
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 02 June 2023
Sec. Viral Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1178355
This article is part of the Research TopicCOVID and Emerging Infectious DiseasesView all 47 articles
 William D. Tolbert1*
William D. Tolbert1* Yaozong Chen1
Yaozong Chen1 Lulu Sun2Mehdi Benlarbi3,4Shilei Ding3Rohini Manickam1†
Lulu Sun2Mehdi Benlarbi3,4Shilei Ding3Rohini Manickam1† Emily Pangaro1Dung N. Nguyen1Suneetha Gottumukkala1Marceline Côté5
Emily Pangaro1Dung N. Nguyen1Suneetha Gottumukkala1Marceline Côté5 Frank J. Gonzalez2Andrés Finzi3,4Zahra R. Tehrani6,7
Frank J. Gonzalez2Andrés Finzi3,4Zahra R. Tehrani6,7 Mohammad M. Sajadi6,7,8
Mohammad M. Sajadi6,7,8 Marzena Pazgier1*
Marzena Pazgier1*SARS-CoV-2, the virus behind the COVID-19 pandemic, has changed over time to the extent that the current virus is substantially different from what originally led to the pandemic in 2019–2020. Viral variants have modified the severity and transmissibility of the disease and continue do so. How much of this change is due to viral fitness versus a response to immune pressure is hard to define. One class of antibodies that continues to afford some level of protection from emerging variants are those that closely overlap the binding site for angiotensin-converting enzyme 2 (ACE2) on the receptor binding domain (RBD). Some members of this class that were identified early in the course of the pandemic arose from the VH 3-53 germline gene (IGHV3-53*01) and had short heavy chain complementarity-determining region 3s (CDR H3s). Here, we describe the molecular basis of the SARS-CoV-2 RBD recognition by the anti-RBD monoclonal antibody CoV11 isolated early in the COVID-19 pandemic and show how its unique mode of binding the RBD determines its neutralization breadth. CoV11 utilizes a heavy chain VH 3-53 and a light chain VK 3-20 germline sequence to bind to the RBD. Two of CoV11’s four heavy chain changes from the VH 3-53 germline sequence, to Ile and to Arg, and some unique features in its CDR H3 increase its affinity to the RBD, while the four light chain changes from the VK 3-20 germline sequence sit outside of the RBD binding site. Antibodies of this type can retain significant affinity and neutralization potency against variants of concern (VOCs) that have diverged significantly from original virus lineage such as the prevalent omicron variant. We also discuss the mechanism by which VH 3-53 encoded antibodies recognize spike antigen and show how minimal changes to their sequence, their choice of light chain, and their mode of binding influence their affinity and impact their neutralization breadth.
Since the beginning of the COVID-19 pandemic, SARS-CoV-2 has been accumulating mutations to enhance infectivity and to avoid immune pressure (1). While SARS-CoV-2 has a much lower mutation rate than other RNA viruses such as influenza or HIV-1, it still mutates in response to its environment and over time due to genetic drift (2). Because SARS-CoV-2 has one major viral protein on its surface accessible to antibodies, the spike glycoprotein that is responsible for both target cell recognition and viral entry, it has become the focus of many of these mutations. The spike protein is a membrane-anchored trimer that is cleaved into S1 and S2 subunits by furin in the expressing cell (3, 4). S1 contains an N-terminal domain and a receptor binding domain (RBD) that recognizes the target cell receptor angiotensin-converting enzyme 2 (ACE2). S2 contains the fusion machinery. After the spike protein binds its target cell, it must be further proteolyzed to expose the fusion peptide in S2, which then inserts into the target cell membrane. This can occur at the cell surface of the target cell by proteases such as TMPRSS2 or in endosomes by proteases such as the cathepsins (5). Cell fusion and viral entry begin with removal of S1 from the trimer, which destabilizes the trimer and causes conformational rearrangements in S2. The initiating event in this process is the binding of the RBD to ACE2, which makes it a prime target for neutralizing antibodies (6–8).
The RBD sits at the top of the trimer and can exist in two different conformations, one with the RBD up and the ACE2 binding site accessible to the solvent and one with the RBD down and the ACE2 binding site occluded within the trimer (4). Each RBD in the trimer can exist in either conformation giving the spike a range of conformations from fully closed with all three RBDs in the down position to fully open with all three RBDs in the up conformation. One of the first mutations in the spike protein to spread and outcompete the original strain was the Asp614-to-Gly (D614G) mutation that removed a stabilizing hydrogen bond that increased the propensity for the RBD to be in the up conformation (9). This mutation likely increased infectivity by increasing the percentage of RBDs accessible to ACE2 for binding (10), but it came at a cost to the virus. It also made the RBD more accessible to neutralizing antibodies. Since then, viral variants such as alpha to the more recent flavors of omicron subvariants have appeared with mutations that can nullify the activity of some of these neutralizing antibodies. This has enabled the virus to continue to infect individuals even when they had been vaccinated or infected by a previous strain.
One group of antibodies that was identified earlier in the pandemic and continues to neutralize the virus and its emerging variants relatively well are those derived from the VH 3-53 (IGHV3-53*01) heavy chain germline gene with short heavy chain complementarity-determining region 3s (CDR H3s) (11). These antibodies closely match the ACE2 binding site on the RBD, explaining in part their continued activity against the virus (12, 13). Initial reports also suggested that they could represent as much as 10% of the neutralizing antibody response elicited by SARS-CoV-2 (11). Structures of the first antibodies of this type were quickly determined and confirmed that they overlap the ACE2 binding site on the RBD (11–20). The receptor binding ridge on the RBD places a boundary on CDR H3 length and provides an explanation for the preference of short CDR H3s. Indeed when examples of VH 3-53 encoded SARS-CoV-2 neutralizing antibodies with longer CDR H3s were determined, many were found to bind the RBD differently (12). The high frequency of this germline gene in the neutralization response to SARS-CoV-2 could be the result of two independent factors, the prevalence of the VH 3-53 (and closely related VH 3-66) germline genes in the general population, one study estimated a frequency of approximately 1% for VH 3-53 in healthy individuals who had not been exposed to SARS-CoV-2 (21), and the low degree of somatic hypermutation necessary to confer specificity, as few as two mutations in the germline sequence was enough to increase one antibody’s affinity to the RBD from 407 nM to 3.6 nM (13).
Here, we report the structure of one example of a VH 3-53 encoded SARS-CoV-2 neutralizing antibody, CoV11, and show how its mode of binding to the SARS-CoV-2 RBD leads to neutralization breadth. CoV11 was isolated from an infected individual early in the course of the pandemic but continues to neutralize many SARS-CoV-2 variants of concern (VOCs) relatively well. While VH 3-53 encoded antibodies have similar sequences and modes of binding to the SARS-CoV-2 RBD, small differences in sequence and their choice of light chain influence their affinity and impact their neutralization breadth.
CoV11 was isolated from a 76-year-old man with history of severe COVID-19, infected in February 2020 with high titer of neutralizing antibodies against the virus. CoV11 (also known as CoVIC-079) has been previously published (22). Briefly, memory B cells were isolated from PBMCs using the EasySep™ Human Memory B Cell Isolation Kit (STEMCELL Technologies). Cells were incubated with biotin-conjugated SARS-CoV-2 Spike antigen (LakePharma) and then, after washing, labeled by incubation with TotalSeq™-C0953 PE Streptavidin (BioLegend). The cell surface labeled single-cell suspension was loaded onto a 10x Genomics Chromium Controller microfluidics chip (10x Genomics) and a VDJ library was prepared based on the manufacturer’s instructions. A subset of cells containing the surface barcode were selected and their VDJ sequences were cloned into IgG1 heavy and light chain vectors. The recombinant plasmids were then co-transfected into FreeStyle-293 cells for expression and the secreted antibodies were purified from culture supernatants after incubations of 1 week by Protein A affinity chromatography. CoV11 was identified as one of the antibodies that strongly neutralized the virus.
SARS-CoV-2 RBD was cloned from an expression plasmid of the SARS-CoV-2 2P soluble spike protein (a gift from Jason McLellan) (residues 319–537) into an expression plasmid with an N-terminal leader sequence and a C-terminal 6-histidine tag. SARS-CoV-2 RDBdelta was made by adding T478K and L452R mutations to the wild-type RBD using a QuikChange mutagenesis kit (Stratagene). CoV11 IgG was produced by transection of CoV11 heavy and light chain plasmids into expi293F cells and SARS-CoV-2 RBDwt or RDBdelta were produced by transfection into GnT1- 293F cells. Cells were then grown in suspension for an additional 7 days at 37°C and 90% humidity. Cells were pelleted by centrifugation and the medium was filtered through a 0.2-micron filter. CoV11 IgG was purified from medium passed over a HiTrap protein A column (Cytiva) equilibrated with phosphate buffered saline (PBS), pH 7.2. The column was washed with PBS and the IgG was eluted with 0.1 M glycine, pH 3.0. Eluted fractions were immediately diluted 10:1 with 1 M tris(hydroxymethyl)aminomethane-HCl (Tris-HCl), pH 8.5. Eluted protein was concentrated to approximately 10 mg/ml and the buffer was exchanged for PBS, pH 7.2. SARS-CoV-2 RBD was purified using a HisTrap column (Cytiva) equilibrated in wash buffer, 25 mM Tris-HCl, pH 8.0, and 500 mM sodium chloride. Medium was passed over the column and the column was washed with wash buffer. Protein was eluted with 25 mM Tris-HCl and 500 mM imidazole, pH 8.0. Eluted protein was concentrated, and the sample was loaded onto a Superdex 200 gel filtration column (Cytiva) equilibrated in 10 mM Tris-HCl, pH 7.2, and 100 mM ammonium acetate. Fractions corresponding to the correct RBD size were concentrated and used for complex formation.
CoV11 Fab was generated from IgG by papain cleavage. IgG in PBS was first exchanged to Fab digest buffer, 10 mM sodium phosphate, pH 7.0, and 5 mM cysteine. IgG (10 mg) at approximately 10 mg/ml was added to 3 ml of papain-linked agarose slurry (Thermo Fisher) previously equilibrated in Fab digest buffer. The digest was incubated at 37°C for 3 h and then filtered to remove the papain. Filtered protein was passed over a protein A column equilibrated in PBS, pH 7.2. Flow through fractions containing the Fab were concentrated and then loaded onto a Superdex 200 gel filtration column (Cytiva) equilibrated in 10 mM Tris-HCl, pH 7.2, and 100 mM ammonium acetate. The Fab elution peak corresponding to a molecular weight of approximately 50 kDa was collected and concentrated for use in complex formation or SPR.
Complexes were made by mixing Fab and RBD at a 1:1 ratio and incubation on ice for 30 min. The sample was then loaded onto a Superdex 200 gel filtration column (Cytiva) equilibrated in 10 mM Tris-HCl, pH 7.2, and 100 mM ammonium acetate. Fractions corresponding to the complex molecular weight were collected and concentrated to approximately 10 mg/ml for use in crystallization trials.
SARS-CoV-2 S-pseudotyped virus-like particles (VLPs) containing the synthetic firefly luciferase (Luc2) reporter were prepared using the BEI NR-52948 kit as described in (22). To generate VLP pseudotyped with spikes of different SARS-CoV-2 VOCs, spike pseudotyping vector plasmids, including WA-1/2020 (WT), D614G (BEI Resources), gamma (a gift from Dr. Robert Petrovich from NIEHS), alpha, beta, epsilon, iota, delta (InvivoGen), and omicron BA.1 (Sino Biological), BQ.1.1 (23, 24) and XBB.1.5 (25) VOCs, were used. Sixteen to 24 h post seeding, 293T cells (Thermo Fisher Scientific) were co-transfected with respective spike plasmid or VSV G (positive control), lentiviral backbone, and three helper plasmids encoding Gag, Tat1b, and Rev1b (BEI Resources). At 72 h post-transfection, the supernatant was harvested and clarified by a 0.45-μm filter. To determine viral titers, hACE2-expressing 293T cells (gift from Dr. Allison Malloy, USUHS) were infected with serial VLP dilutions. Forty-eight to 60 h post-infection, luciferase signal was detected by the Bright-Go Luciferase Assay System (Promega) for titer estimations (26). VLPs were concentrated by a homemade fourfold lentivirus concentrator (protocol of MD Anderson) and stored at 4°C for short-term use or −20°C for longer storage.
For in vitro neutralization assays, 50-μl serial dilutions of CoV11 or synagis with final concentrations from 0.005 to 50 ng/μl were pre-incubated with 50 μl of SARS-CoV-2 spike-pseudotyped VLPs (~106 RLU/ml) of WT or one of eight VOCs in 96-well plates at 37°C for 1 h. Subsequently, Hace2-expressing 293T cells (1.25 × 104 cells/well) in 50 μl of culture medium were added and incubated at 37°C for 48 h before luciferase signal measurement with the Bright-Go Luciferase Assay System (Promega). Data analysis and normalization followed the protocol described previously in (26). Pseudoviral neutralization experiments presented in Figure S3 were performed as we previously reported (23). Briefly, 293T cells were transfected by the calcium phosphate method with the lentiviral vector Pnl4.3 R-E- Luc (NIH AIDS Reagent Program) and a plasmid encoding for SARS-CoV-2 Spikes at a ratio of 5:4. Two days post-transfection, cell supernatants were harvested and stored at –80°C until use. 293T-ACE2 target cells were seeded at a density of 1×104 cells/well in 96-well luminometer-compatible tissue culture plates (Perkin Elmer) 24 h before infection. Recombinant viruses in a final volume of 100 μl were incubated with the indicated concentrations of mAbs for 1 h at 37°C and were then added to the target cells followed by incubation for 48 h at 37°C; cells were lysed by the addition of 30 μl of passive lysis buffer (Promega) followed by one freeze–thaw cycle. An LB941 TriStar luminometer (Berthold Technologies) was used to measure the luciferase activity of each well after the addition of 100 μl of luciferin buffer (15 Mm MgSO4, 15 Mm KPO4 [Ph 7.8], 1 Mm ATP, and 1 Mm dithiothreitol) and 50 μl of 1 Mm d-luciferin potassium salt (Prolume). Relative lighting unit (RLU) for luciferase activity was recorded and the ratio to “no mAb” was calculated. VSV-G pseudoviral particles were used as a specificity control. Surface plasmon resonance (SPR) measurements were carried out as described in (27). All assays were performed on a Biacore 3000 (Cytiva) at room temperature using 10 Mm HEPES, Ph 7.5, 150 Mm NaCl and 0.05% Tween 20 as running buffer. For the kinetic measurement, ~80–200 RU of CoV11 IgG was immobilized on a Protein A sensor chip (Cytiva) and twofold serial dilutions of SARS-CoV-2 RBD from WT or one of six VOCs was then injected as solute analyte with concentrations ranging from 6.25 to 200 Nm. For all kinetic assays, the sensor chip was regenerated using 10 Mm glycine, Ph 2.5, before the next cycle. Sensorgrams were corrected by subtraction of the corresponding blank channel as well as for the buffer background and kinetic constants were determined using a 1:1 Langmuir model with the BIAevaluation software (Cytiva) as shown in Table S2 and Figure S1. Goodness of fit of the curve was evaluated by the χ2 of the fit with a value below 3 considered acceptable.
Binding kinetics were performed using an Octet RED96e system (ForteBio) with shaking at 1,000 RPM. Protein A (ProA) biosensors were hydrated in water prior to use. CoV11 mAb was loaded into ProA biosensors at 12.5 µg/ml in 10X kinetic buffer (Fortebio) for 120 s. Loaded biosensors were placed in 10X kinetic buffer (ForteBio) for 120 s for baseline equilibration. Association of CoV11 mAb (in 10X kinetics buffer) to the different RBD proteins was carried out for 180 s at various RBD concentrations in a twofold dilution series from 200 nM to 6.25 nM, prior to dissociation for 300 s. The data were baseline subtracted prior to fitting performed using a 1:1 binding model and the ForteBio data analysis software. Calculation of on rates (ka), off rates (kd), and affinity constants (KD) were computed using a global fit applied to all data.
Initial crystals were grown from Molecular Dimensions sparse matrix screens, specifically ProPlex Eco and MacroSol Eco screens. Crystals were then reproduced and optimized using the hanging-drop, vapor diffusion method. CoV11 Fab-RBDwt crystals were grown from 15% polyethylene glycol (PEG) 4000, 150 mM ammonium sulfate, and 100 mM 2-(N-morpholino)ethanesulfonic acid (MES), pH 6.0. CoV11 Fab-RBDdelta crystals were grown from 10% PEG 4000, 200 mM sodium acetate, and 100 mM sodium citrate, pH 5.5 (crystal form 1), and 12% PEG 8000 and 100 mM sodium phosphate, pH 6.5 (crystal form 2). Prior to freezing, crystals were briefly soaked in the crystallization condition supplemented with 20% of 2-methyl-2,4-pentanediol (MPD) as cryoprotectant.
Diffraction data were collected at the Stanford Synchrotron Radiation Light Source (SSRL) beamline 12-2 on a Dectris Pilatus 6M area detector. All data were processed and reduced with HKL3000 (28). Structures were solved by molecular replacement with PHASER from the CCP4 suite (29) based on the coordinates from PDB ID 7JMP. Refinement was carried out with Refmac5 (29) and/or Phenix (30) and model building was done with COOT (29). Data collection and refinement statistics are shown in Table 1. Ramachandran statistics were calculated with MolProbity and illustrations were prepared with Pymol Molecular graphics (http://pymol.org).
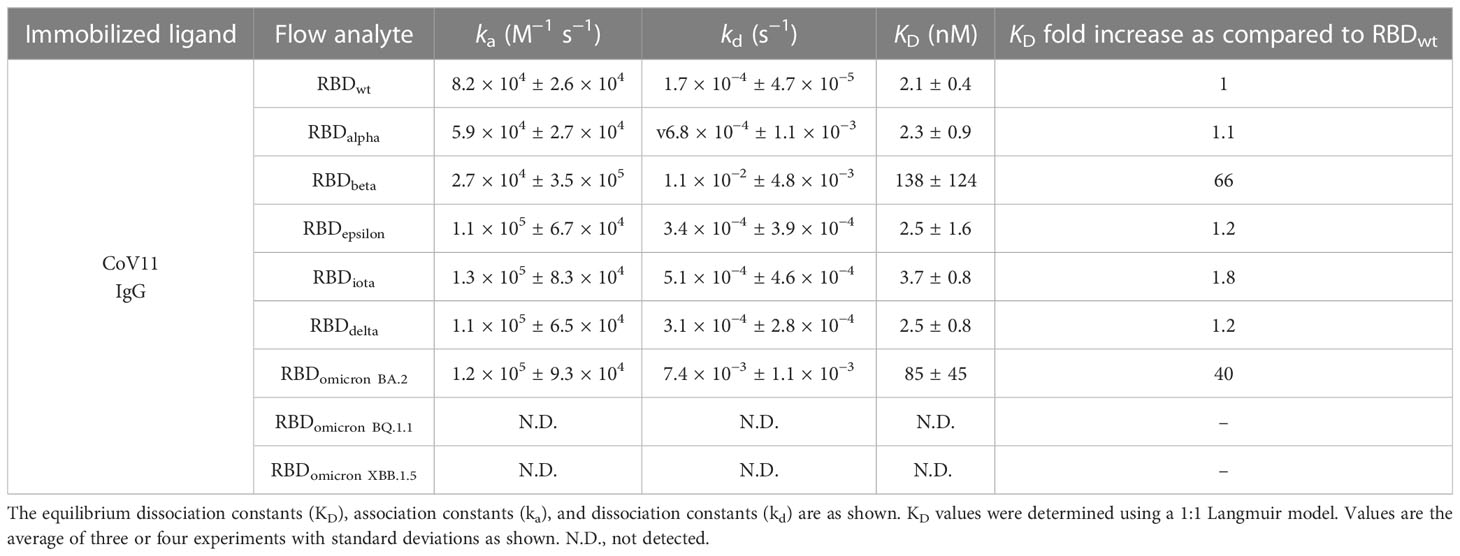
Table 1 Combined kinetic constants (BLI and SPR) of CoV11 binding to the RBD of SARS-CoV-2 wt and selected VOCs.
CoV11 is derived from the heavy chain VH 3-53 (IGHV3-53*01) and light chain VK 3-20 (IGKV3-20*01) germline genes and has four mutations in its heavy chain and four in its light chain relative to germline sequences. It also has a short CDR H3 of 10 amino acids, which is the product of heavy chain joining and diversity germline sequences. When tested for recognition of RBD of different SARS-CoV2 variants, CoV11 binds the original, wild type (wt), SARS-CoV-2 strain RBD with a KD of 2.1 nM and potently neutralizes SARS-CoV-2 wt pseudotyped viruses with an IC50 of 0.003 µg/ml (Tables 1, S2, S3 and Figure 1). CoV11’s affinity to the RBD and neutralization potency are reduced by SARS-CoV-2 VOCs, but to different degrees, most likely due to the avidity of an IgG as compared to a Fab. CoV11’s affinity to alpha (B.1.1.7) RBD is 2.3 nM and that to beta (B.1.351) RBD is 10.4 nM as determined by SPR (Table S2) and 202 nM as determined by bio-layer interferometry (BLI) (Table S3); CoV11’s affinity to epsilon (B.1.427/B.1.429) RBD is 2.5 nM, that to iota (B.1.526) RBD is 3.7 nM, that to delta (B.1.617.2) RBD is 2.5 nM, and that to omicron BA.2 RBD is 24.4 nM as determined by SPR and 105 nM as determined by BLI. We observed higher KDs for beta and omicron BA.2 determinations by BLI as compared to those determined by SPR. This could be in part be due to differences in the two techniques, but for the beta RBD, it could also be due to the use of commercially prepared protein used for BLI measurements versus in-house-produced protein used for SPR measurements. The KDs for the other RBDs were comparable and are averaged (Table 1). In comparison, the IC50 of CoV11 IgG to pseudotyped alpha is 0.02 µg/ml; beta, 1.3 µg/ml; epsilon, 0.01 µg/ml; gamma (P.1), 0.07 µg/ml; iota, 0.04 µg/ml; delta, 0.05 µg/ml; omicron (B.1.1.529.1 or BA.1), 1.9 µg/ml (Figure 1). Thus, the avidity of an IgG relative to a Fab largely mitigates small losses in CoV11’s affinity due to escape mutations at the antibody–RBD interface in VOCs. Of note, the most detrimental mutations in VOCs for CoV11 occur in the beta and omicron RBDs (Figure 1 and Tables 1, S2, and S3).
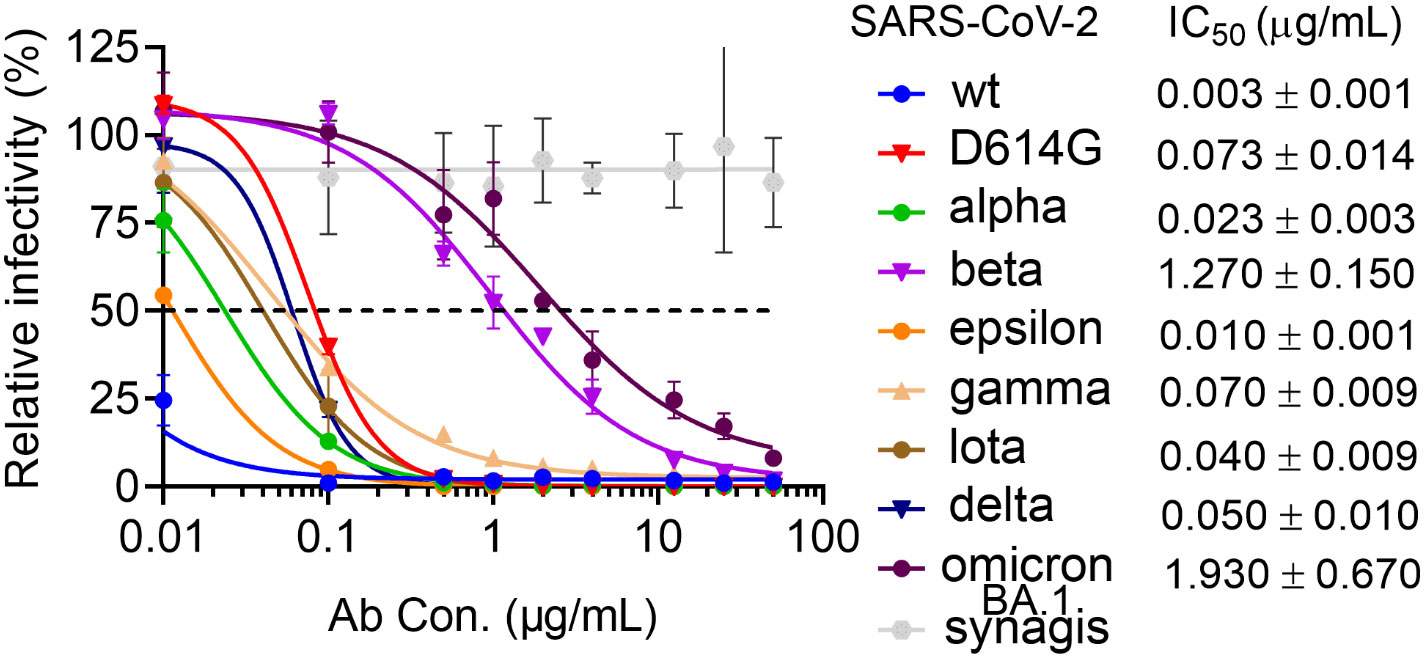
Figure 1 Neutralization activity of COV11 against SARS-CoV-2wt and VOCs. Dose–response neutralization curves of SARS-CoV-2 pseudotyped lentivirus with eight SAR2-CoV-2 S variants. hACE2 expressing 293T cells were infected with different variants of SARS-CoV-2 PsV in the presence of CoV11, Synagis IgG (negative control), or PBS saline. Infectivity was quantified by the cellular luciferase signal 48 h post-infection. Relative infectivity was normalized by the luciferase signal in infected cells without intervention (PBS saline). EC50 values for individual strains are shown to the right.
To get a better understanding of how CoV11 recognizes SARS-CoV2 spike, we determined the crystal structure of CoV11 Fab in complex with the wt and the delta (B.1.617.2) variants of the SARS-CoV-2 RBD. CoV11 Fab-RBDwt crystals belonged to space group P212121 and diffracted to 2.05 Å with one CoV11 Fab-RBDwt complex in the asymmetric unit (Figure 2A and Table 2). The wt RBD complex was refined to an R/Rfree of 0.166/0.198. CoV11 Fab-RBDdelta crystallized in two different space groups, P212121, which diffracted to 2.05 Å, and C2, which diffracted to 2.4 Å. Both crystals had one CoV11 Fab-RBDdelta complex in the asymmetric unit. The orthorhombic delta RBD P212121 crystal form was refined to an R/Rfree of 0.177/0.208 and the monoclinic delta RBD C2 crystal form was refined to an R/Rfree of 0.179/0.224. Complete data collection and refinement statistics can be found in Table 2.
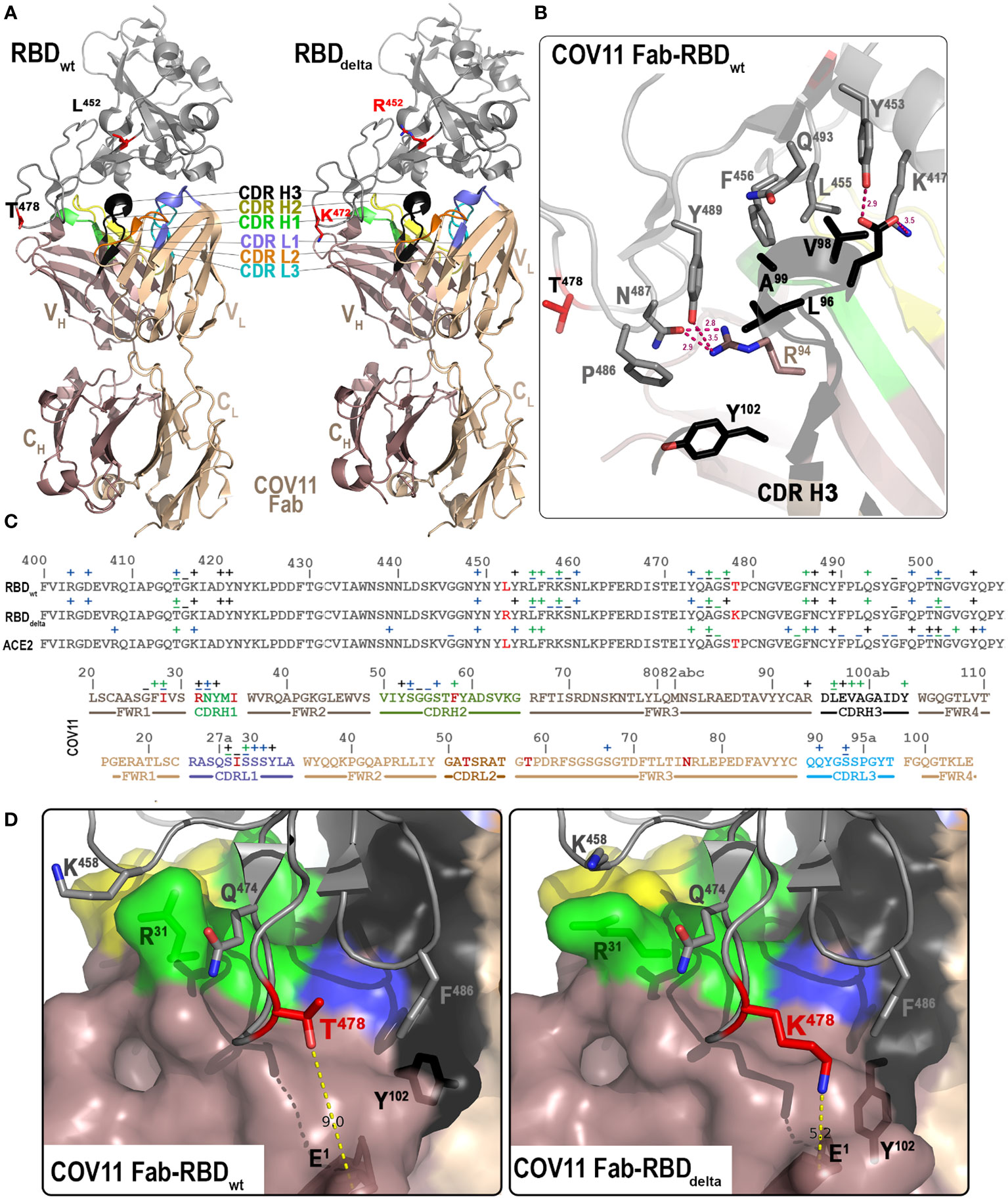
Figure 2 Crystal structures of the CoV11 Fab-RBD complexes. (A) Overall structure of the complex of CoV11 Fab-RBDwt (left) and CoV11 Fab-RBDdelta (right) are shown as a ribbon diagram. The complementarity-determining regions (CDRs) of CoV11 Fab are colored as follows: CDR H1 is green, CDR H2 is yellow, CDR H3 is black, CDR L1 is light blue, CDR L2 is orange, and CDR L3 is cyan. Residues T/K 478 and L/R452 mutated in delta as compared to wt are shown as sticks and colored red. (B) Close-up view into the interaction network mediated by the CoV11 CDR H3. Complex interface residues are shown as sticks and H-bonds are shown as magenta dashed lines. (C) Binding footprints of CoV11 and ACE2 on the RBD and the RBD on the CoV11 heavy and light chains. Contact residues defined by a 5-Å cutoff are marked above the sequence with (+) for side chain and (−) for main chain to indicate the type of contact. Contact types are colored as follows: hydrophilic (blue), hydrophobic (green), and both (black). Residues that differ from the VH 3-53 or VK 3-20 germline sequences on the heavy and light chains respectively are colored red. CDRs are colored as in (A, D) Interaction network around T/K 478. CoV11 and RBD residues in which the conformation differs between CoV11 Fab-RBDwt (left) and CoV11 Fab-RBDdelta (right) complexes are shown as sticks. The distance between T/K 478 of the RBD and N-terminal E1 of CoV11 heavy chain are shown with yellow dashed lines. A molecular surface is displayed over the Fab and the RBD is shown as a ribbon diagram.
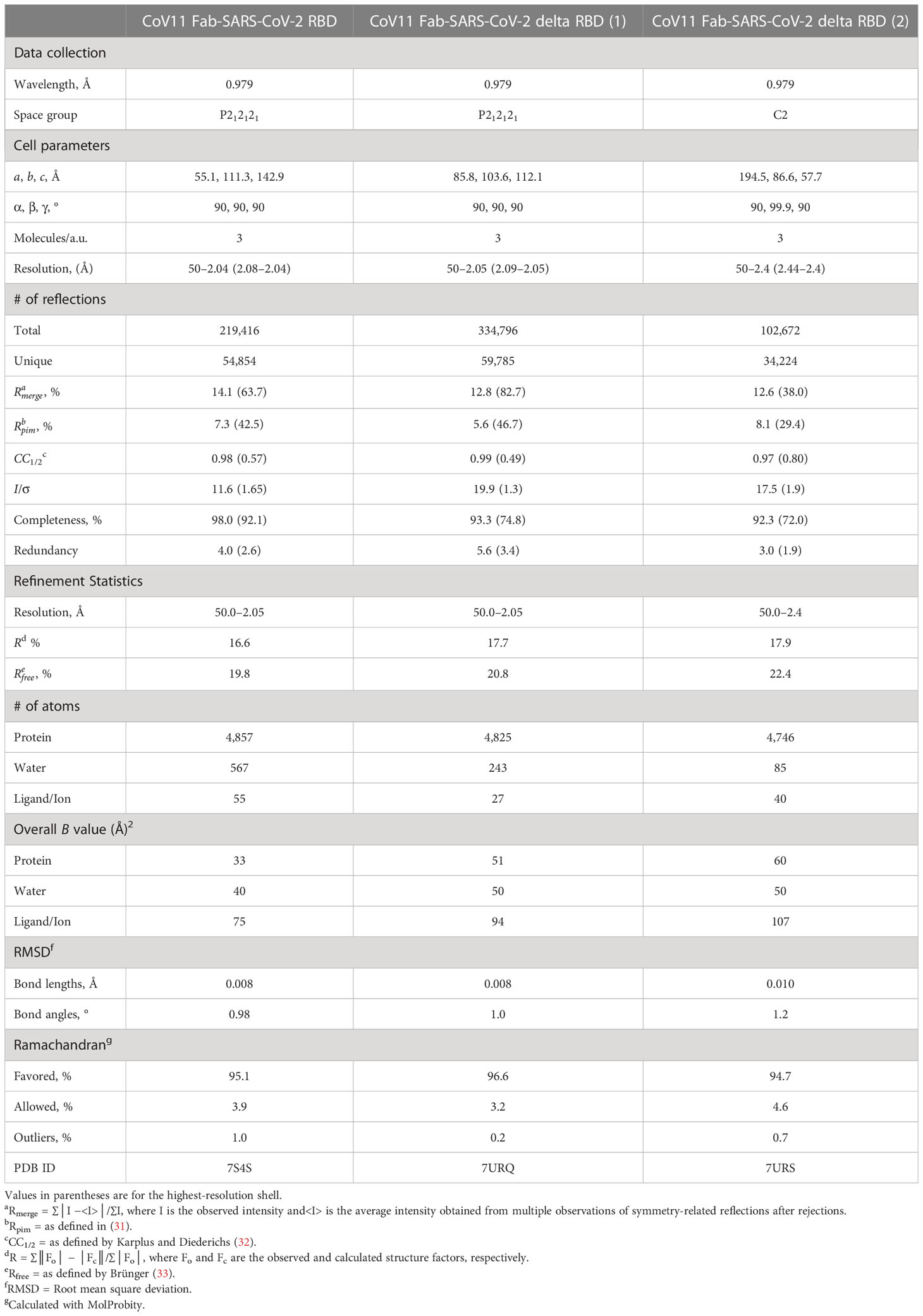
Table 2 Data collection and refinement statistics.
CoV11 binds the RBD in what has become the characteristic mode of binding for VH 3-53 encoded SARS-CoV-2 neutralizing antibodies by closely overlapping the ACE2 binding site on the RBD (Figure 2A). Binding to the RBD is achieved mainly through CDRs H1, H2, H3, and L1 [19.7%, 21.0%, 20.3%, and 19.9% of the total buried surface area (BSA) of CoV11, respectively] with small contributions from framework residues in the heavy chain (12.4%) and CDR L3 (4.9%); light chain framework residues (1.5%) and CDR L2 (0.3%) make almost no contribution to the total CoV11 BSA (Table S1). The bulk of these residues are encoded by the two CoV11 germline genes (IGHV3-53*01 and IGKV3-20*01), i.e., residues outside of the CDR H3, and account for the germline preference associated with this mode of binding to the RBD.
Most of the heavy chain framework residues that contribute to binding occur in framework region 1 preceding CDR H1 (, , and —for clarity, antibody residues are referenced using Kabat numbering (34) in superscript and region in subscript and RBD and ACE2 residues are referenced using the residue number in superscript and the protein in subscript unless implicitly implied in the text) (Figures 2B, C). The carboxyl of makes a main chain hydrogen bond to the nitrogen of RBD residue and the side chains of and make hydrophobic van der Waals contacts to RDB residues and . also makes van der Waals contacts with .
CDR H1 residues , , and increase the number of contacts to RBD residues in this region. makes a main chain hydrogen bond to RBD residue and a side chain-mediated hydrogen bond to in addition to hydrophobic contacts with and . makes a side chain-mediated hydrogen bond to the main chain carbonyl of , and the side chain of makes a hydrogen bond to the main chain carbonyl of (Figures 2B,C).
CDR H2 contributions to the interface are mainly through the side chains of , , and and the main chains of and . makes a strong hydrogen bond to and weaker hydrogen bonds to and . makes a strong hydrogen bond to . The main chain nitrogen of makes a hydrogen bond to while the main chain carbonyl makes a water-mediated hydrogen bond to the main chain of and the main chain nitrogen of makes water-mediated hydrogen bonds with the side chains of and . makes side chain-mediated van der Waals contacts with and (Figure 2C).
CoV11’s CDR H3 interaction with the RBD begins with the arginine preceding the CDR H3, , which makes hydrogen bonds to RBD residues and and van der Waals contacts with (Figure 2B). CDR H3 packs against , , and , while makes hydrogen bonds to and . makes van der Waals contacts with , , and , and makes contacts with , , , and . Finally, at the end of the CDR H3 packs against . Residues from the center of the CDR H3 do not interact with the RBD. Aside from and , most of the CDR H3 interface interactions are hydrophobic (Figure 2C).
The bulk of CoV11’s interactions from the light chain reside in CDR L1 (Figure 2C). forms a hydrogen bond with and van der Waals contacts with and . The main chains of and are mainly involved in hydrophobic contacts with although also forms water-mediated hydrogen bonds with , , and . The remaining contacts in CDR L1 are hydrophilic. forms a water-mediated hydrogen bond with , forms a weak hydrogen bond with , and forms a hydrogen bond with .
CDR L2 does not make any contacts with the RBD, but in the framework region preceding CDR L3 makes a weak hydrogen bond to . All other interface residues reside in CDR L3. The carboxyl of forms a strong hydrogen bond with , the side chain of makes a hydrogen bond to and a water-mediated hydrogen bond to , and at the beginning of the CDR L3 also makes a weak hydrogen bond to (Figure 2C).
The delta SARS-CoV-2 variant (B.1.617.2), which was originally identified in India in late 2020, quickly became the most prevalent SARS-CoV-2 variant in the US and the world in 2021 (35, 36). CoV11 binds to the wild-type RBD with an affinity of 2.1 nM and to the delta RBD with an affinity of 2.5 nM largely due to a slight increase in the koff (kd) for the delta RBD (Table 1); the kon (ka) for both RBDs is roughly equivalent. Delta contained two mutations in the RBD, Leu452 to Arg and Thr478 to Lys. The CoV11 Fab-RBDdelta complex shows that although these mutations sit outside of CoV11’s epitope footprint, they do make small changes to the way CoV11 binds to the RBD. The most notable difference is in how the N-terminus of the Fab sits relative to position 478 in the RBD (Figures 2D, E). The glutamic acid at the N-terminus of CoV11 is approximately 9 Å from but 5 Å from . This is only partly due to the longer side chain on . The change in the position of CoV11’s N-terminus also influences how interacts with . In the wt RBD, and display aromatic π stacking interactions, but in the delta RBD, the closer N-terminus shifts such that it cannot interact with Phe486. The interaction between may partially compensate for this loss. The closer N-terminus also causes some downstream changes to the interface. forms a hydrogen bond to the main chain of in the wt RBD but makes stronger van der Waals interactions with the aliphatic portion of the side chain in the delta RBD. is also closer to in the wt RBD, while is closer to Gln498 in the delta RBD. Most other differences are minor, which is mirrored by the less than twofold reduction in binding affinity for the delta RBD as compared to wt.
There are four sequence changes in the heavy chain of CoV11 relative to the VH 3-53 germline sequence. In the first sequence change, the VH 3-53 germline sequence has Thr at position 28, while CoV11 has Ile (Figure S2). This -to-Ile change in CoV11 increases van der Waals contacts with and . The second sequence difference, a germline -to-Arg change in CDR H1 adds one hydrogen bond to the CoV11–RBD interface; the side chain of is too short to make a similar hydrogen bond (Figure 2C). The third sequence difference, a -to-Ile change in CoV11, is not involved in the CoV11–RBD interface and likely has little if any contribution to CoV11 affinity for the RBD. Finally, the fourth sequence change resides in CDR H2. A -to-Phe change in CoV11 relative to the germline sequence does not also seem to change the interface much except in that it switches the interface to use only the hydrophobic van der Waals interactions of instead of the both hydrophilic and hydrophobic interactions of . Thus, the increases to RBD affinity for CoV11 over the germline gene largely come from two changes in sequence, to Ile and to Arg.
Sequence changes from the germline gene in the light chain contribute even less to the interface. CoV11 utilizes a light chain with four changes in sequence relative to the VK 3-20 germline gene (Figure S2). The only residue that differs from the VK 3-20 germline sequence involved in the CoV11–RBD interface is in CDR L1 ( to Ile) (Figure 2C). , however, only interacts with the RBD via main chain residues, which suggests that the Val-to-Ile change makes little if any contribution to CoV11’s binding affinity. The three other changes in sequence relative to the VK 3-20 germline ( to Thr, to Thr, and to Asn) sit well outside the CoV11–RBD interface. This suggests that the use of an unmodified VK 3-20 sequence might result in an antibody with a similar if not identical binding affinity to the RBD. Thus, the light chain’s contribution to CoV11’s affinity to the RBD is largely the result of the choice of the light chain germline gene and not the somatic mutations within the gene.
CDR H3 residues are not encoded by the VH 3-53 germline sequence but instead arise from heavy chain joining and diversity genes. Anti-SARS-CoV-2 VH 3-53 encoded antibody CDR H3s can therefore differ significantly. The VH 3-53 germline sequence ends at position 94 (Figure S2). makes hydrogen bonds to RBD residues and and van der Waals contacts with (Figure 2B). Only one anti-SARS-CoV-2 VH 3-53 encoded neutralizing antibody with a similar binding mode to the RBD whose structure is available has a residue other than arginine at this position, BG1-22 (PDB ID 7M6F). BG1-22 is also the antibody with the longest CDR H3, 19 amino acids; the shorter side chain at position 94 may facilitate the accommodation of the longer CDR H3 by eliminating hydrogen bonds to or (Figure S2). Many anti-SARS-CoV-2 VH 3-53 encoded neutralizing antibodies also have a hydrophilic residue at position 97, usually Asp, Glu, Tyr, or Gln. CoV11 has . Only the longer side chain of Glu preferentially forms a hydrogen bond with over . This may help explain CoV11’s resistance to gamma (P.1), which has a Lys-to-Thr mutation and beta (B.1.351) and omicron (B.1.1.529.1 or BA.1), which have a Lys-to-Asn mutation.
The remaining interactions of CoV11’s CDR H3 with the RBD are mainly hydrophobic, , , , and . While many other VH 3-53 encoded anti-SARS-CoV-2 neutralizing antibodies also have hydrophobic residues at position 96, some use Asp or Glu to form a salt bridge with . A similar situation is seen for position 98, with many VH 3-53 encoded anti-SARS-CoV-2 neutralizing antibodies also encoding a hydrophobic amino acid. The exceptions are largely due to those antibodies that encode a residue that can form a hydrogen bond or salt bridge with . Position 99 is more universally hydrophobic. Exceptions are those of Tyr, Lys, or Ser that form hydrogen bonds or salt bridges with or , residues that are also mutated in some VOCs (Figures 3A, B). Position 102 at the end of CDR H3 is almost always Tyr, Val, or Ile; however, some antibodies in this class with longer CDR H3s use other CDR H3 residues to make similar contacts with the RBD. Those that do use position 102 make equivalent contacts with the RBD using Ile, Val, or Tyr; CoV11 uses . These CDR H3 differences influence antibody neutralization breadth with those minimizing contacts to , , and , residues mutated in VOCs, having greater neutralization breadth for current VOCs. Therefore, CoV11 is among the anti-SARS-CoV-2 VH 3-53 encoded antibodies with the greatest neutralization breadth for current VOCs.
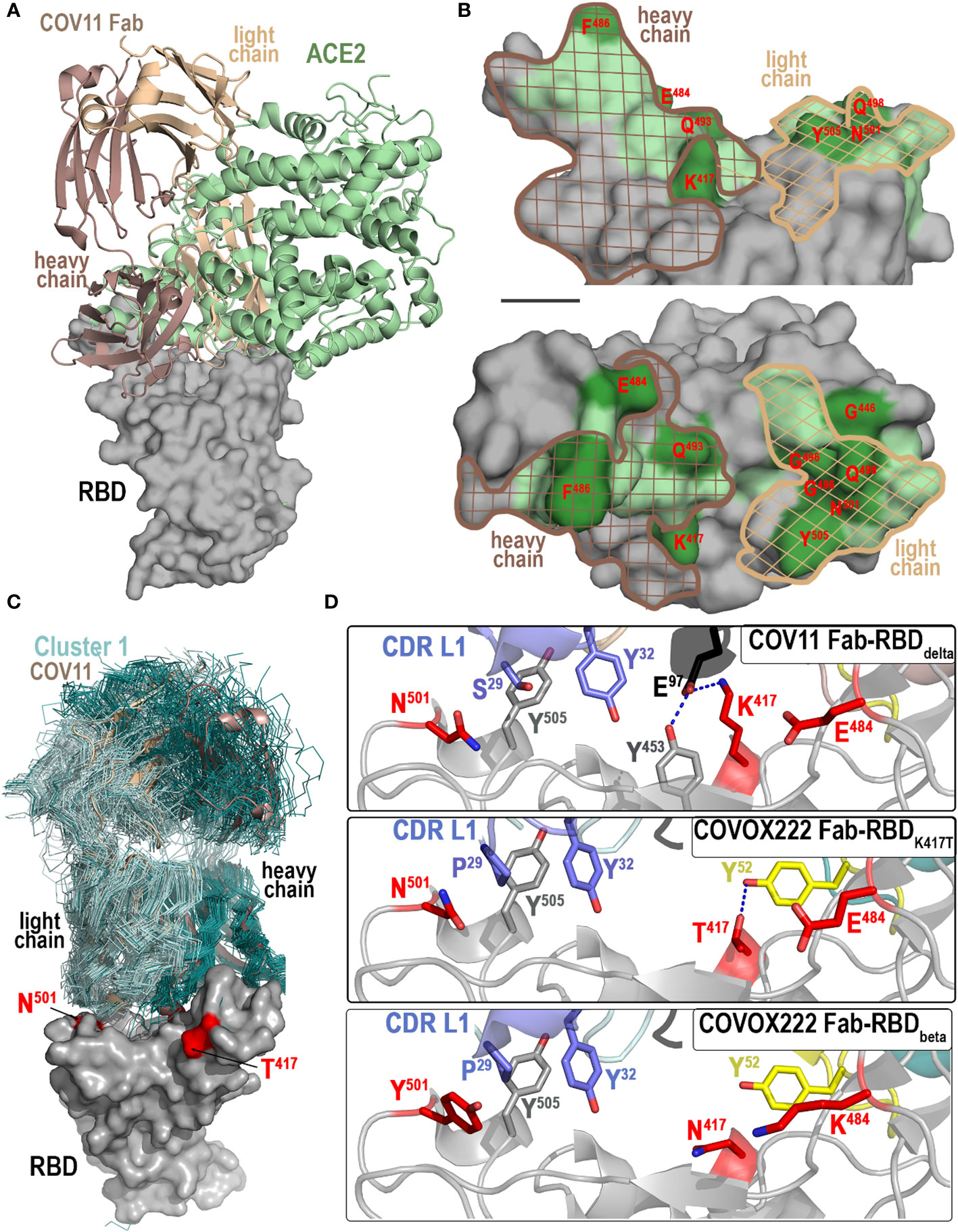
Figure 3 Molecular basis of CoV11’s interaction with the RBD. (A, B) Comparison of the binding modes of CoV11 and ACE2 to the receptor. (A) The structure of the ACE2-RBDwt complex (PBD ID: 6VW1) was overlaid over the CoV11 Fab-RBDwt complex (based on the RBD). Surface is displayed over the RBD molecule with CoV11 and ACE2 shown as cartoons. (B) Epitope footprint of heavy (dark brown mesh) and light (light brown mesh) chain of CoV11 is shown over the ACE2 footprint (green). RBD residues that mutate in VOCs are shown in darker green and labeled in red. (C, D) Recognition of RBD by CoV11 and RBD-specific Cluster 1 VH 3-53 or VH 3-66 germline-derived antibodies. (C) Available crystal structures of Cluster 1 antibodies in complex with the RBD of various SARS-CoV-2 variants (58 total structures) and the CoV11 Fab-RBDdelta complex were superimposed based on the RBD. Fabs of Cluster 1 VH 3-53 or VH 3-66 germline-derived antibodies are shown as ribbons (colored dark and light cyan for heavy and light chain, respectively) and the Fabs of CoV11 are shown as a cartoon (dark and light wheat for heavy and light chain, respectively). The RBD contact residues that mutate in the delta and beta strains are shown in red. (D) Close-up view of the interaction network mediated by CoV11 and COVOX222 (PDB IDs: 7NX8 and 7NXA) to RBD residues 417, 484, and 501 that mutate in VOCs. Complex interface residues are shown as sticks and H-bonds are shown as blue dashed lines.
The CoV11 and ACE2 binding sites on the RBD largely overlap, but there are residues unique to each interface and those that are used by both are not always used in the same way (Figures 3A, B). These differences set a limit on which residues can be changed by the virus and not disrupt receptor binding. CoV11 makes contact with 20 residues in common with ACE2 on the RBD (residues 415, 417, 449, 453, 455, 456, 473, 475–476, 486–487, 489–490, 493, 496, 498, 500–502, and 505) and 11 residues that are not in the ACE2 interface (residues 403, 405, 416, 420–421, 457–460, 474, and 477) (Figure 3C). ACE2, on the other hand, only utilizes nine residues that are absent from the CoV11 epitope (residues 408, 439, 446, 484–485, 492, 499, 503, and 506).
Eleven residues in the ACE2 binding site have changed in VOCs to date (currently omicron BA.5 and XBB1.5), seven of which are also used by CoV11 (Arg408 to Ser [omicron BA.2 and BA.4/5], Lys417 to Asn [beta and omicron variants] or Thr [gamma], Gly446 to Ser [omicron BA.3], Glu484 to Lys [beta and gamma] or Ala [omicron variants], Phe486 to Val [omicron BA.4/5] or Pro [omicron XBB1.5], Phe490 to Ser [omicron XBB1.5], Gln493 to Arg [omicron BA.1, BA.2, BA.3, and XBB1.5], Gly496 to Ser [omicron BA.1], Gln498 to Arg [omicron variants], Asn501 to Tyr [alpha, beta, gamma, and omicron variants], and Tyr505 to His [omicron variants]). In contrast, only three of the residues that are unique to the CoV11 epitope have changed (Asp405 to Asn [omicron BA.2, BA.3, BA.4/5, and XBB1.5], Asn460 to Lys [omicron XBB1.5], and Ser477 to Asn [omicron variants], shown combined in Figure 3B).
The first VOC mutation to occur in the RBD (Alpha, B.1.1.7), Asn501 to Tyr, increased the RBD’s affinity to ACE2 and recapitulated the enhanced infectivity seen in Alpha relative to the original SARS-CoV-2 strain (37); Tyr potentially preserves the polar interactions of Asn (), but strengthens the hydrophobic interactions with the aliphatic portion of other ACE2 interacting residues ( and ). Three other VOC mutations (those at Gly446, Phe490, and Gly496) are predicted to have little or no effect on ACE2 affinity since they are interface contacts solely made to main chain RBD atoms. Most other VOC mutations change ACE2’s affinity to the RBD. Mutations in VOCs that have appeared more recently are more disruptive of the ACE2-RBD interface, suggesting that their function may be geared more towards immune evasion. The Arg408-to-Ser mutation in omicron BA.2 and BA.4/5 potentially removes a hydrogen bond between the RBD and the glycan linked to . The Lys417 mutation to Thr or Asn in gamma or beta and omicron VOCs respectively removes a salt bridge between Lys417 and . Glu484 to Lys or Ala in beta or omicron VOCs removes a salt bridge between Glu484 and . Phe486 to Val or Pro (omicron BA.4/5 or XBB1.5) changes the hydrophobic packing between Phe486 and , , and . Gln493 to Arg (omicron BA.1–3 and XBB1.5) potentially removes a hydrogen bond to but maintains one to . Finally, Tyr505 to His in omicron VOCs potentially maintains a hydrogen bond to but removes ones to and . The observation that some of these mutations have reverted back to wt, e.g., Gln493 to Arg in omicrons BA.1–3 has gone back to Gln in BA.4/5, suggests that the virus is paying a price for these changes. It also suggests that these changes are less drastic than they could be if antibody evasion was their only criteria for incorporation into the RBD. Some of these changes in the RBD may have been in response to antibodies like CoV11.
While it is impossible to predict which changes from the germline sequence are the most important for increasing antibody neutralization breadth, the structure offers some insight. The first is that the heavy chain VH 3-53 and VH 3-66 germline sequences are essentially equivalent for this mode of binding to the RBD. The VH 3-66 germline sequence differs from the VH 3-53 sequence in one position, 12 (Figure S2). in VH 3-66 and in VH 3-53 sit well outside of the epitope footprint. This essentially doubles the number of naïve germline sequences that can serve as starting points for RBD binding. The second is that two changes relative to the heavy chain germline sequence are able to significantly increase CoV11’s affinity to the RBD, to Ile and to Arg. Twelve other anti-SARS-CoV-2 VH 3-53 or VH 3-66 antibodies of this type have a similar mutation at position 28 (13 total of 58 sequences or 22%), and six others have a similar mutation at position 31 (7 total of 58 sequences or 12%) (Figures 3D, S2). Approximately 50% that do not have the -to-Ile mutation have a mutation of the adjacent to a smaller hydrophobic residue, Val, Ile, or Leu, which may imply a similar function and a common mechanism of increasing RBD affinity. In contrast, only one sequence has a change from to anything other than Arg (2%), suggesting that this is a less common means of increasing RBD affinity (14% combined). The only other CoV11 change from the heavy chain germline sequence is the conservative mutation of to Phe, which is present in 28 (48%) of the other heavy chain germline sequences. Only two are mutated to any other residue (Asp, 3%), suggesting that this residue is important for binding the RBD even though its mutation from the germline might not provide much benefit. A large percentage of mutations in the germline sequence occur outside of the epitope footprint. For CoV11, this only amounts to one of the four heavy chain mutations, but for some of the other examples, it can be more than half of the mutations present, which suggests that the process of affinity maturation also introduces many functionally silent mutations.
Sequences outside of the VH 3-53 germline sequence also contribute to neutralization breadth. CoV11 has at position 97 (Figure 2B). Only the longer side chain of Glu preferentially forms a hydrogen bond with over . ’s mutation to Thr or Asn in gamma, beta, and omicron RBDs supports the incorporation of this preference in the CDR H3. Only three other antibodies with structures available of this type have a similar , EH3, COVA2-04, and BG4-25. Most other antibodies of this class are less tolerant of mutations at . The remaining CDR H3 residues of CoV11 used to interact with the RBD are mainly hydrophobic, , , , and . While hydrogen bonds and/or salt bridges contribute to CDR H3 affinity for other antibodies of this type, they generally do so to residues that have mutated in VOCs. The preferential use of hydrophobic van der Waals interactions in the CDR H3 may be a contributing factor to neutralization breadth.
CoV11’s light chain has four mutations relative to its VK 3-20 germline sequence. However, none of these residues is directly involved in the RBD binding interface; only the main chain atoms of residue 28 are used in the CoV11 Fab-RBD interface, making the -to-Ile change of little impact with respect to binding affinity. Also, unlike the case for the heavy chain, the VK 3-20 germline sequence contains the full light chain epitope footprint. This implies that a light chain with an unmutated VK 3-20 germline sequence would serve CoV11 as well as the one it has. Supporting this observation, of the 15 antibodies with VK 3-20 germline sequences paired to a VH 3-53 or VH 3-66 heavy chain, 5 (33%) have unmutated VK 3-20 germline sequences.
There are 58 structures of complexes of VH 3-53 or VH 3-66 heavy chain antibodies in complex with the RBD that bind like CoV11 (Figure 3D and S3). Fifteen of these are paired to a VK 3-20 germline-derived light chain. The most commonly paired light chain gene for these antibodies is VK 1-9 (17 antibodies), followed by VK 3-20 (15), VK 1D-33 (9), and VK 1-39 (6). There are four other kappa light chain germline gene examples with only one example of each, VK 1-12, VK 1-5, VK 1S8, and VK 4-1. Examples with lambda light chain germline genes are rarer with only seven total in the dataset. Two have two examples each, VL 1-40 and VL 3-21. The remaining genes have only one example each, VL 1-9, VL 2-8, and VL 1-44. While these germline genes represent a variety of sequences, the portions that interact with the RBD are surprisingly similar.
As shown in Figure 3D, CoV11 uses the side chain of to form a hydrogen bond with the main chain of . All other antibodies using the VK 3-20 germline sequence share this feature with two exceptions that have deletions of residue 27A (Figure S3). The two VK 3-20 germline exceptions and all but one other antibody with kappa light chains make a similar hydrogen bond when possible (approximately 40% are and cannot) with or . Lambda light chains also tend to shift this interaction to position 28 even though most have longer CDR L1s. in CoV11 and most other VK 3-20 germline sequence light chains pack against . However two VK 3-20 germline sequences have mutated to , BG4-25 and COVOX222 (Figure 3D and S3). Pro at position 29 also packs well against the mutation seen in every VOC since alpha (Figure 2D), which may indicate that these antibodies were elicited against alpha and not wt RBD. Most other kappa light chain antibodies use to pack against , but, possibly because of a shorter CDR L1, form stronger hydrogen bonds between the often common Ser at position 30 and , making the mutation more disruptive. Tyr at position 32 near the end of CDR L1, or Phe or Trp in a few cases, is then used to pack against , ending CDR L1’s interaction with the RBD.
CDR L2 makes little or no contribution to the interface for almost all of these paired light chains, independent of type (Figures 2A, C, Table S1). CDR L3’s interaction with the RBD with the exception of lambda and VK 1-39 light chains usually begins with a hydrogen bond from residue 90 to using Gln with the occasional His, Glu, or Arg. and have the advantage in that they can still form a hydrogen bond to . The exceptions to this rule are mostly the lambda and VK 1-39 light chains, which shift their interaction with the RBD further along their CDR L3s. The remaining contacts in CoV11 are mediated by and . With the exception of COVOX222, which has and , these residues are invariant in VK 3-20 light chains. The residues and sequences used by other light chains are more diverse with fewer common features. Most kappa light chains other than those derived from the VK 3-20 germline gene seem to focus part of their CDR L3 to forming a hydrogen bond or salt bridge with , making them more susceptible to a mutation there. This is less common for the lambda light chains.
In conclusion, the SARS-CoV-2 spike protein has changed over the course of the pandemic to increase infectivity and to evade immune pressure. This has led to mutations in the spike protein that have diminished the neutralization potency of a large number of antibodies elicited either by natural infection or by vaccination. Some of the most potent of these antibodies directly compete with ACE2 in binding the RBD (38–40). The low level of somatic mutation in one group of these antibodies enabled their early identification in B-cell sequences of infected individuals (11). This group of antibodies derived from the heavy chain VH 3-53 or VH 3-66 germline sequences with generally short CDR H3s were later found to almost directly overlap the ACE2 binding site on the RBD, providing a clear explanation for their mode of neutralization of the virus (11, 13–19).
Mutations in the virus in viral variants have affected antibodies of this class as they have done to all SARS-CoV-2 neutralizing antibodies. The first widely spread mutation that out-competed the original strain, to Tyr in the alpha variant (37), mostly affected the light chain of this group of antibodies and many were able to accommodate this change with little change to receptor affinity. Other more recent mutations in VOCs that greatly affected other antibody classes (22), e.g., the to Lys or Ala in the beta, gamma, and omicron variants, have also had little effect on receptor affinity for this group of antibodies. One of the most detrimental for this type of antibodies has been changes from to Thr or Asn in the gamma, beta, and omicron variants. CoV11 is an example of an antibody of this class that depends less strongly on a salt bridge or hydrogen bond to due to unique characteristics of its heavy chain CDR H3. While CoV11’s affinity to RBDs containing mutations of is lower, the fact that both Fabs of a CoV11 IgG can interact with a single spike protein partially counteracts this loss in affinity in neutralization.
The majority of residues that make up CoV11’s epitope footprint are shared with ACE2 in binding the RBD. This ensures that these residues cannot change without also affecting the RBD’s affinity to ACE2 and places a constraint on mutations that can occur in VOCs. CoV11 can still neutralize omicron BA.1 with only a slightly higher IC50 than it does beta, the variant with the next highest IC50 in spite of the fact that omicron BA.1 incorporates many more mutations to the RBD. Small changes to CoV11, e.g., the -to-Pro change used by COVOX222’s light chain, could overcome some of these losses in affinity to VOCs. Additional changes in the most recent VOCs such as the Asn460-to-Lys change in omicron BQ.1.1 or the Phe486-to-Pro, Gln493-to-Arg, and Asn460-to-Lys changes in omicron XBB.1.5 are enough to reduce CoV11’s affinity to the point where it no longer effectively neutralizes the virus (Figures S2 and S3). SPR and BLI data indicate that much of this loss in affinity is due to increases in the off-rate (kd) rather than decreases in the on-rate (ka) of CoV11, which suggests that small changes in CoV11’s sequence could allow CoV11 to compensate. For example, the Phe486-to-Pro change in XBB.1.5 could be ameliorated with a Val2-to-Phe change in CoV11’s heavy chain; Val2 is one of CoV11’s main contact residues with Phe486 and mutating it to Phe restores the contact in XBB.1.5. Other changes to restore binding and increase neutralization breadth in Omicron variants are less straightforward to predict due to the sheer number of amino acid changes possible at each position. CoV11 was elicited early in the course of the pandemic. Continued rounds of somatic mutation and selection to VOCs could easily generate a more potent and broadly neutralizing version that is more adapted to current VOCs.
The high prevalence of VH 3-53 and VH 3-66 germline sequences in the naïve antibody pool has probably helped in the immune response to SARS-CoV-2 in the general population. However, antibody selection and maturation is a stochastic process and not all individuals will utilize these germline sequences in response to SARS-CoV-2. There are many equally good responses to the antigen in its initial incarnation. It is only during the course of the pandemic that some responses have been shown to be better at neutralizing multiple strains than others. Antibodies derived from the VH 3-53 and VH 3-66 germline sequences have fared relatively well to date mainly because they share a majority of residues in common with ACE2 in their epitope footprint. While it is not possible to predict how the virus will continue to change in the future, CoV11 represents a good starting point for the design of a broad neutralizing response to SARS-CoV-2. With some minor changes, it could be made to be an even more broad and potent neutralizing therapeutic antibody for current VOCs and possibly also for the next generation of VOCs.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found here: 7S4S, 7URQ, and 7URS (Protein data bank).
WT, MS, and MP conceptualized this study and designed the experiments. WT, YC, RM, EP, and SG produced, purified, and characterized the proteins. WT and MP solved and analyzed the crystal structures of complexes. YC performed SPR kinetics. YC and LS generated PsVs and performed neutralization assays. MB performed BLI kinetics, SD performed neutralization of omicron variants. MC generated the BQ.1.1 and XBB.1.5 spike-expressing plasmids. ZT and MS isolated the CoV11 antibody. WT, MS, AF, and MP analyzed data. MP, FG, AF, and MS funded the work. WT and MP wrote the manuscript and every author has read, edited, and approved the final manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by USUHS intramural funds to M.P., a Canadian Institutes of Health Research (CIHR) operating grant Pandemic and Health Emergencies Research/Project #177958 to A.F. and M.P., Merit Award # I01 BX005469-01 to M.M.S., and in part by funding to F.J.G. from the Intramural Research Program, National Institutes of Health, National Cancer Institute, Center for Cancer Research. Use of the Stanford Synchrotron Radiation Lightsource, SLAC National Accelerator Laboratory, is supported by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences under Contract No. DE-AC02-76SF00515. The SSRL Structural Molecular Biology Program is supported by the DOE Office of Biological and Environmental Research, and by the National Institutes of Health, National Institute of General Medical Sciences. This work was also supported by an Exceptional Fund COVID-19 from the Canada Foundation for Innovation (CFI) #41027 to A.F. A.F. is the recipient of Canada Research Chair on Retro-viral Entry no. RCHS0235 950-232424. M.B. is the recipient of a CIHR master’s scholarship award.
We thank BEI Resources for sharing the SARS-CoV-2-related reagents, including pseudotyped lentiviral kits (NR-53816 and NR-53817), recombinant SB.1.1.7 (NR-55311), SB.1.351 (NR-55311), SB.1526 (NR-55438), and SP.1 (NR-55307).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The views expressed in this manuscript are those of the authors and do not reflect the official policy or position of the Uniformed Services University, the U.S. Army, the Department of Defense, the National Institutes of Health, the Department of Health and Human Services or the U.S. Government, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1178355/full#supplementary-material
Supplementary Figure 1 | Representative SPR kinetic measurements of RBD antigens binding to immobilized CoV11. All measurement were performed with ~80-200 RU CoV11 IgG immobilized on a Protein A chip. Flow RBD antigens were injected at the indicated concentrations. The background-corrected sensorgrams (colored) were fitted with a 1:1 Langmuir model (grey). Kinetic constants are summarized in Table S2.
Supplementary Figure 2 | Representative BLI kinetic measurements of RBD antigens binding to immobilized CoV11. All measurement were performed with CoV11 IgG immobilized on a Protein A sensor tip. Sensor tips were exposed to RBD antigens at the indicated concentrations. The background-corrected sensorgrams (colored) were fitted with a 1:1 Langmuir model (grey). Kinetic constants are summarized in Table S3.
Supplementary Figure 3 | Neutralization activity of COV11 IgG against SARS-CoV-2 D614G (positive control) or Omicrons BQ.1.1 and XBB.1.5. Dose response neutralization curves of SARS-CoV-2 pseudotyped lentivirus with SAR2-CoV-2 S variants. hACE2 expressing 293T cells were infected with different variants of SARS-CoV-2 PsV or VSVG PsV (negative control) in the presence of CoV11 IgG. Infectivity was quantified by the cellular luciferase signal 48 hours post infection. Relative infectivity was normalized by the luciferase signal in infected cells without intervention (PBS saline).
Supplementary Figure 4 | Heavy chain sequence alignment of VH 3-53 or VH 3-66 IgGs that bind the RBD like CoV11. Antibodies are labeled by PDB ID underscore antibody name and residues are numbered using the Kabat antibody number scheme. Contact residues defined by a 5 Å cutoff are marked above the sequence with (+) for side chain and (-) for main chain to indicate the type of contact. Contact types are colored as follows: hydrophilic (blue), hydrophobic (green) and both (black). Residues that differ from the VH 3-53 or VH 3-66 germline sequence colored red. The one residue difference in the VH 3-66 germline sequence relative to the VH 3-53 germline sequence is colored blue. CDRs are colored as in .
Supplementary Figure 5 | Light chain sequence alignment of IgGs that bind the RBD like CoV11. Antibodies are labeled by PDB ID underscore antibody name and residues are numbered using the Kabat antibody number scheme. Contact residues defined by a 5 Å cutoff are marked above the sequence with (+) for side chain and (-) for main chain to indicate the type of contact. Contact types are colored as follows: hydrophilic (blue), hydrophobic (green) and both (black). Residues that differ from the VK 3-20 germline sequence colored red. Only the first 15 sequences utilize the VK 3-20 germline sequence. The last five use lambda germline sequences and the remaining sequences use kappa germline sequences other than VK 3-20. CDRs are colored as in .
Supplementary Table 1 | Details of the CoV11 Fab-RBDwt, CoV11 Fab-RDBdelta, C098 Fab-RBDwt (germline VH 3-53 and VK 3-20), and ACE2-RBDwt interfaces as calculated by the EBI PISA server (http://www.ebi.ac.uk/msd-srv/prot_int/cgi-bin/piserver).
Supplementary Table 2 | Surface Plasmon Resonance (SPR) kinetic constants of CoV11 binding to the RBD of SARS-CoV-2 wt and selected VOCs. CoV11 IgG was immobilized on Protein A chip and the RBDs of various SARS-CoV-2 strains were injected as flow analytes. The dissociation constants (KD), association constants (ka) and dissociation constants (kd) are as shown. KD values were determined using a 1:1 Langmuir model. Experimental binding curves are shown in Figure S1.
Supplementary Table 3 | Bio-Layer Interferometry (BLI) kinetic constants of CoV11 binding to the RBD of SARS-CoV-2 wt and selected VOCs. CoV11 IgG was immobilized on a Protein A sensor tip and then placed into solutions containing the RBDs of various SARS-CoV-2 strains at different concentrations. The dissociation constants (KD), association constants (ka) and dissociation constants (kd) are as shown. KD values were determined using a 1:1 Langmuir model. Values are the average of two or three experiments with standard deviations as shown. N.D., not detected. Experimental binding curves are shown in Figure S2.
1. Hu B, Guo H, Zhou P, Shi ZL. Characteristics of SARS-CoV-2 and COVID-19. Nat Rev Microbiol (2021) 19:141–54. doi: 10.1038/s41579-020-00459-7
2. Callaway E. Beyond omicron: what's next for COVID's viral evolution. Nature (2021) 600:204–7. doi: 10.1038/d41586-021-03619-8
3. Wrapp D, Wang N, Corbett KS, Goldsmith JA, Hsieh CL, Abiona O, et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science (2020) 367:1260–3. doi: 10.1126/science.abb2507
4. Walls AC, Park YJ, Tortorici MA, Wall A, McGuire AT, Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell (2020) 181:281–292 e6. doi: 10.1016/j.cell.2020.02.058
5. Shang J, Wan Y, Luo C, Ye G, Geng Q, Auerbach A, et al. Cell entry mechanisms of SARS-CoV-2. Proc Natl Acad Sci U.S.A. (2020) 117:11727–34. doi: 10.1073/pnas.2003138117
6. Lan J, Ge J, Yu J, Shan S, Zhou H, Fan S, et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature (2020) 581:215–20. doi: 10.1038/s41586-020-2180-5
7. Yan R, Wang R, Ju B, Yu J, Zhang Y, Liu N, et al. Structural basis for bivalent binding and inhibition of SARS-CoV-2 infection by human potent neutralizing antibodies. Cell Res (2021) 31:517–25. doi: 10.1038/s41422-021-00487-9
8. Shang J, Ye G, Shi K, Wan Y, Luo C, Aihara H, et al. Structural basis of receptor recognition by SARS-CoV-2. Nature (2020) 581:221–4. doi: 10.1038/s41586-020-2179-y
9. Benton DJ, Wrobel AG, Roustan C, Borg A, Xu P, Martin SR, et al. The effect of the D614G substitution on the structure of the spike glycoprotein of SARS-CoV-2. Proc Natl Acad Sci U.S.A. (2021) 118:e2022586118. doi: 10.1073/pnas.2022586118
10. Zhang L, Jackson CB, Mou H, Ojha A, Peng H, Quinlan BD, et al. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat Commun (2020) 11:6013. doi: 10.1038/s41467-020-19808-4
11. Yuan M, Liu H, Wu NC, Lee CD, Zhu X, Zhao F, et al. Structural basis of a shared antibody response to SARS-CoV-2. Science (2020) 369:1119–23. doi: 10.1126/science.abd2321
12. Wu NC, Yuan M, Liu H, Lee CD, Zhu X, Bangaru S, et al. An alternative binding mode of IGHV3-53 antibodies to the SARS-CoV-2 receptor binding domain. Cell Rep (2020) 33:108274. doi: 10.1016/j.celrep.2020.108274
13. Hurlburt NK, Seydoux E, Wan YH, Edara VV, Stuart AB, Feng J, et al. Structural basis for potent neutralization of SARS-CoV-2 and role of antibody affinity maturation. Nat Commun (2020) 11:5413. doi: 10.1038/s41467-020-19231-9
14. Barnes CO, Jette CA, Abernathy ME, Dam KA, Esswein SR, Gristick HB, et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature (2020) 588:682–7. doi: 10.1038/s41586-020-2852-1
15. Zhang Q, Ju B, Ge J, Chan JF, Cheng L, Wang R, et al. Potent and protective IGHV3-53/3-66 public antibodies and their shared escape mutant on the spike of SARS-CoV-2. Nat Commun (2021) 12:4210. doi: 10.1038/s41467-021-24514-w
16. Muecksch F, Weisblum Y, Barnes CO, Schmidt F, Schaefer-Babajew D, Wang Z, et al. Affinity maturation of SARS-CoV-2 neutralizing antibodies confers potency, breadth, and resilience to viral escape mutations. Immunity (2021) 54:1853–1868 e7. doi: 10.1016/j.immuni.2021.07.008
17. Supasa P, Zhou D, Dejnirattisai W, Liu C, Mentzer AJ, Ginn HM, et al. Reduced neutralization of SARS-CoV-2 B.1.1.7 variant by convalescent and vaccine sera. Cell (2021) 184:2201–2211 e7. doi: 10.1016/j.cell.2021.02.033
18. Scheid JF, Barnes CO, Eraslan B, Hudak A, Keeffe JR, Cosimi LA, et al. B cell genomics behind cross-neutralization of SARS-CoV-2 variants and SARS-CoV. Cell (2021) 184:3205–3221 e24. doi: 10.1016/j.cell.2021.04.032
19. Dejnirattisai W, Zhou D, Supasa P, Liu C, Mentzer AJ, Ginn HM, et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell (2021) 184:2939–2954 e9. doi: 10.1016/j.cell.2021.03.055
20. Li W, Chen Y, Prevost J, Ullah I, Lu M, Gong SY, et al. Structural basis and mode of action for two broadly neutralizing antibodies against SARS-CoV-2 emerging variants of concern. Cell Rep (2022) 38:110210. doi: 10.1016/j.celrep.2021.110210
21. Yan Q, He P, Huang X, Luo K, Zhang Y, Yi H, et al. Germline IGHV3-53-encoded RBD-targeting neutralizing antibodies are commonly present in the antibody repertoires of COVID-19 patients. Emerg Microbes Infect (2021) 10:1097–111. doi: 10.1080/22221751.2021.1925594
22. Hastie KM, Li H, Bedinger D, Schendel SL, Dennison SM, Li K, et al. Defining variant-resistant epitopes targeted by SARS-CoV-2 antibodies: a global consortium study. Science (2021) 374:472–8. doi: 10.1126/science.abh2315
23. Tauzin A, Nicolas A, Ding S, Benlarbi M, Medjahed H, Chatterjee D, et al. Spike recognition and neutralization of SARS-CoV-2 omicron subvariants elicited after the third dose of mRNA vaccine. Cell Rep (2023) 42:111998. doi: 10.1016/j.celrep.2023.111998
24. Tauzin A, Benlarbi M, Medjahed H, Gregoire Y, Perreault J, Gendron-Lepage G, et al. Humoral responses against BQ.1.1 elicited after breakthrough infection and SARS-CoV-2 mRNA vaccination. Vaccines (Basel) (2023) 11:242. doi: 10.3390/vaccines11020242
25. Guenthoer J, Lilly M, Starr TN, Dadonaite B, Lovendahl KN, Croft JT, et al. Identification of broad, potent antibodies to functionally constrained regions of SARS-CoV-2 spike following a breakthrough infection. bioRxiv (2023) 29:2022.12.15.520606. doi: 10.1101/2022.12.15.520606
26. Ferrara F, Temperton N. Pseudotype neutralization assays: from laboratory bench to data analysis. Methods Protoc (2018) 1:8. doi: 10.3390/mps1010008
27. Chen Y, Prevost J, Ullah I, Romero H, Lisi V, Tolbert WD, et al. Molecular basis for antiviral activity of two pediatric neutralizing antibodies targeting SARS-CoV-2 spike RBD. iScience (2023) 26:105783. doi: 10.1016/j.isci.2022.105783
28. Otwinowski Z, Minor W, Carter CW Jr. Processing of X-ray diffraction data collected in oscillation mode. Meth Enzymol (1997) 276:307–26. doi: 10.1016/S0076-6879(97)76066-X
29. N. Collaborative Computational Project. The CCP4 suite: programs for protein crystallography. Acta Crystallogr D Biol Crystallogr (1994) 50:760–3. doi: 10.1107/S0907444994003112
30. Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N, et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D Biol Crystallogr D66 (2010 66:213–21. doi: 10.1107/S0907444909052925
31. Weiss MS. Global indicators of X-ray data quality. J Appl Cryst (2001) 34:130–5. doi: 10.1107/S0021889800018227
32. Karplus PA, Diederichs K. Linking crystallographic model and data quality. Science (2012) 336:1030–3. doi: 10.1126/science.1218231
33. Brunger AT. Free r value: cross-validation in crystallography, methods in enzymology. Academic Press (1997) p. 366–96.
35. Samieefar N, Rashedi R, Akhlaghdoust M, Mashhadi M, Darzi P, Rezaei N. Delta variant: the new challenge of COVID-19 pandemic, an overview of epidemiological, clinical, and immune characteristics. Acta BioMed (2022) 93:e2022179. doi: 10.23750/abm.v93i1.12210
36. Kirola L. Genetic emergence of B.1.617.2 in COVID-19. New Microbes New Infect (2021) 43:100929. doi: 10.1016/j.nmni.2021.100929
37. Liu Y, Liu J, Plante KS, Plante JA, Xie X, Zhang X, et al. The N501Y spike substitution enhances SARS-CoV-2 infection and transmission. Nature (2022) 602:294–9. doi: 10.1038/s41586-021-04245-0
38. Brouwer PJM, Caniels TG, van der Straten K, Snitselaar JL, Aldon Y, Bangaru S, et al. Potent neutralizing antibodies from COVID-19 patients define multiple targets of vulnerability. Science (2020) 369:643–50. doi: 10.1126/science.abc5902
39. Rogers TF, Zhao F, Huang D, Beutler N, Burns A, He WT, et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science (2020) 369:956–63. doi: 10.1126/science.abc7520
Keywords: SARS-CoV-2, neutralizing antibody, VH 3-53 germline gene (IGHV3-53*01), receptor binding domain (RBD), variants of concern (VOC)
Citation: Tolbert WD, Chen Y, Sun L, Benlarbi M, Ding S, Manickam R, Pangaro E, Nguyen DN, Gottumukkala S, Côté M, Gonzalez FJ, Finzi A, Tehrani ZR, Sajadi MM and Pazgier M (2023) The molecular basis of the neutralization breadth of the RBD-specific antibody CoV11. Front. Immunol. 14:1178355. doi: 10.3389/fimmu.2023.1178355
Received: 02 March 2023; Accepted: 16 May 2023;
Published: 02 June 2023.
Edited by:
Amol Suryawanshi, Auburn University, United StatesReviewed by:
Rahul K Suryawanshi, Gladstone Institutes, United StatesCopyright © 2023 Tolbert, Chen, Sun, Benlarbi, Ding, Manickam, Pangaro, Nguyen, Gottumukkala, Côté, Gonzalez, Finzi, Tehrani, Sajadi and Pazgier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: William D. Tolbert, d2lsbGlhbS50b2xiZXJ0LmN0ckB1c3Vocy5lZHU=; Marzena Pazgier, bWFyemVuYS5wYXpnaWVyQHVzdWhzLmVkdQ==
†Present address: Rohini Manickam, Department of Biochemistry, Molecular and Cellular Biology (MCB), Graduate Program of Uniformed Services University of the Health Sciences, Bethesda, MD, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.