
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 17 August 2023
Sec. Alloimmunity and Transplantation
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1154650
This article is part of the Research TopicRole of the Immune System in Renal Transplantation: Importance, Mechanism, and TherapyView all 12 articles
 Elena Cuadrado-Payán1,2
Elena Cuadrado-Payán1,2 María José Ramírez-Bajo2,3
María José Ramírez-Bajo2,3 Elisenda Bañón-Maneus2,3
Elisenda Bañón-Maneus2,3 Jordi Rovira2,3
Jordi Rovira2,3 Fritz Diekmann1,2,3
Fritz Diekmann1,2,3 Ignacio Revuelta1,2,3†
Ignacio Revuelta1,2,3† David Cucchiari1,2*†
David Cucchiari1,2*†Antibody-mediated rejection is the leading cause of kidney graft dysfunction. The process of diagnosing it requires the performance of an invasive biopsy and subsequent histological examination. Early and sensitive biomarkers of graft damage and alloimmunity are needed to identify graft injury and eventually limit the need for a kidney biopsy. Moreover, other scenarios such as delayed graft function or interstitial fibrosis and tubular atrophy face the same problem. In recent years, interest has grown around extracellular vesicles, specifically exosomes actively secreted by immune cells, which are intercellular communicators and have shown biological significance. This review presents their potential as biomarkers in kidney transplantation and alloimmunity.
In kidney transplantation, antibody-mediated rejection (ABMR) continues to represent the most significant challenge to be resolved in order to improve graft and patient survival (1, 2). Although acute ABRM is a potentially treatable disease, chronic ABMR has limited therapeutic options. It invariably progresses to end-stage chronic kidney disease (ESKD), representing over 50% of death-censored graft losses. Therefore, early detection of acute ABMR, timely treatment, and prevention of its progression to chronic ABMR are vital to guarantee satisfactory results for kidney transplant recipients, especially in the high immunological risk group. With this premise, some centers have developed a strategy based on protocol kidney graft biopsies. However, a biopsy is an invasive method with potential risk of associated complications; moreover, it presents a high financial and resource cost (1, 2).
For this reason, many transplant centers choose to perform biopsies only “by indication” when some classical parameters are altered, such as creatinine, proteinuria, or the existence of specific anti-Human Leukocyte Antigen (HLA) donor antibodies (DSA) (3). However, these indicators need more sensitivity and change when rejection is established (3). This unmet clinical need led to a quest to discover early and sensitive biomarkers of graft damage, limiting renal graft biopsies’ performance to only those patients with a high likelihood of rejection. Furthermore, beyond the rejection field, other scenarios in kidney transplantation, such as delayed graft function (DGF) or interstitial fibrosis and tubular atrophy (IFTA), run into the same problem without biomarkers that allow for early detection or differentiation from other pathologies.
Among these possible biomarkers, it is worth highlighting circulating extracellular vesicles (EVs), specifically, exosomes actively secreted by immune cells, which are intercellular communicators that carry microRNA, DNA, and proteins with biological significance as intercellular mediators (4).
This review summarizes the current EVs literature in kidney transplantation and their use as biomarkers.
The discovery of EVs dates to the last century. Since then, several names have been attributed to them, and a sharp increase in scientific publications has been evidenced in the last decade (5). In 2014, the Minimal Information for Studies of Extracellular Vesicles (“MISEV”) guidelines were released by the International Society for Extracellular Vesicles (ISEV). Subsequently, an update was proposed in 2018 by consensus of the largest group of EV experts, defining EVs as “the generic term for particles naturally released by cells that are delimited by a lipid bilayer and cannot replicate” (6). One year later, in 2019, another publication released by the corresponding authors of the MISEV guidelines asserted the accuracy and clarity of EV nomenclature to specialists and non-specialists and their use as a scaffold for progressively more detailed designation (7).
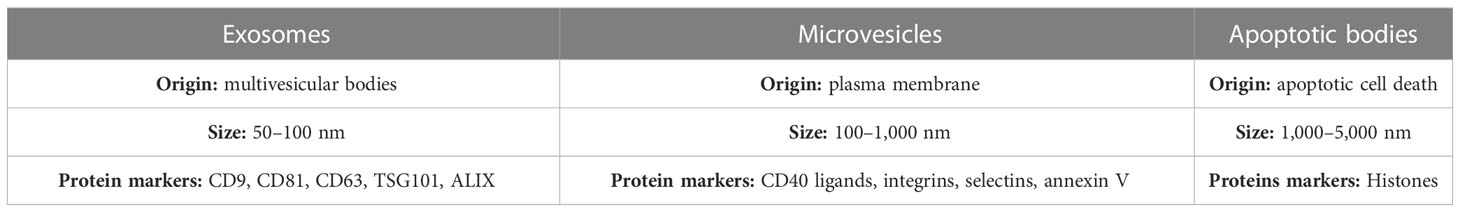
EVs are classified into three categories: exosomes, which are intraluminal vesicles contained in multivesicular bodies (MVBs) and released into the extracellular environment upon fusion of MVBs with the plasma membrane, microvesicles (also called microparticles) budded from the plasma membrane, and apoptotic bodies, the largest of the known vesicles and released during programmed cell death when the plasma membrane blebs (8–12) (Table 1).
Due to the internal origin of exosomes from MVBs, they represent the parental cell’s internal activity and conditional state more closely than other types of EVs (5, 10, 11, 16, 17).
EVs seems that can be secreted by any cell type studied, including immune cells, are believed to play a central role in cell-to-cell communication (5, 6, 11). Their content is diverse and includes protein, lipids, nucleic acids, and other bioactive molecules that are determined according to the type of cell from which they arise (11, 16, 18). These content molecules provide EVs with different capabilities in terms of biogenesis and transport. Moreover, membrane curvature, which begins in the parent cell during membrane budding, determines the shape, composition, and size of each EV and therefore has a role in their physiological function (19–22).
Surface-exposed components and ligands determine EVs’ biodistribution and their binding to target cells or binding to the extracellular matrix, allowing intracellular signaling pathways via simple interaction with the surface of the target cells or by internalization. For instance, proteins such as tetraspanins (CD81, CD82, CD63, or CD9) help penetrate exosomes inside cells, invasion, and fusion, whereas heat shock proteins such as HSP70 and HSP90 are involved in antigen binding and presentation. Other proteins such as Alix, annexins, Rab, or TSG101 are associated with exosome release, membrane transport, and function. Notably, some of these proteins, such as CD63, CD81, HSP70, and TSG101, which are enriched explicitly in exosomes, are generally used as their marker proteins (13, 17, 18)
Lipids such as cholesterol and sphingomyelin enrich EV membranes and, as well as their essential structural role, can also be transferred between cells (23).
The parent cell source and the properties of target cells determine EVs’ biodistribution, and their quantity in circulation is determined by the balance between production and clearance. Clearance occurs via interactions with target cells through endocytosis, phagocytosis, pinocytosis, or membrane fusion (14), and also through the liver, spleen, and lungs (24, 25). Regarding the half-life, different studies have described a predominantly short one, ranging from 2–5 min to 5 h (24–27).
Up to now, there is no unique standardized protocol for EV isolation (28), and obtaining highly pure EVs is necessary to attribute them a specific function or property to be used as biomarkers (6). Their isolation and purification are decisive for most downstream applications due to their overlap with lipoproteins or protein aggregates; these can easily mistake EV detection due to their similar biophysical properties and act as contaminants (5, 15, 28).
EVs can be isolated from many sources, including biological fluids and cell culture supernatants (29, 30). Initial publications on blood-derived EVs prompted the recommendation to preferably conduct plasma studies due to platelet-derived EV released in the serum after blood collection during clot formation (30). In contrast, other studies have found EV isolation from serum to be more reproducible (29) and, in kidney transplantation, the serum content could reflect renal and endothelial recovery functions (31, 32)
According to MISEV 2018, differential ultracentrifugation was the most common isolation technique (6). It consists of consecutive centrifugation steps, each with an increase in centrifugation forces and durations, aiming to isolate smaller from larger particles based on differences in their densities (33). Other procedures, such as size exclusion chromatography (SEC), ultrafiltration, precipitation, or immunoisolation, were used by approximately 5–20% of researchers (6).
Since then, an assortment of techniques has been developed, such as field-flow fractionation (FFF), variations of size exclusion chromatography (SEC), ion exchange chromatography, microfiltration, fluorescence-activated sorting, novel immunoisolation techniques, and microfluidics or precipitation techniques using a plethora of commercial kits (5, 6).
Further information on EV isolation is beyond the scope of this review. All these methods, along with their strengths and weaknesses, are extensively discussed elsewhere (34, 35).
EVs are involved in the regulation of physiological functions such as maintenance of homeostasis, enhanced coagulation (36–38), vessel integrity (39), tissue repair (40), or synaptic plasticity (41). They are also involved in inflammation, angiogenesis, or transmission of oncogenic molecules to neighboring cells, favoring neoplasia propagation and procoagulant capacity (4, 10, 11, 42–44).
Regarding their role in immunology, EVs act in innate and adaptative immune systems. In the innate, their major role has been described as pro-inflammatory mediators secreted by activated macrophages, neutrophilic granulocytes, NK cells, or mast cells in scenarios such as infection (45–47), sepsis (47–49), or chronic inflammation (50). In addition, an anti-inflammatory role also has been described through TGF-β secretion or regulation of endogenous lipid mediators (51).
Regarding adaptive immunity, EVs are a source of antigens for antigen-presenting cells (APC) such as macrophages, dendritic cells (DCs), and B cells. Depending on their cargo and parenting cells, they can induce immunogenic or tolerogenic responses (8, 42, 52). Recipient APCs can release EVs containing peptide-MHC I or II complexes and co-stimulatory molecules that contribute to antigen presentation (53, 54). This release is carried out constitutively, although this process can be increased after stimulation (55). Furthermore, graft-derived exosomes can initiate the immune response in a direct or semi-direct pathway that will end up causing graft rejection (56, 57). The direct pathway consists of exosomes from donor tissue behaving as donor APCs presenting MCH molecules or tissue-specific self-antigens to recipient T cells (58, 59). In contrast, in the semi-direct pathway, graft exosomes are taken up by recipient APCs, presenting intact MHC molecules from these graft exosomes on the surface of APCs, known as MHC cross-dressing (60).
Besides promoting intercellular information exchange via their surface molecules, their role as carriers of soluble mediators such as cytokines has been described. These cytokines include interleukin 1β (IL-1 β), interleukin 1α (IL-1 α), interleukin 18 (IL-18), macrophage migration inhibitory factor (MIF), interleukin 32, membrane-bound tumor necrosis factor (TNF), interleukin 6 (IL-6), vascular endothelial growth factor (VEGF), interleukin 8 (CXCL8), fractalkine (CX3CL1), CCL2, CCL3, CCL4, CCL5, and CCL20, and transforming growth factor β (TGF β) (27, 61).
Nowadays, biomarkers of the different EVs in circulation have been assessed in plenty of publications. The most developed field is oncology, where tumor mass has been linked to the amount of tumor-derived circulating exosomes. In the field of kidney disease, some studies demonstrate the participation of exosomes in different processes, which include acute kidney failure, autoimmune kidney disease, diabetic kidney disease, glomerulonephritis, vasculitis, or thrombotic microangiopathies (16).
Regarding transplantation, EVs in body fluids have been proposed as liquid biopsies. Mainly, publications focus on heart, lung, or pancreatic islet allografts. A profile of blood-derived EVs through multiplex flow cytometric assay using antibody-coated capture beads has been described in heart transplant recipients. A significant increase in the concentration of plasma-derived EVs in patients undergoing both acute cell and ABMR has been confirmed compared with subjects not undergoing them (62). In the lung, circulating exosomes with lung self-antigens can be a viable non-invasive biomarker for identifying patients at risk of developing chronic lung allograft dysfunction (63–65). Regarding pancreatic islet allografts, a human-into-mouse xenogeneic islet transplant model led to a marked decrease in the transplant islet exosome signal in early rejection, and changes in exosomal microRNA and proteomic profiles preceded hyperglycemia (66).
In kidney transplant, a decrease of circulating microparticle levels and their procoagulant activity after graft has been described in comparison to the prior hemodialysis status, hypothesizing that microparticles could be associated with cardiovascular risk improvement after transplant (67, 68). Studies have also been carried out on the urine and plasma of recipients, revealing their potential use as markers of cellular or humoral rejection, DGF, IFTA, mediated drug toxicity, and other non-specific graft injuries (18). Below, we expand the role of EVs in all of these settings (Figure 1). The methods of isolation for each study are summarized in Table 2.
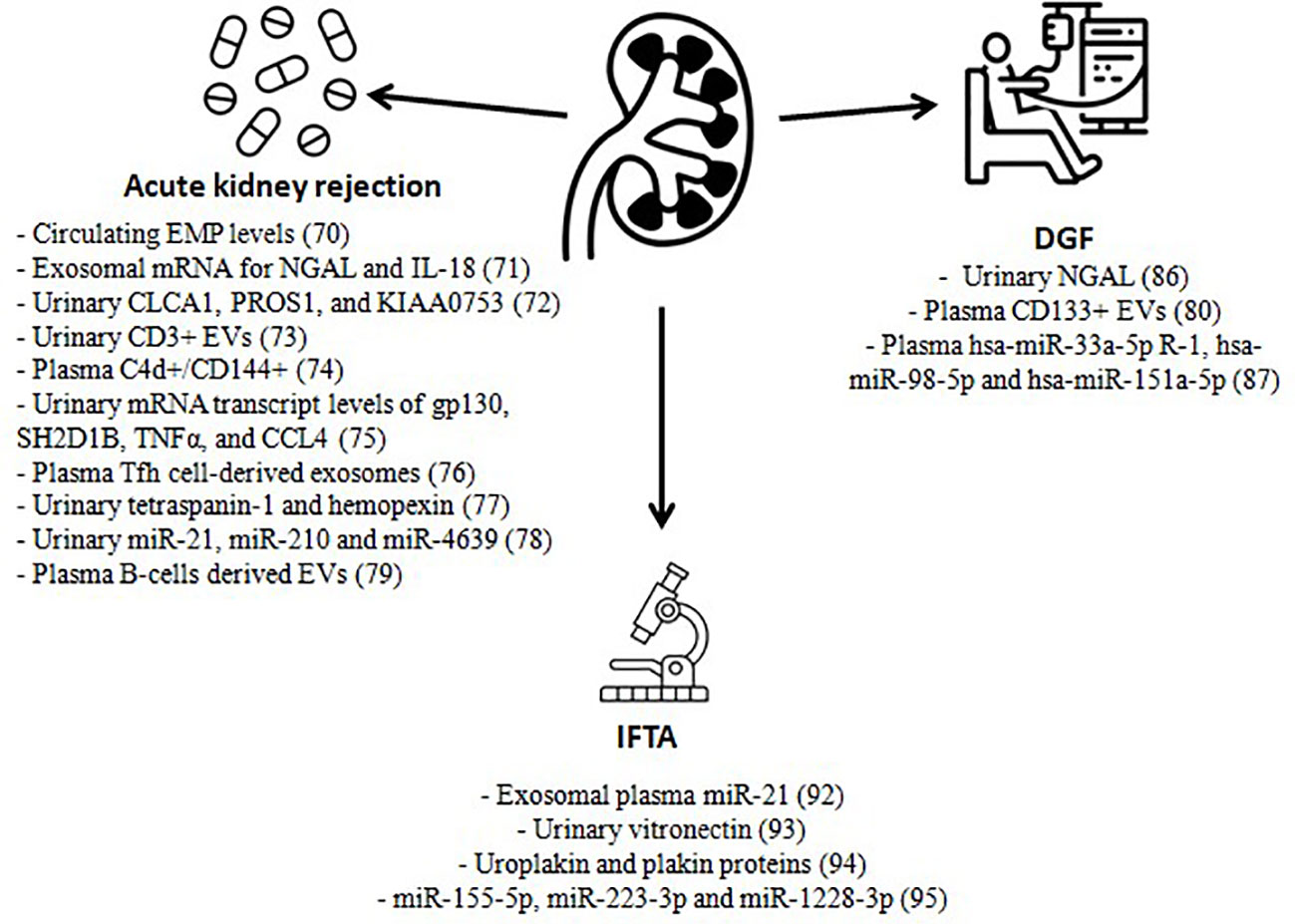
Figure 1 Extracellular vesicles as biomarkers in kidney transplantation. EMP, endothelial microparticles; NGAL, neutrophil gelatinase-associated lipocalin; IL-18, interleukin 18; EVs, extracellular vesicles; Tfh, T follicular helper; gp130, glycoprotein 130; SH2D1B SH2, domain containing 1B, TNFα, tumor necrosis factor alpha; CCL4, chemokine ligands 4; DGF, delayed graft function; IFTA, interstitial fibrosis and tubular atrophy.
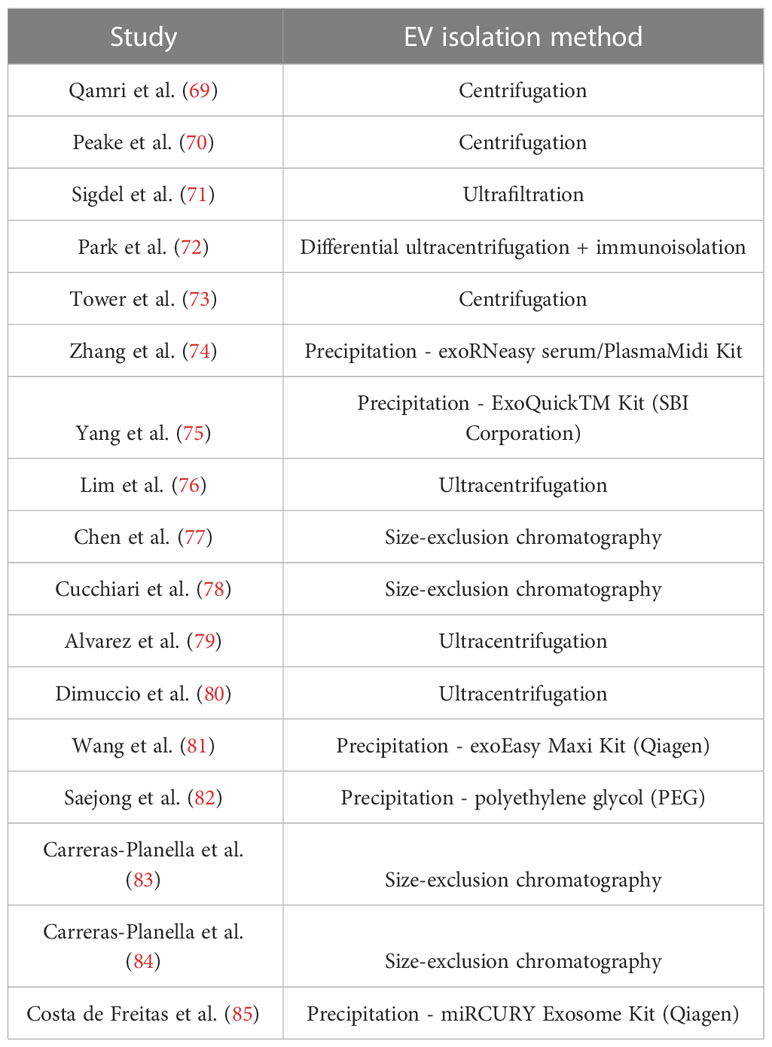
Table 2 Methods of EV isolation in each study mentioned.
In graft rejection, all the immune system components are involved in causing graft injury, including T and B lymphocytes, antibodies, endothelial cells, NK cells, macrophages, polymorphonuclear cells, or complement components (86).
Qamri et al. analyzed early post-transplant changes in circulating endothelial CD31+/CD42b− microparticle (EMP) levels after kidney transplant in 213 kidney recipients and 14 kidney + pancreas recipients. In the first cohort, no changes in circulating EMP levels were observed when graft dysfunction was unrelated to acute rejection. However, when this dysfunction was due to an episode of acute rejection (confirmed through a graft biopsy), an elevation in circulating EMP was detected. At the time of stratification according to PTC C4d staining, in patients with the negative one, a faster decrease in EMP levels was observed in comparison with patients with positive PTC C4d staining. This led the authors to suggest that circulating EMP levels could inform about ongoing endothelial cell injury. Moreover, when analyzing the different etiologies of end-stage kidney disease, a trend was found toward a decline in post-transplant EMP levels in causes due to diabetes mellitus, obstructive/inherited kidney disease, and autoimmune disease (69).
In another study by Peake et al., urinary exosomal mRNA from frequent kidney injury biomarkers such as neutrophil gelatinase-associated lipocalin (NGAL), interleukin-18 (IL-18) and kidney injury molecule-1 (KIM-1), together with the constitutively produced cystatin C, were compared with their corresponding synthesized urinary protein levels as well as with the creatinine reduction ratio (CRR) on post-operatory day 7. The results showed that, although urinary NGAL and IL-18 protein levels did correlate with CRR on day 7, this was not the case for mRNA inside urinary exosomes. The explanation for these findings lay in the selectivity for exosome packaging and does not have to be representative of the parenting cell (70).
One year later, Sigdel et al. described 11 proteins to be enriched in the urinary exosomes of patients with biopsy-proven acute rejection; three of these proteins (CLCA1, PROS1, and KIAA0753) had not been previously identified in healthy urine exosomal proteins (71).
Park et al. reported a urine-based platform, iKEA (integrated kidney exosome analysis), to detect rejection of kidney transplants through T-cell-derived EVs. This platform, based on a magneto-electrochemical sensing system, revealed a higher level of CD3-positive EVs in kidney rejection recipients, with an accuracy of approximately 91.1% (72).
Tower et al. found a correlation between plasma C4d+, especially C4d+/CD144+ microvesicles, and the presence of ABMR and its severity and response to treatment in kidney recipients. Ninety-five kidney recipients with for-cause biopsies performed and twenty-three healthy volunteers were evaluated. After histopathologic examination of the graft biopsies, 28 patients with ABMR were found. In them, the density of C4d+/CD144+ microvesicles was 11-fold greater than in kidney transplant patients without ABMR and 24-fold greater than in healthy volunteers. The densities of C4d+ and C4d+/annexin V+ (C4d+/AVB+) microvesicles were also higher in ABMR recipients. Moreover, C4d+/AVB+ microvesicles correlated with ABMR biopsy severity. Lastly, in nine cases, treatment was associated with a reduction in the densities of C4d+/CD144+ and CD144+ microvesicles, which also showed them to be a treatment response monitoring tool (73).
Zhang et al. selected 21 genes (related to inflammatory and IL-6 signaling events or elevated in renal biopsies of patients with ABMR) whose mRNA transcript levels were increased in plasma exosomes of ABMR kidney recipients compared with cell-mediated rejection and/or no rejection. The authors also generated a gene score with the combination of the transcript levels of four of these genes (gp130, SH2D1B, TNFα, and CCL4), which was significantly higher in the ABMR group than the other groups (74).
Yang et al. suggested a correlation between ABMR and follicular helper T cell (Tfh cell)-derived exosomes through their increase in the circulation of such patients compared with non-ABMR patients. Moreover, Tfh cell-derived exosomes promoted B cell proliferation and differentiation. Moreover, their study reported a decline in CTLA-4 expression on the Tfh cell-derived exosome surface in kidney transplant patients with ABMR. CTLA-4 is a leukocyte differentiation antigen and a transmembrane receptor on T cells, with an established role in alloantigen-driven T cell activation and various autoimmune diseases. CTLA-4 on exosomes inhibited human T cell activation by directly interacting with the molecules CD80 or CD86. Furthermore, intracellular CTLA-4 can inhibit Tfh cell differentiation, reduce IL-21 secretion, and inhibit B cell proliferation and differentiation into plasma cells (75).
Regarding acute T cell-mediated rejection, Lim et al. identified several urinary exosomal biomarker candidates in an Asian population of kidney transplant patients using a proteomics approach. Validation of the findings by western blot assay proved that tetraspanin-1 and hemopexin were significantly higher in TCMR patients (76).
Chen et al. established a circulating exosomal miRNA panel by extracting plasma exosomes from 58 kidney transplant recipients and 27 healthy controls. Exosomal miR-21, miR-210, and miR-4639 could discriminate between subjects with chronic kidney dysfunction and those with normal graft function. At one year follow-up, patients with a low calculated score from this three-miRNA panel revealed a stable recovery of allograft function (77).
Lastly, our group has proposed using B-cells–derived EVs (BEVs) to check B cell proliferation in secondary lymphoid organs and bone marrow after desensitization. BEVs (CD19+ or HLA-II+) were associated with surviving B cells in lymph nodes retrieved upon surgery on patients who received desensitization with Rituximab, plasma exchanges, and immunoglobulins. After the administration of Rituximab, no B cells were circulating. CD19+ or HLA-II+ EVs may reflect the mass of surviving B cells in secondary lymphoid organs that may predispose them to subsequent rejection. This is suggested by the rebound of BEVs in patients who develop ABMR after desensitization (78).
DGF leads to a higher risk of acute rejection and progression to chronic graft dysfunction (80, 87–89). The leading cause of DGF, ischemia-reperfusion injury (IRI), prompts a complex, alloantigen-independent immune response, which includes crosstalk between polymorphonuclear cells, macrophages, and donor cells as well as the release of EVs with pro-inflammatory and anti-inflammatory effects (80, 90, 91). Moreover, endothelial cells and renal tubular epithelial cells release EVs when exposed to oxidative stress, hypoxia, an acidic environment, or inflammation (8, 90, 91).
Among the first studies on exosomes and kidney dysfunction, Alvarez et al. evaluated if the different urine fractions (cellular or exosomal) have different NGAL expression in 15 kidney allograft recipients (eleven living donors and four deceased donors) soon after transplantation. Western blot analysis showed that the average NGAL expression in the exosomal fraction was significantly higher in deceased donor patients from the first post-operatory day and that its expression lasted increased in patients with DGF compared with non-DGF patients (79).
Dimuccio et al. showed lower levels of CD133-positive EVs in urine samples of transplanted patients. This decrease was evidenced from the first post-operatory day until day 7, when an increase was described. However, compared with patients with DGF, these last had a significant rise. Moreover, in patients with severe pre-transplant vascular injury of the allograft, CD133-positive EVs did not increase. The origin of CD133-positive EVs appeared restricted to glomeruli and proximal tubules. These data implicate CD133-positive cells in renal tissue regeneration after injury due to cold ischemia and IRI. Accordingly, no increase was observed in recipients with severe pre-transplant vascular damage, implying an inefficient regeneration of the graft tissue in these recipients (80).
In another study, Wang et al. explored miRNA expression profiling in the DGF process. Fifty-two known and five conserved exosomal miRNAs expressed in kidney-transplanted patients with DGF were identified. Three co-expressed exosomal miRNAs: hsa-miR-33a-5p R-1, hsa-miR-98-5p, and hsa-miR-151a-5p, were further observed to be significantly upregulated in the peripheral blood of DGF patients. Moreover, hsa-miR-151a-5p was positively correlated with the patient’s first-week serum creatinine levels, blood urea nitrogen, and uric acid after transplantation (81).
In the field of kidney transplant, fibrosis serves as the final and irreversible stage of the pathogenic mechanisms that lead to the loss of allograft function (92). For this reason, beyond the invasiveness of renal biopsy, clinical data need to be more specific to allow for early detection (92–95).
Saejong et al. describe the potential use of microRNA (miR)-21 expression in plasma exosomes for non-invasive monitoring of high-grade IFTA in kidney transplant patients. There are already previous studies on the role of exosomal miR-21 as a fibrosis biomarker and its association with TGF-β, a cytokine known to be involved in fibrosis pathogenesis. In the study by Saejong, miR-21 from the plasma exosome fraction (but not from the whole plasma) could discriminate between low- versus high-grade IFTA. It is demonstrated that the released miR-21 decreases phosphatase and tensin homolog (PTEN), which causes the phosphorylation of Protein kinase B (AKT) signaling, in turn reducing the expression of E-cadherin and raising the expression of α-SMA and fibronectin in kidney tubules (82).
More recently, Carreras-Planella et al. describe the search for kidney allograft dysfunction protein biomarkers related to four biopsy-proven diagnoses: IFTA, acute T-cell rejection, calcineurin inhibitors toxicity, and normal kidney function. The authors carried out a proteomic analysis of the urinary EVs (uEVs), discovering some EV-associated proteins that show different expressions depending on whether they come from pathological or normal kidney function allografts. Moreover, a change in the expression of vitronectin (VTN) was also evidenced in recipients with chronic IFTA, suggesting urinary VTN levels as another possible biomarker for monitoring fibrotic changes in kidney transplant patients. For the fibrosis process to occur, VTN must join the potent profibrotic glycoprotein PAI-1, although the precise mechanisms are arguable. VTN has been reported to increase PAI-1 activity in the renal tissue, hindering fibrinolysis. However, other studies described the opposite, highlighting a protective role against fibrosis (83).
The same group also demonstrated the potential role of uEVs as biomarkers of chronic calcineurin inhibitor toxicity (CNIT). Their nephrotoxicity and role in kidney fibrosis are known and have been described in multiple studies and they are first-line agents in the immunosuppressive regimen of kidney transplantation. The problems we continue to face are CNIT diagnosis and management. In this study, the urine from kidney transplant recipients with CNIT diagnosis is compared with recipients with IFTA and without CNIT or normal allograft function (all of them under a similar immunosuppressive scheme that included CNI). After data analysis, members of the uroplakin (UPK1A, UPK1B, UPK2, and UPK3A) and plakin families (periplakin and envoplakin) were significantly upregulated in the CNIT group, suggesting a central role in CNIT development. The binding of uroplakin proteins covers the urothelium’s surface to prevent urine influx from the lumen, also covering the renal pelvis, ureters, urinary bladder, and prostatic urethra. Periplakin and envoplakin function as cell-linker proteins. The upregulation of these proteins in the CNIT recipient’s uEV suggests that the toxic effect of CNI on the urothelium may increase their citolinker activity (84).
Lastly, Costa de Freitas et al. also evaluated the expression of different urinary exosome-derived miRNAs (exo-miRs) in transplant patients on a tacrolimus regimen. As a result, a difference in the expression of 16 exo-miRs was observed. Among them, the marked upregulation of miR-155-5p and downregulation of miR-223-3p and miR-1228-3p stand out. Moreover, it was found that the tacrolimus dose correlated with the expression of miR-155-5p and miR-223-3p, serum creatinine with the expression of miR-223-3p, and the number of blood leukocytes with miR-223-3p and miR-1228-3p (85).
EVs participate in intercellular communication in physiological and pathological processes, and in recent years, interest in them has grown as tools to monitor post-transplant evolution in a non-invasive way. Previous studies on diverse biological samples (blood or urine) include a wide range of pathologies such as kidney graft rejection (both cellular and humoral), DGF, IFTA, and drug toxicity. The limitations we have to consider are that the studies published and presented here do not often include multiple centers, the number of patients included is low, and the results have yet to be validated in larger cohorts. Appropriate method validation studies need to be improved, and the isolation protocol needs standardization to avoid the co-isolation of various vesicles or differences in contamination levels. The most modern technologies will likely offer new opportunities in this field; for instance, the Imaging Flow Cytometry (IFCM)-based methodology for the direct detection (without prior isolation) of donor-derived EVs (dd-EVs) in the plasma of kidney transplant patients based on Human Leukocyte Antigen (HLA) mismatch (96) or further investigation into the proteomic landscape and protein signature in urinary EVs (97).
Despite the promising published data, nowadays, we cannot use EVs as a definitive decision tool. Future studies are required before their analysis could facilitate the decision process in routine clinical practice. We still need basic parameters such as creatinine, proteinuria, or specific anti-HLA donor antibodies, and EV analysis may not replace the invasiveness of graft biopsy as the gold standard.
Future studies will extend our knowledge of the role of EVs as biomarkers in the kidney transplant field. A combination of biomarkers could help us decide whether a biopsy should be done and may have a supportive role when interpreting data provided by an allograft biopsy.
Conceptualization, EC-P, MR-B, EB-M, JR, and DC; resources, EC-P, MR-B, EB-M, JR, and FD; writing—original draft preparation, EC-P and DC; writing—review and editing, IR and DC; supervision, IR. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sellarés J, De Freitas DG, Mengel M, Reeve J, Einecke G, Sis B, et al. Understanding the causes of kidney transplant failure: The dominant role of antibody-mediated rejection and nonadherence. Am J Transpl (2012) 12(2):388–99. doi: 10.1111/j.1600-6143.2011.03840.x
2. Verhoeven JGHP, Boer K, Van Schaik RHN, Manintveld OC, Huibers MMH, Baan CC, et al. Liquid biopsies to monitor solid organ transplant function: A review of new biomarkers. Ther Drug Monit (2018) 40(5):515–25. doi: 10.1097/FTD.0000000000000549
3. Lo DJ, Kaplan B, Kirk AD. Biomarkers for kidney transplant rejection. Nat Rev Nephrol (2014) 10(4):215–25. doi: 10.1038/nrneph.2013.281
4. Van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Nephrol (2014) 10(4):215–25. doi: 10.1038/nrneph.2013.281
5. Stam J, Bartel S, Bischoff R, Wolters JC. Isolation of extracellular vesicles with combined enrichment methods. J Chromatogr B Analyt Technol BioMed Life Sci (2021) 1169:122604. doi: 10.1016/j.jchromb.2021.122604
6. Théry C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles (2018) 7(1):1535750. doi: 10.1080/20013078.2018.1535750
7. Witwer KW, Théry C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J Extracell Vesicles (2019) 8(1):1648167. doi: 10.1080/20013078.2019.1648167
8. Sailliet N, Ullah M, Dupuy A, Silva AKA, Gazeau F, Le Mai H, et al. Extracellular vesicles in transplantation. Front Immunol (2022) 13:800018. doi: 10.3389/fimmu.2022.800018
9. Gholizadeh S, Shehata Draz M, Zarghooni M, Sanati-Nezhad A, Ghavami S, Shafiee H, et al. Microfluidic approaches for isolation, detection, and characterization of extracellular vesicles: Current status and future directions. Biosens Bioelectron (2017) 91:588–605. doi: 10.1016/j.bios.2016.12.062
10. Karpman D, Ståhl AL, Arvidsson I. Extracellular vesicles in renal disease. Nat Rev Nephrol (2017) 13(9):545–62. doi: 10.1038/nrneph.2017.98
11. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol (2014) 30:255–89. doi: 10.1146/annurev-cellbio-101512-122326
12. Sedgwick AE, D’Souza-Schorey C. The biology of extracellular microvesicles. Traffic (2018) 19(5):319–27. doi: 10.1111/tra.12558
13. Jadli AS, Ballasy N, Edalat P, Patel VB. Inside(sight) of tiny communicator: exosome biogenesis, secretion, and uptake. Mol Cell Biochem (2020) 467(1-2):77–94. doi: 10.1007/s11010-020-03703-z
14. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal (2021) 19(1):47. doi: 10.1186/s12964-021-00730-1
15. György B, Módos K, Pállinger É, Pálóczi K, Pásztói M, Misják P, et al. Detection and isolation of cell-derived microparticles are compromised by protein complexes resulting from shared biophysical parameters. Blood (2011) 117(4):e39–48. doi: 10.1182/blood-2010-09-307595
16. Ashcroft J, Leighton P, Elliott TR, Hosgood SA, Nicholson ML, Kosmoliaptsis V. Extracellular vesicles in kidney transplantation: a state-of-the-art review. Kidney Int (2022) 101(3):485–97. doi: 10.1016/j.kint.2021.10.038
17. Colombo M, Moita C, Van Niel G, Kowal J, Vigneron J, Benaroch P, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci (2013) 126(24):5553–65. doi: 10.1242/jcs.128868
18. Gołębiewska JE, Wardowska A, Pietrowska M, Wojakowska A, Dębska-ślizień A. Small extracellular vesicles in transplant rejection. Cells (2021) 10(11):2989. doi: 10.3390/cells10112989
19. Kralj-Iglic VVP. Curvature-induced sorting of bilayer membrane constituents and formation of membrane rafts. Adv Planar Lipid Bilayers Liposomes (2006) 5:129–49. doi: 10.1016/S1554-4516(06)05005-8
20. Perez-Hernandez D, Gutiérrez-Vázquez C, Jorge I, López-Martín S, Ursa A, Sánchez-Madrid F, et al. The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J Biol Chem (2013) 288(17):11649–61. doi: 10.1074/jbc.M112.445304
21. Nazarenko I, Rana S, Baumann A, McAlear J, Hellwig A, Trendelenburg M, et al. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Cancer Res (2010) 70(4):1668–78. doi: 10.1158/0008-5472.CAN-09-2470
22. Batista BS, Eng WS, Pilobello KT, Hendricks-Muñoz KD, Mahal LK. Identification of a conserved glycan signature for microvesicles. J Proteome Res (2011) 10(10):4624–33. doi: 10.1021/pr200434y
23. Kooijmans SAA, Vader P, van Dommelen SM, van Solinge WW, Schiffelers RM. Exosome mimetics: A novel class of drug delivery systems. Int J Nanomed (2012) 7:1525–41. doi: 10.2147/IJN.S29661
24. Morishita M, Takahashi Y, Nishikawa M, Sano K, Kato K, Yamashita T, et al. Quantitative analysis of tissue distribution of the B16BL6-derived exosomes using a streptavidin-lactadherin fusion protein and Iodine-125-Labeled biotin derivative after intravenous injection in mice. J Pharm Sci (2015) 104(2):705–13. doi: 10.1002/jps.24251
25. Takahashi Y, Nishikawa M, Shinotsuka H, Matsui Y, Ohara S, Imai T, et al. Visualization and in vivo tracking of the exosomes of murine melanoma B16-BL6 cells in mice after intravenous injection. J Biotechnol (2013) 165(2):77–84. doi: 10.1016/j.jbiotec.2013.03.013
26. Rank A, Nieuwland R, Crispin A, Grützner S, Iberer M, Toth B, et al. Clearance of platelet microparticles in vivo. Platelets (2011) 22(2):111–6. doi: 10.3109/09537104.2010.520373
27. Yáñez-Mó M, Siljander PRM, Andreu Z, Zavec AB, Borràs FE, Buzas EI, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles J Extracell Vesicles (2015) 4:27066. doi: 10.3402/jev.v4.27066
28. Erdbrügger U, Lannigan J. Analytical challenges of extracellular vesicle detection: A comparison of different techniques. Cytometry A (2016) 89(2):123–34. doi: 10.1002/cyto.a.22795
29. Hill AF, Pegtel DM, Lambertz U, Leonardi T, O’Driscoll L, Pluchino S, et al. ISEV position paper: extracellular vesicle RNA analysis and bioinformatics. J Extracell Vesicles (2013) 2(1):22859. doi: 10.3402/jev.v2i0.22859
30. Witwer KW, Buzás EI, Bemis LT, Bora A, Lässer C, Lötvall J, et al. Standardization of sample collection, isolation and analysis methods in extracellular vesicle research. J Extracell Vesicles (2013) 2(1). doi: 10.3402/jev.v2i0.20360
31. Burrello J, Bolis S, Balbi C, Burrello A, Provasi E, Caporali E, et al. An extracellular vesicle epitope profile is associated with acute myocardial infarction. J Cell Mol Med (2020) 24(17):9945–57. doi: 10.1111/jcmm.15594
32. Burrello J, Monticone S, Burrello A, Bolis S, Cristalli CP, Comai G, et al. Identification of a serum and urine extracellular vesicle signature predicting renal outcome after kidney transplant. Nephrol Dial Transpl (2023) 38(3):764–77. doi: 10.1093/ndt/gfac259
33. Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol (2006). doi: 10.1002/0471143030.cb0322s30
34. Chen J, Li P, Zhang T, Xu Z, Huang X, Wang R, et al. Review on strategies and technologies for exosome isolation and purification. Front Bioeng Biotechnol (2022) 9:811971. doi: 10.3389/fbioe.2021.811971
35. Clos-Sansalvador M, Monguió-Tortajada M, Roura S, Franquesa M, Borràs FE. Commonly used methods for extracellular vesicles’ enrichment: Implications in downstream analyses and use. Eur J Cell Biol (2022) 101(3):151227. doi: 10.1016/j.ejcb.2022.151227
36. Castaman G. A bleeding disorder characterised by isolated deficiency of platelet microvesicle generation. Lancet (1996) 347(9002):700–1. doi: 10.1016/s0140-6736(96)91259-3
37. Stormorken H. A new syndrome: thrombocytopathia, muscle fatigue, asplenia, miosis, migraine, dyslexia and ichthyosis. Clin Genet (1985) 28:367–74. doi: 10.1111/j.1399-0004.1985.tb02209.x
38. Weiss HJ, Vicic WJ, Lages BA, Rogers J. Isolated deficiency of platelet procoagulant activity. Am J Med (1979) 67(2):206–13. doi: 10.1016/0002-9343(79)90392-9
39. Abid Hussein MN, Böing AN, Sturk A, Hau CM, Nieuwland R. Inhibition of microparticle release triggers endothelial cell apoptosis and detachment. Thromb Haemost (2007) 98:1096–107. doi: 10.1160/th05-04-0231
40. Zhang B, Wang M, Gong A, Zhang X, Wu X, Zhu Y, et al. HucMSc-exosome mediated-Wnt4 signaling is required for cutaneous wound healing. Stem Cells (2015) 33(7):2158–68. doi: 10.1002/stem.1771
41. Cano A, Ettcheto M, Bernuz M, Puerta R, de Antonio EE, Sánchez-López E, et al. Extracellular vesicles, the emerging mirrors of brain physiopathology. Int J Biol Sci (2023) 19(3):721–43. doi: 10.7150/ijbs.79063
42. Burrello J, Monticone S, Gai C, Gomez Y, Kholia S, Camussi G. Stem cell-derived extracellular vesicles and immune-modulation. Front Cell Dev Biol (2016) 4:83. doi: 10.3389/fcell.2016.00083
43. Ramirez-Bajo MJ, Rovira J, Lazo-Rodriguez M, Banon-Maneus E, Tubita V, Moya-Rull D, et al. Impact of mesenchymal stromal cells and their extracellular vesicles in a rat model of kidney rejection. Front Cell Dev Biol (2020) 8:10. doi: 10.3389/fcell.2020.00010
44. Lugini L, Valtieri M, Federici C, Cecchetti S, Meschini S, Condello M, et al. Exosomes from human colorectal cancer induce a tumor-like behavior in colonic mesenchymal stromal cells. Oncotarget (2016) 7(31):50086–98. doi: 10.18632/oncotarget.10574
45. Oehmcke S, Mörgelin M, Malmström J, Linder A, Chew M, Thorlacius H, et al. Stimulation of blood mononuclear cells with bacterial virulence factors leads to the release of pro-coagulant and pro-inflammatory microparticles. Cell Microbiol (2012) 14(1):107–19. doi: 10.1111/j.1462-5822.2011.01705.x
46. Wang JG, Williams JC, Davis BK, Jacobson K, Doerschuk CM, Ting JPY, et al. Monocytic microparticles activate endothelial cells in an IL-1β-dependent manner. Blood (2011) 118(8):2366–74. doi: 10.1182/blood-2011-01-330878
47. Tian C, Wang K, Zhao M, Cong S, Di X, Li R. Extracellular vesicles participate in the pathogenesis of sepsis. Front Cell Infect Microbiol (2022) 12:1018692. doi: 10.3389/fcimb.2022.1018692
48. Mastronardi ML, Mostefai HA, Meziani F, Martínez MC, Asfar P, Andriantsitohaina R. Circulating microparticles from septic shock patients exert differential tissue expression of enzymes related to inflammation and oxidative stress. Crit Care Med (2011) 39(7):1739–48. doi: 10.1097/CCM.0b013e3182190b4b
49. Prakash PS, Caldwell CC, Lentsch AB, Pritts TA, Robinson BRH. Human microparticles generated during sepsis in patients with critical illness are neutrophil-derived and modulate the immune response. J Trauma Acute Care Surg (2012) 73(2):401–6. doi: 10.1097/TA.0b013e31825a776d
50. Boilard E, Nigrovic PA, Larabee K, Watts GFM, Coblyn JS, Weinblatt ME, et al. Platelets amplify inflammation in arthritis via collagen-dependent microparticle production. Science (2010) 327(5965):580–3. doi: 10.1126/science.1181928
51. Gasser O, Schifferli JA. Activated polymorphonuclear neutrophils disseminate anti-inflammatory microparticles by ectocytosis. Blood (2004) 104(8):2543–8. doi: 10.1182/blood-2004-01-0361
52. Ratajczak J, Wysoczynski M, Hayek F, Janowska-Wieczorek A, Ratajczak MZ. Membrane-derived microvesicles: Important and underappreciated mediators of cell-to-cell communication. Leukemia (2006) 20(9):1487–95. doi: 10.1038/sj.leu.2404296
53. Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV, Melief C, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med (1996) 183(3):1161–72. doi: 10.1084/jem.183.3.1161
54. Giri PK, Schorey JS. Exosomes derived from M. bovis BCG infected macrophages activate antigen-specific CD4+ and CD8+ T cells in vitro and in vivo. PloS One (2008) 3(6):e2461. doi: 10.1371/journal.pone.0002461
55. Obregon C, Rothen-Rutishauser B, Gitahi SK, Gehr P, Nicod LP. Exovesicles from human activated dendritic cells fuse with resting dendritic cells, allowing them to present alloantigens. Am J Pathol (2006) 169(6):2127–36. doi: 10.2353/ajpath.2006.060453
56. Ramis JM. Extracellular vesicles in cell biology and medicine. Sci Rep (2020) 10(1):8667. doi: 10.1038/s41598-020-65826-z
57. Pêche H, Heslan M, Usal C, Amigorena S, Cuturi MC. Presentation of donor major histocompatibility complex antigens by bone marrow dendritic cell-derived exosomes modulates allograft rejection. Transplantation (2003) 76(10):1503–10. doi: 10.1097/01.TP.0000092494.75313.38
58. Segura E, Amigorena S, Théry C. Mature dendritic cells secrete exosomes with strong ability to induce antigen-specific effector immune responses. Blood Cells Mol Dis (2005) 35(2):89–93. doi: 10.1016/j.bcmd.2005.05.003
59. Pêche H, Renaudin K, Beriou G, Merieau E, Amigorena S, Cuturi MC. Induction of tolerance by exosomes and short-term immunosuppression in a fully MHC-mismatched rat cardiac allograft model. Am J Transpl (2006) 6(7):1541–50. doi: 10.1111/j.1600-6143.2006.01344.x
60. Mastoridis S, Londoño MC, Kurt A, Kodela E, Crespo E, Mason J, et al. Impact of donor extracellular vesicle release on recipient cell “cross-dressing” following clinical liver and kidney transplantation. Am J Transpl (2021) 21(7):2387–98. doi: 10.1111/ajt.16123
61. Gulinelli S, Salaro E, Vuerich M, Bozzato D, PizzIrani C, Bolognesi G, et al. IL-18 associates to microvesicles shed from human macrophages by a LPS/TLR-4 independent mechanism in response to P2X receptor stimulation. Eur J Immunol (2012) 42(12):3334–45. doi: 10.1002/eji.201142268
62. Castellani C, Burrello J, Fedrigo M, Burrello A, Bolis S, Di Silvestre D, et al. Circulating extracellular vesicles as non-invasive biomarker of rejection in heart transplant. J Heart Lung Transpl (2020) 39(10):1136–48. doi: 10.1016/j.healun.2020.06.011
63. Sharma M, Gunasekaran M, Ravichandran R, Fisher CE, Limaye AP, Hu C, et al. Circulating exosomes with lung self-antigens as a biomarker for chronic lung allograft dysfunction: A retrospective analysis. J Heart Lung Transpl (2020) 39(11):1210–9. doi: 10.1016/j.healun.2020.07.001
64. Gunasekaran M, Xu Z, Nayak DK, Sharma M, Hachem R, Walia R, et al. Donor-derived exosomes with lung self-antigens in human lung allograft rejection. Am J Transpl (2017) 17(2):474–84. doi: 10.1111/ajt.13915
65. Gunasekaran M, Sharma M, Hachem R, Bremner R, Smith MA, Mohanakumar T. Circulating exosomes with distinct properties during chronic lung allograft rejection. J Immunol (2018) 200(8):2535–41. doi: 10.4049/jimmunol.1701587
66. Vallabhajosyula P, Korutla L, Habertheuer A, Yu M, Rostami S, Yuan CX, et al. Tissue-specific exosome biomarkers for noninvasively monitoring immunologic rejection of transplanted tissue. J Clin Invest (2017) 127(4):1375–91. doi: 10.1172/JCI87993
67. Al-Massarani G, Vacher-Coponat H, Paul P, Arnaud L, Loundou A, Robert S, et al. Kidney transplantation decreases the level and procoagulant activity of circulating microparticles. Am J Transpl (2009) 9(3):550–7. doi: 10.1111/j.1600-6143.2008.02532.x
68. Al-Massarani G, Vacher-Coponat H, Paul P, Widemann A, Arnaud L, Loundou A, et al. Impact of immunosuppressive treatment on endothelial biomarkers after kidney transplantation. Am J Transpl (2008) 8(11):2360–7. doi: 10.1111/j.1600-6143.2008.02399.x
69. Qamri Z, Pelletier R, Foster J, Kumar S, MOmani H, Ware K, et al. Early posttransplant changes in circulating endothelial microparticles in patients with kidney transplantation. Transpl Immunol (2014) 31(2):60–4. doi: 10.1016/j.trim.2014.06.006
70. Peake PW, Pianta TJ, Succar L, Fernando M, Pugh DJ, McNamara K, et al. A comparison of the ability of levels of urinary biomarker proteins and exosomal mRNA to predict outcomes after renal transplantation. PloS One (2014) 9(6):e98644. doi: 10.1371/journal.pone.0098644
71. Sigdel KR, Cheng A, Wang Y, Duan L, Zhang YL. The emerging functions of long noncoding RNA in immune cells: autoimmune diseases. J Immunol Res (2015) 2015:848790. doi: 10.1155/2015/848790
72. Park J, Lin HY, Assaker JP, Jeong S, Huang CH, Kurdi A, et al. Integrated kidney exosome analysis for the detection of kidney transplant rejection. ACS Nano (2017) 11(11):11041–6. doi: 10.1021/acsnano.7b05083
73. Tower CM, Reyes M, Nelson K, Leca N, Kieran N, Muczynski K, et al. Plasma C4d+ endothelial microvesicles increase in acute antibody-mediated rejection. Transplantation (2017) 101(9):2235–43. doi: 10.1097/TP.0000000000001572
74. Zhang H, Huang E, Kahwaji J, Nast CC, Li P, Mirocha J, et al. Plasma exosomes from HLA-sensitized kidney transplant recipients contain mRNA transcripts which predict development of antibody-mediated rejection. Transplantation (2017) 101(10):2419–28. doi: 10.1097/TP.0000000000001834
75. Yang J, Bi L, He X, Wang Z, Qian Y, Xiao L, et al. Follicular helper T cell derived exosomes promote B cell proliferation and differentiation in antibody-mediated rejection after renal transplantation. BioMed Res Int (2019) 2019:6387924. doi: 10.1155/2019/6387924
76. Lim JH, Lee CH, Kim KY, Jung HY, Choi JY, Cho JH, et al. Novel urinary exosomal biomarkers of acute T cell-mediated rejection in kidney transplant recipients: A cross-sectional study. PloS One (2018) 13(9):e0204204. doi: 10.1371/journal.pone.0204204
77. Chen Y, Han X, Sun Y, He X, Xue D. A circulating exosomal microRNA panel as a novel biomarker for monitoring post-transplant renal graft function. J Cell Mol Med (2020) 24(20):12154–63. doi: 10.1111/jcmm.15861
78. Cucchiari D, Tubita V, Rovira J, Ramirez-Bajo MJ, Banon-Maneus E, Lazo-Rodriguez M, et al. B cell-derived extracellular vesicles reveal residual B cell activity in kidney graft recipients undergoing pre-transplant desensitization. Front Med (Lausanne) (2021) 8:781239. doi: 10.3389/fmed.2021.781239
79. Alvarez S, Suazo C, Boltansky A, Ursu M, Carvajal D, Innocenti G, et al. Urinary exosomes as a source of kidney dysfunction biomarker in renal transplantation. Transplant Proc (2013) 45(10):3719–23. doi: 10.1016/j.transproceed.2013.08.079
80. Dimuccio V, Ranghino A, Barbato LP, Fop F, Biancone L, Camussi G, et al. Urinary CD133+ extracellular vesicles are decreased in kidney transplanted patients with slow graft function and vascular damage. PloS One (2014) 9(8):e104490. doi: 10.1371/journal.pone.0104490
81. Wang J, Li X, Wu X, Wang Z, Zhang C, Cao G, et al. Expression profiling of exosomal miRNAs derived from the peripheral blood of kidney recipients with DGF using high-throughput sequencing. BioMed Res Int (2019) 2019:1759697. doi: 10.1155/2019/1759697
82. Saejong S, Townamchai N, Somparn P, Tangtanatakul P, Ondee T, HIrankarn N, et al. MicroRNA-21 in plasma exosome, but not from whole plasma, as a biomarker for the severe interstitial fibrosis and tubular atrophy (IF/TA) in post-renal transplantation. Asian Pac J Allergy Immunol (2022) 40(1):94–102. doi: 10.12932/AP-101019-0656
83. Carreras-Planella L, Cucchiari D, Cañas L, Juega J, Franquesa M, Bonet J, et al. Urinary vitronectin identifies patients with high levels of fibrosis in kidney grafts. J Nephrol (2021) 34(3):861–74. doi: 10.1007/s40620-020-00886-y
84. Carreras-Planella L, Juega J, Taco O, Cañas L, Franquesa M, Lauzurica R, et al. Proteomic characterization of urinary extracellular vesicles from kidney-transplanted patients treated with calcineurin inhibitors. Int J Mol Sci (2020) 21(20):1–14. doi: 10.3390/ijms21207569
85. Costa De Freitas RC, Bortolin RH, Vecchia Genvigir FD, Bonezi V, Crespo Hirata TD, Felipe CR, et al. Differentially expressed urinary exo-miRs and clinical outcomes in kidney recipients on short-term tacrolimus therapy: A pilot study. Epigenomics (2020) 12(22):2019–34. doi: 10.2217/epi-2020-0160
86. Monguió-Tortajada M, Lauzurica R, Borràs FE. Tolerance in organ transplantation: From conventional immunosuppression to extracellular vesicles. Front Immunol (2014) 5:416. doi: 10.3389/fimmu.2014.00416
87. Perico N, Cattaneo D, Sayegh M, Remuzzi G. Delayed graft function in kidney transplantation. Lancet (2004) 364(9447):1814–27. doi: 10.1016/S0140-6736(04)17406-0
88. Ponticelli C. Ischaemia-reperfusion injury: A major protagonist in kidney transplantation. Nephrol Dial Transpl (2014) 29(6):1134–40. doi: 10.1093/ndt/gft488
89. Eltzschig HK, Eckle T. Ischemia and reperfusion-from mechanism to translation. Nat Med (2011) 17(11):1391–401. doi: 10.1038/nm.2507
90. Quaglia M, Dellepiane S, Guglielmetti G, Merlotti G, Castellano G, Cantaluppi V. Extracellular vesicles as mediators of cellular crosstalk between immune system and kidney graft. Front Immunol (2020) 11:74. doi: 10.3389/fimmu.2020.00074
91. Duan P, Tan J, Miao Y, Zhang Q. Potential role of exosomes in the pathophysiology, diagnosis, and treatment of hypoxic diseases. Am J Transl Res (2019) 11(3):1184–201.
92. Boor P, Floege J. Renal allograft fibrosis: Biology and therapeutic targets. Am J Transpl (2015) 15(4):863–86. doi: 10.1111/ajt.13180
93. Eulalio A, Huntzinger E, Izaurralde E. Getting to the root of miRNA-mediated gene silencing. Cell (2008) 132(1):9–14. doi: 10.1016/j.cell.2007.12.024
94. Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem (2010) 79:351–79. doi: 10.1146/annurev-biochem-060308-103103
95. Gallo A, Tandon M, Alevizos I, Illei GG. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PloS One (2012) 7(3):e30679. doi: 10.1371/journal.pone.0030679
96. Woud WW, Hesselink DA, Hoogduijn MJ, Baan CC, Boer K. Direct detection of circulating donor-derived extracellular vesicles in kidney transplant recipients. Sci Rep (2022) 12(1):21973. doi: 10.1038/s41598-022-26580-6
Keywords: extracellular vesicles, exosomes, kidney, transplant, kidney transplant, biomarker
Citation: Cuadrado-Payán E, Ramírez-Bajo MJ, Bañón-Maneus E, Rovira J, Diekmann F, Revuelta I and Cucchiari D (2023) Physiopathological role of extracellular vesicles in alloimmunity and kidney transplantation and their use as biomarkers. Front. Immunol. 14:1154650. doi: 10.3389/fimmu.2023.1154650
Received: 30 January 2023; Accepted: 21 July 2023;
Published: 17 August 2023.
Edited by:
Long Zheng, The Second Affiliated Hospital of Zhejiang University School of Medicine, ChinaReviewed by:
Vincenzo Cantaluppi, University of Eastern Piedmont, ItalyCopyright © 2023 Cuadrado-Payán, Ramírez-Bajo, Bañón-Maneus, Rovira, Diekmann, Revuelta and Cucchiari. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Cucchiari, Y3VjY2hpYXJpQGNsaW5pYy5jYXQ=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.