
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol., 19 January 2024
Sec. T Cell Biology
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1344078
 Antonio Bensussen1*
Antonio Bensussen1* José Antonio Torres-Magallanes1
José Antonio Torres-Magallanes1 Elena R. Álvarez-Buylla2,3*Elena Roces de Álvarez-Buylla1*
Elena R. Álvarez-Buylla2,3*Elena Roces de Álvarez-Buylla1*CD4+ T lymphocytes have been classified into several lineages, according to their gene expression profiles and their effector responses. Interestingly, recent evidence is showing that many lineages could yield hybrid phenotypes with unique properties and functions. It has been reported that such hybrid lineages might underlie pathologies or may function as effector cells with protection capacities against molecular threats. In this work, we reviewed the characteristics of the hybrid lineages reported in the literature, in order to identify the expression profiles that characterize them and the markers that could be used to identify them. We also review the differentiation cues that elicit their hybrid origin and what is known about their physiological roles.
Helper T lymphocytes (Th), also known as CD4+ T cells, are considered fundamental components of the immune system. In response to antigen presenting cells (APC), CD4+ T lymphocytes secrete a set of specific cytokines to coordinate immune responses. The cytokine signature is sustained by specific gene expression programs, that allows CD4+ T cells to be classified in particular subsets, that, in turn underlie their responses to certain types of threats and determined physiological functions (1). Two main CD4+ T cells groups have been distinguished: the regulatory cells (Treg) and the effector cells (Teff). Regarding to canonical effector lineages, they are divided in Th1, Th2, Th9, Th17, Th22 and TFH (T follicular lymphocytes) subsets, which can be identified as T-bet+ IFN-γ+ (Th1), GATA3+ IL-4+ (Th2), PU.1+ IL-9+ (Th9), RORγT+ IL-17+ (Th17), AHR+ IL-22+ (Th22) and BCL6+ IL-21+ (TFH) (Figure 1A). On the other hand, regulatory subsets are generally identified by expressing high levels of transcriptional factor FoxP3, as well as cytokines like TGF-β and IL-10 (Figure 1A). Each phenotype has a specific physiological role. For instance, Th1 is specialized in promoting an inflammatory response in response to viral infections, intracellular bacterial infections, neoplasms, or intracellular deregulations. In contrast, other subsets such as the iTreg downregulate these effector functions and are responsible for immune resolution and subsequent restoration of physiological homeostasis (2). It has been proposed that there are specific Treg lineages for every single CD4+ T effector subset (Figure 1A). The latter implies that all T helper cells can be specifically modulated (3).
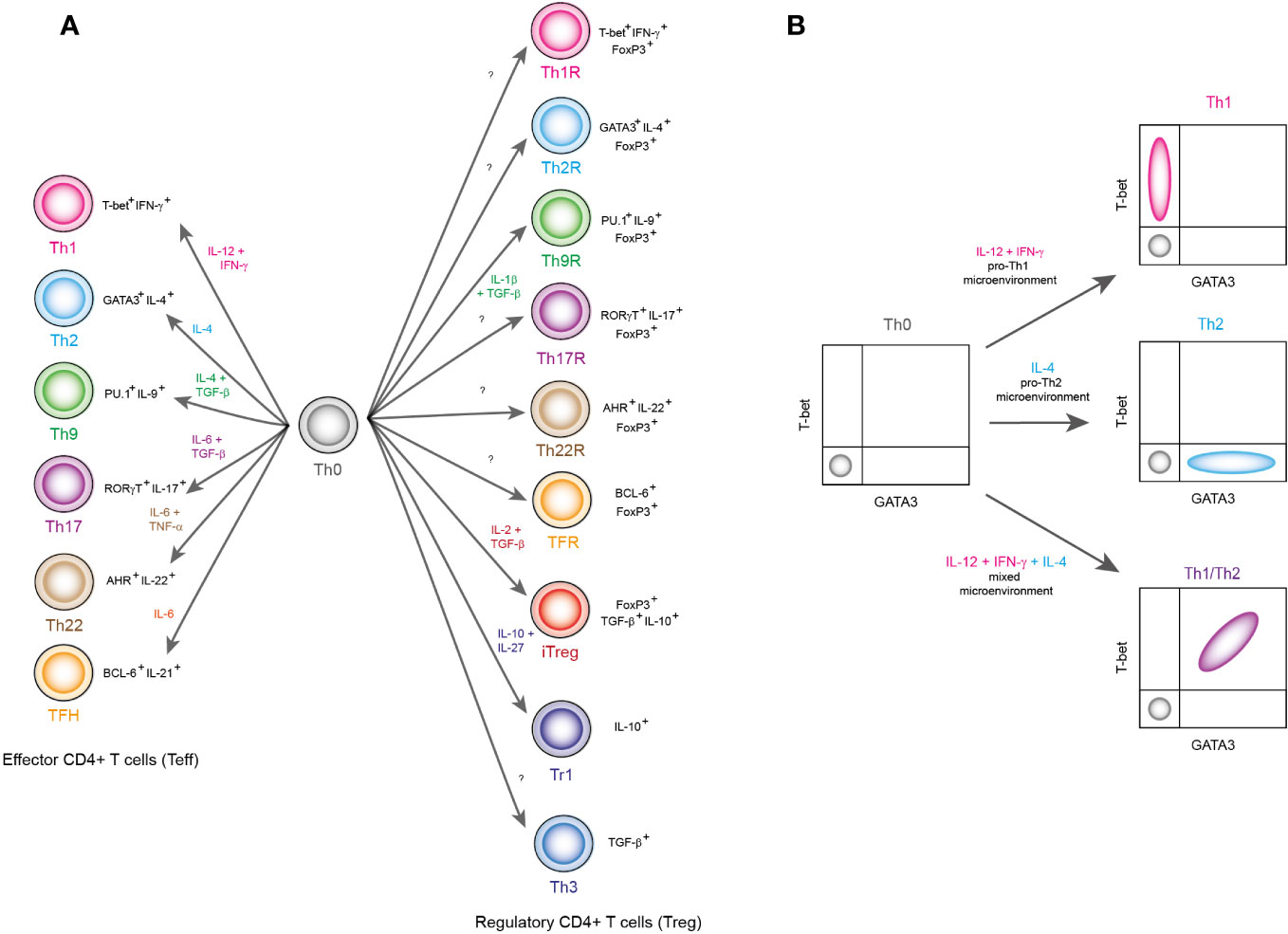
Figure 1 Classification of CD4+ T lymphocyte subpopulations. (A) Lymphocyte subpopulations are classified based on the transcriptional factors and cytokines they express. In this panel, differentiation environment is reported with colored labels as well as the name of each lineage. Also, the signature transcription factors and cytokines are shown in black labels. (B) This figure shows a schematic representation of flow cytometry results that show the emergence of hybrid lineages, which are outside the pure definitions of classical phenotypes Th1 and Th2. Here, the combination of the cytokines IFN-γ, IL-12 and IL-4 produces a population of T-bet+ GATA3+ hybrid cells, which correspond to a Th1/Th2 hybrid lineage.
As data accumulate, it has been shown that such a strict categorical approach is limited. For instance, when studying the ratio of Th1 (T-bet+) versus Th2 (GATA3+) cells under different conditions (4) (Figure 1B), there are scenarios in which both groups prevail, producing T-bet+ GATA3+ (Th1/Th2) double positive cells (Figure 1B). This hybrid subpopulation is able to maintain its phenotype in the form of memory cells and resist cell reprogramming (4), but it has been generally ignored because they have been found at low frequencies. Nevertheless, it is not clear whether these cells have a low populational frequency because in nature there are very few scenarios that elicit their differentiation, or they are rare because the conditions that are used in the laboratory are rare in nature. The differentiation processes that have been described under laboratory conditions are indeed relevant. But in order to understand the conditions that elicit hybrid lineages and their biological role, a wider range of physiological should be studied. Here, we review the physiological effects of CD4+ T cell hybrid lineages and what is currently known about them.
These hybrid lymphocytes are characterized by being T-bet+ GATA3+ cells, as well as by expressing Th1 and Th2 cytokines. These cells were first characterized in mice infected by the helminths S. mansoni and H. polygyrus (4), but they also appeared in infections caused by the nematode S. ratti. These hybrids have been characterized in humans due to infections of the nematode S. stercoralis (5). Regarding its differentiation, Th1/Th2 hybrids appear due to the combination of IL-12, IL-4 and IFN-γ (4) (Figure 1B). In fact, they may present heterogeneous expression levels of T-bet and GATA3 due to variations on IFN-γ levels (4). Interestingly, these cells produce memory cells that are not susceptible to being reprogrammed after differentiation (4). Furthermore, in humans it was found that they present a predominantly Th1 profile (5) but in mice it is known that they maintain a balance between Th1 and Th2 (4). It is thought that at a physiological level, these cells act as immunomodulators without having to resort to immunosuppression, since they prevent the exacerbation of the Th2 response, which could potentially protect against allergies (4).
This cell lineage can be identified as T-bet+ IL-9+ or alternatively as IFN-γ+ IL-9+, and it was first observed in humans as a byproduct of stimulation of Th17 cells with TGF-β (6). Years later, the Th1/Th9 hybrid was observed again in mice (7). Conventionally, it has been accepted that the way to induce Th9 polarization is using the combination of IL-4 plus TGF-β. However, when testing the combination IL-4 plus IL-1β, Th9 cells appeared that were T-bet+ IL-9+ (7). In addition, it was proven that they had strong antitumor activity when tested against the B16 melanoma cell line (7).
This population of cells can be identified as T-bet+ RORγT+ or alternatively as IFN-γ+ IL-17A+. This lineage was first characterized in humans in the context of multiple sclerosis, and various studies have shown that it has a pathogenic effect (8). In addition, they have also been associated with severe orbitopathy of graves (9), type 1 diabetes, autoimmune uveitis and dry disease, intestine of Crohn’s disease, Sjogren’s disease, psoriasis (8) and cancer (10). Contradictorily, there are studies that suggest that these hybrids could have therapeutic effects. In mice, it has been reported that these cells present strong antitumor activity (11) and in humans, it has been seen that they promote recovery from sarcoidosis (12). Likewise, in observations in a murine model of colitis it was found that these hybrids can have a protective effect, since by hindering this hybrid population by deleting T-bet, the expression of IL-17 and IL-22 is triggered, drastically increasing the pathogenicity of inflammation of the intestine (13). Molecularly, Th1/Th17 hybrids are highly proliferative in response to stimulation by the TCR, they also increase their expression of GLUT1 and they can be stimulated by the action of IL-1β (8). Regarding their differentiation, in mice it was possible to obtain them in environments with IL-6, IL-23, IL-1β, IL-12 and low levels of TGF-β (11). Ex vivo observations show that in mice, IL-23 is essential to obtain these hybrids without affecting the differentiation of the Th1 and Th17 base phenotypes (13). On the other hand, in humans it is known that the secretion of IL-21 by central memory cells prevents the appearance of Th1/Th17 hybrids (13).
This lineage is identifiable as T-bet+ AhR+ IL-22+, or alternatively as IFN-γ+ IL-22+. These cells were first found as a byproduct of murine Th22 lymphocyte differentiation in response to C. rodentium infection (14). Regarding their differentiation conditions, in mice it is known that Th22 cells obtained by the combination of cytokines TNF-α and IL-6 in an inflammatory context rich in IL-12, can transition to pure Th1 and Th1/Th22 hybrids (15). At the genetic level, experimental evidence suggests that this hybrid originates from a balance between the mutually exclusive regulation of AhR and T-bet (15), since the absence of any of these transcriptional factors hinders the expression of IL-22 (15). In humans, it has been found that this population appears before the worsening of multiple sclerosis begins (16). Furthermore, it was found that this cell lineage has autoreactive properties, and is insensitive to treatment with IFN-β, consolidating itself as a fundamental factor to hinder the treatment of multiple sclerosis (16).
Th1/TFH hybrids are characterized as T-bet+ BCL6+ CXCR5+, although they can also be labeled as T-bet+ IL-6+ or IFN-γ+ IL-21+ or IFN-γ+ BCL6+. These cells were characterized in mice infected with P. chabaudi (17, 18). Concerning to differentiation, these cells appear in environments rich in IFN-γ and IL-27, the latter being responsible for biasing differentiation towards the TFH side (17). Interestingly, it was found that in response to prolonged P. chabaudi infections, Th1/TFH hybrids tend to be biased towards the TFH side, increasing the stimulation of antibody production (17). However, brief infections bias differentiation towards the Th1 side, which favors the Th1/TFH memory being able to protect against reinfection by P. chabaudi. This suggests that the emergence of the Th1/TFH hybrid population could have beneficial effects in the short term, but may represent a subsequent risk (18). Additionally, these cells were found in human nasal polyps. In humans, this lineage appears through the action of IL-12; and these hybrids were found to be responsible for the inflammation of nasal polyps (19). Counterintuitively, it is suggested that the role of this Th1/TFH population is protective, since differentiation towards the TFH pole protects from damage produced by an enhanced Th1 response (19).
Th2/Th9 hybrids identifiable as IL-4+ IL-9+ or alternatively GATA3+ PU.1+, were observed in patients as a product of mutations in STAT1 or STAT3. It is speculated that these hybrids could make patients prone to contracting candidiasis, cancer and sinopulmonary infection (20). Similarly, these hybrid cells were observed at low frequencies in E. granulosus infection (21). In mice, these hybrids were observed as a byproduct of papain-induced airway inflammation (22). However, the conditions of its differentiation and its exact physiological function are unknown.
This hybrid population can be identified as GATA3+ RORγT+; or alternatively, as IL-4+ IL-17A+ or GATA3+ CD161+. Th2/Th17 hybrids were found in patients with asthma, and have the ability to induce IgE secretion (23). Interestingly, it was not possible to obtain these hybrids under in vitro conditions, but CCR6+ CD161+ memory cells produced these hybrids in IL-4-rich environments, suggesting that the Th2/Th17 population could appear in this context (23). Furthermore, this population was also found in a murine model of asthma, and it was found that they had the ability to exacerbate the inflammation produced in asthma (24). Years later it was found that in the context of asthma in mice, these hybrids could differentiate based on the activity of dendritic cells when repeatedly exposed to pro-inflammatory treatments with ovalbumin (25). Memory cells with Th2/Th17 phenotype were recently found, and they were linked to the appearance of Palmoplantar pustulosis (26). Regarding differentiation of these hybrids in mice, Th2/Th17 cells were detected in low quantities as byproducts of an unconventional pro-Th17 differentiation pathway, in which the combination of IL-9 plus TGF-β was used (27).
These hybrids have been identified as IL-9+ IL-17+. Experimentally, it has been seen that Th9/Th17 hybrids appear due to the joint stimulation of the cytokines TGF-β, IL-1β, IL-6, IL-21 and IL-23 (6). Alternatively, it was also found that they could differentiate in the presence of the combination of cytokines TGF-β plus IL-12, or TGF-β plus IL-1β, or by the classic Th9 differentiation conditions, TGF-β plus IL-4 (6). Although, they have also appeared due to the combination of cytokines TGF-β, IL-4, IL-1β and IL-23 (28). This lineage was first characterized in patients with autoimmune diabetes; being significantly more frequent than what is observed in healthy people (6). Later these cells were found again in both humans and mice, in the context of autoimmune gastritis (28). However, trials showed that the decrease in the Th9/Th17 hybrid population severely worsened the development of the disease (28). Therefore, it is suggested that these cells could have a protective effect against damage caused by the Th17 response (28).
This hybrid line, defined by the presence of the IL-9+ BCL6+ markers, should ideally not exist. This is because Th9 differentiation conditions inhibit BCL6 activity, yet a tiny fraction of this hybrid lineage was found in mice (29). Experimental data suggest that IL-2 in the presence of a classical pro-Th9 environment could stabilize this hybrid population, albeit at low levels (29). It is not known if it has any specific physiological function.
Th17/Th22 hybrids can be identified as IL-17+ IL-22+ or alternatively as RORγT+ BCL6+. Originally, it was thought that these cells were pathogenic, since in murine models of colitis, increasing their frequency triggers the severity of intestinal inflammation (13). However, recent evidence suggests that these cells play an important role in the regulation of intestinal homeostasis; since in rhesus macaques infected with simian immunodeficiency virus (SIV), decreasing the frequency of these hybrids, enhanced the severity of SIV infection (30). There are no reports on the conditions that promote their differentiation, but it was observed that the presence of IL-23 increases the differentiation of these hybrids (13). Additionally, it has been reported that the absence of T-bet drastically increases its amount (31).
Recently, experimental evidence was found for the existence of a hybrid Th1/Th2-like regulatory lineage in humans (32). This lineage was identified as T-bet+ GATA3+ FoxP3+, and its prevalence was observed to decrease in patients with cardiovascular disease (32). The differentiation conditions of this phenotype are unknown, and it is thought that it may have a protective effect against cardiovascular disease.
In humans, a regulatory hybrid Th1/Th17-like lineage was found; labeled as T-bet+ RORγT+ FoxP3+ (33). This lineage was found as part of memory cells and was seen to have a high proliferative capacity compared to other regulatory cells. At the systemic level, these hybrids were predominantly observed in the colon. Interestingly, in the context of cancer, a significant decrease in this cell population was observed (33). Furthermore, it was observed that in the context of cancer, Th2-like regulatory cells (GATA3+ FoxP3+) are attracted to the tumor, and inhibit regulatory Th1/Th17-like cells, so it is thought that they may have a protective role against the tumors (33). The conditions that favor their differentiation in vitro are unknown.
In addition to the limitations that hybrid lineages represent for the conventional classification system, there are effector cells whose classification is impossible. For example, effector cells were found in mice, whose phenotypes were IL-2+, TNF+, IL-2+ TNF+ IL- 22+ IL-5+, GM-CSF+ IL-5+ IL-13+ and IL-2+ IFN-γ+ TNF+ IL-22+ IL-5+ (34). In addition, it was also shown that even the traditionally defined lineages might present notable variations, for example, Th1 cells identified as IFN-γ+, and IFN-γ+ TNF+ GM-CSF+ IL-22+ have been described (34). Physiologically, these polyfunctional lymphocytes, i.e., CD4+ T cells that express more than one biologically active cytokine, have been found to have completely different functions from each other (35). Indeed, polyfunctional Th1 lymphocytes identified as IFN-γ+ TNF-α+, IL-2+ provide effective protection against COVID-19 (36), unlike conventional Th1 lymphocytes (IFN-γ+). Interestingly, Th17/Th22 hybrid cells were found to be polyfunctional in healthy Rhesus macaques, because they express up to four additional cytokines (30). In the presence of SIV, not only did the Th17/Th22 population decrease, but polyfunctional hybrid cells also decreased in frequency (30). Furthermore, increments of these cells have been found to be correlated with the reduction of SIV viremia levels, thus suggesting their role in health (30).
Traditionally, CD4+ T lymphocytes were thought to have clearly distinguishable phenotypes, with specific functions and regulatory pathways. Nowadays, several experimental results suggest the opposite. In fact, the evidence shows that the differentiation of CD4+ T lymphocytes occurs in the form of a gradient, which implies that hybrid subpopulations can be limitless (Table 1). The open question is to understand how such lineages emerge. The answer requires dealing with the complexity of the microenvironments in which the differentiation of CD4+ T cells occurs, including intracellular alterations such as epigenetic changes. In this sense, systems biology allows to visualize the effect of intrinsic and extrinsic variables that impact the differentiation of CD4+ T cells, in conditions that are not feasible to achieve experimentally. For example, by using multistable models, it has been possible to reproduce the molecular mechanisms that originate all differentiation patterns of the canonical lineages of CD4+ T lymphocytes (37). In addition, these models predicted that stochastic intracellular perturbations in the signaling and the expression of signature transcription factors caused the emergence of non-canonical lineages (37).
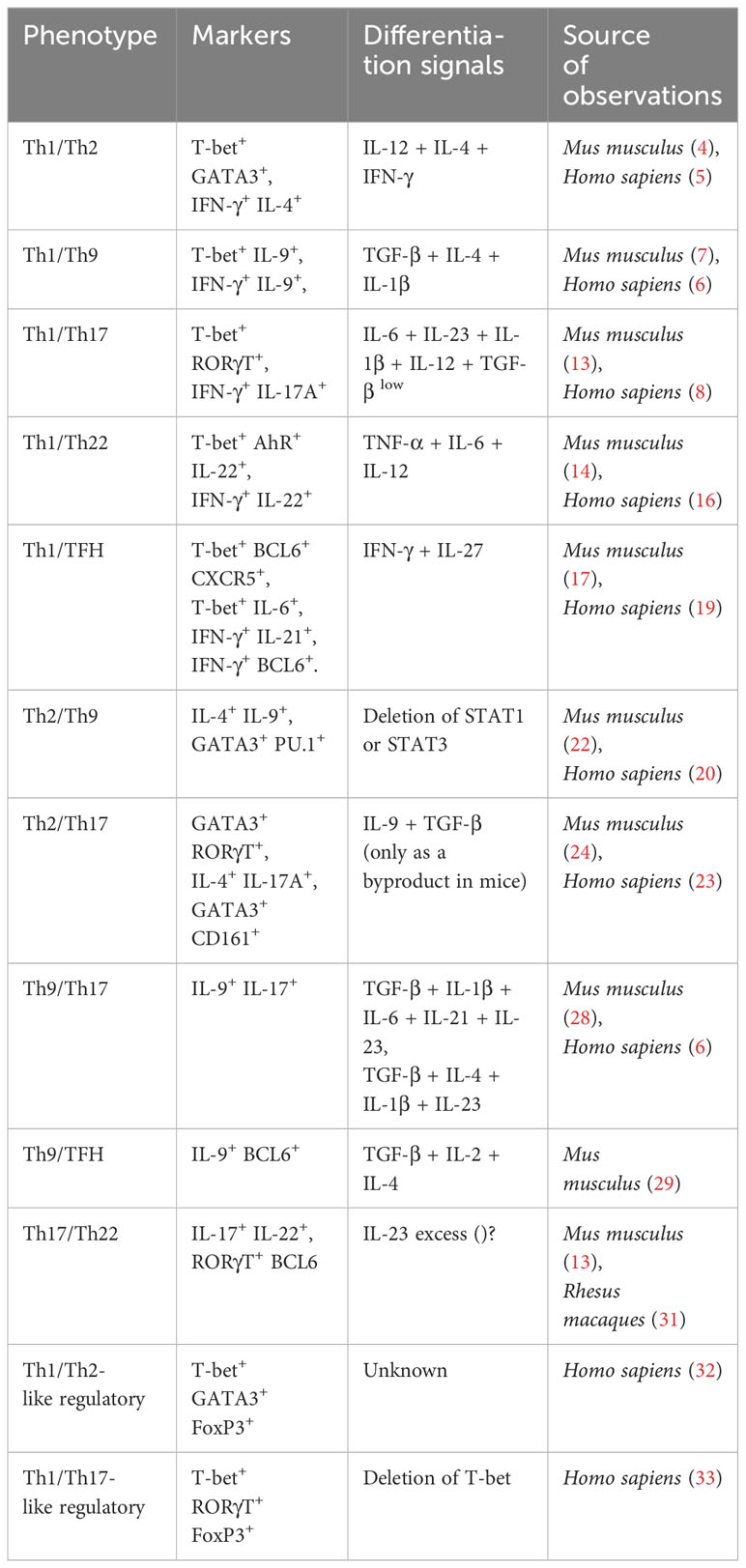
Table 1 Hybrid lineages of CD4+ T cells.
Moreover, continuous approaches of multistable models showed that the majority of Th phenotypes follow bimodal dynamics, but depending on the levels of polarizing cytokine, non-canonical phenotypes may emerge (38). Indeed, the model predicts that microenvironments mixed with IL-4+ and IFN-γ+ affect cell differentiation depending on their concentration. To be precise, high levels of exocrine IFN-γ or IL-4 produces either Th1 or Th2 cells that are not plastic, which means that even in presence of the contrary cytokine, they maintain their phenotype (38). On the contrary, low levels of exocrine IFN-γ or IL-4 generates plastic Th1 or Th2 phenotypes (38). Molecularly, the model predicts that Th2 lineage requires to produce autocrine IL-4 to maintain its phenotype. But if the autocrine production of this cytokine is reduced, then it may appear Th1/Th2 hybrids (38), which implies that plasticity is governed by the microenvironment and the intracellular state. Concerning to the effect of other external influences, multistable models reveled that, systemic hormones such as insulin (39) or local bioactive signals like adipokines (40) are able to bias CD4+ T cell differentiation towards inflammatory phenotypes, reducing the presence of regulatory lineages even in anti-inflammatory microenvironments.
Recently, independent studies have demonstrated these predictions. Indeed, inhibiting the enzyme ornithine decarboxylase increased epigenomic alterations in histone acetylation, which augments transcriptional noise as well as the frequency of hybrid lineages (41). Similarly, it has been reported that the majority of genes responsible for the emergence of Th1 and Th2, present bimodal dynamics, and only a fraction of them present mixed behaviors (42). Furthermore, it has been demonstrated that high exposition to polarizing cytokines IL-12 and IFN-γ during the infection of P. chabaudi, reduces the frequency of Th1/TFH hybrids (18). On the contrary, low exposition to such cytokines, increases the hybrid subset (18). Perhaps, these mechanisms could be extrapolated to other immune cells, like CD8+ T lymphocytes. In fact, Tc1/Tc17 lymphocytes (identified as IFN-γ+ IL-17+ CD8+) have been described as important downregulators of encephalitis (43). In conclusion, the study of hybrid lineages of CD4+ T lymphocytes is important to further understand the complex dynamics of immune systems and their plastic responses under various cues. Understanding such complexity will necessarily require the use of formal and computer modeling approaches.
AB: Conceptualization, Data curation, Investigation, Visualization, Writing – original draft. JT-M: Conceptualization, Data curation, Investigation, Visualization, Writing – original draft. EÁ-B: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing. EdA-B: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by research grant (project CF 2019 2096023) from Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT) in Mexico City.
We thank to Dr. Oxana Dobrovinskaya, Universidad de Colima, for her comments. AB thanks to CONAHCYT for his postdoctoral fellowship. JT-M thanks to CONAHCYT for his doctoral fellowship.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Tay RE, Richardson EK, Toh HC. Revisiting the role of CD4+ T cells in cancer immunotherapy—new insights into old paradigms. Cancer Gene Ther (2021) 28(1):5–17. doi: 10.1038/s41417-020-0183-x
2. Negi V, Paul D, Das S, Bajpai P, Singh S, Mukhopadhyay A, et al. Altered expression and editing of miRNA-100 regulates iTreg differentiation. Nucleic Acids Res (2015) 43(16):8057–65. doi: 10.1093/nar/gkv752
3. Cretney E, Kallies A, Nutt SL. Differentiation and function of Foxp3+ effector regulatory T cells. Trends Immunol (2013) 34:74–80. doi: 10.1016/j.it.2012.11.002
4. Peine M, Rausch S, Helmstetter C, Fröhlich A, Hegazy AN, Kühl AA, et al. Stable T-bet+GATA-3+ Th1/Th2 hybrid cells arise in vivo, can develop directly from naive precursors, and limit immunopathologic inflammation. PloS Biol (2013) 11(8):1–17. doi: 10.1371/journal.pbio.1001633
5. Bock CN, Babu S, Breloer M, Rajamanickam A, Boothra Y, Brunn ML, et al. Th2/1 hybrid cells occurring in murine and human Strongyloidiasis share effector functions of Th1 cells. Front Cell Infect Microbiol (2017) 7(261):1–15. doi: 10.3389/fcimb.2017.00261
6. Beriou G, Bradshaw EM, Lozano E, Costantino CM, Hastings WD, Orban T, et al. TGF-β Induces IL-9 production from human Th17 cells. J Immunol (2010) 185(1):46–54. doi: 10.4049/jimmunol.1000356
7. Xue G, Jin G, Fang J, Lu Y. IL-4 together with IL-1β induces antitumor Th9 cell differentiation in the absence of TGF-β signaling. Nat Commun (2019) 10(1):1–10. doi: 10.1038/s41467-019-09401-9
8. Stadhouders R, Lubberts E, Hendriks RW. A cellular and molecular view of T helper 17 cell plasticity in autoimmunity. J Autoimmun (2018) 87:1–15. doi: 10.1016/j.jaut.2017.12.007
9. Fang S, Zhang S, Huang Y, Wu Y, Lu Y, Zhong S, et al. Evidence for associations between Th1/Th17 “hybrid” phenotype and altered lipometabolism in very severe graves orbitopathy. J Clin Endocrinol Metab (2020) 105(6):1851–67. doi: 10.1210/clinem/dgaa124
10. Guéry L, Hugues S. Th17 cell plasticity and functions in cancer immunity. BioMed Res Int (2015) 314620:1–11. doi: 10.1155/2015/314620
11. Chatterjee S, Daenthanasanmak A, Chakraborty P, Wyatt MW, Dhar P, Selvam SP, et al. CD38-NAD + Axis regulates immunotherapeutic anti-tumor T cell response. Cell Metab (2018) 27(1):84–100. doi: 10.1016/j.cmet.2017.10.006
12. Kaiser Y, Lepzien R, Kullberg S, Eklund A, Smed-Sörensen A, Grunewald J. Expanded lung T-bet+RORγT+ CD4+ T-cells in sarcoidosis patients with a favourable disease phenotype. Eur Respir J (2016) 48(2):484–94. doi: 10.1183/13993003.00092-2016
13. Krausgruber T, Schiering C, Adelmann K, Harrison OJ, Chomka A, Pearson C, et al. T-bet is a key modulator of IL-23-driven pathogenic CD4 + T cell responses in the intestine. Nat Commun (2016) 7(May):1–12. doi: 10.1038/ncomms11627
14. Basu R, O’Quinn DB, Silberger DJ, Schoeb TR, Fouser L, Ouyang W, et al. Th22 cells are an important source of IL-22 for host protection against enteropathogenic bacteria. Immunity (2012) 37(6):1061–75. doi: 10.1016/j.immuni.2012.08.024
15. Plank MW, Kaiko GE, Maltby S, Weaver J, Tay HL, Shen W, et al. Th22 cells form a distinct Th lineage from Th17 cells in vitro with unique transcriptional properties and Tbet-dependent Th1 plasticity. J Immunol (2017) 198(5):2182–90. doi: 10.4049/jimmunol.1601480
16. Rolla S, Bardina V, De Mercanti S, Quaglino P, De Palma R, Gned D, et al. Th22 cells are expanded in multiple sclerosis and are resistant to IFN-β. J Leukoc Biol (2014) 96(6):1155–64. doi: 10.1189/jlb.5A0813-463RR
17. Carpio VH, Aussenac F, Puebla-Clark L, Wilson KD, Villarino AV, Dent AL, et al. T helper plasticity is orchestrated by STAT3, bcl6, and blimp-1 balancing pathology and protection in malaria. iScience (2020) 23(7):1–18. doi: 10.1016/j.isci.2020.101310
18. Carpio VH, Opata MM, Montañez ME, Banerjee PP, Dent AL, Stephens R. IFN-γ and IL-21 Double Producing T Cells Are Bcl6-Independent and Survive into the Memory Phase in Plasmodium chabaudi Infection. PloS One (2015) 10(12):1–19. doi: 10.1371/journal.pone.0144654
19. Xiao L, Jia L, Zhang Y, Yu S, Wu X, Yang B, et al. Human IL-21 +IFN-γ+CD4+ T cells in nasal polyps are regulated by IL-12. Sci Rep (2015) 5(12781):1–12. doi: 10.1038/srep12781
20. Zhang Y, Siegel AM, Sun G, Dimaggio T, Freeman AF, Milner JD. Human TH9 differentiation is dependent on signal transducer and activator of transcription (STAT) 3 to restrain STAT1-mediated inhibition. J Allergy Clin Immunol (2019) 143(3):1108–1118.e4. doi: 10.1016/j.jaci.2018.06.036
21. Pang N, Zhang F, Ma X, Zhang Z, Zhao H, Xin Y, et al. Th9/IL-9 profile in human echinococcosis: Their involvement in immune response during infection by Echinococcus granulosus. Mediators Inflamm (2014) 2014(781649):1–11. doi: 10.1155/2014/781649
22. Wilhelm C, Hirota K, Stieglitz B, Van Snick J, Tolaini M, Lahl K, et al. An IL-9 fate reporter demonstrates the induction of an innate IL-9 response in lung inflammation. Nat Immunol (2011) 12(11):1071–7. doi: 10.1038/ni.2133
23. Cosmi L, Maggi L, Santarlasci V, Capone M, Cardilicchia E, Frosali F, et al. Identification of a novel subset of human circulating memory CD4+ T cells that produce both IL-17A and IL-4. J Allergy Clin Immunol (2010) 125(1–3):222–30. doi: 10.1016/j.jaci.2009.10.012
24. Wang YH, Voo KS, Liu B, Chen CY, Uygungil B, Spoede W, et al. A novel subset of CD4+ TH2 memory/ effector cells that produce inflammatory IL-17 cytokine and promote the exacerbation of chronic allergic asthma. J Exp Med (2010) 207(11):2479–91. doi: 10.1084/jem.20101376
25. Raymond M, Van VQ, Wakahara K, Rubio M, Sarfati M. Lung dendritic cells induce TH17 cells that produce T H2 cytokines, express GATA-3, and promote airway inflammation. J Allergy Clin Immunol (2011) 128(1):192–201.e6. doi: 10.1016/j.jaci.2011.04.029
26. McCluskey D, Benzian-Olsson N, Mahil SK, Hassi NK, Wohnhaas CT, Burden AD, et al. Single-cell analysis implicates TH17-to-TH2 cell plasticity in the pathogenesis of palmoplantar pustulosis. J Allergy Clin Immunol (2022) 150(4):882–93. doi: 10.1016/j.jaci.2022.04.027
27. Elyaman W, Bradshaw EM, Uyttenhove C, Dardalhon V, Awasthi A, Imitola J, et al. IL-9 induces differentiation of TH17 cells and enhances function of FoxP3+ natural regulatory T cells. Proc Natl Acad Sci USA (2009) 106(31):12885–90. doi: 10.1073/pnas.0812530106
28. Stephens GL, Swerdlow B, Benjamin E, Coyle AJ, Humbles A, Kolbeck R, et al. IL-9 is a Th17-derived cytokine that limits pathogenic activity in organ-specific autoimmune disease. Eur J Immunol (2011) 41(4):952–62. doi: 10.1002/eji.201040879
29. Liao W, Spolski R, Li P, Du N, West EE, Ren M, et al. Opposing actions of IL-2 and IL-21 on Th9 differentiation correlate with their differential regulation of BCL6 expression. Proc Natl Acad Sci USA (2014) 111(9):3508–13. doi: 10.1073/pnas.1301138111
30. Ryan ES, Micci L, Fromentin R, Paganini S, McGary CS, Easley K, et al. Loss of function of intestinal IL-17 and IL-22 producing cells contributes to inflammation and viral persistence in SIV-infected Rhesus Macaques. PloS Pathog (2016) 12(2):1–22. doi: 10.1371/journal.ppat.1005412
31. Barnes JL, Plank MW, Asquith K, Maltby S, Sabino LR, Kaiko GE, et al. T-helper 22 cells develop as a distinct lineage from Th17 cells during bacterial infection and phenotypic stability is regulated by T-bet. Mucosal Immunol (2021) 14(5):1077–87. doi: 10.1038/s41385-021-00414-6
32. Kimura T, Kobiyama K, Winkels H, Tse K, Miller J, Vassallo M, et al. Regulatory CD4+ T cells recognize major histocompatibility complex class II molecule-restricted peptide epitopes of apolipoprotein B. Circulation (2018) 138(11):1130–43. doi: 10.1161/CIRCULATIONAHA.117.031420
33. Halim L, Romano M, McGregor R, Correa I, Pavlidis P, Grageda N, et al. An atlas of human regulatory T helper-like cells reveals features of Th2-like tregs that support a tumorigenic environment. Cell Rep (2017) 20(3):757–70. doi: 10.1016/j.celrep.2017.06.079
34. Tortola L, Jacobs A, Pohlmeier L, Obermair FJ, Ampenberger F, Bodenmiller B, et al. High-dimensional T helper cell profiling reveals a broad diversity of stably committed effector states and uncovers interlineage relationships. Immunity (2020) 53(3):597–613. doi: 10.1016/j.immuni.2020.07.001
35. Burel JG, Apte SH, Groves PL, McCarthy JS, Doolan DL. Polyfunctional and IFN-γ monofunctional human CD4+ T cell populations are molecularly distinct. JCI Insight (2017) 2(3):1–13. doi: 10.1172/jci.insight.87499
36. Zheng HY, Zhang M, Yang CX, Zhang N, Wang XC, Yang XP, et al. Elevated exhaustion levels and reduced functional diversity of T cells in peripheral blood may predict severe progression in COVID-19 patients. Cell Mol Immunol (2020), 1–3. doi: 10.1038/s41423-020-0401-3
37. Martinez-Sanchez ME, Mendoza L, Villarreal C, Alvarez-Buylla ER. A minimal regulatory network of extrinsic and intrinsic factors recovers observed patterns of CD4+ T cell differentiation and plasticity. PloS Comput Biol (2015) 11(6):e1004324. doi: 10.1371/journal.pcbi.1004324
38. Martinez-Sanchez ME, Huerta L, Alvarez-Buylla ER, Luján CV. Role of cytokine combinations on CD4+ T cell differentiation, partial polarization, and plasticity: Continuous network modeling approach. Front Physiol (2018) 9(AUG):1–14. doi: 10.3389/fphys.2018.00877
39. Martinez-Sanchez ME, Hiriart M, Alvarez-Buylla ER. The CD4+ T cell regulatory network mediates inflammatory responses during acute hyperinsulinemia: A simulation study. BMC Syst Biol (2017) 11(1):64. doi: 10.1186/s12918-017-0436-y
40. Bensussen A, Torres-Magallanes JA, Roces de Álvarez-Buylla E. Molecular tracking of insulin resistance and inflammation development on visceral adipose tissue. Front Immunol (2023) 14(March):1–17. doi: 10.3389/fimmu.2023.1014778
41. Puleston DJ, Baixauli F, Sanin DE, Edwards-Hicks J, Villa M, Kabat AM, et al. Polyamine metabolism is a central determinant of helper T cell lineage fidelity. Cell (2021) 184(16):4186–4202.e20. doi: 10.1016/j.cell.2021.06.007
42. Burt P, Peine M, Peine C, Borek Z, Serve S, Floßdorf M, et al. Dissecting the dynamic transcriptional landscape of early T helper cell differentiation into Th1, Th2, and Th1/2 hybrid cells. Front Immunol (2022) 13:1–11. doi: 10.3389/fimmu.2022.928018
Keywords: CD4+ T cells, hybrid lineages, plasticity, differentiation, immune response
Citation: Bensussen A, Torres-Magallanes JA, Álvarez-Buylla ER and de Álvarez-Buylla ER (2024) Hybrid lineages of CD4+ T cells: a handbook update. Front. Immunol. 15:1344078. doi: 10.3389/fimmu.2024.1344078
Received: 24 November 2023; Accepted: 04 January 2024;
Published: 19 January 2024.
Edited by:
Laura Patrussi, University of Siena, ItalyReviewed by:
Namrata Gautam, Moffitt Cancer Center, United StatesCopyright © 2024 Bensussen, Torres-Magallanes, Álvarez-Buylla and de Álvarez-Buylla. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Bensussen, YW50b25pby5iZW5zdXNzZW5AZ21haWwuY29t; Elena R. Álvarez-Buylla, ZWFidXlsbGFAZ21haWwuY29t; Elena Roces de Álvarez-Buylla, cmFiQHVjb2wubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.