
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 23 May 2023
Sec. Vaccines and Molecular Therapeutics
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1130539
 Shanti Pather1*
Shanti Pather1* Shabir A. Madhi2
Shabir A. Madhi2 Benjamin J. Cowling3
Benjamin J. Cowling3 Paul Moss4Jeremy P. Kamil5
Paul Moss4Jeremy P. Kamil5 Sandra Ciesek6Alexander Muik1Özlem Türeci1
Sandra Ciesek6Alexander Muik1Özlem Türeci1The highly transmissible Omicron (B.1.1.529) variant of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) was first detected in late 2021. Initial Omicron waves were primarily made up of sub-lineages BA.1 and/or BA.2, BA.4, and BA.5 subsequently became dominant in mid-2022, and several descendants of these sub-lineages have since emerged. Omicron infections have generally caused less severe disease on average than those caused by earlier variants of concern in healthy adult populations, at least, in part, due to increased population immunity. Nevertheless, healthcare systems in many countries, particularly those with low population immunity, have been overwhelmed by unprecedented surges in disease prevalence during Omicron waves. Pediatric admissions were also higher during Omicron waves compared with waves of previous variants of concern. All Omicron sub-lineages exhibit partial escape from wild-type (Wuhan-Hu 1) spike-based vaccine-elicited neutralizing antibodies, with sub-lineages with more enhanced immuno-evasive properties emerging over time. Evaluating vaccine effectiveness (VE) against Omicron sub-lineages has become challenging against a complex background of varying vaccine coverage, vaccine platforms, prior infection rates, and hybrid immunity. Original messenger RNA vaccine booster doses substantially improved VE against BA.1 or BA.2 symptomatic disease. However, protection against symptomatic disease waned, with reductions detected from 2 months after booster administration. While original vaccine-elicited CD8+ and CD4+ T-cell responses cross-recognize Omicron sub-lineages, thereby retaining protection against severe outcomes, variant-adapted vaccines are required to expand the breadth of B-cell responses and improve durability of protection. Variant-adapted vaccines were rolled out in late 2022 to increase overall protection against symptomatic and severe infections caused by Omicron sub-lineages and antigenically aligned variants with enhanced immune escape mechanisms.
The Omicron variant (Pango lineage B.1.1.529, GISAID clade GR/484A) of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) was first detected in November 2021 (1, 2). This first Omicron sub-lineage, now known as BA.1, contains numerous mutations, particularly in the spike protein, resulting in enhanced transmissibility, partial escape from previously established neutralizing antibodies to wild-type (Wuhan-Hu-1) SARS-CoV-2, and risk of re-infection (3–5). A sister lineage, BA.2, emerged soon after. Omicron was not the first variant of concern (VOC) to exhibit neutralizing antibody evasion; the Beta variant in particular exhibited partial resistance to neutralization by antibodies elicited by initial pandemic waves and first-generation vaccines (6). Unlike the Beta variant, which circulated for only a short time in a limited number of countries, the high reproductive rate of BA.1 and BA.2 allowed these variants to rapidly disseminate around the world. Although primary infection caused by BA.1 or BA.2 appeared to be generally less severe than previous VOCs, with case numbers doubling far quicker than previous waves, the emergence of these variants has been considered a turning point in the pandemic.
Owing to the large number of individuals infected with BA.1 or BA.2 and the high transmissibility of these variants, several sub-lineages of Omicron have since emerged (2, 7). This led to further waves of infection with sub-lineages BA.4 and BA.5 in many countries; BA.5 descendent lineages are now dominant globally (8). In this review, we describe the characteristics of Omicron B.1.1.529 and its sub-lineages, the associated burden of disease, and the impact of this group of variants on vaccine effectiveness (VE) in the context of evolving infection- and vaccine-induced population immunity. This review was written at a time when the BA.4, BA.5, and BA.2.75 sub-lineages were dominant in most regions and bivalent variant-adapted vaccines were being rolled out. Newer sub-lineages have since emerged and key data on these sub-lineages were added during peer review. The initial literature search was performed on June 20, 2022, with additional searches conducted on a case-by-case basis, owing to the rapidly evolving nature of this topic.
There has been significant intra-variant evolution since BA.1 was first detected in November 2021. BA.1 was supplanted by other Omicron sub-lineages, as was BA.3. Most initial BA.2 sub-lineages were supplanted by descendant sub-lineages such as BA.2.75, BA.4, and BA.5. Omicron sub-lineages currently being monitored by the World Health Organization are BA.5 sub-lineages BQ.1 (including descendent lineage BQ.1.1) and BF.7, BA.2 descendent lineages BA.2.75 and CH.1.1, and XBB (including descendent lineage XBB.1.5) (2). XBB is a recombinant of BA.2.10.1 and BA.2.75 (2). The term ‘Omicron’ encompasses BA.1, BA.2, BA.3, and all subsequent B.1.1.529 sub-lineages (2). The Omicron group of variants forms a new phylogenetically distinct clade that is not directly descended from other SARS-CoV-2 VOCs (9) (Figure 1). Of all the previous VOCs, BA.1 is most closely related to the Alpha and Gamma variants, underscoring those ancestors of the Omicron variant sub-lineages that remained in circulation during the Delta variant wave (9).
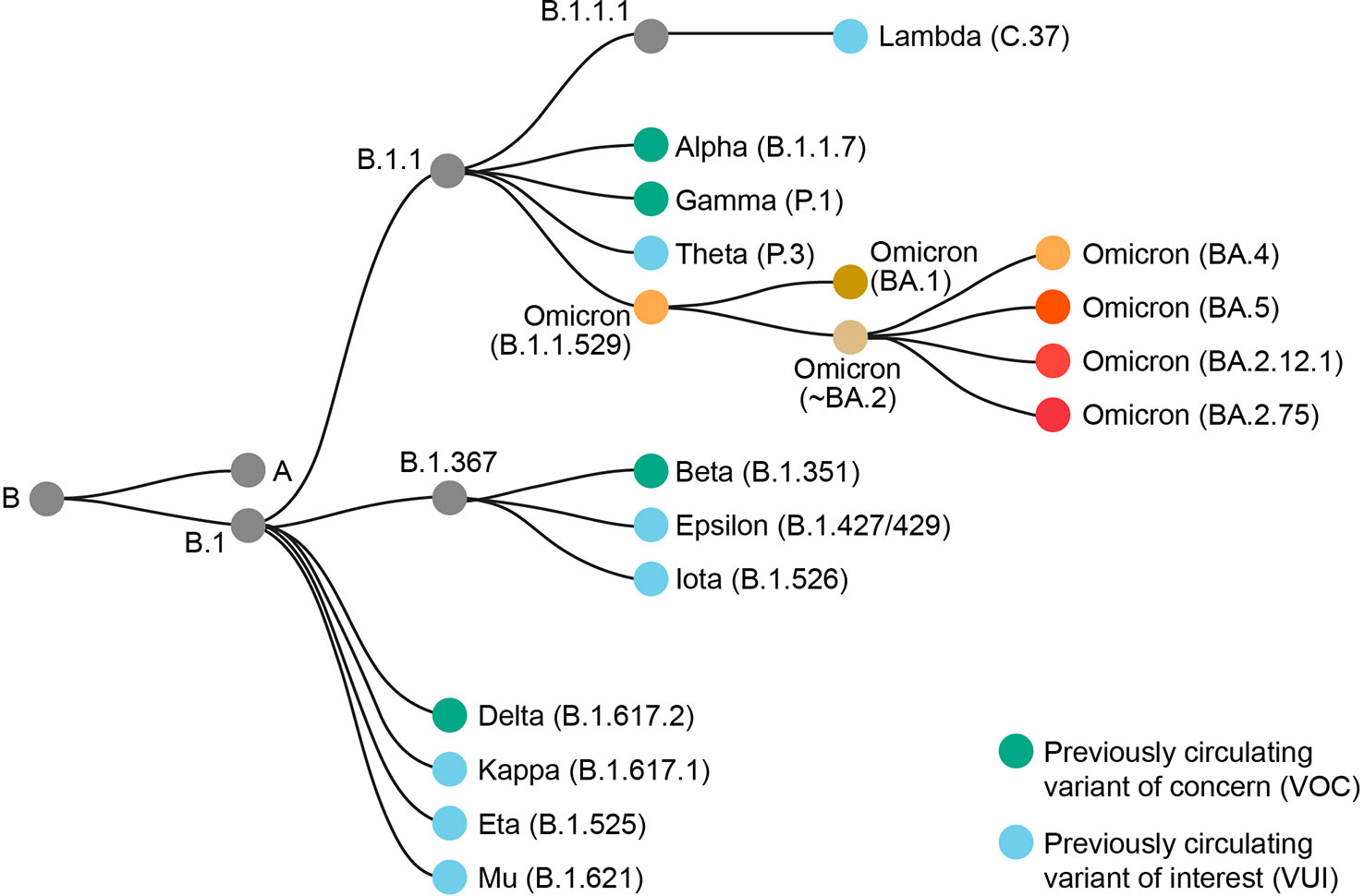
Figure 1 Phylogenetic relationship of Omicron B.1.1.529 and sub-lineages to other SARS-CoV-2 variants. Adapted from Nextstrain (7).
Key substitutions and deletions in the genomes of Omicron sub-lineages with known functions are shown in Table 1. A large number of these are linked to evasion of receptor-binding domain (RBD)- or N-terminal domain (NTD)-directed neutralizing antibodies (10, 12, 15, 16, 28), or enhanced binding to the angiotensin-converting enzyme 2 receptor (14, 17–19, 22). However, results from at least two studies in mice suggest that mutations affecting viral genes outside of S play critical roles in the reduced pathogenicity of Omicron lineages relative to earlier VOCs such as Delta or the ‘ancestral’ 2019 Wuhan lineage (29, 30). Alterations in non-S genes have been linked to changes in protein stability (24), increased host immune suppression (25), and enhanced sub-genomic RNA expression leading to increased viral load (26, 27, 31).
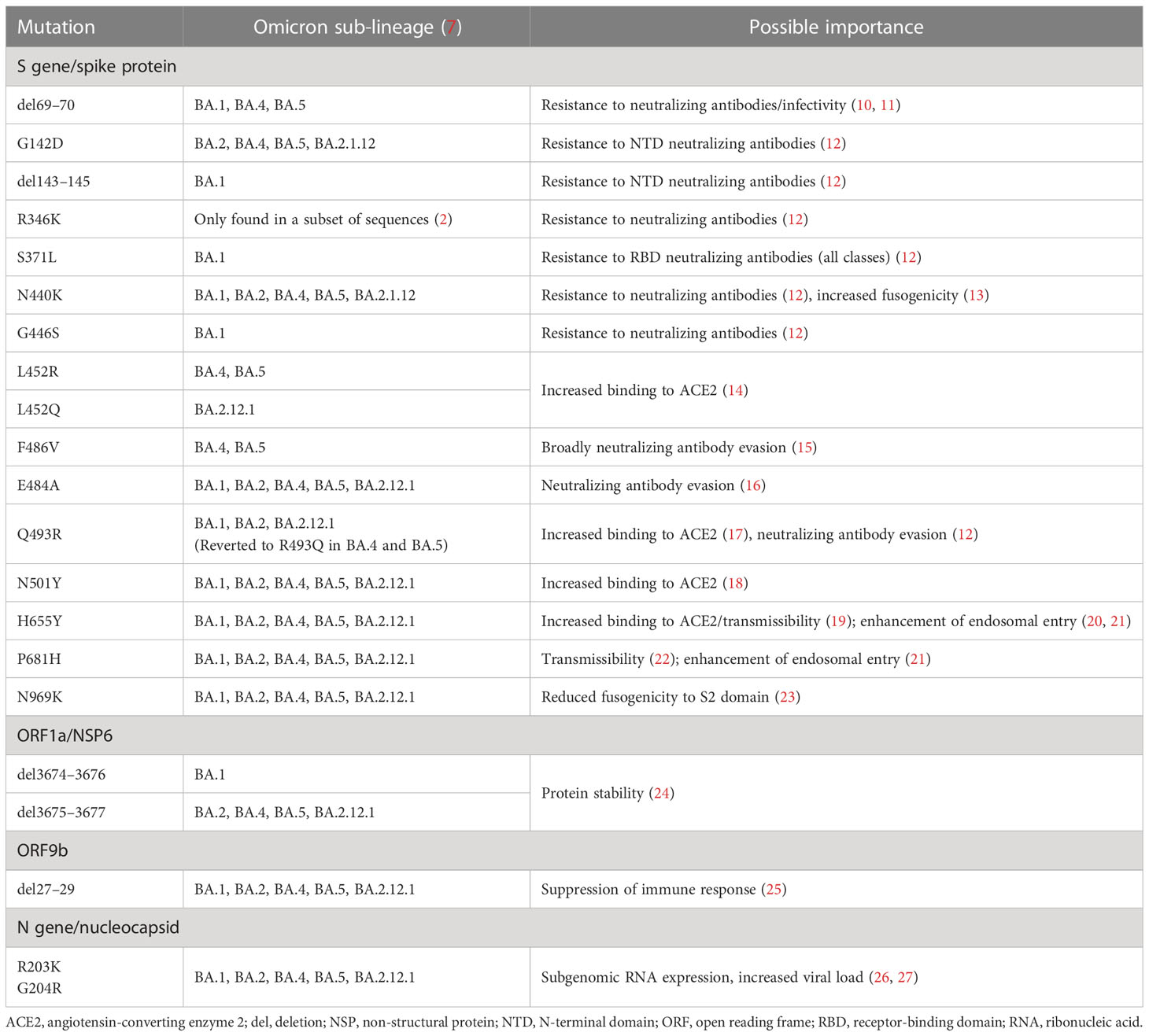
Table 1 Key genetic substitutions and deletions of Omicron BA.1 and sub-lineages BA.2, BA.4, BA.5, and BA.2.12.1.
Importantly, alterations to the S gene of the BA.1 and BA.2 sub-lineages have led to a fundamental change in the entry route of Omicron into host cells. While previous VOCs, such as Delta, were able to enter host cells via surface fusion following transmembrane protease serine 2-mediated proteolysis, BA.1 and BA.2 exhibit reduced fusogenicity, proteolysis, and syncytia formation, and have therefore switched entry pathway preference towards cathepsin-dependent fusion within the endosome (Figure 2) (20, 23, 28, 33). Enhanced endosomal entry and reduced fusogenicity have occurred as a result of alterations at the S1/S2 cleavage site and in the S2 domain, leading to reduced furin pre-processing, and retaining stabilization of the spike protein in the ‘closed’ conformation (20, 21, 28), which makes the RBD less accessible to neutralizing antibodies. As such, BA.1 and BA.2 have altered cell tropism compared with previous VOCs, favoring the upper respiratory tract (33).
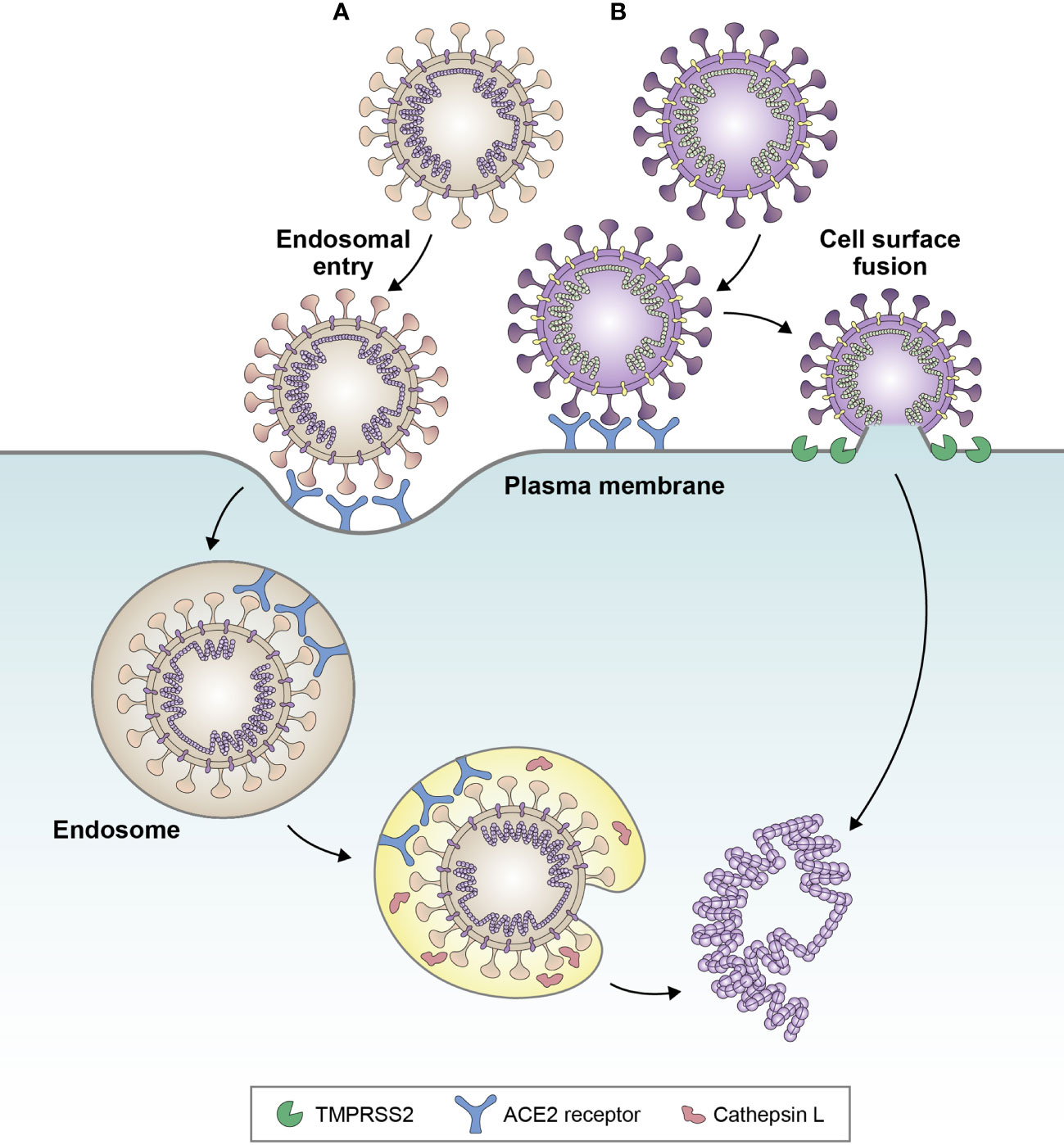
Figure 2 Favored cell entry pathways of (A) BA.1 and (B) Delta variant. Delta favors cell surface fusion, whereas BA.1 favors endosomal entry. Evidence suggests that BA.4 and BA.5 sub-lineages may be partially reverting back towards cell surface fusion, due to increased fusogenicity compared with BA.1. Adapted from Tang et al. Antiviral Res (2020);178:104792 (32).
Later Omicron sub-lineages, BA.4 and BA.5, have been shown to exhibit increased fusogenicity compared with BA.2, which has been attributed to the L452R and N440K substitutions (13, 34). Sub-lineages BA.4.6, BQ.1, and BQ.1.1 exhibit enhanced fusogenicity compared with BA.4 and BA.5 (13). The loss, and subsequent reattainment, of fusogenicity by Omicron sub-lineages is consistent with the Canyon Hypothesis (35), in which sustained viral transmission in seropositive populations leads to the emergence of variants with closed RBD configurations (such as BA.1/BA.2), reducing susceptibility to neutralizing antibodies. Circulation in populations with decreasing neutralizing antibody levels then leads to the emergence of variants with viral entry proteins favoring open configurations for more efficient cell entry (36). The F486V mutation of BA.4 and BA.5 has been shown to provide resistance to class 1 and class 2 RBD antibodies (15) and may have facilitated the move back towards the open configuration. In line with this, infection experiments in a guinea pig model have linked increased fusogenicity of BA.4 and BA.5 versus BA.2 with increased pathogenicity (37).
Antigenic drift has occurred rapidly with SARS-CoV-2 since the pandemic began and has led to a quick succession of changes in viral fitness. The emergence of recombinant variants, such as XBB and its descendants, demonstrates that antigenic shift may also occur, resulting in viruses that are antigenically distant from previously circulating variants and that have more evolved immuno-evasive mechanisms (38). The high frequency of changes in the genome of SARS-CoV-2 and the possibility of antigenic shift suggest that regular updates to coronavirus disease 2019 (COVID-19) vaccines may be required in the future, in order to provide protection against emerging antigenically distinct variants.
When compared with previous VOCs, neutralizing antibodies induced by original COVID-19 vaccines to BA.1 are reduced, as is memory B-cell recognition (39). BA.2 evades infection- and original vaccine-induced neutralizing antibodies with a comparable level of efficiency to BA.1 (40). Both BA.1 and BA.2 have been shown to evade neutralizing antibodies elicited by a primary series of messenger RNA (mRNA; mRNA-1273 or BNT162b2), vector-based (Ad26.COV2, Sputnik V, or ChAdOx1 nCoV-19), subunit (NVX-CoV2373), and inactivated (BBIBP-CorV) vaccines (41), although some activity is retained from mRNA or ChAdOx1 nCoV-19 vaccination. Original vaccine booster doses can restore neutralizing antibody activity against these Omicron sub-lineages to an extent, depending on the vaccine platforms used. Homologous vaccination with three doses of original mRNA vaccine (mRNA-1273 or BNT162b2), or heterologous vaccination with a vector-based vaccine (Ad26.COV2.S, ChAdOx1 nCoV-19, or Sputnik V) followed by an original mRNA booster, improves neutralization potency against BA.1 and BA.2 (41–43). In individuals vaccinated with a primary series of inactivated vaccine, a homologous booster enhances neutralizing antibody responses to BA.1, but to a lesser extent than heterologous mRNA vaccine boosters (44–47). Neutralizing antibody responses to BA.1 and BA.2 after original mRNA vaccine booster dosing have been shown to decline over time, reaching low levels at 4–6 months post-booster (48, 49). The rate of neutralizing antibody decay seems to be similar to that of wild-type SARS-CoV-2 (48). T-cell responses elicited by original mRNA vaccines against BA.1 are generally maintained for longer than humoral responses, suggesting that protection against severe disease may be preserved despite neutralizing antibody decay (50); however, T-cell responses may begin to wane from 6 months after infection (51).
Evidence suggests that hybrid immunity (vaccine-induced immunity in individuals who have also been infected) after a primary series of inactivated vaccine plus single original mRNA vaccine booster may generate higher neutralizing antibody responses to BA.2 than a second booster of either original mRNA or inactivated vaccine in individuals without previous infection (49).
Later Omicron sub-lineages, such as BA.4 and BA.5, seem to evade cross-protection from BA.1 infection. In individuals vaccinated with original vaccines, breakthrough BA.1 disease results in strong neutralizing antibody activity against BA.1 (52), reactivating memory B cells (53), and a less substantial increase in neutralizing antibody titers against BA.4 and BA.5 (52). Neutralizing antibody levels against BA.4 and BA.5 in non-vaccinated individuals with prior BA.1 infection are lower than those in individuals vaccinated with original vaccines and with BA.1 breakthrough infection (54). Neutralizing antibody titers are lower against BA.4, BA.5, and BA.2.12.1 than against BA.1 in individuals with prior BA.1 infection (17, 34, 54, 55). Exposure to BA.2 results in greater neutralizing activity against BA.4 and BA.5 than exposure to BA.1, driven by antibodies targeting the NTD of the spike protein (56). This is likely due to the fact that BA.2 is more closely phylogenetically related to BA.4 and BA.5 than BA.1 is (56, 57). BA.4 and BA.5 are antigenically distinct and are as distant from BA.1 as BA.1 is from the wild-type virus (38).
BA.4, BA.5, and BA.2.12.1 also escape original vaccine-elicited neutralizing antibodies to a greater extent than BA.1 or BA.2 (17, 34, 54, 58). BA.4 and BA.5 escape sera from individuals who received a primary series and booster of original mRNA vaccine to a greater extent than BA.2.12.1 (4.2-fold vs. 1.8-fold greater than BA.2 in one study (15) and by a factor of 3.3 vs. 2.2 compared with BA.1 in another (58)). However, individuals vaccinated with three doses of original mRNA vaccine with BA.4/BA.5 breakthrough infections have been shown to exhibit broad and robust neutralizing activity against BA.1, BA.2, BA.2.12.1, BA.4, and BA.5 (57). Nonetheless, BA.2.75.2, BQ.1, BQ.1.1, XBB, XBB.1, and CH.1.1 have been shown to exhibit lower neutralization sensitivity than BA.4/BA.5, indicating further neutralization escape with newly emerging sub-lineages (13, 38, 59–62). This is due to the phylogenetic distance between the XBB and BQ sub-lineages and BA.4/5 (38).
Owing to an extensive number of substitutions in the S gene relative to the ‘ancestral’ 2019 virus, Omicron variants are epistatically poised for escape from a variety of different antibodies (63). Notably, the efficacy of several monoclonal antibody therapies against Omicron sub-lineages BA.2.12.1, BA.4, and BA.5, as well as BQ and XBB sub-lineages, has been shown to be limited, although small-molecule antivirals retain activity (38, 64). There is also evidence that Omicron sub-lineages, as well as other VOCs, have developed relative resistance to interferons, affecting the host innate immune response (65).
Overall, neutralizing activity conferred by original COVID-19 vaccines is limited against newly emerging, antigenically distant sub-lineages; a pattern similar to that observed with influenza. The current trajectory suggests that SARS-CoV-2 will continue to evolve towards more immune-evasive variants, further affecting the effectiveness of vaccines and therapeutics. This further supports the need for regular updates to COVID-19 vaccine composition and booster vaccination to restore protection against circulating variants.
The transmissibility of BA.1 is clearly greater than that of previous VOCs, as demonstrated by its rapid global spread. BA.2 has been shown to be more transmissible than BA.1 (66), possibly attributable to an increased viral load in the upper pharynx (67). BA.4, BA.5, and BA.2.12.1 may be more transmissible than BA.2 (17, 68); however, owing to their increased fusogenicity, they also spread more efficiently in human lung cells, which suggests that they may be more likely to manifest as a lower respiratory tract infection (37). XBB.1.5 has an additional growth advantage over other Omicron sub-lineages, with a doubling time of 9 days (69). This is due to a higher angiotensin-converting enzyme 2 binding affinity compared with earlier variants, such as XBB.1 and BQ.1.1, which is conferred by the S486P mutation (70).
The risk of re-infection is increased with Omicron sub-lineages versus other VOCs (71). In Qatar, prior infection with pre-Omicron variants has been shown to provide less protection against symptomatic BA.1 infection than against symptomatic disease caused by other VOCs (56% vs. 85–92%, respectively) (72). In a study in Scotland, the proportion of BA.1 cases that were possible re-infections was >10 times greater than the proportion of Delta cases that were re-infections (7.6% vs. 0.7%, respectively) (73). This may be due to the fact that Omicron variants have become dominant, with prolonged circulation and resurgences compared with Delta and other previous VOCs. Nevertheless, prior infection with pre-Omicron variants does seem to provide a similar level of protection against severe, critical, or fatal disease due to re-infection with BA.1 versus other VOCs (72). This may also be partially due to the greater propensity of BA.1 for upper respiratory tract infection compared with other VOCs.
Immunity from prior infection can have a large role in protection against Omicron sub-lineages in countries with low vaccination rates. For example, in South Africa, high seroprevalence of SARS-CoV-2 immunoglobulin G after the Delta wave led to an apparent decoupling of infection from hospitalizations during the first Omicron wave (74). However, as new antigenically distinct sub-lineages with greater capacity for immune evasion emerge, the benefits of prior infection may be reduced, especially with waning of immunity over time. For example, in Qatar, effectiveness of pre-Omicron infection against BA.2.75 was 6%, and effectiveness of BA.1 or BA.2 infection against BA.2.75 was 50% (75). Protection conferred by prior infection against more antigenically distant and immune-evasive BQ sub-lineages is further reduced, as demonstrated by the increased re-infection risk with XBB sub-lineages (76).
The high transmissibility of Omicron sub-lineages and relative neutralizing antibody evasiveness have led to a rapid and substantial increase in disease prevalence, in terms of number of infections, compared with the pre-Omicron period, which has been sustained through continued replacement by emerging sub-lineages. Globally, the BA.1 wave began in November/December 2021 and peaked in January/February 2022 (77). BA.1 was then largely replaced by BA.2, and BA.2 was subsequently displaced by BA.4 and BA.5 around August 2022; the BA.2 descendent sub-lineage BA.2.75 also began to circulate at this time (77). Newer sub-lineages with enhanced immuno-evasive properties, including BQ.1.1, CH.1.1, and XBB.1.5, are now circulating in multiple countries (8). As Omicron sub-lineages are now dominant, it is reasonably likely that future VOCs will evolve from recent Omicron sub-lineages. It is also still possible that new VOCs will arise from non-dominant circulating viruses or earlier branches of the SARS-CoV-2 phylogenetic tree, as was observed with Omicron (9).
In many countries, the severity of infections in Omicron BA.1 waves has been milder than that of previous VOCs in terms of risk of hospitalization (73, 78–80), need for mechanical ventilation, and death (81, 82). These data must be interpreted in the context of continually evolving population immunity derived from infection and/or vaccination. Similarly, it is difficult to compare the clinical severity of each Omicron sub-lineage on this background of increasing and geographically variable population immunity. The current evidence does not indicate any significant change in disease severity associated with BA.2.12.1, BA.4, or BA.5 compared with BA.2, indeed, the reduced severity of BA.1 and BA.2 versus previous VOCs has persisted with BA.4 and BA.5 (83, 84), although BA.4 and BA.5 did not circulate at a time of significant circulation of other respiratory viruses in the Northern Hemisphere. There are no data to suggest an increase in disease severity with currently circulating sub-lineages, such as BQ.1.1 or XBB.1.5 (76, 85). There have been considerable regional differences in the severity of Omicron infections, likely linked to differences in population immunity. In South Africa, where infection-induced immunity was high and vaccine coverage was low, the BA.4/BA.5 wave has resulted in less severe infections than the BA.1 wave (86), with limited impact on healthcare services (74). In the United Kingdom, where vaccination coverage was high, the rise in prevalence during the BA.1 and BA.2 waves was associated with increases in hospitalizations and deaths, but at lower levels than previous waves (87). The impact has been greater in countries where vaccination rates or prior infection rates are low. The BA.2.2 wave in Hong Kong, where both vaccination coverage in older adults and prior infection rates were low, resulted in more than 1 million cases and close to 10,000 deaths (88, 89), and the BA.5 wave in New Zealand, where infection-induced immunity was low, led to a peak 7-day rolling average of 25 deaths per day, placing additional strain on the capacity of hospitals already overburdened with high caseloads (90). Estimates suggest that in China, which previously had very low rates of prior infection, up to 248 million people (18% of the population) may have been infected during the first 20 days of December 2022 (91).
Elderly, immunocompromised, comorbid, and unvaccinated populations remain at high risk of severe disease resulting from Omicron infection (78, 92). In addition, pediatric infections seem to be more frequent with Omicron sub-lineages versus earlier variants. Pediatric admissions were higher during the BA.1 wave than with previous waves (93–95), and SARS-CoV-2 seroprevalence in pediatric populations substantially increased, reaching 68–77% across age groups in the United States by February 2022 (96). In some countries, this has translated to increased severity in children, whereas in others, severity in children has been similar to, or lower than, previous waves. In Hong Kong, 2% of unvaccinated children hospitalized with BA.2 required admission to pediatric intensive care and two deaths were recorded (97). In the United Kingdom, the risk of hospital admission with Omicron BA.1 infection in children <10 years of age was comparable to that of Delta (98). In South Africa, although SARS-CoV-2 seropositivity in children <12 years of age reached 84% after the BA.1 wave (86), and the incidence of pediatric hospitalizations was similar to previous waves, mortality was lower (74). These differences are likely due to geographical variations in vaccination coverage and prior infection in pediatric populations. Pediatric admissions have also increased during circulation of the BQ.1.1 sub-lineage in the United States, with many States reporting that 90% of pediatric beds are occupied. However, other seasonal respiratory viruses such as respiratory syncytial virus and influenza have also been circulating in children during this period (99, 100).
Some evidence suggests a change in the rate of COVID-19 complications in pediatric populations with Omicron sub-lineages compared with previous VOCs. BA.1 has been associated with a significant increase in upper airway infection in children (4.1% during an Omicron-dominant period vs. 1.5% in the pre-Omicron period in the United States; p < 0.001) (101), although this may be partially attributable to increased testing rates. Of patients with croup at a US hospital, 48% were infected with SARS-CoV-2 during the BA.1 wave compared with only 3% during the Delta wave (102). Conversely, the relative risk of multisystem inflammatory syndrome after BA.1 infection in unvaccinated children in Denmark was significantly reduced compared with the Delta wave (0.12; 95% confidence interval [CI]: 0.06–0.23; p < 0.001) (103).
An increase in infections in pregnant women has also been observed during the BA.1 wave compared with Delta and pre-Delta periods (median 138 vs. 14 and 17 cases per week, respectively, in a study in the United States), with the majority occurring in unvaccinated individuals (104). The majority of these cases were non-severe, with an odds ratio for severe or critical illness versus the pre-Delta period of 0.20 (95% CI: 0.05–0.83). Severity of BA.1 may have been mitigated by evolving population immunity.
Omicron has also resulted in numerous healthcare worker (HCW) absences, further exacerbating pressure on healthcare systems. In England, approximately 40% of HCW absences in the last week of January 2022 (a BA.1-dominant period) were due to COVID-19 (105). Persistent symptoms are common in people recovering from COVID-19 and can hinder their ability to work (106). Thus, the presence of persistent symptoms may result in increased and/or prolonged HCW absences. Omicron outbreaks also increased stress among HCWs, potentially leading to further absences. In a survey of HCWs in Saudi Arabia, uncertainties around Omicron during the BA.1 wave were significantly correlated with stress, leading to reduced resilience and ability to cope (107).
Measures that countries have taken to support healthcare systems during Omicron waves include postponing elective surgeries to free up staff and beds, deploying military personnel to support hospitals, and recalling retired HCWs (108).
In summary, the emergence of the highly transmissible Omicron variant and its subsequent sub-lineages resulted in an increase in hospitalizations and HCW absenteeism, placing additional burden on already overstretched healthcare systems. The current evidence demonstrates that populations such as the elderly, people with co-morbidities, and pregnant women, remain at high risk of severe outcomes of Omicron infection, including hospitalization and death. This supports the need for continued booster vaccinations in these populations as SARS-CoV-2 continues to evolve.
As described earlier, Omicron sub-lineages exhibit partial escape from humoral immunity induced by current vaccines. In vaccinated individuals, the acute B-cell response to BA.1 breakthrough is mediated by vaccine-induced B-cell clones with a bias toward recognition of ancestral SARS-CoV-2 (52, 53). BA.1 breakthrough infection induces a shift towards the formation of memory B cells against epitopes that are broadly conserved across variants, with a robust recall response (52, 53). When compared with BA.1 convalescent patients, vaccinated individuals have been reported to have lower levels of BA.1-reactive B cells, as well as lower levels of neutralizing antibodies in bronchoalveolar lavage fluid (109). This may suggest that infection induces a greater mucosal immune response than vaccination. In line with these data, hybrid immunity has been shown to provide greater protection against symptomatic BA.1 or BA.2 disease than vaccination alone (110). In Qatar, effectiveness of three doses of BNT162b2 against symptomatic BA.2 infection in individuals with no prior infection at a median of 43 days post-booster was 52%, while effectiveness in individuals with a prior infection was 77% (110). Hybrid immunity from infection and booster vaccination seems to confer the greatest neutralization capacity (111), with broader activity and cross-reactive antibody affinity maturation against BA.1 and BA.2 versus infection-naïve booster-vaccinated individuals (112). Individuals vaccinated with three doses of mRNA vaccine with BA.4/BA.5 breakthrough infections exhibit broad and robust neutralizing activity against BA.1, BA.2, BA.2.12.1, BA.4, and BA.5 (57).
Although Omicron sub-lineages evade vaccine-elicited humoral responses to varying degrees (41, 54, 58), cell-mediated immunity to Omicron sub-lineages remains robust in vaccinated individuals. T-cell responses induced by different vaccine platforms, including mRNA (mRNA-1273 and BNT162b2), vector-based (Ad26.COV2.S), and subunit vaccines (NVX-CoV2373), cross-recognize variants from Alpha to BA.1 (39, 113–115). In vaccinees from the United States and South Africa, CD8+ T-cell responses to the spike protein of BA.1 induced by BNT162b2 or Ad26.COV2.S were >76% and CD4+ T-cell responses were >70% that of T-cell responses to the wild-type spike protein (114, 115). Some variation has been observed by vaccine platform; for example, in vaccinated individuals from Hong Kong, T-cell responses to BA.1 were higher in those who received two doses of BNT162b2 than in those who received two doses of the inactivated vaccine CoronaVac (CD8+: 81.8% vs. 71.4%; CD4+: 96.7% vs. 82.1%, respectively) (116). Conservation of the cell-mediated immune response in the lung may be associated with prevention of severe disease (109). This may partially explain why, despite the greater neutralization capacity of hybrid immunity versus vaccine-induced immunity, limited differences in VE against severe outcomes were observed between individuals with three mRNA vaccine doses with or without prior infection (110).
In VE studies across different settings, a booster dose of mRNA vaccine resulted in transient improved VE versus symptomatic disease during BA.1- or BA.2-dominant periods (117–122), but VE waned quickly over time. In England, VE against symptomatic BA.1 disease after a primary series of BNT162b2, mRNA-1273, or ChAdOx1 nCoV-19 followed by an mRNA booster had decreased at 5–9 weeks post-booster and further declined at >10 weeks (117). VE against symptomatic BA.2 disease decreased from 74% at 7 days post-booster to 44% at ≥15 weeks (119). In Qatar, effectiveness of a booster dose of BNT162b2 against symptomatic Omicron infection (any sub-lineage during a period of BA.1 and BA.2 dominance) decreased from 56% at 2–3 weeks post-booster to 22% at ≥14 weeks. Similarly, effectiveness of an mRNA-1273 booster decreased from 54% at 2–3 weeks post-booster to 35% at ≥6 weeks (121). In the United States, VE of mRNA-1273 against BA.1 infection decreased from 71.6% after 14–60 days post-booster to 47.4% after >60 days (118), and VE of BNT162b2 against BA.1 infection in HCWs decreased from 75% within 8 weeks to 55% at >16 weeks (120). However, VE against severe outcomes of BA.1, BA.2, or BA.4/5 infection has been very high following an mRNA booster dose (119, 122–127). In the United States, VE of three doses of mRNA vaccine against invasive mechanical ventilation or in-hospital death during the BA.1 wave was 94% (124). In Canada, VE against severe outcomes of BA.1 infection was 95% ≥7 days post-mRNA booster (122). In England, VE against BA.2 hospitalization post-mRNA booster peaked at 89% (119). A booster dose was also associated with a reduction in the risk of hospitalization and death due to BA.5 infection in Portugal (128). In children 5–11 years of age, effectiveness of a third dose of BNT162b2 against Omicron-related emergency department or urgent care encounters (any sub-lineage) was 77% after a median of 43 days post-booster (129). In the United States, effectiveness of three doses of BNT162b2 against BA.4/BA.5-related emergency department visits was 71% <3 months post-booster (127), and, in South Africa, effectiveness of three doses of BNT162b2 against hospitalization during the BA.4/BA.5 wave was 69% at 1−2 months post-booster (126).
VE against severe outcomes of BA.1, BA.2, and BA.4/BA.5 infection also wanes from 3–4 months after administration of the booster dose, albeit to a lesser extent than VE against symptomatic disease (94, 119, 126, 127, 130, 131). In some countries, a fourth vaccine dose has been administered to the elderly and/or HCWs, in order to provide additional protection against Omicron sub-lineages for these high-risk populations. In Israel, during a BA.1-dominant period, the rate of confirmed infection was reduced 2-fold and the rate of severe illness by 3.5-fold at 4 weeks after a fourth dose of BNT162b2 in adults >60 years of age compared with those who had only received three doses (132). Breakthrough COVID-19 infection rates in HCWs in Israel who received a fourth dose were lower than in those who received three doses during the BA.1 wave (7% vs. 20%, respectively) (133). In Ontario, Canada, VE of mRNA vaccines against symptomatic infection and severe outcomes in long-term care residents ≥60 years of age during a BA.1-dominant period was 69% and 86%, respectively, ≥7 days after the fourth dose (134). Given that booster doses provide additional protection against severe outcomes of Omicron infection, including hospitalization and death, continued vaccination of high-risk populations is likely.
Duration of neutralizing antibody protection against infection is a function both of sustained activity over time and variant sensitivity to neutralizing antibody activity. Based on the observed waning of VE against infection after booster administration (119, 130), and the fact that neutralizing antibody titers against Omicron sub-lineages are lower than those against prior VOCs after a booster dose (5, 43, 135), boosters against prior circulating VOCs may not address the complex needs posed by waning immune responses and new variants with enhanced transmissibility or pathogenicity. Therefore, adaptation of vaccines to include new VOCs, thus allowing increased duration and breadth of protection against infection, is highly desirable.
Variant-adapted vaccines are vaccines that have been updated to provide improved immune responses against a specific variant or variants (136). As well as addressing the specific variant/variants, these vaccines have the potential to increase the breadth of primary neutralizing antibody responses against other Omicron sub-lineages and prior VOCs when compared with the original vaccines (136, 137), through the formation of memory B cells against the new variant and robust recall of old memory B responses (52). Preservation and expansion of the T-cell response may potentially provide more durable protection against severe disease and deaths over time (138).
The need for variant-adapted vaccines was recognized after the emergence of BA.1, and mRNA vaccine manufacturers subsequently initiated the development of BA.1-adapted vaccines (139, 140). By the time data on the BA.1-adapted vaccines had been generated, sub-lineages BA.4 and BA.5 had increased in prevalence and were expected to become dominant. Data from BA.1 and BA.2 convalescent serum from triple mRNA-vaccinated individuals showed low neutralizing antibody titers against BA.4 and BA.5, suggesting that BA.1-adapted vaccines may not provide high levels of protection against BA.4/BA.5 infection (56). However, data from BA.4/BA.5-infected and triple mRNA-vaccinated individuals revealed robust neutralization of BA.4/BA.5 and all previously circulating Omicron sub-lineages (57). Manufacturers therefore also initiated development of BA.4/BA.5-adapted mRNA vaccines. Based on preliminary data on a prototype BA.4/BA.5-adapted vaccine, in July 2022, the United States Food and Drug Administration (FDA) recommended that modified vaccines should have a BA.4/BA.5 spike protein component in addition to the existing composition to create a bivalent booster for use in both adult and pediatric populations in Q3/Q4 2022 (141, 142).
Both the FDA and European Medicines Agency (EMA) have provided guidance on regulatory requirements for variant-adapted vaccines, stating that effectiveness can be established on the basis of immunogenicity bridging studies demonstrating superiority of the neutralizing antibody response elicited by the adapted vaccine compared with the prototype vaccine in terms of geometric mean titer ratios (143, 144). Based on such data, the EMA has granted full marketing authorization for a bivalent original BNT162b2/Omicron BA.1 vaccine in individuals ≥12 years of age, and for a bivalent original BNT162b2/Omicron BA.4-BA.5 vaccine in individuals 5–11 years of age (pediatric formulation) and ≥12 years of age (140). Bivalent mRNA-1273/Omicron BA.1 and mRNA-1273/Omicron BA.4-5 vaccines have received EMA approval for use in individuals ≥12 years of age (139). The FDA has granted Emergency Use Authorization for original BNT162b2/Omicron BA.4-BA.5 vaccines in individuals ≥5 years of age and mRNA-1273/Omicron BA.4-5 vaccines in individuals ≥6 years of age (145).
Early clinical data show that the bivalent BA.4-5 BNT162b2 vaccine induces greater pan-Omicron neutralizing activity and substantially higher neutralizing antibody titers against BA.4/BA.5 compared with the original vaccine in adults ≥55 years of age (136). The bivalent BA.4-5 BNT162b2 vaccine elicits T-cell responses to emerging immuno-evasive variants, such as XBB.1.5, albeit slightly reduced when compared with responses to the wild-type virus (146). Real-world data will further strengthen evidence for the effectiveness of these variant-adapted vaccines. Emerging data from the US Centers for Disease Control and Prevention (CDC) indicate a 2.4-times-lower risk of death due to COVID-19 in vaccinated individuals who received an Omicron BA.4-5 vaccine booster compared with those who did not (147). Although evidence suggests that the XBB.1.5 sub-lineage can partially escape neutralizing antibodies elicited by bivalent BA.4-5 boosters, VE estimates are similar to those against BA.5: CDC estimates suggest effectiveness against symptomatic infection in adults of 37–52% against BA.5 and 43–39% against XBB/XBB.1.5 at ≥2 weeks post-booster (61, 148).
Although the bivalent vaccines provide protection against currently circulating variants, given the continuing emergence of more antigenically distant sub-lineages and the increasing resistance of these sub-lineages to neutralizing antibodies, it is likely that further variant-adapted vaccines will be required as SARS-CoV-2 continues to evolve. In line with this, the Vaccines and Related Biological Products Advisory Committee anticipate conducting vaccine composition evaluations for COVID-19 vaccines at least annually, with a potential variant change to be selected in May 2023 (149). Bivalent vaccines may be implemented in vaccine-naïve populations as well as being administered as boosters, based on EMA Emergency Task Force guidance (150). Specific areas that still require global decision-making for future variant-adapted vaccines include the rationale for taking a monovalent versus a bivalent approach, and the rationale for requiring an updated vaccine, as well as the specific variant that should be included.
Ultimately, the value in developing a variant-adapted vaccine will always be mitigated by the time taken to detect the variant and develop and approve the vaccine (151), as well as the challenges to improving durability of protection during the frequent evolution of SARS-CoV-2. Several approaches to the development of novel vaccines that elicit a broader immune response and increased durability of protection against current and future variants are being researched (151–156), including candidates with enhanced prefusion spike proteins to improve magnitude and breadth of immune response (156), and candidates that target other non-spike antigens (153, 155). T-cell-enhanced vaccines composed of T-cell antigens encoding non-spike proteins that are conserved across variants are being evaluated and may have potential to expand protection against severe disease (157). Novel adjuvants designed to trigger specific components of innate immunity are being explored (158), and intranasal vaccines to elicit an enhanced immune response at the nasal mucosa are also in development (159).
Pan-sarbecovirus vaccines that provide broad and durable protection against all members of the sarbecovirus subgenus (i.e. SARS-CoV-2, SARS-CoV-1, and various non-human coronaviruses) are particularly desirable, and several approaches to this are under investigation (153, 155). Several mosaic nanoparticle vaccines, composed of numerous copies of the RBD or prefusion spike protein from SARS-CoV-2 and related viruses, such as Middle East Respiratory Syndrome coronavirus or coronaviruses circulating in bats, are in early development. A mosaic nanoparticle vaccine containing the RBD from SARS-CoV-2 and seven animal sarbecoviruses was shown to elicit broad immunity in preclinical studies (160). Several unanswered questions remain around the clinical development of pan-sarbecovirus vaccines, including the type of immunogenicity data that will be required in addition to vaccine efficacy and the correlates of immunity that can be used (particularly for cell-mediated immunogenicity studies). It remains to be seen whether such approaches will result in improved breadth and duration of protection compared with current bivalent COVID-19 vaccines.
Since the emergence of BA.1 in late 2021, new Omicron sub-lineages have continued to arise, superseding the previous Omicron sub-lineage approximately every 3 months. Later sub-lineages have had enhanced immuno-evasive properties and higher reproductive rates. Omicron sub-lineages continue to cause a substantial healthcare burden due to increased transmissibility, leading to a high prevalence of disease, and relative evasion of immunity, leading to re-infection and reduced VE. Novel vaccine strategies, such as variant-adapted vaccines or next-generation T-cell- enhanced approaches, are required to increase overall protection and durability of protection against symptomatic and severe infections caused by current and future Omicron sub-lineages, as well as other VOCs emerging from previously circulating variants.
All authors contributed to the manuscript conception, writing, and review process, and approved the final version for submission.
Medical writing support, including assisting authors with the development of the outline and initial draft and incorporation of comments was provided by Rachel Wright, PhD, and editorial support was provided by Ian Norton, PhD, all of Scion, London, UK, supported by BioNTech SE according to Good Publication Practice guidelines (Link).
SAM’s institution has received grants from the Bill and Melinda Gates Foundation and South African Medical Research Council, and grants for COVID-19 vaccine studies from Novavax, Gritsone, Providence, and ImmunityBio. BJC consults for AstraZeneca, Fosun Pharma, GlaxoSmithKline, Moderna, Pfizer, Roche, and Sanofi Pasteur. JPK has a grant from the Rockefeller Foundation to increase equity and representativeness in SARS-CoV-2 sequencing, has served on a BioNTech advisory panel, and holds stock in BioNTech, Pfizer, and Moderna, who manufacture COVID-19 vaccines. SC has received an honorarium for serving on a clinical advisory board for BioNTech. ÖT is a management board member and employee at BioNTech SE Mainz, Germany and co-founder of the company. AM and SP are employees at BioNTech SE. AM and ÖT are inventors on patents and patent applications related to RNA technology and COVID-19 vaccines. AM, SP, and ÖT have securities from BioNTech SE.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Viana R, Moyo S, Amoako DG, Tegally H, Scheepers C, Althaus CL, et al. Rapid epidemic expansion of the SARS-CoV-2 omicron variant in southern Africa. Nature (2022) 603(7902):679–86. doi: 10.1038/s41586-022-04411-y
2. World Health Organization. Tracking SARS-CoV-2 variants (2023). Available at: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (Accessed 03 February 2023).
3. Torjesen I. COVID-19: Omicron may be more transmissible than other variants and partly resistant to existing vaccines, scientists fear. BMJ (2021) 375:n2943. doi: 10.1136/bmj.n2943
4. World Health Organization (WHO). Update on Omicron (2021). Available at: https://www.who.int/news/item/28-11-2021-update-on-omicron. (Accessed 14 June 2022).
5. Wilhelm A, Widera M, Grikscheit K, Toptan T, Schenk B, Pallas C, et al. Limited neutralisation of the SARS-CoV-2 omicron subvariants BA.1 and BA.2 by convalescent and vaccine serum and monoclonal antibodies. EBioMedicine (2022) 82:104158. doi: 10.1016/j.ebiom.2022.104158
6. McLean G, Kamil J, Lee B, Moore P, Schulz TF, Muik A, et al. The impact of evolving SARS-CoV-2 mutations and variants on COVID-19 vaccines. mBio (2022) 13(2):e0297921. doi: 10.1128/mbio.02979-21
7. CoVariants. CoVariants (2022). Available at: https://covariants.org/ (Accessed 10 August 2022).
8. World Health Organization. Weekly epidemiological update on COVID-19 - 8 February 2023 (2023). Available at: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-COVID-19---8-february-2023. (Accessed 03 February 2023).
9. Kandeel M, Mohamed MEM, Abd El-Lateef HM, Venugopala KN, El-Beltagi HS. Omicron variant genome evolution and phylogenetics. J Med Virol (2022) 94(4):1627–32. doi: 10.1002/jmv.27515
10. McCarthy KR, Rennick LJ, Nambulli S, Robinson-McCarthy LR, Bain WG, Haidar G, et al. Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape. Science (2021) 371(6534):1139–42. doi: 10.1126/science.abf6950
11. Meng B, Kemp SA, Papa G, Datir R, Ferreira I, Marelli S, et al. Recurrent emergence of SARS-CoV-2 spike deletion H69/V70 and its role in the alpha variant B.1.1.7. Cell Rep (2021) 35(13):109292. doi: 10.1016/j.celrep.2021.109292
12. Liu L, Iketani S, Guo Y, Chan JF, Wang M, Liu L, et al. Striking antibody evasion manifested by the omicron variant of SARS-CoV-2. Nature (2022) 602(7898):676–81. doi: 10.1038/s41586-021-04388-0
13. Qu P, Evans JP, Faraone J, Zheng Y-M, Carlin C, Anghelina M, et al. Distinct neutralizing antibody escape of SARS-CoV-2 omicron subvariants BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2. bioRxiv (2022). doi: 10.1101/2022.10.19.512891
14. Motozono C, Toyoda M, Zahradnik J, Saito A, Nasser H, Tan TS, et al. SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microbe (2021) 29(7):1124–36 e11. doi: 10.1016/j.chom.2021.06.006
15. Wang Q, Guo Y, Iketani S, Nair MS, Li Z, Mohri H, et al. Antibody evasion by SARS-CoV-2 omicron subvariants BA.2.12.1, BA.4 and BA.5. Nature (2022) 608(7923):603–8. doi: 10.1038/s41586-022-05053-w
16. Greaney AJ, Loes AN, Crawford KHD, Starr TN, Malone KD, Chu HY, et al. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe (2021) 29(3):463–76 e6. doi: 10.1016/j.chom.2021.02.003
17. Cao Y, Yisimayi A, Jian F, Song W, Xiao T, Wang L, et al. BA.2.12.1, BA.4 and BA.5 escape antibodies elicited by omicron infection. Nature (2022) 608(7923):593–602. doi: 10.1038/s41586-022-04980-y
18. Zahradnik J, Marciano S, Shemesh M, Zoler E, Harari D, Chiaravalli J, et al. SARS-CoV-2 variant prediction and antiviral drug design are enabled by RBD in vitro evolution. Nat Microbiol (2021) 6(9):1188–98. doi: 10.1038/s41564-021-00954-4
19. Gong SY, Chatterjee D, Richard J, Prevost J, Tauzin A, Gasser R, et al. Contribution of single mutations to selected SARS-CoV-2 emerging variants spike antigenicity. Virology (2021) 563:134–45. doi: 10.1016/j.virol.2021.09.001
20. Yamamoto M, Tomita K, Hirayama Y, Inoue J-i, Kawaguchi Y, Gohda J. SARS-CoV-2 omicron spike H655Y mutation is responsible for enhancement of the endosomal entry pathway and reduction of cell surface entry pathways. bioRxiv (2022). doi: 10.1101/2022.03.21.485084
21. Hoffmann M, Kleine-Weber H, Pohlmann S. A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells. Mol Cell (2020) 78(4):779–84 e5. doi: 10.1016/j.molcel.2020.04.022
22. Rambaut A, Loman N, Pybus OG, Barclay W, Barrett J, Carabelli A, et al. Preliminary genomic characterisation of an emergent SARS-CoV-2 lineage in the UK defined by a novel set of spike mutations (2021). Available at: https://virological.org/t/preliminary-genomic-characterisation-of-an-emergent-SARS-cov-2-lineage-in-the-uk-defined-by-a-novel-set-of-spike-mutations/563. (Accessed 14 September 2021).
23. Peacock TP, Brown JC, Zhou J, Thakur N, Sukhova K, Newman J, et al. The altered entry pathway and antigenic distance of the SARS-CoV-2 omicron variant map to separate domains of spike protein. bioRxiv (2022). doi: 10.1101/2021.12.31.474653
24. Benvenuto D, Angeletti S, Giovanetti M, Bianchi M, Pascarella S, Cauda R, et al. Evolutionary analysis of SARS-CoV-2: how mutation of non-structural protein 6 (NSP6) could affect viral autophagy. J Infect (2020) 81(1):e24–e7. doi: 10.1016/j.jinf.2020.03.058
25. Thorne LG, Bouhaddou M, Reuschl AK, Zuliani-Alvarez L, Polacco B, Pelin A, et al. Evolution of enhanced innate immune evasion by SARS-CoV-2. Nature (2022) 602:487–95. doi: 10.1038/s41586-021-04352-y
26. Leary S, Gaudieri S, Parker MD, Chopra A, James I, Pakala S, et al. Generation of a novel SARS-CoV-2 sub-genomic RNA due to the R203K/G204R variant in nucleocapsid: homologous recombination has potential to change SARS-CoV-2 at both protein and RNA level. Pathog Immun (2021) 6(2):27–49. doi: 10.20411/pai.v6i2.460
27. Mourier T, Shuaib M, Hala S, Mfarrej S, Alofi F, Naeem R, et al. SARS-CoV-2 genomes from Saudi Arabia implicate nucleocapsid mutations in host response and increased viral load. Nat Commun (2022) 13(1):601. doi: 10.1038/s41467-022-28287-8
28. Meng B, Abdullahi A, Ferreira I, Goonawardane N, Saito A, Kimura I, et al. Altered TMPRSS2 usage by SARS-CoV-2 omicron impacts infectivity and fusogenicity. Nature (2022) 603(7902):706–14. doi: 10.1038/s41586-022-04474-x
29. Chen D-Y, Kenney D, Chin CV, Tavares AH, Khan N, Conway HL, et al. Role of spike in the pathogenic and antigenic behavior of SARS-CoV-2 BA.1 omicron. bioRxiv (2022). doi: 10.1101/2022.10.13.512134
30. Simon-Loriere E, Montagutelli X, Lemoine F. Rapid characterization of a delta-omicron SARS-CoV-2 recombinant detected in Europe. Res Square (2022). doi: 10.21203/rs.3.rs-1502293/v1
31. Mears HV, Young GR, Sanderson T, Harvey R, Crawford M, Snell DM, et al. Emergence of new subgenomic mRNAs in SARS-CoV-2. bioRxiv (2022). doi: 10.1101/2022.04.20.488895
32. Tang T, Bidon M, Jaimes JA, Whittaker GR, Daniel S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antiviral Res (2020) 178:104792. doi: 10.1016/j.antiviral.2020.104792
33. Willett BJ, Grove J, MacLean OA, Wilkie C, De Lorenzo G, Furnon W, et al. SARS-CoV-2 omicron is an immune escape variant with an altered cell entry pathway. Nat Microbiol (2022) 7(8):1161–79. doi: 10.1038/s41564-022-01143-7
34. Qu P, Faraone JN, Evans JP, Zou X, Zheng Y-M, Carlin C, et al. Differential evasion of delta and omicron immunity and enhanced fusogenicity of SARS-CoV-2 omicron BA.4/5 and BA.2.12.1 subvariants. bioRxiv (2022). doi: 10.1101/2022.05.16.492158
35. Rossmann MG. The canyon hypothesis. hiding the host cell receptor attachment site on a viral surface from immune surveillance. J Biol Chem (1989) 264(25):14587–90.
36. Wolf KA, Kwan JC, Kamil JP. Structural dynamics and molecular evolution of the SARS-CoV-2 spike protein. mBio (2022) 13(2):e0203021. doi: 10.1128/mbio.02030-21
37. Kimura I, Yamasoba D, Tamura T, Nao N, Oda Y, Mitoma S, et al. Virological characteristics of the novel SARS-CoV-2 omicron variants including BA.2.12.1, BA.4 and BA.5. bioRxiv (2022). doi: 10.1101/2022.05.26.493539
38. Wang Q, Iketani S, Li Z, Liu L, Guo Y, Huang Y, et al. Alarming antibody evasion properties of rising SARS-CoV-2 BQ and XBB subvariants. Cell (2023) 186(2):279–86 e8. doi: 10.1016/j.cell.2022.12.018
39. Tarke A, Coelho CH, Zhang Z, Dan JM, Yu ED, Methot N, et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from alpha to omicron. Cell (2022) 185(5):847–59 e11. doi: 10.1016/j.cell.2022.01.015
40. Arora P, Zhang L, Rocha C, Sidarovich A, Kempf A, Schulz S, et al. Comparable neutralisation evasion of SARS-CoV-2 omicron subvariants BA.1, BA.2, and BA.3. Lancet Infect Dis (2022) 22(6):766–7. doi: 10.1016/S1473-3099(22)00224-9
41. Bowen JE, Addetia A, Dang HV, Stewart C, Brown JT, Sharkey WK, et al. Omicron spike function and neutralizing activity elicited by a comprehensive panel of vaccines. Science (2022) 377(6608):890–94. doi: 10.1126/science.abq0203
42. Leung NHL, Cheng SMS, Martin-Sanchez M, Au NYM, Ng YY, Luk LLH, et al. Immunogenicity of a third dose of BNT162b2 to ancestral SARS-CoV-2 & omicron variant in adults who received two doses of inactivated vaccine. Clin Infect Dis (2022) 76(33):e299–307. doi: 10.1093/cid/ciac458
43. Muik A, Lui BG, Wallisch AK, Bacher M, Muhl J, Reinholz J, et al. Neutralization of SARS-CoV-2 omicron by BNT162b2 mRNA vaccine-elicited human sera. Science (2022) 375(6581):678–80. doi: 10.1126/science.abn7591
44. Cheng SMS, Mok CKP, Leung YWY, Ng SS, Chan KCK, Ko FW, et al. Neutralizing antibodies against the SARS-CoV-2 omicron variant BA.1 following homologous and heterologous CoronaVac or BNT162b2 vaccination. Nat Med (2022) 28(3):486–9. doi: 10.1038/s41591-022-01704-7
45. Perez-Then E, Lucas C, Monteiro VS, Miric M, Brache V, Cochon L, et al. Neutralizing antibodies against the SARS-CoV-2 delta and omicron variants following heterologous CoronaVac plus BNT162b2 booster vaccination. Nat Med (2022) 28(3):481–5. doi: 10.1038/s41591-022-01705-6
46. Costa Clemens SA, Weckx L, Clemens R, Almeida Mendes AV, Ramos Souza A, Silveira MBV, et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001): a phase 4, non-inferiority, single blind, randomised study. Lancet (2022) 399(10324):521–9. doi: 10.1016/S0140-6736(22)00094-0
47. Zuo F, Abolhassani H, Du L, Piralla A, Bertoglio F, de Campos-Mata L, et al. Heterologous immunization with inactivated vaccine followed by mRNA-booster elicits strong immunity against SARS-CoV-2 omicron variant. Nat Commun (2022) 13(1):2670. doi: 10.1038/s41467-022-30340-5
48. Xia H, Zou J, Kurhade C, Cai H, Yang Q, Cutler M, et al. Neutralization and durability of 2 or 3 doses of the BNT162b2 vaccine against omicron SARS-CoV-2. Cell Host Microbe (2022) 30(4):485–8 e3. doi: 10.1016/j.chom.2022.02.015
49. Cowling BJ, Cheng SMS, Martin-Sanchez M, Au NYM, Chan KCK, Li JKC, et al. Slow waning of antibodies following a third dose of BNT162b2 in adults who had previously received two doses of inactivated vaccine. J Infect Dis (2022) 227(2):251–5. doi: 10.1093/infdis/jiac380
50. GeurtsvanKessel CH, Geers D, Schmitz KS, Mykytyn AZ, Lamers MM, Bogers S, et al. Divergent SARS-CoV-2 omicron-reactive T and B cell responses in COVID-19 vaccine recipients. Sci Immunol (2022) 7(69):eabo2202. doi: 10.1126/sciimmunol.abo2202
51. Moss P. The T cell immune response against SARS-CoV-2. Nat Immunol (2022) 23(2):186–93. doi: 10.1038/s41590-021-01122-w
52. Quandt J, Muik A, Salisch N, Lui BG, Lutz S, Kruger K, et al. Omicron BA.1 breakthrough infection drives cross-variant neutralization and memory B cell formation against conserved epitopes. Sci Immunol (2022) 7(75):eabq2427. doi: 10.1126/sciimmunol.abq2427
53. Kaku CI, Bergeron AJ, Ahlm C, Normark J, Sakharkar M, Forsell MNE, et al. Recall of preexisting cross-reactive B cell memory after omicron BA.1 breakthrough infection. Sci Immunol (2022) 7(73):eabq3511. doi: 10.1126/sciimmunol.abq3511
54. Khan K, Karim F, Ganga Y, Bernstein M, Jule Z, Reedoy K, et al. Omicron BA.4/BA.5 escape neutralizing immunity elicited by BA.1 infection. Nat Commun (2022) 13(1):4686. doi: 10.1038/s41467-022-32396-9
55. Kurhade C, Zou J, Xia H, Liu M, Yang Q, Cutler M, et al. Neutralization of omicron sublineages and deltacron SARS-CoV-2 by three doses of BNT162b2 vaccine or BA.1 infection. Emerg Microbes Infect (2022) 11(1):1828–32. doi: 10.1080/22221751.2022.2099305
56. Muik A, Lui BG, Bacher M, Wallisch AK, Toker A, Finlayson A, et al. Omicron BA.2 breakthrough infection enhances cross-neutralization of BA.2.12.1 and BA.4/BA.5. Sci Immunol (2022) 7(77):eade2283. doi: 10.1126/sciimmunol.ade2283
57. Muik A, Lui BG, Bacher M, Wallisch AK, Toker A, Couto CIC, et al. Exposure to BA.4/5 s protein drives neutralization of omicron BA.1, BA.2, BA.2.12.1, and BA.4/5 in vaccine-experienced humans and mice. Sci Immunol (2022) 7(78):eade9888. doi: 10.1126/sciimmunol.ade9888
58. Hachmann NP, Miller J, Collier AY, Ventura JD, Yu J, Rowe M, et al. Neutralization escape by SARS-CoV-2 omicron subvariants BA.2.12.1, BA.4, and BA.5. N Engl J Med (2022) 387(1):86–8. doi: 10.1056/NEJMc2206576
59. Kurhade C, Zou J, Xia H, Liu M, Chang HC, Ren P, et al. Low neutralization of SARS-CoV-2 omicron BA.2.75.2, BQ.1.1 and XBB.1 by parental mRNA vaccine or a BA.5 bivalent booster. Nat Med (2022) 29(2):344–47. doi: 10.1038/s41591-022-02162-x
60. Jiang N, Wang L, Hatta M, Feng C, Currier M, Lin X, et al. Bivalent mRNA vaccine improves antibody-mediated neutralization of many SARS-CoV-2 omicron lineage variants. bioRxiv (2023). doi: 10.1101/2023.01.08.523127
61. Qu P, Faraone JN, Evans JP, Zheng Y-M, Carlin C, Anghelina M, et al. Extraordinary evasion of neutralizing antibody response by omicron XBB.1.5, CH.1.1 and CA.3.1 variants. bioRxiv (2023). doi: 10.1101/2023.01.16.524244
62. Miller J, Hachmann NP, Collier AY, Lasrado N, Mazurek CR, Patio RC, et al. Substantial neutralization escape by SARS-CoV-2 omicron variants BQ.1.1 and XBB.1. N Engl J Med (2023) 388(7):662–64. doi: 10.1056/NEJMc2214314
63. Witte L, Baharani V, Schmidt F, Wang Z, Cho A, Raspe R, et al. Epistasis lowers the genetic barrier to SARS-CoV-2 neutralizing antibody escape. bioRxiv (2022). doi: 10.1101/2022.08.17.504313
64. Takashita E, Yamayoshi S, Simon V, van Bakel H, Sordillo EM, Pekosz A, et al. Efficacy of antibodies and antiviral drugs against omicron BA.2.12.1, BA.4, and BA.5 subvariants. N Engl J Med (2022) 387(5):468–70. doi: 10.1056/NEJMc2207519
65. Guo K, Barrett BS, Morrison JH, Mickens KL, Vladar EK, Hasenkrug KJ, et al. Interferon resistance of emerging SARS-CoV-2 variants. Proc Natl Acad Sci USA (2022) 119(32):e2203760119. doi: 10.1073/pnas.2203760119
66. Lyngse FP, Kirkeby CT, Denwood M, Christiansen LE, Molbak K, Moller CH, et al. Household transmission of SARS-CoV-2 omicron variant of concern subvariants BA.1 and BA.2 in Denmark. Nat Commun (2022) 13(1):5760. doi: 10.1038/s41467-022-33498-0
67. Lentini A, Pereira A, Winqvist O, Reinius B. Monitoring of the SARS-CoV-2 omicron BA.1/BA.2 lineage transition in the Swedish population reveals increased viral RNA levels in BA.2 cases. Med (2022) 3(9):636–43. doi: 10.1016/j.medj.2022.07.007
68. Tegally H, Moir M, Everatt J, Giovanetti M, Scheepers C, Wilkinson E, et al. Emergence of SARS-CoV-2 omicron lineages BA.4 and BA.5 in South Africa. Nat Med (2022) 28(9):1785–90. doi: 10.1038/s41591-022-01911-2
69. European Centre for Disease Prevention and Control (ECDC). Update on SARS-CoV-2 variants: ECDC assessment of the XBB.1.5 sub-lineage (2023). Available at: https://www.ecdc.europa.eu/en/news-events/update-SARS-cov-2-variants-ecdc-assessment-xbb15-sub-lineage. (Accessed 13 February 2023).
70. Yue C, Song W, Wang L, Jian F, Chen X, Gao F, et al. Enhanced transmissibility of XBB.1.5 is contributed by both strong ACE2 binding and antibody evasion. bioRxiv (2023). doi: 10.1101/2023.01.03.522427
71. Pulliam JRC, van Schalkwyk C, Govender N, von Gottberg A, Cohen C, Groome MJ, et al. Increased risk of SARS-CoV-2 reinfection associated with emergence of omicron in South Africa. Science (2022) 376(6593):eabn4947. doi: 10.1126/science.abn4947
72. Altarawneh HN, Chemaitelly H, Hasan MR, Ayoub HH, Qassim S, AlMukdad S, et al. Protection against the omicron variant from previous SARS-CoV-2 infection. N Engl J Med (2022) 386(13):1288–90. doi: 10.1056/NEJMc2200133
73. Sheikh A, Kerr S, Woolhouse M, McMenamin J, Robertson C. EAVE II collaborators. Severity of omicron variant of concern and effectiveness of vaccine boosters against symptomatic disease in Scotland (EAVE II): a national cohort study with nested test-negative design. Lancet Infect Dis (2022) 22(7):959–66. doi: 10.1016/S1473-3099(22)00141-4
74. Madhi SA, Kwatra G, Myers JE, Jassat W, Dhar N, Mukendi CK, et al. Population immunity and COVID-19 severity with omicron variant in South Africa. N Engl J Med (2022) 386(14):1314–26. doi: 10.1056/NEJMoa2119658
75. Chemaitelly H, Tang P, Coyle P, Yassine HM, Al-Khatib HA, Smatti MK, et al. Protection against reinfection with the omicron BA.2.75 subvariant. N Engl J Med (2023) 388(7):665–67. doi: 10.1056/NEJMc2214114
76. World Health Organization. TAG-VE statement on omicron sublineages BQ.1 and XBB (2022). Available at: https://www.who.int/news/item/27-10-2022-tag-ve-statement-on-omicron-sublineages-bq.1-and-xbb. (Accessed 01 December 2022).
77. CoVariants. Overview of Variants/Mutations (2022). Available at: https://covariants.org/per-variant. (Accessed 10 August 2022).
78. Auvigne V, Vaux S, Strat YL, Schaeffer J, Fournier L, Tamandjou C, et al. Severe hospital events following symptomatic infection with SARS-CoV-2 omicron and delta variants in France, December 2021-January 2022: a retrospective, population-based, matched cohort study. EClinicalMedicine (2022) 48:101455. doi: 10.1016/j.eclinm.2022.101455
79. Veneti L, Boas H, Brathen Kristoffersen A, Stalcrantz J, Bragstad K, Hungnes O, et al. Reduced risk of hospitalisation among reported COVID-19 cases infected with the SARS-CoV-2 omicron BA.1 variant compared with the delta variant, Norway, December 2021 to January 2022. Euro Surveill (2022) 27(4):2200077. doi: 10.2807/1560-7917.ES.2022.27.4.2200077
80. Emani VR, Pallipuram VK, Goswami KK, Maddula KR, Reddy R, Nakka AS, et al. Increasing SARS-CoV2 cases, hospitalizations and deaths among the vaccinated elderly populations during the omicron (B.1.1.529) variant surge in UK. medRxiv (2022). doi: 10.1101/2022.06.28.22276926
81. Maslo C, Friedland R, Toubkin M, Laubscher A, Akaloo T, Kama B. Characteristics and outcomes of hospitalized patients in South Africa during the COVID-19 omicron wave compared with previous waves. JAMA (2022) 327(6):583–4. doi: 10.1001/jama.2021.24868
82. Modes ME, Directo MP, Melgar M, Johnson LR, Yang H, Chaudhary P, et al. Clinical characteristics and outcomes among adults hospitalized with laboratory-confirmed SARS-CoV-2 infection during periods of B.1.617.2 (Delta) and B.1.1.529 (Omicron) variant predominance - one hospital, California, July 15-September 23, 2021, and December 21, 2021-January 27, 2022. MMWR Morb Mortal Wkly Rep (2022) 71(6):217–23. doi: 10.15585/mmwr.mm7106e2
83. World Health Organization (WHO). Weekly epidemiological update on COVID-19 - 6 July 2022 (2022). Available at: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-COVID-19—6-july-2022. (Accessed 11 July 2022).
84. Lewnard JA, Hong V, Tartof SY. Association of SARS-CoV-2 BA.4/BA.5 omicron lineages with immune escape and clinical outcome. medRxiv (2022). doi: 10.1101/2022.07.31.22278258
85. World Health Organization. XBB.1.5 updated rapid risk assessment, 25 January 2023 (2023). Available at: https://www.who.int/docs/default-source/coronaviruse/25012023xbb.1.pdf?sfvrsn=c3956081_1. (Accessed 13 February 2023).
86. Madhi SA, Kwatra G, Myers JE, Jassat W, Dhar N, Mukendi CK, et al. SARS-CoV-2 infections during omicron (BA.1) dominant wave and subsequent population immunity in Gauteng, South Africa. medRxiv (2022). doi: 10.1101/2022.07.13.22277575
87. Elliott P, Eales O, Steyn N, Tang D, Bodinier B, Wang H, et al. Twin peaks: the omicron SARS-CoV-2 BA.1 and BA.2 epidemics in England. Science (2022) 376(6600):eabq4411. doi: 10.1126/science.abq4411
88. McMenamin ME, Nealon J, Lin Y, Wong JY, Cheung JK, Lau EHY, et al. Vaccine effectiveness of one, two, and three doses of BNT162b2 and CoronaVac against COVID-19 in Hong Kong: a population-based observational study. Lancet Infect Dis (2022) 22(10):1435–43. doi: 10.1016/S1473-3099(22)00345-0
89. Mefsin YM, Chen D, Bond HS, Lin Y, Cheung JK, Wong JY, et al. Epidemiology of infections with SARS-CoV-2 omicron BA.2 variant, Hong Kong, January-march 2022. Emerg Infect Dis (2022) 28(9):1856–58. doi: 10.3201/eid2809.220613
90. Lustig A, Vattiato G, Maclaren O, Watson LM, Datta S, Plank MJ. Modelling the impact of the omicron BA.5 subvariant in New Zealand. J R Soc Interface (2023) 20(199):20220698. doi: 10.1098/rsif.2022.0698
91. Bloomberg UK. China estimates Covid surge is infecting 37 million people a day (2022). Available at: https://www.bloomberg.com/news/articles/2022-12-23/china-estimates-COVID-surge-is-infecting-37-million-people-a-day. (Accessed 14 February 2023).
92. Malahe SRK, Hoek RAS, Dalm V, Broers AEC, den Hoed CM, Manintveld OC, et al. Clinical characteristics and outcome of immunocompromised patients with COVID-19 caused by the omicron variant: a prospective observational study. Clin Infect Dis (2022) 227(3):364–70. doi: 10.1093/cid/ciac571
93. Centers for Disease Control and Prevention (CDC). COVID data tracker: new hospital admissions (2022). Available at: https://COVID.cdc.gov/COVID-data-tracker/#new-hospital-admissions. (Accessed 16 June 2022).
94. Public Health England. SARS-CoV-2 variants of concern and variants under investigation in England: technical briefing 34 (2002). Available at: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1050236/technical-briefing-34-14-january-2022.pdf. (Accessed 16 June 2022).
95. Wang L, Berger NA, Kaelber DC, Davis PB, Volkow ND, Xu R. Incidence rates and clinical outcomes of SARS-CoV-2 infection with the omicron and delta variants in children younger than 5 years in the US. JAMA Pediatr (2022) 176(8):811–3. doi: 10.1001/jamapediatrics.2022.0945
96. Clarke KEN, Kim Y, Jones J, Lee A, Deng Y, Nycz E, et al. Pediatric infection-induced SARS-CoV-2 seroprevalence increases and seroprevalence by type of clinical care-September 2021-February 2022. J Infect Dis (2022) 227(3):364–70. doi: 10.1093/infdis/jiac423
97. Tso WWY, Kwan MYW, Wang YL, Leung LK, Leung D, Chua GT, et al. Severity of SARS-CoV-2 omicron BA.2 infection in unvaccinated hospitalized children: comparison to influenza and parainfluenza infections. Emerg Microbes Infect (2022) 11(1):1742–50. doi: 10.1080/22221751.2022.2093135
98. Nyberg T, Ferguson NM, Nash SG, Webster HH, Flaxman S, Andrews N, et al. Comparative analysis of the risks of hospitalisation and death associated with SARS-CoV-2 omicron (B.1.1.529) and delta (B.1.617.2) variants in England: a cohort study. Lancet (2022) 399(10332):1303–12. doi: 10.1016/S0140-6736(22)00462-7
99. Centers for Disease Control and Prevention (CDC). COVID data tracker: variant proportions (2023). Available at: https://COVID.cdc.gov/COVID-data-tracker/#variant-proportions. (Accessed 09 February 2023).
100. Wietecha M, Del Monte M. Open letter by Mark Wietecha, CEO of Children's Hospital Association and Mark Del Monte, JD, CEO of American Academy of Pediatrics (2022). Available at: https://downloads.aap.org/DOFA/Final%20AAP-CHA%20emergency%20declaration%20request%2011-14-22.pdf. (Accessed 09 February 2023).
101. Martin B, DeWitt PE, Russell S, Sanchez-Pinto LN, Haendel MA, Moffitt R, et al. Acute upper airway disease in children with the omicron (B.1.1.529) variant of SARS-CoV-2-A report from the US national COVID cohort collaborative. JAMA Pediatr (2022) 176(8):819–21. doi: 10.1001/jamapediatrics.2022.1110
102. Tun EM, Koid Jia Shin C, Usoro E, Thomas-Smith SE, Trehan I, Migita RT, et al. Croup during the coronavirus disease 2019 omicron variant surge. J Pediatr (2022) 247:147–49. doi: 10.1016/j.jpeds.2022.05.006
103. Holm M, Espenhain L, Glenthoj J, Schmidt LS, Nordly SB, Hartling UB, et al. Risk and phenotype of multisystem inflammatory syndrome in vaccinated and unvaccinated Danish children before and during the omicron wave. JAMA Pediatr (2022) 176(8):821–3. doi: 10.1001/jamapediatrics.2022.2206
104. Adhikari EH, MacDonald L, SoRelle JA, Morse J, Pruszynski J, Spong CY. COVID-19 cases and disease severity in pregnancy and neonatal positivity associated with delta (B.1.617.2) and omicron (B.1.1.529) variant predominance. JAMA (2022) 327(15):1500–2. doi: 10.1001/jama.2022.4356
105. Iacobucci G. COVID-19: staff absences are continuing to stretch NHS hospitals, say leaders. BMJ (2022) 376:o350. doi: 10.1136/bmj.o350
106. Lambert N, Survivor C, El-Azab SA, Ramrakhiani NS, Barisano A, Yu L, et al. The other COVID-19 survivors: timing, duration, and health impact of post-acute sequelae of SARS-CoV-2 infection. J Clin Nurs (2022). doi: 10.1111/jocn.16541
107. Temsah MH, Alenezi S, Alarabi M, Aljamaan F, Alhasan K, Assiri R, et al. Healthcare workers' SARS-CoV-2 omicron variant uncertainty-related stress, resilience, and coping strategies during the first week of the world health organization's alert. Int J Environ Res Public Health (2022) 19(4):1944. doi: 10.3390/ijerph19041944
108. Euractiv. With peak yet to come, Europe’s healthcare groans under omicron’s swift spread. Available at: https://www.euractiv.com/section/coronavirus/news/with-peak-yet-to-come-europes-healthcare-groans-under-omicrons-swift-spread/. (Accessed 17 June 2022).
109. Tang J, Zeng C, Cox TM, Li C, Son YM, Cheon IS, et al. Respiratory mucosal immunity against SARS-CoV-2 following mRNA vaccination. Sci Immunol (2022) 7(76):eadd4853. doi: 10.1126/sciimmunol.add4853
110. Altarawneh HN, Chemaitelly H, Ayoub HH, Tang P, Hasan MR, Yassine HM, et al. Effects of previous infection and vaccination on symptomatic omicron infections. N Engl J Med (2022) 387(1):21–34. doi: 10.1056/NEJMoa2203965
111. Zaballa M-E, Perez-Saez J, de Mestral C, Pullen N, Lamour J, Turelli P, et al. Seroprevalence of anti-SARS-CoV-2 antibodies and cross-variant neutralization capacity after the omicron BA.2 wave in Geneva, Switzerland. medRxiv (2022). doi: 10.1101/2022.07.27.22278126
112. Bellusci L, Grubbs G, Zahra FT, Forgacs D, Golding H, Ross TM, et al. Antibody affinity and cross-variant neutralization of SARS-CoV-2 omicron BA.1, BA.2 and BA.3 following third mRNA vaccination. Nat Commun (2022) 13(1):4617. doi: 10.1038/s41467-022-32298-w
113. De Marco L, D'Orso S, Pirronello M, Verdiani A, Termine A, Fabrizio C, et al. Assessment of T-cell reactivity to the SARS-CoV-2 omicron variant by immunized individuals. JAMA Netw Open (2022) 5(4):e2210871. doi: 10.1001/jamanetworkopen.2022.10871
114. Keeton R, Tincho MB, Ngomti A, Baguma R, Benede N, Suzuki A, et al. T cell responses to SARS-CoV-2 spike cross-recognize omicron. Nature (2022) 603(7901):488–92. doi: 10.1038/s41586-022-04460-3
115. Liu J, Chandrashekar A, Sellers D, Barrett J, Jacob-Dolan C, Lifton M, et al. Vaccines elicit highly conserved cellular immunity to SARS-CoV-2 omicron. Nature (2022) 603(7901):493–6. doi: 10.1038/s41586-022-04465-y
116. Peng Q, Zhou R, Wang Y, Zhao M, Liu N, Li S, et al. Waning immune responses against SARS-CoV-2 variants of concern among vaccinees in Hong Kong. EBioMedicine (2022) 77:103904. doi: 10.1016/j.ebiom.2022.103904
117. Andrews N, Stowe J, Kirsebom F, Toffa S, Rickeard T, Gallagher E, et al. COVID-19 vaccine effectiveness against the omicron (B.1.1.529) variant. N Engl J Med (2022) 386(16):1532–46. doi: 10.1056/NEJMoa2119451
118. Tseng HF, Ackerson BK, Luo Y, Sy LS, Talarico CA, Tian Y, et al. Effectiveness of mRNA-1273 against SARS-CoV-2 omicron and delta variants. Nat Med (2022) 28(5):1063–71. doi: 10.1038/s41591-022-01753-y
119. Kirsebom FCM, Andrews N, Stowe J, Toffa S, Sachdeva R, Gallagher E, et al. COVID-19 vaccine effectiveness against the omicron (BA.2) variant in England. Lancet Infect Dis (2022) 22(7):931–3. doi: 10.1016/S1473-3099(22)00309-7
120. Richterman A, Behrman A, Brennan PJ, O'Donnell JA, Snider CK, Chaiyachati KH. Durability of severe acute respiratory syndrome coronavirus 2 messenger RNA booster vaccine protection against Omicron among healthcare workers with a vaccine mandate. Clin Infect Dis (2023) 76(3):e319–26. doi: 10.1093/cid/ciac454
121. Chemaitelly H, Ayoub HH, AlMukdad S, Coyle P, Tang P, Yassine HM, et al. Duration of mRNA vaccine protection against SARS-CoV-2 omicron BA.1 and BA.2 subvariants in Qatar. Nat Commun (2022) 13(1):3082. doi: 10.1038/s41467-022-30895-3
122. Buchan SA, Chung H, Brown KA, Austin PC, Fell DB, Gubbay JB, et al. Estimated effectiveness of COVID-19 vaccines against omicron or delta symptomatic infection and severe outcomes. JAMA Netw Open (2022) 5(9):e2232760. doi: 10.1001/jamanetworkopen.2022.32760
123. Lauring AS, Tenforde MW, Chappell JD, Gaglani M, Ginde AA, McNeal T, et al. Clinical severity of, and effectiveness of mRNA vaccines against, COVID-19 from omicron, delta, and alpha SARS-CoV-2 variants in the United States: prospective observational study. BMJ (2022) 376:e069761. doi: 10.1136/bmj-2021-069761
124. Tenforde MW, Self WH, Gaglani M, Ginde AA, Douin DJ, Talbot HK, et al. Effectiveness of mRNA vaccination in preventing COVID-19-associated invasive mechanical ventilation and death - United States, March 2021-January 2022. MMWR Morb Mortal Wkly Rep (2022) 71(12):459–65. doi: 10.15585/mmwr.mm7112e1
125. Tartof SY, Slezak JM, Puzniak L, Hong V, Frankland TB, Ackerson BK, et al. Effectiveness of a third dose of BNT162b2 mRNA COVID-19 vaccine in a large US health system: a retrospective cohort study. Lancet Reg Health Am (2022) 9:100198. doi: 10.1016/j.lana.2022.100198
126. Collie S, Nayager J, Bamford L, Bekker LG, Zylstra M, Gray G. Effectiveness and durability of the BNT162b2 vaccine against omicron sublineages in South Africa. N Engl J Med (2022) 387(14):1332–3. doi: 10.1056/NEJMc2210093
127. Tartof SY, Slezak JM, Puzniak L, Hong V, Frankland TB, Ackerson BK, et al. BNT162b2 vaccine effectiveness against SARS-CoV-2 omicron BA.4 and BA.5. Lancet Infect Dis (2022) 22(12):1663–5. doi: 10.1016/S1473-3099(22)00692-2
128. Kislaya I, Casaca P, Borges V, Sousa C, Ferreira BI, Fernandes E, et al. SARS-CoV-2 BA.5 vaccine breakthrough risk and severity compared with BA.2: a case-case and cohort study using electronic health records in Portugal. medRxiv (2022). doi: 10.1101/2022.07.25.22277996
129. Tartof SY, Frankland TB, Puzniak L, Slezak JM, Hong V, Takhar H, et al. BNT162b2 against COVID-19-associated emergency department and urgent care visits among children 5-11 years of age: a test negative design. J Pediatr Infect Dis Soc (2023) 12(3):177–79. doi: 10.1093/jpids/piad005
130. Ferdinands JM, Rao S, Dixon BE, Mitchell PK, DeSilva MB, Irving SA, et al. Waning 2-dose and 3-dose effectiveness of mRNA vaccines against COVID-19-associated emergency department and urgent care encounters and hospitalizations among adults during periods of delta and omicron variant predominance - VISION network, 10 states, August 2021-January 2022. MMWR Morb Mortal Wkly Rep (2022) 71(7):255–63. doi: 10.15585/mmwr.mm7107e2
131. Moller Kirsebom FC, Andrews N, Stowe J, Groves N, Chand M, Ramsay M, et al. Effectiveness of the COVID-19 vaccines against hospitalisation with omicron sub-lineages BA.4 and BA.5 in England. Lancet Reg Health Eur (2022) 23:100537. doi: 10.1016/j.lanepe.2022.100537
132. Bar-On YM, Goldberg Y, Mandel M, Bodenheimer O, Amir O, Freedman L, et al. Protection by a fourth dose of BNT162b2 against omicron in Israel. N Engl J Med (2022) 386(18):1712–20. doi: 10.1056/NEJMoa2201570
133. Cohen MJ, Oster Y, Moses AE, Spitzer A, Benenson S. Israeli-Hospitals 4th vaccine working group. association of receiving a fourth dose of the BNT162b vaccine with SARS-CoV-2 infection among health care workers in Israel. JAMA Netw Open (2022) 5(8):e2224657. doi: 10.1001/jamanetworkopen.2022.24657
134. Grewal R, Kitchen SA, Nguyen L, Buchan SA, Wilson SE, Costa AP, et al. Effectiveness of a fourth dose of COVID-19 mRNA vaccine against the omicron variant among long term care residents in Ontario, Canada: test negative design study. BMJ (2022) 378:e071502. doi: 10.1136/bmj-2022-071502
135. Nemet I, Kliker L, Lustig Y, Zuckerman N, Erster O, Cohen C, et al. Third BNT162b2 vaccination neutralization of SARS-CoV-2 omicron infection. N Engl J Med (2022) 386(5):492–4. doi: 10.1056/NEJMc2119358
136. Zou J, Kurhade C, Patel S, Kitchin N, Tompkins K, Cutler M, et al. Neutralization of BA.4–BA.5, BA.4.6, BA.2.75.2, BQ.1.1, and XBB.1 with bivalent vaccine. New Engl J Med (2023) 388(9):854–57. doi: 10.1056/NEJMc2214916
137. Pfizer, BioNTech. Pfizer/BioNTech COVID-19 omicron-modified vaccine options (2022). Available at: https://www.fda.gov/media/159496/download. (Accessed 3 October 2022).
138. Vardhana S, Baldo L, Morice WG, Wherry EJ. Understanding T cell responses to COVID-19 is essential for informing public health strategies. Sci Immunol (2022) 7(71):eabo1303. doi: 10.1126/sciimmunol.abo1303
139. European Medicines Agency. Spikevax. Summary of product characteristics (2022). Available at: https://www.ema.europa.eu/en/medicines/human/EPAR/spikevax. (Accessed 13 October 2022).
140. European Medicines Agency. Comirnaty COVID-19 vaccine. Summary of product characteristics (2022). Available at: https://www.ema.europa.eu/en/documents/product-information/comirnaty-epar-product-information_en.pdf. (Accessed 07 September 2022).
141. United States Food and Drug Administration (FDA). Coronavirus (COVID-19) update: FDA recommends inclusion of omicron BA.4/5 component for COVID-19 vaccine booster doses (2022). Available at: https://www.fda.gov/news-events/press-announcements/coronavirus-COVID-19-update-fda-recommends-inclusion-omicron-ba45-component-COVID-19-vaccine-booster. (Accessed 12 July 2022).
142. United States Centers for Disease Control and Prevention. CDC Fall vaccination operational planning guide – information for the fall vaccine campaign, including upcoming bivalent COVID-19 vaccine booster doses (2022). Available at: https://www.cdc.gov/vaccines/COVID-19/downloads/CDC-Fall-Vaccination-Operational-Planning-Guide.pdf. (Accessed 16 August 2022).
143. U.S. Food and Drug Administration. Emergency use authorization for vaccines to prevent COVID-19: guidance for industry (2022). Available at: https://www.fda.gov/media/142749/download (Accessed 12 July 2022).
144. European Medicines Agency. Reflection paper on the regulatory requirements for vaccines intended to provide protection against variant strain(s) of SARS-CoV-2 (2021). Available at: https://www.ema.europa.eu/en/documents/scientific-guideline/reflection-paper-regulatory-requirements-vaccines-intended-provide-protection-against-variant_en.pdf. (Accessed 23 February 2021).
145. US Food & Drug Administration. Coronavirus (COVID-19) update: FDA authorizes Moderna and Pfizer-BioNTech bivalent COVID-19 vaccines for use as a booster dose in younger age groups (2022). Available at: https://www.fda.gov/news-events/press-announcements/coronavirus-COVID-19-update-fda-authorizes-moderna-and-pfizer-biontech-bivalent-COVID-19-vaccines. (Accessed 13 October 2022).
146. Lasrado N, Collier A-r, Miller J, Hachmann NP, Liu J, Sciacca M, et al. Waning immunity against XBB.1.5 following bivalent mRNA boosters. bioRxiv (2023). doi: 10.1101/2023.01.22.525079
147. Prevention CfDCa. Rates of COVID-19 cases and deaths by vaccination status (2023). Available at: https://COVID.cdc.gov/COVID-data-tracker/#rates-by-vaccine-status. (Accessed 03 February 2023).
148. Link-Gelles R, Ciesla AA, Roper LE, Scobie HM, Ali AR, Miller JD, et al. Early estimates of bivalent mRNA booster dose vaccine effectiveness in preventing symptomatic SARS-CoV-2 infection attributable to omicron BA.5- and XBB/XBB.1.5-related sublineages among immunocompetent adults - increasing community access to testing program, United States, December 2022-January 2023. MMWR Morb Mortal Wkly Rep (2023) 72(5):119–24. doi: 10.15585/mmwr.mm7205e1
149. Vaccines and Related Biological Products Advisory Committee. Vaccines and related biological products advisory committee meeting January 26, 2023 (2023). Available at: https://www.fda.gov/media/165307/download. (Accessed 29 March 2023).
150. European Medicines Agency. ETF statement on the use of the EMA approved bivalent original/Omicron BA.4-5 mRNA vaccines for primary series (2022). Available at: https://www.ema.europa.eu/en/documents/other/etf-statement-use-ema-approved-bivalent-original/omicron-ba4-5-mrna-vaccines-primary-series_en.pdf. (Accessed 07 December 2022).
151. Stokel-Walker C. How are vaccines being adapted to meet the changing face of SARS-CoV-2? BMJ (2022) 377:o1257. doi: 10.1136/bmj.o1257
152. Morens DM, Taubenberger JK, Fauci AS. Universal coronavirus vaccines - an urgent need. N Engl J Med (2022) 386(4):297–9. doi: 10.1056/NEJMp2118468
153. Altmann DM, Boyton RJ. COVID-19 vaccination: the road ahead. Science (2022) 375(6585):1127–32. doi: 10.1126/science.abn1755
154. Dai L, Gao GF. Viral targets for vaccines against COVID-19. Nat Rev Immunol (2021) 21(2):73–82. doi: 10.1038/s41577-020-00480-0
155. Dolgin E. Pan-coronavirus vaccine pipeline takes form. Nat Rev Drug Discovery (2022) 21(5):324–6. doi: 10.1038/d41573-022-00074-6
156. Pfizer Inc, BioNTech SE. Pfizer and BioNTech advance COVID-19 vaccine strategy with study start of next-generation vaccine candidate based on enhanced spike protein Design (2022). Available at: https://www.pfizer.com/news/announcements/pfizer-and-biontech-advance-COVID-19-vaccine-strategy-study-start-next. (Accessed 27 July 2022).
157. BioNTech. Pfizer and BioNTech advance next-generation COVID-19 vaccine strategy with study start of candidate aimed at enhancing breadth of T cell responses and duration of Protection (2022). Available at: https://investors.biontech.de/news-releases/news-release-details/pfizer-and-biontech-advance-next-generation-COVID-19-vaccine. (Accessed 29 March 2023).
158. Tomalka JA, Suthar MS, Deeks SG, Sekaly RP. Fighting the SARS-CoV-2 pandemic requires a global approach to understanding the heterogeneity of vaccine responses. Nat Immunol (2022) 23(3):360–70. doi: 10.1038/s41590-022-01130-4
159. Chavda VP, Vora LK, Pandya AK, Patravale VB. Intranasal vaccines for SARS-CoV-2: from challenges to potential in COVID-19 management. Drug Discov Today (2021) 26(11):2619–36. doi: 10.1016/j.drudis.2021.07.021
Keywords: sub-lineage, BA.1, vaccine, disease burden, Omicron
Citation: Pather S, Madhi SA, Cowling BJ, Moss P, Kamil JP, Ciesek S, Muik A and Türeci Ö (2023) SARS-CoV-2 Omicron variants: burden of disease, impact on vaccine effectiveness and need for variant-adapted vaccines. Front. Immunol. 14:1130539. doi: 10.3389/fimmu.2023.1130539
Received: 23 December 2022; Accepted: 05 May 2023;
Published: 23 May 2023.
Edited by:
Daniela Hozbor, Institute of Biotechnology and Molecular Biology (IBBM), ArgentinaReviewed by:
Hayley Gans, Stanford University, United StatesCopyright © 2023 Pather, Madhi, Cowling, Moss, Kamil, Ciesek, Muik and Türeci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shanti Pather, U2hhbnRpLlBhdGhlckBiaW9udGVjaC5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.