 Ana Banko1*
Ana Banko1* Andja Cirkovic2
Andja Cirkovic2 Rada Miskovic3,4
Rada Miskovic3,4 Ivica Jeremic4,5
Ivica Jeremic4,5 Milka Grk6Milica Basaric5
Milka Grk6Milica Basaric5 Ivana Lazarevic1
Ivana Lazarevic1 Sanvila Raskovic3,4
Sanvila Raskovic3,4 Aleksa Despotovic2
Aleksa Despotovic2 Danijela Miljanovic1
Danijela Miljanovic1- 1Institute of Microbiology and Immunology, Faculty of Medicine, University of Belgrade, Belgrade, Serbia
- 2Institute for Medical Statistics and Informatics, Faculty of Medicine, University of Belgrade, Belgrade, Serbia
- 3Clinic of Allergy and Immunology, University Clinical Center of Serbia, Belgrade, Serbia
- 4Faculty of Medicine, University of Belgrade, Belgrade, Serbia
- 5Institute of Rheumatology, Faculty of Medicine, University of Belgrade, Belgrade, Serbia
- 6Institute of Human Genetics, Faculty of Medicine, University of Belgrade, Belgrade, Serbia
Introduction: The relationship between Systemic lupus erythematosus (SLE) and Epstein-Barr virus (EBV) infection has been suggested for decades, but the underlying mechanism of the EBV influence on SLE development remains to be elucidated.
Methods: The goals of this research, which included 103 SLE patients and 99 controls, were to investigate the association of the parameters of EBV infection and SLE, to explore whether pooled demographic, clinical and EBV markers achieve a more significant effect on SLE development than each of them individually, and to evaluate EBV nuclear antigen 1 (EBNA1) and latent membrane protein 1 (LMP1) gene polymorphisms in isolates from SLE patients.
Results: Comprehensive results related to serological, molecular and sequence markers of EBV infection in SLE patients demonstrated even 24 times higher possibility of having SLE if there is the presence of anti-EBV-EA(D) (early antigen) IgG antibodies (OR=24.086 95%CI OR=2.86-216.07, p=0.004). There was the same distribution of glucocorticoids (p=0.130), antimalarials (p=0.213), and immunosuppressives (p=0.712) in anti-EBV-EA(D) IgG positive and negative SLE patients. Further, higher anti-EBV-EA(D) IgG antibodies titers were identified as independent factors associated with lymphopenia, hematological SLE manifestation (OR=1.041, 95%CI OR=1.01-1.08, p=0.025, while a higher titer of anti-CA (viral capsid antigen) IgG antibodies (OR=1.015, 95%CI OR=1.01-1.03, p=0.019) and positive RF (rheumatoid factors) (OR=4.871, 95%CI OR=1.52-15.61, p=0.008) were identified as independent factors associated with alopecia within SLE. Finally, novel data on EBV EBNA1 and LMP1 gene polymorphisms in lupus are reported.
Conclusion: The results support further investigation targeting EBV as a prognostic marker and therapeutic goal for lupus.
1 Introduction
Systemic lupus erythematosus (SLE) is a female predominant systemic autoimmune disease. It is characterized by the involvement of multiple organs and significant morbidity through pathogenic autoantibody production caused by the autoactivation of T and B cells. SLE includes repeated episodes of exacerbation, known as flares, and remission. Similar to other autoimmune disorders, the etiology of SLE still needs to be clarified entirely. The interplay between various genetic and environmental risk factors contributes to the onset and perpetuation of SLE. From external factors, infection, particularly Epstein-Barr virus (EBV) infection, has multiple potential roles in driving autoimmunity (1).
EBV is one of the most common human viruses. As a member of the Human Herpesviruses family, it latently infects up to 99% of the world’s population with the ability to survive in cells for life (2). With such unique capacities, it represents a constant challenge to the host. After the primary lytic infection of epithelial cells, EBV maintains latency in B lymphocytes with the expression of a limited number of genes defined with different types of latency (3). However, it occasionally reactivates the lytic cycle resulting in shifts between different sets of expressed genes (4). The humoral response includes the synthesis of antiviral antibodies against antigens of both the lytic and latent phases. Firstly, anti-EA (early antigen) and anti-CA (viral capsid antigen), during the first 3-4 weeks and at the time of the onset of the clinical symptoms, respectively (5). Although anti-CA IgG levels persist for life, the levels are higher during lytic infection (primary or reactivation) (6). On the other hand, EA IgG could be found in 85% of the acute infection, with usual persistence of 3 months, but with detectable levels years after primary infection in some cases (5). It is estimated that up to 30% of healthy subjects with a history of EBV have EA (D) IgG (5). Moreover, these antibodies could be detectable during reactivations or in immunocompromised patients (5, 6). Anti-EBNA-1 IgG is usually undetectable during the first month after the onset of clinical symptoms and is therefore indicative of past infection (5).
The relationship between SLE and EBV infection has been suggested for decades, but the underlying mechanism of the EBV influence on SLE progression remains to be elucidated. Several models have been proposed, among which antibody production through typical molecular mimicry and epitope spreading is the most important. The EBV genome encodes human homolog proteins such as latent protein, EBV nuclear antigen 1 (EBNA1), and the lytic protein, viral IL-10, that in SLE patients stimulate humoral and inflammatory immune responses (7). In particular, EBNA1 epitopes induce the production of autoantibodies against C1q and cross-react with SLE autoantigens, including Sm and Ro (7, 8). Based on the previously described knowledge, it was suggested that impaired control of EBV infection and cell-mediated immunity, with altered T cell function and the presence of multiple antibodies, leads to polyclonal expansion of EBV-infected B cells (9). Finally, autoreactive B cell activation during flares leads to frequent relapsing-remitting episodes of EBV infection, contributing to a vicious circle of repeated immune stimulations (10). Recent examples of more frequent EBV reactivations in SLE patients include the presence and elevation of serological markers CA IgG and EA IgG, higher frequencies of EBV-infected cells, higher viral loads and blood mononuclear cells, and increased expression of lytic genes (11). Moreover, in SLE patients, EBV-specific cytotoxic T and dendritic cells have reduced functionality, decreasing interferon levels after EBV infection (12, 13). The inverse relationship between decreased EBV lytic cycle responsive T cells and levels of anti-EBV antibodies is also followed by a negative correlation with the SLE disease activity index (SLEDAI) (14).
The balance that EBV establishes with the immune system and the host’s capacity to control cell proliferation in some EBV-associated diseases could be braked by the consequences influenced by specific variability of EBV genes (15, 16). In this regard, EBNA1 and latent membrane protein 1 (LMP1) genes are particularly interesting, with EBNA1 as the most prominently expressed EBV gene and LMP1 as the crucial viral oncogene. Their nucleotide variants and sequence specificities in previous studies showed potential contributions to the malignancy development (15, 17, 18). However, investigation of EBV gene variability and whether any mutation in viral genes contributes to SLE pathology has never been published before. Moreover, understanding this diversity in EBV-associated diseases revealed the possibility of predicting potential viral epitope targets to determine a treatment strategy (19).
Guided by the knowledge gaps and numerous unsolved hypotheses about SLE evolution, the goals of this research were set: a) to investigate the association between the respondents’ characteristics, including the parameters of EBV infection and SLE; b) to explore whether pooled markers achieve a more significant effect on SLE development than each of them individually, c) to evaluate EBV EBNA1 and LMP1 gene polymorphisms in isolates from SLE patients.
2 Materials and methods
2.1 Study design and participants
This prospective cohort study included 103 SLE patients diagnosed and treated at the Clinic for Allergology and Immunology University Clinical Center of Serbia and the Institute of Rheumatology, Belgrade, between June 2020 and November 2021. All SLE patients met at least 4 ACR SLE 1997 classification criteria (20). Patients with significant comorbidities (severe cardiac, pulmonary, and psychiatric diseases or dementia) and those unable to cooperate were excluded from the study. After detailed clinical interviews and physical examination of patients, all relevant demographic, clinical, and laboratory data were collected from their medical records. Disease activity was assessed using SLEDAI 2K scale (21) and Physician Global Assessment (PGA) (21), while fatigue was evaluated by the FACIT Fatigue Scale (version 4) (22). Patients with SLEDAI 2K≥6 were considered to have active SLE. Lupus Low Disease Activity State (LLDAS) was defined as SLEDAI-2K score ≤4, no activity in major organs, and PGA <1 (23, 24), while SLEDAI-2K score = 0 defined remission. The control group consisted of 99 participants. After a thorough interview and physical examination, we excluded those with positive personal or family histories of systemic autoimmune diseases and suggestive signs and symptoms of systemic autoimmune or active malignant disease. The exclusion criteria for both study cohorts were age under 18, unfeasibility to cooperate, and malignancies. All participants provided written informed consent. The study was performed in accordance with the Declaration of Helsinki and was approved by the Ethical Board of the Faculty of Medicine, University of Belgrade (No 1550/IX-14).
2.2 Samples
SLE patients’ serum and whole blood samples were collected at the Clinic for Allergology and Immunology University Clinical Center of Serbia and at the Institute of Rheumatology, Belgrade. Control group serum and whole blood samples were obtained from volunteers at the Institute of Rheumatology, Belgrade and the Institute of Microbiology and Immunology, Faculty of Medicine, University of Belgrade. After collecting 5 ml of blood in plain vacutainers, sera were separated by centrifugation. Plasma was also separated by centrifugation from 5 ml of ethylenediaminetetraacetic acid (EDTA) blood collected in vacutainer tubes. Two tubes (sera and plasma) from each patient were immediately tested or stored at -70°C until further analysis.
2.3 EBV serological testing
Anti-EBV antibodies against CA (IgG), CA (IgM), EA (IgG), EA (IgM), and EBNA1 (IgG) were identified and measured using commercial ELISAs according to manufacturer’s instructions in collected sera (Euroimmun, Lubeck, Germany). The assays used recombinant [EBNA1 and EA(D)] or purified (CA) antigens. Standard calibrators were used in each assay to calculate index values/optical density (OD) ratios, which serve as a quantitative measure of IgG antibody levels or a semi-quantitative measurement of IgM antibody levels. All assays met pre-determined quality control measures based on positive, negative, and blank controls. The positivity of IgG antibody presence was defined by a cut-off value of 20 relative units (RU/ml). For the sample values that were out of linearity defined by the used assays, re-measurement was done as following: samples were remeasured in a new test run at a dilution of e.g. 1:400 and results in RU/ml read from the calibration curve were multiplied by a factor 4. The positivity of IgM antibody presence was defined as OD ratio ≥1.1. Absorbances were recorded on a Multiscan FC microplate reader (Thermo Scientific, Massachusetts, USA) using a wavelength of 405 nm with background subtraction at 650 nm.
2.4 EBV DNA detection
Viral DNA was isolated from 200 μL plasma using a PureLink Genomic DNA Mini Kit (Invitrogen by Thermo Fisher Scientific, Massachusetts, USA) according to the manufacturer’s instructions. Two hundred and two DNA isolates were further used in a nested-PCR method to amplify two EBV genes: EBNA1 and LMP1. Amplifications of the C terminus of EBNA1 and C terminus of LMP1 were performed by nested PCRs as described previously (25), using primers that were reported by Lorenzetti et al. and Li et al. (26, 27).
2.5 EBV EBNA1 and LMP1 carboxy-terminal regions sequencing with sequence analysis
For purification, performing cycle sequencing reactions and sequencing of EBNA1 and LMP1, both sense and antisense strands, principles described before were used (25).
LMP1 and EBNA1 nucleotide sequences 506-bp and 329-bp long were separately aligned and compared with a reference sequence in Bioedit 7.0.5.3 software (28). EBNA1 subtypes and sub-variants were defined after inspection of signature amino acid changes at the following positions: 471, 475, 476, 479, 487, 492, 499, 500, 502, 517, 520, 524, 525, 528, and 533. Characterization of LMP1 variants was performed using the same software by examination of amino acid changes described by Edwards et al. (29). The sequence used in both alignments as the reference was downloaded from the GenBank/EMBL7DDBJ database under the accession number V01555.
2.6 Data analysis
Numerical data were described with the arithmetic mean and standard deviation or median and interquartile range (IQR), depending on the data distribution. Normal distribution was evaluated by mathematical (Kolmogorov-Smirnov and Shapiro-Wilk test, skewness and kurtosis, and coefficient of variation) and graphical methods (histogram, box-plot, Q-Q diagram). Categorical variables were presented as absolute and relative numbers in n (%). Student t-test for two independent samples or Mann-Whitney U test was used to compare numerical data between study cohorts, depending on the data distribution. The chi-square test was used to test the difference in the distribution of categories of two independent samples.
To evaluate all possible factors influencing a person’s probability of having SLE, univariate logistic regression analysis was performed first, with multivariate analysis after that with significant variables from the previous analysis in the model. Variance Inflation Factor (VIF) multicollinearity testing method and correlation coefficients were used. All variables with VIF>5 were eliminated from the multivariate model. Also, according to the co-variance matrix and correlation coefficients, one of two correlated variables with a lower p-value in univariate logistic regression was eliminated. Forward Wald regression method was applied, and only the last step was presented in the results. The odds ratio (OR), 95% confidence interval of odds ratio (95% CI OR), and p-value were reported. Complete statistical analysis was performed in SPSS Statistics for Windows Version 26.0 (IBM Corp. Armonk, NY).
The sample size was calculated by G Power 3.1.9.2 for the major outcome – the presence of anti-EBV-EA(D) IgG antibodies in SLE patients and control according to previously known data (7) and for the effect size of 0.3, error of the I type (α) of 0.05, and statistical power of 0.8. It was obtained that 88 observation units per study group would be enough to determine the differences between them.
3 Results
3.1 Characteristics of the study cohorts
This study included 103 SLE patients, mostly women (91.3%), with an average age of 45.42 ± 12.9 years. The most common manifestations were lymphopenia (50%), then alopecia (39%), rash (32%), arthritis (32%), and leucopenia (32%). Lupus nephritis was present in 15%, neutropenia in 13%, mucosal ulcerations in 12%, and thrombocytopenia in 11%. Almost 40% of SLE patients had secondary Sjogren syndrome, and less than 10% had secondary antiphospholipid syndrome. Of 99 participants in the control group, there were 88% women with an average age of 55.43 ± 13.62 years. More detailed demographic, clinical and laboratory characteristics of study cohorts are shown in Table 1. SLE patients were treated with corticosteroids (87%), antimalarials (83%), immunosuppressants (29%), and pulse therapy (13%).
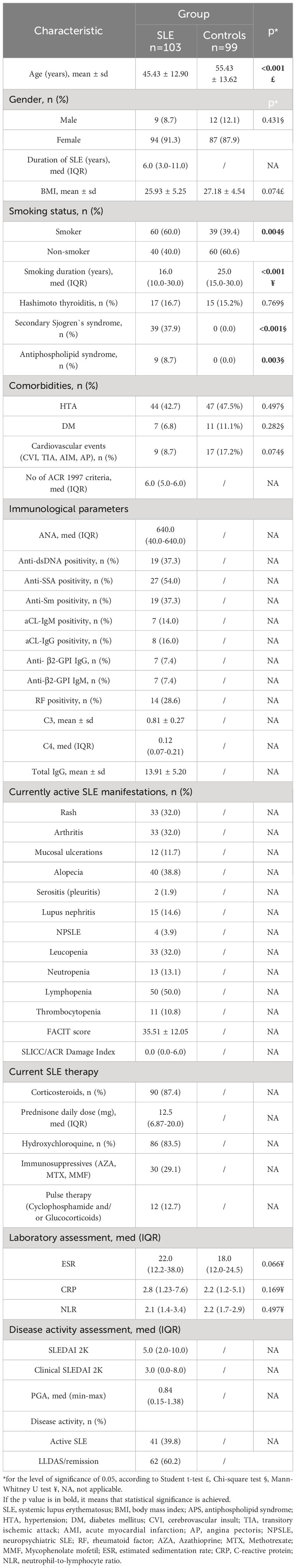
Table 1 Baseline characteristics of the study cohorts.
3.2 EBV infection status in SLE patients and controls
The prevalence of EBV DNA (EBNA1 and/or LMP1 gene) was 11% in the plasma of SLE patients, without statistically significant difference in comparison with controls (p=0.893). Anti-EBV-CA IgM, anti-EBV-EA(D) IgG, and anti-EBV-EA(D) IgM antibodies were more prevalent in SLE patients than in controls (p<0.001, p<0.001, and p=0.008, respectively). The status of EBV infection was determined based on the results from both EBV DNA and anti-EBV antibody testing. The active EBV infection was declared if EBV DNA and/or anti-EBV IgM antibodies were present. SLE patients more often had active EBV infection than controls (42% vs 18%, p<0.001) (Table 2). SLE patients with latent EBV infection were significantly more often treated with immunosuppressive drugs than those with active infection (37% vs. 19%, p=0.047), while the distribution of other medications (corticosteroids, antimalarials, pulse therapy) was the same between SLE patients with active and latent EBV infection (p=0.797, p=0.306, and p=0.773, respectively) (Available in Supplementary Material Table 1).
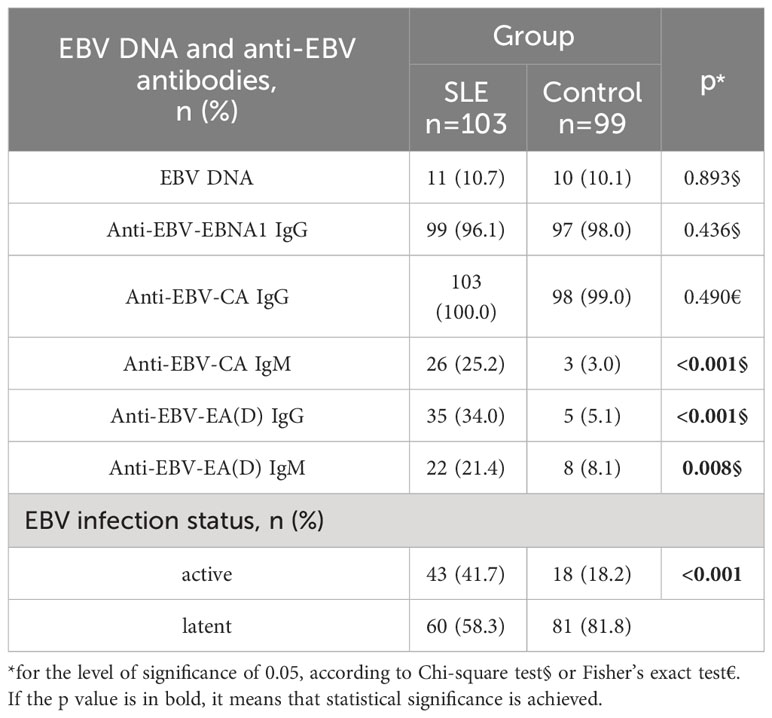
Table 2 Prevalence of EBV DNA, anti-EBV antibodies and active EBV infection.
The titers of all evaluated anti-EBV antibodies in SLE patients and controls were further compared (Figures 1–5). SLE patients had significantly higher titers of anti-EBV-EBNA1 IgG, anti-EBV-CA IgM, anti-EBV-EA(D) IgG, anti-EBV-EA(D) IgM antibodies than controls (p<0.001, p<0.001, p<0.001, and p=0.012, respectively), while the titer of anti-EBV-CA IgG antibodies was significantly higher in controls than in SLE patients (p=0.003).
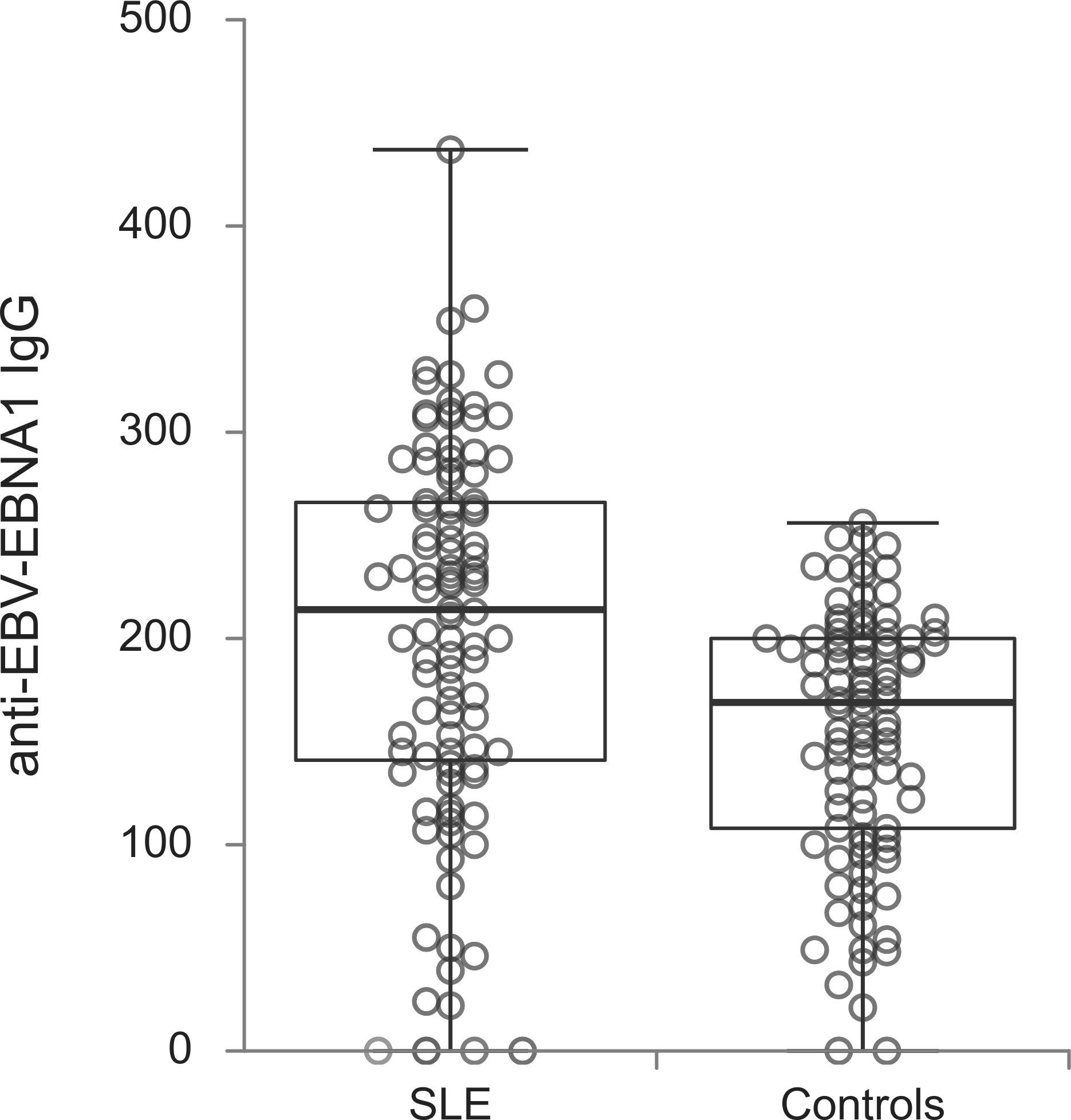
Figure 1 Anti-EBV-EBNA1 IgG antibodies in SLE patients and controls (p<0.001).
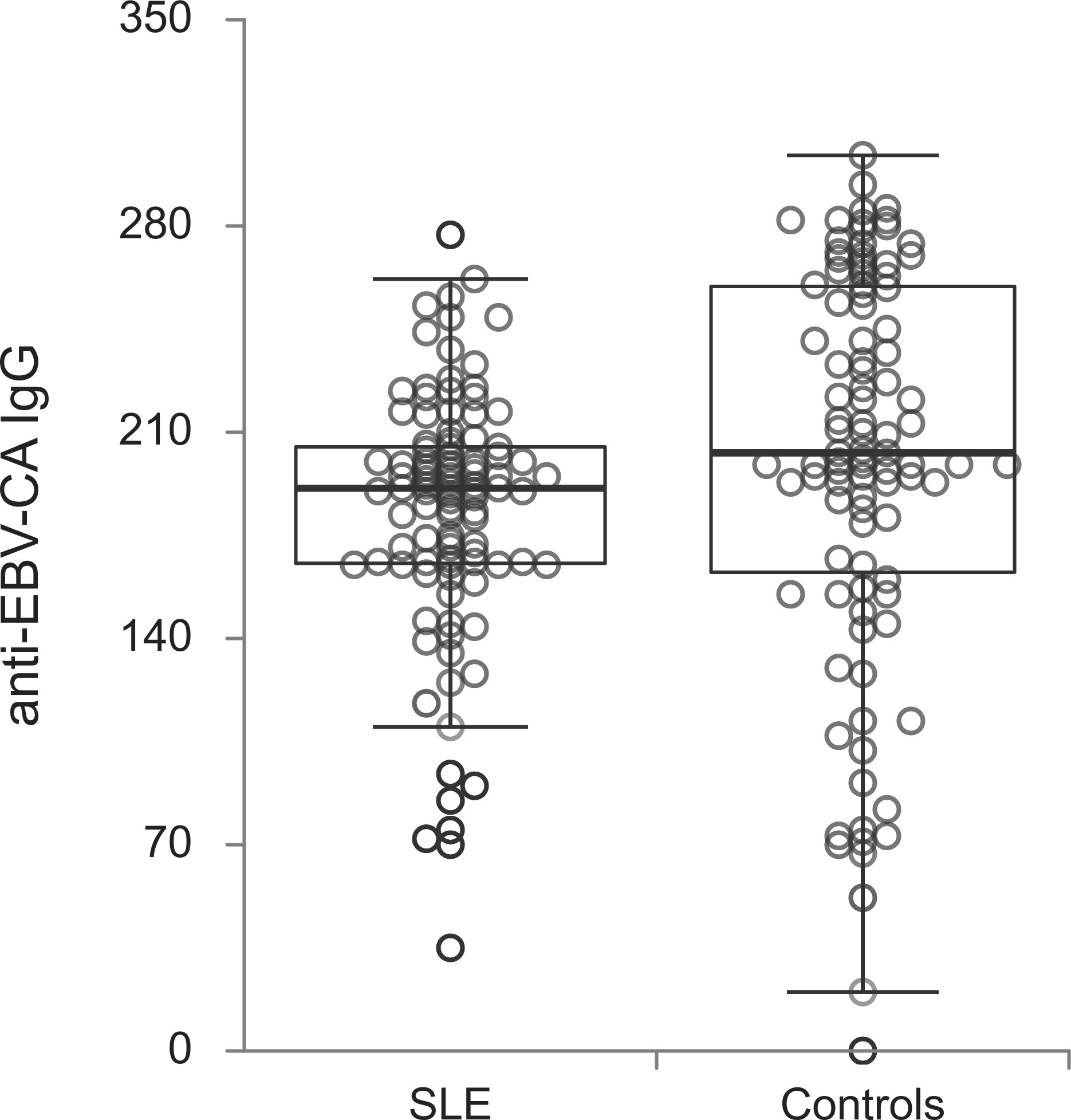
Figure 2 Anti-EBV-CA IgG antibodies in SLE patients and controls (p=0.003).
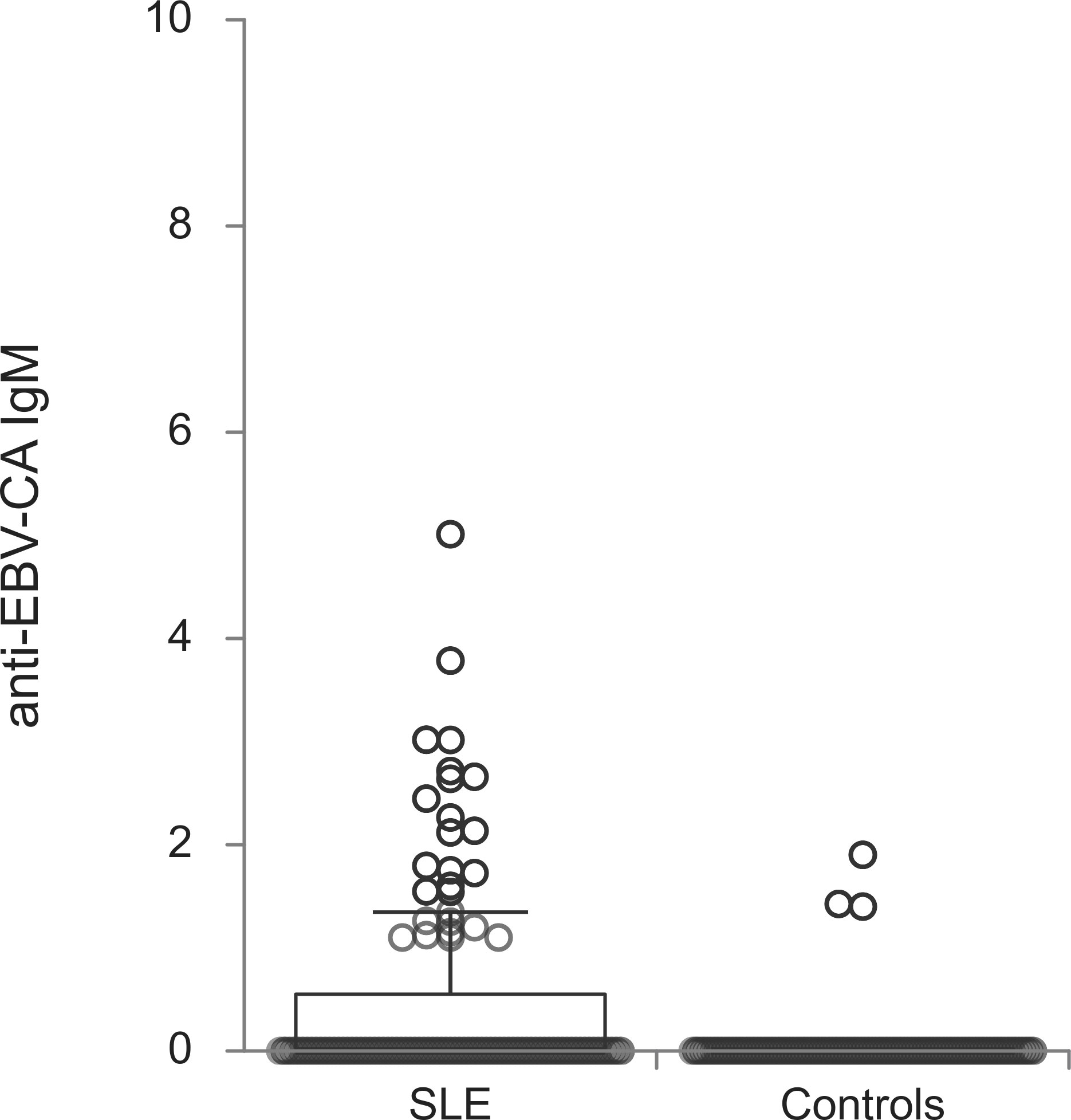
Figure 3 Anti-EBV-CA IgM antibodies in SLE patients and controls (p<0.001).
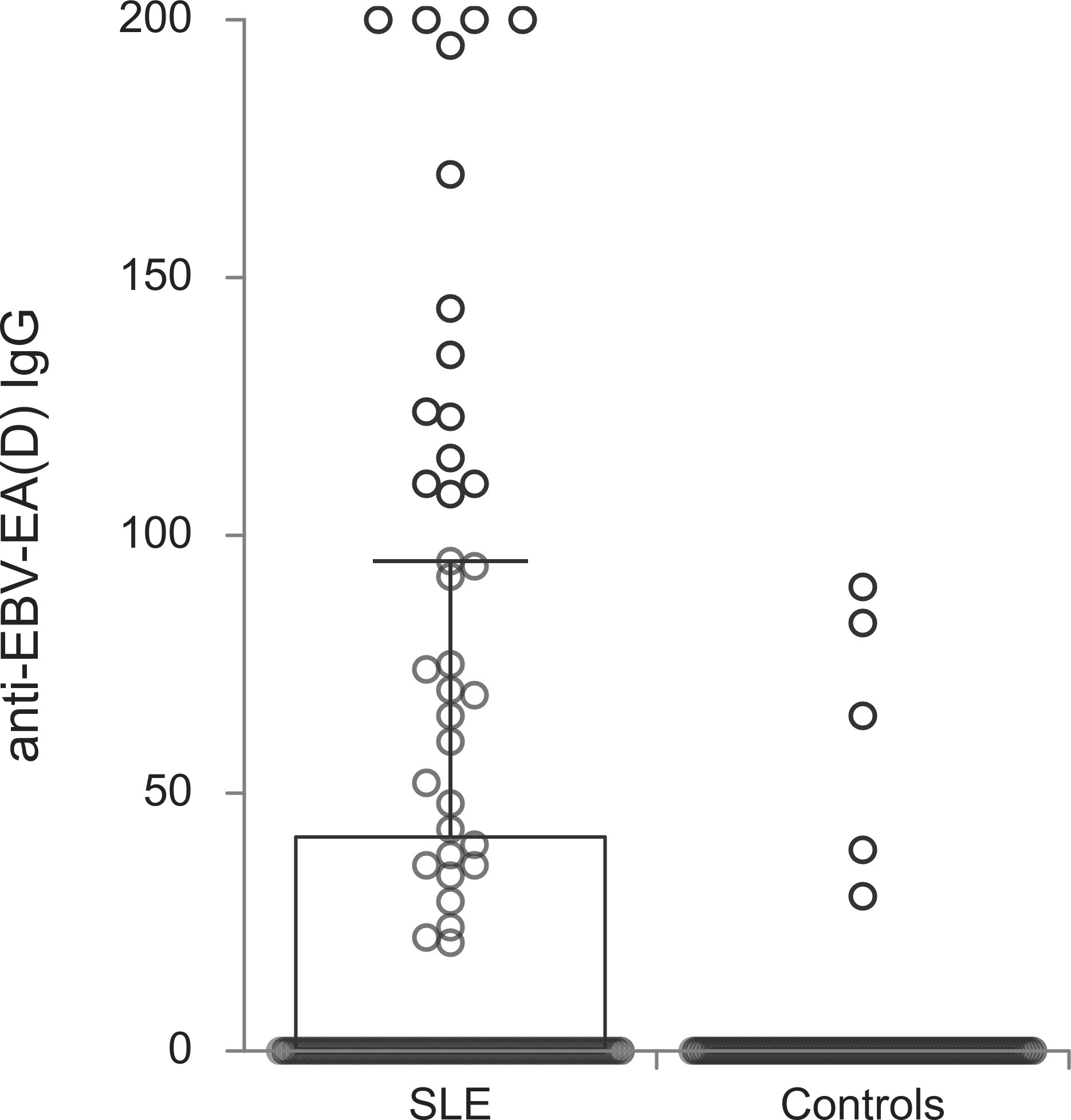
Figure 4 Anti-EBV-EA(D) IgG antibodies in SLE patients and controls (p<0.001).
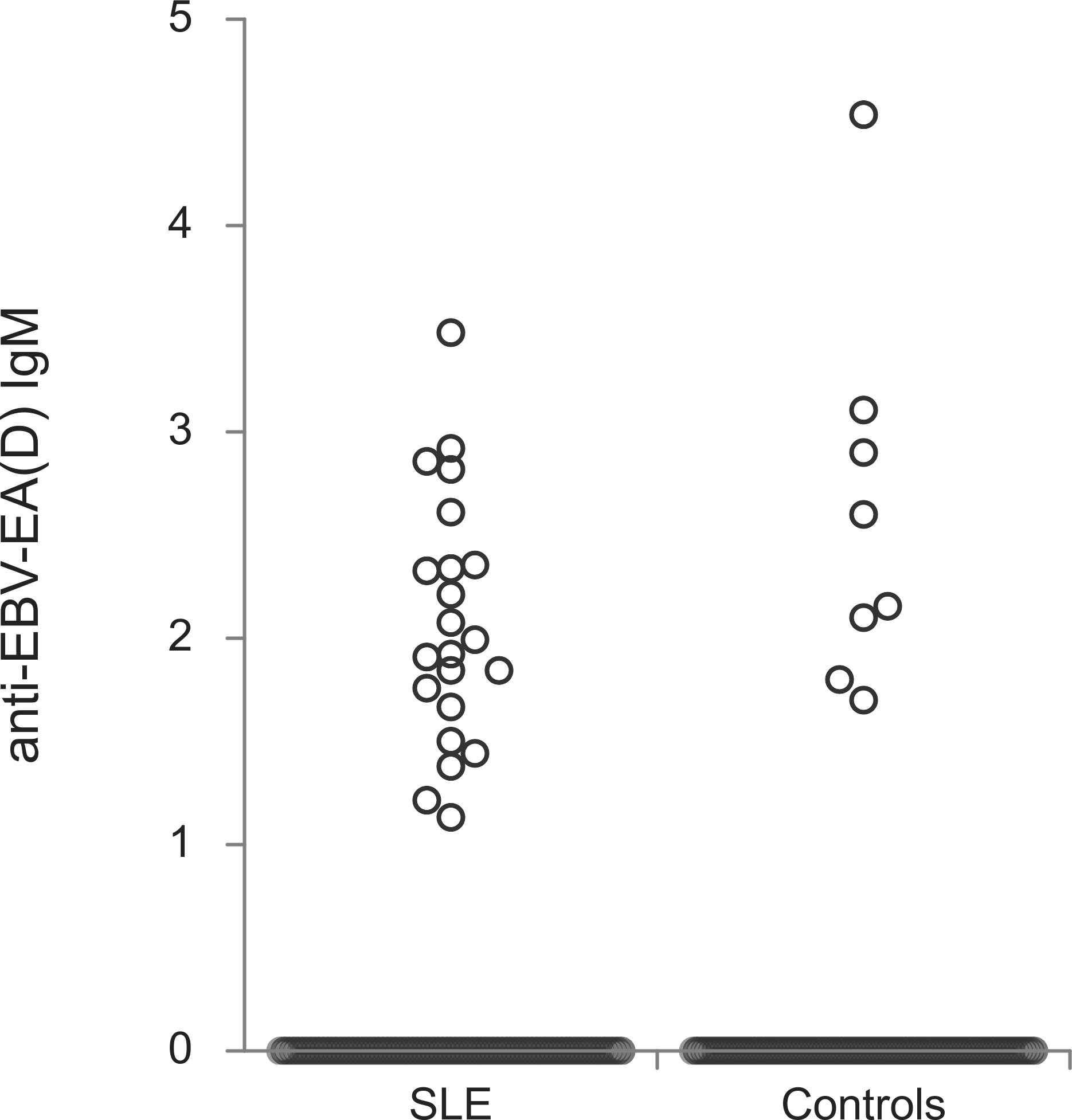
Figure 5 Anti-EBV-EA(D) IgM antibodies in SLE patients and controls (p=0.012).
3.3 Evaluation of factors associated with SLE
All applicable EBV and clinicodemographic parameters were evaluated for defined outcomes (presence of SLE, the active form of SLE disease, and different clinical manifestations of SLE: rash, arthritis, ulcerations, alopecia, nephritis, leucopenia, lymphopenia, thrombocytopenia). Univariate, then multivariate regression analysis was performed. All variables with p<0.05 in univariate regression were included in the multivariate model taking into consideration collinearity. The complete analysis is available in the Supplementary Material Tables 2-12.
3.3.1 Baseline characteristics and markers of EBV infection in association with systemic lupus erythematosus
According to multivariate regression modeling, younger age (OR=0.814, 95%CI OR=0.74-0.89, p<0.001), the presence of anti-EBV-EA(D) IgG antibodies (OR=24.086, 95%CI OR=2.68-216.07, p=0.004), higher titers of anti-EBNA1 IgG antibodies (OR=1.023, 95%CI OR=1.01-1.03 p<0.001) and anti-EBV-CA IgM antibodies (OR=3.105, 95%CI OR=1.07-8.98, p=0.036), and lower titer of anti-EBV-CA IgG antibodies (OR=0.984, 95%CI OR=0.97-0.99, p=0.013) were independently associated with the presence of systemic lupus erythematosus (Table 3).
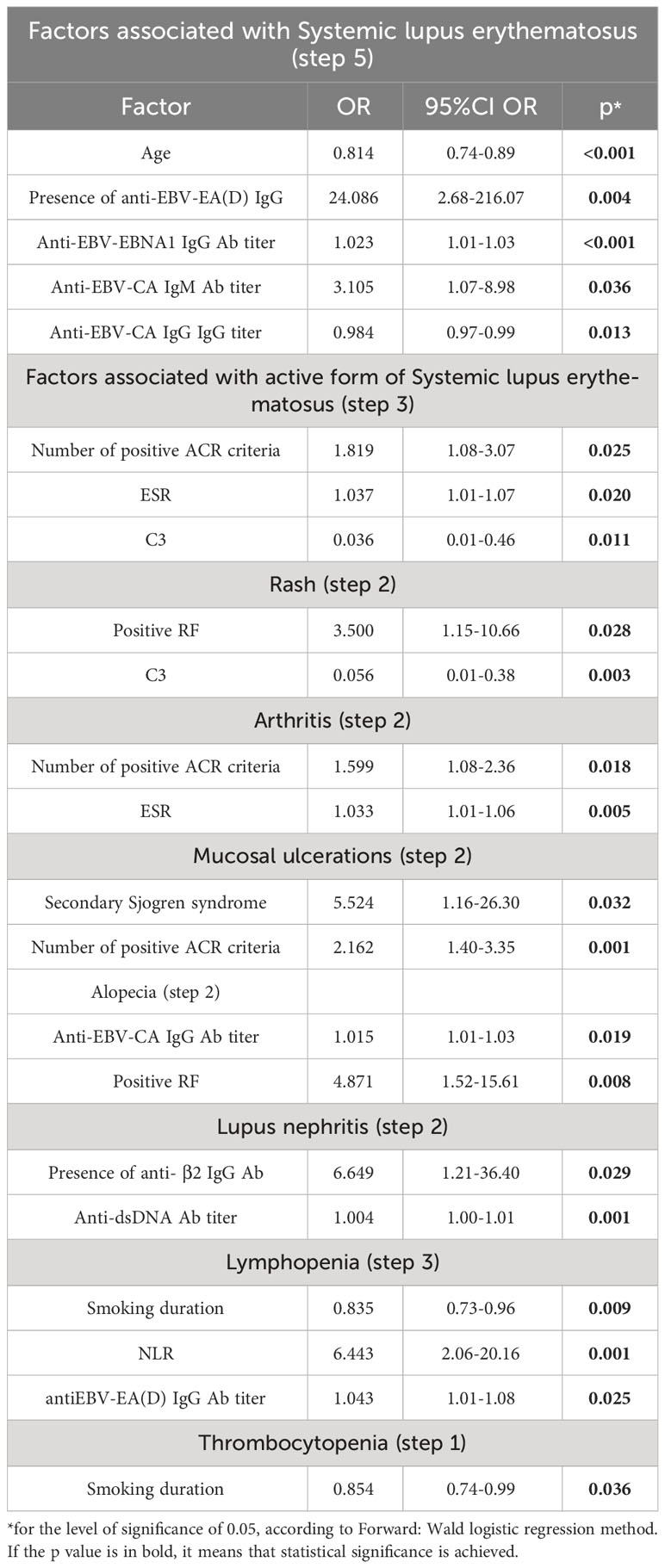
Table 3 Factors associated with Systemic lupus erythematosus and its manifestations.
3.3.2 Association between baseline characteristics and markers of EBV infection and active SLE disease
According to multivariate regression modeling, a greater number of positive criteria (OR=1.819, 95%CI OR=1.08-3.07, p=0.025), higher ESR (OR=1.037, 05%CI OR=1.01-1.07, p=0.020), and lower C3 complement level (OR=0.036, 95% CI OR=0.01-0.46, p=0.011) were independent factors associated with active SLE disease (Table 3).
3.3.3 Association between baseline characteristics and markers of EBV infection and lupus manifestations
3.3.3.1 Rash
Lower C3 level (OR=0.056, 95%CI OR=0.01-0.38, p=0.003) and positive rheumatoid factor (OR=3.500, 95%CI OR=1.15-10.66, p=0.028) were independent factors associated with rash lupus manifestation (Table 3).
3.3.3.2 Arthritis
Greater number of positive criteria (OR=1.599, 95%CI OR=1.08-2.36, p=0.018) and higher ESR (OR=1.033, 95%CI OR=1.01-1.06, p=0.005) were independent factors associated with arthritis within SLE (Table 3).
3.3.3.3 Mucosal ulcerations
Greater number of positive ACR criteria (OR=2.162, 95%CI OR=1.40-3.35, p=0.001) and presence of secondary Sjogren syndrome (OR=5.524, 95%CI OR=1.16-26.30, p=0.032) were factors independently associated with mucosal ulcerations within SLE (Table 3).
3.3.3.4 Alopecia
Higher titer of anti-EBV-CA IgG antibodies (OR=1.015, 95%CI OR=1.01-1.03, p=0.019) and positive RF (OR=4.871, 95%CI OR=1.52-15.61, p=0.008) were independent factors related to alopecia within SLE (Table 3).
3.3.3.5 Lupus nephritis
Positive anti-b2-GP I IgG (OR=6.649, 95%CI OR=1.21-36.40, p=0.029) and higher titer of anti-dsDNA (OR=1.004, 95%CI OR=1.00-1.01, p=0.001) were independent factors associated with lupus nephritis (Table 3).
3.3.3.6 Leucopenia
The presence of anti-SSA antibodies was the only factor potentially associated with leucopenia (OR=4.676, 95%CI OR=1.18-18.51, p=0.028).
3.3.3.7 Lymphopenia
Shorter smoking duration (OR=0.835, 95%CI OR=0.73-0.96, p=0.009), higher neutrophil-to-lymphocyte ratio (NLR) (OR=6.443, 95%CI OR=2.06-20.16, p=0.001), and higher titer of anti-EBV-EA(D) IgG antibodies (OR=1.041, 95%CI R=1.01-1.08, p=0.025) were independent factors associated with Lymphopenia (Table 3).
3.3.3.8 Thrombocytopenia
Shorter smoking duration (OR=0.854, 95%CI OR=0.74-0.99, p=0.036) was an independent factor associated with thrombocytopenia (Table 3).
3.4 Variations of EBV EBNA1 and LMP1 carboxy-terminal regions sequences
Eight sequences of the EBNA1 gene fragment (coordinates 109261-109590) from 8 SLE patients were analyzed and compared with the B95-8 prototype strain. Inspection included signature amino acid changes at the following positions: 471, 475, 476, 479, 487, 492, 499, 500, 502, 517, 520, 524, 525, 528, and 533. Based on amino acid substitutions, isolates were grouped into two prototype subtypes, P-Ala and P-Thr (Table 4). Since none of the obtained sequences fully corresponded to the prototype sequences, further analysis of additional nucleotide variability and amino acid substitutions was performed. Three subvariants (sv) were identified: P-Ala-sv-2 and -3; P-Thr-sv-7. All three subvariants were previously found in Serbian isolates, however, P-Ala-sv-3 and P-Thr-sv-7 were described as newly identified (18). P-Ala-sv-3 differed from P-Ala-sv-2 in position 499, where the new subvariant retained the same nucleotide combination as the prototype sequence. P-Thr-sv-7 was characterized by four changes compared to the prototype sequence (positions 476, 487, 492, and 524) or two changes compared to the P-Thr sequence (499 and 520).
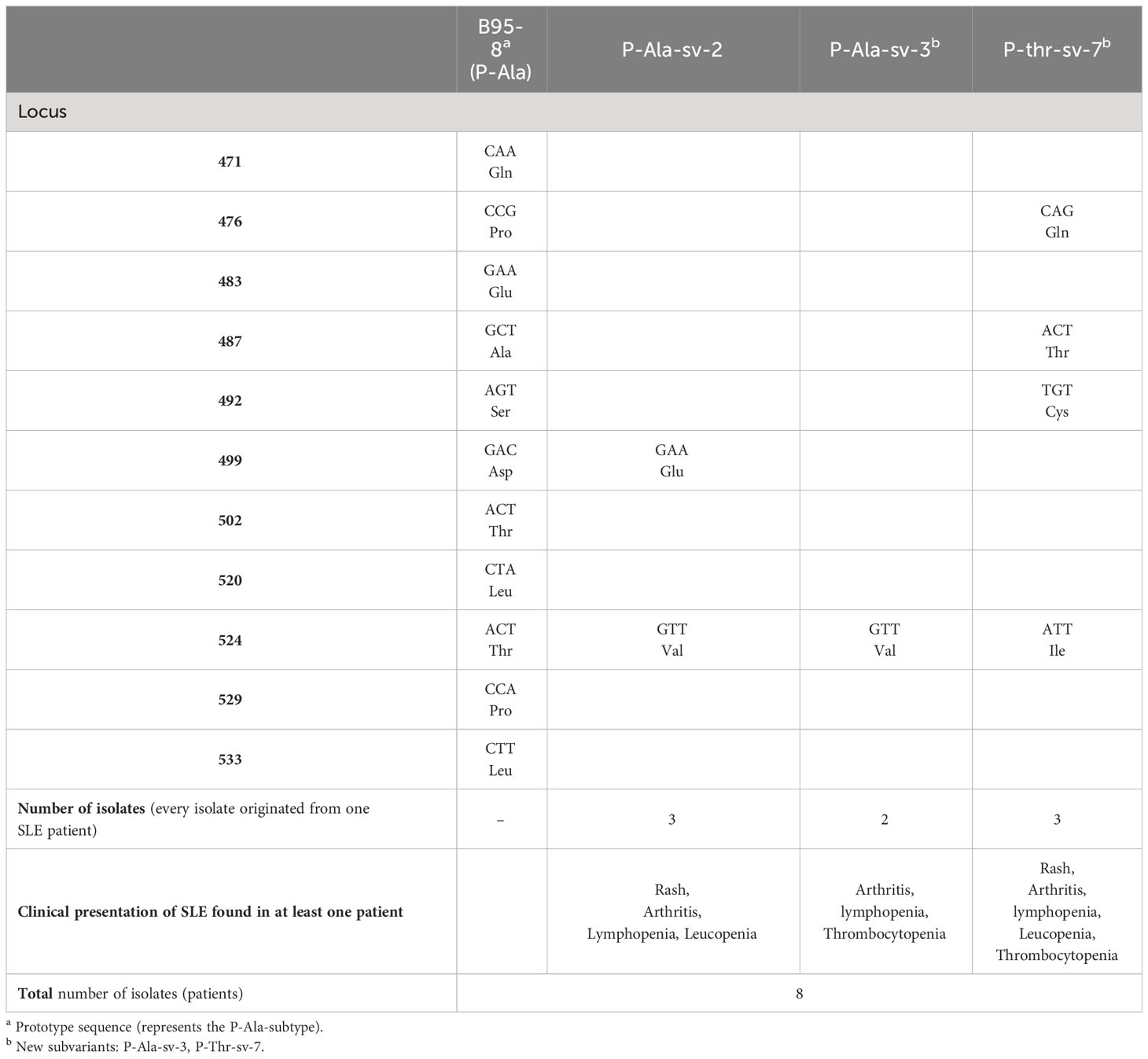
Table 4 EBNA1 C-terminal nucleotide and amino acid changes found in EBV isolates in SLE patients.
Eight sequences of the LMP1 gene fragment (coordinates 168719-168213) from 8 SLE patients were aligned and compared with the B95-8 prototype strain. Characterization of LMP1 variants was performed using by well-known examination of amino acid changes described by Edwards et al. (29). Due to divergence from the reference sequence, presence of deletions, number of 11-amino acid repeats and total amino acid changes, isolates were divided into four variants: B95-8, North Carolina, Mediterranean and China1 (Table 5).

Table 5 Distribution of three LMP1 characteristics in EBV isolates in SLE patients.
4 Discussion
After numerous attempts to prove a cause-and-effect relationship between EBV infection and SLE, more data on EBV-dependent features still need to be included for this evidence (30). The only fact is that the association between SLE and EBV increased reactivation due to insufficient latency regulation or enhanced transition from latent to lytic phase (30, 31). This confirmation was established by a few dozen studies based on detecting EBV infection markers in lupus patients (9). However, most studies measured a limited number of viral markers, quite often without a control group to compare findings. Thus, the latest publications did not include IgM antibodies to CA and EA, free viral DNA or gene sequence variability (7). Moreover, European research was limited primarily to Denmark and Italy, with rare exceptions from France, Netherlands and Hungary (9). To our knowledge, comprehensive analysis of EBV markers in lupus patients and their potential relationship with characteristics, different clinical presentations of SLE or therapeutic modalities is rare among literature data. Therefore, identifying independent EBV factors for SLE and our study’s first molecular analysis of EBV genes represent a valuable contribution to this still under-investigated scientific field.
It is known that occasional EBV reactivations can be diagnosed serologically and by molecular detection of viral DNA. Although the specific serological interpretation of EBV reactivation in SLE patients may be missed because of a possible reflection of a non-specific antibody increase due to an autoimmune background, the available results say otherwise. The antibody responses towards other herpesviruses, including cytomegalovirus (CMV) and herpes simplex virus (HSV), are similar between SLE patients and controls (30). Interestingly, however, the dominant EBV latency type maintained during lupus could not be clearly defined using commonly known criteria. In particular, the combination of BZLF1 and LMP1 in SLE patients probably represents dysregulated latent gene expression in the direction of intermediate latency form between latency II and III (30). On the other hand, EBV reactivation in these patients does not have to be a consequence of immune dysregulation but perhaps immunosuppression (11).
This research emphasized the hypothesis that the immune response to EBV activity represents an important element in the sequence of events that follows the clinical course of SLE. According to the results, there is undoubtedly a highly significant EBV activity compared to the control group. Firstly, anti-EBV-CA IgM, anti-EBV-EA(D) IgG, and anti-EBV-EA(D) IgM antibodies were more prevalent in SLE patients than controls. Moreover, all of the mentioned antibodies together with anti-EBV-EBNA1 IgG had also significantly higher titers in SLE patients than controls. Previous research showed similar results, including prospective investigation of SLE disease transition (7, 11). Based on all processed viral parameters, active EBV infection was more often in the lupus group than in the control group, corroborating previous descriptions of the delicate relationship between the virus and SLE pathogenesis (7, 10, 11). Besides SLE, our research group obtained similar results in patients with rheumatoid arthritis (RA), another systemic autoimmune disease. Active or recent EBV infection was proven in the same percentage among RA patients (42%) and more prevalent than controls (32). Finally, approximately equal seroprevalence of anti-EBV-EBNA1-IgG and anti-EBV-CA IgG antibodies in both study groups showed the same level of prior EBV exposure. These findings suggested the specificity of selected serological markers that proved viral activity in SLE patients.
EBV, in particular EBNA1, is a candidate for the heteroimmune response (11). According to the hypothesis, this response represents the substrate for generating SLE autoantibodies. Thus anti-EBV-EBNA1 IgG antibodies should be present at a higher rate in lupus patients (31). However, very few of the latest studies dealing only with seroprevalence, have managed to show this (9, 31, 33). Their authors suggested that the anti-EBNA1 response independently carries a risk for SLE which was documented with the higher prevalence of anti-EBNA1 antibodies in lupus patients. On the other hand, more publications including one of the latest meta-analyses nor reach this conclusion (9). Following the recent findings from Laurynenka et al. (9, 31, 33) and their suggestion that anti-EBNA1 heteroimmune response is a foundation from which pathogenic SLE autoimmunity develops, our results could be an important contributor to this theory. Although in the subjects from our study, regardless of the presence of SLE, anti-EBNA1 IgG seroprevalence was equal, their titers were significantly higher in lupus patients. Thus, these additional data on higher titer values could be a consequence of progressed anti-EBNA1 response, its cross-reaction with SLE autoantigens led by molecular mimicry, autoantibody epitope spreading and development of clinically manifested lupus (9, 31, 33).
The reports about the seroprevalence of anti-EBV-CA IgG between lupus patients and controls are conflicting. Our study showed the same level of prior EBV exposure between groups, which was in concordance with publications from Filipinos, India, the USA and more (7, 34, 35). On the other hand, some literature data described the higher prevalence of anti-EBV-CA IgG and its association with the development of SLE (9, 11). The authors even noted the potential influence of race on this association (36). Regardless of the mentioned differences in seroprevalence, most works including our, were uniform about the titer results with proved significant differences in the anti-EBV-CA IgG titers in SLE compared to the control groups (7, 11). It is known that EBV reactivation increases anti-EBV-CA IgG levels suggesting a more frequent and significant uncontrolled viral replication in lupus compared to the control group (6). However in other systemic autoimmune diseases the case was not the same, and our research group did not prove the mentioned difference in the anti-EBV-CA IgG titers when RA patients were taken into consideration and compared with a control group (32).
One of the most noticeable results confirmed the strong association between anti-EBV-EA(D) IgG antibodies and SLE. Thus, this study, for the first time, revealed a 24 times higher possibility of having SLE, if there is a presence of anti-EBV-EA(D) IgG antibodies. According to our knowledge, only one study so far obtained a higher index of association between EBV infection and autoimmune disease based on serological EBV status. Unlike ours, it was a huge longitudinal study that proved a 32-fold increase in multiple sclerosis risk after infection with EBV (37). In addition, younger age, higher titers of anti-EBNA1 IgG antibodies and lower titers of anti-EBV-CA IgG antibodies in our study were also found to be independently associated with the presence of lupus, but with negligible OR values when compared with anti-EBV-EA(D) IgG antibodies. On the basis of this result, it could become clear that the presence of anti-EBV-EA(D) IgG antibodies is a property that characterizes lupus. This probably reflected the chronic production of antibodies against cells expressing EA(D) and released EA(D) from lysed EBV-infected cells (38). Due to different types of infected cells, enhanced production of anti-EA(D) antibodies includes not only IgG (lymphocytes) but also IgA (epithelial cells), in addition to autoantibodies as EA(D) bound to dsDNA functions as EBV-DNA polymerase (38). More anti-EA(D) antibody types could suggest disseminated infection with a higher capacity to induce cell lysis and consequential autoimmunity. In addition to this, it is important to mention an apparent contradiction in how the treatment of SLE, which is based on immunosuppression, could be successful against a virus-induced autoimmune disease. This finding was also obtained in our study. The presence and elevation of anti-EA(D) IgG antibodies were independent of the applied type of immunosuppressive therapy. Supported by previous publications also, this paradox might be explained by the biology of EBV, its tropism for B lymphocytes and the capacity of immunosuppression to limit the spreading of the virus and infected B cells in addition to its alleviation of inflammation (38–42). Relying on the suggestions from the literature data of the importance of anti-EBV-EA(D) IgG for discrimination of SLE patients, together with results from our study, it is indicative that anti-EBV-EA(D) IgG could be considered as a marker of the presence of the disease (43, 44). Moreover, it could also be used to design future targeted therapy, as mentioned in the literature data (7, 45). One of the most recent research about SLE therapy identified blocking EBV lytic phase switching as a critical (8).
Interestingly, some researchers have even demonstrated an association between EBV activity and the severity of lupus disease (7, 46–48). Others failed to find any association between EBV antibody profile, viral load, and clinical manifestations (12, 35). By analyzing the association between different EBV infection markers and individual clinical presentations of lupus patients from Serbia, this study identified viral serological predictors for alopecia and specific hematological SLE manifestation for the first time. In particular, only one RU/ml higher titer of anti-EBV-EA(D) IgG antibodies carries a 4% higher chance for lymphopenia. Although the shorter smoking duration and higher NLR were also factors associated with lymphopenia, elevated anti-EBV-EA(D) IgG titers could be helpful during the laboratory monitoring of lupus and indicate the possibility of developing a specific clinical manifestation. As this result was not reported earlier, properly explaining the underlying mechanism could be difficult. However, it might be essential to interpret it together with the results of Wood et al. (7). In one of their latest paper, anti-EBV-EA IgG antibodies were higher when expression of interferon (IFN) and inflammatory or lymphoid and monocyte responses were higher. In addition, those patients with higher anti-EBV-EA(D) IgG responses were more likely to have renal involvement. Another publication that described this potential association with lupus nephritis detected the expression of EBV genes in renal tissue (49). Finally, in our study, a higher titer of anti-CA IgG antibodies and positive RF revealed independent factors related to alopecia within SLE. This finding should be analyzed in further investigations also.
The results, including the interplay between smoking and EBV in autoimmunity development, are scarce and conflicting. In those reported in multiple sclerosis (MS) patients, there are indications of competing antagonism, where mentioned exposures compete to affect the outcome (50). This study did not reveal any similar findings. However, shorter smoking duration was shown as an independent factor associated with lymphopenia in lupus patients.
According to the last systemic review with meta-analysis, only seven studies tested and compared EBV DNA presence between SLE and the control group (9). In addition to high heterogeneity, the positive rate for EBV DNA in SLE and control group was 55.1% and 20.7%, respectively. This difference was not described as significant due to the small number of studies, participants and heterogeneity. Results from our study did not reveal any difference between patients and healthy participants, as it was also published lately by Han et al. (46). Many authors proved a higher viral load in lupus of up to 40-fold greater increase regardless of immunosuppressive therapy (38). On the other hand, some authors did not find any viral genome at all (51). Thus, the large discrepancy in the stratification of SLE patients and the selection of samples for DNA testing must be addressed. Unfortunately, previous reports refer to peripheral B cells, PBMC and even sera (7, 38, 46, 48). Unlike them, our study tested viral DNA from free cell blood compartments depicting the amount of virus released during active replication and tendentially eliminating the amount that rests in latency. It was estimated that healthy carriers have approximately 1 to 50 infected cells per million leukocytes or 1 to 30 copies of EBV DNA per million leukocytes in whole blood. Thus, besides B cells or PBMC samples, any biopsy tissue may contain B lymphocytes and harbor amplifiable EBV DNA, too (52).
In our small group of positive EBV DNA isolates from SLE patients, we reported characteristics of EBNA1 and LMP1 gene sequences as an additional novelty. In need of more data on EBV gene sequences in lupus, reports on EBV gene sequences in other systemic autoimmune diseases are also extremely rare. They could be only found in RA or multiple sclerosis (MS) (19, 53, 54). In addition, those reports included only EBNA1 and EBNA2 variants. Similar to our study, previous authors did not prove the correlation of EBV variability with the risk of autoimmune disease (19, 54). However, some assumptions exist that specific virus genetic variants contribute to autoimmune development (53). EBNA1 was described as an unusual immunogen and antigen (31). Gene sequences were assessed to identify potentially specific EBNA1 gene variability associated with those irregularities. Together with the previous reports from Serbian EBNA1 isolates, the dominance of P-Ala and P-Thr subtypes again confirmed one of the earliest hypotheses of European distribution in non-malignant diseases and geographically specific distribution (18) (55). On the other hand, EBNA1 variability patterns in this study did not at all correlate with previous findings in RA isolates (54). Interesting is that only P-Ala-sv-3 isolates lacked mucocutaneous presentation. This finding could be further investigated on a more significant number of isolates.
Although the LMP1 gene is significantly heterogeneous with greater variability than most other EBV genes, there is no literature on LMP1 polymorphisms in systemic autoimmune diseases. Even though this EBV oncogene is primarily significant in EBV-related cancers, its immunogenic potential, dependent on its variability, should be addressed. Thus, LMP1 variants with 30-bp deletion correlate with higher tumorigenic activity and lower immunogenic potential of EBV (15). One of those polymorphisms characterizes China1, identified in two of our isolates. According to some suggestions, China1-specific changes in Human leukocyte antigen (HLA) virus epitopes (within LMP1) could enable LMP1 expression, which leads to the inability to be recognized by LMP1-specific CD8+ cytotoxic T lymphocytes (56). It would be essential to follow whether patients with these isolates would develop some EBV-related cancers. With the identification of 4 different LMP1 variants, sequences from this study showed expected diversity for this geographic area. Due to the small sample and the diversity shown, specific LMP1 variants did not suggest any association with the clinical presentation of lupus. However, the largest group consisted of NC isolates, which should be investigated further because of the earlier hypothesis that the NC variant cannot inhibit T-cell proliferation and natural killer cytotoxicity due to amino acid substitutions responsible for immunosuppressive functions (56).
When it comes to study limitations, it would be more potent if the design was prospective with longitudinal follow-up of exposed and unexposed populations from the period without disease until the moment of SLE diagnosis. A follow-up period after the diagnosis of SLE would yield valuable results too. The same could be added to monitoring the influence of individual EBV gene polymorphisms on the potential development of the underlying disease and EBV-associated tumors. However, this kind of study would require large cohorts and would last long with an unpredictable outcome. There are more aspects related to other autoimmune diseases or other viruses and novel diagnostic tools that could be addressed also: lack of EBV sequences from other autoimmune diseases, serology data for other viruses, and droplet digital PCR for detection of low concentrations of EBV. Moreover, as immunological parameters were part of routine laboratory diagnosis and monitoring of SLE patients, these data for the study were collected from the patients’ medical records. Without additional immunological testing of the healthy participants, the analysis included immunological parameters only in the SLE population.
5 Conclusion
This study obtained a comprehensive analysis of serological, molecular and sequence markers of EBV infection in SLE patients, providing additional evidence and new hypotheses about the role of EBV infection in SLE development, particularly anti-EBV immune response. The results revealed even 24 times higher possibility of having SLE if there is the presence of anti-EBV-EA(D) IgG antibodies. Additionally, higher titers of anti-EBNA1 IgG antibodies and lower titers of anti-EBV-CA IgG antibodies were also found to be independently associated with the presence of lupus. Identification of independent viral factors associated with specific lupus manifestations for the first time revealed anti-EBV-EA(D) IgG antibodies as an independent factor associated with lymphopenia and a higher titer of anti-CA IgG antibodies as an independent factor associated with alopecia. Finally, the novel data on EBV EBNA1 and LMP1 gene polymorphisms in lupus have been reported.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Ethical Board of the Faculty of Medicine, University of Belgrade (No 1550/IX-14). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AB: Conceptualization, Project administration, Supervision, Writing – original draft, Writing – review & editing. AC: Formal analysis, Methodology, Writing – review & editing. RM: Resources, Writing – review & editing. IJ: Methodology, Validation, Writing – review & editing. MG: Resources, Validation, Writing – original draft. MB: Investigation, Writing – original draft. IL: Data curation, Formal analysis, Writing – review & editing. SR: Resources, Writing – review & editing. AD: Data curation, Visualization, Writing – original draft. DM: Conceptualization, Investigation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The research was funded by the Science Fund of the Republic of Serbia, PROMIS, grant number 6060866, ROLERS.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1307589/full#supplementary-material
References
1. Harley BJ, Harley ITW, Guthridge JM, James JA. The curiously suspicious: A role for Epstein-Barr virus in lupus. Lupus (2006) 15(11):768–77. doi: 10.1177/0961203306070009
2. Sternbæk L, Draborg AH, Østerlund MT, Iversen LV, Troelsen L, Theander E, et al. Increased antibody levels to stage-specific Epstein–Barr virus antigens in systemic autoimmune diseases reveal a common pathology. Scand J Clin Lab Invest. (2019) 79(1–2):7–16. doi: 10.1080/00365513.2018.1550807
3. Balandraud N, Roudier J. Epstein-Barr virus and rheumatoid arthritis. Jt Bone Spine. (2018) 85(2):165–70. doi: 10.1016/j.jbspin.2017.04.011
4. Thorley-Lawson D. Epstein-Barr virus exploiting the immune system. Nat Rev Immunol (2001) 1(1):75–82. doi: 10.1038/35095584
5. De Paschale M. Serological diagnosis of Epstein-Barr virus infection: Problems and solutions. World J Virol (2012) 1(1):31–43. doi: 10.5501/wjv.v1.i1.31
6. Gulley ML. Review molecular diagnosis of epstein-barr virus-related diseases. J Mol Diagn. (2001) 3(1):1–10. doi: 10.1016/S1525-1578(10)60642-3
7. Wood RA, Guthridge L, Thurmond E, Guthridge CJ, Kheir JM, Bourn RL, et al. Serologic markers of Epstein-Barr virus reactivation are associated with increased disease activity, inflammation, and interferon pathway activation in patients with systemic lupus erythematosus. J Transl Autoimmun (2021) 4:100117. doi: 10.1016/j.jtauto.2021.100117
8. Afrasiabi A, Keane JT, Ong LTC, Alinejad-Rokny H, Fewings NL, Booth DR, et al. Genetic and transcriptomic analyses support a switch to lytic phase in Epstein Barr virus infection as an important driver in developing Systemic Lupus Erythematosus. J Autoimmun (2022) 127:102781. doi: 10.1016/j.jaut.2021.102781
9. Li ZX, Zeng S, Wu HX, Zhou Y. The risk of systemic lupus erythematosus associated with Epstein–Barr virus infection: a systematic review and meta-analysis. Clin Exp Med (2019) 19(1):23–36. doi: 10.1007/s10238-018-0535-0
10. Draborg AH, Lydolph MC, Westergaard M, Larsen SO, Nielsen CT, Duus K, et al. Elevated concentrations of serum immunoglobulin free light chains in systemic lupus erythematosus patients in relation to disease activity, inflammatory status, B cell activity and Epstein-Barr virus antibodies. PloS One (2015) 10(9):e0138753. doi: 10.1371/journal.pone.0138753
11. Jog NR, Young KA, Munroe ME, Harmon MT, Guthridge JM, Kelly JA, et al. Association of Epstein-Barr virus serological reactivation with transitioning to systemic lupus erythematosus in at-risk individuals. Ann Rheum Dis (2019) 78(9):1235–41. doi: 10.1136/annrheumdis-2019-215361
12. Larsen M, Sauce D, Deback C, Arnaud L, Mathian A, Miyara M, et al. Exhausted cytotoxic control of epstein-barr virus in human lupus. PloS Pathog (2011) 7(10):e1002328. doi: 10.1371/journal.ppat.1002328
13. Kang I, Quan T, Nolasco H, Park S-H, Hong MS, Crouch J, et al. Defective control of latent epstein-barr virus infection in systemic lupus erythematosus. J Immunol (2004) 172(2):1287–94. doi: 10.4049/jimmunol.172.2.1287
14. Draborg AH, Jacobsen S, Westergaard M, Mortensen S, Larsen JL, Houen G, et al. Reduced response to Epstein-Barr virus antigens by T-cells in systemic lupus erythematosus patients. Lupus Sci Med (2014) 1(1):e000015. doi: 10.1136/lupus-2014-000015
15. Knecht H, Bachmann E, Brousset P, Sandvej K, Nadal D, Bachmann F, et al. Deletions within the LMP1 oncogene of Epstein-Barr virus are clustered in Hodgkin’s disease and identical to those observed in nasopharyngeal carcinoma. Blood (1993) 82:2937–42. doi: 10.1182/blood.V82.10.2937.2937
16. Ai J, Xie Z, Liu C, Huang Z, Xu J. Analysis of EBNA-1 and LMP-1 variants in diseases associated with EBV infection in Chinese children. Virol J (2012) 9:13. doi: 10.1186/1743-422X-9-13
17. Puchhammer-Stöckl E, Görzer I. Cytomegalovirus and Epstein-Barr virus subtypes-The search for clinical significance. J Clin Virol (2006) 36(4):239–48. doi: 10.1016/j.jcv.2006.03.004
18. Banko AV, Lazarevic IB, Karalic DZ, Djukic VB, Cupic MD, Stevanovic G, et al. The sequence analysis of Epstein–Barr virus EBNA1 gene: could viral screening markers for nasopharyngeal carcinoma be identified? Med Microbiol Immunol (2019) 208(1):81–8. doi: 10.1007/s00430-018-0561-2
19. Tschochner M, Leary S, Cooper D, Strautins K, Chopra A, Clark H, et al. Identifying patient-specific Epstein-Barr Nuclear Antigen-1 genetic variation and potential autoreactive targets relevant to multiple sclerosis pathogenesis. PloS One (2016) 11(2):e0147567. doi: 10.1371/journal.pone.0147567
20. Hochberg M. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheumatol (1997) 40(9):1725. doi: 10.1002/art.1780400928
21. Matteo P, Chessa E, Morand EF, Ugarte-gil MF, Tektonidou M, Van Vollenhoven R, et al. Physician global assessment international standardisation COnsensus in systemic lupus erythematosus: the PISCOS. Lancet Rheumatol (2022) 4(6):e441–9. doi: 10.1016/S2665-9913(22)00107-2
22. Holloway L, Humphrey L, Heron L, Pilling C, Kitchen H, Højbjerre L, et al. Patient-reported outcome measures for systemic lupus erythematosus clinical trials: A review of content validity, face validity and psychometric performance. Health Qual Life Outcomes. (2014) 12:116. doi: 10.1186/s12955-014-0116-1
23. Tselios K, Gladman DD, Urowitz MB. How can we define low disease activity in systemic lupus erythematosus? Semin Arthritis Rheum (2019) 48(6):1035–40. doi: 10.1016/j.semarthrit.2018.10.013
24. Zen M, Iaccarino L, Gatto M, Saccon F, Larosa M, Ghirardello A, et al. Lupus low disease activity state is associated with a decrease in damage progression in Caucasian patients with SLE, but overlaps with remission. Ann Rheum Dis (2018) 77(1):104–10. doi: 10.1136/annrheumdis-2017-211613
25. Banko AV, Lazarevic IB, Folic MM, Djukic VB, Cirkovic AM, Karalic DZ, et al. Characterization of the variability of Epstein-Barr virus genes in nasopharyngeal biopsies: Potential predictors for carcinoma progression. PloS One (2016) 11(4):0153498. doi: 10.1371/journal.pone.0153498
26. Lorenzetti MA, Altcheh J, Moroni S, Moscatelli G, Chabay PA, Preciado MV. EBNA1 sequences in Argentinean pediatric acute and latent epstein-barr virus infection reflect circulation of novel South American variants. J Med Virol (2010) 82(10):1730–8. doi: 10.1002/jmv.21871
27. Li DJ, Bei JX, Mai SJ, Xu JF, Chen LZ, Zhang RH, et al. The dominance of China 1 in the spectrum of Epstein-Barr virus strains from cantonese patients with nasopharyngeal carcinoma. J Med Virol (2009) 81(7):1253–60. doi: 10.1002/jmv.21503
28. Hall T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95, 98, NT, 2000, XP. Nucl Acids Symp (1999) 41:95–8.
29. Edwards RH, Seillier-Moiseiwitsch F, Raab-Traub N. Signature amino acid changes in latent membrane protein 1 distinguish epstein-barr virus strains. Virology (1999) 261:79–95. doi: 10.1006/viro.1999.9855
30. Jog NR, James JA. Epstein barr virus and autoimmune responses in systemic lupus erythematosus. Front Immunol (2021) 11:623944. doi: 10.3389/fimmu.2020.623944
31. Laurynenka V, Ding L, Kaufman KM, James JA, Harley JB. A high prevalence of anti-EBNA1 heteroantibodies in systemic lupus erythematosus (SLE) supports anti-EBNA1 as an origin for SLE autoantibodies. Front Immunol (2022) 13:830993. doi: 10.3389/fimmu.2022.830993
32. Miljanovic D, Cirkovic A, Jermic I, Basaric M, Lazarevic I, Grk M, et al. Markers of epstein–barr virus infection in association with the onset and poor control of rheumatoid arthritis: A prospective cohort study. Microorganisms (2023) 11(8):1958. doi: 10.3390/microorganisms11081958
33. McClain MT, Poole BD, Bruner BF, Kaufman KM, Harley JB, James JA. An altered immune response to Epstein-Barr nuclear antigen 1 in pediatric systemic lupus erythematosus. Arthritis Rheumatol (2006) 54(1):360–8. doi: 10.1002/art.21682
34. Vista ES, Weisman MH, Ishimori ML, Chen H, Bourn RL, Bruner BF, et al. Strong viral associations with SLE among Filipinos. Lupus Sci Med (2017) 4(1):e000214. doi: 10.1136/lupus-2017-000214
35. Chougule D, Nadkar M, Rajadhyaksha A, Pandit-Shende P, Surve P, Dawkar N, et al. Association of clinical and serological parameters of systemic lupus erythematosus patients with Epstein-Barr virus antibody profile. J Med Virol (2018) 90(3):559–63. doi: 10.1002/jmv.24904
36. Parks CG, Cooper GS, Hudson LL, Dooley MA, Treadwell EL, St.Clair EW, et al. Association of Epstein-Barr virus with systemic lupus erythematosus: Effect modification by race, age, and cytotoxic T lymphocyte-associated antigen 4 genotype. Arthritis Rheumatol (2005) 52(4):1148–59. doi: 10.1002/art.20997
37. Bjornevik K, Cortese M, Healy BC, Kuhle J, Mina MJ, Leng Y, et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Sci (80- ). (2022) 375(6578):296–301. doi: 10.1126/science.abj8222
38. Quaglia M, Merlotti G, De Andrea M, Borgogna C, Cantaluppi V. Review viral infections and systemic lupus erythematosus: New players in an old story. Viruses (2021) 13(2):277. doi: 10.3390/v13020277
39. Mok CC. Current role of rituximab in systemic lupus erythematosus. Int J Rheum Dis (2015) 18(2):154–63. doi: 10.1111/1756-185X.12463
40. Draborg A, Izarzugaza JMG, Houen G. How compelling are the data for Epstein-Barr virus being a trigger for systemic lupus and other autoimmune diseases? Curr Opin Rheumatol (2016) 28(4):398–404. doi: 10.1097/BOR.0000000000000289
41. França SA de S, Viana JBG de O, Góes HCA, Fonseca RR de S, Laurentino RV, Costa IB, et al. Epidemiology of the epstein–barr virus in autoimmune inflammatory rheumatic diseases in northern Brazil. Viruses (2022) 14(4):694. doi: 10.3390/v14040694
42. Gross AJ, Hochberg D, Rand WM, Thorley-Lawson DA. EBV and systemic lupus erythematosus: A new perspective. J Immunol (2005) 174(11):6599–607. doi: 10.4049/jimmunol.174.11.6599
43. Trier NH, Holm BE, Heiden J, Slot O, Locht H, Lindegaard H, et al. Antibodies to a strain-specific citrullinated Epstein-Barr virus peptide diagnoses rheumatoid arthritis. Sci Rep (2018) 8(1):3684. doi: 10.1038/s41598-018-22058-6
44. Draborg AH, Jørgensen JM, Müller H, Nielsen CT, Jacobsen S, Iversen LV, et al. Epstein-Barr virus early antigen diffuse (EBV-EA/D)-directed immunoglobulin A antibodies in systemic lupus erythematosus patients. Scand J Rheumatol (2012) 41(4):280–9. doi: 10.3109/03009742.2012.665944
45. Ramos-Casals M, Sanz I, Bosch X, Stone JH, Khamashta MA. B-cell-depleting therapy in systemic lupus erythematosus. Am J Med (2012) 125(4):327–36. doi: 10.1016/j.amjmed.2011.09.010
46. Han L, Zhang Y, Wang Q, Xin M, Yang K, Lei K, et al. Epstein–Barr virus infection and type I interferon signature in patients with systemic lupus erythematosus. Lupus (2018) 27(6):947–54. doi: 10.1177/0961203317753069
47. Truszewska A, Wirkowska A, Gala K, Truszewski P, Krzemień-Ojak Ł, Mucha K, et al. EBV load is associated with cfDNA fragmentation and renal damage in SLE patients. Lupus (2021) 30(8):1214–25. doi: 10.1177/09612033211010339
48. Piroozmand A, Kashani HH, Zamani B. Correlation between Epstein-Barr virus infection and disease activity of systemic lupus erythematosus: A cross-sectional study. Asian Pacific J Cancer Prev (2017) 18(2):523–7. doi: 10.22034/APJCP.2017.18.2.523
49. Yu XX, Yao CW, Tao JL, Yang C, Luo MN, Li SM, et al. The expression of renal Epstein-Barr virus markers in patients with lupus nephritis. Exp Ther Med (2014) 7(5):1135–40. doi: 10.3892/etm.2014.1578
50. Bjørnevik K, Riise T, Bostrom I, Casetta I, Cortese M, Granieri E, et al. Negative interaction between smoking and EBV in the risk of multiple sclerosis: The EnvIMS study. Mult Scler. (2017) 23(7):1018–24. doi: 10.1177/1352458516671028
51. Katz BZ, Salimi B, Kim S, Nsiah-Kumi P, And BA, Wagner-Weiner L. Epstein-Barr virus burden in adolescents with systemic lupus erythematosus. Pediatr Infect Dis J (2001) 20(2):148–53. doi: 10.1097/00006454-200102000-00006
52. Gulley ML, Tang W. Laboratory assays for Epstein-Barr virus-related disease. J Mol Diagnostics. (2008) 10(4):279–92. doi: 10.2353/jmoldx.2008.080023
53. Mechelli R, Manzari C, Policano C, Annese A, Picardi E, Umeton R, et al. Epstein-Barr virus genetic variants are associated with multiple sclerosis. Neurology (2015) 84(13):1362–8. doi: 10.1212/WNL.0000000000001420
54. Masuoka S, Kusunoki N, Takamatsu R, Takahashi H, Tsuchiya K, Kawai S, et al. Epstein-Barr virus infection and variants of Epstein-Barr nuclear antigen-1 in synovial tissues of rheumatoid arthritis. PloS One (2018) 13(12):e0208957. doi: 10.1371/journal.pone.0208957
55. Gutiérrez M, Raj A, Spangler G, Sharma A, Hussain A, Judde J, et al. Sequence variations in EBNA-1 may dictate restriction of tissue distribution of Epstein-Barr virus in normal and tumour cells. J Gen Virol (1997) 78:1663–70. doi: 10.1099/0022-1317-78-7-1663
Keywords: Epstein-Barr virus (EBV), systemic lupus erythematosus (SLE), marker, EA(D) IgG, anti-EBV antibodies, EBNA1, LMP1
Citation: Banko A, Cirkovic A, Miskovic R, Jeremic I, Grk M, Basaric M, Lazarevic I, Raskovic S, Despotovic A and Miljanovic D (2023) Epstein-Barr virus infection as potential indicator of the occurrence and clinical presentation of systemic lupus erythematosus. Front. Immunol. 14:1307589. doi: 10.3389/fimmu.2023.1307589
Received: 05 October 2023; Accepted: 20 November 2023;
Published: 07 December 2023.
Edited by:
Anne-Sophie Korganow, Hôpitaux Universitaires de Strasbourg, FranceReviewed by:
Jelka Pohar, National Institute of Biology (NIB), SloveniaGerson D. Keppeke, Universidad Católica del Norte, Chile
Gunnar Houen, University of Copenhagen, Denmark
Copyright © 2023 Banko, Cirkovic, Miskovic, Jeremic, Grk, Basaric, Lazarevic, Raskovic, Despotovic and Miljanovic. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Banko, YW5hLmJhbmtvQG1lZC5iZy5hYy5ycw==