developing effective therapeutic interventions, especially in chronic or impaired situations, to enhance closure and minimize scarring.
Neutrophils, comprising 50% to 70% of leukocytes in adults, are crucial in initiating inflammation at sites of acute inflammation or infection. They exist in a dormant state and have a lifetime of 8-12 hours while circulating in circulation and 1-2 days when residing in tissues. Macrophages play a crucial role in initiating the inflammatory phase at wound sites, triggered by stimuli such as growth factors, chemokines, and N-formyl peptides. The buildup of neutrophils at wound sites reaches its maximum during the early phase of inflammation and decreases after four days (67–70). Neutrophils in wounds facilitate the leukocyte recruitment cascade towards inflamed tissue, secreting chemoattractants, releasing ROS and antibacterial proteins, effectively eradicating infections (71, 72). Neutrophils regulate innate and adaptive immune responses through intercellular communication with chemokines, cytokines, and immune cells. Their presence of proteases enhances antibacterial efficacy and tissue penetration. However, the overproduction of proteolytic enzymes can damage receptors, growth factors, and ECM and impede vascular processes, highlighting the potential dangers of neutrophil overproduction (73).
Neutrophils, activated and producing neutrophil extracellular traps (NETs), are crucial in trapping and removing pathogens. They are discharged through suicidal and NETosis mechanisms, allowing neutrophils to survive and participate in tasks like phagocytosis. Prompt clearance of neutrophils is essential for inflammation resolution, as failure may lead to chronic wounds like venous leg ulcers, diabetic foot ulcers, and pressure ulcers (70, 72, 74, 75). This highlights the significance of prompt clearance for good wound healing and reduction of inflammation in situations both normal and pathological circumstances.
Monocytes are essential for the human immune system, acting as a defense mechanism against infections (74). Originating from the bone marrow, they differentiate into macrophages and dendritic cells, each with distinct immune responses. Monocytes can be classified into three subsets: CD14++CD16− classical monocytes, CD14++CD16+ intermediate monocytes, and CD14+CD16++ nonclassical monocytes, each with unique functions, size, morphology, and transcriptional profiles (76).
Research using deuterium labeling has provided insights into the life cycle of monocytes in the bone marrow. Monocyte precursors differentiate into classical monocytes, followed by a postmitotic maturation phase (77). Their lifespan is around one day before apoptosis or circulation exit. A smaller percentage matures into intermediate monocytes, lasting four days (65). A significant proportion of intermediate monocytes transform into nonclassical monocytes, lasting seven days before cell death (78). Monocyte migration is a regular process, allowing them to persist in tissues, acquire antigen-presenting capabilities, and differentiate into macrophages (79). The dynamic nature and adaptability of immune responses are crucial in understanding monocyte-macrophage populations in skin tissue (65, 77–79).
Following tissue injury, macrophages identify molecular patterns linked to damage or infections, activating monocytes and neutrophils to migrate toward the wound site. Monocytes secrete chemokines and cytokines, causing neutrophils to chemotaxis towards the wound location. Neutrophils expel their granules, attracting inflammatory monocytes that differentiate into macrophages (78). These immune cells have flexibility, allowing them to differentiate into diverse phenotypes based on the wound site’s conditions. The recognition of patterns initiates a series of interconnected reactions, including the synthesis of inflammatory chemicals and pro-inflammatory cytokines (80). Macrophages and neutrophils are essential components in wound healing, actively participating in the removal of pathogens, deceased cells, and tissue remnants. Both exhibit similar capabilities, including phagocytosis, killing intracellular mechanisms, and generating NETs (80, 81). After debris clearance, neutrophils collaborate with macrophages to terminate inflammation and restore homeostasis. The shift towards an anti-inflammatory state promotes tissue restoration and restructuring during wound healing (78, 82–84).
Innate lymphoid cells (ILCs) are essential in the innate immune response, distinguishing them from T cells, B cells, and NK cells. The ILC family consists of three subsets: Group 1, NK cells responsible for secreting IFN-γ and TNF-α, and ILC2 cells involved in wound healing (85). The activation of ILC2s by IL-33 promotes reepithelialization and wound healing (86). Invariant NK T cells significantly impact skin wound healing by increasing IFN-γ production, releasing growth factors like VEGF and TGF-β, enhancing collagen accumulation, facilitating myofibroblast differentiation, and stimulating blood vessel formation. Invariant NK T cells also help mitigate neutrophil-organized inflammatory responses, expediting wound healing. These various innate lymphocyte subsets are crucial contributors to skin wound healing and the immune system environment (39, 87, 88).
Skin injuries trigger an immune response using pattern recognition receptors (PRRs), including TLR, NOD-like receptors, and C-type lectin receptors. This triggers the release of pro-inflammatory cytokines and the influx of macrophages and neutrophils (89). Skin wounds cause DCs to break down PAMPs and DAMPs, delivering them to naïve CD8+ T lymphocytes in draining lymph nodes (89). T cells differentiate into effector and central memory T cells, and apoptosis occurs when skin-homing receptor-expressing TEM moves to wound sites and releases immune mediators (90, 91).
Secondary lymphoid organs express lymph node-homing receptors by TCM cells, leading to their differentiation into TEM cells. These TEM cells travel to distal lymph nodes, establishing systemically immune memory. Local DCs move towards inflamed skin areas, offering antigens to skin-resident CD8+ Trm cells. This interaction leads to the proliferation of CD8+ Trm cells and the recruitment of effector memory T cells (TEM), contributing to pathogen clearance (92). CD8+ Trm is localized in the epidermis, serving as the first defense mechanism against reinfections (93, 94). The skin’s immune systems respond efficiently to external assaults, facilitating wound healing and establishing immunological memory. Understanding these physiological mechanisms could help develop therapeutic interventions for enhancing immune system responses and wound healing.
The skin’s immune system relies heavily on CD4+Foxp3+ Tregs, which regulate immune responses and contribute to the skin’s microenvironment (95, 96). These cells, which express CCR6 and CLA, move and aggregate in skin hair follicles, promoting wound reepithelialization and modulating inflammation. They also limit IFN-γ production and control inflammatory macrophages (39, 97).
Tregs, or regulatory T cells, persist in the skin after pathogen clearance and play a crucial role in maintaining long-term immunological control in healthy adult skin (98). CD4+ T cells secrete cytokines that induce inflammatory responses, maintaining immunological homeostasis and defense against infections. Their diverse roles highlight their importance in skin protection (99).
B cells are essential in the humoral immune response, transforming into plasma cells that produce antibodies. They also transport antigens to T cells and regulate local immunological responses through the secretion of cytokines and growth factors (100). Studies have shown that mice with spleen deprivation experience a delay in wound healing after injuries (101). B cells treat wounds by generating antibodies against injured tissue and producing cytokines that repair wounds (102).
A study by Sirbulescu et al. found that mature B cells accelerate wound healing, reducing healing time by 2-3 days. This highlights the potential of B lymphocytes in wound healing. B cells are integral to the immune system and play a pivotal role in various immunological responses (103). Recent studies have highlighted the importance of antibodies in wound healing, as they generate antibodies, present antigens, and regulate immune responses by releasing cytokines. Understanding the multifaceted involvement of B cells in wound healing not only advances our understanding of immunity but also presents opportunities for treatments (6).
During wound healing, mast cells secrete antimicrobial peptides, enzymes, VEGF, and histamines, which promote neutrophil influx and vascular permeability, preventing skin infections (104). Histamine from mast cells encourages keratinocyte growth and re-epithelialization, while histamine and tryptase boost fibroblast proliferation and collagen production, contributing to wound contraction (6, 105). A model of fetal wound healing found that a rise in mast cell numbers is associated with skin fibrosis and scarring. In 15 embryonic day wounds, injection of mast cell lysate causes scar development, while deletion of mast cells lowers scar development in 18 embryonic day wounds (106). However, further characterization is needed to understand the precise processes involved in mast cell participation in scar development. Mast cells have functional variability depending on their tissue and microenvironment, leading to several subsets with unique activities in wound healing (Figure 2) (107). Further investigation is needed to understand the distinct functions performed by different subsets of mast cells in wound healing, particularly in cases with prolonged duration or compromised healing capacity.
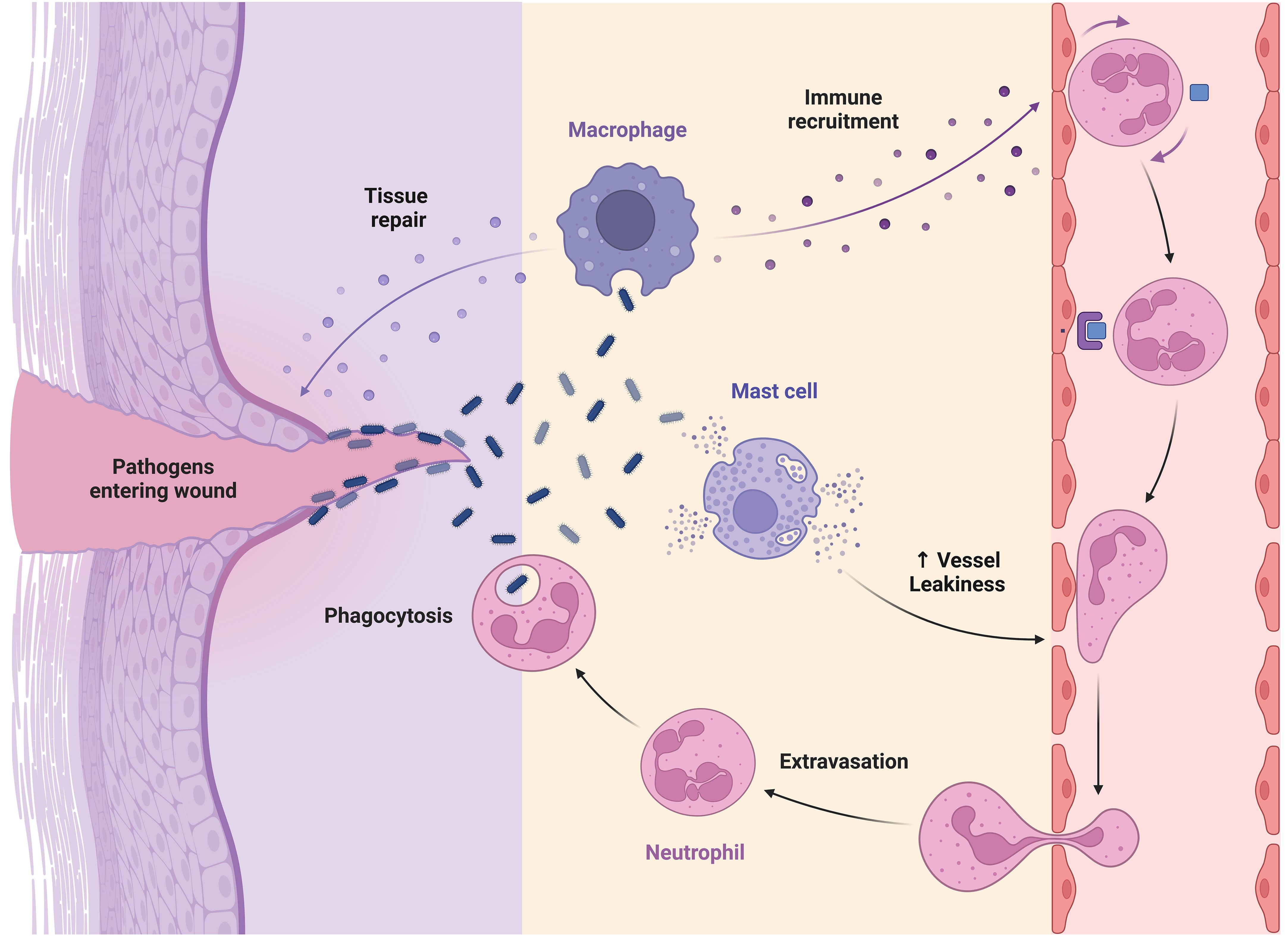
Figure 2 Role of macrophage, mast cell, and neutrophil in wound healing.
Skin-derived cells (pDCs) are essential in the immune system, facilitating T-cell responses and presenting antigens. Langerhans cells, first discovered in the 19th century, are derived from early myeloid progenitors and continue to exist into maturity (6, 108). They are responsible for exposing antigens to T cells inside the dermis and relocating to the discharging lymphatic system to initiate T-cell responses (109).
In the dermis of mice, there are two primary subtypes of resident DCs: CD11b DCs and CD103 DCs. CD103 DCs are capable of cross-presenting antigens, detecting deceased cells, and perceiving stimuli like F-actin and DAMPs, initiating immunological responses (110). The significance of skin-derived cells (pDCs) in wound healing has received less attention than their role in fighting infection. Researchers can use single-cell technology to understand better the roles of different types of immune cells in immune responses and wound healing. The classification of Langerhans cell types and dermal DCs is a subject of discussion (111).
Apoptosis, a regulated form of cell death, serves a critical function in the stages of tissue healing, as shown by its complex involvement in the resolution of these phases (112). The occurrence of apoptosis in inflammatory cells may be noticed as early as 12 hours after the occurrence of a lesion, as shown by the findings of Brown et al. in their research conducted in 1997. Additional investigations conducted on rats have demonstrated the occurrence of apoptosis in scab-inflammatory cells and myofibroblasts, which play a critical role in wound healing and tissue regeneration. This process was seen to begin around the twelfth day, reach its highest point on the twentieth day, and then resolve by the sixtieth day (113).
The research findings demonstrate a temporal association between the process of myofibroblast apoptosis and the healing of wounds, explicitly highlighting the role of stromal keratocytes (114). The initiation of cellular processes crucial for corneal wound healing is facilitated by apoptosis, whereas a delay in healing might occur due to delayed phagocytosis of apoptotic neutrophils by macrophages. The investigation also examines the involvement of apoptotic cells in the release of growth signals that induce cell proliferation. The fundamental determinant of wound healing is the process of apoptosis in immune cells, which prompts inquiries about the principal component (115).
The process of necrosis, which involves tissue healing and inflammation, is improbable to serve as the primary mechanism for lowering cellularity in wounds. The method of emigration is characterized by inefficiency and energy expenditure, hence making apoptosis the most rational strategy for reducing cell numbers throughout the stages of healing. Apoptosis serves as a ubiquitous mechanism for the elimination of unnecessary cells and tissues while concurrently mitigating the risk of inducing further inflammatory responses. The induction of apoptosis in inflammatory cells may be triggered by the presence of invading organisms and nonviable tissue. Similarly, fibroblasts engaged in the process of collagen deposition can also suffer apoptosis. Following the process of wound maturation, it has been shown that ECs and any residual fibroblasts undergo a gradual disappearance without any noticeable signs or symptoms (116).
Brown et al.’s mouse dermal wound investigation showed that apoptosis is essential for tissue healing. After the injury, inflammatory cells apoptosis within 12 hours, indicating epithelial factors inhibit inflammation. Clinicians have found that wound closure or grafting reduces inflammation (113).
Esmouliere et al. demonstrated that apoptosis reduces fibroblasts and aids wound healing. The shift from granulation to scar tissue, a crucial tissue healing step, was studied. Inflammatory cells of exposed wound scabs showed early apoptosis. The research found that myofibroblasts, important contractile cells in wound healing, undergo apoptosis beginning at day 12, peaking at day 20, and ending at day 60 (117). As the wound progressed, cells creating new blood vessels showed a comparable apoptotic pattern, indicating a coordinated reduction in cellularity. This indicates cell-to-cell communication. Covering a 10-day-old incision with a particular skin flap caused myofibroblast death within hours (118).
A comprehensive understanding of the mechanism by which apoptosis drives the decline in fibroblast and myofibroblast function is of utmost importance in the context of wound healing. Failure to regulate this process in a timely manner may result in persistent apoptotic dysfunction, aberrant collagen production, and the creation of excessive scar tissue (117–119). The use of growth factor therapy expedites the progression of apoptotic patterns and the healing of wounds, hence reducing the duration of the inflammatory phase. Animals with diabetes exhibit a delay in the healing process of wounds and an increase in apoptotic cells, which may be attributed to several processes associated with diabetes and variable degrees of damage in the healing process (118, 120).
The cellular process of autophagy, which involves the degradation and recycling of cellular components, is known to have a significant impact on wound healing. It actively participates in several phases of the healing process, contributing to its overall efficiency and effectiveness (121). The early phase of inflammation after an injury serves to commence the process of wound healing by recruiting monocytes and neutrophils to the site of damage. Neutrophils are essential in the defense against microorganisms and the facilitation of inflammation by using antibacterial and proinflammatory processes such as phagocytosis, formation of ROS, degranulation, and the release of neutrophil extracellular traps (122). Autophagy is of great importance in the context of neutrophil-specific activities since it has been seen to enhance the phagocytic activity of human neutrophils when subjected to Streptococcus pneumoniae. The reduction of autophagy leads to a decrease in rates of phagocytosis. The investigation using mice that lack specific autophagy-related genes (Atg5/7) demonstrates a reduction in neutrophil degranulation and levels of ROS, therefore emphasizing the complex relationship between autophagy and neutrophil functionality (Figure 3) (123).
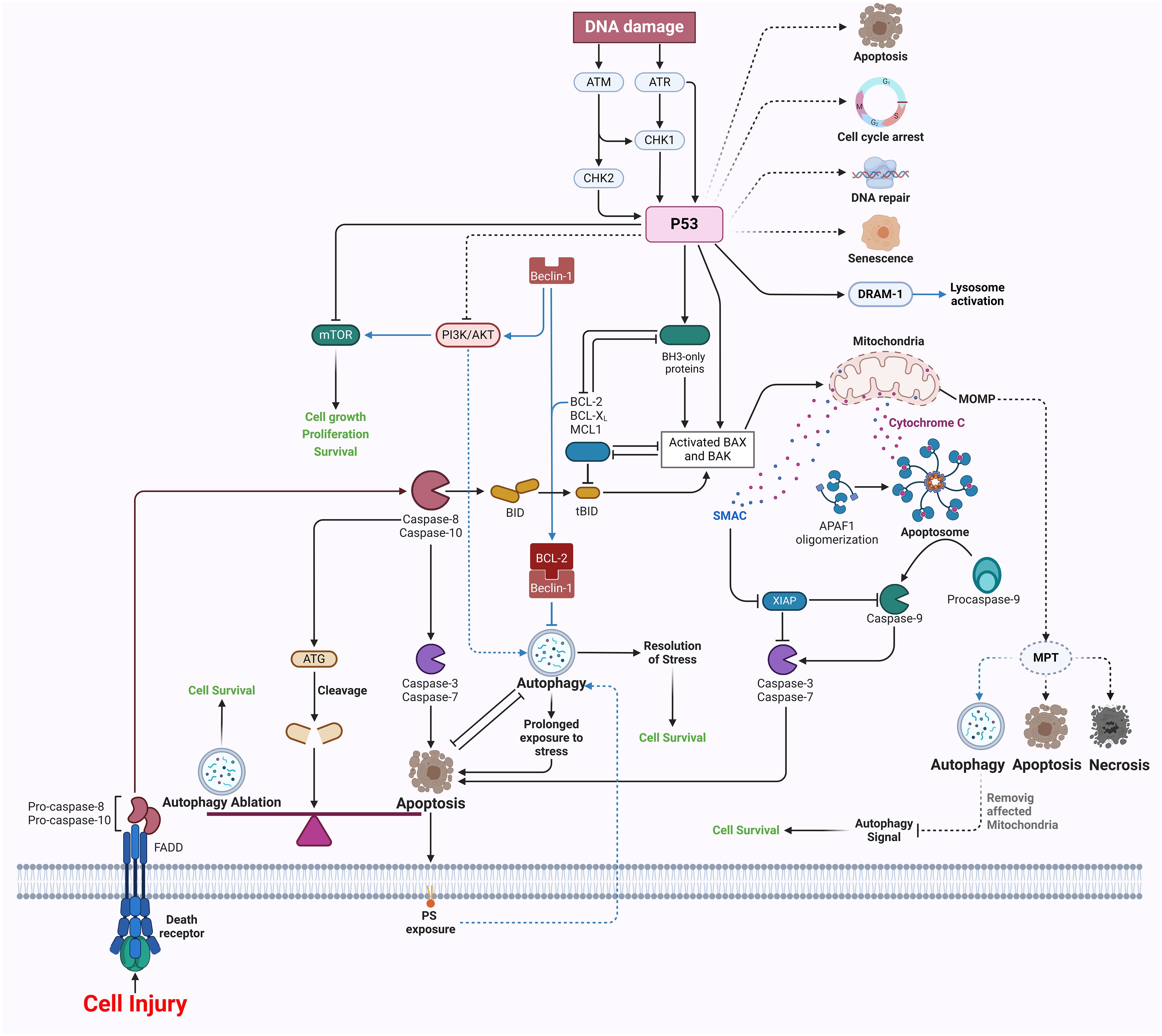
Figure 3 Apoptosis and autophagy in wound healing.
Moreover, the stimulation of autophagy enhances the creation of NETs. It increases the survival rate in mice afflicted with sepsis, therefore emphasizing its significance in the immune response. Furthermore, the impact of autophagy on macrophages is evident in the observation that the administration of 3-methyladenine amplifies their capacity to engulf pathogens, therefore emphasizing the intricate interplay between autophagy and macrophage functionality (121, 124). Moreover, the impairment of autophagy in macrophages results in their polarization towards the M1 phenotype, while the stimulation of autophagy favors the M2 phenotype, hence diminishing inflammation and facilitating tissue regeneration. However, the precise underlying mechanism remains ambiguous and needs additional research (125).
Recent research has demonstrated that MSCs have crucial functions in the process of tissue regeneration and wound healing. These functions include encouraging the growth of new blood vessels (angiogenesis), controlling inflammatory responses, and improving the formation of new epithelial tissue (126). Prior research has shown that controlling autophagy could be a successful approach to enhance the survival of MSCs and enhance the results of wound healing (127). A prior investigation discovered that palmitate stimulates the programmed cell death of MSCs by causing an increase in the levels of ROS inside the cells. However, the activation of autophagy, which occurs via the ROS–JNK–p38 MAPK signaling pathway, safeguards MSCs from undergoing apoptosis (128). Overexpression of hypoxia-inducible factor-1α enhances the survival of MSCs in low oxygen circumstances by stimulating autophagy via the suppression of PI3K/AKT/mTOR signaling (129).
Furthermore, the serine/threonine kinase aurora kinase A triggers autophagy by specifically targeting FOXO3a in order to safeguard adipose-derived stem cells from death caused by hyperglycemia. Other research has shown that blocking microRNA (miR)-34a enhances the effectiveness of MSCs in treating diabetic wounds by activating the sirtuin-1/FOXO3a pathway-mediated autophagy (130). Furthermore, An et al. demonstrated that autophagy inducer-pretreated MSCs injected subcutaneously stimulate VEGF production by triggering MSC-specific paracrine signaling via the ERK1/2 pathway, which in turn improves wound healing (121, 126).
Ferroptosis is characterized by the absence of nucleosomal DNA condensate, chromatin condensate, and apoptotic body development, distinguishing it from conventional necrosis features such as organelle enlargement and cytoplasmic rupture (131). Ferroptosis is a metabolic anomaly characterized by a diminution in the size of mitochondria, a reduction in cristae density, and an increase in membrane density. Notably, the nucleus remains unaffected in terms of its size. The condition is based on a metabolic imbalance that relies on iron, resulting in the excessive buildup of lipid ROS and subsequent cell death. Cysteine metabolism is the fundamental process (132). The cellular uptake of cystine occurs through a cysteine-glutamate antiporter known as system xc-, while the efflux of glutamate takes place. Thioredoxin reductase 1 (TrxR1) facilitates the reduction of cystine, leading to its conversion into glutathione (GSH) via the enzymatic actions of GCL and GSS (133). The lipid-repairing enzyme GPX4 is responsible for the conversion of phospholipid hydroperoxides into non-peroxide forms, therefore effectively suppressing their pro-oxidative properties. The interference of Erastin with the uptake of cystine by the system xc- transporter results in the depletion of glutathione, thus inhibiting the activity of GPX4. Consequently, this disruption leads to the buildup of ROS in lipids, an excess of iron, and an enhanced vulnerability to ferroptosis (134). The condition of iron overload leads to the occurrence of cellular ferroptosis, a process in which external Fe3+ molecules bind to ferritin, subsequently entering the cell via the transferrin receptor 1 (TFR1) and undergoing reduction to Fe2+ facilitated by STEAP3. The introduction of an excessive amount of Fe2+ ions into the Fenton reaction leads to the occurrence of ferroptosis (133, 135).
There is a strong correlation between ferroptosis and skin wounds (136). Multiple investigations have shown that the use of ferroptosis inhibitors may enhance the healing of diabetic wounds by suppressing the process of ferroptosis. For instance, when ferrostatin-1 (a substance that inhibits ferroptosis) is applied directly to a wound in rats with diabetes, it may speed up the healing process by blocking ferroptosis via the activation of the PI3K/Akt signaling pathway (137). Furthermore, several studies have shown that the use of DFO, an alternative form of ferroptosis inhibitors, might enhance the healing process of diabetic wounds. Gao and colleagues discovered that the simultaneous use of DFO and hydroxysafflor yellow A in a hydrogel may expedite the recovery of diabetic wounds in rats by promoting angiogenesis (138).
Excessive exposure to radiation may injure the nearby blood vessels and lead to the development of angiosclerosis. This occurs due to the detrimental effects on the structure and functions of proteins and DNA, resulting in a delay in the healing process of wounds (139). Gan et al. have shown that injecting plasma-derived exosomes (RP-Exos) locally may enhance the healing of rat irradiation wounds by increasing the growth, movement, cell division, and survival of fibroblasts. Additionally, it has been shown that RP-Exos can interfere with the ferroptosis pathway, hence preventing ferroptosis in irradiated fibroblasts (140). In addition to irradiation treatment options for tumors, excess exposure to UV radiation, particularly UVA and UVB, may also result in UV-induced cutaneous wounds. Kavita Vats and her colleagues have shown that an overabundance of UVB radiation may trigger inflammation and the death of human keratinocytes by generating ferroptosis. This process can be suppressed by ferrostatin-1. In addition, an overabundance of UV radiation may lead to a deficiency in GSH, which in turn disrupts the balance of redox reactions in the body (Figure 4) (136, 141).
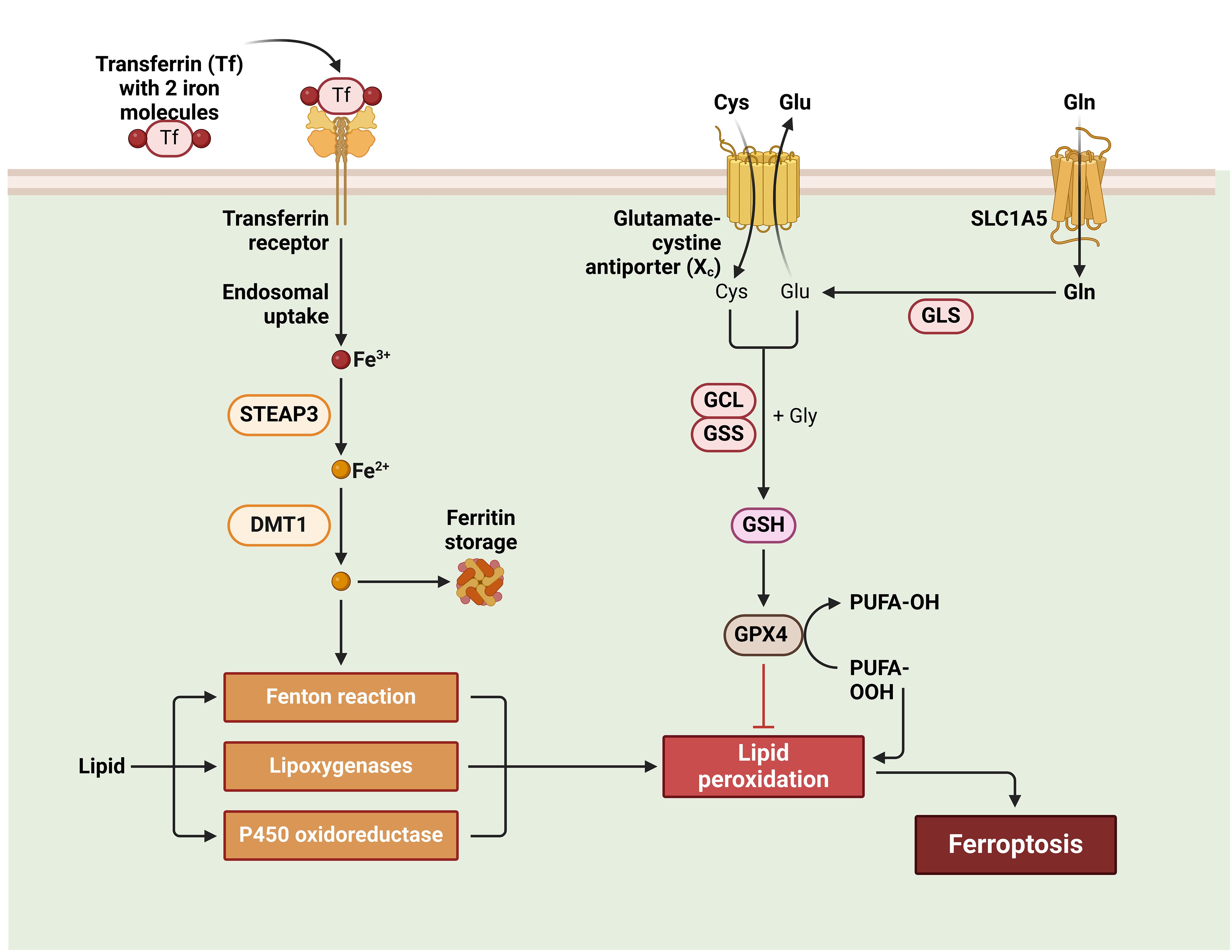
Figure 4 Ferroptosis in wound healing.
Ferroptosis, which involves lipid metabolism, requires lipid peroxidation. Free polyunsaturated fatty acids (PUFAs) are converted into PUFAs-phosphatidylethanolamine (PUFAs-PE), especially arachidonic acid (AA) and adrenaline, which are substrates for lipid peroxidation. ACSL4 and LPCAT3 are critical enzymes in this pathway (136, 142, 143). ACSL4 and LPCAT3 regulate Ferroptosis. LPCAT3 creates PUFAs-PE from PUFAs-CoA, while ACSL4 creates PUFAs-CoA. Ferroptosis results from PUFAs-PE oxidation and lipid peroxidation. ACSL4 is expressed more in ferroptosis-sensitive cells like HepG2 and HL60. MLE cells without LPCAT3 are more resistant to ferroptosis, lowering cell death (Figure 5) (136, 144).
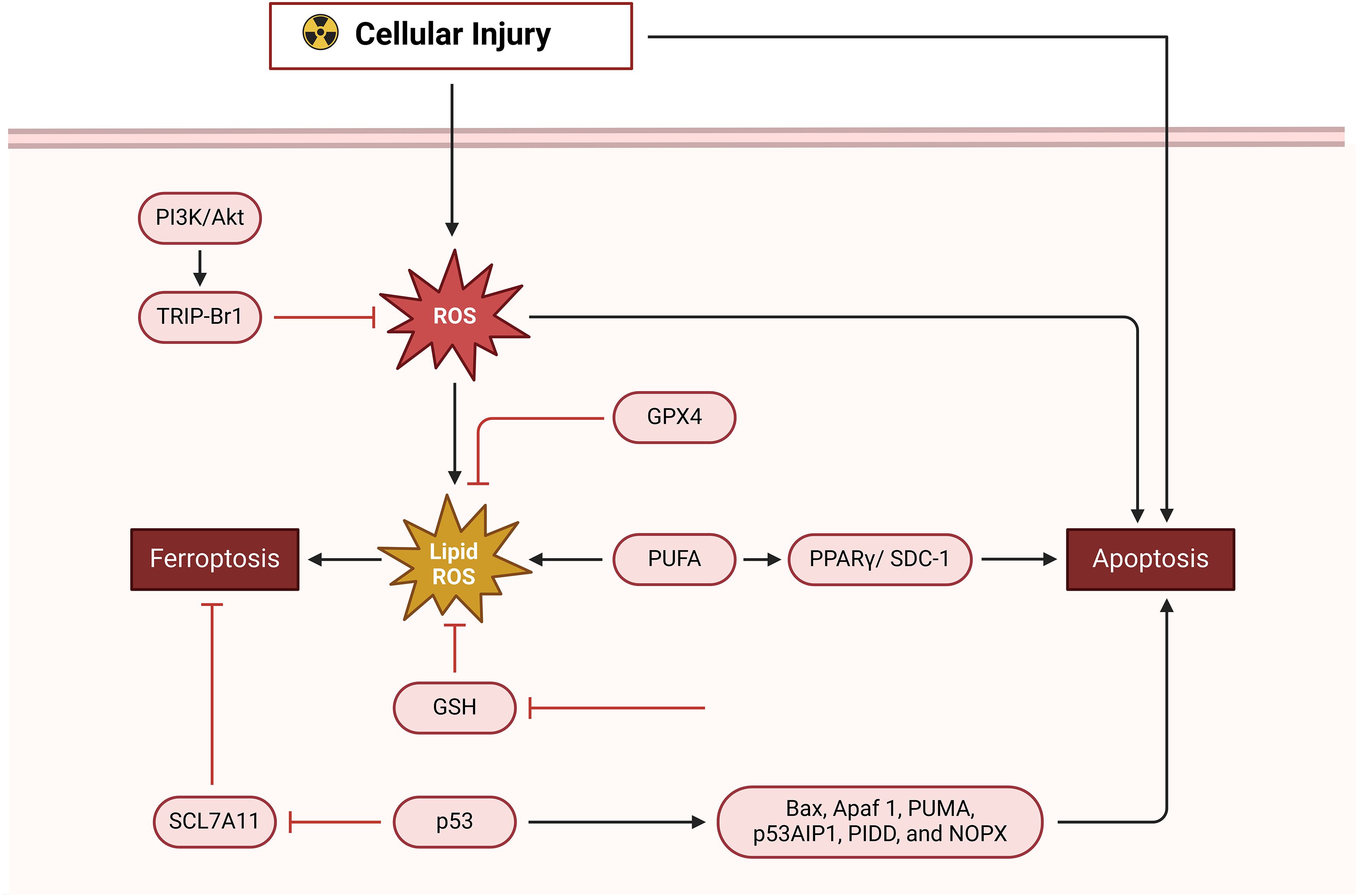
Figure 5 Apoptosis and ferroptosis in wound healing.
Chronic wounds are characterized by an overactive inflammatory response, which hinders the normal progression of the wound-healing process. This disruption ultimately results in the development of scars since it interferes with the sequential progression of the four phases of wound healing (145). The NLRP3 inflammasome, which is primarily located in cutaneous epithelial tissues, plays a pivotal role in orchestrating the immunological response of the organism. Recent studies have demonstrated that the administration of mulberry leaf and fruit extract (MLFE) has the potential to augment the process of wound healing, with a specific focus on its impact on the NLRP3 inflammasome. The combination of MLFE with mulberry leaf extract has been shown to exhibit enhanced anti-inflammatory and anti-obesity properties (146). The research proposes that MLFE can restore NLRP3 inflammasome levels in individuals with obesity, which might facilitate prompt wound healing. This finding underscores the possible associations between MLFE and obesity (147, 148).
TFNAs, DNA nanomaterials, promote corneal and skin healing with their angiogenic, antioxidant, anti-inflammatory, anti-fibrotic, and low-toxicity functions (146, 149). TFNAs heal diabetic wounds faster, decrease skin fibrosis, and block pyroptotic pathways (150). Bioactive glass (BG) may work for wound healing and soft tissue restoration. It controls the Cx43/ROS signaling pathway to suppress EC pyroptosis and improve wound healing (151). BG inhibits caspase-1 activation and perforation, slows ROS generation, and regulates connexin 43 expression, promoting blood vessel development and wound healing (148, 152).
The interaction between inflammasome, pyroptosis, and wound healing is complicated and needs additional study (153). Novel carriers and Chinese herbal extracts are being tested for therapeutic use. Recent studies have shown that the TFNAs, NLRP3 inflammasome, and BG improve healing and reduce problems. Understanding these relationships may help design chronic wound treatments and individualized wound healing techniques (148, 154).
Pyroptosis can be induced with caspase-1, NLPR3, ILβ and IL18. The natural healing process of wounds depends upon NALP3 signaling. Elevated pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, facilitate wound healing. Multiple research studies show that the signaling pathway of the NALP3 inflammasome is of significant importance in the process of skin wound healing (155). Research conducted both in vitro and in vivo has shown that TFNAs improve corneal transparency, hasten the process of wound reepithelialization, and have a beneficial effect on the healing of corneal epithelial wounds (150). TFNAs have advantages not only in promoting the healing of corneal wounds but also in facilitating the healing of skin wounds. The research findings demonstrated that the use of TFNAs expedited the skin wound healing process and mitigated scarring. The first study indicates that nanophase materials with nucleic acid biological characteristics have the potential to expedite wound healing and minimize scarring. These findings suggest that TFNA might be used to facilitate the regeneration of skin tissue (156). Studies have shown that TFNA suppresses the pyroptotic pathway, which in turn lowers levels of inflammatory cytokines and increases the amount of collagen in the skin. Following treatment with TFNA, levels of NLRP3 inflammasome and pro-caspase-1 were found to be decreased. This suggests that the inflammasome was reduced together with the active form of caspase-1, which led to a subsequent decrease in N-terminal GSDMD levels. According to the findings, TFNA has both anti-inflammatory and anti-fibrotic properties, although it does not cause cytotoxicity (152). Pro-caspase-1, caspase-1, NLRP3, GSDMD, and a number of other proteins associated with the pyroptosis and inflammasome signaling pathways were shown to be present in this research. In summary, TFNAs have essential scientific relevance for skin wound healing, and it has been established that they are strongly associated with pyroptosis and inflammasome pathways. This suggests that TFNAs may play a role in the activation of these pathways (Figure 6) (148, 152).
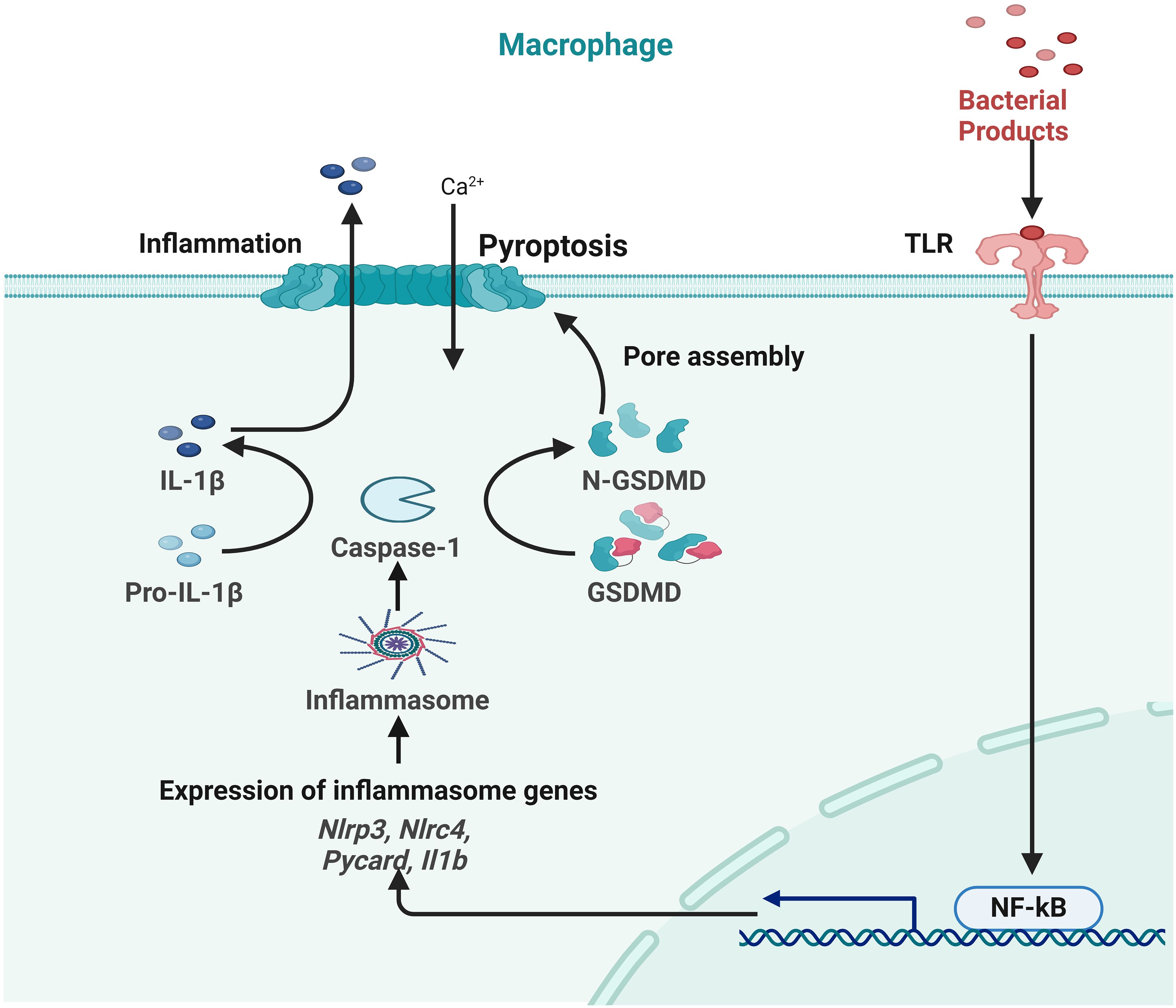
Figure 6 Role of pyroptosis in inflammation during skin wound healing.
Tsvetkov et al. discovered “cuproptosis,” a cellular death process characterized by the transportation of Cu to mitochondria (157). The finding mentioned above has inspired investigations into the control of mitochondrial copper and its possible implications in cancer treatment. Previous studies have shown a connection between the toxicity of elesclomol in cancer cells and many parameters, such as the levels of ferredoxin-1 and the rates of mitochondrial respiration, which are influenced by the availability of Cu (157).
Cuproptosis, a particular type of cell death that is reliant on Cu and occurs inside mitochondria, exhibits distinct characteristics when compared to well-established processes such as ferroptosis, apoptosis, or necroptosis. FDX1, an essential participant in this mechanism, functions as a ligand for elesclomol, facilitating the conversion of Cu3+ ions to Cu2+ ions and subsequent release inside the matrix of mitochondria (158). Research indicates that FDX1, metabolic enzymes, and targets of lipoylation proteins within the PDH complex mediate Cu ionophore toxicity. Posttranslational lipoylation adds lipoic acid to mitochondrial proteins, reducing Cu-induced cell death. FDX1-dependent cancer cells require lipoic acid pathway components for survival, and modifications reduce metabolic enzyme lipoylation, making some cells susceptible to elesclomol-induced cuproptosis (157, 159, 160).
Cuproptosis refers to the phenomenon in which mitochondrial Cu undergoes a process of substitution with proteins, hence playing a crucial role in elucidating the physiological significance of Cu in the human body. Cu is often located inside the matrix of mitochondria and is integrated into the assembling process of cytochrome c oxidase. Copper shortages have been shown to be associated with the occurrence of developmental problems. However, it has been observed that the administration of elesclomol can effectively mitigate these defects (161). The use of Cu metalloallostery, a mechanism that governs several cellular processes, such as proliferation and autophagy, has been under scrutiny due to its possible harmful effects. Cuproptosis is a phenomenon characterized by cellular demise, which is intricately associated with copper biology (162, 163).
According to Tsvetkov’s study findings, cuproptosis might potentially play a role in the development of Wilson’s illness, which is defined by an abnormal buildup of Cu in the body. Increased Cu concentrations in liver cells trigger the activation of Cu metalloallostery, which can remove impaired mitochondria and alleviate cuproptosis (157). The buildup of metals generated by ions affects cellular homeostasis, hence impacting the stability of proteins. Histone H3 can attach to Cu and function as a Cu reductase, therefore indicating the presence of many protein binding sites that may accommodate exchangeable Cu ions (164). Cuproptosis has the potential to induce cellular demise via the process of copper-mediated aggregation (162, 165).
Cuproptosis, controlled cell death using copper buildup and reactive oxygen species, is associated with cancer neurological, and cardiovascular problems. Recent research suggests it may aid wound healing (162, 165). Research shows that wound healing may be accelerated by upregulating cuproptosis-related genes and delayed by suppressing it. Cuproptosis is an essential process in the context of wound healing as it facilitates the elimination of impaired cells, recruits inflammatory cells, and governs the orchestration of new tissue generation. Nevertheless, the specific contributions of the subject in question continue to be a topic of continuous scholarly investigation, whereby the possible advantages and disadvantages are contingent upon the particular circumstances (165).
Necrosis takes place when an illness remains untreated or when harm to tissue reaches an irreversible state. The condition can advance into the deeper layers of tissue, impacting the integrity of bone tissue and possibly resulting in the development of bacteremia and sepsis (7). Skin necrosis may be due to either extrinsic causes or vascular blockage. Necrosis is a pathological condition that is distinguished by cellular or tissue demise, often leading to a discoloration of the skin in shades of purple, blue, or black. Frequently seen necrotizing disorders include necrotizing fasciitis, ecthyma, and meningococcemia (166).
The proliferative stage of wound healing is a complicated process that includes the creation of neovascularization, re-epithelialization, granulation tissue, and immune system modulation. This phase of wound healing occurs after the inflammatory stage of wound repair. Granulation tissue is predominantly activated fibroblasts, as described by Alexis Carrel and John Hunter. These fibroblasts produce new ECM, which helps to contract wounds and forms cellular and structural elements (30, 167).
Neovascularization is necessary for the healing of wounds, the transport of nutrients, the preservation of oxygen balance, the multiplication of cells, and the regeneration of tissue (30). During the early stages of embryonic development, endothelial progenitor cells (EPCs), also known as angioblasts, give rise to the first blood vessels. Angiogenesis is the primary process that leads to the development of new blood vessels during adult tissue healing. This process involves the activation of local microvascular ECs in response to hypoxia and growth factors such as PDGF and VEGF. Proliferation, migration, and the development of capillaries are all processes that ECs go through (168).
ECs and pericytes are involved in the angiogenesis procedure. These cells react to hypoxic injury settings by beginning the cycle of angiogenesis, which includes tearing down the ECM and generating blood vessels (169). Pericytes maintain the integrity of these vessels. Granulation tissue may also be formed owing to the participation of circulating progenitor cells and several subtypes of fibroblasts in the formation process. Understanding these systems is necessary for the development of therapeutic strategies for wound healing (6).
Angiogenesis refers to the physiological process through which new blood vessels are formed, mainly via the involvement of ECs (170). The initiation of the procedure is influenced by a multitude of variables, such as the presence of proteolytic enzymes and growth factors (171). Enzymes such as these facilitate the migration of ECs via clots that are rich in fibrin and fibronectin, hence promoting the repair of injured regions. ECs also exhibit a response to hypoxia and engage in interactions with perivascular cells, therefore emphasizing the complicated structure of their stimulation. This mechanism has a critical role in both physiological and pathological contexts (172). VEGF, FGF, PDGF-B, TGF, and angiopoietins stimulate EC to migrate and proliferate to start angiogenesis (171–173). Some ECs become lead tip cells and extend filopodia toward pro-angiogenic growth factors, while others become trailing stem cells (174).
The Notch signaling pathway is of significant importance in the determination of ECs as either tip or stalk cells. The regulation of this pathway is governed by VEGF, which is synthesized by several cellular populations inside the wound microenvironment. VEGF-A, which belongs to the VEGF family, plays a crucial role in the process of angiogenesis by directing tip cells towards regions of higher concentration gradients and promoting the proliferation of stalk cells in a concentration-dependent manner. The complicated interaction among ECs, growth factors, and signaling pathways exemplifies the sophisticated and orchestrated nature of vascular development (171, 175, 176).
EPCs have garnered significant scientific attention since their first characterization in 1997. Initial investigations have shown that HSCs, as well as EPCs, are involved in the process of blood vessel regeneration. EPCs undergo a sequential three-step mechanism in order to get into ischemic tissues (177). This process involves their mobilization from the bone marrow, subsequent navigation via the circulatory system, and eventual integration into sprouting endothelium (178). Nevertheless, new research has cast uncertainty on the actual function of EPCs in the processes of ischemia-responsive vasculogenesis and blood vessel restoration (179). In mouse models, it has been shown that circulating progenitor cells do not undergo differentiation into ECs at sites of damage or tumor formation (180).
The identification and characterization of circulating progenitor in blood vessels have posed significant challenges, mostly stemming from the complexities associated with their proper isolation from the circulatory system or the surrounding microenvironment (178, 179, 181). The use of single-cell technologies has excellent potential in elucidating the identification and function of cells involved in the process of wound healing, hence facilitating a more accurate comprehension of the mechanisms behind blood vessel regeneration and repair. The acquisition of this information has the potential to enhance the efficacy of wound healing mechanisms and perhaps facilitate the development of specific therapeutic approaches for vascular restoration (6, 181).
Fibroblasts are an assortment of cells located in the dermal layers, demonstrating notable versatility and serving a range of functions (6, 182). Fibroblasts respond to soluble extracellular signals, including growth factors and cytokines, by undergoing activation, leading to cellular proliferation and the modulation of metalloproteinases. During the process of healing a wound, fibroblasts that have reached maturity move toward the granulation tissue, therefore initiating the production of collagen and substituting the temporary fibrin matrix (183, 184). Myofibroblasts are generated, leading to an increase in the accumulation of collagen and the initiation of contraction of the wound. Fibroblasts possess the ability to perceive both the magnitude and direction of mechanical loads, after which they convert this sensory input into appropriate adaptive responses (183, 185). To illustrate fibroblast behavior, consider the intermediary filament vimentin, which inhibits fibroblast growth, promotes the accumulation of collagen, and stimulates keratinocyte mesenchymal reepithelialization and differentiation. It is essential for the study of wound healing procedures and, possibly, for the development of focused treatments in wound care to have a firm grasp on the complex mechanisms governing fibroblast activity (186).
The process of wound healing encompasses wound contraction. This phenomenon leads to a reduction in the surface area of the wound and an improvement in the mechanical strength of the surrounding tissue. The procedure mentioned above converts migratory fibroblasts into myofibroblasts that express α-SMA, leading to the deposition of ECM and facilitating the process of wound healing (187). Myofibroblasts, which exhibit contractile properties similar to smooth muscle cells, contribute to wound healing by depositing ECM components such as collagen type I and III. They achieve wound contraction by connecting to polymerized fibronectin and collagen fibrils (188). The ability to exhibit alternative protein contractility and induce apoptosis upon the restoration of the integrity of tissues has been observed (6, 189). Nevertheless, under pathological circumstances such as hypertrophic scarring, the presence of myofibroblasts might endure, hence playing a role in the excessive formation of scar tissue. The strategic targeting of myofibroblasts has significant potential in the advancement of therapeutic approaches for fibrosis and damage, rendering them a critical focal point for therapeutic interventions. Comprehending the dynamics and control of myofibroblasts is essential in the progression of wound healing procedures and the formulation of tailored medicines to enhance results in healing wounds and fibrosis management (Figure 7) (6, 189).
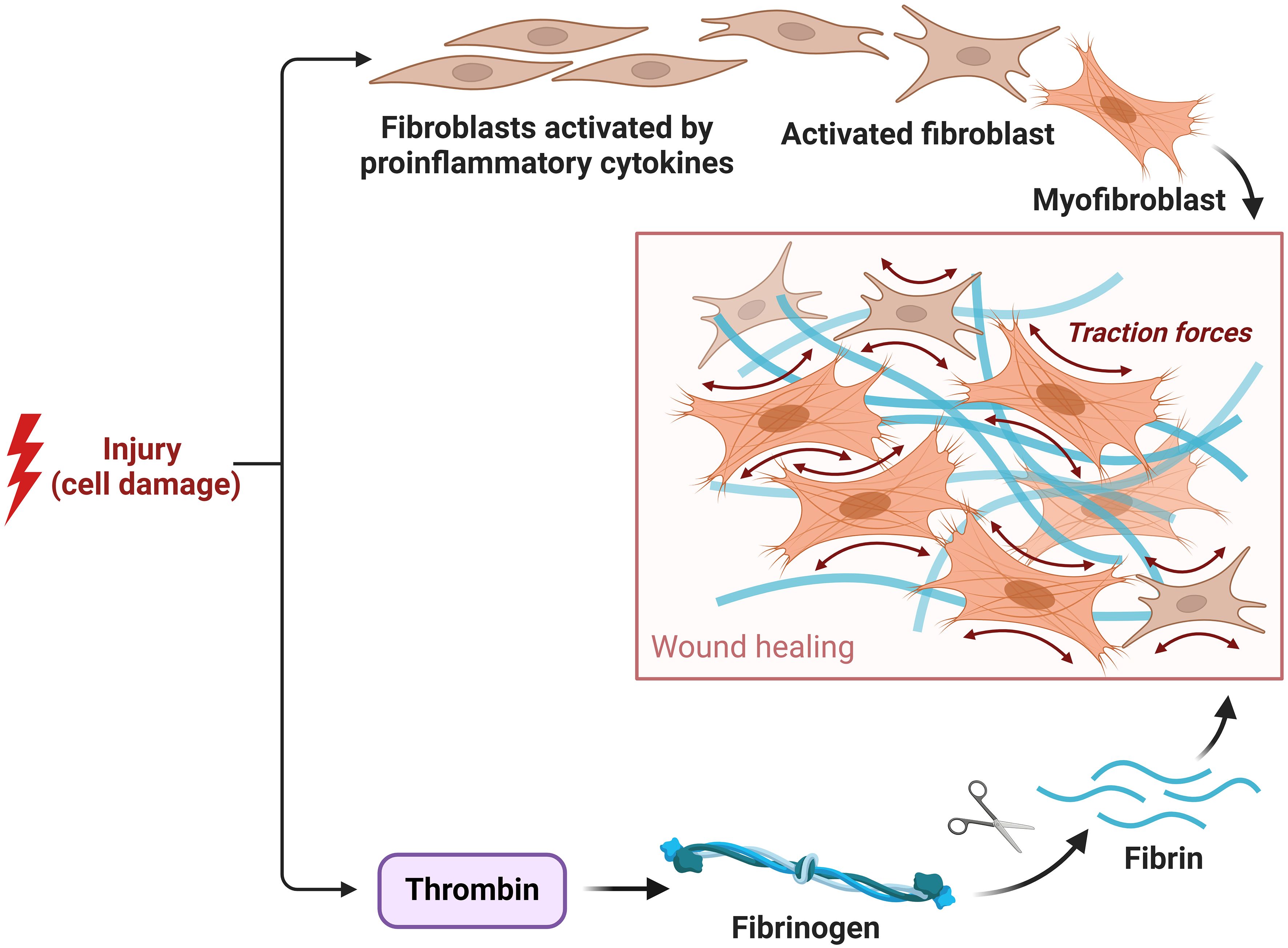
Figure 7 Role of fibroblast and myofibroblast in the proliferative phase.
The epidermis, an integral layer of the skin, serves as a vital protective barrier from external factors and plays a critical role in the process of wound healing. The structure in question is comprised of many distinct layers, including the basal, granular, spinous, and stratum corneum layers. Its main constituents are keratinocytes, which are interconnected by desmosomes. The basal layer is in direct association with the basement membrane, housing many components such as hair follicles, immune cells, sebaceous, and sweat glands. The presence of stem cells inside the epidermis plays a vital role in the processes of repair and maintenance (6, 43, 190). Lineage tracing investigations have been conducted to discern the presence of stem cell populations inside different skin structures, such as the interfollicular epidermis (IFE), hair follicles, sebaceous glands, sweat glands, and melanocytes. The concept known as the EPU explains the processes of regeneration and repair. In this model, slow-cycling stem cells located in the basal layer of the epidermis generate transit-amplifying cells via a process of asymmetric proliferation (191). iPSCs exhibit a phenomenon known as inflammatory memory, which facilitates their ability to mount rapid reactions when encountering subsequent challenges. The turnover of epidermal appendages, such as sebaceous glands, hair follicles, and sweat glands, is a continuous process that involves lineage-restricted stem cells dedicated to the maintenance of their respective cell types. Gaining a comprehensive understanding of the dynamics and characteristics of these cells provides valuable insights into the processes behind wound healing. Such insights have the potential to pave the way for the development of precise wound-healing techniques and regenerative medicines (6, 43, 192–194).
When a person has traumatic damage to a peripheral nerve, the homeostatic function of their skin is disrupted. Collateral reinnervation and nerve regeneration are both necessary steps in the procedure of repairing neurological functionality (195). The peripheral nervous system in adults is capable of regenerating nerve activity via the process of growing back the terminals of myelinated nerve and rejoining the wounded nerve (196). Schwann cells, monocytes, macrophages, and fibroblasts are essential players in the process of regenerating nerves. Schwann cells are distinguished by their exceptional plasticity, which enables them to redifferentiate into progenitor-like cells after damage. This facilitates the rebuilding of axons (39, 195, 197).
The process of Schwann cell differentiation initiates the secretion of monocyte chemoattractant protein-1, PAP-III, IL-1α, and IL-1β, which in turn pull monocytes/macrophages to the location of damage. Macrophages are known to secrete VEGF and HIF, hence facilitating the process of angiogenesis (198). The newly reorganized vasculature functions as a supportive structure for Schwann cell cords, reducing the guidance of axon development. The complex nature of this process guarantees the restoration of nerve function after an injury, and comprehending these processes is of utmost importance in the development of specific therapeutic approaches for traumatic damage to the peripheral nerve (199).
The procedure of wound healing encompasses a prolonged phase of tissue remodeling or maturation, which may last for many months or possibly extend over years. The period, as mentioned above, has a substantial impact on the probability of scar formation and wound recurrence. The remodeling phase encompasses the regression of neovasculature and the remodeling of tissue from granulation into scar tissue (52). The presence of collagen III is first seen in granulation tissue, and over time, it undergoes a progressive replacement by collagen I. During the progression of wound remodeling, the myofibroblasts present in the granulation tissue actively produce matrix metalloproteinases (MMPs) and their inhibitors (TIMPs), which play a crucial role in selectively degrading particular ECM components (52, 200). An aberrant production of TIMPs and MMPs may give rise to an asymmetry in ECM modification, which may, therefore, contribute to the development of chronic wounds (52, 117). Macrophages are of significant importance in the process of wound remodeling as they are responsible for the degradation of excessive ECM and the phagocytosis of debris (201).
During the remodeling stage of wound repair, the blood vessels that are generated during the process of angiogenesis exhibit increased permeability and a deficiency in tight intercellular junctions. This characteristic facilitates the infiltration of immune cells into the wound site. The process of neovessel pruning is essential for the development of stable and well-perfused blood vessels, which involves the programmed cell death of ECs. Vessel pruning may also be influenced by the process of re-epithelialization (30). ECs possess negative feedback systems that regulate their response to VEGF (202). These processes include the stimulation of intracellular Sprouty proteins and Vasohibin. During the advanced phase of wound healing, endothelial cells demonstrate the expression of CXCR3, which subsequently hinders the process of endothelial tube formation. The comprehension and manipulation of these mechanisms have the potential to result in specific therapies, enhanced results in wound healing, and decreased scarring (202).
In response to injuries, the human body undergoes a series of physiological processes that include wound healing and the subsequent creation of scars. Scars are tissue characterized by an effective neo-formation process, which involves the replacement of lost tissue with an ECM that is mainly composed of fibronectin and collagen types I and III. Specific components of the skin, including hair follicles, subepidermal appendages, and glands, have little or no regenerative capacity after significant damage (203). The scar tissue matrix, which is denoted by granulation tissue, is distinguished by a considerable concentration of fibroblasts, capillaries, macrophages, granulocytes, and collagen fibers (204). During the first stage of scar formation, known as the primordial scar tissue phase, the process of angiogenesis is still ongoing but not fully developed. The predominant cell type during this phase is fibroblasts (205). The process of scar development concludes during the remodeling phase, commencing on day 21 and continuing for one year subsequent to the occurrence of an injury. During this particular stage, the ECM constituents experience continuous modifications, characterized by the replacement of collagen III with type I collagen and the involvement of myofibroblasts in the process of wound contraction. Excessive use of scarring results in a disruption of the equilibrium between the processes of biosynthesis and degradation, which subsequently gives rise to a prolonged inflammatory phase, an extended proliferative phase, and a diminished remodeling process (204–206). Abundant microvessels are present in hypertrophic scars, and aberrant scarring is also characterized by alterations in the ECM and epithelial tissue (7).
The predominant kind of stem cells used in medical applications are adult stem cells, which are obtained from several sources, such as bone marrow and adipose tissue (207). According to previous research, MSCs exhibit anti-inflammatory and immunomodulatory characteristics during the inflammation phase while also promoting the activation of fibroblasts, keratinocytes, and ECs during the proliferative phase. This cellular response ultimately leads to an expedited healing of wounds (207). Research has shown that the use of MSC treatment has the potential to decrease the duration of wound healing, minimize wound contraction, enhance the formation of new blood vessels (angiogenesis), and expedite the process of epithelialization (208, 209). The administration of stem cell treatment may occur by topical or systemic means. Topically, MSCs can be applied in the kind of spray, aiding in the retention of these cells throughout the wound (210). In order to enhance the retention of stem cells inside a wound, it has become common practice to provide these cells with suitable support or scaffold material, such as collagen or skin substitutes. This phenomenon contributes to the preservation of cellular functionality and promotes the movement of cells inside the wound site (35, 211).
Traditional methods of wound care include a range of interventions, including surgical interventions, non-surgical therapies, and the use of pharmaceutical agents to facilitate the healing process of skin wounds. Surgical interventions include debridement, skin grafts, and skin flaps, while non-surgical modalities involve the use of topical medications, wound dressings, and skin replacements (212). Surgical debridement is often regarded as the preferred method for wound treatment because of its high efficacy and expeditiousness. However, it is essential to acknowledge that this approach is not without its inherent hazards, including the administration of general anesthesia and potential tissue damage (213).
Skin grafts are often used in surgical procedures to control wounds, taking into consideration the specific characteristics and causes of the wound. The two main categories of skin grafts are split-thickness skin grafts and full-thickness skin grafts. Split-thickness grafts are appropriate for treating tiny lesions that only include damage to the epidermis. Full-thickness grafts are a valuable treatment option for extensive and deep wounds due to their ability to facilitate healing in both the epidermal and dermal layers, leading to the formation of healed tissue without visible scarring (7). Nevertheless, the use of allografts is restricted due to the scarcity of available donor skin and the need for a wound bed with adequate vascularization. Skin grafts may be categorized as autografts, allografts, and xenografts, depending on the source of the donor. Autografts are often regarded as the preferred method for skin restoration since they include the use of healthy skin obtained from the same individual to restore the integrity of the skin effectively. Allografts have the advantage of readily accessible donor skin and the capacity to be stored for extended periods for potential future use. In contrast, xenografts refer to skin grafts acquired from distinct species and then implanted into human recipients (214). Nevertheless, both allografts and xenografts are subject to some restrictions, including immunological rejection, the potential for disease transmission, the occurrence of painful healing, and the formation of scars (7, 215).
Traditional non-surgical methods for treating wounds include a range of interventions, such as the use of dressings for wounds, topical medications, scaffolds/hydrogels-based skin transplants, and skin replacements. The objective of these treatments is to achieve equilibrium in moisture levels, address issues related to inflammation and infection, and regulate the processes of contraction and re-epithelialization (216). The skin is most often targeted for medication delivery via topical formulations, which include gels, emulsions, pastes, creams, foams, lotions, and sprays. Topical antibiotics, such as silver sulfadiazine and neomycin, have shown efficacy in combating bacterial infections. Nevertheless, it is advisable to halt their use in order to prevent the development of hypersensitivity and allergic responses (217). Topical products have been subjected to experimentation in order to assess the efficacy of growth factors in promoting wound healing. Growth factors have significant potential in treating skin wound healing. They stimulate cell proliferation, promoting the formation of new tissue and wound closure. They also enhance angiogenesis, blood supply, and nutrient and oxygen delivery to the healing tissue (218). Growth factors can modulate inflammation, reduce inflammation, and promote tissue repair (219). They regulate the synthesis and remodeling of the extracellular matrix, enhancing tissue strength and integrity (220). They facilitate the migration and differentiation of cells, promoting the differentiation of specialized cell types for tissue repair (221). Growth factors may also reduce scar formation and improve the aesthetic outcome of wound healing (222). Some growth factors have antimicrobial properties, reducing the risk of wound infection and promoting a more favorable healing environment (223). Advances in delivery systems, like hydrogels, nanoparticles, and scaffolds, allow for controlled delivery of growth factors at the wound site, optimizing their therapeutic effects while minimizing adverse effects (224). Overall, growth factors hold promise as a therapeutic approach to enhance skin wound healing by promoting faster and more efficient tissue repair processes (225).
However, the restricted bioavailability of growth factors, caused by their quick elimination from the wound site, imposes limitations on their practical use. Nanoparticle-encapsulated growth factors are now being used in topical applications as novel methods to enhance collagen production and angiogenesis (226).
Cytokeratins, specifically KRT16 and KRT17, play a crucial role in skin wound healing. They are upregulated during the process of wound healing, providing structural support to keratinocytes, the predominant cell type in the epidermis. They also regulate cell proliferation and differentiation, with KRT16 and KRT17 promoting keratinocyte proliferation, essential for the formation of new epithelial tissue (227). Cytokeratins may also modulate the inflammatory response during wound healing, interacting with immune cells and signaling molecules involved in inflammation. Changes in cytokeratin expression patterns have been proposed as biomarkers for monitoring wound healing progression and assessing therapeutic interventions’ effectiveness (228). Dysregulation of cytokeratin expression or function has been linked to the pathogenesis of chronic wounds, where impaired re-epithelialization contributes to delayed healing. Understanding the roles of cytokeratins in wound healing may offer insights into novel therapeutic strategies for chronic wound management (229).
Natural biomaterials, particularly keratins, play pivotal roles in wound healing, offering promising therapeutic avenues in both healthy individuals and those with conditions such as diabetes. Keratin-based biomaterials possess inherent biocompatibility and bioactivity, serving as effective scaffolds for cell attachment, proliferation, and differentiation, thereby promoting tissue regeneration. They also exhibit modulatory effects on the inflammatory response, fostering a shift towards a pro-healing environment, which is particularly relevant in chronic wounds observed in diabetic patients (229). Keratin-based dressings help maintain moisture balance in the wound environment, preventing excessive drying or exudate accumulation and promoting cell migration, proliferation, and ECM deposition (230). They also promote angiogenesis, a hallmark of chronic wounds, by providing cues for endothelial cell proliferation and migration, enhancing blood vessel formation and tissue perfusion in the wound bed (218). Keratin-based biomaterials have been investigated for their potential to reduce hypertrophic scars or keloids, which can impair function and aesthetics. They can promote more organized ECM deposition and reduce excessive fibrosis during the healing process. By being biodegradable and biocompatible, keratin-based biomaterials are suitable for various wound healing applications, including in diabetic patients with compromised healing capacity (Table 2) (238).
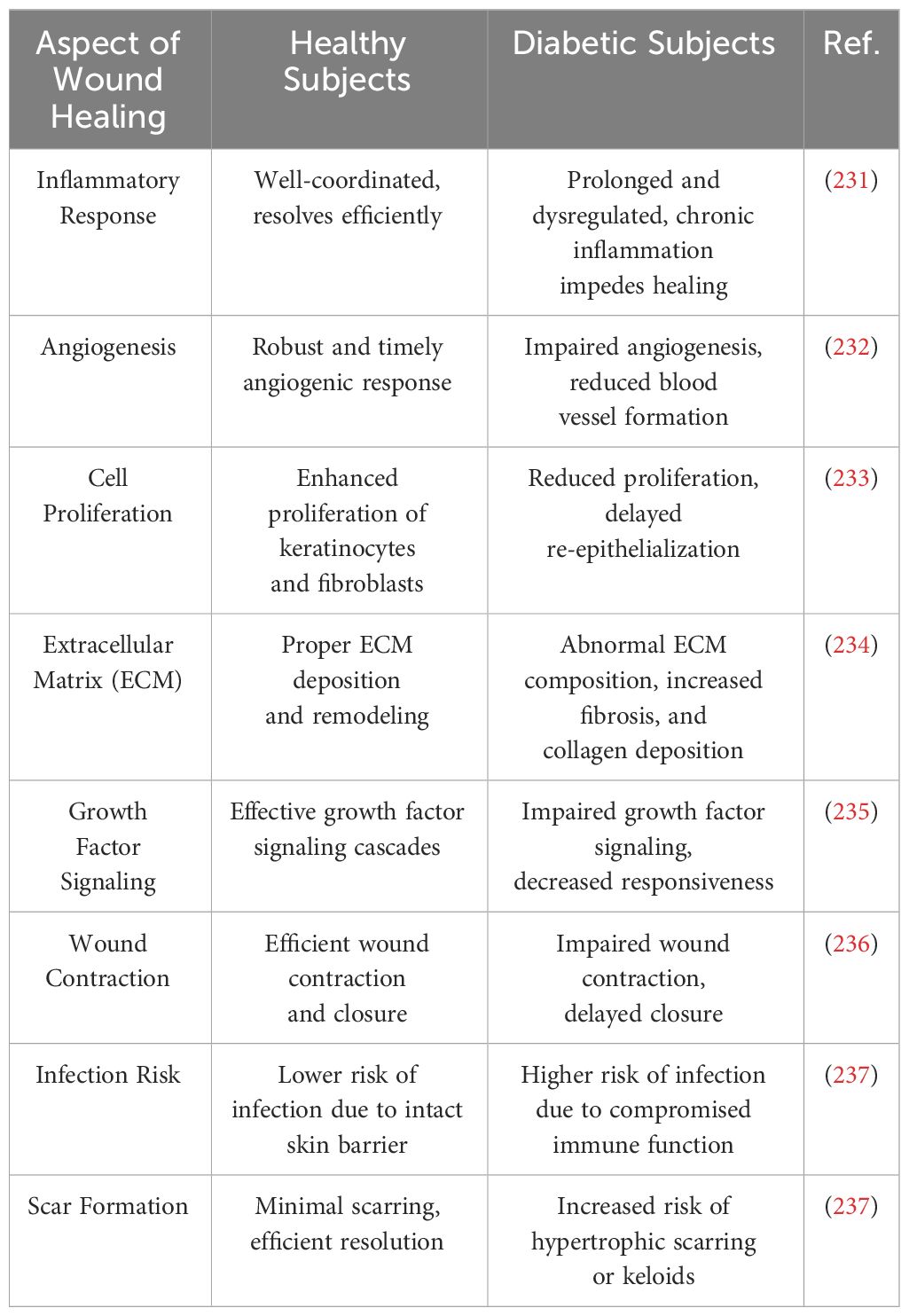
Table 2 Summary of the differences in wound healing in healthy and diabetic subjects.
The acronym TIME, which stands for Tissue, Infection, Moisture, and Epithelial, serves as a complete framework for wound care, including several elements that impede the healing process (215, 239). The framework has four distinct components, including tissue assessment, infection evaluation, moisture management, and epithelium edge development. The primary objective of TIME is to analyze devitalized or non-self-material in the wound bed, evaluate the origin and treatment of infection, manage wound exudate, and check the advancement of edges as well as surrounding skin status (240). The comprehension of the relationship between TIME and the molecular biology of injuries has resulted in significant progress in the field of chronic wound care interventions and technology (7).
Negative pressure wound treatment (NPWT) is a wound healing technique that leverages the application of differential vacuum or suction to augment the healing process. The use of this treatment modality is often seen in the management of both acute and chronic wounds, including pressure ulcers, diabetic foot ulcers (DFUs), postoperative incisions, lower extremity wounds, traumatic wounds, infected wounds, burns, necrotizing fasciitis, infected sternal injuries, and post-skin grafting interventions. Significant advancements have been seen in individuals suffering from vascular lower leg ulcers and DFUs who possess adequate blood circulation (241, 242). NPWT devices are comprised of three essential components, as described in the literature (243). These components include a porous substance that fills the wound site, a drainage port that is coupled to a vacuum pump under regulated conditions, and an adhesive film dressing that effectively seals the wound. The porous substance often used is polyurethane foam, while the adhesive dressing is composed of polyurethane owing to its occlusive qualities (241, 244, 245).
NPWT facilitates the process of wound healing by using four main mechanisms: microdeformation, macrodeformation, elimination of wound fluid and decrease of edema, and modification of the wound microenvironment (242). Notwithstanding its efficacy, the use of this intervention has inherent dangers and possible consequences. There are three primary problems associated with this condition, including infection, bleeding, and the retention of foam dressing (241, 246). Bleeding occurs as a result of physical trauma to the underlying tissues, a condition that may be further exacerbated by factors such as necrotic tissue, infection, or coagulopathy. The use of NPWT in near proximity to accessible blood vessels, nerves, organs, or anastomotic sites is discouraged due to the heightened likelihood of fistulae development (245, 246). It is recommended to provide treatment for infections prior to the administration of NPWT. Additionally, it is essential to note that the non-dissolving nature of the dressing materials used in NPWT may potentially lead to complications such as bleeding or infection caused by foam retention. Additional concerns include the presence of discomfort during dressing changes and the occurrence of patient allergies to the adhesive bandage or foam substance (241, 243, 245, 247).
Enterocutaneous fistulas, which were formerly regarded as contraindications for non-operative percutaneous wound closure, are currently being managed with favorable outcomes. The device achieved fistula closure in a majority of the documented patients, namely those with low-output fistulas, comprising almost two-thirds of the total sample size of forty (248). Trauma patients, as well as those with abdominal compartment syndrome, are managed by using an open abdomen technique with the use of a negative pressure wound therapy device. This approach serves as a temporary measure by providing coverage, eliminating intraabdominal contaminants and exudates, and reducing visceral edema (249).
The use of NPWD closure has yielded substantial advancements in the management of severe orthopedic injuries. In the past, significant injuries to the extremities were managed by the use of extensive debridement, which necessitated the use of free flaps for rapid wound covering. The NPWD generates a hermetically sealed and safeguarded environment that effectively eliminates swelling and accumulation of blood outside blood vessels, hence enhancing blood flow and optimizing the preservation of the area around the wound. Serial debridements are conducted, and ultimate rebuilding is carried out in a stable wound in a planned manner (250). The contraindication of exposed tendons, joints, or bones has been reconsidered due to the formation of granulation tissue over these structures, which enables the possibility of skin grafting if deemed essential (251).
Historically, the two most critical oxygen-based treatments for wound care were known as hyperbaric oxygen therapy and topical hyperbaric oxygen therapy. However, recent developments in CDOT and Transdermal Oxygen Therapy have led to the introduction of a novel category of oxygen-based wound healing devices. These devices provide persistent treatment of wounds with oxygenation and are part of the oxygen-based wound care industry. Although these techniques promote wound healing in a manner that is analogous to one another, there are significant technical and medical distinctions between them (252).
HBOT is a medical intervention in which a patient inhales pure oxygen at a pressure more significant than that of the surrounding atmosphere. This treatment has been shown to enhance the process of neovascularization, promote the production of ECM, and reduce inflammation (252). The use of this treatment has been shown to be advantageous in the management of late radiation tissue damage, acute wounds, chronic ulcers, and burns (241, 253). Neovascularization is initiated by an upregulation of ROS and RNS at the local level, leading to the activation of growth factors such as TGF-β, VEGF, and angiopoietin 2 (252). HBOT has been shown to induce the activation and specialization of circulating SPCs originating from the bone marrow, leading to the formation of blood vessels through the process of vasculogenesis. HBOT has been shown to enhance the production of fibroblast growth factors, which subsequently promotes the migration and proliferation of fibroblasts. The augmentation of oxygen levels serves as a stimulus for the proliferation of fibroblasts, leading to an increased production of collagen and subsequent enhancement of tissue tensile strength. In conclusion, HBOT has been shown to effectively decrease inflammation by reducing edema, inhibiting pro-inflammatory cytokines, promoting macrophage chemotaxis, enhancing leukocyte bactericide activity, and blocking neutrophil β2 integrin (251, 253, 254).
The progression of HBOT into THOT involves the use of a chamber that is placed around a wounded region to avoid leaks and to be filled with oxygen at high flow rates to provide a rich source of oxygen at the surface of the wound. When compared with HBOT, THOT is associated with a reduced risk of adverse effects, may treat a wider variety of therapeutic patients, and can be used in a wider variety of locations at a cheaper cost. However, in comparison to HBOT, it does not have the identical oxygen potential since collagen formation and cross-linking are lower, and the pace at which wounds are closed is slower. The effects of THOT are comparable to those of HBOT, and it was shown to speed healing in wounds that are persistent (255).
CDOT devices offer a convenient and portable solution for supplying a continuous stream of oxygen to wound sites. This technology serves as an alternative to THOT and HBOT methods. The delivery of oxygen at atmospheric pressure and slower flow rates to the wound bed necessitates the use of a wet dressing to facilitate diffusion. The term “THOT,” although characterized by high flow rates, is associated with intermittent treatment and necessitates a humid atmosphere (256). Although research is scarce in this area, several studies have demonstrated that THOT may have potential benefits in the healing of DFUs, ulcers associated with sickle cell disease, as well as in the treatment of stubborn and painful wounds when used as an adjuvant (255, 257).
Healing wounds is a complicated process at the moment because various kinds of wounds have distinct molecular and cellular processes that must be addressed. Our knowledge of wound assessment and treatment methods has been significantly expanded as a result of developments in technology, which has resulted in a transition away from the use of traditional dry dressings and toward the use of modern moist dressings, growth factor therapies, responsive dressings, tissue-engineered skin, nanotherapeutics, gene therapy, and stem cell therapy. The road to individualized wound healing has been paved with the introduction of cutting-edge technologies such as 3D bioprinting, platelet-rich plasma treatment, and ECM-based techniques. However, the treatment of chronic wounds is still a problem, which calls for the advancement of unique and imaginative treatment methods that take into account their efficacy, the benefits-risks equilibrium, and the affordability and efficacy of the options (258).
The process of wound healing may be complicated for doctors to manage. Still, nanotechnology has swiftly generated nanomaterials that are employed in medical, pharmacy, chemical manufacturing, and even in the military. Because of their one-of-a-kind physicochemical features, these materials produce effects that are localized to their surfaces and have a limited scale. The rising need for nanomaterial coverings such as nanofibers, hydrogels, and films has sped up the advancement of these materials, which has led to their increased use in a wide variety of disciplines (259). Advancements in nanotechnology and biomedicine have facilitated the use of nanomaterials in the healing of wounds. These materials have shown their efficacy in several aspects of the wound healing process, including hemostasis, antibacterial activity, inflammatory modulation, and promotion of cell proliferation. The augmentation of these characteristics may be accomplished by using certain technologies, such as the integration of nanoparticles with sodium alginate and gelatin, to produce a composite hydrogel (259). The antibacterial effectiveness may be enhanced by incorporating antibiotics into nanomaterials (260). The use of bionic nanofiber scaffolds has been shown to effectively mimic the ECM of the skin, hence facilitating the process of adhesion of cells and proliferation (261).
Nanoparticles are of significant importance in the context of wound healing applications, as they include inherent characteristics that expedite the process of wound healing. Additionally, researchers have been actively involved in the development of bioengineered drug delivery systems, which aim to provide sustained and regulated release of therapeutic molecules for enhanced wound healing outcomes. Metal and metal oxide nanoparticles, polymer nanomaterials, and several other nanotherapeutics have been extensively applied to the treatment of chronic wounds owing to their inherent benefits. Metallic nanoparticles, including gold, silver, zinc, and zinc oxide, have antibacterial characteristics that promote the process of wound healing (262). In addition to the substances mentioned above, cerium, nitric oxide, bioactive glass, and carbon-based nanoparticles are also known to possess inherent bioactivity.
Nevertheless, it is essential to consider the potential toxicity associated with metal-based nanotherapeutics prior to their in vivo use. The reduction of hazards in metal nanotherapeutics may be achieved by optimizing the size and characteristics (263). Polymeric nanostructures are now being used in the field of healing wounds and regeneration of the skin. The inclusion of antibiotics and growth factors inside polymeric nanoparticles has the potential to mitigate wound infection and expedite the process of wound healing (264). Table 3 provides a summary of a complete list of nanotherapeutic-based techniques for the management of acute and chronic wounds. These strategies make use of inorganic as well as organic nanomaterials.
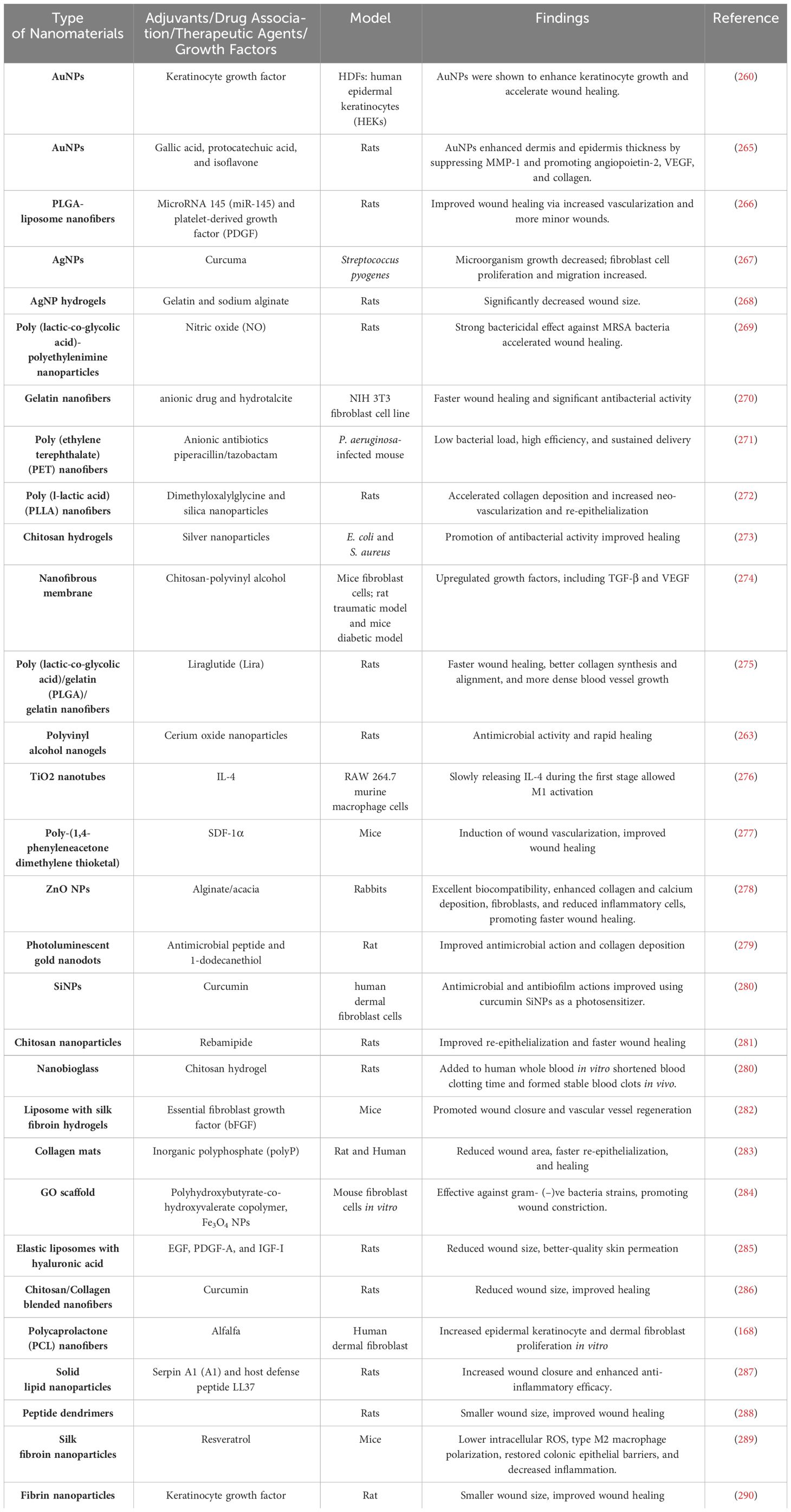
Table 3 Nanotherapeutics used for the management of wound healing.
The use of various nanomaterials in the regulation of the inflammatory state shows promise in the treatment of chronic wounds, making nanotherapeutics a potentially effective method. Burn wounds exhibit an upregulation of inflammatory mediators such as TNF-α, IL-6, and IL-1β. Conversely, diabetic wounds have a considerable expression of IL-18. Nevertheless, previous studies have shown that the application of silver nanoparticles and polymeric nanofibers resulted in a decrease in inflammatory markers, but it did not exhibit significant effects on IL-18 levels. This suggests that these interventions may have limited efficacy in reducing inflammation in the context of diabetic wounds (260, 262, 291). The identification of the inflammatory phase presents a challenge because of its overlap with the proliferative stages. Hence, there is an urgent requirement to produce nanomaterials that possess remarkable anti-inflammatory characteristics in order to facilitate the immunoregulation of many forms of chronic wounds. A plausible approach might include the use of immunomodulatory nanomaterials to identify distinctive indicators that demarcate the shift from the inflammatory phase to the proliferative phase. Nanotherapeutic-based techniques provide a potential avenue for the clinical management of chronic wounds. These approaches demonstrate notable antibacterial properties, effectively mitigate bacterial medication resistance, diminish the inflammatory phase, and expedite the wound healing process.
Nevertheless, a detailed summary of their processes has yet to be undertaken. This study critically examines the probable mechanisms and recent advancements in the use of nanoparticles for wound healing. It specifically focuses on the potential toxicity associated with these materials and identifies the present limits in clinical use and fundamental research (258).
Cell therapy encompasses a range of methodologies that use viable cells for therapeutic objectives, with the intention of rectifying, substituting, or reinstating the physiological functionality of a compromised tissue or organ. Stem cells are a kind of cells that have not yet undergone differentiation and can self-renew and develop into precursor or progenitor cells of one or several distinct cell lineages (292). The application of stem cells from adults is prevalent in the field of regenerative medicine. This is primarily because of their accessibility via in vitro growing techniques and the absence of ethical dilemmas (293). Bone marrow is well recognized as a prevalent source of adult stem cells that play a crucial role in the regeneration or repair of several tissues, such as the cartilage, bone, tendons, heart, and skin (294). Cell-based treatment for prolonged wound repair involves the use of many processes, including the interaction and impact of growth factors, the management of inflammatory procedures, and the activation of immunological processes to expedite re-epithelialization and vascularization (295). In the past few years, there have been many clinical and preclinical investigations that have shown significant effects on the improvement of wound healing quality by the use of stem cells (296). The beneficial effect of stem cell-based wound healing is primarily ascribed to its capacity to release pro-regenerative cytokines as well as growth factors, which facilitate skin regeneration in the management of chronic wounds (294, 297). Autologous stem cells possess remarkable differentiation capabilities, promote the formation of new blood vessels, and are generally well-tolerated by patients, exhibiting minimal adverse responses. The primary therapeutic objective of stem cell-based treatment is to specifically address the enhancement of wound healing quality in the context of wound care (298).
The primary sources of stem cells used for wound healing and skin regeneration are adult MSCs, ESCs, and iPSCs, which have been more recently investigated. The use of ESCs in wound healing has been limited owing to ethical problems that are linked to their use (299). MSCs play a crucial role in all four stages of wound healing, including facilitating the migration of cells to the site of injury, supporting the formation of new blood vessels (angiogenesis), releasing growth factors and cytokines, and aiding in the process of re-epithelialization. In the first human trial, researchers used MSCs generated from bone marrow to treat severe skin burn injuries, which were afterward followed by skin grafting. The study documented notable enhancements in neo-vascularization and pain alleviation (300). According to a research investigation conducted on burn injuries, the application of BMSCs into the wound site resulted in a reduction in wound contraction, modulation of EC activity, and an enhancement of angiogenesis (301). In a previous investigation, a comparable research endeavor used a biodegradable collagen membrane known as Coladerm, together with autologous bone marrow-derived MSCs and skin fibroblasts, to address the issue of chronic non-healing wounds, namely diabetic ulcers. The findings of this study revealed a notable reduction in wound size and an improvement in vascularization after a combined treatment period of 29 days (302).
According to recent research, it has been shown that exosomes formed from stem cells, which include growth factors and cytokines, can expedite the process of wound healing by several mechanisms such as facilitating cell proliferation, migration, re-epithelialization, angiogenesis, and activating signaling pathways (303). Adipose-derived MSCs are often used in the context of wound healing owing to their ease of access, limited invasiveness, and absence of ethical constraints (304). Umbilical cord mesenchymal stem cells (UC-MSC) have shown encouraging therapeutic outcomes in the treatment of chronic wounds in individuals with diabetes, exhibiting increased cellular proliferation and collagen deposition (305, 306). The use of MSCs in conjunction with natural substances has shown augmented therapeutic attributes in third-degree burn injuries, exhibiting heightened levels of angiogenesis, wound closure, and re-epithelialization as compared to control groups. The transplantation of human Wharton’s jelly stem cells (HWJSC) with acellular dermal matrix has been shown to result in improved healing of burn wounds, according to previous studies (307, 308).
Conversely, Cell reprogramming technology encompasses the process of transforming mature somatic cells into iPSCs. This technique has considerable promise for many applications within the pharmaceutical sector, clinical settings, and laboratory environments. iPSCs have comparable attributes to ESCs, but they are not exempt from ethical considerations. Autologous cells have the potential for expansion and use, hence circumventing problems associated with immune rejection (302). The use of iPSCs in producing human autologous cells for progressive persistent wound therapy and degenerative skin illnesses has been extensively explored via in vitro and in vivo research conducted on mice models, revealing significant promise (309). iPSCs may be produced using many methodologies, one of which involves the transduction of somatic cells with a mixture of reprogramming factors. Nevertheless, the process of reprogramming adult somatic cells and promoting differentiation in the target cell line has proven to be challenging (310, 311). The utilization of iPSC methodologies can generate a substantial quantity of human autologous cells, rendering them suitable for implementation in genome editing methodologies. Autologous iPSC has the potential to serve as a viable and enduring therapeutic option for addressing chronic injuries that arise from genetic predisposition (303). Nevertheless, there is a need for more comprehension of these cellular classifications to ensure the safeguarding of patient well-being (298).
In recent years, there has been a significant focus on the enhancement of cutaneous wound healing. This has led researchers to investigate the fundamental processes involved and devise novel treatment strategies to improve the overall quality of wound repair. The use of nanotherapeutic-based methodologies has garnered significant interest in the medical community due to their potential efficacy in the treatment of many forms of chronic wounds, owing to their advantageous characteristics. Nevertheless, nanotherapeutics have several obstacles in terms of biological safety, the danger of transdermal toxicities, the variety of persistent wounds, and financial limitations (312, 313). In order to tackle these issues, scholars are now investigating many tactics, including the modification of the physical and chemical characteristics of nanomaterials, the use of stabilizers, and the enhancement of production procedures and nanoformulations. Moreover, a comprehensive comprehension of the metabolic and functional characteristics of animals and humans is essential in the advancement of tailored nanomaterials designed for various wound classifications (314–317). The financial implications associated with nanoformulations provide a significant constraint on the widespread use of nanomaterials in healthcare contexts. The strategies used include dose reduction, incorporation of cost-effective adjuvants, and utilization of microneedles and layered self-assembly techniques to provide regulated and prolonged drug delivery. The decrease in manufacturing costs may be achieved by the optimization of fabrication methods and the development of nanoformulations. The use of nanomaterials in diverse therapeutic formulations and application techniques has been seen, although the underlying processes by which they facilitate wound healing remain inadequately investigated. The TGF-β1/SMAD signaling pathway is well recognized as one of the predominant signaling channels involved in the proliferation phase of wound healing. Additionally, macrophage polarization has been identified as another critical signaling pathway during the inflammatory phase of wound healing (318). There is a need for modeling approaches that are both stimulated and controlled in order to accurately depict the underlying mechanics of wound healing across various kinds of wounds (258).
Stem cell-based treatments have emerged as a prevalent and productive approach to facilitating wound healing. Several tactics, including the investigation of novel stem cell sources, use of stem cell-derived exosomes, and genetic alterations of stem cells, achieve this. Nevertheless, there are still some problems associated with stem cell sources, including genetic instability, immunogenicity, infection risk, carcinogenesis risk, and high processing costs. Autologous iPSC, which are generated from non-viral vectors, are being employed as a strategy to address the challenges posed by immunogenicity and cancer concerns (313, 319). In summary, the potential for enhancing the therapeutic applicability of stem cell-derived stem cells for wound healing may be realized via future developments in GMP-compliant scaling-up technologies (320).
The maintenance of physiological homeostasis relies heavily on the presence of healthy skin. However, there are several cases when inadequate healing occurs, requiring further intervention. Despite the considerable amount of research conducted and the increased understanding of wound healing procedures, the management of difficult-to-heal chronic wounds and more extensive wounds continues to pose a significant obstacle in the fields of skin regeneration and wound care. Wound healing is a complex process that involves several cellular and molecular systems, hence necessitating the use of multiple treatments rather than relying on a single approach. In order to address these constraints, researchers have investigated a range of novel and developing therapeutic approaches, including nanotherapeutics and stem cell research. These treatments may be used alone or in combination to expedite the process of wound healing, hence offering potential solutions and prospects for managing complicated wounds that are difficult to heal in patients. The challenges pertaining to these unique methodologies and the likely outlook for the future have been deliberated over in order to augment wound care and its administration. In general, the progress made in many technological fields and the discovery of novel and inventive methodologies have significant prospects for enhancing the efficacy of wound healing.
AM: Writing – original draft, Software, Methodology, Conceptualization, Writing – review & editing, Visualization, Investigation, Formal Analysis. CS: Resources, Project administration, Writing – review & editing, Visualization, Methodology, Investigation, Formal Analysis. PG: Software, Writing – review & editing, Project administration, Methodology, Investigation. SW: Writing – original draft, Visualization, Validation, Supervision, Funding acquisition, Formal Analysis, Writing – review & editing, Investigation. JX: Writing – original draft, Writing – review & editing, Supervision, Investigation, Funding acquisition, Formal Analysis, Conceptualization.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This review was supported by the Zhejiang Provincial Natural Science Foundation of China (LQ21H090001), the National Natural Science Foundation of China (Grants 82172428 and 81972150), the Post-Doctoral Research Start-up Fund of Lishui People’s Hospital, Zhejiang, China (2023bsh001) and the Public Welfare Technology Research Funding Project of Zhejiang (LTGY24H100002 and LTGY23H160034), and Key Research and Development Project of Lishui (2022ZDYF23).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Díaz-García D, Filipová A, Garza-Veloz I, Martinez-Fierro ML. A beginner’s introduction to skin stem cells and wound healing. Int J Mol Sci. (2021) 22:11030. doi: 10.3390/ijms222011030
2. Aragona M, Dekoninck S, Rulands S, Lenglez S, Mascré G, Simons BD, et al. Defining stem cell dynamics and migration during wound healing in mouse skin epidermis. Nat Commun. (2017) 8:14684. doi: 10.1038/ncomms14684
3. Chou WC, Takeo M, Rabbani P, Hu H, Lee W, Chung YR, et al. Direct migration of follicular melanocyte stem cells to the epidermis after wounding or UVB irradiation is dependent on Mc1r signaling. Nat Med. (2013) 19:924–9. doi: 10.1038/nm.3194
4. Leclère FM. The use of integra® Dermal regeneration template versus flaps for reconstruction of full-thickness scalp defects involving the calvaria: A cost–benefit analysis. Aesthetic Plast Surg. (2017) 41:472–3. doi: 10.1007/s00266-016-0765-z
5. Boyce ST, Lalley AL. Tissue engineering of skin and regenerative medicine for wound care. Burn Trauma. (2018) 6:4. doi: 10.1186/s41038-017-0103-y
6. Rodrigues M, Kosaric N, Bonham CA, Gurtner GC. Wound healing: A cellular perspective. Physiol Rev. (2019) 99:665–706. doi: 10.1152/physrev.00067.2017
7. Tottoli EM, Dorati R, Genta I, Chiesa E, Pisani S, Conti B. Skin wound healing process and new emerging technologies for skin wound care and regeneration. Pharmaceutics. (2020) 12:1–30. doi: 10.3390/pharmaceutics12080735
8. Nussbaum SR, Carter MJ, Fife CE, DaVanzo J, Haught R, Nusgart M, et al. An economic evaluation of the impact, cost, and medicare policy implications of chronic nonhealing wounds. Value Heal. (2018) 21:27–32. doi: 10.1016/j.jval.2017.07.007
9. Dreifke MB, Jayasuriya AA, Jayasuriya AC. Current wound healing procedures and potential care. Mater Sci Eng C. (2015) 48:651–62. doi: 10.1016/j.msec.2014.12.068
10. Armstrong DG, Swerdlow MA, Armstrong AA, Conte MS, Padula WV, Bus SA. Five year mortality and direct costs of care for people with diabetic foot complications are comparable to cancer. J Foot Ankle Res. (2020) 13:16. doi: 10.1186/s13047-020-00383-2
11. Sorg H, Tilkorn DJ, Hager S, Hauser J, Mirastschijski U. Skin wound healing: an update on the current knowledge and concepts. Eur Surg Res. (2017) 58:81–94. doi: 10.1159/000454919
12. Holgate ST. Innate and adaptive immune responses in asthma. Nat Med. (2012) 18:673–83. doi: 10.1038/nm.2731
13. Sinno H, Prakash S. Complements and the wound healing cascade: an updated review. Plast Surg Int. (2013) 2013:1–7. doi: 10.1155/2013/146764
14. Korting HC, Schöllmann C, White RJ. Management of minor acute cutaneous wounds: Importance of wound healing in a moist environment. J Eur Acad Dermatol Venereol. (2011) 25:130–7. doi: 10.1111/j.1468-3083.2010.03775.x
15. Margolis DJ, Hoffstad O, Nafash J, Leonard CE, Freeman CP, Hennessy S, et al. Location, location, location: Geographic clustering of lower-extremity amputation among medicare beneficiaries with diabetes. Diabetes Care. (2011) 34:2363–7. doi: 10.2337/dc11-0807
16. Okur ME, Karantas ID, Şenyiğit Z, Üstündağ Okur N, Siafaka PI. Recent trends on wound management: New therapeutic choices based on polymeric carriers. Asian J Pharm Sci. (2020) 15:661–84. doi: 10.1016/j.ajps.2019.11.008
17. Han G, Ceilley R. Chronic wound healing: A review of current management and treatments. Adv Ther. (2017) 34:599–610. doi: 10.1007/s12325-017-0478-y
18. Hosseini M, Shafiee A. Engineering bioactive scaffolds for skin regeneration. Small. (2021) 17:2101384. doi: 10.1002/smll.202101384
19. Monavarian M, Kader S, Moeinzadeh S, Jabbari E. Regenerative scar-free skin wound healing. Tissue Eng - Part B Rev. (2019) 25:294–311. doi: 10.1089/ten.teb.2018.0350
20. L L, Y Y, H Y, C J, D H, C W, et al. Human umbilical cord mesenchymal stem cells transplantation promotes cutaneous wound healing of severe burned rats. PloS One. (2014) 9:1–17. doi: 10.1371/journal.pone.0088348
21. Lorenz P, Leavitt T, Hu M, Marshall C, Barnes L, Longaker M. Stem cells and chronic wound healing: state of the art. Chronic Wound Care Manag Res. (2016) 7:7–27. doi: 10.2147/cwcmr.s84369
22. Krisp C, Jacobsen F, Mckay MJ, Molloy MP, Steinstraesser L, Wolters DA. Proteome analysis reveals antiangiogenic environments in chronic wounds of diabetes mellitus type 2 patients. Proteomics. (2013) 13:2670–81. doi: 10.1002/pmic.201200502
23. Chang HK, Kim PH, Cho HM, Yum SY, Choi YJ, Son YS, et al. Inducible HGF-secreting human umbilical cord blood-derived MSCs produced via TALEN-mediated genome editing promoted angiogenesis. Mol Ther. (2016) 24:1644–54. doi: 10.1038/mt.2016.120
24. Ennis WJ, Lee C, Gellada K, Corbiere TF, Koh TJ. Advanced technologies to improve wound healing: Electrical stimulation, vibration therapy, and ultrasound-what is the evidence? Plast Reconstr Surg. (2016) 138:94S–104S. doi: 10.1097/PRS.0000000000002680
25. Huang C, Leavitt T, Bayer LR, Orgill DP. Effect of negative pressure wound therapy on wound healing. Curr Probl Surg. (2014) 51:301–31. doi: 10.1067/j.cpsurg.2014.04.001
26. Beldon P. Basic science of wound healing. Surgery. (2010) 28:409–12. doi: 10.1016/j.mpsur.2010.05.007
27. Krafts KP. Tissue repair: The hidden drama. Organogenesis. (2010) 6:225–33. doi: 10.4161/org.6.4.12555
28. Rittié L, Fisher GJ. UV-light-induced signal cascades and skin aging. Ageing Res Rev. (2002) 1:705–20. doi: 10.1016/S1568-1637(02)00024-7
29. Martin P. Wound healing - Aiming for perfect skin regeneration. Sci (80- ). (1997) 276:75–81. doi: 10.1126/science.276.5309.75
30. Gurtner GC, Werner S, Barrandon Y, Longaker MT. Wound repair and regeneration. Nature. (2008) 453:314–21. doi: 10.1038/nature07039
31. Ribatti D, Nico B, Crivellato E, Vacca A. Macrophages and tumor angiogenesis. Leukemia. (2007) 21:2085–9. doi: 10.1038/sj.leu.2404900
32. Italiano JE, Shivdasani RA. Megakaryocytes and beyond: The birth of platelets. J Thromb Haemost. (2003) 1:1174–82. doi: 10.1046/j.1538-7836.2003.00290.x
33. Ullah I, Subbarao RB, Rho GJ. Human mesenchymal stem cells - Current trends and future prospective. Biosci Rep. (2015) 35:e00191. doi: 10.1042/BSR20150025
34. Eming SA, Krieg T, Davidson JM. Inflammation in wound repair: Molecular and cellular mechanisms. J Invest Dermatol. (2007) 127:514–25. doi: 10.1038/sj.jid.5700701
35. Thiruvoth F, Mohapatra D, Sivakumar D, Chittoria R, Nandhagopal V. Current concepts in the physiology of adult wound healing. Plast Aesthetic Res. (2015) 2:250. doi: 10.4103/2347-9264.158851
36. Ebner JA, Maytin EV. Cutaneous wound healing. Dermatologic Surg. (2008) 90:81–100. doi: 10.1016/B978-0-7020-3049-9.00006-3
37. Suzuki-Inoue K. [Activation and inhibitory mechanisms of blood platelets]. Nihon Rinsho. (2014) 72:1212–7.
38. Martins-Green M, Petreaca M, Wang L. Chemokines and their receptors are key players in the orchestra that regulates wound healing. Adv Wound Care. (2013) 2:327–47. doi: 10.1089/wound.2012.0380
39. Luis Cañedo D, Mara Cañedo A. Skin acute wound healing : A comprehensive review. Int J Inflam. (2019) 2019:1–15. doi: 10.1155/2019/3706315
40. Tennent GA, Brennan SO, Stangou AJ, O’Grady J, Hawkins PN, Pepys MB. Human plasma fibrinogen is synthesized in the liver. Blood. (2007) 109:1971–4. doi: 10.1182/blood-2006-08-040956
41. Karra R, Walter AO, Wu SM. The relationship between cardiac endothelium and fibroblasts: It’s complicated. J Clin Invest. (2017) 127:2892–4. doi: 10.1172/JCI95492
42. Furie B, Furie BC. Mechanisms of thrombus formation. Mech Disease. N Engl J Med. (2008) 359:938–49. doi: 10.1056/NEJMra0801082
43. Irena P, Olivera S CYN, Horacio R GNA, Andrew S BPS, Laiqua K RIR, Marjana T-C. Epithelialization in wound healing: A comprehensive review. Adv Wound Care. (2014) 3:445–64. doi: 10.1089/wound.2013.0473
44. Goffin JM, Pittet P, Csucs G, Lussi JW, Meister JJ, Hinz B. Focal adhesion size controls tension-dependent recruitment of α-smooth muscle actin to stress fibers. J Cell Biol. (2006) 172:259–68. doi: 10.1083/jcb.200506179
45. Park JE, Barbul A. Understanding the role of immune regulation in wound healing. Am J Surg. (2004) 187:S11–6. doi: 10.1016/S0002-9610(03)00296-4
46. Woo YC, Park SS, Subieta AR, Brennan TJ. Changes in tissue pH and temperature after incision indicate acidosis may contribute to postoperative pain. Anesthesiology. (2004) 101:468–75. doi: 10.1097/00000542-200408000-00029
47. Greco V, Chen T, Rendl M, Schober M, Pasolli HA, Stokes N, et al. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell. (2009) 4:155–69. doi: 10.1016/j.stem.2008.12.009
48. Pradhan S, Khatlani T, Nairn AC, Vijayan KV. The heterotrimeric G protein G1 interacts with the catalytic subunit of protein phosphatase 1 and modulates G protein–coupled receptor signaling in platelets. J Biol Chem. (2017) 292:13133–42. doi: 10.1074/jbc.M117.796656
49. Goren I, Allmann N, Yogev N, Schürmann C, Linke A, Holdener M, et al. A transgenic mouse model of inducible macrophage depletion: Effects of diphtheria toxin-driven lysozyme m-specific cell lineage ablation on wound inflammatory, angiogenic, and contractive processes. Am J Pathol. (2009) 175:132–47. doi: 10.2353/ajpath.2009.081002
50. Berndt MC, Shen Y, Dopheide SM, Gardiner EE, Andrews RK. The vascular biology of the glycoprotein Ib-IX-V complex. Thromb Haemost. (2001) 86:178–88. doi: 10.1055/s-0037-1616216
51. Neligan PC. Craniofacial, head and neck surgery and pediatric plastic surgery. Plast Surg. (2012) 3:1170.
52. Caley MP, Martins VLC, O’Toole EA. Metalloproteinases and wound healing. Adv Wound Care. (2015) 4:225–34. doi: 10.1089/wound.2014.0581
53. FitzGerald GA. Mechanisms of platelet activation: Thromboxane A2 as an amplifying signal for other agonists. Am J Cardiol. (1991) 68:11–15. doi: 10.1016/0002-9149(91)90379-Y
54. Gillitzer R, Goebeler M. Chemokines in cutaneous wound healing. J Leukoc Biol. (2001) 69:513–21. doi: 10.1189/jlb.69.4.513
55. H S, B M, B N, J H, W NH. The direct thrombin inhibitors (argatroban, bivalirudin and lepirudin) and the indirect Xa-inhibitor (danaparoid) increase fibrin network porosity and thus facilitate fibrinolysis. Thromb Haemost. (2010) 103:1076–84. doi: 10.1160/TH09-05-0306
56. Tuan TL, Wu H, Huang EY, Chong SSN, Laug W, Messadi D, et al. Increased plasminogen activator inhibitor-1 in keloid fibroblasts may account for their elevated collagen accumulation in fibrin gel cultures. Am J Pathol. (2003) 162:1579–89. doi: 10.1016/S0002-9440(10)64292-7
57. Werner S, Grose R. Regulation of wound healing by growth factors and cytokines. Physiol Rev. (2003) 83:835–70. doi: 10.1152/physrev.2003.83.3.835
58. Nami N, Feci L, Napoliello L, Giordano A, Lorenzini S, Galeazzi M, et al. Crosstalk between platelets and PBMC: New evidence in wound healing. Platelets. (2016) 27:143–8. doi: 10.3109/09537104.2015.1048216
59. Petreaca ML, Yao M, Liu Y, DeFea K, Martins-Green M. Transactivation of vascular endothelial growth factor receptor-2 by interleukin-8 (IL-8/CXCL8) is required for IL-8/CXCL8-induced endothelial permeability. Mol Biol Cell. (2007) 18:5014–23. doi: 10.1091/mbc.E07-01-0004
60. Schraufstatter IU, Chung J, Burger M. IL-8 activates endothelial cell CXCR1 and CXCR2 through Rho and Rac signaling pathways. Am J Physiol - Lung Cell Mol Physiol. (2001) 280:1094–103. doi: 10.1152/ajplung.2001.280.6.l1094
61. Nathan C. Neutrophils and immunity: challenges and opportunities. Nat Rev Immunol. (2006) 6:173–82. doi: 10.1038/nri1785
62. Conus S, Perozzo R, Reinheckel T, Peters C, Scapozza L, Yousefi S, et al. Caspase-8 is activated by cathepsin D initiating neutrophil apoptosis during the resolution of inflammation. J Exp Med. (2008) 205:685–98. doi: 10.1084/jem.20072152
63. Simpson DM, Ross R. The neutrophilic leukocyte in wound repair a study with antineutrophil serum. J Clin Invest. (1972) 51:2009–23. doi: 10.1172/JCI107007
64. Daley JM, Reichner JS, Mahoney EJ, Manfield L, Henry WL, MastroFrancesco B, et al. Modulation of macrophage phenotype by soluble product(s) released from neutrophils. J Immunol. (2005) 174:2265–72. doi: 10.4049/jimmunol.174.4.2265
65. Jakubzick C, Gautier EL, Gibbings SL, Sojka DK, Schlitzer A, Johnson TE, et al. Minimal differentiation of classical monocytes as they survey steady-state tissues and transport antigen to lymph nodes. Immunity. (2013) 39:599–610. doi: 10.1016/j.immuni.2013.08.007
66. Roberts HR, Tabares AH. Overview of the coagulation reactions. Mol Basis Thromb Hemost. (1995) 58:35–50. doi: 10.4103/0019-5049.144643
67. Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol. (2013) 13:159–75. doi: 10.1038/nri3399
68. Su Y, Richmond A. Chemokine regulation of neutrophil infiltration of skin wounds. Adv Wound Care. (2015) 4:631–40. doi: 10.1089/wound.2014.0559
69. Mantovani A, Cassatella MA, Costantini C, Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol. (2011) 11:519–31. doi: 10.1038/nri3024
70. Futosi K, Fodor S, Mócsai A. Reprint of Neutrophil cell surface receptors and their intracellular signal transduction pathways. Int Immunopharmacol. (2013) 17:1185–97. doi: 10.1016/j.intimp.2013.11.010
71. Schnoor M, Alcaide P, Voisin MB, Van Buul JD. Crossing the vascular wall: common and unique mechanisms exploited by different leukocyte subsets during extravasation. Mediators Inflammation. (2015) 2015:946509. doi: 10.1155/2015/946509
72. Mayadas TN, Cullere X, Lowell CA. The multifaceted functions of neutrophils. Annu Rev Pathol Mech Dis. (2014) 9:181–218. doi: 10.1146/annurev-pathol-020712-164023
73. Brinkmann V, R U, G C, F B, U Y, W DS, et al. Neutrophil extracellular traps kill bacteria. Sci (80- ). (2004) 303:1532–5. doi: 10.1126/science.1092385
74. Boyette LB, MacEdo C, Hadi K, Elinoff BD, Walters JT, Ramaswami B, et al. Phenotype, function, and differentiation potential of human monocyte subsets. PloS One. (2017) 12:e0176460. doi: 10.1371/journal.pone.0176460
75. Levin R, Grinstein S, Canton J. The life cycle of phagosomes: formation, maturation, and resolution. Immunol Rev. (2016) 273:156–79. doi: 10.1111/imr.12439
76. Gordon S, Taylor PR. Monocyte and macrophage heterogeneity. Nat Rev Immunol. (2005) 5:953–64. doi: 10.1038/nri1733
77. Patel AA, Zhang Y, Fullerton JN, Boelen L, Rongvaux A, Maini AA, et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J Exp Med. (2017) 214:1913–23. doi: 10.1084/jem.20170355
78. Willenborg S, Eming SA. Macrophages-sensors and effectors coordinating skin damage and repair. JDDG - J Ger Soc Dermatol. (2014) 12:214–21. doi: 10.1111/ddg.12290
79. Ginhoux F, Schultze JL, Murray PJ, Ochando J, Biswas SK. New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat Immunol. (2016) 17:34–40. doi: 10.1038/ni.3324
80. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. (2008) 8:958–69. doi: 10.1038/nri2448
81. West HC, Bennett CL. Redefining the role of langerhans cells as immune regulators within the skin. Front Immunol. (2018) 8:1941. doi: 10.3389/fimmu.2017.01941
82. Deckers J, Hammad H, Hoste E. Langerhans cells: Sensing the environment in health and disease. Front Immunol. (2018) 9:93. doi: 10.3389/fimmu.2018.00093
83. Schaible UE, Weiss G. Macrophage defense mechanisms against intracellular bacteria. Immunol Rev. (2015) 264:182–203. doi: 10.1111/imr.12266
84. Buckley CD, Gilroy DW, Serhan CN. Proresolving lipid mediators and mechanisms in the resolution of acute inflammation. Immunity. (2014) 40:315–27. doi: 10.1016/j.immuni.2014.02.009
85. Bonefeld CM, Geisler C. The role of innate lymphoid cells in healthy and inflamed skin. Immunol Lett. (2016) 179:25–8. doi: 10.1016/j.imlet.2016.01.005
86. Rak GD, Osborne LC, Siracusa MC, Kim BS, Wang K, Bayat A, et al. IL-33-dependent group 2 innate lymphoid cells promote cutaneous wound healing. J Invest Dermatol. (2016) 136:487–96. doi: 10.1038/JID.2015.406
87. Shalaby KH. Pattern recognition receptors and aging. Immunol Aging. (2014), 87–143. doi: 10.1007/978-3-642-39495-9_8
88. Chou C, Li MO. Tissue-resident lymphocytes across innate and adaptive lineages. Front Immunol. (2018) 9:2104. doi: 10.3389/fimmu.2018.02104
89. Strbo N, Yin N, Stojadinovic O. Innate and adaptive immune responses in wound epithelialization. Adv Wound Care. (2014) 3:492–501. doi: 10.1089/wound.2012.0435
90. Chen L, Dipietro LA. Toll-like receptor function in acute wounds. Adv Wound Care. (2017) 6:344–55. doi: 10.1089/wound.2017.0734
91. Hofmann U, Frantz S. Role of lymphocytes in myocardial injury, healing, and remodeling after myocardial infarction. Circ Res. (2015) 116:354–67. doi: 10.1161/CIRCRESAHA.116.304072
92. Mackay LK, Stock AT, Ma JZ, Jones CM, Kent SJ, Mueller SN, et al. Long-lived epithelial immunity by tissue-resident memory T (TRM) cells in the absence of persisting local antigen presentation. Proc Natl Acad Sci U.S.A. (2012) 109:7037–42. doi: 10.1073/pnas.1202288109
93. Li J, Tan J, Martino MM, Lui KO. Regulatory T-cells: Potential regulator of tissue repair and regeneration. Front Immunol. (2018) 9:585. doi: 10.3389/fimmu.2018.00585
94. Clark RA. Skin-resident T cells: The ups and downs of on site immunity. J Invest Dermatol. (2010) 130:362–70. doi: 10.1038/jid.2009.247
95. Rosenblum MD, Way SS, Abbas AK. Regulatory T cell memory. Nat Rev Immunol. (2016) 16:90–101. doi: 10.1038/nri.2015.1
96. Sharma A, Rudra D. Emerging functions of regulatory T cells in tissue homeostasis. Front Immunol. (2018) 9:883. doi: 10.3389/fimmu.2018.00883
97. Ali N, Rosenblum MD. Regulatory T cells in skin. Immunology. (2017) 152:372–81. doi: 10.1111/imm.12791
98. Giganti G, Atif M, Mohseni Y, Mastronicola D, Grageda N, Povoleri GAM, et al. Treg cell therapy: How cell heterogeneity can make the difference. Eur J Immunol. (2021) 51:39–55. doi: 10.1002/eji.201948131
99. Eyerich K, Dimartino V, Cavani A. IL-17 and IL-22 in immunity: Driving protection and pathology. Eur J Immunol. (2017) 47:607–14. doi: 10.1002/eji.201646723
100. Egbuniwe IU, Karagiannis SN, Nestle FO, Lacy KE. Revisiting the role of B cells in skin immune surveillance. Trends Immunol. (2015) 36:102–11. doi: 10.1016/j.it.2014.12.006
101. van de Veen W, Stanic B, Wirz OF, Jansen K, Globinska A, Akdis M. Role of regulatory B cells in immune tolerance to allergens and beyond. J Allergy Clin Immunol. (2016) 138:654–65. doi: 10.1016/j.jaci.2016.07.006
102. Raziyeva K, Kim Y, Zharkinbekov Z, Kassymbek K, Jimi S, Saparov A. Immunology of acute and chronic wound healing. Biomolecules. (2021) 11:700. doi: 10.3390/biom11050700
103. Sîrbulescu RF, Boehm CK, Soon E, Wilks MQ, Ilieş I, Yuan H, et al. Mature B cells accelerate wound healing after acute and chronic diabetic skin lesions. Wound Repair Regener. (2017) 25:774–91. doi: 10.1111/wrr.12584
104. Foley TT, Ehrlich HP. Through gap junction communications, co-cultured mast cells and fibroblasts generate fibroblast activities allied with hypertrophic scarring. Plast Reconstr Surg. (2013) 131:1036–44. doi: 10.1097/PRS.0b013e3182865c3f
105. Garbuzenko E, Nagler A, Pickholtz D, Gillery P, Reich R, Maquart FX, et al. Human mast cells stimulate fibroblast proliferation, collagen synthesis and lattice contraction: A direct role for mast cells in skin fibrosis. Clin Exp Allergy. (2002) 32:237–46. doi: 10.1046/j.1365-2222.2002.01293.x
106. Nakano T, Sonoda T, Hayashi C, Yamatodani A, Kanayama Y, Yamamura T, et al. Fate of bone marrow-derived cultured mast cells after intracutaneous, intraperitoneal, and intravenous transfer into genetically mast cell-deficient W/Wν mice: Evidence that cultured mast cells can give rise to both connective tissue type and mucosal mast. J Exp Med. (1985) 162:1025–43. doi: 10.1084/jem.162.3.1025
107. Rao KN, Brown MA. Mast cells: Multifaceted immune cells with diverse roles in health and disease. Ann N Y Acad Sci. (2008) 1143:83–104. doi: 10.1196/annals.1443.023
108. Romani N, Holzmann S, Tripp CH, Koch F, Stoitzner P. Langerhans cells - Dendritic cells of the epidermis. Apmis. (2003) 111:725–40. doi: 10.1034/j.1600-0463.2003.11107805.x
109. Komi DEA, Khomtchouk K, Santa Maria PL. A review of the contribution of mast cells in wound healing: involved molecular and cellular mechanisms. Clin Rev Allergy Immunol. (2020) 58:298–312. doi: 10.1007/s12016-019-08729-w
110. Dong J, Chen L, Zhang Y, Jayaswal N, Mezghani I, Zhang W, et al. Mast cells in diabetes and diabetic wound healing. Adv Ther. (2020) 37:4519–37. doi: 10.1007/s12325-020-01499-4
111. Malissen B, Tamoutounour S, Henri S. The origins and functions of dendritic cells and macrophages in the skin. Nat Rev Immunol. (2014) 14:417–28. doi: 10.1038/nri3683
112. Ellis S, Lin EJ, Tartar D. Immunology of wound healing. Curr Dermatol Rep. (2018) 7:350–8. doi: 10.1007/s13671-018-0234-9
113. Brown DL, Kao WWY, Greenhalgh DG. Apoptosis down-regulates inflammation under the advancing epithelial wound edge: Delayed patterns in diabetes and improvement with topical growth factors. Surgery. (1997) 121:372–80. doi: 10.1016/S0039-6060(97)90306-8
114. Kamil S, Mohan RR. Corneal stromal wound healing: Major regulators and therapeutic targets. Ocul Surf. (2021) 19:290–306. doi: 10.1016/j.jtos.2020.10.006
115. Wu YS, Chen SN. Apoptotic cell: Linkage of inflammation and wound healing. Front Pharmacol. (2014) 5 JAN:1. doi: 10.3389/fphar.2014.00001
116. Miller MA, Zachary JF. Mechanisms and morphology of cellular injury, adaptation, and death. Pathol Basis Vet Dis Expert Consult. (2017) 66:2–43.e19. doi: 10.1016/B978-0-323-35775-3.00001-1
117. Desmouliere A, Redard M, Darby I, Gabbiani G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am J Pathol. (1995) 146:56–66.
118. Greenhalgh DG. The role of apoptosis in wound healing. Int J Biochem Cell Biol. (1998) 30:1019–30. doi: 10.1016/S1357-2725(98)00058-2
119. Avramović V, Petrović V, Jović M, Vlahović P. Quantification of cells expressing markers of proliferation and apoptosis in chronic tonsilitis. Acta Otorhinolaryngol Ital. (2015) 35:277–84.
120. Darby IA, Bisucci T, Hewitson TD, MacLellan DG. Apoptosis is increased in a model of diabetes-impaired wound healing in genetically diabetic mice. Int J Biochem Cell Biol. (1997) 29:191–200. doi: 10.1016/S1357-2725(96)00131-8
121. Ren H, Zhao F, Zhang Q, Huang X, Wang Z. Autophagy and skin wound healing. Burn Trauma. (2022) 10:tkac003. doi: 10.1093/burnst/tkac003
122. Hill D, Cosgarea I, Reynolds N, Lovat P, Armstrong J. Research techniques made simple: analysis of autophagy in the skin. J Invest Dermatol. (2021) 141:5–9.e1. doi: 10.1016/j.jid.2020.10.004
123. Ullah I, Ritchie ND, Evans TJ. The interrelationship between phagocytosis, autophagy and formation of neutrophil extracellular traps following infection of human neutrophils by Streptococcus pneumoniae. Innate Immun. (2017) 23:413–23. doi: 10.1177/1753425917704299
124. Zhu Y, Li H, Ding S, Wang Y. Autophagy inhibition promotes phagocytosis of macrophage and protects mice from methicillin-resistant staphylococcus aureus pneumonia. J Cell Biochem. (2018) 119:4808–14. doi: 10.1002/jcb.26677
125. Kawano A, Ariyoshi W, Yoshioka Y, Hikiji H, Nishihara T, Okinaga T. Docosahexaenoic acid enhances M2 macrophage polarization via the p38 signaling pathway and autophagy. J Cell Biochem. (2019) 120:12604–17. doi: 10.1002/jcb.28527
126. An Y, Liu WJ, Xue P, Ma Y, Zhang LQ, Zhu B, et al. Autophagy promotes MSC-mediated vascularization in cutaneous wound healing via regulation of VEGF secretion article. Cell Death Dis. (2018) 9:58. doi: 10.1038/s41419-017-0082-8
127. Jakovljevic J, Harrell CR, Fellabaum C, Arsenijevic A, Jovicic N, Volarevic V. Modulation of autophagy as new approach in mesenchymal stem cell-based therapy. BioMed Pharmacother. (2018) 104:404–10. doi: 10.1016/j.biopha.2018.05.061
128. Liu Y, Wang N, Zhang S, Liang Q. Autophagy protects bone marrow mesenchymal stem cells from palmitate-induced apoptosis through the ROS-JNK/p38 MAPK signaling pathways. Mol Med Rep. (2018) 18:1485–94. doi: 10.3892/mmr.2018.9100
129. Lv B, Hua T, Li F, Han J, Fang J, Xu L, et al. Hypoxia-inducible factor 1 α protects mesenchymal stem cells against oxygen-glucose deprivation-induced injury via autophagy induction and PI3K/AKT/mTOR signaling pathway. Am J Transl Res. (2017) 9:2492–9.
130. Zhang F, Gao F, Wang K, Liu X, Zhang Z. MiR-34a inhibitor protects mesenchymal stem cells from hyperglycaemic injury through the activation of the SIRT1/FoxO3a autophagy pathway. Stem Cell Res Ther. (2021) 12:115. doi: 10.1186/s13287-021-02183-2
131. Zhao YG, Codogno P, Zhang H. Machinery, regulation and pathophysiological implications of autophagosome maturation. Nat Rev Mol Cell Biol. (2021) 22:733–50. doi: 10.1038/s41580-021-00392-4
132. Skouta R, Dixon SJ, Wang J, Dunn DE, Orman M, Shimada K, et al. Ferrostatins inhibit oxidative lipid damage and cell death in diverse disease models. J Am Chem Soc. (2014) 136:4551–6. doi: 10.1021/ja411006a
133. Huang Y, Ding Y, Wang B, Ji Q, Peng C, Tan Q. Neutrophils extracellular traps and ferroptosis in diabetic wounds. Int Wound J. (2023) 20:3840–54. doi: 10.1111/iwj.14231
134. Mandal PK, Seiler A, Perisic T, Kölle P, Canak AB, Förster H, et al. System xc- and thioredoxin reductase 1 cooperatively rescue glutathione deficiency. J Biol Chem. (2010) 285:22244–53. doi: 10.1074/jbc.M110.121327
135. Tang L, Zhang Y, Qian Z, Shen X. The mechanism of Fe2+-initiated lipid peroxidation in liposomes: The dual function of ferrous ions, the roles of the pre-existing lipid peroxides and the lipid peroxyl radical. Biochem J. (2000) 352:27–36. doi: 10.1042/0264-6021:3520027
136. Bi M, Li D, Zhang J. Research progress and insights on the role of ferroptosis in wound healing. Int Wound J. (2023) 20:2473–81. doi: 10.1111/iwj.14102
137. Shuangwen L, Yuan L, Ziyu W, Zhongming W, Hongjuan F. Diabetic ferroptosis plays an important role in triggering on inflammation in diabetic wound. Am J Physiol Endocrinol Metab. (2021) 321:E509–20. doi: 10.1152/ajpendo.00042.2021
138. Gao SQ, Chang C, Li JJ, Li Y, Niu XQ, Zhang DP, et al. Co-delivery of deferoxamine and hydroxysafflor yellow A to accelerate diabetic wound healing via enhanced angiogenesis. Drug Delivery. (2018) 25:1779–89. doi: 10.1080/10717544.2018.1513608
139. Ye LF, Chaudhary KR, Zandkarimi F, Harken AD, Kinslow CJ, Upadhyayula PS, et al. Radiation-induced lipid peroxidation triggers ferroptosis and synergizes with ferroptosis inducers. SSRN Electron J. (2019) 15:469–84. doi: 10.2139/ssrn.3474050
140. Gan F, Wang R, Lyu P, Li Y, Fu R, Du Y, et al. Plasma-derived exosomes boost the healing of irradiated wound by regulating cell proliferation and ferroptosis. J BioMed Nanotechnol. (2021) 17:100–14. doi: 10.1166/jbn.2021.3008
141. Vats K, Kruglov O, Mizes A, Samovich SN, Amoscato AA, Tyurin VA, et al. Keratinocyte death by ferroptosis initiates skin inflammation after UVB exposure. Redox Biol. (2021) 47:102143. doi: 10.1016/j.redox.2021.102143
142. Stockwell BR. Ferroptosis: Death by lipid peroxidation. Free Radic Biol Med. (2018) 120:S7. doi: 10.1016/j.freeradbiomed.2018.04.034
143. Su LJ, Zhang JH, Gomez H, Murugan R, Hong X, Xu D, et al. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid Med Cell Longev. (2019) 2019:5080843. doi: 10.1155/2019/5080843
144. Friedmann Angeli JPJP, Schneider M, Proneth B, Tyurina YY, Tyurin VA, Hammond VJ, et al. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat Chem Biol. (2017) 10:232–43. doi: 10.1038/nchembio.2238
145. Weinheimer-Haus EM, Mirza RE, Koh TJ. Nod-like receptor protein-3 inflammasome plays an important role during early stages of wound healing. PloS One. (2015) 10:e0119106. doi: 10.1371/journal.pone.0119106
146. Li Y, Tang Y, Shi S, Gao S, Wang Y, Xiao D, et al. Tetrahedral framework nucleic acids ameliorate insulin resistance in type 2 diabetes mellitus via the PI3K/Akt pathway. ACS Appl Mater Interfaces. (2021) 13:40354–64. doi: 10.1021/acsami.1c11468
147. Chen F, Wei G, Xu J, Ma X, Wang Q. Naringin ameliorates the high glucose-induced rat mesangial cell inflammatory reaction by modulating the NLRP3 Inflammasome. BMC Complement Altern Med. (2018) 18. doi: 10.1186/s12906-018-2257-y
148. Mu X, Wu X, He W, Liu Y, Wu F, Nie X. Pyroptosis and inflammasomes in diabetic wound healing. Front Endocrinol (Lausanne). (2022) 13:950798. doi: 10.3389/fendo.2022.950798
149. Chen Y, Shi S, Li B, Lan T, Yuan K, Yuan J, et al. Therapeutic effects of self-assembled tetrahedral framework nucleic acids on liver regeneration in acute liver failure. ACS Appl Mater Interfaces. (2022) 14:13136–46. doi: 10.1021/acsami.2c02523
150. Liu N, Zhang X, Li N, Zhou M, Zhang T, Li S, et al. Tetrahedral framework nucleic acids promote corneal epithelial wound healing in vitro and in vivo. Small. (2019) 15:e1901907. doi: 10.1002/smll.201901907
151. Lin S, Zhang Q, Li S, Zhang T, Wang L, Qin X, et al. Antioxidative and angiogenesis-promoting effects of tetrahedral framework nucleic acids in diabetic wound healing with activation of the Akt/Nrf2/HO-1 pathway. ACS Appl Mater Interfaces. (2020) 12:11397–408. doi: 10.1021/acsami.0c00874
152. Jiang Y, Li S, Zhang T, Zhang M, Chen Y, Wu Y, et al. Tetrahedral framework nucleic acids inhibit skin fibrosis via the pyroptosis pathway. ACS Appl Mater Interfaces. (2022) 14:15069–79. doi: 10.1021/acsami.2c02877
153. Ma JW, Ji DD, Li QQ, Zhang T, Luo L. Inhibition of connexin 43 attenuates oxidative stress and apoptosis in human umbilical vein endothelial cells. BMC Pulm Med. (2020) 20:19. doi: 10.1186/s12890-019-1036-y
154. Zhang K, Chai B, Ji H, Chen l, Ma Y, Zhu L, et al. Bioglass promotes wound healing by inhibiting endothelial cell pyroptosis through regulation of the connexin 43/reactive oxygen species (ROS) signaling pathway. Lab Investig. (2022) 102:90–101. doi: 10.1038/s41374-021-00675-6
155. Dai J, Zhang X, Wang Y, Chen H, Chai Y. ROS-activated NLRP3 inflammasome initiates inflammation in delayed wound healing in diabetic rats. Int J Clin Exp Pathol. (2017) 10:9902–9. doi: 10.3390/ijms23126820
156. Zhu J, Zhang M, Gao Y, Qin X, Zhang T, Cui W, et al. Tetrahedral framework nucleic acids promote scarless healing of cutaneous wounds via the AKT-signaling pathway. Signal Transduct Target Ther. (2020) 5:120. doi: 10.1038/s41392-020-0173-3
157. Tsvetkov P, Coy S, Petrova B, Dreishpoon M, Verma A, Abdusamad M, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Sci (80- ). (2022) 375:1254–61. doi: 10.1126/science.abf0529
158. Wiebelhaus N, Zaengle-Barone JM, Hwang KK, Franz KJ, Fitzgerald MC. Protein Folding Stability Changes across the Proteome Reveal Targets of Cu Toxicity in E. coli. ACS Chem Biol. (2021) 16:214–24. doi: 10.1021/acschembio.0c00900
159. Joshi PR, Sadre S, Guo XA, McCoy JG, Mootha VK. Lipoylation is dependent on the ferredoxin FDX1 and dispensable under hypoxia in human cells. J Biol Chem. (2023) 299:105075. doi: 10.1016/j.jbc.2023.105075
160. Rowland EA, Snowden CK, Cristea IM. Protein lipoylation: an evolutionarily conserved metabolic regulator of health and disease. Curr Opin Chem Biol. (2018) 42:76–85. doi: 10.1016/j.cbpa.2017.11.003
161. Guthrie LM, Soma S, Yuan S, Silva A, Zulkifli M, Snavely TC, et al. Elesclomol alleviates Menkes pathology and mortality by escorting Cu to cuproenzymes in mice. Sci (80- ). (2020) 368:620–5. doi: 10.1126/science.aaz8899
162. Cobine PA, Brady DC. Cuproptosis: Cellular and molecular mechanisms underlying copper-induced cell death. Mol Cell. (2022) 82:1786–7. doi: 10.1016/j.molcel.2022.05.001
163. Zhu X, Boulet A, Buckley KM, Phillips CB, Gammon MG, Oldfather LE, et al. Mitochondrial copper and phosphate transporter specificity was defined early in the evolution of eukaryotes. Elife. (2021) 10:1–65. doi: 10.7554/eLife.64690
164. Attar N, Campos OA, Vogelauer M, Cheng C, Xue Y, Schmollinger S, et al. The histone H3-H4 tetramer is a copper reductase enzyme. Sci (80- ). (2020) 369:59–64. doi: 10.1126/science.aba8740
165. Tang D, Chen X, Kroemer G. Cuproptosis: a copper-triggered modality of mitochondrial cell death. Cell Res. (2022) 32:417–8. doi: 10.1038/s41422-022-00653-7
166. Ferguson MWJ, O’Kane S. Scar-free healing: From embryonic mechanism to adult therapeutic intervention. Philos Trans R Soc B Biol Sci. (2004) 359:839–50. doi: 10.1098/rstb.2004.1475
167. Ferguson JE, Kelley RW, Patterson C. Mechanisms of endothelial differentiation in embryonic vasculogenesis. Arterioscler Thromb Vasc Biol. (2005) 25:2246–54. doi: 10.1161/01.ATV.0000183609.55154.44
168. Ahn S, Ardoña HAM, Campbell PH, Gonzalez GM, Parker KK. Alfalfa nanofibers for dermal wound healing. ACS Appl Mater Interfaces. (2019) 11:33535–47. doi: 10.1021/acsami.9b07626
169. Eilken HM, Adams RH. Dynamics of endothelial cell behavior in sprouting angiogenesis. Curr Opin Cell Biol. (2010) 22:617–25. doi: 10.1016/j.ceb.2010.08.010
170. Gerhardt H, Golding M, Fruttiger M, Ruhrberg C, Lundkvist A, Abramsson A, et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J Cell Biol. (2003) 161:1163–77. doi: 10.1083/jcb.200302047
171. Khoury CC, Ziyadeh FN. Angiogenic factors. Contrib Nephrol. (2011) 170:83–92. doi: 10.1159/000324950
172. Cook-Mills JM, Deem TL. Active participation of endothelial cells in inflammation. J Leukoc Biol. (2005) 77:487–95. doi: 10.1189/jlb.0904554
173. Muller WA, Weigl SA, Deng X, Phillips DM. PECAM-1 is required for transendothelial migration of leukocytes. J Exp Med. (1993) 178:449–60. doi: 10.1084/jem.178.2.449
174. Nagaoka T, Kaburagi Y, Hamaguchi Y, Hasegawa M, Takehara K, Steeber DA, et al. Delayed wound healing in the absence of intercellular adhesion molecule-1 or L-selectin expression. Am J Pathol. (2000) 157:237–47. doi: 10.1016/S0002-9440(10)64534-8
175. Inducible PIS, Tumor BY, Factor N, Weller A, Isenmann S, Vestwebers D. Cloning of the Mouse Endothelial Selectins. J Biol Chem. (1992) 267(21):15176–83. doi: 10.1016/S0021-9258(18)42162-X
176. Silver FH, Freeman JW, Devore D. Viscoelastic properties of human skin and processed dermis. Ski Res Technol. (2001) 7:18–23. doi: 10.1034/j.1600-0846.2001.007001018.x
177. Collins CA, Kretzschmar K, Watt FM. Reprogramming adult dermis to a neonatal state through epidermal activation of β-catenin. Development. (2011) 138:5189–99. doi: 10.1242/dev.064592
178. Davies PF. Hemodynamic shear stress and the endothelium in cardiovascular pathophysiology. Nat Clin Pract Cardiovasc Med. (2009) 6:16–26. doi: 10.1038/ncpcardio1397
179. Grant MB, May WS, Caballero S, Brown GAJ, Guthrie SM, Mames RN, et al. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med. (2002) 8:607–12. doi: 10.1038/nm0602-607
180. Grunewald M, Avraham I, Dor Y, Bachar-Lustig E, Itin A, Yung S, et al. VEGF-induced adult neovascularization: Recruitment, retention, and role of accessory cells. Cell. (2006) 124:175–89. doi: 10.1016/j.cell.2005.10.036
181. Kokovay E, Li L, Cunningham LA. Angiogenic recruitment of pericytes from bone marrow after stroke. J Cereb Blood Flow Metab. (2006) 26:545–55. doi: 10.1038/sj.jcbfm.9600214
182. Tracy LE, Minasian RA, Caterson EJ. Extracellular matrix and dermal fibroblast function in the healing wound. Adv Wound Care. (2016) 5:119–36. doi: 10.1089/wound.2014.0561
183. Wynn TA, Barron L. Macrophages: Master regulators of inflammation and fibrosis. Semin Liver Dis. (2010) 30:245–57. doi: 10.1055/s-0030-1255354
184. Greaves NS, Ashcroft KJ, Baguneid M, Bayat A. Current understanding of molecular and cellular mechanisms in fibroplasia and angiogenesis during acute wound healing. J Dermatol Sci. (2013) 72:206–17. doi: 10.1016/j.jdermsci.2013.07.008
185. Eckes B, Zweers MC, Zhang ZG, Hallinger R, Mauch C, Aumailley M, et al. Mechanical tension and integrin α2β1 regulate fibroblast functions. J Investig Dermatol Symp Proc. (2006) 11:66–72. doi: 10.1038/sj.jidsymp.5650003
186. Cheng F, Shen Y, Mohanasundaram P, Lindström M, Ivaska J, Ny T, et al. Vimentin coordinates fibroblast proliferation and keratinocyte differentiation in wound healing via TGF-β-Slug signaling. Proc Natl Acad Sci U.S.A. (2016) 113:E4320–7. doi: 10.1073/pnas.1519197113
187. Barbosa FL, Chaurasia SS, Cutler A, Asosingh K, Kaur H, de Medeiros FW, et al. Corneal myofibroblast generation from bone marrow-derived cells. Exp Eye Res. (2010) 91:92–6. doi: 10.1016/j.exer.2010.04.007
188. Gabbiani G. The myofibroblast in wound healing and fibrocontractive diseases. J Pathol. (2003) 200:500–3. doi: 10.1002/path.1427
189. Grinnell F. Fibroblasts, myofibroblasts, and wound contraction. J Cell Biol. (1994) 124:401–4. doi: 10.1083/jcb.124.4.401
190. Rousselle P, Montmasson M, Garnier C. Extracellular matrix contribution to skin wound re-epithelialization. Matrix Biol. (2019) 75–76:12–26. doi: 10.1016/j.matbio.2018.01.002
191. Stone RC, Pastar I, Ojeh N, Chen V, Liu S, Garzon KI, et al. Epithelial-mesenchymal transition in tissue repair and fibrosis. Cell Tissue Res. (2016) 365:495–506. doi: 10.1007/s00441-016-2464-0
192. Watt FM. Mammalian skin cell biology: At the interface between laboratory and clinic. Sci (80- ). (2014) 346:937–40. doi: 10.1126/science.1253734
193. Fujiwara H, Ferreira M, Donati G, Marciano DK, Linton JM, Sato Y, et al. The basement membrane of hair follicle stem cells is a muscle cell niche. Cell. (2011) 144:577–89. doi: 10.1016/j.cell.2011.01.014
194. Fujiwara T, Kubo T, Kanazawa S, Shingaki K, Taniguchi M, Matsuzaki S, et al. Direct contact of fibroblasts with neuronal processes promotes differentiation to myofibroblasts and induces contraction of collagen matrix in vitro. Wound Repair Regener. (2013) 21:588–94. doi: 10.1111/wrr.12059
195. Diamond J, Holmes M, Coughlin M. Endogenous NGF and nerve impulses regulate the collateral sprouting of sensory axons in the skin of the adult rat. J Neurosci. (1992) 12:1454–66. doi: 10.1523/jneurosci.12-04-01454.1992
196. Zochodne DW. The challenges and beauty of peripheral nerve regrowth. J Peripher Nerv Syst. (2012) 17:1–18. doi: 10.1111/j.1529-8027.2012.00378.x
197. Cámara-Lemarroy CR, Guzmán-De La Garza FJ, Fernández-Garza NE. Molecular inflammatory mediators in peripheral nerve degeneration and regeneration. Neuroimmunomodulation. (2010) 17:314–24. doi: 10.1159/000292020
198. Chen P, Piao X, Bonaldo P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. (2015) 130:605–18. doi: 10.1007/s00401-015-1482-4
199. Cattin AL, Burden JJ, Van Emmenis L, MacKenzie FE, Hoving JJA, Garcia Calavia N, et al. Macrophage-induced blood vessels guide Schwann cell-mediated regeneration of peripheral nerves. Cell. (2015) 162:1127–39. doi: 10.1016/j.cell.2015.07.021
200. Bos JD, Teunissen MB, Cairo I, Krieg SR, Kapsenberg ML, Das PK, et al. T-cell receptor gamma delta bearing cells in normal human skin. J Invest Dermatol. (1990) 94:37–42. doi: 10.1111/1523-1747.ep12873333
201. Gill SE, Pape MC, Khokha R, Watson AJ, Leco KJ. A null mutation for Tissue Inhibitor of Metalloproteinases-3 (Timp-3) impairs murine bronchiole branching morphogenesis. Dev Biol. (2003) 261:313–23. doi: 10.1016/S0012-1606(03)00318-X
202. Hinz B, Phan SH, Thannickal VJ, Prunotto M, Desmoulire A, Varga J, et al. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am J Pathol. (2012) 180:1340–55. doi: 10.1016/j.ajpath.2012.02.004
203. Hinz B. Formation and function of the myofibroblast during tissue repair. J Invest Dermatol. (2007) 127:526–37. doi: 10.1038/sj.jid.5700613
204. Verhaegen PDHM, Van Zuijlen PPM, Pennings NM, Van Marle J, Niessen FB, van der Horst CMAM, et al. Differences in collagen architecture between keloid, hypertrophic scar, normotrophic scar, and normal skin: An objective histopathological analysis. Wound Repair Regener. (2009) 17:649–56. doi: 10.1111/j.1524-475X.2009.00533.x
205. Xi-Qiao W, Ying-Kai L, Chun Q, Shu-Liang L. Hyperactivity of fibroblasts and functional regression of endothelial cells contribute to microvessel occlusion in hypertrophic scarring. Microvasc Res. (2009) 77:204–11. doi: 10.1016/j.mvr.2008.08.007
206. Morton LM, Phillips TJ. Wound healing and treating wounds Differential diagnosis and evaluation of chronic wounds. J Am Acad Dermatol. (2016) 74:589–605. doi: 10.1016/j.jaad.2015.08.068
207. Koźlik M, Wójcicki P. The use of stem cells in plastic and reconstructive surgery. Adv Clin Exp Med. (2014) 23:1011–7. doi: 10.17219/acem/37360
208. da Silva Meirelles L, Caplan AI, Nardi NB. In search of the in vivo identity of mesenchymal stem cells. Stem Cells. (2008) 26:2287–99. doi: 10.1634/stemcells.2007-1122
209. S NG, C AI. Mesenchymal stem cells: Mechanisms of inflammation. Annu Rev Pathol Mech Dis. (2011) 6:457–78. doi: 10.1146/annurev-pathol-011110-130230
210. Falanga V, Iwamoto S, Chartier M, Yufit T, Butmarc J, Kouttab N, et al. Autologous bone marrow-derived cultured mesenchymal stem cells delivered in a fibrin spray accelerate healing in murine and human cutaneous wounds. Tissue Eng. (2007) 13:1299–312. doi: 10.1089/ten.2006.0278
211. Sorrell JM, Caplan AI. Topical delivery of mesenchymal stem cells and their function in wounds. Stem Cell Res Ther. (2010) 1:30. doi: 10.1186/scrt30
212. Hu BY, Weick JP, Yu J, Ma LX, Zhang XQ, Thomson JA, et al. Neural differentiation of human induced pluripotent stem cells follows developmental principles but with variable potency. Proc Natl Acad Sci U.S.A. (2010) 107:4335–40. doi: 10.1073/pnas.0910012107
213. Pellegrini G, Traverso CE, Franzi AT, Zingirian M, Cancedda R, De Luca M, et al. Exosomes released from human induced pluripotent stem cells-derived MSCs facilitate cutaneous wound healing by promoting collagen synthesis and angiogenesis. J Transl Med. (2013) 13:49. doi: 10.1186/s12967-015-0417-0
214. Yang R, Zheng Y, Burrows M, Liu S, Wei Z, Nace A, et al. Generation of folliculogenic human epithelial stem cells from induced pluripotent stem cells. Nat Commun. (2014) 5:3071. doi: 10.1038/ncomms4071
215. Ehrenreich M, Ruszczak Z. Update on tissue-engineered biological dressings. Tissue Eng. (2006) 12:2407–24. doi: 10.1089/ten.2006.12.2407
216. Powers JG, Higham C, Broussard K, Phillips TJ. Wound healing and treating wounds Chronic wound care and management. J Am Acad Dermatol. (2016) 74:607–25. doi: 10.1016/j.jaad.2015.08.070
217. Varkey M, Ding J, Tredget E. Advances in skin substitutes—Potential of tissue engineered skin for facilitating anti-fibrotic healing. J Funct Biomater. (2015) 6:547–63. doi: 10.3390/jfb6030547
218. Veith AP, Henderson K, Spencer A, Sligar AD, Baker AB. Therapeutic strategies for enhancing angiogenesis in wound healing. Adv Drug Delivery Rev. (2019) 146:97–125. doi: 10.1016/j.addr.2018.09.010
219. Soliman AM, Barreda DR. Acute inflammation in tissue healing. Int J Mol Sci. (2023) 24:641. doi: 10.3390/ijms24010641
220. Wilgus TA. Growth factor–extracellular matrix interactions regulate wound repair. Adv Wound Care. (2012) 1:249–54. doi: 10.1089/wound.2011.0344
221. Jin R, Song G, Chai J, Gou X, Yuan G, Chen Z. Effects of concentrated growth factor on proliferation, migration, and differentiation of human dental pulp stem cells. vitro. J Tissue Eng. (2018) 9:2041731418817505. doi: 10.1177/2041731418817505
222. Guo S, DiPietro LA. Critical review in oral biology & medicine: Factors affecting wound healing. J Dent Res. (2010) 89:219–29. doi: 10.1177/0022034509359125
223. Negut I, Grumezescu V, Grumezescu AM. Treatment strategies for infected wounds. Molecules. (2018) 23:2392. doi: 10.3390/molecules23092392
224. Johnson N, Wang Y. Drug delivery systems for wound healing. Curr Pharm Biotechnol. (2015) 16:621–9. doi: 10.2174/1389201016666150206113720
225. Park JW, Hwang SR, Yoon IS. Advanced growth factor delivery systems in wound management and skin regeneration. Molecules. (2017) 22:1259. doi: 10.3390/molecules22081259
226. Xu K, Chai B, Zhang K, Xiong J, Zhu Y, Xu J, et al. Topical application of fibroblast growth factor 10-PLGA microsphere accelerates wound healing via inhibition of ER stress. Oxid Med Cell Longev. (2020) 2020:8586314. doi: 10.1155/2020/8586314
227. Zhang X, Yin M, Zhang LJ. Keratin 6, 16 and 17—critical barrier alarmin molecules in skin wounds and psoriasis. Cells. (2019) 8:807. doi: 10.3390/cells8080807
228. Krizanova O, Penesova A, Sokol J, Hokynkova A, Samadian A, Babula P. Signaling pathways in cutaneous wound healing. Front Physiol. (2022) 13:1030851. doi: 10.3389/fphys.2022.1030851
229. Konop M, Rybka M, Drapała A. Keratin biomaterials in skin wound healing, an old player in modern medicine: A mini review. Pharmaceutics. (2021) 13:2029. doi: 10.3390/pharmaceutics13122029
230. Saghazadeh S, Rinoldi C, Schot M, Kashaf SS, Sharifi F, Jalilian E, et al. Drug delivery systems and materials for wound healing applications. Adv Drug Delivery Rev. (2018) 127:138–66. doi: 10.1016/j.addr.2018.04.008
231. Janakiram NB, Valerio MS, Goldman SM, Dearth CL. The role of the inflammatory response in mediating functional recovery following composite tissue injuries. Int J Mol Sci. (2021) 22:13552. doi: 10.3390/ijms222413552
232. Johnson KE, Wilgus TA. Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair. Adv Wound Care. (2014) 3:647–61. doi: 10.1089/wound.2013.0517
233. Landén NX, Li D, Ståhle M. Transition from inflammation to proliferation: a critical step during wound healing. Cell Mol Life Sci. (2016) 73:3861–85. doi: 10.1007/s00018-016-2268-0
234. Diller RB, Tabor AJ. The role of the extracellular matrix (ECM) in wound healing: A review. Biomimetics. (2022) 7:87. doi: 10.3390/biomimetics7030087
235. Wu Y, Wu C, Ye L, Wang B, Yuan Y, Liu Y, et al. Exogenous fibroblast growth factor 1 ameliorates diabetes-induced cognitive decline via coordinately regulating PI3K/AKT signaling and PERK signaling. Cell Commun Signal. (2020) 18:81. doi: 10.1186/s12964-020-00588-9
236. Spampinato SF, Caruso GI, De Pasquale R, Sortino MA, Merlo S. The treatment of impaired wound healing in diabetes: Looking among old drugs. Pharmaceuticals. (2020) 13:60. doi: 10.3390/ph13040060
237. Dasari N, Jiang A, Skochdopole A, Chung J, Reece EM, Vorstenbosch J, et al. Updates in diabetic wound healing, inflammation, and scarring. Semin Plast Surg. (2021) 35:153–8. doi: 10.1055/s-0041-1731460
238. Ren S, Guo S, Yang L, Wang C. Effect of composite biodegradable biomaterials on wound healing in diabetes. Front Bioeng Biotechnol. (2022) 10:1060026. doi: 10.3389/fbioe.2022.1060026
239. Natarajan VT, Ganju P, Ramkumar A, Grover R, Gokhale RS. Multifaceted pathways protect human skin from UV radiation. Nat Chem Biol. (2014) 10:542–51. doi: 10.1038/nchembio.1548
240. Menon GK, Dryer L, Kalafsky R. Approaches to the development of cosmetic products to counter the effects of skin aging. Ski Aging Handb Integr Approach to Biochem Prod Dev. (2008), 265–90. doi: 10.1016/B978-0-8155-1584-5.50015-6
241. Oliveira A, Simões S, Ascenso A, Reis CP. Therapeutic advances in wound healing. J Dermatolog Treat. (2022) 33:2–22. doi: 10.1080/09546634.2020.1730296
242. Labanaris AP, Polykandriotis E, Horch RE. The effect of vacuum-assisted closure on lymph vessels in chronic wounds. J Plast Reconstr Aesthetic Surg. (2009) 62:1068–75. doi: 10.1016/j.bjps.2008.01.006
243. Panayi AC, Leavitt T, Orgill DP. Evidence based review of negative pressure wound therapy. World J Dermatol. (2017) 6:1. doi: 10.5314/wjd.v6.i1.1
244. Orgill DP, M EK, S BE, Lee RC, A CE, G GC, et al. The mechanisms of action of vacuum assisted closure: More to learn. Surgery. (2009) 146:40–51. doi: 10.1016/j.surg.2009.02.002
245. Scherer SS, Pietramaggiori G, Mathews JC, Prsa MJ, Huang S, Orgill DP. The mechanism of action of the vacuum-assisted closure device. Plast Reconstr Surg. (2008) 122:786–97. doi: 10.1097/PRS.0b013e31818237ac
246. Mattox EA. Reducing risks associated with negative-pressure wound therapy: Strategies for clinical practice. Crit Care Nurse. (2017) 37:67–77. doi: 10.4037/ccn2017308
247. Beral D, Adair R, Peckham-Cooper A, Tolan D, Botterill I. Chronic wound sepsis due to retained vacuum assisted closure foam. BMJ Br Med J (Overseas Retired Dr Ed. (2009) 339:571–3. doi: 10.1136/bmj.b2269
248. V ML, A CE, M AN, H CL, G KS. Mechanisms and clinical applications of the vacuum-assisted closure (VAC) device: A review. Am J Clin Dermatol. (2005) 6:185–94. doi: 10.2165/00128071-200506030-00005
249. Quah HM, Maw A, Young T, Hay DJ. Vacuum-assisted closure in the management of the open abdomen: a report of a case and initial experiences. J Tissue Viability. (2004) 14:59–62. doi: 10.1016/S0965-206X(04)42003-8
250. Haldipur N, Cooper B, Sanyal S. Managing the open abdomen. J R Army Med Corps. (2006) 152:143–7. doi: 10.1136/jramc-152-03-06
251. Murphy PS, Evans GRD. Advances in wound healing: A review of current wound healing products. Plast Surg Int. (2012) 2012:1–8. doi: 10.1155/2012/190436
252. H MA, A R, E KK, M TA. Oxygen and wound care: A review of current therapeutic modalities and future direction. Wound Repair Regener. (2013) 21:503–11. doi: 10.1111/wrr.12069
253. Dauwe PB, Pulikkottil BJ, Lavery L, Stuzin JM, Rohrich RJ. Does hyperbaric oxygen therapy work in facilitating acute wound healing: A systematic review. Plast Reconstr Surg. (2014) 133:208–15. doi: 10.1097/01.prs.0000436849.79161.a4
254. Gurtner GC. Hyperbaric oxygen: its mechanisms and efficacy. Yearb Plast Aesthetic Surg. (2012) 2012:228–9. doi: 10.1016/j.yprs.2011.02.040
256. Lowell D, Nicklas B, Weily W, Johnson F, Lyons MC II. Transdermal continuous oxygen therapy as an adjunct for treatment of recalcitrant and painful wounds. Foot Ankle Online J. (2009) 32:417–8. doi: 10.3827/faoj.2009.0209.0004
257. Niederauer MQ, Michalek JE, Liu Q, Papas KK, Lavery LA, Armstrong DG. Continuous diffusion of oxygen improves diabetic foot ulcer healing when compared with a placebo control: A randomised, double-blind, multicentre study. J Wound Care. (2018) 27:S30–45. doi: 10.12968/jowc.2018.27.Sup9.S30
258. Kolimi P, Narala S, Nyavanandi D, Youssef AAA, Dudhipala N. Innovative treatment strategies to accelerate wound healing: trajectory and recent advancements. Cells. (2022) 11:2439. doi: 10.3390/cells11152439
259. Wang M, Huang X, Zheng H, Tang Y, Zeng K, Shao L, et al. Nanomaterials applied in wound healing: Mechanisms, limitations and perspectives. J Control Release. (2021) 337:236–47. doi: 10.1016/j.jconrel.2021.07.017
260. Pan A, Zhong M, Wu H, Peng Y, Xia H, Tang Q, et al. Topical application of keratinocyte growth factor conjugated gold nanoparticles accelerate wound healing. Nanomedicine Nanotechnology Biol Med. (2018) 14:1619–28. doi: 10.1016/j.nano.2018.04.007
261. De La Harpe KM, Kondiah PPD, Choonara YE, Marimuthu T, Du Toit LC, Pillay V. The hemocompatibility of nanoparticles: A review of cell-nanoparticle interactions and hemostasis. Cells. (2019) 8:1209. doi: 10.3390/cells8101209
262. Shaikh S, Nazam N, Rizvi SMD, Ahmad K, Baig MH, Lee EJ, et al. Mechanistic insights into the antimicrobial actions of metallic nanoparticles and their implications for multidrug resistance. Int J Mol Sci. (2019) 20:2468. doi: 10.3390/ijms20102468
263. Cao L, Shao G, Ren F, Yang M, Nie Y, Peng Q, et al. Cerium oxide nanoparticle-loaded polyvinyl alcohol nanogels delivery for wound healing care systems on surgery. Drug Delivery. (2021) 28:390–9. doi: 10.1080/10717544.2020.1858998
264. Gainza G, Villullas S, Pedraz JL, Hernandez RM, Igartua M. Advances in drug delivery systems (DDSs) to release growth factors for wound healing and skin regeneration. Nanomedicine Nanotechnology Biol Med. (2015) 11:1551–73. doi: 10.1016/j.nano.2015.03.002
265. Xiao Y, Xu D, Song H, Shu F, Wei P, Yang X, et al. Cuprous oxide nanoparticles reduces hypertrophic scarring by inducing fibroblast apoptosis. Int J Nanomedicine. (2019) 14:5989–6000. doi: 10.2147/IJN.S196794
266. Hu K, Xiang L, Chen J, Qu H, Wan Y, Xiang D. PLGA-liposome electrospun fiber delivery of miR-145 and PDGF-BB synergistically promoted wound healing. Chem Eng J. (2021) 422:129951. doi: 10.1016/j.cej.2021.129951
267. Maghimaa M, Alharbi SA. Green synthesis of silver nanoparticles from Curcuma longa L. and coating on the cotton fabrics for antimicrobial applications and wound healing activity. J Photochem Photobiol B Biol. (2020) 204:111806. doi: 10.1016/j.jphotobiol.2020.111806
268. Diniz FR, Maia RCAP, Rannier L, Andrade LN, Chaud MV, da Silva CF, et al. Silver nanoparticles-composing alginate/gelatine hydrogel improves wound healing in vivo. Nanomaterials. (2020) 10:4071. doi: 10.3390/nano10020390
269. Nurhasni H, Cao J, Choi M, Kim I, Lee BL, Jung Y, et al. Nitric oxide-releasing poly(lactic-co-glycolic acid)-polyethylenimine nanoparticles for prolonged nitric oxide release, antibacterial efficacy, and in vivo wound healing activity. Int J Nanomedicine. (2015) 10:3065–80. doi: 10.2147/IJN.S82199
270. Vimala Devi M, Liji Sobhana SS, Shiny PJ, Ramanathan G, Grace Felciya SJ, Poornima V, et al. Durable nanofibrous matrices augmented with hydrotalcite-like compounds for cutaneous regeneration of burn wounds. Appl Clay Sci. (2020) 187:105476. doi: 10.1016/j.clay.2020.105476
271. Liu S, Fukushima K, Venkataraman S, Hedrick JL, Yang YY. Supramolecular nanofibers self-assembled from cationic small molecules derived from repurposed poly(ethylene teraphthalate) for antibiotic delivery. Nanomedicine Nanotechnology Biol Med. (2018) 14:165–72. doi: 10.1016/j.nano.2017.09.007
272. Ren X, Han Y, Wang J, Jiang Y, Yi Z, Xu H, et al. An aligned porous electrospun fibrous membrane with controlled drug delivery – An efficient strategy to accelerate diabetic wound healing with improved angiogenesis. Acta Biomater. (2018) 70:140–53. doi: 10.1016/j.actbio.2018.02.010
273. Rodríguez-Acosta H, Tapia-Rivera JM, Guerrero-Guzmán A, Hernández-Elizarraráz E, Hernández-Díaz JA, Garza-García JJO, et al. Chronic wound healing by controlled release of chitosan hydrogels loaded with silver nanoparticles and calendula extract. J Tissue Viability. (2022) 31:173–9. doi: 10.1016/j.jtv.2021.10.004
274. Chen Q, Wu J, Liu Y, Li Y, Zhang C, Qi W, et al. Electrospun chitosan/PVA/bioglass Nanofibrous membrane with spatially designed structure for accelerating chronic wound healing. Mater Sci Eng C. (2019) 105:110083. doi: 10.1016/j.msec.2019.110083
275. Yu M, Huang J, Zhu T, Lu J, Liu J, Li X, et al. Liraglutide-loaded PLGA/gelatin electrospun nanofibrous mats promote angiogenesis to accelerate diabetic wound healing: Via the modulation of miR-29b-3p. Biomater Sci. (2020) 8:4225–38. doi: 10.1039/D0BM00442A
276. Li M, Gao L, Chen J, Zhang Y, Wang J, Lu X, et al. Controllable release of interleukin-4 in double-layer sol-gel coatings on TiO 2 nanotubes for modulating macrophage polarization. BioMed Mater. (2018) 13:045008. doi: 10.1088/1748-605X/aa9526
277. Tang T, Jiang H, Yu Y, He F, Ji SZ, Liu YY, et al. A new method of wound treatment: Targeted therapy of skin wounds with reactive oxygen species-responsive nanoparticles containing SDF-1α. Int J Nanomedicine. (2015) 10:6571–85. doi: 10.2147/IJN.S88384
278. Manuja A, Raguvaran R, Kumar B, Kalia A, Tripathi BN. Accelerated healing of full thickness excised skin wound in rabbits using single application of alginate/acacia based nanocomposites of ZnO nanoparticles. Int J Biol Macromol. (2020) 155:823–33. doi: 10.1016/j.ijbiomac.2020.03.221
279. Chen WY, Chang HY, Lu JK, Huang YC, Harroun SG, Tseng YT, et al. Self-assembly of antimicrobial peptides on gold nanodots: against multidrug-resistant bacteria and wound-healing application. Adv Funct Mater. (2015) 25:7189–99. doi: 10.1002/ADFM.201503248
280. Mirzahosseinipour M, Khorsandi K, Hosseinzadeh R, Ghazaeian M, Shahidi FK. Antimicrobial photodynamic and wound healing activity of curcumin encapsulated in silica nanoparticles. Photodiagnosis Photodyn Ther. (2020) 29:101639. doi: 10.1016/j.pdpdt.2019.101639
281. Sun M, Deng Z, Shi F, Zhou Z, Jiang C, Xu Z, et al. Rebamipide-loaded chitosan nanoparticles accelerate prostatic wound healing by inhibiting M1 macrophage-mediated inflammation: Via the NF-κB signaling pathway. Biomater Sci. (2020) 8:912–25. doi: 10.1039/C9BM01512D
282. Xu HL, Chen PP, ZhuGe DL, Zhu QY, Jin BH, Shen BX, et al. Liposomes with silk fibroin hydrogel core to stabilize bFGF and promote the wound healing of mice with deep second-degree scald. Adv Healthc Mater. (2017) 6:1700344. doi: 10.1002/adhm.201700344
283. Schepler H, Neufurth M, Wang S, She Z, Schröder HC, Wang X, et al. Acceleration of chronic wound healing by bio-inorganic polyphosphate: In vitro studies and first clinical applications. Theranostics. (2022) 27:18–34. doi: 10.7150/THNO.67148
284. Pramanik N, Bhattacharya S, Rath T, De J, Adhikary A, Basu RK, et al. Polyhydroxybutyrate-co-hydroxyvalerate copolymer modified graphite oxide based 3D scaffold for tissue engineering application. Mater Sci Eng C. (2019) 94:534–46. doi: 10.1016/j.msec.2018.10.009
285. Perez-Amodio S, Rubio N, Vila OF, Navarro-Requena C, Castaño O, Sanchez-Ferrero A, et al. Polymeric composite dressings containing calcium-releasing nanoparticles accelerate wound healing in diabetic mice. Adv Wound Care. (2021) 10:301–16. doi: 10.1089/wound.2020.1206
286. Jirofti N, Golandi M, Movaffagh J, Ahmadi FS, Kalalinia F. Improvement of the wound-healing process by curcumin-loaded chitosan/collagen blend electrospun nanofibers: in vitro and in vivo studies. ACS Biomater Sci Eng. (2021) 7:3886–97. doi: 10.1021/acsbiomaterials.1c00131
287. Fumakia M, Ho EA. Nanoparticles encapsulated with LL37 and serpin A1 promotes wound healing and synergistically enhances antibacterial activity. Mol Pharm. (2016) 13:2318–31. doi: 10.1021/acs.molpharmaceut.6b00099
288. Deng X, Li X, Chen W, Zhao T, Huang W, Qian H. Design, synthesis and biological evaluation of peptide dendrimers with wound healing promoting activity. Med Chem Res. (2017) 26:580–6. doi: 10.1007/s00044-016-1777-6
289. Ma Y, Duan L, Sun J, Gou S, Chen F, Liang Y, et al. Oral nanotherapeutics based on Antheraea pernyi silk fibroin for synergistic treatment of ulcerative colitis. Biomaterials. (2022) 282:121410. doi: 10.1016/j.biomaterials.2022.121410
290. Muhamed I, Sproul EP, Ligler FS, Brown AC. Fibrin nanoparticles coupled with keratinocyte growth factor enhance the dermal wound-healing rate. ACS Appl Mater Interfaces. (2019) 11:3771–80. doi: 10.1021/acsami.8b21056
291. Mohammadi Z, Sharif Zak M, Majdi H, Mostafavi E, Barati M, Lotfimehr H, et al. The effect of chrysin–curcumin-loaded nanofibres on the wound-healing process in male rats. Artif Cells Nanomedicine Biotechnol. (2019) 47:1642–52. doi: 10.1080/21691401.2019.1594855
292. Yolanda M-M. Adult stem cell therapy in chronic wound healing. J Stem Cell Res Ther. (2014) 04:1000162. doi: 10.4172/2157-7633.1000162
293. Strioga M, Viswanathan S, Darinskas A, Slaby O, Michalek J. Same or not the same? comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. (2012) 21:2724–52. doi: 10.1089/scd.2011.0722
294. Salem HK, Thiemermann C. Mesenchymal stromal cells: Current understanding and clinical status. Stem Cells. (2010) 28:585–96. doi: 10.1002/stem.269
295. Kølle SFT, Fischer-Nielsen A, Mathiasen AB, Elberg JJ, Oliveri RS, Glovinski PV, et al. Enrichment of autologous fat grafts with ex-vivo expanded adipose tissue-derived stem cells for graft survival: A randomised placebo-controlled trial. Lancet. (2013) 382:1113–20. doi: 10.1016/S0140-6736(13)61410-5
296. Tsai HW, Wang PH, Tsui KH. Mesenchymal stem cell in wound healing and regeneration. J Chin Med Assoc. (2018) 81:223–4. doi: 10.1016/j.jcma.2017.06.011
297. Duscher D, Barrera J, Wong VW, Maan ZN, Whittam AJ, Januszyk M, et al. Stem cells in wound healing: the future of regenerative medicine? A mini-review. Gerontology. (2016) 62:216–25. doi: 10.1159/000381877
298. Mousaei Ghasroldasht M, Seok J, Park HS, Liakath Ali FB, Al-Hendy A. Stem cell therapy: from idea to clinical practice. Int J Mol Sci. (2022) 23:2850. doi: 10.3390/ijms23052850
299. Dash BC, Xu Z, Lin L, Koo A, Ndon S, Berthiaume F, et al. Stem cells and engineered scaffolds for regenerative wound healing. Bioengineering. (2018) 5:23. doi: 10.3390/bioengineering5010023
300. Mazini L, Rochette L, Admou B, Amal S, Malka G. Hopes and limits of adipose-derived stem cells (ADSCs) and mesenchymal stem cells (MSCs) in wound healing. Int J Mol Sci. (2020) 21:1306. doi: 10.3390/ijms21041306
301. Xu Y, Huang S, Fu X. Autologous transplantation of bone marrow-derived mesenchymal stem cells: A promising therapeutic strategy for prevention of skin-graft contraction. Clin Exp Dermatol. (2012) 37:497–500. doi: 10.1111/ced.2012.37.issue-5
302. Aboulhoda BE, Abd el Fattah S. Bone marrow-derived versus adipose-derived stem cells in wound healing: value and route of administration. Cell Tissue Res. (2018) 374:285–302. doi: 10.1007/s00441-018-2879-x
303. O’Brien FJ. Biomaterials & scaffolds for tissue engineering. Mater Today. (2011) 14:88–95. doi: 10.1016/S1369-7021(11)70058-X
304. Kocan B, Maziarz A, Tabarkiewicz J, Ochiya T, Banaś-Ząbczyk A. Trophic activity and phenotype of adipose tissue-derived mesenchymal stem cells as a background of their regenerative potential. Stem Cells Int. (2017) 2017:1653254. doi: 10.1155/2017/1653254
305. Jung JA, Yoon YD, Lee HW, Kang SR, Han SK. Comparison of human umbilical cord blood-derived mesenchymal stem cells with healthy fibroblasts on wound-healing activity of diabetic fibroblasts. Int Wound J. (2018) 15:133–9. doi: 10.1111/iwj.12849
306. Shrestha C, Zhao L, Chen K, He H, Mo Z. Enhanced healing of diabetic wounds by subcutaneous administration of human umbilical cord derived stem cells and their conditioned media. Int J Endocrinol. (2013) 2013:592454. doi: 10.1155/2013/592454
307. Afzali L, Mirahmadi-Babaheydari F, Shojaei-Ghahrizjani F, Rahmati S, Shahmoradi B, Banitalebi-Dehkordi M. The effect of encapsulated umbilical cord-derived mesenchymal stem cells in PRPCryogel on regeneration of grade-II burn wounds. Regener Eng Transl Med. (2022) 8:75–85. doi: 10.1007/s40883-020-00188-6
308. Nazempour M, Mehrabani D, Mehdinavaz-Aghdam R, Hashemi SS, Derakhshanfar A, Zare S, et al. The effect of allogenic human Wharton’s jelly stem cells seeded onto acellular dermal matrix in healing of rat burn wounds. J Cosmet Dermatol. (2020) 19:995–1001. doi: 10.1111/jocd.13109
309. Kim HS, Sun X, Lee JH, Kim HW, Fu X, Leong KW. Advanced drug delivery systems and artificial skin grafts for skin wound healing. Adv Drug Delivery Rev. (2019) 146:209–39. doi: 10.1016/j.addr.2018.12.014
310. Trounson A, McDonald C. Stem cell therapies in clinical trials: progress and challenges. Cell Stem Cell. (2015) 17:11–22. doi: 10.1016/j.stem.2015.06.007
311. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. (2007) 131:861–72. doi: 10.1016/j.cell.2007.11.019
312. Mofazzal Jahromi MA, Sahandi Zangabad P, Moosavi Basri SM, Sahandi Zangabad K, Ghamarypour A, Aref AR, et al. Nanomedicine and advanced technologies for burns: Preventing infection and facilitating wound healing. Adv Drug Delivery Rev. (2018) 123:33–64. doi: 10.1016/j.addr.2017.08.001
313. Hashempour S, Ghanbarzadeh S, Maibach HI, Ghorbani M, Hamishehkar H. Skin toxicity of topically applied nanoparticles. Ther Delivery. (2019) 10:383–96. doi: 10.4155/tde-2018-0060
314. Kang H, Buchman JT, Rodriguez RS, Ring HL, He J, Bantz KC, et al. Stabilization of silver and gold nanoparticles: preservation and improvement of plasmonic functionalities. Chem Rev. (2019) 119:664–99. doi: 10.1021/acs.chemrev.8b00341
315. Xu L, Chu Z, Wang H, Cai L, Tu Z, Liu H, et al. Electrostatically assembled multilayered films of biopolymer enhanced nanocapsules for on-demand drug release. ACS Appl Bio Mater. (2019) 2:3429–38. doi: 10.1021/acsabm.9b00381
316. Chen G, Chen Z, Wen D, Wang Z, Li H, Zeng Y, et al. Transdermal cold atmospheric plasma-mediated immune checkpoint blockade therapy. Proc Natl Acad Sci U.S.A. (2020) 117:3687–92. doi: 10.1073/pnas.1917891117
317. Dukhinova MS, Prilepskii AY, Vinogradov VV, Shtil AA. Metal oxide nanoparticles in therapeutic regulation of macrophage functions. Nanomaterials. (2019) 9:1631. doi: 10.3390/nano9111631
318. Ma Z, Li S, Wang H, Cheng W, Li Y, Pan L, et al. Advanced electronic skin devices for healthcare applications. J Mater Chem B. (2019) 7:173–97. doi: 10.1039/C8TB02862A
319. Okano H, Nakamura M, Yoshida K, Okada Y, Tsuji O, Nori S, et al. Steps toward safe cell therapy using induced pluripotent stem cells. Circ Res. (2013) 112:523–33. doi: 10.1161/CIRCRESAHA.111.256149
Keywords: skin, skin wound healing, immune cell-wound healing, wound physical therapy, stem cell therapy
Citation: Mamun AA, Shao C, Geng P, Wang S and Xiao J (2024) Recent advances in molecular mechanisms of skin wound healing and its treatments. Front. Immunol. 15:1395479. doi: 10.3389/fimmu.2024.1395479
Received: 04 March 2024; Accepted: 03 May 2024;
Published: 21 May 2024.
Edited by:
Ronghua Yang, Guangzhou First People’s Hospital, ChinaReviewed by:
Biao Cheng, General Hospital of Southern Theater Command of PLA, ChinaCopyright © 2024 Mamun, Shao, Geng, Wang and Xiao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jian Xiao, eGZ4ajIwMDBAMTI2LmNvbQ==; Shuanghu Wang, d2FuZ3NodWFuZ2h1QGxzdS5lZHUuY24=
†ORCID: Abdullah Al Mamun, orcid.org/0000-0001-9936-3925
Peiwu Geng, orcid.org/0000-0002-9714-7660
Chuxiao Shao, orcid.org/0000-0002-7154-1723
Jian Xiao, orcid.org/0000-0001-7374-6506
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.