
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 21 July 2023
Sec. Parasite Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1193256
This article is part of the Research Topic Immune tolerance and human malaria View all 8 articles
 Suelen Queiroz Diniz1,2Maria Marta Figueiredo1
Suelen Queiroz Diniz1,2Maria Marta Figueiredo1 Pedro Augusto Carvalho Costa1
Pedro Augusto Carvalho Costa1 Olindo Assis Martins-Filho3Andrea Teixeira-Carvalho3
Olindo Assis Martins-Filho3Andrea Teixeira-Carvalho3 Dhélio Batista Pereira4Mauro Shugiro Tada4
Dhélio Batista Pereira4Mauro Shugiro Tada4 Luis Carlos Crocco Afonso5Markus Kohlhoff6
Luis Carlos Crocco Afonso5Markus Kohlhoff6 Carlos Leomar Zani6Ricardo Tostes Gazzinelli1,2
Carlos Leomar Zani6Ricardo Tostes Gazzinelli1,2 Fabiano Oliveira7
Fabiano Oliveira7 Lis Ribeiro Antonelli1,2*
Lis Ribeiro Antonelli1,2*Background: Plasmodium spp. infection triggers the production of inflammatory cytokines that are essential for parasite control, and conversely responsible for symptoms of malaria. Monocytes play a role in host defense against Plasmodium vivax infection and represent the main source of inflammatory cytokines and reactive oxygen species. The anti-inflammatory cytokine IL-10 is a key regulator preventing exacerbated inflammatory responses. Studies suggested that different clinical presentations of malaria are strongly associated with an imbalance in the production of inflammatory and anti-inflammatory cytokines.
Methods: A convenience sampling of peripheral blood mononuclear cells from Plasmodium vivax-infected patients and healthy donors were tested for the characterization of cytokine and adenosine production and the expression of ectonucleotidases and purinergic receptors.
Results: Here we show that despite a strong inflammatory response, monocytes also bear a modulatory role during malaria. High levels of IL-10 are produced during P. vivax infection and its production can be triggered in monocytes by P. vivax-infected reticulocytes. Monocytes express high levels of ectonucleotidases, indicating their important role in extracellular ATP modulation and consequently in adenosine production. Plasmatic levels of adenosine are not altered in patients experiencing acute malaria; however, their monocyte subsets displayed an increased expression of P1 purinergic receptors. In addition, adenosine decreases Tumor Necrosis Factor production by monocytes, which was partially abolished with the blockage of the A2a receptor.
Conclusion: Monocytes have a dual role, attempting to control both the P. vivax infection and the inflammatory response. Purinergic receptor modulators emerge as an untapped approach to ameliorate clinical malaria.
Malaria is an infectious disease caused by protozoan parasites of the genus Plasmodium. Despite recent successes of several control actions implemented in endemic regions, malaria remains one of the greatest public health problems worldwide. Plasmodium vivax is the most widely distributed causative agent of human malaria, and is responsible for most cases in Latin America and Asia (1). Controlling the parasite burden without causing major host pathology benefits from a refined balance between inflammatory and regulatory immune responses, which failure is strongly associated with different clinical manifestations of the disease (2). Early production of pro-inflammatory cytokines, such as Tumor Necrosis Factor (TNF), Interleukin(IL)-6 and IL-1β, supports parasite clearance (3, 4) and, concomitantly, regulatory cytokines, such as IL-10, are also produced (2). IL-10 is a multifunctional cytokine with anti-inflammatory effect on most hematopoietic cell types, including monocytes and macrophages (5). A pre-clinical study reported that African children with severe anemia caused by P. falciparum display lower IL-10 levels than patients with moderate anemia, suggesting that IL-10 plays an important role in preventing severe cases (6). Moreover, the literature shows evidence that IL-10 may be protective by inhibiting TNF activity (7). Despite being considered a highly inflammatory disease, P. vivax infection triggers substantial IL-10 production (8) and although the immune response mediated by IL-10 during malaria is well established, the main sources, and what triggers its production are not fully understood.
In stress situations, such as cellular injury and infections, Adenosine Triphosphate (ATP) is released to the extracellular environment and act as a danger signal (9), binding to P2 purinergic receptors (10), stimulating robust inflammation mainly through the activation of monocytes, macrophages and dendritic cells (11). Therefore, extracellular ATP concentration is tightly regulated by the ectonucleotidases CD39 and CD73, which in cooperation hydrolyze ATP to adenosine (12). Adenosine binding to P1 purinergic receptors, A2a and A2b (13), triggers inhibition of pro-inflammatory cytokines (14) and stimulates the production of IL-10 (15). Considering these findings, we evaluated whether adenosine contributes to the regulation of P. vivax infection inducing inflammation.
We demonstrate that monocytes express high levels of ectonucleotidases, indicating their role in extracellular ATP modulation and, consequently, in adenosine production. Despite not detecting alterations on the plasmatic levels of adenosine in patients experiencing acute malaria caused by P. vivax, their monocytes displayed an increased expression of P1 purinergic receptors. In addition, adenosine decreases TNF production by monocytes, which is partially abolished with the blockage of the adenosine A2a receptor.
A total of 48 patients with uncomplicated malaria caused by P. vivax infection were enrolled in this study at Centro de Pesquisa de Medicina Tropical de Rondônia (CEPEM) in Porto Velho, Rondônia, an endemic area for malaria in the Amazon region of Brazil. The group consisted of eight females (16.66%) and 40 males (83.66%) age range from 18 to 69 years (35 ± 11.13) (Table S1). Up to 100 mL of peripheral blood was collected after confirmation of P. vivax infection by thick blood smear film. Patients were treated according to the Brazilian Ministry of Health guideline. The clinical manifestations of acute malaria were fever, chills, nausea, vomit, diarrhea, myalgia, headache, and arthralgia. Peripheral blood was also collected from 11 healthy donors (HD) living in the same endemic area, and 20 individuals from Belo Horizonte/MG, nonendemic area for malaria. All the HD were tested for P. vivax infection and were negative.
These studies were performed under protocols reviewed and approved by the Ethical Committees on Human Experimentation from Centro de Pesquisa em Medicina Tropical de Rondônia (CEP-CEPEM 095/2009) and Instituto René Rachou, Fundação Oswaldo Cruz (CEP-IRR 2004), the National Ethical Committee (CONEP 15652) from Ministry of Health, Brazil. Only adults, 18 years and older, were enrolled in the study. All patients enrolled in this study provided written informed consent.
Peripheral blood mononuclear cells (PBMC) were prepared from heparinized venous blood of adult volunteers by Ficoll-Hypaque density gradient centrifugation (GE Healthcare Life Sciences). The blood samples were diluted in sterile 0.9% saline (vol/vol). Thirty-five microliters of the diluted blood was added gently into a tube containing 15 mL of Ficoll-Hypaque and centrifuged. PBMC were collected, transferred to a 50 mL conical tube, and washed three times. The cell concentration was determined using a Neubauer chamber and adjusted according to the test to be performed.
The red blood cell pellet from the Ficoll-Hypaque density gradient centrifugation was harvested and washed three times in Phosphate Buffered Saline (PBS) and then resuspended in RPMI 1640 medium (Sigma Aldrich) to a final hematocrit of 10%. Five milliliters of this suspension was overlaid on 5 mL of a 45% Percoll (Sigma Aldrich) solution in a 15 mL tube. After centrifugation, floating mature P. vivax-infected reticulocytes (Pv-RET) were collected, washed three times, and then labeled with FITC (200 µg/mL) (Sigma Aldrich). In brief, 106 Pv-RET were resuspended in PBS and incubated for 30 minutes in the presence of Fluorescein-5-isothiocyanate (FITC) at room temperature, protected from light, and under constant agitation. Cells were washed twice and resuspended according to protocols to be used.
The plasmatic adenosine levels were measured by targeted LC-MS analysis using a Nexera ultra-high performance liquid chromatography (UHPLC) system (Shimadzu, Japan) hyphenated to a maXis ETD high resolution ESI-qTOF mass spectrometer (Bruker, Germany) and controlled by the Bruker Compass v1.5 software Package. The peripheral blood (10 mL) was collected in a tube with heparin (12.4 U/mL) and a “stop solution” containing dipyridamole (75 μM), EHNA (15 μM), and ethylenediaminetetraacetic acid (3mM) were immediately added to the blood to prevent degradation/consumption of adenosine. After centrifugation (800 x g/10 min), the plasma was collected, and protein precipitation was performed using acetonitrile. Samples were centrifugated, and the supernatants were collected and stored at -80°C until analysis. A phenacetin solution (50 ng/mL) was prepared in acetonitrile (precipitation solution) and used as an internal standard to evaluate the efficiency of adenosine extraction. The separation of the compounds present in the supernatant was done using a Shimadzu Nexera UPLC coupled to a Bruker MaXis qTOF mass spectrometer. The column Shimadzu Shim-Pack XR-ODS III (C18, 2.2 μm, 80 A, and 2.0 x 200 mm) was used, and the amount of sample injected into the chromatograph was 10 μL. A binary gradient of mobile phase A (water) and B (acetonitrile) was programmed at a flow rate of 200 μL/min, both with 0.1% formic acid. In intervals of 0–0.5 min, 5% eluent B; 0.5–13 min, 5–100% B; 13–14 min, 100% B; 14–15 min, 100–5% B; and 15–20 min, 5% B were used. Throughout the process, the temperature was maintained at 40°C. Mass spectrometry was performed using the positive MS method with 0.5 Hz frequency and with a range of 40–400 m/z. A high precision calibration (HPC) was performed at the beginning of each run by injecting 20 μL of a 10 mM sodium formate/acetate solution.
CD14+CD16−, CD14+CD16+, and CD14lowCD16+ monocytes from P. vivax-infected patients (Pv) were sorted with a FACSAria II cell sorter (BD Biosciences), using the following antibodies: anti-CD14 (clone 61D3)-BV450, anti-CD16 (clone 3G8)-PECy7, and anti-CD66b (clone G10F5)-FITC. Anti-CD66b was used to exclude contaminations with neutrophils.
The analysis of the expression of messenger RNA (mRNA) was performed using the method of nCounter NanoString. A total of 104 cells of each monocyte subpopulation were lysed in RLT buffer (QIAGEN) supplemented with β-mercaptoethanol and then frozen at -80°C. These lysates were hybridized with specific capture and reporter probes for 16 hours and loaded onto the nCounter prep station. The mRNA quantification was performed in the nCounter Digital Analyzer using 600 fields to detect the hybridized probes. Data were normalized in two ways described previously. Briefly, the first normalization was for small variations utilizing the internal positive controls present in each CodeSet. Then the samples were normalized with seven housekeeping genes that were included in the CodeSet. The data were analyzed with nSolver software.
Phenotypic characterization of peripheral blood leukocytes was performed using conventional and image flow cytometry. We evaluated the expression of the molecules of interest, CD39 (clone eBioA1 (A1))-PE CD73 (clone AD2)-PerCP-Cy5.5 in monocyte subsets, CD4+ and CD8+ cells, B cells and neutrophis. For these analyses, PBMC were incubated with surface antibodies (anti-CD14, anti-CD16, and anti-CD66b, anti-CD19 (clone HIB19)-FITC, anti-CD3 (clone OKT3)-FITC, anti-CD4 (clone SK3)-PE, anti-CD8 (clone RPA-T8)-eFluor780) and evaluated by conventional (BD FACSCelesta) and image flow cytometry (ImageStreamX Mark II, Merck-Millipore).
After purification of monocyte subsets by cell sorter, they were plated in Hanks’ Balanced Salt Solution (HBSS) medium and cultured for 3 hours in the absence (Pv-RET-) or in the presence (Pv-RET+) of P. vivax-infected reticulocytes labeled with FITC in a 1:1 ratio. After culture, the cells were washed and stained with surface antibodies (anti-CD39-PE and anti-CD73-PerCP-Cy5.5) for 30 minutes. After this period, the cells were rewashed, resuspended in 50 µL of staining buffer (PBS 2% fetal calf serum), and analyzed on the image flow cytometer (ImageStreamX Mark II, Merck-Millipore), which, in addition to generating quality images, allows statistical analysis. The analysis was performed using the IDEAS v6.1 software.
Plasma levels of IL-10 were measured using the Th1/Th2/Th17 Cytometric Bead Array kit in plasma from Pv during acute disease following the manufacturer’s specifications. Intracellular IL-10 and TNF production by monocytes was assessed by conventional flow cytometry. Monocytes were stimulated with Pv-RET (1:1 ratio) or LPS (100 ng/mL) in the presence of brefeldin A (0.2 µL/200 µL) for 3 hours. After culture, cells were fixed, and permeabilized, and then incubated with antibodies anti-IL-10-PE and anti-TNF-FITC. The procedure was performed according to the manufacturer’s description (BD Cytofix/Cytoperm™).
To assess the ability of monocytes from Pv to modulate the immune response when stimulated by adenosine (10 nM), they were stimulated with lipopolysaccharides (LPS) (100 ng/mL) in the absence and the presence of adenosine. Subsequently, the intracellular levels of IL-10 and TNF were evaluated as described above. In specific experiments, ZM241385 (4-(2-(7-amino-2-(2-furyl)(1,2,4)triazole(2,3-α)(1,3,5) triazine-5-ylamino)ethyl)phenol) and MRS1754 (N-(4-cyanophenyl)-2(4-(2,3,6,7tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purine-8yl)phenoxy)acetamide) (Tocris Bioscience), the ADORA2a and A2b antagonists respectively, were used. The cells were cultured for 30 minutes with the adenosine receptors antagonists (5 µM) before LPS (100 ng/mL) and adenosine (10 nM) were added.
The statistical analysis was performed, first considering the variables of independence, normality, and variance to define the data as parametric or nonparametric. Nonparametric data were evaluated using the Mann-Whitney test for unpaired samples or Wilcoxon matched-pair for paired samples. Parametric data were assessed by analysis of the unpaired or paired t-test. In all cases, the differences were considered statistically significant at p < 0.05. The GraphPad Prism 8.0 software was used for data analysis.
IL-10 levels were measured in the plasma from acute P. vivax-infected patients (Pv) as well as from HD (Figure 1A). Previous studies showed increased levels of circulating IL-10 during the acute phase of P. vivax infection (16). Likewise, we observe that Pv displayed statistically significant higher levels of IL-10 than HD (Figure 1A, left panel). IL-10 production was also induced by co-culturing monocytes with Pv-infected reticulocytes (Pv-RET) (Figure 1A, right panel).
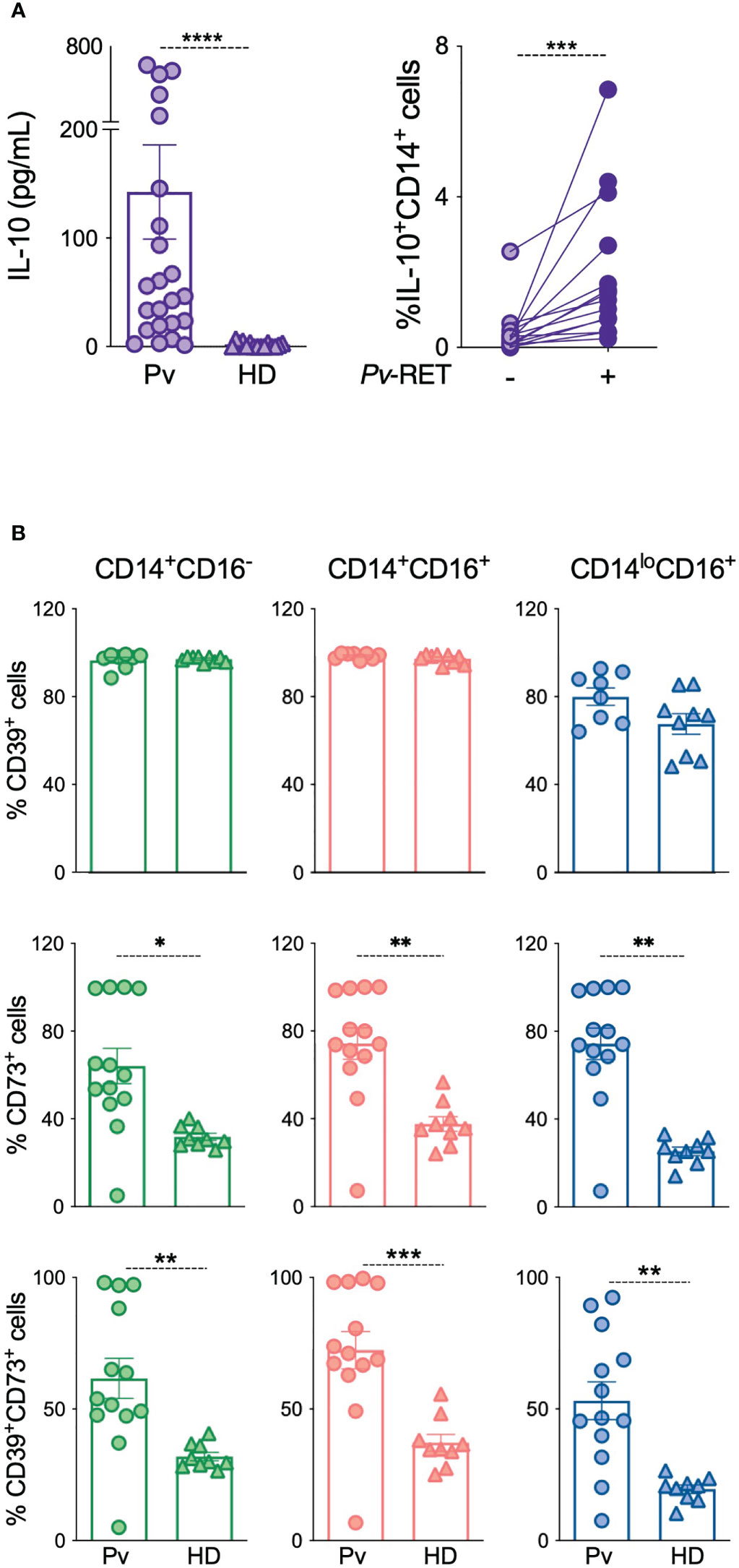
Figure 1 IL-10 and ectonucleotidases are upregulated in patients with acute P. vivax infection. (A) Plasma levels of IL-10 were measured using Th1/Th2/Th17 CBA kit in patients during acute malaria (Pv) (n = 23) and healthy donors (HD) (n = 13) (left panel). Intracellular IL-10 production by monocytes with (Pv-RET) and without (-) P. vivax-infected reticulocytes was assessed by flow cytometry (right panel) (n = 14) (right panel). (B) CD39, CD73 and their co-expression were determined in monocytes from Pv (n = 13) and HD (n = 9) using conventional flow cytometry in classical (green, left panel), inflammatory (red, middle panel) and patrolling monocytes (blue, right panel). Scatter plots with bars representing mean ± SEM. *p ≤ 0.05 **p ≤ 0.01 *** or ****p ≤ 0.001.
Since IL-10 production can be triggered by adenosine, we investigated the expression of the ectonucleotidases CD39 and CD73, molecules responsible for extracellular adenosine production on circulating leukocytes from our cohort using flow cytometry (Figure 1B and Figure S1). No changes were detected on the CD39 expression in monocyte subsets from Pv compared to HD (Figure 1B, top panel), but its expression was increased on T cells during acute malaria (Figure S1A). Conversely, CD73 expression was higher on monocyte subsets from Pv compared to HD (Figure 1B, middle panel, but no changes were observed on T, B or neutrophils (Figure S1B). CD39 and CD73 are sequentially involved in the hydrolysis of extracellular ATP to adenosine, and their co-expression increases its efficiency. Therefore, we analyzed the co-expression of the ectonucleotidases, which followed the CD73 expression pattern, being induced during acute malaria (Figure 1B, bottom panel).
Image flow cytometry was used to define the levels of ectonucleotidases expression and their location on monocyte subsets. As shown by conventional flow cytometry, high frequencies of ectonucleotidases expressing cells were found among all monocyte subsets and were higher among inflammatory monocytes (Figures 2A, B, top and middle panels). Higher frequencies of CD39+CD73+ cells are also found in inflammatory monocytes from Pv than their other counterparts (Figures 2A, B, bottom panel, and Figure S2A).
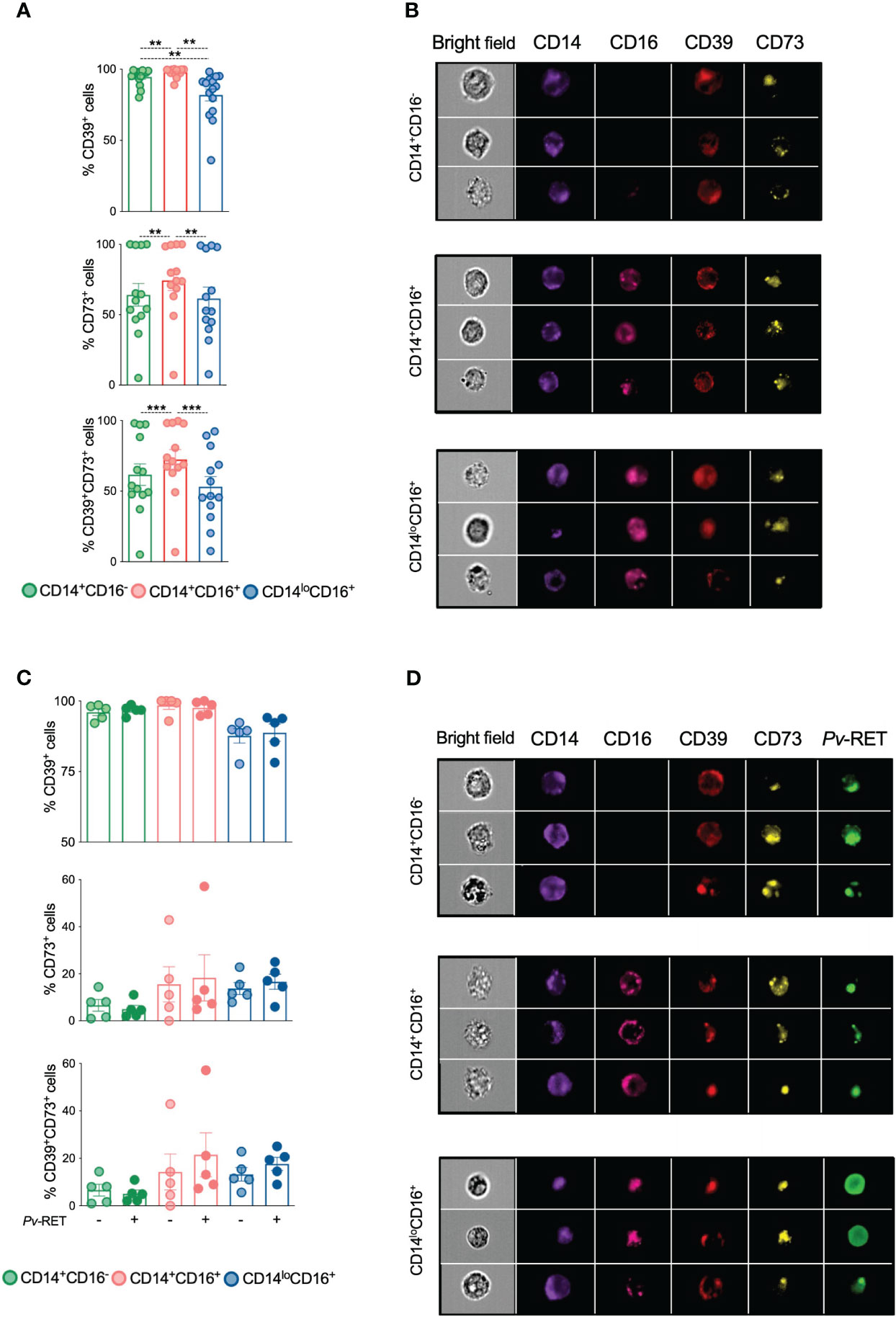
Figure 2 Ectonucleotidases CD39 and CD73 colocalize within monocyte subsets. CD39, CD73 and their co-expression were determined by imaging flow cytometry on monocyte subsets ex vivo (n = 13) (A) and after culture with (+) or without (-) P. vivax-infected reticulocytes (C) (n = 5). **p ≤ 0.01 ***p ≤ 0.001. Scatter plots with bars representing mean ± SEM. Green, red and blue symbols represent classical (CD14+CD16-), inflammatory (CD14+CD16+) and patrolling (CD14loCD16+) monocytes (A, C), respectively. Representative images of the expression of CD39 and CD73 (B) and P. vivax-infected reticulocytes and CD39 and CD73 (D) on monocyte subsets.
We next asked if adding P. vivax-infected reticulocytes would alter the expression of ectonucleotidases on monocyte subpopulations in vitro. We observed that all three monocyte subsets were able to phagocytose Pv-infected reticulocytes (Figures S2B, C), which did not impact the expression of CD39 and CD73 (Figures 2C, D).
Extracellular ATP and adenosine act mainly through P2 and P1 purinergic receptor (10, 13), respectively. To determine whether P. vivax infection alter the expression of purinergic receptors, we analyzed the number of mRNA copies of selected genes in monocyte subsets from Pv and HD using nCounter NanoString. Fourteen genes were assessed: Adenosine deaminase (ADA) (purple), ADORA1 (pink), ADORA2a (yellow), ADORA2b (orange), ADORA3 (dark pink), IL-10 receptor (CD210) (blue), P2X1 (green), P2X3 (dark orange), P2X4 (dark green), P2X7 (red), P2Y1 (dark blue), P2Y11 (black), P2Y2 (brown), and P2Y4 (gray) (Figure 3A) (Table S2). The data obtained were organized in radar graphs, whose differences in shapes indicate that P. vivax infection changes the expression of purinergic receptors (Figure 3A). The expression of ADORA2a and ADORA2b were increased respectively in inflammatory and patrolling monocytes and classical and inflammatory monocytes from Pv compared with HD (Figure 3B). P. vivax infection also increased the expression of ADA in patrolling monocytes. Higher levels of ADORA2b are found in classical than in patrolling monocytes and higher levels of ADA are found in the patrolling subset followed by their inflammatory and classical counterparts. The significant changes observed in the expression of ectonucleotidases and the adenosine receptors ADORA2a and ADORA2b in monocyte subsets, suggest a role for adenosine in malaria. Fewer significant differences were found in P2 purinergic receptors (Table S2).
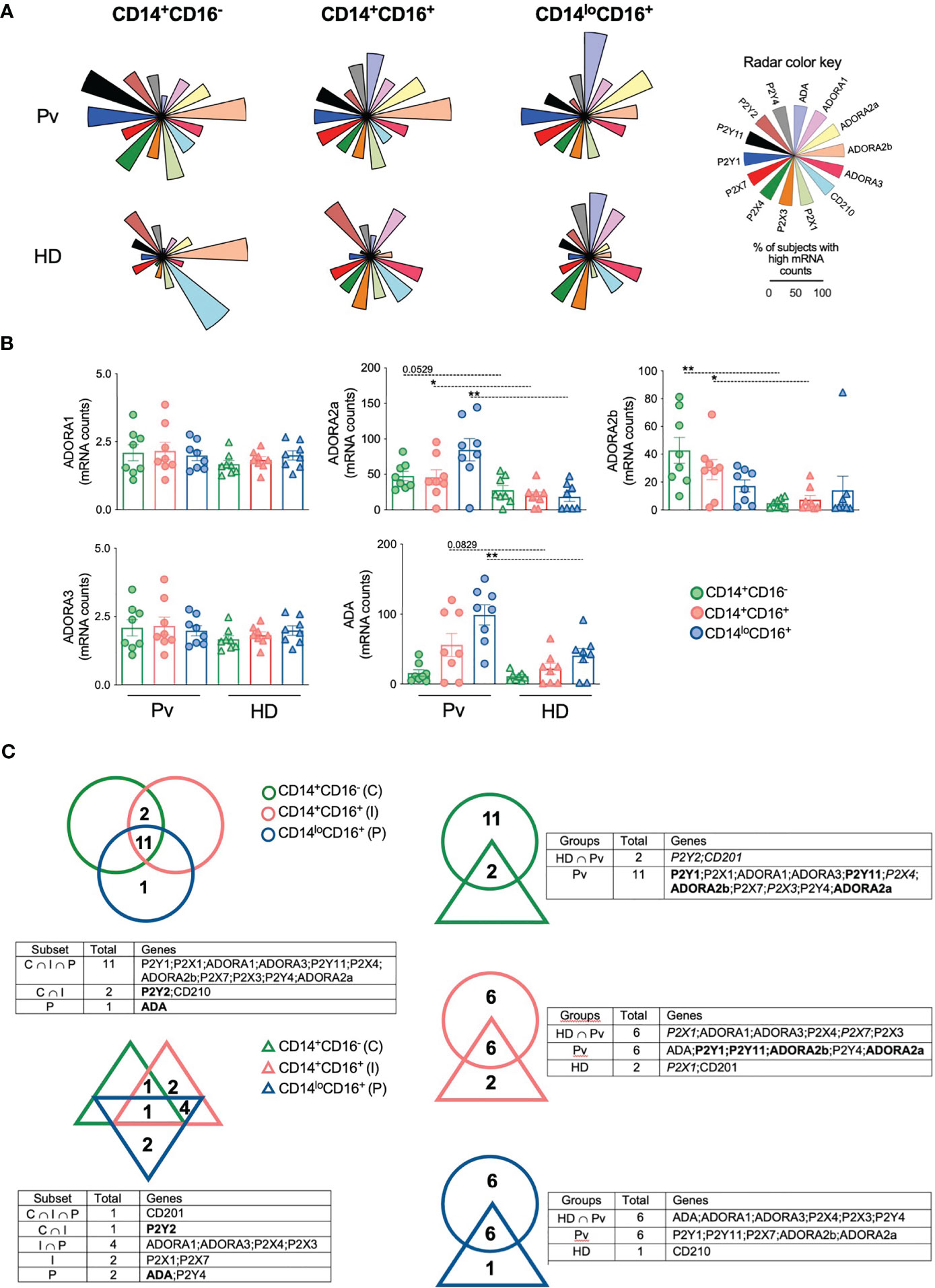
Figure 3 Plasmodium vivax infection alters the expression of adenosine and purinergic receptors in monocyte subsets. Gene expression was performed in monocyte subsets from HD (n = 8) and 8 Pv (n = 8) by NanoString analysis. (A) The proportion of subjects displaying high mRNA counts for each parameter was assessed using the overall median value as the cut-off, and radar graphs were assembled. Genes evaluated were ADA (purple); ADORA1 (pink); ADORA2a (yellow); ADORA2b (orange); ADORA3 (dark pink); CD210 (blue); P2X1 (green); P2X3 (dark orange); P2X4 (dark green); P2X7 (red); P2Y1 (dark blue); P2Y11 (black); P2Y2 (brown); and P2Y4 (gray). (B) Comparison of mRNA counts of adenosine receptors from monocyte subsets from Pv (circles) and HD (triangles). Scatter plots with bars representing mean ± SEM. *p ≤ 0.05 **p ≤ 0.01. (C) Venn diagrams of genes expressed in 50% or more subjects were built to identify common gene expression between each group. Gene expression among monocyte subsets from Pv (top, left panel) and HD (bottom, left panel). Genes in bold were commonly expressed by patients and HD in the same subset. Gene expression in each monocyte subset between Pv (circle) and HD (triangle) (right panel). Genes in bold were commonly expressed by Pv and HD or intersection group among the monocyte subsets analyzed. Genes in italic were exclusively expressed in a specific monocyte subset by Pv, HD or intersection group.
Venn diagrams were built to investigate the similarities and differences among monocyte subsets within the same context. Monocyte subsets from HD expressed only 10 genes in 50% or more subjects, and only CD210 was highly expressed in all subsets (Figure 3C, left lower panel). Noteworthy, 11 genes were expressed by the three subsets from Pv (Figure 3C, left top panel). When each monocyte subset from Pv and HD was compared, we observed that despite expressing a considerable number of genes in common, most of the genes analyzed are highly expressed only in the monocytes from Pv (Figure 3C, right panel), corroborating the hypothesis that Pv infection causes substantial changes in the expression of purinergic receptors by monocyte.
To better explore the impact of malaria on the expression of purinergic and IL-10 receptors, the proportion of subjects displaying each gene was represented as an ascending signature built using each gene’s overall median value (Figure S3). Those parameters observed in 50% or more of the subjects analyzed were highlighted for each monocyte subset. At least 50% of Pv express most of the genes analyzed above the overall median, but higher frequencies of lower expressors are observed in HD (Figure S3). Almost all genes analyzed were highly expressed in monocyte subsets among Pv, except for ADA in classical monocytes; and CD210 and P2Y2 in inflammatory and patrolling monocytes.
The significant changes observed in the expression of ectonucleotidases, and purinergic receptors suggest a role for adenosine during P. vivax infection. Systemic levels of adenosine, measured in the plasma, from Pv and HD (Figure 4A) were similar suggesting that their main role is in the cell microenvironment (Figure 4A). When monocytes from Pv were stimulated with LPS we observed a reduction on IL-10 levels that was not rescued by adenosine (Figure S4). However, adenosine triggered a decrease in the frequencies of TNF-producing monocytes from Pv stimulated with LPS (Figure 4B). In addition, the reduction of TNF production by monocytes induced by adenosine was reverted using ZM241385, a blocker for ADORA2a. The same was not observed with MRS1754, an ADORA2b antagonist (Figure 4B). These results indicate that adenosine act through ADORA2a diminishing the production of the inflammatory cytokine TNF by monocytes.
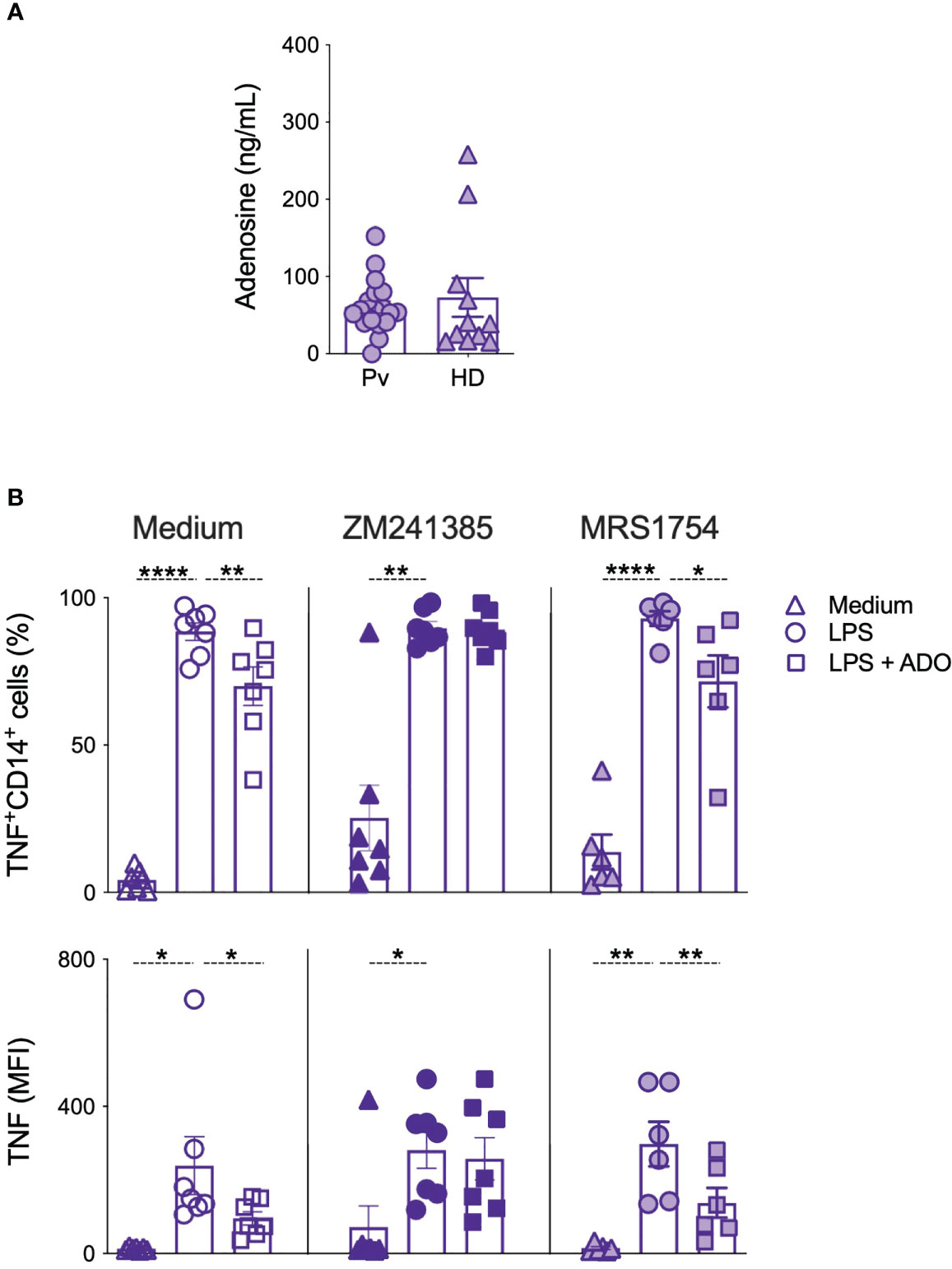
Figure 4 Adenosine decreases TNF production by monocytes in an adenosine 2a receptor depend-manner. (A) Adenosine levels were measured in the plasma of Pv (n = 30) and HD (n = 11) by UHPLC. (B) Frequencies (top panels) and mean fluorescence intensity (MFI) of TNF-producing monocytes were measured after culture with medium alone (triangles), LPS (circles), and LPS + adenosine (squares) and in the absence (open symbols) or presence of adenosine 2a receptor blocker (ZM241385, solid symbols) or adenosine 2b receptor blocker (MRS1754, gradient symbols) (n = 6–7). Scatter plots with bars representing mean ± SEM. *p ≤ 0.05 **p ≤ 0.01 ****p ≤ 0.001.
Plasmodium spp. triggers the production of high levels of inflammatory cytokines and their signaling are essential for both parasite control and symptoms observed during acute disease (2). Induction of IL-10 production is frequently observed in Pv-infected patients and previous studies have suggested a role for this cytokine in controlling immunopathology in human malaria. Individuals infected with P. vivax produce higher levels of circulating IL-10 than patients infected with P. falciparum (8), and different clinical manifestations of the disease are strongly associated with an imbalance in the production of inflammatory and regulatory cytokines (17).
Adenosine is a regulatory molecule associated with the control of the inflammatory responses by stimulating IL-10 production (15). Adenosine measurements in blood/plasma are technically difficult and unreliable due to its short life and rapid clearance (18). No adenosine systemic levels have been reported in malaria, and we detected no measurable increases in the adenosine levels during P. vivax infection. Nevertheless, measuring the expression of ectonucleotidases and adenosine receptors can be used as a substitute for direct measurements of adenosine levels.
The expression of ectonucleotidases is upregulated by pro-inflammatory cytokines, oxidative stress and hypoxia (19), conditions commonly observed during malaria. Previous studies showed that CD39 is heterogeneously expressed in leukocytes (20), being expressed in approximately 90% of neutrophils, monocytes, and B cells. Our results confirm these findings in both Pv-infected and healthy individuals. Classical and inflammatory monocytes expressed higher levels of CD39, whose expression is not altered by P. vivax infection. However, higher frequencies of CD73 and CD39/CD73 expressing cells were found among monocyte subsets from Pv-infected compared to HD. Taken together, CD73 overexpression in monocytes during P. vivax malaria may contribute to adenosine production. Moreover, since CD39 and CD73 correspond to sequential steps of hydrolysis of ATP to adenosine, it is plausible to think that their co-expression increases their efficiency. Our group demonstrated monocytes were also able to produce IL-10 when stimulated with Pv-RET (16). Interestingly, the Pv-RET phagocytosis did not alter the expression of ectonucleotidases in monocyte subsets, suggesting that the induction in the CD73 expression in monocytes from P. vivax-infected patients was due to the environment caused by the P. vivax infection.
Adenosine signals through surface P1 purinergic receptors expressed by immune cells (10, 13). We demonstrated that during acute P. vivax infection, monocyte subsets displayed distinct pattern of expression of P1 receptors. The role of P1 purinergic receptors has been extensively associated with the function of cells from the immune system (21), whereas TNF-induced A2a receptor as a negative feedback control (22). The binding of adenosine to A2a and A2b receptors inhibited TNF production (23) and increased IL-10 (15) release by macrophages. Accordingly, it has been shown that A1 receptors activation on monocytes enhanced Fcγ receptor-mediated phagocytosis, whereas A2 receptors reduced phagocytosis in the same cells (24). In addition, A2b activation also stimulated IL-10 and inhibited IL-12 and LPS-induced IL-18 production by human monocytes (15, 25). P. vivax infection increased the expression of A2a and A2b receptors in monocyte subsets, supporting the role for the adenosine system in immunomodulation during malaria. Adenosine inhibits LPS-induced TNF production in mouse macrophage cell lines (26). Our results confirmed these findings in human monocytes from Pv-infected individuals and that CD14+TNF+ cell frequency is reconstituted by blocking A2a receptor signaling by an antagonist. Notably, the same was not observed by using the A2b antagonist. Previous studies have shown that a high concentration of adenosine is necessary to induce IL-10 compared to the concentration capable of inhibiting TNF production (27). This might be one of the reasons we were not able to detect the modulation of IL-10 production by A2a receptor signaling.
Our work raise several questions that we hope to address in the future, such as the potential role of adenosine pathways in different clinical presentations of malaria, including correlation with the subject parasitemia, number of acute malaria episodes and even in asymptomatic Pv infections. Also it remains to be identified the mechanistic nature by which ectonucleotidases expression is modulated by the environment or the systemic alterations caused by Pv infection.
Altogether, our findings demonstrated that monocyte subsets expressed high levels of the ectonucleotidases CD39 and CD73, suggesting a role for these cells in adenosine production and consequently in the modulation of inflammatory response stimulated by malaria parasites. Despite not detecting differences in plasmatic circulating adenosine levels, we demonstrate that monocyte subsets from Pv-infected patients displayed augmented expression of A2a and A2b receptors, indicating a potential increase in the consumption of adenosine. In addition, we demonstrated that adenosine modulates inflammatory response by decreasing TNF production by monocytes, which is restored with the blockage of ADORA2a. Ultimately, the present study attempted to contribute to the knowledge of immunoregulation during malaria.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by Committees on Human Experimentation from Centro de Pesquisa em Medicina Tropical de Rondônia (CEP-CEPEM 095/2009) and Instituto René Rachou, Fundação Oswaldo Cruz (CEP-IRR 2004), the National Ethical Committee (CONEP 15652) from Ministry of Health, Brazil. The patients/participants provided their written informed consent to participate in this study.
SD, LVA designed the research. SD, AC, MF, PC, MK, CZ performed experiments and discussed data. RG, LCA discussed data and provided reagents. DP, MT participated in the enrollment and provided clinical care. SD, OM-F, RG, FO, LVA performed data analysis. SQD, FO, LA wrote the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (422694/2016-9), INCT-Vacinas (465293/2014-0, CBB-APQ-03608-17), and by the intramural research program of the NIAID, NIH, and USA (FO). LVA, AC, OM-F, and RG are CNPq fellows (PQ). OM-F and AC are research fellows from FAPEAM (PVN-II, PRÓ-ESTADO Program #005/2019. Additionally, the authors thank Yolanda L. Jones, NIH Library Editing Services, for editing assistance. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
We thank the collaborating nurses for patient care and to Clecia O.A. Vieira, Ana Beatriz Queiroz, Cristiane P. Gomes, and Patricia Palhares for excellent technical assistance. We thank Cristina T. Fonseca and Rosiane A. Pereira for scientific discussions. We acknowledge the Program for Technological Development in Tools for Health‐FIOCRUZ for the use of the Flow Cytometry Platform. NanoString nCounter digital gene expression was performed by the CCR Genomics Core at National Cancer Institute/NIH (Bethesda, MD).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1193256/full#supplementary-material
ADP, Adenosine diphosphate; AMP, Adenosine monophosphate; ATP, Adenosine triphosphate; cAMP, Cyclic adenosine monophosphate; CBA, Cytometric bead array; CEPEM, Centro de Pesquisa de Medicina Tropical de Rondônia; HD, Healthy donors; HPC, High precision calibration; UHPLC, Ultra-High high performance liquid chromatography; IL, Interleukin; LPS, Lipopolysaccharides; MS, Mass spectrometry; PBMC, Peripheral blood mononuclear cells; Pv-RET, Plasmodium vivax-infected reticulocytes; TNF, Tumor necrosis factor.
1. W.H.O. World malaria report: 2018. (2018). Available at: https://apps.who.int/iris/handle/10665/275867.
2. Kumar R, Ng S, Engwerda C. The role of il-10 in malaria: a double edged sword. Front Immunol (2019) 10:229. doi: 10.3389/fimmu.2019.00229
3. Stevenson MM, Riley EM. Innate immunity to malaria. Nat Rev Immunol (2004) 4(3):169–80. doi: 10.1038/nri1311
4. Walther M, Woodruff J, Edele F, Jeffries D, Tongren JE, King E, et al. Innate immune responses to human malaria: heterogeneous cytokine responses to blood-stage plasmodium falciparum correlate with parasitological and clinical outcomes. J Immunol (2006) 177(8):5736–45. doi: 10.4049/jimmunol.177.8.5736
5. Couper KN, Blount DG, Riley EM. Il-10: the master regulator of immunity to infection. J Immunol (2008) 180(9):5771–7. doi: 10.4049/jimmunol.180.9.5771
6. Jensen AT, Gasim S, Ismail A, Gaafar A, Kurtzhals JA, Kemp M, et al. Humoral and cellular immune responses to synthetic peptides of the leishmania donovani kinetoplastid membrane protein-11. Scandinavian J Immunol (1998) 48(1):103–9. doi: 10.1046/j.1365-3083.1998.00370.x
7. Othoro C, Lal AA, Nahlen B, Koech D, Orago AS, Udhayakumar V. A low interleukin-10 tumor necrosis factor-alpha ratio is associated with malaria anemia in children residing in a holoendemic malaria region in Western Kenya. J Infect Dis (1999) 179(1):279–82. doi: 10.1086/314548
8. Goncalves RM, Scopel KK, Bastos MS, Ferreira MU. Cytokine balance in human malaria: does plasmodium vivax elicit more inflammatory responses than plasmodium falciparum? PloS One (2012) 7(9):e44394. doi: 10.1371/journal.pone.0044394
9. Di Virgilio F. Purinergic mechanism in the immune system: a signal of danger for dendritic cells. Purinergic Signal (2005) 1(3):205–9. doi: 10.1007/s11302-005-6312-z
10. Abbracchio MP, Burnstock G. Purinoceptors: are there families of P2x and P2y purinoceptors? Pharmacol Ther (1994) 64(3):445–75. doi: 10.1016/0163-7258(94)00048-4
11. Schnurr M, Then F, Galambos P, Scholz C, Siegmund B, Endres S, et al. Extracellular atp and tnf-alpha synergize in the activation and maturation of human dendritic cells. J Immunol (2000) 165(8):4704–9. doi: 10.4049/jimmunol.165.8.4704
12. Antonioli L, Pacher P, Vizi ES, Hasko G. Cd39 and Cd73 in immunity and inflammation. Trends Mol Med (2013) 19(6):355–67. doi: 10.1016/j.molmed.2013.03.005
13. Hasko G, Linden J, Cronstein B, Pacher P. Adenosine receptors: therapeutic aspects for inflammatory and immune diseases. Nat Rev Drug Discovery (2008) 7(9):759–70. doi: 10.1038/nrd2638
14. Hasko G, Kuhel DG, Chen JF, Schwarzschild MA, Deitch EA, Mabley JG, et al. Adenosine inhibits il-12 and tnf-[Alpha] production Via adenosine A2a receptor-dependent and independent mechanisms. FASEB J (2000) 14(13):2065–74. doi: 10.1096/fj.99-0508com
15. Nemeth ZH, Lutz CS, Csoka B, Deitch EA, Leibovich SJ, Gause WC, et al. Adenosine augments il-10 production by macrophages through an A2b receptor-mediated posttranscriptional mechanism. J Immunol (2005) 175(12):8260–70. doi: 10.4049/jimmunol.175.12.8260
16. Antonelli LR, Leoratti FM, Costa PA, Rocha BC, Diniz SQ, Tada MS, et al. The Cd14+Cd16+ inflammatory monocyte subset displays increased mitochondrial activity and effector function during acute plasmodium vivax malaria. PloS Pathog (2014) 9:e1004393. doi: 10.1371/journal.ppat.1004393
17. Andrade BB, Reis-Filho A, Souza-Neto SM, Clarencio J, Camargo LM, Barral A, et al. Severe plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar J (2010) 9:13. doi: 10.1186/1475-2875-9-13
18. Ramakers BP, Pickkers P, Deussen A, Rongen GA, van den Broek P, van der Hoeven JG, et al. Measurement of the endogenous adenosine concentration in humans in vivo: methodological considerations. Curr Drug Metab (2008) 9(8):679–85. doi: 10.2174/138920008786049249
19. Deaglio S, Robson SC. Ectonucleotidases as regulators of purinergic signaling in thrombosis, inflammation, and immunity. Adv Pharmacol (2011) 61:301–32. doi: 10.1016/B978-0-12-385526-8.00010-2
20. Koziak K, Sevigny J, Robson SC, Siegel JB, Kaczmarek E. Analysis of Cd39/Atp diphosphohydrolase (Atpdase) expression in endothelial cells, platelets and leukocytes. Thromb Haemost (1999) 82(5):1538–44. doi: 10.1055/s-0037-1614868
21. Xaus J, Mirabet M, Lloberas J, Soler C, Lluis C, Franco R, et al. Ifn-gamma up-regulates the A2b adenosine receptor expression in macrophages: a mechanism of macrophage deactivation. J Immunol (1999) 162(6):3607–14. doi: 10.4049/jimmunol.162.6.3607
22. Murphree LJ, Sullivan GW, Marshall MA, Linden J. Lipopolysaccharide rapidly modifies adenosine receptor transcripts in murine and human macrophages: role of nf-kappab in a(2a) adenosine receptor induction. Biochem J (2005) 391(Pt 3):575–80. doi: 10.1042/BJ20050888
23. Kreckler LM, Wan TC, Ge ZD, Auchampach JA. Adenosine inhibits tumor necrosis factor-alpha release from mouse peritoneal macrophages Via A2a and A2b but not the A3 adenosine receptor. J Pharmacol Exp Ther (2006) 317(1):172–80. doi: 10.1124/jpet.105.096016
24. Le Vraux V, Chen YL, Masson I, De Sousa M, Giroud JP, Florentin I, et al. Inhibition of human monocyte tnf production by adenosine receptor agonists. Life Sci (1993) 52(24):1917–24. doi: 10.1016/0024-3205(93)90632-d
25. Chen L, Zhang ZH, Watanabe T, Yamashita T, Kobayakawa T, Kaneko A, et al. The involvement of neutrophils in the resistance to leishmania major infection in susceptible but not in resistant mice. Parasitol Int (2005) 54(2):109–18. doi: 10.1016/j.parint.2005.02.001
26. McWhinney CD, Dudley MW, Bowlin TL, Peet NP, Schook L, Bradshaw M, et al. Activation of adenosine A3 receptors on macrophages inhibits tumor necrosis factor-alpha. Eur J Pharmacol (1996) 310(2-3):209–16. doi: 10.1016/0014-2999(96)00272-5
Keywords: malaria, Plasmodium vivax, ectonucleotidases, adenosine, regulation
Citation: Diniz SQ, Figueiredo MM, Costa PAC, Martins-Filho OA, Teixeira-Carvalho A, Pereira DB, Shugiro Tada M, Crocco Afonso LC, Kohlhoff M, Zani CL, Gazzinelli RT, Oliveira F and Antonelli LR (2023) Adenosine pathway regulates inflammation during Plasmodium vivax infection. Front. Immunol. 14:1193256. doi: 10.3389/fimmu.2023.1193256
Received: 24 March 2023; Accepted: 19 June 2023;
Published: 21 July 2023.
Edited by:
Katherine Rose Dobbs, Case Western Reserve University, United StatesReviewed by:
George Hasko, Columbia University, United StatesCopyright © 2023 Diniz, Figueiredo, Costa, Martins-Filho, Teixeira-Carvalho, Pereira, Shugiro Tada, Crocco Afonso, Kohlhoff, Zani, Gazzinelli, Oliveira and Antonelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lis Ribeiro Antonelli, bGlzLmFudG9uZWxsaUBmaW9jcnV6LmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.