
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 29 August 2023
Sec. Viral Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1239572
 Patrick Douglas Corrêa Pereira1†
Patrick Douglas Corrêa Pereira1† Daniel Guerreiro Diniz2,3†
Daniel Guerreiro Diniz2,3† Emanuel Ramos da Costa1,3
Emanuel Ramos da Costa1,3 Nara Gyzely de Morais Magalhães1
Nara Gyzely de Morais Magalhães1 Anderson de Jesus Falcão da Silva1Jéssica Gizele Sousa Leite1
Anderson de Jesus Falcão da Silva1Jéssica Gizele Sousa Leite1 Natan Ibraim Pires Almeida1Kelle de Nazaré Cunha1
Natan Ibraim Pires Almeida1Kelle de Nazaré Cunha1 Mauro André Damasceno de Melo1Pedro Fernando da Costa Vasconcelos4,5José Antonio Picanço Diniz2
Mauro André Damasceno de Melo1Pedro Fernando da Costa Vasconcelos4,5José Antonio Picanço Diniz2 Dora Brites6,7
Dora Brites6,7 Daniel Clive Anthony8
Daniel Clive Anthony8 Cristovam Wanderley Picanço Diniz3*
Cristovam Wanderley Picanço Diniz3* Cristovam Guerreiro-Diniz1
Cristovam Guerreiro-Diniz1Normally, the host immunological response to viral infection is coordinated to restore homeostasis and protect the individual from possible tissue damage. The two major approaches are adopted by the host to deal with the pathogen: resistance or tolerance. The nature of the responses often differs between species and between individuals of the same species. Resistance includes innate and adaptive immune responses to control virus replication. Disease tolerance relies on the immune response allowing the coexistence of infections in the host with minimal or no clinical signs, while maintaining sufficient viral replication for transmission. Here, we compared the virome of bats, rodents and migratory birds and the molecular mechanisms underlying symptomatic and asymptomatic disease progression. We also explore the influence of the host physiology and environmental influences on RNA virus expression and how it impacts on the whole brain transcriptome of seemingly healthy semipalmated sandpiper (Calidris pusilla) and spotted sandpiper (Actitis macularius). Three time points throughout the year were selected to understand the importance of longitudinal surveys in the characterization of the virome. We finally revisited evidence that upstream and downstream regulation of the inflammatory response is, respectively, associated with resistance and tolerance to viral infections.
The devastating Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) pandemic has killed more than 6.88 million people worldwide [for details see COVID 19 online data https://www.worldometers.info/coronavirus/and (1, 2)]. SARS-CoV-2 was first identified in China, in December 2019 (World Health Organization, WHO, March 28, 2023). On March 11, 2020, WHO declared COVID-19 a pandemic, and on December 18, 2020, Pfizer/BioNTech vaccines and Moderna received emergency use authorization from the Food and Drug Administration (FDA). However, by this time, the virus had already caused 1.7 million deaths and had spread to 216 countries (3). Globally, there have been severe knock-on effects on the economy, owing to the restrictive measures put in place to attempt to prevent SARS-CoV-2 from spreading, which have increased unemployment, poverty, hunger, and inequalities (4), particularly in low and middle income countries (5–10).
Arguably, the extent the pandemic was controlled by the administration of non-replicating RNA-vaccines that were designed to induce immunoprotection in survivors (11–14).
If the ongoing pandemic could have been anticipated, few scientists would have expected the magnitude and speed of its spread (15) or the number of people that are still experiencing post-COVID-19 syndrome or post-acute sequelae of SARS-Cov-2 (16) and still need ongoing medical attention (17–19). These individuals regularly require the input of costly multidisciplinary teams for individualized rehabilitation and therapy (20, 21).
The Pandemic has encouraged government throughout the world to prepare public health policies to anticipate zoonotic diseases and those transmitted by arthropods, the main causes of worldwide outbreaks (22). Ongoing international efforts indicate that the most likely profile for the next pandemic will be similar to the last where another zoonotic RNA virus passes from non-human animal to humans and then to sustained human-to-human transmission (23, 24).
Understanding interspecies viral transmission and the ability to model the environmental factors that contribute to the spread of viral diseases to humans is central to the prevention of future pandemics (25). To begin to understand these processes, a broader characterization of the animal virome and an improved understanding of the host response to the presence of the virus is essential (26, 27). Unbiased metagenomic next-generation sequencing (mNGS) is helping to shed new light on virus evolution (28). mNGS applied to bats (29), migratory birds (30, 31), rodents (32) and arthropods (33) to characterize their viromes, and the transcriptomic analysis of gene expression of these large groups of non-human animals is well underway. Here we reanalyzed the virome of bats, rodents and birds to search for mechanistic insight into tolerance and resistance to virus infection. We also highlight the importance of understanding and regulating a dysfunctional innate immune system and inflammasome to control tissue damage (34).
In the wild, an enormous number of viral species circulate in specific niches and very little is known about their biology and transmission (35–39). Many of these viral species have a severe impact on livelihoods and the trade in livestock (40). The commercial trade in wildlife that is associated with hunting and consumption of wild animals has been recognized as an important route for the emergence of human zoonotic diseases (41, 42).
In a viral ‘reservoir’ species there is normally an organized immunological response to the virus, but often there is no apparent disease activity (43–46). In these situations, there is usually a low level of viral infection, and the host is able to tolerate some viral replication. Indeed, as mentioned above, many viral ‘passengers’ are carried by hosts for a lifetime (47).
The pandemic placed bat immunology in the spotlight and it has been suggested that further detailed study may help prevent future pandemics (48). Bats harbor many viruses and share human and domestic animal environments. They have a long-life expectancy and their diversity and winged locomotion favors the emergence and dispersal of new viral species (49). As bats are the only autonomous flying mammals, they are, possibly, the only non-human mammals to occupy almost all regions of the planet, except the Arctic and Antarctic poles (50).
Bats may (like any other mammal) become sick when infected by certain viral species, and the spread of new pathogens into bat colonies be fatal for them (51). However, previous studies on the diversity and evolution of viruses in bats have revealed that they exhibit unique immunological approaches to enable coexistence with viral infections, showing minimal or no overt clinical signs, while allowing enough replication for transmission (26, 52–57). Many of these bat-borne viruses induce several zoonotic human diseases (e.g. Ebola virus, Marburg virus, Nipah virus (NiV), Hendra virus (HeV), SARS-coronavirus (SARS-CoV), MERS coronavirus and (MERS-CoV) (38, 58–60) and all viral species associated with these diseases were found in apparently healthy bats (61).
Experimental infection with Egyptian rousette bats with Marburg virus (MARV) or Ebola virus followed by mRNA expression analysis, revealed the concentration of virus transcripts in the liver and many transcriptional changes in multiple tissues (62). These tissue changes included a robust overexpression of the ANXA1 gene, involved in the regulation of inflammation and cell signaling pathways (63), as well as of MRC1 (CD206) gene associated to a subset of tissue resident macrophages regulating inflammation (64). MARV severe disease includes inflammatory gene dysregulation (cytokine storm) and immune response suppression followed by systemic damage and death in humans, which is avoided in the Egyptian rousette bats by the upregulation of antiviral genes IFN-stimulated gene 15 (ISG15), IFN Induced Protein With Tetratricopeptide Repeats 1 (IFIT1) and 2′,5′-oligoadenylate synthase (OAS) 3 gene in the absence of any significant induction of proinflammatory genes, such as C-C Motif Chemokine Ligand 8 (CCL8), FAS (also called CD95 or APO-1 or TNFRSF6) and interleukin(IL)-6 (27). Because these viruses do not elicit a robust immune responses, it seems to prevent immunopathology, but it also prevents viral clearance that (65) leads to immunological tolerance. Thus, while it has been suggested that viral zoonotic risk seems to be homogeneous among mammalian and avian reservoirs hosts (66), it has been demonstrated that bats harbor significantly higher proportion of zoonotic viruses than all other mammalian orders (49, 67). A broad metagenomic analysis, recently conducted in China, to screen the virome in pharyngeal and anal swab samples of 4440 apparently healthy bats (no overt signs of disease) from 40 major different bat species revealed the presence of a diverse set of viruses (59). The most widely distributed families of mammalian viruses found were Herpesviridae, Papillomaviridae, Retroviridae, Adenoviridae and Astroviridae (~61% of the total viral sequence reads). Many reads related to other families were also found and included Circoviridae, Paramyxoviridae, Coronaviridae, Caliciviridae, Polyomaviridae, Rhabdoviridae, Hepeviridae, Bunyaviridae, Reoviridae, Flaviviridae and Picornaviridae, and the subfamily Parvovirinae. These authors identified in the swabs of a few genera, Rhinolophus spp. Miniopterus spp., and Myotis spp., 16, 10 and 13 virus families respectively, suggesting that these three genera are the major reservoirs for mammalian viruses in China (59).
These data confirmed previous findings that bats are indeed unique reservoirs of diverse virus with immune tolerance (15, 61, 68).
The ability of bats to control intracellular pathogens does not mean that they are not susceptible to extracellular infections (69). Indeed, environmental exposure of small, hibernating brown bats to Pseudogymnoascus destructans has been shown to cause infection and high mortality in colonies associated with white nose syndrome (70).
Free-living migratory birds can also be zoonotic reservoirs and can contribute to the dispersion of microorganisms as biological carriers, mechanical carriers, or carriers of infected hematophagous ectoparasites (71–73). As for bats, migratory birds also host viruses and can be the long-distance transmitters or local amplifiers of a variety of viruses that may induce severe disease in humans, domestic animals, and other wildlife (74–78).
Virome investigations of free-living birds and exploration of the impact of environmental factors on viral infection in the hosts may help to detect virus species with potential translation into human risk (79, 80). In keeping with these ideas, the virome of cloacal swab specimens collected from 3182 birds including more than 87 different species (mostly wild species), from 10 different avian orders was investigated (81). They identified 707 viral genomes from 18 defined families and 4 unclassified virus groups, with 265 virus genomes comprising new virus families, genera, or species. The authors provided evidence for the potential cross-species transmission of certain viruses in wild birds, showing > 95% amino acid sequence identity with previously reported viruses in domestic birds.
Several avian species sharing the variants of the Influenza A virus, except H17N10 and H18N11 subtypes, have been isolated exclusively from bats (82). Influenza viruses of types A and B lead to seasonal influenza epidemics, but only type A is linked to pandemics (83, 84). Migratory birds directly contribute to the maintenance and dissemination of avian influenza A virus within the Northern Atlantic flyway of North America (85).
Owing to the detection of rearrangements and mutations of the Influenza virus in wild birds, public concern about the potential pandemic risk posed by this viral species and its mutations is growing significantly (77, 86, 87). Indeed, evidence of transmission from avian viral species to humans has previously been described for A/H7N9, A/H6N1 and A/H10N8 variants, and approximately 35% of patients infected with A/H7N9 succumbed to zoonotic infection (87).
Although most people with influenza exhibit acute respiratory symptoms and muscle aches, with or without fever, which disappear within 1 week (88), avian species do not always have symptomatic infection, facilitating human exposure to the virus. Wild birds also play significant roles in the ecology and circulation of Eastern and Western Equine Encephalomyelitis and Sindbis alphaviruses, West Nile, Usutu, and St. Louis Encephalitis flaviviruses (71, 89, 90).
The inflammatory response to viral infections in migratory birds has also been well documented in several different domestic and aquatic birds (91). Gallinaceous poultry species, domestic ducks and various aquatic and terrestrial birds are vulnerable to avian influenza viruses (92–96) and host inflammatory response seems to contribute to morbidity and mortality in all species investigated so far (96, 97). Avian influenza (AI) viruses for example, have been detected in more than 105 species of birds of different taxa but the species most frequently found harboring influenza virus belong to Charadriiforms (gulls, terns and shorebirds) and Anseriformes (ducks, geese and swans) orders, where no apparent disease signs or lesions were found (97). It has been suggested that the recognition and elimination of invading pathogens (resistance)(98, 99) or the control of the infection associated tissue damage (tolerance)(100) may explain asymptomatic or minimally symptomatic infections.
Virus infections were associated with liver diseases in chickens raised in scaled farms causing significant economic losses and this was associated with multiple virus infections. Indeed, panvirome profiling of livers, spleens, kidneys, and recta revealed coinfection of chicken infectious anemia virus, avian leukemia, avian encephalomyelitis virus (AEV) and multiple fowl adenoviruses, with a higher abundance of the last two virus species in the liver (101). Many other virus infections with novel strains and variants caused economic losses to the poultry industry worldwide and these included infectious bronchitis virus (102), bursal disease virus (103), H5Nx avian influenza viruses in Europe, Africa and the Americas (104–106), H5N8 and H5N1 avian influenza viruses (107–111). Similarly, the host response to pigeon paramyxovirus type I (PPMV-1) infection (associated with New Castle disease, a great threat to the pigeon industry), induces strong innate immune responses and intense inflammatory responses at an early stage and this is associated with viral pathogenesis (112). After infection, pigeons showed upregulated expression of toll-like receptors (TLRs), such as TLR2, TLR3 and TLR15, together with interferon (IFN) gamma (IFNɣ) and IL-6, whereas IL-18 expression was found down- regulated (112). PPMV-1 is now worldwide disseminated causing extensive infections in domestic and feral pigeons, wild birds and poultry (113).
Differential host responses to avian influenza viruses in avian species with differing susceptibilities were analyzed by transcriptomics, and susceptible birds showed strong neuro-inflammatory responses associated with greater viral load (114). Ducks are natural hosts and reservoirs of influenza A virus and transcriptomic analysis in infected individuals showed this condition associated with downregulation of a distinct set of proinflammatory cytokines in lung, key elements of leukocyte recruitment and complement in intestine (115).
Comparative analysis of chickens and ducks following high pathogenic avian influenza virus infection was associated with increased proinflammatory responses in chickens whereas downregulation of inflammatory response in ducks was associated with mild or asymptomatic profiles (116). In line with these observations mallard ducks are permissive to low pathogenic avian influenza viruses in their intestinal tissues without overt disease signs, thus limiting the duration of proinflammatory cytokine expression, whereas in chickens, respiratory and systemic disease is associated with enhanced virus replication and associated tissue damage (117). While the co-evolution of Mallard ducks with influenza virus have allowed host-pathogen interaction and resistance through retinoic acid-inducible gene I (RIG-I) pathway, providing asymptomatic or minimally symptomatic infections with robust IFN response, chickens lacking the key sensors of RIG-I pathway, with compromised host response, show significant inflammatory changes, associated clinical signs and lesions to the skin, respiratory, digestive, reproductive and nervous systems following high pathogenic influenza virus (HPAIV) infections (117–119).
Rodentia is the most diverse order of mammals, representing more than 40% of mammalian species including 33 families and 2,600 species (120). They are natural virus reservoirs of a variety of species associated with human severe diseases (121–124).
At the molecular level, Cohn and collaborators performed transcriptional and physiological monitoring across 33 mouse strains during in vivo infection with the influenza virus (125). They identified two host-defense gene programs associated with disease tolerance and resistance respectively, providing a paradigm for exploring these immune responses in different species.
At the cellular level, a key subset of specialized T cells for regulation of immune responses and maintenance of the immune tolerance in the periphery, the Treg cells, are recruited and expanded, undergoing functional and molecular reprogramming (126). These changes may protect against exacerbated immune responses while providing tissue homeostasis (127).
Dual RNAseq differential expression analysis following HIN1 infection in two mouse strains with contrasting resistance to influenza virus infections enabled investigation of both the viral and host gene expression profile in the same individual and revealed that host genes were involved in the development of severe pathology, viral replication and immune responses (128). From this report it emerges that IFN Regulatory Factor 7 (Irf7) gene deletion (obtained in the knock-out line on a C57BL/6J background) was associated with greater susceptibility to H1N1 infection with significant body weight losses and increased mortality compared to wild controls (128).
Furthermore, it has been demonstrated using lymphocytic choriomeningitis and Influenza A virus models in mice, that the Treg-dependent immunosuppression is associated with the effector state of Treg cells, which maintain stability and functionality while exposed to IFNɣ. The stability of regulatory T cells may be diminished or reinforced by different Type-1 cytokines (127). The deletion restricted to regulatory T (Treg) cells of the IFNɣ receptor, but not to the IL-12 receptor, prevents Th1-type polarization and promotes Th2-type polarization, avoiding long inflammatory process, and adequate response from memory T cells (129).
It is important to highlight, however, that although many significant immunological discoveries have been obtained in laboratory mice, they do not directly mimic the physiology of wild mice living in their natural environment. To circumscribe this limitation, a new murine model named “wilding” was generated by transferring embryos of C57BL/6 into wild mice (130). This approach revealed that the immune profiles of wilding mice, laboratory mice, and wild-type mice showed that wilding individuals and laboratory mice exhibit a different epithelial barrier microbiome, gut microbiome, virome, and repertoire of pathogens. Interestingly, immunological phenotypes of wildings and wild-type mice were closer to and seemingly a better mimic of the human immune system. In agreement, the first phase 1 clinical trial in multiple laboratory mouse models of autoimmune/inflammatory diseases, using CD28 monoclonal antibodies, which are able to activate regulatory T cells, resulted in human life threatening activation of inflammatory T cells and cytokine storms (131). Similarly, laboratory mice born from wild mice (wilding mice) after treatment with a CD28-superagonist showed an inflammatory cytokine response and an absence of Treg expansion (130).
In addition to the sustained virome surveillance of bats, rodents and migratory birds, arthropods are another essential source of information for the future control of pandemics (132–134). As in mammals and birds, arthropods also react to potential threatens with an innate immune system response comprising both cellular and humoral components (135). Arthropods harbor numerous arboviruses that may spread through migratory birds and bats, at stopover sites, to other species, or to other individuals of the same species, and many of these arboviruses are of great epidemiological interest (136, 137). For example, mosquito-borne and tick-borne viruses cause some of the most severe diseases with high fatality rates in humans and animals and continue to threat human and livestock health globally (135, 138, 139).
To understand the emergence of arboviruses and the dynamics of potential outbreaks, it is essential to reconstruct the virome of the species of interest and understand its composition and potential modulatory actions on arbovirus transmission (140, 141). For example, mosquito vector competence for dengue is modulated by insect-specific viruses (142). Using virome analysis of 815 urban Aedes mosquitoes collected from 12 countries worldwide it was demonstrated that two mosquito-specific viruses were the most abundant (Phasi Charoen-like virus and Humaita Tubiacanga virus). The spatiotemporal analysis of virus circulation in an endemic urban area revealed a 200% increase in chances of having DENV in wild A. aegypti mosquitoes when both Phasi Charoen-like virus and Humaita Tubiacanga virus were present (142).
Metagenomic analysis of mosquitoes can provide valuable information, including new or unsuspected arboviruses, as well as non-arboviral pathogens ingested from hosts on which they feed (143). Next generation sequencing of RNA extracted from excreta of Aedes vigilax and Culex annulirostris reversed for cDNA and sequenced, allowed the assembly of near-full-length viromes of Australian Anopheles totivirus, Wuhan insect virus 33, and Hubei odonate virus 5 and 7 new viruses, closely related to members of the order Picornavirales and to previously described, but unclassified RNA viruses (143).
At least 500 arboviruses have been identified so far, 100 of which may induce human diseases and 40 of them cause domestic animal diseases (144, 145), and the Amazon is a spotlight for emergence of new arboviruses and this has been associated with disturbance of the natural ecosystem (146).
Emergent viruses may involve urbanization - in which humans have become the amplification hosts - and peridomestic mosquitoes (mainly Aedes aegypti) mediate human-to-human transmission (dengue, yellow fever, chikungunya and Zika viruses) (147, 148). Secondary amplification, for example, has occurred in outbreaks of Japanese encephalitis, Venezuelan equine encephalitis, and Rift Valley fever viruses (149, 150). Alternatively, simple spillover from enzootic cycles may occur as it is observed in West Nile fever (151), Crimean-Congo hemorrhagic fever (152, 153), tick-borne encephalitis (154, 155) and Alkhurma hemorrhagic fever virus (156, 157).
Vector-borne viruses enter the vector, infect, and replicate. Escaping from the gut and reaching the salivary glands, they infect vertebrate host through saliva during blood feeding (136, 158, 159). Sialotranscriptomes of unfed and fed ticks showed that blood feeding alters the expression profiles to facilitate the feeding process and pathogen transmission (160).
Human pathogens often originate from other species that act as reservoirs/hosts and from where eventual spillover and sustained human-to-human transmission takes place (161). Indeed, around 89% of the 180 recognized RNA viruses with the potential to harm humans are zoonotic (162) and the global effort for emergent infectious disease surveillance and investigation is currently in the wrong place and must migrate to hotspots in lower latitudes, such as tropical Africa, Latin America and Asia (163). The spread of humans into new ecosystems owing to the requirement for more land, climate change, loss of biodiversity, and the trade and consumption of wild animals have all enhanced the risk of transmission of these pathogens from animals to humans and increases the likelihood of pandemics (41, 164, 165).
To identify the intermediate host from where viruses may spillover, pathogen screening of animals, animal products and their movements as well as diagnostic of emerging infectious disease is required along the human-wildlife interface (157, 162). In these surveillance studies (birds, bats, rodents or arthropods viromes), the collection of ecological and physiological parameters in different environments and at different times throughout the year is also essential to understand whether a host response is resistant or tolerant of a virus (166). Indeed, host and environmental factors throughout the year influence seasonal virus transmission and highlighting the importance of longitudinal surveys at multiple time points in many hosts (167). Stopovers in bird may contribute to the recovery of constitutive immune function, which is compromised during migration (168). For example a non-stop transatlantic flight may not allow the innate immune system to recover before arrival at the final destination and may affect the ability of the bird, in this instance, to clear a virus (169).
Virome descriptions are generally based on cross-sectional space-time sampling limited to a single time point (166, 167), thus precluding how intraspecific variation and host physiology and ecology affect viral communities (170). As few reports have highlighted the importance of these influences on virome studies, as an exemplar, we show here how viral material in the brain of two long-distance migratory birds with contrasting migratory flights (Calidris pusilla and Actitis macularius) has provided an exciting opportunity to explore phenotypic variations influencing host physiology and environmental factors on the virome. The semipalmated sandpiper (C. pusilla) leaves the Bay of Fundy in Canada and arrives on the coast of South America 5-6 days later in a single non-stop flight with no opportunity to feed (171). While the Actitis macularius arrives at the wintering places with overland flights, after multiple stopovers for feeding and rest (migratory map at https://explorer.audubon.org/explore/species/1566/spotted-sandpiper/migration?sidebar=collapse).
We compared differential gene expression of viral species quantifying virus transcripts in the brain of different individuals of these trans-oceanic and trans-continental overland long-distance migratory birds. All individuals were captured within three-time windows of the wintering period. Four birds were collected between September and October (recently arrived birds), four between April and May (pre-migratory birds) and four in the intermediate time window (wintering birds). All birds were collected in Otelina Island (0°45’42.57”S and 46°55’51.86”W), in the mangrove area of the Amazon estuary at Bragança municipality, Brazil.
We searched for virus transcripts using the pipeline VIRTUS (v.1.2.1) and VIRTUS2 (v.2.0) using as reference the genome and transcriptome of Calidris pugnax (Accession code ASM143184v1) followed by NCBI Ref-Seq Viral Genomes (172).
C. pusilla and A. macularius telencephalon transcriptomes (FASTQ files) subjected to VIRTUS 2 (v.2.0) pipeline, revealed the presence of 370 and 626 virus species respectively (see Supplementary Tables S1, S2). The differential virus transcripts expressions were quantified with VIRTUS (v.1.2.1) by comparison of gene expressions in the three time points: recently arrived, intermediate and pre-migratory birds are shown in Figure 1. Table S3 indicates capture dates for each bird.
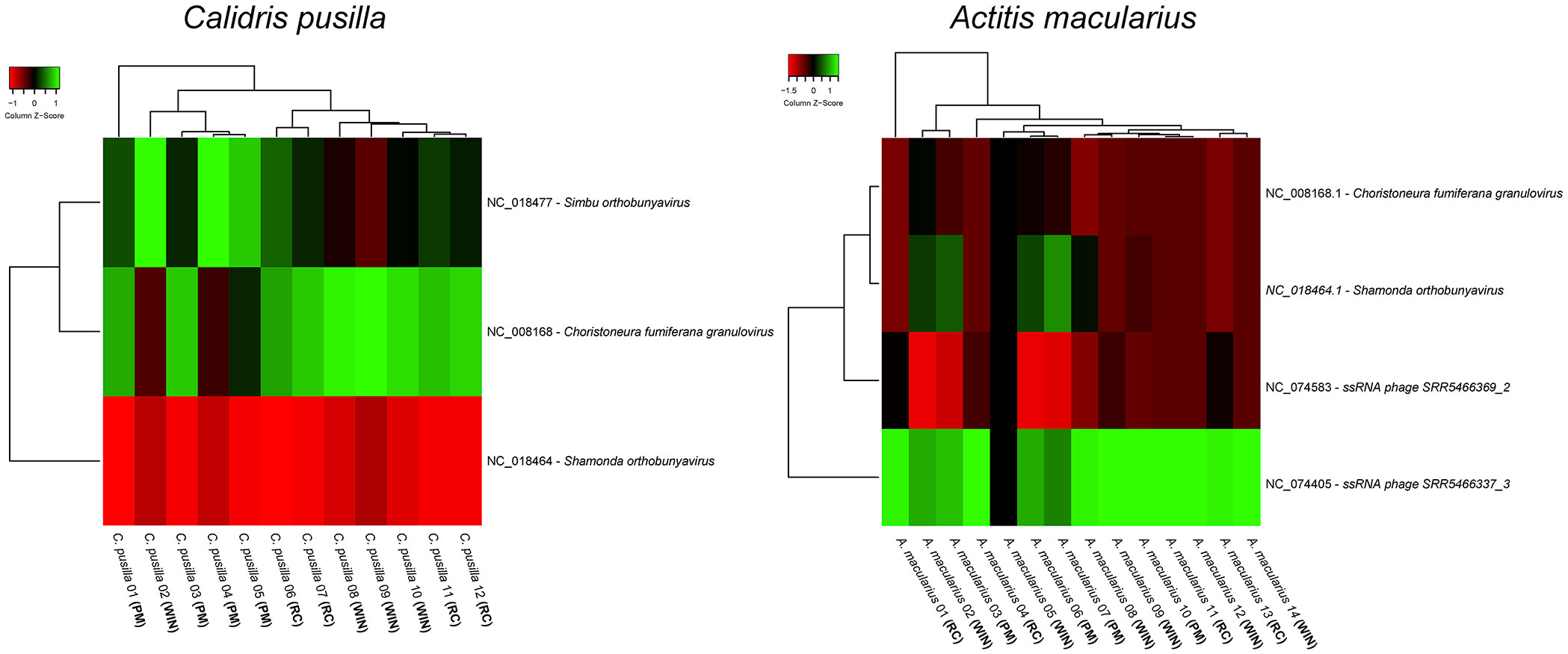
Figure 1 Differential expression of virus transcripts in the whole brain transcriptome of different birds captured at three-time windows of the wintering period: recently arrived, intermediate (wintering) and premigratory. Note the existence of marked differences in the heat maps of Actitis macularius (left) and Calidris pusilla (right). We divided the wintering period into three equal time windows: newcomers (August–November), wintering (December–March), and pre-migration (April–July). RC = recently arrived birds (captured between August and November); PM = premigratory birds (captured between April and July); WIN = wintering birds (captured between December and March).
While stopover may contribute to recover constitutive immune function, that is compromised during migration (168), non-stop transatlantic flight may not allow innate immune recovering before arrival (169).
In addition, adaptive mutations in the viral genome over time as it occurred with SARS-CoV-2, may result in the emergence of multiple variants which may alter virus interactions with host cell receptors for better or worse (173, 174).
Avoiding, resisting, or tolerating viruses are different mechanisms that ultimately must act to protect organisms from the homeostatic associated imbalance owing to infections (98, 99).
Avoidance requires that the pathogen must be detected before host infection and this may be acquired through social information and chemical signals to recognize infected conspecifics (175–177). Olfactory cues that induce aversive behavior exude from animals with acute inflammatory processes or infected, pushing away their conspecifics (178, 179). Formyl peptide receptor 3 is a G protein-coupled receptor expressed in subsets of sensory neurons of murine vomeronasal organ that act as pathogen sensor (180). Indeed, Fpr3 expression in the immune system is upregulated after lipopolysaccharide and bacterial MgrB peptide, driving avoidance behavior through the stimulation of a subset of vomeronasal sensory neurons (179).
The primary function of the immune system is, however, the recognition and elimination of invading pathogens (resistance) (98, 99) or alternatively, the control of the damage induced by a given burden (tolerance) (100).
When the pathogen is a virus species, its early detection in host cells is accompanied by activation of antiviral effector mechanisms, including type I IFN production (181, 182), natural killer (NK) cell activation (183), and B and T cell responses (184, 185).
Bats, as other eutherian mammals, synthesize anti-viral effector molecules including type I IFN, T and B cell responses, and innate responses to pathogen-associated molecular patterns (PAMPs) derived from viruses, bacteria or parasites (61, 186, 187). RNA sequencing in tissues and cells from different bats confirmed, at molecular level, these similarities (188), and this included receptors and molecules associated with innate and adaptive immunity, and microRNAs (189).
The IFN system provides the first line of defense against viral infection in vertebrates with type I IFN promoting humoral immunity (190) and both type I and III IFNs associated with the adoption of an anti-viral state in infected and neighboring cells (53, 191). However, the innate and adaptive immune systems that detect and respond to invading pathogens in eutherian mammals must be integrated to guarantee efficient protection. The role of integrating these two systems in both bats and humans belongs to the lymphocytes of the innate immune response, which share both adaptive and innate features, and are abundant in the peripheral circulation and in barrier tissues (192, 193).
Innate lymphocytes (“unconventional” or “innate-like T cells”), including innate lymphoid cells, natural killer cells, ɣδ T cells and mucosal-associated invariant T cells (194), were detected in a number of bat species (192).
Sequencing, assembly and the analysis of the genome of Rousettus aegyptiacus (Marburg virus reservoir) demonstrated the differential expansion of NK cell receptors, MHC class I genes, and type I IFN, as compared to other mammals, suggesting an inhibitory immune state (tolerance), rather than enhanced antiviral defenses (resistance) in bat immune system (195). However, many aspects of innate and adaptive immune responses of bats differ from human response, and these include a reduction of several immunoglobulin subclasses and contraction (fewer IFN genes compared with any other sequenced mammal genomes) of the type I IFN locus, and unusual constitutive expression of IFN-α in tissues and cells, unaffected by viral infection. This unique constitutive expression of IFN-stimulated subset of genes associated with anti-viral activity and resistance to DNA damage, may contribute to the coexistence of bats with viruses (53).
As previously described, humans and bats react in contrasting ways to the presence of SARS-CoV-2 (15, 196). While the inflammatory response is the major cause of deaths associated with the SARS-CoV-2 infection in humans, infected bats downregulate the genes associated with the inflammatory response avoiding its deleterious effects (26, 61, 197, 198).
Macrophages of the bat Myotis myotis and the domestic mouse (Mus musculus) when stimulated with polycytidylic acid, Poly (I:C) (ligand of toll-like receptor 3 – TLR3) mimicking a virus infection, or lipopolysaccharides from bacterial membrane (ligands of toll-like receptor 4 - TLR4) showed high levels of mRNA of IFNβ, tumor necrosis factor (TNF) and IL-1β. However, the bats exhibited sustained high-level transcription of the anti-inflammatory cytokine IL-10, which was not detected in the house mouse (197).
In addition, bats produce the innate potent IFN-inducible restriction factor tetherin that restricts the replication of diverse enveloped viruses (including retro-, filo-, herpes- and arenaviruses). As a consequence, the release of budding virions from infected cells may be inhibited (199–201) that would normally stimulate antiviral IFN response through the nuclear-factor kappa B (NF-KB) signaling pathway (202, 203).
More than two years after its emergence, the SARS-CoV-2 virus continues to affect a subgroup of patients. These patients, who have recovered from the acute phase of the disease, report a diverse repertoire of symptoms that characterize chronic or long COVID-19 (204, 205). During the acute phase, as part of the virus replicative cycle, SARS-CoV-2 induces the death and injury of virus-infected cells and tissues, followed by a wave of local inflammation associated with increased secretion of the pro-inflammatory cytokines and chemokines IL-6, IFNɣ, MCP1 (monocyte chemoattractant protein-1) and IP-10 (interferon gamma inducible protein) (206). In most individuals, the recruited cells clear the infection in the lung, the immune response subsides, and patients recover (207). However, in some patients, a dysfunctional immune response occurs, that is characterized by augmented cytokine production, that leads to widespread lung inflammation and multi-organ failure (207–209).
Indeed, severe acute respiratory syndrome induced by SARS-CoV-2 induces proinflammatory immune responses in the periphery and/or in the brain via classical TL receptor inflammatory pathway and this event is crucial for the onset of acute and chronic neuroinflammation and sickness-related behavior (210). Dysregulated immune responses resulting in lymphopenia and increased proinflammatory cytokine production seems to be the principal cause of the pathophysiology in COVID-19 and patients. High concentrations of IL-6 accompany the most severe cases (211).
The humoral immune response by B-lymphocytes is part of the adaptive immune mechanisms, controlling viral infections through the neutralizing antibodies and antibody-dependent cellular toxicity (212, 213). The elimination of viral infection and the apoptosis of virally infected cells is also provided by B lymphocytes through effector molecules, exhibiting antibody-independent functions (214). However, lack of B cells has been associated with a mild form of COVID-19 suggesting that inflammatory cytokines, especially IL-6, may have a central role in the disease severity (213). In fact, patients with agammaglobulinemia (caused by a lack of B lymphocytes), show mild symptoms, short duration, and no need of treatment by IL-6 inhibitors, while patients exhibiting variable immune deficiencies with dysfunctional B lymphocytes, (also showing significant low levels of immunoglobulins in the serum), show severe outcomes and require mechanical ventilation, antiretroviral agents and IL-6-blocking agents (215).
Survivors from acute phase may exhibit a form of post-acute sequelae characterized by atypical chronic persistence of symptoms that may last for months resulted in the long-COVID syndrome (204). Long COVID appears to be a multi-organ disease and with a spectrum of symptoms (systemic, neuropsychiatric, cardiac, and respiratory) have been associated with persistent inflammation, induced autoimmunity, and putative organ reservoirs of SARS-CoV-2 RNA or antigens (216–219). Recent evidence supports, however, the hypothesis that the persistence of the inflammatory response for a long period after the acute phase is closely correlated with the post-acute sequelae of COVID-19 (PASC). Actually, the combination of digital epidemiology with selective biobanking recent analysis suggested that instead of autoantibodies, the elevated plasma levels of IL-6, IL-1β and TNF-α is in the core of the clinical symptoms of PASC COVID-19 (220). While rheumatoid factor, antinuclear antibodies, and antiphospholipid antibodies showed no correlation with post-acute COVID-19 sequelae, analysis of blood samples from participants with ongoing post-COVID19 sequelae, revealed at 6 and 8 months after infection, increased levels of the pro-inflammatory cytokines TNF-α, IL-1β and IL-6 in close correlation with symptoms of the post-acute COVID-19 syndrome (220).
The important inflammasome sensor, NLR family pyrin domain containing protein 3 (NLRP3), has been linked to both viral-induced and age-related inflammation (221). Inflammasomes activate inflammatory caspases promoting the maturation of IL-1β and IL-18, while inducing cell death by pyroptosis (222). Dysregulation of the inflammasome is associated with several autoinflammatory syndromes and autoimmune diseases. Interestingly, ASC2 that negatively regulates inflammasome activation in both sterile and infectious settings was identified in myeloid cell genomes of 13 bat species, whereas none or minimal expression was found in most human tissues (223). Indeed, bats dampen inflammation by enhanced innate immune tolerance with implications for longevity and unique viral reservoir status (221). In contrast, the NLRP3 inflammasome was found activated in response to SARS-CoV-2 infection and identified in peripheral blood mononuclear cells (PBMCs) and postmortem tissues of COVID-19 patients (224). Moreover, NLRP3 inflammasome activation and hyperinflammation is associated with COVID-19 severity, and Pan and collaborators demonstrated that SARS-CoV-2 nucleocapsid protein can directly interact with NLRP3 to promote the assembly and activation of NLRP3 inflammasome (225). Though some inflammasome genes were also expressed in birds, little is known about the role of inflammasomes in avian responses (226). However, we may speculate that NLRP3 inflammasome is a potential target for therapy. Some similarities between the inflammasomes of humans, rodents, and other animal species suggest that the development of veterinary therapeutics may consider the inflammasome modulation by small molecules once the recombinant proteins used for human therapeutics are not economically practical. Recently, it has been demonstrated that ACS2 also suppresses SARS-CoV-2-immune-complex-induced inflammasome activation (101). In recent years, food bioactive compounds, in particular fruit and vegetables rich in flavonoids and antioxidants, as NLRP3 inflammasome modulators, have been discussed as a whole, rather than a single nutrient or functional compound (227). However, there is a lack of human data on their activity on the NLRP3 activation and there is a widespread opinion that the inhibition of the inflammasome activation may only have a minimal effect, and would only contribute more significantly if an autoimmune response is already established (228).
Virus species must introduce their RNA or DNA genomes into the infected cell to replicate, and a number of nuclei acid sensors (229) including the retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) are activated leading to convergent signaling cascade and transcriptional expression of genes encoding Type I IFNs (230, 231). RLRs are RNA sensor molecules that play essential roles in innate antiviral immunity (232) Type I IFNs bind to IFN-α/β receptor (IFNAR) which activates the Janus kinases (JAKs) and the signal transducer and activator of transcription (STAT) 1 and 2, which in turn lead to the expression of ISGs with a variety of anti-virus effects (231).
Frequent spillover of animal influenza viruses to humans, including, swine influenza viruses (233) and avian influenza virus (234) represents risks of zoonotic outbreak and pandemic (235, 236).
Birds are important reservoirs for RNA viruses and the innate defense against RNA viruses in birds involves detection of viral RNA including RIG-1 receptors, endosomal TLRs and their downstream adaptor proteins (237). Aquatic birds and shorebirds are common synanthropic species most commonly associated with avian influenza viruses (238). Because of the complete absence of any detectable RIG-I sequences in several galliform species, including the domestic fowl (Gallus gallus) (239), waterfowl and shorebirds that occasionally are associated with poultry facilities may become a source of significant damage to poultry farming (238).
Although RIG-I is involved mainly in antiviral signaling activated by RNA viruses, DNA virus infections (e.g. duck virus enteritis virus) may also increase the expression of IFN- β and RIG-I (240). RIG-I in infected cells by influenza A binds to virus genome and these single-stranded viral genomes are the natural RIG-I agonists that triggers a cascade of events leading to antiviral cytokines production (241). In contrast, the Middle East respiratory syndrome coronavirus (MERS-CoV), strongly inhibits both MDA5- and RIG-I-mediated activation of IFN-β promoter activity, while downstream signaling molecules are left largely unaffected (242, 243).
RIG-I and MDA5 (melanoma differentiation-associated gene 5), collectively known as RLRs, differentially recognized viral double-stranded RNA virus. For example, RIG-I seems to be essential for IFNs production in response to paramyxoviruses (244), influenza A virus (245) and Japanese encephalitis virus (246, 247), whereas MDA5 is critical for picornavirus detection (248) which is confirmed by high susceptibility of RIG-I-/- and MDA5-/- mice to these viral infections (249).
Because of the bat immune system has the ability to clear or maintain a number of viral infections without apparent clinical signs, bat cell lines have been used to characterize the IFN response to different viral infections in bat reservoirs (52, 250–253). Following infection with Marburg virus (MARV) and Ebola virus (EBOV) Egyptian fruit bats survived with no overt disease (254, 255) and it was demonstrated that MARV and EBOV replication was inhibited in the Egyptian fruit bat cell line R06EJ, transfected with Egyptian fruit bats innate immune genes. The overexpression of Type I, II and III IFNs, as well as DDX58 (RIG-I), IFH1 and IRF1 were associated with virus inhibition and maybe responsible for viral clearance (256).
IFNs act as extracellular cytokines activating cell surface receptors followed by intracellular kinase signal transduction and phosphorylation of transcription factors leading to the induction of ISGs allowing antiviral defense, antiproliferative activities, and stimulation of adaptive immunity (257). All type I IFNs, type II IFN (IFNɣ) and type III IFNs exhibit well-described viral inhibitory properties (258–261).
Infectious bronchitis virus (IBV) is a single strand, positive sense RNA virus (belonging to gammacoronaviruses) (262) that induces infectious bronchitis and is responsible for significant economic losses within the global poultry industry (263). Infectious bronchitis is a good example of the effects of IFN-I suppression where IBV – encoded nucleocapsid protein acts as an antagonist of IFN-I, compromising the expression of IFN-stimulated genes allowing IBV evasion from avian innate immune responses (264).
The host innate immune response mediated by IFN-1 results in the up-regulation of hundreds of ISGs and provides early protection against viral infection. Comparative studies have revealed that while many ISGs are common to all mammals, each species displays its own distinct repertoire of ISGs (265). Antiviral IFNs are responsible for the high tolerance of bats to zoonotic viruses (266), with reduced inflammatory phenotypes and bat species-specific adaptations affecting innate immune responses, where genomic and functional studies revealed unique subsets of ISGs (53, 265). For example, camelids and bats are tolerant to MERS-CoV replication and display no signs of sickness behavior. When the cervical lymph node cells from MERS-CoV convalescent llamas were pulsed with viral strains (clades B and C), although viral replication was not supported in the lymph node cells, a cellular immune response was mounted. Th1 responses (IFN-γ, IL-2, IL-12) with a transient peak of antiviral responses (type I IFNs, IFN-λ3, ISGs, PRRs and TFs). In addition, significant expression of inflammatory cytokines (TNF-α, IL-1β, IL-6, IL-8) and inflammasome components (NLRP3, CASP1, PYCARD) were suppressed (267). It was suggested that IFN-λ3 counterbalances the inflammatory processes and integrate innate and adaptive immune responses.
Recent studies in bats indicate that the unique balance between host defense and immune tolerance may explain why bats are so special virus reservoirs (15, 195, 221, 268). Previous studies demonstrate that the innate immune system of unstimulated bat tissues remain switched on due to constitutive expression of three IFN-α gene that limit viral replication without the presence of high antibodies titre (53, 65, 268). Moreover it has been demonstrated that bat microbiome from Great Himalayan Leaf-nosed bats (Hipposideros armiger) transplanted into H1N1 infected mice, reduces the inflammatory response and increases survival rate being that associated with increased production of flavonoid and isoflavones as well as with the quick innate immune response induced by the bat fecal transplanted microbiota, thus conferring mouse tolerance to influenza virus (H1N1) infection (269).
Cytokines are secreted glycoproteins that act as intercellular messengers essential for proliferation, differentiation and growth or apoptosis of the target cells, being these process initiated after binding to cell surface receptors and activation of intracellular cascades including the JAK-STAT pathway (270, 271). Most cytokines use JAK and signal transducers and activators of transcription (JAK/STAT) pathway to promote gene transcriptional regulation resulting in the expression of hematopoietic growth factors and immunomodulatory and inflammatory cytokines (271). Their signals are attenuated by multiple mechanisms and these include the suppressor cytokine signaling (SOCS) family of proteins, which is the principal negative regulation mechanism for the JAK-STAT pathway (272). An important negative-feedback inhibitor of signaling induced by cytokines that act via the JAK/STAT pathway is the SOCS family of proteins (273). When a virus infects the host cells, the innate immune receptors identify, distinguish, and react to the invader using nucleic acid or viral protein sensors (274) inducing infected cells to produce type I IFNs and proinflammatory cytokines (275). Cytokines stimulate the expression of SOCS proteins, intracellular inducible inhibitors that limit the signal magnitude of cytokines employing JAK and STAT pathways (276, 277) or target ubiquitinated signal transduction factors, avoiding potential tissue damage caused by excessive secretion of cytokines, maintaining homeostasis (275). Several viruses, however, have developed mechanisms to induce robust host SOCS protein expression increasing inhibition of protective antiviral signaling pathways allowing viruses to evade host immune response (276, 278). Indeed, the hijacking and subsequent upregulation of the SOCS proteins upon viral infection, suppress the associated JAK-STAT signaling activities, followed by reduction of the host antiviral response and viral replication (279). SOCS proteins increased expression was found in many viruses including SARS-CoV (279), herpes simplex virus (280), hepatitis B virus (281), hepatitis C virus (282), Zika virus (283), respiratory syncytial virus (284) and influenza A virus (285).
Overexpression of SOCS1 leads to a decrease in phosphorylation levels of JAK1, tyrosine kinase 2 (TYK2) while also inhibits antiviral and antiproliferative responses induced by IFN-I. Consistent with this, SOCS1 ablated cells and SOCS1−/− mice are resistant to viral infection (286; 275). Similarly, porcine reproductive and respiratory syndrome virus (PRRSV) (major threaten to global swine industry) increases SOCS3 production via activation of p38/AP-1 signaling pathway to promote viral replication and persistent infection (287).To obtain a favorable outcome for viral infections, avoiding the installation of an inflammatory pathology with eventual fatal evolution, the host must have strict control of its expression (288, 289). For this, the innate immune system recognizes molecules that identify many pathogens associated molecular patterns (PAMPs) or damaged associated molecular patterns (DAMPs) and initiates a rapid response producing inflammatory mediators that activate programmed cell death pathways including pyroptosis, apoptosis and necroptosis with significant crosstalk among them (290–292).
Few scientists would have expected the magnitude and speed of the spread of the COVID-19 pandemic. While the deaths, rightly, have attracted the headlines, an unknown number of carriers of post-acute sequelae of SARS-CoV-2 still require medical attention. To anticipate pandemics in the future and avoid unnecessary morbidity and mortality, it has been suggested that a broader characterization of the animal virome, host physiology and the underlying mechanisms associated with disease tolerance and anti-viral defenses are essential. We reviewed the literature concerning the virome of bats, rodents and migratory birds and highlighted the potential mechanisms of tolerance and resistance to virus infection in these groups. It is now clear that the type of host inflammatory response to the presence of virus is central to these mechanisms. We also emphasize the importance of understanding and controlling the dysfunctional innate immune system and inflammasome to deal with future outbreaks in the period before we have effective vaccines.
CD, CG-D, PP, DD, NM and MM contributed to conception and design of the study. PP organized the database and performed statistical analysis. DD optimized figures and adapted submission to the journal requirements. CD wrote the first draft. DB wrote section of the manuscript. CD, DA, DD and DB contributed to final edition. All authors contributed to the article and approved the submitted version.
CD was supported by Brazilian National Research Council - CNPq Process No 407075/2021-6 and No. 301268/2019-3, UFPA/EBSERH and UFPA/FINEP/FADESPSOS Equipment 2021 and AMT was supported by CAPES doctoral fellowship. PFCV was supported by the National Institute for Science and Technology for Emerging and Reemerging Viruses (INCT-VER grant CNPq process 406360/2022-7) and by CNPq process 310295/2021-1. DB was supported by Fundação para a Ciência e Tecnologia (FCT-PTDC/MED-NEU/2382/2021, LISBOA-01-0145-FEDER-031395, UIDB/04138/2020 and UIDP/04138/2020). Programa de Apoio à publicação qualificada - PROPESP/UFPA (PAPQ). DD was supported by the Brazilian National Research Council-CNPq (PDS fellowship CNPq Proc No.168555(2021-2)
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1239572/full#supplementary-material
1. Kaul R, Devi S. Coronavirus - A crippling affliction to humans. Recent Pat Biotechnol (2022) 16:226–42. doi: 10.2174/1872208316666220404103033
2. Akbar SMF, Al Mahtab M, Khan S. Cellular and molecular mechanisms of pathogenic and protective immune responses to SARS-CoV-2 and implications of COVID-19 vaccines. Vaccines (Basel) (2023) 11. doi: 10.3390/vaccines11030615
3. Meo SA, Bukhari IA, Akram J, Meo AS, Klonoff DC. COVID-19 vaccines: comparison of biological, pharmacological characteristics and adverse effects of Pfizer/BioNTech and Moderna Vaccines. Eur Rev Med Pharmacol Sci (2021) 25:1663–9. doi: 10.26355/eurrev_202102_24877
4. Schippers MC, Ioannidis JPA, Joffe AR. Aggressive measures, rising inequalities, and mass formation during the COVID-19 crisis: An overview and proposed way forward. Front Public Health (2022) 10:950965. doi: 10.3389/fpubh.2022.950965
5. Onyeaka H, Anumudu CK, Al-Sharify ZT, Egele-Godswill E, Mbaegbu P. COVID-19 pandemic: A review of the global lockdown and its far-reaching effects. Sci Prog (2021) 104:368504211019854. doi: 10.1177/00368504211019854
6. Maredia MK, Adenikinju A, Belton B, Chapoto A, Faye NF, Liverpool-Tasie S, et al. COVID-19's impacts on incomes and food consumption in urban and rural areas are surprisingly similar: Evidence from five African countries. Glob Food Sec (2022) 33:100633. doi: 10.1016/j.gfs.2022.100633
7. Mishrif A, Khan A. Examining the effects of COVID-19 on logistics and supply chain networks in Oman. J Emerg Manag (2023) 21:289–309. doi: 10.5055/jem.0721
8. Ogier EM, Smith DC, Breen S, Gardner C, Gaughan DJ, Gorfine HK, et al. Initial impacts of the COVID-19 pandemic on Australian fisheries production, research organisations and assessment: shocks, responses and implications for decision support and resilience. Rev Fish Biol Fish (2023) 33:513–34. doi: 10.1007/s11160-023-09760-z
9. Takyi PO, Dramani JB, Akosah NK, Aawaar G. Economic activities' response to the COVID-19 pandemic in developing countries. Sci Afr (2023) 20:e01642. doi: 10.1016/j.sciaf.2023.e01642
10. Yeboah H, Yaya S. Health and economic implications of the ongoing coronavirus disease (COVID-19) pandemic on women and children in Africa. Reprod Health (2023) 20:70. doi: 10.1186/s12978-023-01616-w
11. Rauch S, Jasny E, Schmidt KE, Petsch B. New vaccine technologies to combat outbreak situations. Front Immunol (2018) 9:1963. doi: 10.3389/fimmu.2018.01963
12. Pollard AJ, Bijker EM. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol (2021) 21:83–100. doi: 10.1038/s41577-020-00479-7
13. Szabó GT, Mahiny AJ, Vlatkovic I. COVID-19 mRNA vaccines: Platforms and current developments. Mol Ther (2022) 30:1850–68. doi: 10.1016/j.ymthe.2022.02.016
14. Guo X, Liu D, Huang Y, Deng Y, Wang Y, Mao J, et al. Revolutionizing viral disease vaccination: the promising clinical advancements of non-replicating mRNA vaccines. Virol J (2023) 20:64. doi: 10.1186/s12985-023-02023-0
15. Irving AT, Ahn M, Goh G, Anderson DE, Wang LF. Lessons from the host defences of bats, a unique viral reservoir. Nature (2021) 589:363–70. doi: 10.1038/s41586-020-03128-0
16. Merad M, Blish CA, Sallusto F, Iwasaki A. The immunology and immunopathology of COVID-19. Science (2022) 375:1122–7. doi: 10.1126/science.abm8108
17. Lechner-Scott J, Levy M, Hawkes C, Yeh A, Giovannoni G. Long COVID or post COVID-19 syndrome. Mult Scler Relat Disord (2021) 55:103268. doi: 10.1016/j.msard.2021.103268
18. Ceban F, Ling S, Lui LMW, Lee Y, Gill H, Teopiz KM, et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav Immun (2022) 101:93–135. doi: 10.1016/j.bbi.2021.12.020
19. Desai AD, Lavelle M, Boursiquot BC, Wan EY. Long-term complications of COVID-19. Am J Physiol Cell Physiol (2022) 322:C1–C11. doi: 10.1152/ajpcell.00375.2021
20. Greenhalgh T, Knight M, A'Court C, Buxton M, Husain L. Management of post-acute covid-19 in primary care. BMJ (2020) 370:m3026. doi: 10.1136/bmj.m3026
21. Venkatesan P. NICE guideline on long COVID. Lancet Respir Med (2021) 9:129. doi: 10.1016/S2213-2600(21)00031-X
22. Gazal S, Sharma N, Tikoo M, Shikha D, Badroo GA, Rashid M, et al. Nipah and hendra viruses: Deadly zoonotic paramyxoviruses with the potential to cause the next pandemic. Pathogens (2022) 11. doi: 10.3390/pathogens11121419
23. Thompson AJ, Paulson JC. Adaptation of influenza viruses to human airway receptors. J Biol Chem (2021) 296:100017. doi: 10.1074/jbc.REV120.013309
24. Farlow A, Torreele E, Gray G, Ruxrungtham K, Rees H, Prasad S, et al. The future of epidemic and pandemic vaccines to serve global public health needs. Vaccines (Basel) (2023) 11. doi: 10.3390/vaccines11030690
25. Albery GF, Eskew EA, Ross N, Olival KJ. Predicting the global mamMalian viral sharing network using phylogeography. Nat Commun (2020) 11:2260. doi: 10.1038/s41467-020-16153-4
26. Prescott J, Guito JC, Spengler JR, Arnold CE, Schuh AJ, Amman BR, et al. Rousette bat dendritic cells overcome marburg virus-mediated antiviral responses by upregulation of interferon-related genes while downregulating proinflammatory disease mediators. mSphere (2019) 4. doi: 10.1128/mSphere.00728-19
27. Guito JC, Prescott JB, Arnold CE, Amman BR, Schuh AJ, Spengler JR, et al. Asymptomatic infection of marburg virus reservoir bats is explained by a strategy of immunoprotective disease tolerance. Curr Biol (2021) 31:257–270.e255. doi: 10.1016/j.cub.2020.10.015
28. Harvey E, Holmes EC. Diversity and evolution of the animal virome. Nat Rev Microbiol (2022) 20:321–34. doi: 10.1038/s41579-021-00665-x
29. Van Brussel K, Holmes EC. Zoonotic disease and virome diversity in bats. Curr Opin Virol (2022) 52:192–202. doi: 10.1016/j.coviro.2021.12.008
30. Vibin J, Chamings A, Collier F, Klaassen M, Nelson TM, Alexandersen S. Metagenomics detection and characterisation of viruses in faecal samples from Australian wild birds. Sci Rep (2018) 8:8686. doi: 10.1038/s41598-018-26851-1
31. Meng W, Yang Q, Vrancken B, Chen Z, Liu D, Chen L, et al. New evidence for the east-west spread of the highly pathogenic avian influenza H5N1 virus between Central Asian and east Asian-Australasian flyways in China. Emerg Microbes Infect (2019) 8:823–6. doi: 10.1080/22221751.2019.1623719
32. Wu Z, Lu L, Du J, Yang L, Ren X, Liu B, et al. Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases. Microbiome (2018) 6:178. doi: 10.1186/s40168-018-0554-9
33. Ribeiro GO, Monteiro FJC, Rego MODS, Ribeiro ESD, Castro DF, Caseiro MM, et al. Detection of RNA-dependent RNA polymerase of hubei reo-like virus 7 by next-generation sequencing in. Viruses (2019) 11. doi: 10.3390/v11020147
34. Ayres JS. Surviving COVID-19: A disease tolerance perspective. Sci Adv (2020) 6:eabc1518. doi: 10.1126/sciadv.abc1518
35. Daniels PW, Halpin K, Hyatt A, Middleton D. Infection and disease in reservoir and spillover hosts: determinants of pathogen emergence. Curr Top Microbiol Immunol (2007) 315:113–31. doi: 10.1007/978-3-540-70962-6_6
36. Mackenzie JS, Jeggo M. Reservoirs and vectors of emerging viruses. Curr Opin Virol (2013) 3:170–9. doi: 10.1016/j.coviro.2013.02.002
37. Hemmatzadeh F, Boardman W, Alinejad A, Hematzade A, Moghadam MK. Molecular and serological survey of selected viruses in free-ranging wild ruminants in Iran. PloS One (2016) 11:e0168756. doi: 10.1371/journal.pone.0168756
38. Wang LF, Anderson DE. Viruses in bats and potential spillover to animals and humans. Curr Opin Virol (2019) 34:79–89. doi: 10.1016/j.coviro.2018.12.007
39. Celone M, Okech B, Han BA, Forshey BM, Anyamba A, Dunford J, et al. A systematic review and meta-analysis of the potential non-human animal reservoirs and arthropod vectors of the Mayaro virus. PLoS Negl Trop Dis (2021) 15:e0010016. doi: 10.1371/journal.pntd.0010016
40. Jones BA, Mahapatra M, Mdetele D, Keyyu J, Gakuya F, Eblate E, et al. Peste des Petits Ruminants Virus Infection at the Wildlife-Livestock Interface in the Greater Serengeti Ecosystem 2015-2019. Viruses (2021) 13. doi: 10.3390/v13050838
41. Keatts LO, Robards M, Olson SH, Hueffer K, Insley SJ, Joly DO, et al. Implications of zoonoses from hunting and use of wildlife in North American Arctic and boreal biomes: Pandemic potential, monitoring, and mitigation. Front Public Health (2021) 9:627654. doi: 10.3389/fpubh.2021.627654
42. Petrovan SO, Aldridge DC, Bartlett H, Bladon AJ, Booth H, Broad S, et al. Post COVID-19: a solution scan of options for preventing future zoonotic epidemics. Biol Rev Camb Philos Soc (2021) 96:2694–715. doi: 10.1111/brv.12774
43. Easterbrook JD, Klein SL. Immunological mechanisms mediating hantavirus persistence in rodent reservoirs. PLoS Pathog (2008) 4:e1000172. doi: 10.1371/journal.ppat.1000172
44. Watanabe S, Masangkay JS, Nagata N, Morikawa S, Mizutani T, Fukushi S, et al. Bat coronaviruses and experimental infection of bats, the Philippines. Emerg Infect Dis (2010) 16:1217–23. doi: 10.3201/eid1608.100208
45. Munster VJ, Adney DR, van Doremalen N, Brown VR, Miazgowicz KL, Milne-Price S, et al. Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis). Sci Rep (2016) 6:21878. doi: 10.1038/srep21878
46. Williams EP, Spruill-Harrell BM, Taylor MK, Lee J, Nywening AV, Yang Z, et al. Common themes in zoonotic spillover and disease emergence: Lessons learned from bat- and rodent-borne RNA viruses. Viruses (2021) 13. doi: 10.3390/v13081509
47. Virgin HW, Wherry EJ, Ahmed R. Redefining chronic viral infection. Cell (2009) 138:30–50. doi: 10.1016/j.cell.2009.06.036
48. Mallapaty S. Bats play host to a horde of nasty viruses - can studying their immunity help stop pandemics? Nature (2023) 615:576–9. doi: 10.1038/d41586-023-00791-x
49. Dutheil F, Clinchamps M, Bouillon-Minois JB. Bats, pathogens, and species richness. Pathogens (2021) 10. doi: 10.3390/pathogens10020098
50. Mann DL. SARS-CoV-2 and bats: From flight to fighting COVID-19. JACC Basic Transl Sci (2020) 5:545–6. doi: 10.1016/j.jacbts.2020.04.012
51. Mühldorfer K, Speck S, Kurth A, Lesnik R, Freuling C, Müller T, et al. Diseases and causes of death in European bats: dynamics in disease susceptibility and infection rates. PLoS One (2011) 6:e29773. doi: 10.1371/journal.pone.0029773
52. Glennon NB, Jabado O, Lo MK, Shaw ML. Transcriptome profiling of the virus-induced innate immune response in pteropus vampyrus and its attenuation by nipah virus interferon antagonist functions. J Virol (2015) 89:7550–66. doi: 10.1128/JVI.00302-15
53. Zhou P, Tachedjian M, Wynne JW, Boyd V, Cui J, Smith I, et al. Contraction of the type I IFN locus and unusual constitutive expression of IFN-α in bats. Proc Natl Acad Sci U S A (2016) 113:2696–701. doi: 10.1073/pnas.1518240113
54. Xie J, Li Y, Shen X, Goh G, Zhu Y, Cui J, et al. Dampened STING-dependent interferon activation in bats. Cell Host Microbe (2018) 23:297–301.e294. doi: 10.1016/j.chom.2018.01.006
55. Mendenhall IH, Wen DLH, Jayakumar J, Gunalan V, Wang L, Mauer-Stroh S, et al. Diversity and evolution of viral pathogen community in cave nectar bats. Viruses (2019) 11. doi: 10.3390/v11030250
56. Tan CW, Yang X, Anderson DE, Wang LF. Bat virome research: the past, the present and the future. Curr Opin Virol (2021) 49:68–80. doi: 10.1016/j.coviro.2021.04.013
57. Wiederkehr MA, Qi W, Schoenbaechler K, Fraefel C, Kubacki J. Virus diversity, abundance, and evolution in three different bat colonies in Switzerland. Viruses (2022) 14. doi: 10.3390/v14091911
58. Han HJ, Wen HL, Zhou CM, Chen FF, Luo LM, Liu JW, et al. Bats as reservoirs of severe emerging infectious diseases. Virus Res (2015) 205:1–6. doi: 10.1016/j.virusres.2015.05.006
59. Wu Z, Yang L, Ren X, He G, Zhang J, Yang J, et al. Deciphering the bat virome catalog to better understand the ecological diversity of bat viruses and the bat origin of emerging infectious diseases. ISME J (2016) 10:609–20. doi: 10.1038/ismej.2015.138
60. Letko M, Seifert SN, Olival KJ, Plowright RK, Munster VJ. Bat-borne virus diversity, spillover and emergence. Nat Rev Microbiol (2020) 18:461–71. doi: 10.1038/s41579-020-0394-z
61. Banerjee A, Baker ML, Kulcsar K, Misra V, Plowright R, Mossman K. Novel insights into immune systems of bats. Front Immunol (2020) 11:26. doi: 10.3389/fimmu.2020.00026
62. Jayaprakash AD, Ronk AJ, Prasad AN, Covington MF, Stein KR, Schwarz TM, et al. Marburg and ebola virus infections elicit a complex, muted inflammatory state in bats. Viruses (2023) 15. doi: 10.3390/v15020350
63. Han PF, Che XD, Li HZ, Gao YY, Wei XC, Li PC. Annexin A1 involved in the regulation of inflammation and cell signaling pathways. Chin J Traumatol (2020) 23:96–101. doi: 10.1016/j.cjtee.2020.02.002
64. Ibrahim AM, Moss MA, Gray Z, Rojo MD, Burke CM, Schwertfeger KL, et al. Diverse macrophage populations contribute to the inflammatory microenvironment in preMalignant lesions during localized invasion. Front Oncol (2020) 10:569985. doi: 10.3389/fonc.2020.569985
65. Schountz T, Baker ML, Butler J, Munster V. Immunological control of viral infections in bats and the emergence of viruses highly pathogenic to humans. Front Immunol (2017) 8:1098. doi: 10.3389/fimmu.2017.01098
66. Mollentze N, Streicker DG. Viral zoonotic risk is homogenous among taxonomic orders of mamMalian and avian reservoir hosts. Proc Natl Acad Sci U S A (2020) 117:9423–30. doi: 10.1073/pnas.1919176117
67. Olival KJ, Hosseini PR, Zambrana-Torrelio C, Ross N, Bogich TL, Daszak P. Host and viral traits predict zoonotic spillover from mammals. Nature (2017) 546:646–50. doi: 10.1038/nature22975
68. Skirmuntt EC, Escalera-Zamudio M, Teeling EC, Smith A, Katzourakis A. The potential role of endogenous viral elements in the evolution of bats as reservoirs for zoonotic viruses. Annu Rev Virol (2020) 7:103–19. doi: 10.1146/annurev-virology-092818-015613
69. Brook CE, Dobson AP. Bats as 'special' reservoirs for emerging zoonotic pathogens. Trends Microbiol (2015) 23:172–80. doi: 10.1016/j.tim.2014.12.004
70. Hicks AC, Darling SR, Flewelling JE, von Linden R, Meteyer CU, Redell DN, et al. Environmental transmission of Pseudogymnoascus destructans to hibernating little brown bats. Sci Rep (2023) 13:4615. doi: 10.1038/s41598-023-31515-w
71. Hubálek Z. An annotated checklist of pathogenic microorganisms associated with migratory birds. J Wildl Dis (2004) 40:639–59. doi: 10.7589/0090-3558-40.4.639
72. Siljic M, Sehovic R, Jankovic M, Stamenkovic G, Loncar A, Todorovic M, et al. Evolutionary dynamics of Usutu virus: Worldwide dispersal patterns and transmission dynamics in Europe. Front Microbiol (2023) 14:1145981. doi: 10.3389/fmicb.2023.1145981
73. Wang Y, Wang M, Zhang H, Zhao C, Zhang Y, Shen J, et al. Prevalence, evolution, replication and transmission of H3N8 avian influenza viruses isolated from migratory birds in eastern China from 2017 to 2021. Emerg Microbes Infect (2023) 12:2184178. doi: 10.1080/22221751.2023.2184178
74. Reperant LA, Kuiken T, Osterhaus AD. Influenza viruses: from birds to humans. Hum Vaccin Immunother (2012) 8:7–16. doi: 10.4161/hv.8.1.18672
75. Verhagen JH, van Dijk JG, Vuong O, Bestebroer T, Lexmond P, Klaassen M, et al. Migratory birds reinforce local circulation of avian influenza viruses. PLoS One (2014) 9:e112366. doi: 10.1371/journal.pone.0112366
76. Chan JF, To KK, Chen H, Yuen KY. Cross-species transmission and emergence of novel viruses from birds. Curr Opin Virol (2015) 10:63–9. doi: 10.1016/j.coviro.2015.01.006
77. Wang Y, Wang M, Zhang H, Zhao C, Zhang Y, He G, et al. Emergence, evolution, and biological characteristics of H10N4 and H10N8 avian influenza viruses in migratory wild birds detected in Eastern China in 2020. Microbiol Spectr (2022) 10:e0080722. doi: 10.1128/spectrum.00807-22
78. Ortiz L, Geiger G, Ferreri L, Moran D, Mendez D, Gonzalez-Reiche AS, et al. Blue-winged teals in Guatemala and their potential role in the ecology of H14 subtype influenza a viruses. Viruses (2023) 15. doi: 10.3390/v15020483
79. Rahman MM, Talukder A, Chowdhury MMH, Talukder R, Akter R. Coronaviruses in wild birds - A potential and suitable vector for global distribution. Vet Med Sci (2021) 7(1):264–72. doi: 10.1002/vms3.360
80. Wang YXG, Voutilainen L, Aminikhah M, Helle H, Huitu O, Laakkonen J, et al. The impact of wildlife and environmental factors on hantavirus infection in the host and its translation into human risk. Proc Biol Sci (2023) 290:20222470. doi: 10.1098/rspb.2022.2470
81. Shan T, Yang S, Wang H, Zhang J, Gong G, Xiao Y, et al. Virome in the cloaca of wild and breeding birds revealed a diversity of significant viruses. Microbiome (2022) 10:60. doi: 10.1186/s40168-022-01246-7
82. Shi Y, Wu Y, Zhang W, Qi J, Gao GF. Enabling the 'host jump': structural determinants of receptor-binding specificity in influenza A viruses. Nat Rev Microbiol (2014) 12:822–31. doi: 10.1038/nrmicro3362
83. Yoon SW, Webby RJ, Webster RG. Evolution and ecology of influenza A viruses. Curr Top Microbiol Immunol (2014) 385:359–75. doi: 10.1007/82_2014_396
84. Wille M, Holmes EC. The ecology and evolution of influenza viruses. Cold Spring Harb Perspect Med (2020) 10. doi: 10.1101/cshperspect.a038489
85. Prosser DJ, Chen J, Ahlstrom CA, Reeves AB, Poulson RL, Sullivan JD, et al. Maintenance and dissemination of avian-origin influenza A virus within the northern Atlantic Flyway of North America. PLoS Pathog (2022) 18:e1010605. doi: 10.1371/journal.ppat.1010605
86. Kim SM, Kim YI, Pascua PN, Choi YK. Avian influenza A viruses: Evolution and zoonotic infection. Semin Respir Crit Care Med (2016) 37:501–11. doi: 10.1055/s-0036-1584953
87. Li YT, Linster M, Mendenhall IH, Su YCF, Smith GJD. Avian influenza viruses in humans: lessons from past outbreaks. Br Med Bull (2019) 132:81–95. doi: 10.1093/bmb/ldz036
88. Uyeki TM, Hui DS, Zambon M, Wentworth DE, Monto AS. Influenza. Lancet (2022) 400:693–706. doi: 10.1016/S0140-6736(22)00982-5
89. Michel F, Sieg M, Fischer D, Keller M, Eiden M, Reuschel M, et al. Evidence for West Nile virus and usutu virus infections in wild and resident birds in Germany 2017 and 2018. Viruses (2019) 11. doi: 10.3390/v11070674
90. Mancuso E, Cecere JG, Iapaolo F, Di Gennaro A, Sacchi M, Savini G, et al. West nile and usutu virus introduction via migratory birds: A retrospective analysis in Italy. Viruses (2022) 14. doi: 10.3390/v14020416
91. Wang Z, Loh L, Kedzierski L, Kedzierska K. Avian influenza viruses, inflammation, and CD8(+) T cell immunity. Front Immunol (2016) 7:60. doi: 10.3389/fimmu.2016.00060
92. Normile D. Avian influenza. Evidence points to migratory birds in H5N1 spread. Science (2006) 311:1225. doi: 10.1126/science.311.5765.1225
93. Munster VJ, Baas C, Lexmond P, Waldenström J, Wallensten A, Fransson T, et al. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog (2007) 3:e61. doi: 10.1371/journal.ppat.0030061
94. Verhagen JH, Herfst S, Fouchier RA. Infectious disease. How a virus travels the world. Science (2015) 347:616–7. doi: 10.1126/science.aaa6724
95. Verhagen JH, Höfle U, van Amerongen G, van de Bildt M, Majoor F, Fouchier RA, et al. Long-term effect of serial infections with H13 and H16 low-pathogenic avian influenza viruses in black-headed gulls. J Virol (2015) 89:11507–22. doi: 10.1128/JVI.01765-15
96. Mishra A, Vijayakumar P, Raut AA. Emerging avian influenza infections: Current understanding of innate immune response and molecular pathogenesis. Int Rev Immunol (2017) 36:89–107. doi: 10.1080/08830185.2017.1291640
97. França MS, Brown JD. Influenza pathobiology and pathogenesis in avian species. Curr Top Microbiol Immunol (2014) 385:221–42. doi: 10.1007/82_2014_385
98. Medzhitov R, Schneider DS, Soares MP. Disease tolerance as a defense strategy. Science (2012) 335:936–41. doi: 10.1126/science.1214935
99. Rivas FV, Chervonsky AV, Medzhitov R. ART and immunology. Trends Immunol (2014) 35:451. doi: 10.1016/j.it.2014.09.002
100. Råberg L, Graham AL, Read AF. Decomposing health: tolerance and resistance to parasites in animals. Philos Trans R Soc Lond B Biol Sci (2009) 364:37–49. doi: 10.1098/rstb.2008.0184
101. Ahn M, Chen VC, Rozario P, Ng WL, Kong PS, Sia WR, et al. Bat ASC2 suppresses inflammasomes and ameliorates inflammatory diseases. Cell (2023) 186:2144–2159.e2122. doi: 10.1016/j.cell.2023.03.036
102. Wang C, Hou B. A pathogenic and recombinant infectious bronchitis virus variant (CK/CH/GX/202109) with multiorgan tropism. Vet Res (2023) 54:54. doi: 10.1186/s13567-023-01182-w
103. Li K, Niu X, Jiang N, Zhang W, Wang G, Huang M, et al. Comparative pathogenicity of three strains of infectious bursal disease virus closely related to poultry industry. Viruses (2023) 15. doi: 10.3390/v15061257
104. Lee DH, Bertran K, Kwon JH, Swayne DE. Evolution, global spread, and pathogenicity of highly pathogenic avian influenza H5Nx clade 2.3.4.4. J Vet Sci (2017) 18:269–80. doi: 10.4142/jvs.2017.18.S1.269
105. Engelsma M, Heutink R, Harders F, Germeraad EA, Beerens N. Multiple introductions of reassorted highly pathogenic avian influenza H5Nx viruses clade 2.3.4.4b causing outbreaks in wild birds and poultry in the Netherlands 2020-2021. Microbiol Spectr (2022) 10:e0249921. doi: 10.1128/spectrum.02499-21
106. Kwon JH, Bertran K, Lee DH, Criado MF, Killmaster L, Pantin-Jackwood MJ, et al. Diverse infectivity, transmissibility, and pathobiology of clade 2.3.4.4 H5Nx highly pathogenic avian influenza viruses in chickens. Emerg Microbes Infect (2023) 12:2218945. doi: 10.1080/22221751.2023.2218945
107. Rehman S, Effendi MH, Witaningruma AM, Nnabuikeb UE, Bilal M, Abbas A, et al. Avian influenza (H5N1) virus, epidemiology and its effects on backyard poultry in Indonesia: a review. F1000Res (2022) 11:1321. doi: 10.12688/f1000research.125878.1
108. Dey P, Ahuja A, Panwar J, Choudhary P, Rani S, Kaur M, et al. Immune control of avian influenza virus infection and its vaccine development. Vaccines (Basel) (2023) 11. doi: 10.3390/vaccines11030593
109. Ntakiyisumba E, Lee S, Park BY, Tae HJ, Won G. Prevalence, seroprevalence and risk factors of avian influenza in wild bird populations in Korea: A systematic review and meta-analysis. Viruses (2023) 15. doi: 10.3390/v15020472
110. Takadate Y, Tsunekuni R, Kumagai A, Mine J, Kikutani Y, Sakuma S, et al. Different infectivity and transmissibility of H5N8 and H5N1 high pathogenicity avian influenza viruses isolated from chickens in Japan in the 2021/2022 season. Viruses (2023) 15. doi: 10.3390/v15020265
111. Yang XY, Gong QL, Li YJ, Ata EB, Hu MJ, Sun YY, et al. The global prevalence of highly pathogenic avian influenza A (H5N8) infection in birds: A systematic review and meta-analysis. Microb Pathog (2023) 176:106001. doi: 10.1016/j.micpath.2023.106001
112. Wang F, Gao M, Han Z, Hou Y, Zhang L, Ma Z, et al. Innate immune responses of domestic pigeons to the infection of pigeon paramyxovirus type 1 virus. Poult Sci (2021) 100:603–14. doi: 10.1016/j.psj.2020.11.045
113. Aldous EW, Fuller CM, Ridgeon JH, Irvine RM, Alexander DJ, Brown IH. The evolution of pigeon paramyxovirus type 1 (PPMV-1) in Great Britain: a molecular epidemiological study. Transbound Emerg Dis (2014) 61:134–9. doi: 10.1111/tbed.12006
114. Morris KM, Mishra A, Raut AA, Gaunt ER, Borowska D, Kuo RI, et al. The molecular basis of differential host responses to avian influenza viruses in avian species with differing susceptibility. Front Cell Infect Microbiol (2023) 13:1067993. doi: 10.3389/fcimb.2023.1067993
115. Campbell LK, Fleming-Canepa X, Webster RG, Magor KE. Tissue specific transcriptome changes upon influenza A virus replication in the duck. Front Immunol (2021) 12:786205. doi: 10.3389/fimmu.2021.786205
116. Kuchipudi SV, Tellabati M, Sebastian S, Londt BZ, Jansen C, Vervelde L, et al. Highly pathogenic avian influenza virus infection in chickens but not ducks is associated with elevated host immune and pro-inflammatory responses. Vet Res (2014) 45:118. doi: 10.1186/s13567-014-0118-3
117. Evseev D, Magor KE. Innate immune responses to avian influenza viruses in ducks and chickens. Vet Sci (2019) 6. doi: 10.3390/vetsci6010005
118. Pantin-Jackwood MJ, Swayne DE. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev Sci Tech (2009) 28:113–36. doi: 10.20506/rst.28.1.1869
119. Kalthoff D, Globig A, Beer M. (Highly pathogenic) avian influenza as a zoonotic agent. Vet Microbiol (2010) 140:237–45. doi: 10.1016/j.vetmic.2009.08.022
120. D’Elía G, Fabre PH, Lessa EP. Rodent systematics in an age of discovery: Recent advances and prospects. J Mammalogy (2019) 100:852–71. doi: 10.1093/jmammal/gyy179
121. Gravinatti ML, Barbosa CM, Soares RM, Gregori F. Synanthropic rodents as virus reservoirs and transmitters. Rev Soc Bras Med Trop (2020) 53:e20190486. doi: 10.1590/0037-8682-0486-2019
122. Kumar N, Acharya A, Gendelman HE, Byrareddy SN. The 2022 outbreak and the pathobiology of the monkeypox virus. J Autoimmun (2022) 131:102855. doi: 10.1016/j.jaut.2022.102855
123. Garry RF. Lassa fever - the road ahead. Nat Rev Microbiol (2023) 21:87–96. doi: 10.1038/s41579-022-00789-8
124. Obando-Rico CJ, Valencia-Grajales YF, Bonilla-Aldana DK. Prevalence of orthohantavirus in rodents: A systematic review and meta-analysis. Travel Med Infect Dis (2023) 51:102504. doi: 10.1016/j.tmaid.2022.102504
125. Cohn O, Yankovitz G, Peshes-Yaloz N, Steuerman Y, Frishberg A, Brandes R, et al. Distinct gene programs underpinning disease tolerance and resistance in influenza virus infection. Cell Syst (2022) 13:1002–15.e1009. doi: 10.1016/j.cels.2022.11.004
126. Schlöder J, Shahneh F, Schneider FJ, Wieschendorf B. Boosting regulatory T cell function for the treatment of autoimmune diseases - That's only half the battle! Front Immunol (2022) 13:973813. doi: 10.3389/fimmu.2022.973813
127. Piconese S, Barnaba V. Stability of regulatory T cells undermined or endorsed by different type-1 cytokines. Adv Exp Med Biol (2015) 850:17–30. doi: 10.1007/978-3-319-15774-0_2
128. Wilk E, Pandey AK, Leist SR, Hatesuer B, Preusse M, Pommerenke C, et al. RNAseq expression analysis of resistant and susceptible mice after influenza A virus infection identifies novel genes associated with virus replication and important for host resistance to infection. BMC Genomics (2015) 16:655. doi: 10.1186/s12864-015-1867-8
129. Gocher-Demske AM, Cui J, Szymczak-Workman AL, Vignali KM, Latini JN, Pieklo GP, et al. IFNγ-induction of T. Nat Immunol (2023) 24(5):841–54. doi: 10.1038/s41590-023-01453-w
130. Rosshart SP, Herz J, Vassallo BG, Hunter A, Wall MK, Badger JH, et al. Laboratory mice born to wild mice have natural microbiota and model human immune responses. Science (2019) 365. doi: 10.1126/science.aaw4361
131. Hünig T. The rise and fall of the CD28 superagonist TGN1412 and its return as TAB08: a personal account. FEBS J (2016) 283:3325–34. doi: 10.1111/febs.13754
132. Musso D, Rodriguez-Morales AJ, Levi JE, Cao-Lormeau VM, Gubler DJ. Unexpected outbreaks of arbovirus infections: lessons learned from the Pacific and tropical America. Lancet Infect Dis (2018) 18:e355–61. doi: 10.1016/S1473-3099(18)30269-X
133. Xu Z, Feng Y, Chen X, Shi M, Fu S, Yang W, et al. Virome of bat-infesting arthropods: Highly divergent viruses in different vectors. J Virol (2022) 96:e0146421. doi: 10.1128/jvi.01464-21
134. Faizah AN, Kobayashi D, Matsumura R, Watanabe M, Higa Y, Sawabe K, et al. Blood meal source identification and RNA virome determination in Japanese encephalitis virus vectors collected in Ishikawa Prefecture, Japan, show distinct avian/mamMalian host preference. J Med Entomol (2023) 60(3):620–8. doi: 10.1093/jme/tjad028
135. Perveen N, Muhammad K, Muzaffar SB, Zaheer T, Munawar N, Gajic B, et al. Host-pathogen interaction in arthropod vectors: Lessons from viral infections. Front Immunol (2023) 14:1061899. doi: 10.3389/fimmu.2023.1061899
136. Maqbool M, Sajid MS, Saqib M, Anjum FR, Tayyab MH, Rizwan HM, et al. Potential mechanisms of transmission of tick-borne viruses at the virus-tick interface. Front Microbiol (2022) 13:846884. doi: 10.3389/fmicb.2022.846884
137. Hoffman T, Olsen B, Lundkvist Å. The biological and ecological features of northbound migratory birds, ticks, and tick-borne microorganisms in the African-Western Palearctic. Microorganisms (2023) 11. doi: 10.3390/microorganisms11010158
138. Labuda M, Nuttall PA. Tick-borne viruses. Parasitology (2004) 129 Suppl:S221–245. doi: 10.1017/S0031182004005220
139. Bartíková P, Holíková V, Kazimírová M, Štibrániová I. Tick-borne viruses. Acta Virol (2017) 61:413–27. doi: 10.4149/av_2017_403
140. de Almeida JP, Aguiar ER, Armache JN, Olmo RP, Marques JT. The virome of vector mosquitoes. Curr Opin Virol (2021) 49:7–12. doi: 10.1016/j.coviro.2021.04.002
141. Moonen JP, Schinkel M, van der Most T, Miesen P, van Rij RP. Composition and global distribution of the mosquito virome - A comprehensive database of insect-specific viruses. One Health (2023) 16:100490. doi: 10.1016/j.onehlt.2023.100490
142. Olmo RP, Todjro YMH, Aguiar ERGR, de Almeida JPP, Ferreira FV, Armache JN, et al. Mosquito vector competence for dengue is modulated by insect-specific viruses. Nat Microbiol (2023) 8:135–49. doi: 10.1038/s41564-022-01289-4
143. Ramírez AL, Colmant AMG, Warrilow D, Huang B, Pyke AT, McMahon JL, et al. Metagenomic analysis of the virome of mosquito excreta. mSphere (2020) 5. doi: 10.1128/mSphere.00587-20
144. Hubálek Z, Rudolf I, Nowotny N. Arboviruses pathogenic for domestic and wild animals. Adv Virus Res (2014) 89:201–75. doi: 10.1016/B978-0-12-800172-1.00005-7
145. García-Romero C, Carrillo Bilbao GA, Navarro JC, Martin-Solano S, Saegerman C. Arboviruses in mammals in the neotropics: A systematic review to strengthen epidemiological monitoring strategies and conservation medicine. Viruses (2023) 15. doi: 10.3390/v15020417
146. Vasconcelos PFC, Travassos da Rosa AP, Rodrigues SG, Travassos da Rosa ES, Degallier N, Travassos da Rosa JF. Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses. Cad. Saúde Pública (2001) 17:155–64. doi: 10.1590/S0102-311X2001000700025
147. Nunes JAL, Sousa JR, Smith VC, Quaresma JS, Vasconcelos PFDC, Chiang JO. Immunological impact of cytokines on the chikungunya virus pathophysiology: A literature narrative review. Rev Med Virol (2023) 33(4):e2441. doi: 10.1002/rmv.2441
148. Ribeiro YP, Falcão LFM, Smith VC, de Sousa JR, Pagliari C, Franco ECS, et al. Comparative analysis of human hepatic lesions in dengue, yellow fever, and chikungunya: Revisiting histopathological changes in the light of modern knowledge of cell pathology. Pathogens (2023) 12. doi: 10.3390/pathogens12050680
149. Weaver SC, Charlier C, Vasilakis N, Lecuit M. Zika, chikungunya, and other emerging vector-borne viral diseases. Annu Rev Med (2018) 69:395–408. doi: 10.1146/annurev-med-050715-105122
150. Van den Eynde C, Sohier C, Matthijs S, De Regge N. Japanese encephalitis virus interaction with mosquitoes: A review of vector competence, vector capacity and mosquito immunity. Pathogens (2022) 11. doi: 10.3390/pathogens11030317
151. Hadfield J, Brito AF, Swetnam DM, Vogels CBF, Tokarz RE, Andersen KG, et al. Twenty years of West Nile virus spread and evolution in the Americas visualized by Nextstrain. PLoS Pathog (2019) 15:e1008042. doi: 10.1371/journal.ppat.1008042
152. Fillâtre P, Revest M, Tattevin P. Crimean-Congo hemorrhagic fever: An update. Med Mal Infect (2019) 49:574–85. doi: 10.1016/j.medmal.2019.09.005
153. Bernard C, Holzmuller P, Bah MT, Bastien M, Combes B, Jori F, et al. Systematic review on crimean-congo hemorrhagic fever enzootic cycle and factors favoring virus transmission: Special focus on France, an apparently free-disease area in Europe. Front Vet Sci (2022) 9:932304. doi: 10.3389/fvets.2022.932304
154. Yoshii K. Epidemiology and pathological mechanisms of tick-borne encephalitis. J Vet Med Sci (2019) 81:343–7. doi: 10.1292/jvms.18-0373
155. Liebig K, Boelke M, Grund D, Schicht S, Bestehorn-Willmann M, Chitimia-Dobler L, et al. The stable matching problem in TBEV enzootic circulation: How important is the perfect tick-virus match? Microorganisms (2021) 9. doi: 10.3390/microorganisms9010196
156. Al-Tawfiq JA, Memish ZA. Alkhurma hemorrhagic fever virus. Microbes Infect (2017) 19:305–10. doi: 10.1016/j.micinf.2017.04.004
157. Bhatia B, Feldmann H, Marzi A. Kyasanur forest disease and alkhurma hemorrhagic fever virus-two neglected zoonotic pathogens. Microorganisms (2020) 8. doi: 10.3390/microorganisms8091406
158. Šimo L, Kazimirova M, Richardson J, Bonnet SI. The essential role of tick salivary glands and saliva in tick feeding and pathogen transmission. Front Cell Infect Microbiol (2017) 7:281. doi: 10.3389/fcimb.2017.00281
159. Nuttall PA. Tick saliva and its role in pathogen transmission. Wien Klin Wochenschr (2019) 135(7-8):165–76. doi: 10.1007/s00508-019-1500-y
160. Esteves E, Maruyama SR, Kawahara R, Fujita A, Martins LA, Righi AA, et al. Analysis of the salivary gland transcriptome of unfed and partially fed. Front Cell Infect Microbiol (2017) 7:476. doi: 10.3389/fcimb.2017.00476
161. Zinsstag J, Utzinger J, Probst-Hensch N, Shan L, Zhou XN. Towards integrated surveillance-response systems for the prevention of future pandemics. Infect Dis Poverty (2020) 9:140. doi: 10.1186/s40249-020-00757-5
162. Watsa M, Group WDSF. Rigorous wildlife disease surveillance. Science (2020) 369:145–7. doi: 10.1126/science.abc0017
163. Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature (2008) 451:990–3. doi: 10.1038/nature06536
164. Khetan AK. COVID-19: why declining biodiversity puts us at greater risk for emerging infectious diseases, and what we can do. J Gen Intern Med (2020) 35:2746–7. doi: 10.1007/s11606-020-05977-x
165. Sánchez A, Contreras A, Corrales JC, de la Fe C. [In the beginning it was zoonosis: One Health to combat this and future pandemics. SESPAS Report 2022]. Gac Sanit (2022) 36 Suppl 1:S61–7. doi: 10.1016/j.gaceta.2022.01.012
166. Wallau GL, Barbier E, Tomazatos A, Schmidt-Chanasit J, Bernard E. The virome of bats inhabiting Brazilian biomes: Knowledge gaps and biases towards zoonotic viruses. Microbiol Spectr (2023) 11:e0407722. doi: 10.1128/spectrum.04077-22
167. Raghwani J, Faust CL, François S, Nguyen D, Marsh K, Raulo A, et al. Seasonal dynamics of the wild rodent faecal virome. Mol Ecol (2022). doi: 10.1111/mec.16778
168. Eikenaar C, Hegemann A, Packmor F, Kleudgen I, Isaksson C. Not just fuel: energy stores are correlated with immune function and oxidative damage in a long-distance migrant. Curr Zool (2020) 66:21–8. doi: 10.1093/cz/zoz009
169. Schmaljohann H, Eikenaar C, Sapir N. Understanding the ecological and evolutionary function of stopover in migrating birds. Biol Rev Camb Philos Soc (2022) 97(4):1231–52. doi: 10.1111/brv.12839
170. Bergner LM, Orton RJ, Benavides JA, Becker DJ, Tello C, Biek R, et al. Demographic and environmental drivers of metagenomic viral diversity in vampire bats. Mol Ecol (2020) 29:26–39. doi: 10.1111/mec.15250
171. Brown S. The Remarkable Odyssey of a Semipalmated Sandpiper Shorebird Science. Canada: Manomet Soaring Solutions Grounded Science (2014).
172. Brister JR, Ako-Adjei D, Bao Y, Blinkova O. NCBI viral genomes resource. Nucleic Acids Res (2015) 43:D571–577. doi: 10.1093/nar/gku1207
173. Wang R, Chen J, Gao K, Wei GW. Vaccine-escape and fast-growing mutations in the United Kingdom, the United States, Singapore, Spain, India, and other COVID-19-devastated countries. Genomics (2021) 113:2158–70. doi: 10.1016/j.ygeno.2021.05.006
174. Tragni V, Preziusi F, Laera L, Onofrio A, Mercurio I, Todisco S, et al. Modeling SARS-CoV-2 spike/ACE2 protein-protein interactions for predicting the binding affinity of new spike variants for ACE2, and novel ACE2 structurally related human protein targets, for COVID-19 handling in the 3PM context. EPMA J (2022) 13:149–75. doi: 10.1007/s13167-021-00267-w
175. Kavaliers M, Choleris E, Agmo A, Pfaff DW. Olfactory-mediated parasite recognition and avoidance: linking genes to behavior. Horm Behav (2004) 46:272–83. doi: 10.1016/j.yhbeh.2004.03.005
176. Rivière S, Challet L, Fluegge D, Spehr M, Rodriguez I. Formyl peptide receptor-like proteins are a novel family of vomeronasal chemosensors. Nature (2009) 459:574–7. doi: 10.1038/nature08029
177. Curtis VA. Infection-avoidance behaviour in humans and other animals. Trends Immunol (2014) 35:457–64. doi: 10.1016/j.it.2014.08.006
178. Boillat M, Challet L, Rossier D, Kan C, Carleton A, Rodriguez I. The vomeronasal system mediates sick conspecific avoidance. Curr Biol (2015) 25:251–5. doi: 10.1016/j.cub.2014.11.061
179. Bufe B, Teuchert Y, Schmid A, Pyrski M, Pérez-Gómez A, Eisenbeis J, et al. Bacterial MgrB peptide activates chemoreceptor Fpr3 in mouse accessory olfactory system and drives avoidance behaviour. Nat Commun (2019) 10:4889. doi: 10.1038/s41467-019-12842-x
180. Stempel H, Jung M, Pérez-Gómez A, Leinders-Zufall T, Zufall F, Bufe B. Strain-specific loss of formyl peptide receptor 3 in the murine vomeronasal and immune systems. J Biol Chem (2016) 291:9762–75. doi: 10.1074/jbc.M116.714493
181. Sadler AJ, Williams BR. Interferon-inducible antiviral effectors. Nat Rev Immunol (2008) 8:559–68. doi: 10.1038/nri2314
182. Lang R, Li H, Luo X, Liu C, Zhang Y, Guo S, et al. Expression and mechanisms of interferon-stimulated genes in viral infection of the central nervous system (CNS) and neurological diseases. Front Immunol (2022) 13:1008072. doi: 10.3389/fimmu.2022.1008072
183. Björkström NK, Strunz B, Ljunggren HG. Natural killer cells in antiviral immunity. Nat Rev Immunol (2022) 22:112–23. doi: 10.1038/s41577-021-00558-3
184. Jansen JM, Gerlach T, Elbahesh H, Rimmelzwaan GF, Saletti G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J Clin Virol (2019) 119:44–52. doi: 10.1016/j.jcv.2019.08.009
185. Shah VK, Firmal P, Alam A, Ganguly D, Chattopadhyay S. Overview of immune response during SARS-CoV-2 infection: Lessons from the past. Front Immunol (2020) 11:1949. doi: 10.3389/fimmu.2020.01949
186. Baker ML, Schountz T, Wang LF. Antiviral immune responses of bats: a review. Zoonoses Public Health (2013) 60:104–16. doi: 10.1111/j.1863-2378.2012.01528.x
187. Mandl JN, Schneider C, Schneider DS, Baker ML. Going to bat(s) for studies of disease tolerance. Front Immunol (2018) 9:2112. doi: 10.3389/fimmu.2018.02112
188. Shaw TI, Srivastava A, Chou WC, Liu L, Hawkinson A, Glenn TC, et al. Transcriptome sequencing and annotation for the Jamaican fruit bat (Artibeus jamaicensis). PloS One (2012) 7:e48472. doi: 10.1371/journal.pone.0048472
189. Cowled C, Stewart CR, Likic VA, Friedländer MR, Tachedjian M, Jenkins KA, et al. Characterisation of novel microRNAs in the Black flying fox (Pteropus alecto) by deep sequencing. BMC Genomics (2014) 15:682. doi: 10.1186/1471-2164-15-682
190. Zhong C, Liu F, Hajnik RJ, Yao L, Chen K, Wang M, et al. Type I interferon promotes humoral immunity in viral vector vaccination. J Virol (2021) 95:e0092521. doi: 10.1128/JVI.00925-21
191. de Weerd NA, Nguyen T. The interferons and their receptors–distribution and regulation. Immunol Cell Biol (2012) 90:483–91. doi: 10.1038/icb.2012.9
192. Leeansyah E, Hey YY, Sia WR, Ng JHJ, Gulam MY, Boulouis C, et al. MR1-restricted T cells with MAIT-like characteristics are functionally conserved in the pteropid bat. iScience (2020) 23:101876. doi: 10.1016/j.isci.2020.101876
193. Sia WR, Zheng Y, Han F, Chen S, Ma S, Wang LF, et al. Exploring the role of innate lymphocytes in the immune system of bats and virus-host interactions. Viruses (2022) 14. doi: 10.3390/v14010150
194. Godfrey DI, Koay HF, McCluskey J, Gherardin NA. The biology and functional importance of MAIT cells. Nat Immunol (2019) 20:1110–28. doi: 10.1038/s41590-019-0444-8
195. Pavlovich SS, Lovett SP, Koroleva G, Guito JC, Arnold CE, Nagle ER, et al. The Egyptian rousette genome reveals unexpected features of bat antiviral immunity. Cell (2018) 173:1098–1110.e1018. doi: 10.1016/j.cell.2018.03.070
196. Nagaraja S, Jain D, Kesavardhana S. Inflammasome regulation in driving COVID-19 severity in humans and immune tolerance in bats. J Leukoc Biol (2022) 111:497–508. doi: 10.1002/JLB.4COVHR0221-093RR
197. Kacprzyk J, Hughes GM, Palsson-McDermott EM, Quinn SR, Puechmaille SJ, O'Neill LAJ, et al. A potent anti-inflammatory response in bat macrophages may be linked to extended longevity and viral tolerance. Acta Chiropterologica (2017) 19:219–28. doi: 10.3161/15081109ACC2017.19.2.001
198. Subudhi S, Rapin N, Misra V. Immune system modulation and viral persistence in bats: Understanding viral spillover. Viruses (2019) 11. doi: 10.3390/v11020192
199. Neil SJ, Zang T, Bieniasz PD. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature (2008) 451:425–30. doi: 10.1038/nature06553
200. Heusinger E, Kluge SF, Kirchhoff F, Sauter D. Early vertebrate evolution of the host restriction factor tetherin. J Virol (2015) 89:12154–65. doi: 10.1128/JVI.02149-15
201. Hayward JA, Tachedjian M, Johnson A, Irving AT, Gordon TB, Cui J, et al. Unique evolution of antiviral tetherin in bats. J Virol (2022) 96:e0115222. doi: 10.1128/jvi.01152-22
202. Cocka LJ, Bates P. Identification of alternatively translated Tetherin isoforms with differing antiviral and signaling activities. PloS Pathog (2012) 8:e1002931. doi: 10.1371/journal.ppat.1002931
203. Li SX, Barrett BS, Guo K, Santiago ML. Tetherin/BST-2: restriction factor or immunomodulator? Curr HIV Res (2016) 14:235–46. doi: 10.2174/1570162X14999160224102752
204. Nalbandian A, Sehgal K, Gupta A, Madhavan MV, McGroder C, Stevens JS, et al. Post-acute COVID-19 syndrome. Nat Med (2021) 27:601–15. doi: 10.1038/s41591-021-01283-z
205. Ramakrishnan RK, Kashour T, Hamid Q, Halwani R, Tleyjeh IM. Unraveling the mystery surrounding post-acute sequelae of COVID-19. Front Immunol (2021) 12:686029. doi: 10.3389/fimmu.2021.686029
206. Tay MZ, Poh CM, Rénia L, MacAry PA, Ng LFP. The trinity of COVID-19: immunity, inflammation and intervention. Nat Rev Immunol (2020) 20:363–74. doi: 10.1038/s41577-020-0311-8
207. Wu Z, McGoogan JM. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese center for disease control and prevention. JAMA (2020) 323:1239–42. doi: 10.1001/jama.2020.2648
208. Chowdhury MA, Hossain N, Kashem MA, Shahid MA, Alam A. Immune response in COVID-19: A review. J Infect Public Health (2020) 13:1619–29. doi: 10.1016/j.jiph.2020.07.001
209. Nazerian Y, Ghasemi M, Yassaghi Y, Nazerian A, Hashemi SM. Role of SARS-CoV-2-induced cytokine storm in multi-organ failure: Molecular pathways and potential therapeutic options. Int Immunopharmacol (2022) 113:109428. doi: 10.1016/j.intimp.2022.109428
210. Frank MG, Fleshner M, Maier SF. Exploring the immunogenic properties of SARS-CoV-2 structural proteins: PAMP:TLR signaling in the mediation of the neuroinflammatory and neurologic sequelae of COVID-19. Brain Behav Immun (2023) 111:259–69. doi: 10.1016/j.bbi.2023.04.009
211. Tang Y, Liu J, Zhang D, Xu Z, Ji J, Wen C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front Immunol (2020) 11:1708. doi: 10.3389/fimmu.2020.01708
212. Yu Y, Wang M, Zhang X, Li S, Lu Q, Zeng H, et al. Antibody-dependent cellular cytotoxicity response to SARS-CoV-2 in COVID-19 patients. Signal Transduct Target Ther (2021) 6:346. doi: 10.1038/s41392-021-00759-1
213. Mansourabadi AH, Aghamajidi A, Dorfaki M, Keshavarz F, Shafeghat Z, Moazzeni A, et al. B lymphocytes in COVID-19: a tale of harmony and discordance. Arch Virol (2023) 168:148. doi: 10.1007/s00705-023-05773-y
214. Upasani V, Rodenhuis-Zybert I, Cantaert T. Antibody-independent functions of B cells during viral infections. PLoS Pathog (2021) 17:e1009708. doi: 10.1371/journal.ppat.1009708
215. Quinti I, Lougaris V, Milito C, Cinetto F, Pecoraro A, Mezzaroma I, et al. A possible role for B cells in COVID-19? Lesson from patients with agammaglobulinemia. J Allergy Clin Immunol (2020) 146:211–13.e214. doi: 10.1016/j.jaci.2020.04.013
216. Peluso MJ, Deitchman AN, Torres L, Iyer NS, Munter SE, Nixon CC, et al. Long-term SARS-CoV-2-specific immune and inflammatory responses in individuals recovering from COVID-19 with and without post-acute symptoms. Cell Rep (2021) 36:109518. doi: 10.1016/j.celrep.2021.109518
217. Mehandru S, Merad M. Pathological sequelae of long-haul COVID. Nat Immunol (2022) 23:194–202. doi: 10.1038/s41590-021-01104-y
218. Chen B, Julg B, Mohandas S, Bradfute SB, Force RMPT. Viral persistence, reactivation, and mechanisms of long COVID. Elife (2023) 12. doi: 10.7554/eLife.86015
219. Sherif ZA, Gomez CR, Connors TJ, Henrich TJ, Reeves WB, Force RMPT. Pathogenic mechanisms of post-acute sequelae of SARS-CoV-2 infection (PASC). Elife (2023) 12. doi: 10.7554/eLife.86002
220. Schultheiß C, Willscher E, Paschold L, Gottschick C, Klee B, Henkes SS, et al. The IL-1β, IL-6, and TNF cytokine triad is associated with post-acute sequelae of COVID-19. Cell Rep Med (2022) 3:100663. doi: 10.1016/j.xcrm.2022.100663
221. Ahn M, Anderson DE, Zhang Q, Tan CW, Lim BL, Luko K, et al. Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host. Nat Microbiol (2019) 4:789–99. doi: 10.1038/s41564-019-0371-3
222. Broz P, Dixit VM. Inflammasomes: mechanism of assembly, regulation and signalling. Nat Rev Immunol (2016) 16:407–20. doi: 10.1038/nri.2016.58
223. Minton K. Negative regulation of inflammasome activation in bats. Nat Rev Immunol (2023) 1. doi: 10.1038/s41577-023-00892-8
224. Rodrigues TS, de Sá KSG, Ishimoto AY, Becerra A, Oliveira S, Almeida L, et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J Exp Med (2021) 218. doi: 10.1084/jem.20201707
225. Pan P, Shen M, Yu Z, Ge W, Chen K, Tian M, et al. SARS-CoV-2 N protein promotes NLRP3 inflammasome activation to induce hyperinflammation. Nat Commun (2021) 12:4664. doi: 10.1038/s41467-021-25015-6
226. Vrentas CE, Schaut RG, Boggiatto PM, Olsen SC, Sutterwala FS, Moayeri M. Inflammasomes in livestock and wildlife: Insights into the intersection of pathogens and natural host species. Vet Immunol Immunopathol (2018) 201:49–56. doi: 10.1016/j.vetimm.2018.05.008
227. Spano M, Di Matteo G, Ingallina C, Ambroselli D, Carradori S, Gallorini M, et al. Modulatory properties of food and nutraceutical components targeting NLRP3 inflammasome activation. Nutrients (2022) 14. doi: 10.3390/nu14030490
228. Ke Q, Greenawalt AN, Manukonda V, Ji X, Tisch RM. The regulation of self-tolerance and the role of inflammasome molecules. Front Immunol (2023) 14:1154552. doi: 10.3389/fimmu.2023.1154552
229. Hartmann G. Nucleic acid immunity. Adv Immunol (2017) 133:121–69. doi: 10.1016/bs.ai.2016.11.001
230. McNab F, Mayer-Barber K, Sher A, Wack A, O'Garra A. Type I interferons in infectious disease. Nat Rev Immunol (2015) 15:87–103. doi: 10.1038/nri3787
231. Rehwinkel J, Gack MU. RIG-I-like receptors: their regulation and roles in RNA sensing. Nat Rev Immunol (2020) 20:537–51. doi: 10.1038/s41577-020-0288-3
232. Onomoto K, Onoguchi K, Yoneyama M. Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors. Cell Mol Immunol (2021) 18:539–55. doi: 10.1038/s41423-020-00602-7
233. Myers KP, Olsen CW, Gray GC. Cases of swine influenza in humans: a review of the literature. Clin Infect Dis (2007) 44:1084–8. doi: 10.1086/512813
234. Shi J, Zeng X, Cui P, Yan C, Chen H. Alarming situation of emerging H5 and H7 avian influenza and effective control strategies. Emerg Microbes Infect (2023) 12:2155072. doi: 10.1080/22221751.2022.2155072
235. Olsen B, Munster VJ, Wallensten A, Waldenström J, Osterhaus AD, Fouchier RA. Global patterns of influenza a virus in wild birds. Science (2006) 312:384–8. doi: 10.1126/science.1122438
236. Abdelwhab EM, Mettenleiter TC. Zoonotic animal influenza virus and potential mixing vessel hosts. Viruses (2023) 15. doi: 10.3390/v15040980
237. Magor KE. Evolution of RNA sensing receptors in birds. Immunogenetics (2022) 74:149–65. doi: 10.1007/s00251-021-01238-1
238. Shriner SA, Root JJ. A review of avian influenza A virus associations in synanthropic birds. Viruses (2020) 12. doi: 10.3390/v12111209
239. Krchlíková V, Hron T, Těšický M, Li T, Ungrová L, Hejnar J, et al. Dynamic evolution of avian RNA virus sensors: Repeated loss of RIG-I and RIPLET. Viruses (2022) 15. doi: 10.3390/v15010003
240. Huo H, Wang Y, Wang D, Chen X, Zhao L, Chen H. Duck RIG-I restricts duck enteritis virus infection. Vet Microbiol (2019) 230:78–85. doi: 10.1016/j.vetmic.2019.01.014
241. Rehwinkel J, Tan CP, Goubau D, Schulz O, Pichlmair A, Bier K, et al. RIG-I detects viral genomic RNA during negative-strand RNA virus infection. Cell (2010) 140:397–408. doi: 10.1016/j.cell.2010.01.020
242. Yang Y, Ye F, Zhu N, Wang W, Deng Y, Zhao Z, et al. Middle East respiratory syndrome coronavirus ORF4b protein inhibits type I interferon production through both cytoplasmic and nuclear targets. Sci Rep (2015) 5:17554. doi: 10.1038/srep17554
243. Lee JY, Bae S, Myoung J. Middle East respiratory syndrome coronavirus-encoded ORF8b strongly antagonizes IFN-β promoter activation: its implication for vaccine design. J Microbiol (2019) 57:803–11. doi: 10.1007/s12275-019-9272-7
244. Goodbourn S, Randall RE. The regulation of type I interferon production by paramyxoviruses. J Interferon Cytokine Res (2009) 29:539–47. doi: 10.1089/jir.2009.0071
245. Liu G, Zhou Y. Cytoplasm and beyond: Dynamic innate immune sensing of influenza A virus by RIG-I. J Virol (2019) 93. doi: 10.1128/JVI.02299-18
246. Nazmi A, Dutta K, Basu A. RIG-I mediates innate immune response in mouse neurons following Japanese encephalitis virus infection. PloS One (2011) 6:e21761. doi: 10.1371/journal.pone.0021761
247. Jiang R, Ye J, Zhu B, Song Y, Chen H, Cao S. Roles of TLR3 and RIG-I in mediating the inflammatory response in mouse microglia following Japanese encephalitis virus infection. J Immunol Res (2014) 2014:787023. doi: 10.1155/2014/787023
248. Buskiewicz IA, Koenig A, Huber SA, Budd RC. Caspase-8 and FLIP regulate RIG-I/MDA5-induced innate immune host responses to picornaviruses. Future Virol (2012) 7:1221–36. doi: 10.2217/fvl.12.115
249. Kato H, Takeuchi O, Sato S, Yoneyama M, Yamamoto M, Matsui K, et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature (2006) 441:101–5. doi: 10.1038/nature04734
250. Siu KL, Yeung ML, Kok KH, Yuen KS, Kew C, Lui PY, et al. Middle east respiratory syndrome coronavirus 4a protein is a double-stranded RNA-binding protein that suppresses PACT-induced activation of RIG-I and MDA5 in the innate antiviral response. J Virol (2014) 88:4866–76. doi: 10.1128/JVI.03649-13
251. Sarkis S, Lise MC, Darcissac E, Dabo S, Falk M, Chaulet L, et al. Development of molecular and cellular tools to decipher the type I IFN pathway of the common vampire bat. Dev Comp Immunol (2018) 81:1–7. doi: 10.1016/j.dci.2017.10.023
252. Tarigan R, Shimoda H, Doysabas KCC, Ken M, Iida A, Hondo E. Role of pattern recognition receptors and interferon-beta in protecting bat cell lines from encephalomyocarditis virus and Japanese encephalitis virus infection. Biochem Biophys Res Commun (2020) 527:1–7. doi: 10.1016/j.bbrc.2020.04.060
253. Tarigan R, Katta T, Takemae H, Shimoda H, Maeda K, Iida A, et al. Distinct interferon response in bat and other mamMalian cell lines infected with Pteropine orthoreovirus. Virus Genes (2021) 57:510–20. doi: 10.1007/s11262-021-01865-6
254. Jones ME, Schuh AJ, Amman BR, Sealy TK, Zaki SR, Nichol ST, et al. Experimental inoculation of Egyptian Rousette bats (Rousettus aEgyptiacus) with viruses of the ebolavirus and marburgvirus genera. Viruses (2015) 7:3420–42. doi: 10.3390/v7072779
255. Paweska JT, Storm N, Grobbelaar AA, Markotter W, Kemp A, Jansen van Vuren P. Experimental inoculation of Egyptian fruit bats (Rousettus aEgyptiacus) with ebola virus. Viruses (2016) 8. doi: 10.3390/v8020029
256. Kuzmin IV, Ramanathan P, Basler CF, Bukreyev A. Effects of overexpression of the Egyptian fruit bat innate immune genes on filovirus infections in the host cells. Front Virol (2021) 1. doi: 10.3389/fviro.2021.759655
257. Schoggins JW. Interferon-stimulated genes: What do they all do? Annu Rev Virol (2019) 6:567–84. doi: 10.1146/annurev-virology-092818-015756
258. Kang S, Brown HM, Hwang S. Direct antiviral mechanisms of interferon-gamma. Immune Netw (2018) 18:e33. doi: 10.4110/in.2018.18.e33
259. Schoggins JW. Recent advances in antiviral interferon-stimulated gene biology. F1000Res (2018) 7:309. doi: 10.12688/f1000research.12450.1
260. Wells AI, Coyne CB. Type III interferons in antiviral defenses at barrier surfaces. Trends Immunol (2018) 39:848–58. doi: 10.1016/j.it.2018.08.008
261. Mesev EV, LeDesma RA, Ploss A. Decoding type I and III interferon signalling during viral infection. Nat Microbiol (2019) 4:914–24. doi: 10.1038/s41564-019-0421-x
262. King AMQ, Lefkowitz EJ, Mushegian AR, Adams MJ, Dutilh BE, Gorbalenya AE, et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch Virol (2018) 163:2601–31. doi: 10.1007/s00705-018-3847-1
263. Cook JK, Jackwood M, Jones RC. The long view: 40 years of infectious bronchitis research. Avian Pathol (2012) 41:239–50. doi: 10.1080/03079457.2012.680432
264. Huang M, Liu Y, Xia Y, Wang J, Zheng X, Cao Y. Infectious bronchitis virus nucleocapsid protein suppressed type I interferon production by interfering with the binding of MDA5-dsRNA and interacting with LGP2. Vet Microbiol (2023) 284:109798. doi: 10.1016/j.vetmic.2023.109798
265. Shaw AE, Hughes J, Gu Q, Behdenna A, Singer JB, Dennis T, et al. Fundamental properties of the mamMalian innate immune system revealed by multispecies comparison of type I interferon responses. PloS Biol (2017) 15:e2004086. doi: 10.1371/journal.pbio.2004086
266. Gorbunova V, Seluanov A, Kennedy BK. The world goes bats: Living longer and tolerating viruses. Cell Metab (2020) 32:31–43. doi: 10.1016/j.cmet.2020.06.013
267. Rodon J, Te N, Segalés J, Vergara-Alert J, Bensaid A. Enhanced antiviral immunity and dampened inflammation in llama lymph nodes upon MERS-CoV sensing: bridging innate and adaptive cellular immune responses in camelid reservoirs. Front Immunol (2023) 14:1205080. doi: 10.3389/fimmu.2023.1205080
268. Hayman DTS. Bat tolerance to viral infections. Nat Microbiol (2019) 4:728–9. doi: 10.1038/s41564-019-0430-9
269. Liu B, Chen X, Zhou L, Li J, Wang D, Yang W, et al. The gut microbiota of bats confers tolerance to influenza virus (H1N1) infection in mice. Transbound Emerg Dis (2022) 69:e1469–87. doi: 10.1111/tbed.14478
270. Krebs DL, Hilton DJ. SOCS proteins: negative regulators of cytokine signaling. Stem Cells (2001) 19:378–87. doi: 10.1634/stemcells.19-5-378
271. Morris R, Kershaw NJ, Babon JJ. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci (2018) 27:1984–2009. doi: 10.1002/pro.3519
272. Faraji J, Karimi M, Lawrence C, Mohajerani MH, Metz GAS. Non-diagnostic symptoms in a mouse model of autism in relation to neuroanatomy: the BTBR strain reinvestigated. Transl Psychiatry (2018) 8:234. doi: 10.1038/s41398-018-0280-x
273. Liau NPD, Laktyushin A, Lucet IS, Murphy JM, Yao S, Whitlock E, et al. The molecular basis of JAK/STAT inhibition by SOCS1. Nat Commun (2018) 9:1558. doi: 10.1038/s41467-018-04013-1
274. Koyama S, Ishii KJ, Coban C, Akira S. Innate immune response to viral infection. Cytokine (2008) 43:336–41. doi: 10.1016/j.cyto.2008.07.009
275. Huang S, Liu K, Cheng A, Wang M, Cui M, Huang J, et al. SOCS proteins participate in the regulation of innate immune response caused by viruses. Front Immunol (2020) 11:558341. doi: 10.3389/fimmu.2020.558341
276. Baetz A, Zimmermann S, Dalpke AH. Microbial immune evasion employing suppressor of cytokine signaling (SOCS) proteins. Inflammation Allergy Drug Targets (2007) 6:160–7. doi: 10.2174/187152807781696446
277. Croker BA, Kiu H, Nicholson SE. SOCS regulation of the JAK/STAT signalling pathway. Semin Cell Dev Biol (2008) 19:414–22. doi: 10.1016/j.semcdb.2008.07.010
278. Akhtar LN, Benveniste EN. Viral exploitation of host SOCS protein functions. J Virol (2011) 85:1912–21. doi: 10.1128/JVI.01857-10
279. Low ZY, Wen Yip AJ, Chow VTK, Lal SK. The Suppressor of Cytokine Signalling family of proteins and their potential impact on COVID-19 disease progression. Rev Med Virol (2022) 32:e2300. doi: 10.1002/rmv.2300
280. Alston CI, Dix RD. SOCS and herpesviruses, with emphasis on cytomegalovirus retinitis. Front Immunol (2019) 10:732. doi: 10.3389/fimmu.2019.00732
281. Du LY, Cui YL, Chen EQ, Cheng X, Liu L, Tang H. Correlation between the suppressor of cytokine signaling-1 and 3 and hepatitis B virus: possible roles in the resistance to interferon treatment. Virol J (2014) 11:51. doi: 10.1186/1743-422X-11-51
282. Miyoshi H, Fujie H, Shintani Y, Tsutsumi T, Shinzawa S, Makuuchi M, et al. Hepatitis C virus core protein exerts an inhibitory effect on suppressor of cytokine signaling (SOCS)-1 gene expression. J Hepatol (2005) 43:757–63. doi: 10.1016/j.jhep.2005.03.028
283. Seong RK, Lee JK, Shin OS. Zika virus-induction of the suppressor of cytokine signaling 1/3 contributes to the modulation of viral replication. Pathogens (2020) 9. doi: 10.3390/pathogens9030163
284. Xu X, Zheng J, Zheng K, Hou Y, Zhao F, Zhao D. Respiratory syncytial virus NS1 protein degrades STAT2 by inducing SOCS1 expression. Intervirology (2014) 57:65–73. doi: 10.1159/000357327
285. Pauli EK, Schmolke M, Wolff T, Viemann D, Roth J, Bode JG, et al. Influenza A virus inhibits type I IFN signaling via NF-kappaB-dependent induction of SOCS-3 expression. PloS Pathog (2008) 4:e1000196. doi: 10.1371/journal.ppat.1000196
286. Fenner JE, Starr R, Cornish AL, Zhang JG, Metcalf D, Schreiber RD, et al. Suppressor of cytokine signaling 1 regulates the immune response to infection by a unique inhibition of type I interferon activity. Nat Immunol (2006) 7:33–9. doi: 10.1038/ni1287
287. Luo X, Chen XX, Qiao S, Li R, Lu Q, Geng R, et al. Porcine reproductive and respiratory syndrome virus increases SOCS3 production via activation of p38/AP-1 signaling pathway to promote viral replication. Vet Microbiol (2021) 257:109075. doi: 10.1016/j.vetmic.2021.109075
288. Hartshorn KL. Innate immunity and influenza A virus pathogenesis: Lessons for COVID-19. Front Cell Infect Microbiol (2020) 10:563850. doi: 10.3389/fcimb.2020.563850
289. Liu PP, Blet A, Smyth D, Li H. The science underlying COVID-19: Implications for the cardiovascular system. Circulation (2020) 142:68–78. doi: 10.1161/CIRCULATIONAHA.120.047549
290. Zheng M, Kanneganti TD. The regulation of the ZBP1-NLRP3 inflammasome and its implications in pyroptosis, apoptosis, and necroptosis (PANoptosis). Immunol Rev (2020) 297:26–38. doi: 10.1111/imr.12909
291. Place DE, Lee S, Kanneganti TD. PANoptosis in microbial infection. Curr Opin Microbiol (2021) 59:42–9. doi: 10.1016/j.mib.2020.07.012
Keywords: virome, disease tolerance, resistance, inflammatory response, sickness behavior
Citation: Pereira PDC, Diniz DG, da Costa ER, Magalhães NGdM, da Silva AdJF, Leite JGS, Almeida NIP, Cunha KdN, de Melo MAD, Vasconcelos PFdC, Diniz JAP, Brites D, Anthony DC, Diniz CWP and Guerreiro-Diniz C (2023) Genes, inflammatory response, tolerance, and resistance to virus infections in migratory birds, bats, and rodents. Front. Immunol. 14:1239572. doi: 10.3389/fimmu.2023.1239572
Received: 13 June 2023; Accepted: 14 August 2023;
Published: 29 August 2023.
Edited by:
Alexander Bukreyev, University of Texas Medical Branch at Galveston, United StatesReviewed by:
Abhishek N. Prasad, University of Texas Medical Branch at Galveston, United StatesCopyright © 2023 Pereira, Diniz, da Costa, Magalhães, da Silva, Leite, Almeida, Cunha, de Melo, Vasconcelos, Diniz, Brites, Anthony, Diniz and Guerreiro-Diniz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristovam Wanderley Picanço Diniz, Y3dwZGluaXpAZ21haWwuY29t
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.