
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 13 March 2023
Sec. T Cell Biology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1124018
This article is part of the Research Topic Women in T Cell Biology View all 16 articles
 Lídia Sans-de San Nicolàs1
Lídia Sans-de San Nicolàs1 Ignasi Figueras-Nart2
Ignasi Figueras-Nart2 Irene García-Jiménez1
Irene García-Jiménez1 Montserrat Bonfill-Ortí2Antonio Guilabert3
Montserrat Bonfill-Ortí2Antonio Guilabert3 Laia Curto-Barredo4Marta Bertolín-Colilla4Marta Ferran4Esther Serra-Baldrich5
Laia Curto-Barredo4Marta Bertolín-Colilla4Marta Ferran4Esther Serra-Baldrich5 Ramon M. Pujol4
Ramon M. Pujol4 Luis F. Santamaria-Babí1*
Luis F. Santamaria-Babí1*Background: The role of allergen sensitization in IL-31 production by T cells and specifically in the clinical context of atopic dermatitis (AD) has not been characterized.
Methods: The response to house dust mite (HDM) in purified memory T cells cocultured with epidermal cells from AD patients (n=58) and control subjects (n=11) was evaluated. AD-associated cytokines from culture supernatants, plasma proteins and mRNA expression from cutaneous lesions were assessed and related with the clinical features of the patients.
Results: HDM-induced IL-31 production by memory T cells defined two subsets of AD patients according to the presence or absence of IL-31 response. Patients in the IL-31 producing group showed a more inflammatory profile, and increased HDM-specific (sp) and total IgE levels compared to the IL-31 non-producing group. A correlation between IL-31 production and patient’s pruritus intensity, plasma CCL27 and periostin was detected. When the same patients were analyzed based on sp IgE and total IgE levels, an increased IL-31 in vitro response, as well as type 2 markers in plasma and cutaneous lesions, was found in patients with sp IgE levels > 100 kUA/L and total IgE levels > 1000 kU/L. The IL-31 response by memory T cells was restricted to the cutaneous lymphocyte-associated antigen (CLA)+ T-cell subset.
Conclusion: IgE sensitization to HDM allows stratifying IL-31 production by memory T cells in AD patients and relating it to particular clinical phenotypes of the disease.
IL-31 is a clinically relevant neuroimmune cytokine involved in epidermal barrier disruption, pruritus, inflammation and tissue remodeling in atopic dermatitis (AD) (1–3). It is enhanced in the sera from AD patients and correlates with disease severity (4).
The major source of IL-31 are CD4+ T cells associated to a Th2 phenotype (5–8), but it is also produced by dendritic cells, mast cells, group 2 innate lymphoid cells, basophils, eosinophils or M2 macrophages (9–14). Its receptor, IL-31RA, is expressed by multiple cell types, including skin sensory neurons, keratinocytes and immune cells, activating the JAK/STAT, PI3K/AKT and MAPK/JNK pathways (9, 15, 16).
IL-31 and IL-31RA expression is increased in the AD skin and are part of the “core” signature characterized by the activation of the IL-31/IL-1 signaling pathway (17). The epidermal barrier function is affected by IL-31 modulation of keratinocyte differentiation directly or via other mediators including IL-20, IL-24, IL-33 and IL-1α (18–23). IL-31 binding to its receptor in sensory neurons stimulates itch signaling, neuronal branching and increases of cutaneous nerve fiber density (9, 24). A clear improvement of pruritus has been observed after IL-31RA blockade in moderate-to-severe AD patients (25–27). IL-31 initiates and maintains inflammation by promoting the release of proinflammatory cytokines and chemokines by myeloid cells, eosinophils, basophils and keratinocytes (10, 15, 18–20, 28–31).
Aeroallergens such as house dust mite (HDM) are associated to disease severity and total IgE levels in AD (32, 33). The relationship between IL-31 and allergens has been poorly characterized and mainly limited to canine models: HDM-sensitized dogs showed IL-31 expression in CD3+ CD4+ T cells in the skin lesions and production of IL-31 by Th2-polarized peripheral blood mononuclear cells (PBMC) stimulated with HDM (7, 34). Non-lesional skin from AD patients challenged with HDM induced IL-31 mRNA expression in three out of six HDM allergic patients (21). Increased frequencies of IL-31+ CD4+ and CD8+ T cells in PBMC from HDM-sensitized patients have also been detected (35). Nevertheless, no studies have addressed the role of allergen in IL-31 production by memory T cells and its possible relationship with particular clinical features in AD patients.
In this study, HDM-induced IL-31 by cutaneous lymphocyte-associated antigen (CLA)+ memory T cells correlated with patient’s pruritus. Patients with elevated HDM-specific (sp) IgE (> 100 kUA/L) and total IgE levels (> 1000 kU/L) showed an increased IL-31 production by memory T cells compared to patients with lower sp and total IgE levels. These results show for the first time a relationship between allergen-specific T-cell-mediated IL-31 production, clinical status of the patients and the levels of allergen sensitization that can be of help to guide patient’s identification in response to IL-31-directed therapies.
Peripheral blood and two skin biopsies from active lesional areas were collected from 58 consented moderate-to-severe AD patients and 11 consented controls under institutional review board-approved protocols at the Hospital de Bellvitge, Hospital General de Granollers, Hospital del Mar and Hospital de la Santa Creu I Sant Pau (Spain). Exclusion criteria included topical or systemic anti-inflammatories for the last 2 or 4 weeks prior to the study, respectively.
Plasma samples were assessed for total IgE (kU/L) and HDM-specific IgE (response (OD) and kUA/L) by ImmunoCAP (Thermo Fisher Scientific, Waltham, MA, USA). Serum samples were used for lactate dehydrogenase (LDH) measurement (U/L) in a diagnostic laboratory. Patients’ characteristics are summarized in Table S1.
Central and effector memory CD45RA- T lymphocytes were purified from PBMC after Ficoll (GE Healthcare, Princeton, NJ, USA) gradient and three consecutive immunomagnetic separations (Miltenyi Biotech, Bergisch Gladbach, Germany) (36). First CD14+ and CD19+ cells were depleted, then CD16+ and CD45RA+ lymphocytes were depleted, and finally CD45RA- T cells were divided into CLA+ and CLA- T-cell subpopulations using the Miltenyi product ref 130-092-464. Sample purity was >95% of CLA-positivity for the CLA+ T-cell subpopulation and <10% of CLA-positivity for the CLA- T-cell subpopulation (36). Terminal differentiated memory TEMRA cells were not studied with this approach because of CD45RA+ cells depletion.
Punch skin biopsy samples were incubated in dispase solution (Corning, Corning, NY, USA) overnight at 4°C, and the epidermal sheet was peeled off from the dermis. The epidermis was cut into pieces and incubated with trypsin (Biological Industries, Kibbutz Beit Haemek, Israel) for 15 minutes at 37°C. Then, the epidermal tissue was mechanically disaggregated by pipetting and the cell suspension was transferred to fresh culture media (1:1 volume) (RPMI supplemented with 10% FBS (Thermo Fisher Scientific) and 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA)). Finally, the epidermal cells suspension (Epi) was obtained by means of centrifugation. It contained all cell types at the same proportion present in the epidermis, being keratinocytes the major cell type.
The coculture system was performed by seeding 3 x 104 autologous epidermal cells with 5 x 104 circulating CLA+ or CLA- memory T cells (CLA+/Epi or CLA-/Epi, respectively) in a 96-well U-bottom plate (Falcon, Corning, Corning, NY, USA), in the culture media described above. Whole (CD4+ and CD8+) CLA+ memory T cells were used; however, we assume that results obtained are due to CD4+CLA+ memory T cells, since the majority of CLA+ memory T cells are CD4+ (80% CD4+ and 20% CD8+) and IL-31 is mainly produced by CD4+CLA+ T cells (6, 23). Cocultures were left untreated (M) or activated for 5 days with HDM extract kindly provided by LETI Pharma at 10 µg/mL or 24 hours with staphylococcal enterotoxin B (SEB) (Sigma-Aldrich) at 100 ng/mL final well concentration (37). In cultures containing only T cells or epidermal cells, the mentioned amounts of each cell type were used, and activation with HDM was performed in the same way. Collected supernatants were kept at -20°C.
For blocking assays, HLA-A/B/C (class I) (BioLegend, San Diego, CA, USA), HLA-DR (class II) (BioLegend), CD1a (Bio X Cell, Lebanon, NH, USA), IL-33 (R & D Systems, Minneapolis, Minnesota, USA) neutralizing antibodies, or respective mouse IgG2a isotype control (BioLegend), mouse IgG1 (BioLegend) and goat IgG isotype control (R & D Systems) were added to cocultures at 10 µg/mL (for HLA and CD1a blocking assays) or 1 µg/mL (for IL-33 blocking assay) final concentration.
5 x 104 purified CLA+ or CLA- memory T cells were added to the top chamber of 0.4 µm pore size polycarbonate 96-well HTS Transwell culture insert (Corning) containing 3 x 104 autologous lesional epidermal cells stimulated with HDM at 10 µg/mL final well concentration in the bottom chamber. Supernatants were collected at day 5 of culture.
ProcartaPlex immunoassay (Invitrogen, Waltham, MA, USA) was used to measure IL-31, IL-13, IL-4, IL-5, IL-17A, IL-22 and IFN-γ concentrations in the culture supernatants with the MAGPIX instrument (Invitrogen) and analysed with ProcartaPlex Analyst software version 1.0 (Invitrogen). Values below the lower limit of quantification (LLOQ) were treated as zero. Plasma CCL22 levels were quantified with ProcartaPlex.
Pre-coated ELISA kits were used for quantification of plasma levels of CCL27, CCL17, CCL18, sIL-2R (Invitrogen) and periostin (AdipoGen Life Sciences, San Diego, CA, USA).
Lesional skin biopsy specimens frozen in Tissue-Tek O.C.T. Compound (Sakura Finetek, Alphen aan den Rijn, The Netherlands) at -80°C were used for RNA isolation by using the TRIzol Reagent (Invitrogen).
For qRT-PCR, cDNA was obtained with the High Capacity cDNA Reverse Transcription kit (Applied Biosystems, Waltham, MA, USA) and preamplified with the TaqMan PreAmp Master Mix (2x) (Applied Biosystems). Taqman Gene Expression Master Mix and FAM-labelled probes (Table S2) (Applied Biosystems) were used for qRT-PCR with an ABI Prism 7900HT instrument (Applied Biosystems). Data was processed by SDS analysis software version 2.4.1 (Applied Biosystems) and gene expression was calculated by using the Δ - Δ cycle threshold (Ct) method (with the mean cycle threshold value for RPLP0 and the gene of interest for each sample). The equation 1.8e (CtRPLP0 - Ctgene of interest) x 104 was used for normalizing the values (38).
For gene-array analysis, quality control of concentration and integrity of the isolated RNA was performed with the NanoDrop One (Thermo Fisher Scientific) and the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). PrimeView Human Gene Expression Arrays (Applied Biosystems) were processed at the Functional Genomics Facility of IRB Barcelona (Barcelona, Spain) and raw data (CEL files) were processed with Transcriptome Analysis Console version 4.0 (Applied Biosystems). Raw data were deposited in the Gene Expression Omnibus repository, accession number GSE226073. Genes with a fold change (FCH) of 1.5 or greater and a p value of less than .05 were considered differentially expressed genes (DEG). Pathway enrichment analysis was performed using g:Profiler web server (https://biit.cs.ut.ee/gprofiler/gost) with Gene Ontology (GO) biological process database and a Benjamini-Hochberg FDR of .05 or greater.
Data analysis and representation were performed with GraphPad Prism software version 8 (GraphPad Software Corporation, San Diego, CA, USA). Data are represented as the median ± 95% confidence interval (CI). Wilcoxon test was used to compare two conditions within the same group and Mann-Whitney test was used to compare two groups. In tables, sample median (25th-75th percentiles) and Mann-Whitney test were used for continuous variables, raw numbers (percentages) and Fisher’s exact test were used for categorical variables, and bold values indicate significant data. Correlations were examined using Spearman coefficient and represented with linear regression. Differences were considered significant at a P-value of less than .05 and showed as: (*) p <.05; (**) p <.01; (***) p <.001; (****) p <.0001.
AD-derived cocultures containing circulating memory CLA+ T cells and autologous lesional epidermal cells (CLA+/Epi) in the presence of HDM led to IL-31 production, whereas CLA- T-cell AD cocultures (CLA-/Epi) and control (C)-derived cocultures did not produced IL-31 upon stimulation (Figure 1A). Epidermal cells suspension promoted HDM-induced CLA+ T-cell-derived IL-31 response (Figure S1A) and it was produced in a time-dependent manner (Figure S1B).
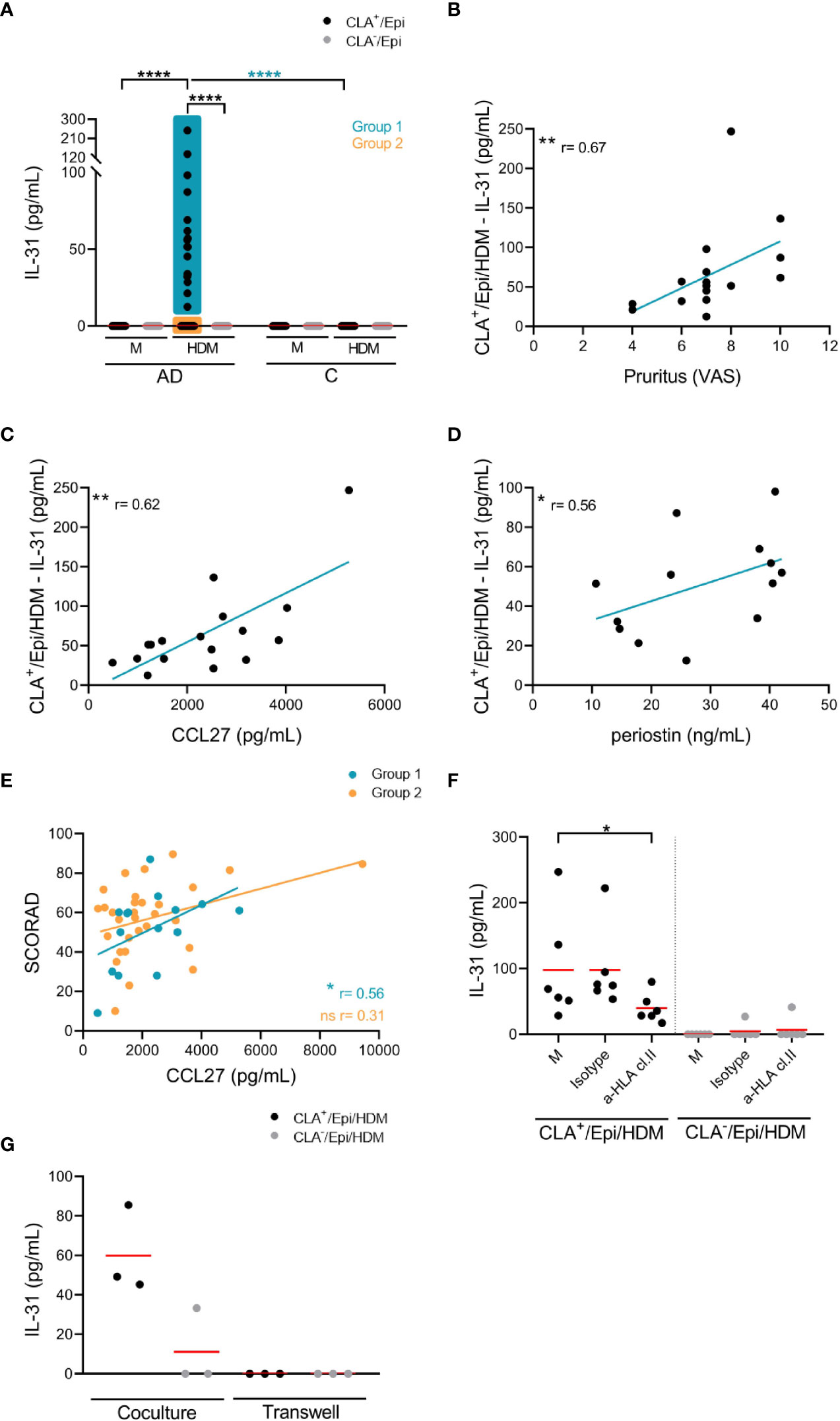
Figure 1 HDM-induced IL-31 by memory T cells correlates with patient’s pruritus and plasma CCL27 and periostin, partially depends on HLA class II molecules and the cell-cell contact with epidermal cells, and is restricted to CLA+ T-cells subset. (A) IL-31 (pg/mL) produced by CLA+/- T-cell cocultures in basal conditions (M) or stimulated with HDM in AD-(n=58) and C-(n=11) derived samples. Correlations of CLA+/Epi/HDM IL-31 production in group 1 with (B) pruritus (n=17), and plasma (C) CCL27 (n=17) and (D) periostin (n=13) levels. (E) Correlation between plasma CCL27 levels and SCORAD in group 1 (n=15) and group 2 (n=31) of patients. (F) HDM-activated CLA+/- T-cell cocultures were treated with HLA class II neutralizing antibody or control IgG isotype at day 0, and IL-31 production (pg/mL) at day 5 was compared with respect to isotype values (n=6). Red bar indicates the mean. (G) CLA+/- T cells were added in the upper chamber and HDM-stimulated epidermal cells suspension in the lower chamber of the 96-well transwell plate and IL-31 (pg/mL) was measured at day 5. Data for three representative patients. AD, atopic dermatitis; C, control subjects; CLA, cutaneous lymphocyte-associated antigen T cells; Epi, epidermal cells suspension; HDM, house dust mite; M, untreated; SCORAD, scoring atopic dermatitis. ns: p >.05; *p <.05; **p <.01; ****p <.0001.
AD patients were classified into two groups to further characterize clinical profiles based on the differential response to HDM: those producing IL-31 by CLA+ T cells were arranged into group 1 (n=17), and those with no IL-31 production were arranged into group 2 (n=41) (Figure 1A).
Within group 1 of patients, HDM-induced IL-31 response directly correlated with pruritus (r = 0.67, p = .0036; Figure 1B) and plasma levels of CCL27 (r = 0.62, p = .0090; Figure 1C) and periostin (r = 0.56, p = .050; Figure 1D). Correlations within group 2 of patients could not be performed due to undetectable IL-31 production in the cocultures. As shown in Figure 1E, there was direct correlation between plasma CCL27 levels and SCORAD in group 1 (r = 0.56, p = .032) but not in group 2 (r = 0.31, p = .091). Additionally, HDM-induced CLA+ T-cell IL-31 nearly correlated with SCORAD in group 1 of patients (r = 0.49, p = .066; not shown). There was no correlation between IL-31 in vitro response to HDM and plasma levels of HDM-specific (sp) IgE and total IgE, although there was a tendency for the former (r = 0.46, p = .068; Figure S2).
The IL-31 production by CLA+ memory T cells was blocked by 59% by a neutralizing HLA class II, but not HLA class I, antibody (Figures 1F; S3A). Blocking of IL-33 and CD1a molecules did not affect IL-31 production (Figures S3B, C), but cell-cell contact between CLA+ T cells and epidermal cells was necessary for IL-31 production as demonstrated by transwell cultures (Figure 1G).
IL-31 was simultaneously measured with other AD-derived mediators (IL-13, IL-4, IL-5, IL-17A, IL-22 and IFN-γ) and we found that IL-13, IL-4, IL-5, IL-17A and IL-22 production by HDM-induced CLA+ T-cell AD cocultures was higher than that by C cocultures (Figure S4A). HDM-activated CLA+/Epi cocultures from group 1 showed increased production of IL-13, IL-4, IL-5, IL-17A and IL-22 compared to group 2 (Figures 2A; S4B), revealing a more inflammatory phenotype in group 1 versus group 2. Also, in group 1 IL-31 production directly correlated with IL-13 (r = 0.62, p = .0093) and IL-4 (r = 0.85, p <.0001), but no correlation with IL-5, IL-17A, IL-22 and IFN-γ was observed (Figure 2B). Although group 2 of patients were defined for their null production of IL-31 by HDM-stimulated CLA+ T cells, there was an heterogeneous IL-13, IL-4, IL-5, IL-17A, IL-22 and IFN-γ response in the CLA+ T-cell cocultures of this group (Figure S5). All correlations were tested with and without outlier patient data.
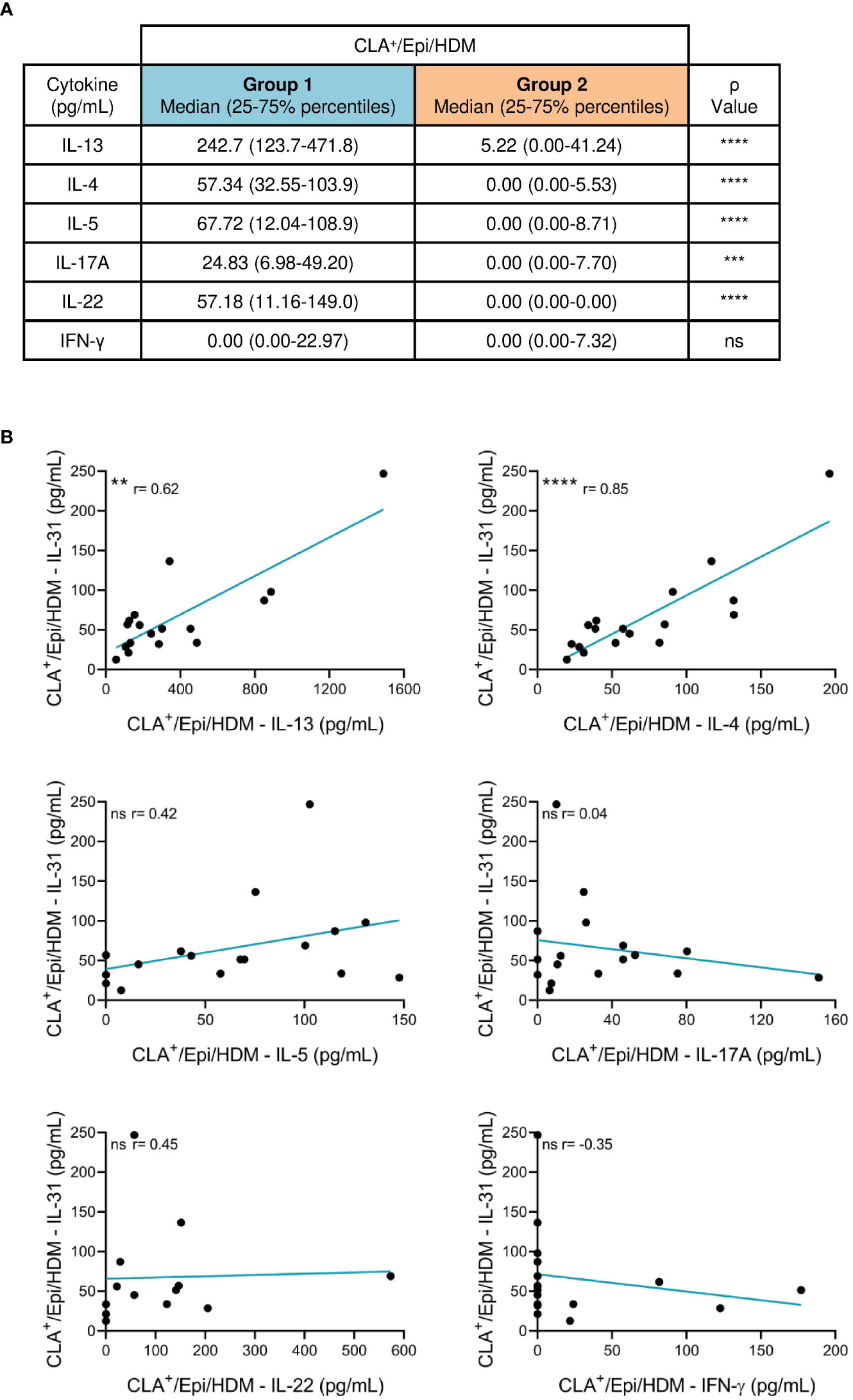
Figure 2 Patients producing IL-31 (group 1) by HDM-stimulated CLA+ T cells have a more inflammatory phenotype than patients with undetectable IL-31 (group 2). (A) IL-13, IL-4, IL-5, IL-17A, IL-22 and IFN-γ (pg/mL) produced by HDM-induced CLA+ T-cell cocultures was compared between groups 1 (n=13-17) and 2 (n=35-41) of patients. (B) In group 1, HDM-stimulated CLA+ T-cell-dependent IL-31 production was correlated with IL-13 (n=17), IL-4 (n=17), IL-5 (n=17), IL-17A (n=17), IL-22 (n=13) and IFN-γ (n=17). CLA, cutaneous lymphocyte-associated antigen T cells; Epi, epidermal cells suspension; HDM, house dust mite. ns: p >.05; **p <.01; ***p <.001; ****p <.0001.
When stimulating cocultures with Staphylococcus aureus superantigen B (SEB), another clinically relevant stimulus of AD, IL-31 was specifically produced by CLA+ T cells but no significant differences were found between group 1 and group 2 of patients (Figure S6A). Additionally, therewas no correlation between HDM- and SEB-induced IL-31 production by memory T cells (Figure S6B).
Patients from group 1 displayed increased sp IgE and total IgE levels compared with patients from group 2 (Table 1). No differences were found for severity, pruritus, eosinophilia, years since diagnosis and comorbidities. Although group 2 of patients had a reduced allergen sensitization degree when compared to group 1 of patients, they showed higher total IgE and sp IgE levels than controls (total IgE (kU/L): group 2 median = 568.00, C median = 21.10, p <.0001; sp IgE response (OD): group 2 median = 5338, C median = 27.00, p <.0001).
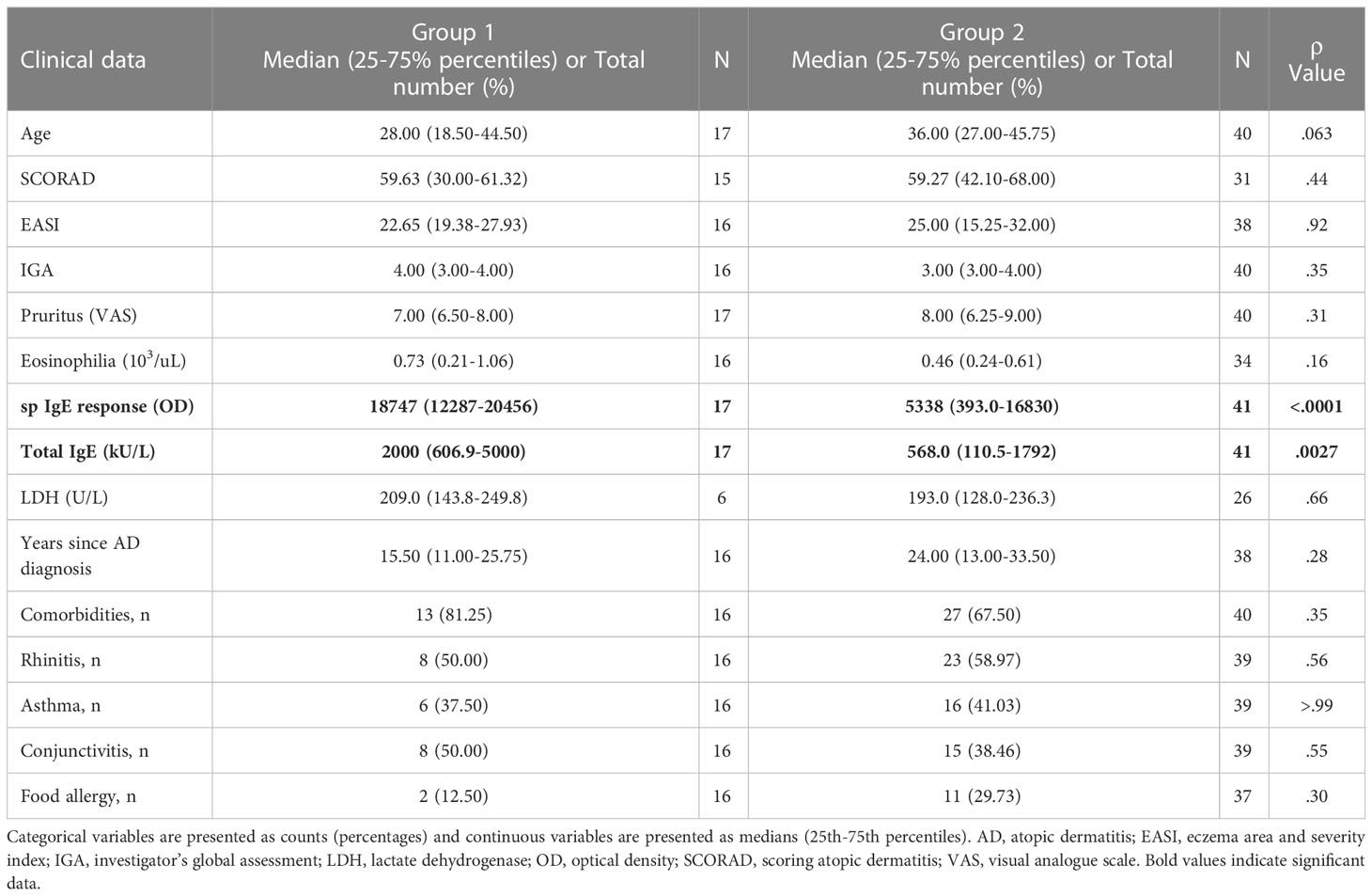
Table 1 Differences in clinical characteristics between group 1 and group 2.
To further characterize both groups, we evaluated changes in lesional skin tissue from patients belonging to group 1 and group 2 using gene-array analysis. We identified two hundred six probe-sets (143 unique genes) up-regulated and one hundred probe-sets (80 gens) down-regulated in group 1 skin compared with group 2 skin (Figure S7A; Table S3). Among the upregulated genes there were IL-20, IL-24, CCL20, CXCL1, CCL2 and EGR1. Only for IL-20 was FDR of less than .05. Enrichment tests of DEG revealed that group 1 lesional skin was enriched for biological processes related to the immune system such as response to external stimulus, inflammatory response and cellular response to chemokine (Figure S7B; Table S4). On the contrary, group 2 lesional skin was enriched for developmental processes (Figure S7C; Table S5).
Interestingly, difference in IL-20 expression between group 1 and group 2 of patients by gene-array analysis was confirmed by qRT-PCR (Figure S7D), but higher number of patients should be analyzed to confirm this data due to significant overlap between both groups. Additionally, increase in IL-17A and IL-21 mRNA expression was detected in group 1 compared to group 2 (Figure S7D), and no differences were found for IL-31 mRNA expression in cutaneous lesions between both groups.
An alternative analysis in the same patients revealed that those with high levels of sp IgE (> 100 kUA/L) showed increased HDM-induced IL-31 response by memory T cells, only in the CLA+ T-cell subset, than those with low levels of sp IgE (< 100 kUA/L) (Figure 3A). There was a trend towards positive correlation between IL-31 response by HDM-stimulated CLA+ T-cell cocultures and SCORAD in the high sp IgE group (r = 0.28, p = .28), whereas this trend was negative for the low sp IgE group (r = -0.32, p = .094; Figure 3B). Patients from the high sp IgE group showed increased eosinophilia and total IgE levels compared to patients from the low sp IgE group, as expected (Table 2). Additionally, in the high sp IgE there was a tendency towards more patients with conjunctivitis, a sign of HDM clinical symptom (p <.1).
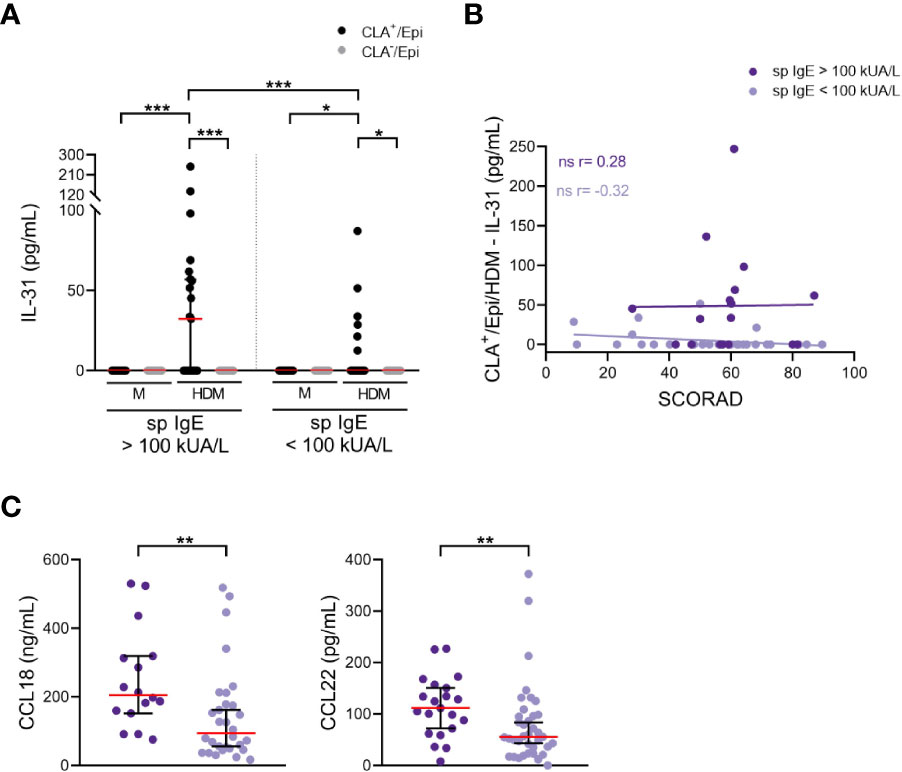
Figure 3 Enhanced IL-31 in vitro response and plasma levels of CCL18 and CCL22 in patients with sp IgE > 100 kUA/L. (A) IL-31 (pg/mL) from AD culture supernatants separated according to sp IgE levels (> 100 kUA/L n=21, < 100 kUA/L n=37). (B) Correlation between CLA+/Epi/HDM IL-31 and SCORAD in high (n=17) and low (n=29) sp IgE groups. (C) Plasma levels of CCL18 (ng/mL) and CCL22 (pg/mL) were compared between high (n=16-21) and low (n=30-37) sp IgE groups. CLA, cutaneous lymphocyte-associated antigen T cells; Epi, epidermal cells suspension; HDM, house dust mite; M, untreated; sp IgE, specific IgE to house dust mite; SCORAD, scoring atopic dermatitis. ns: p >.05; *p <.05; **p <.01; ***p <.001.
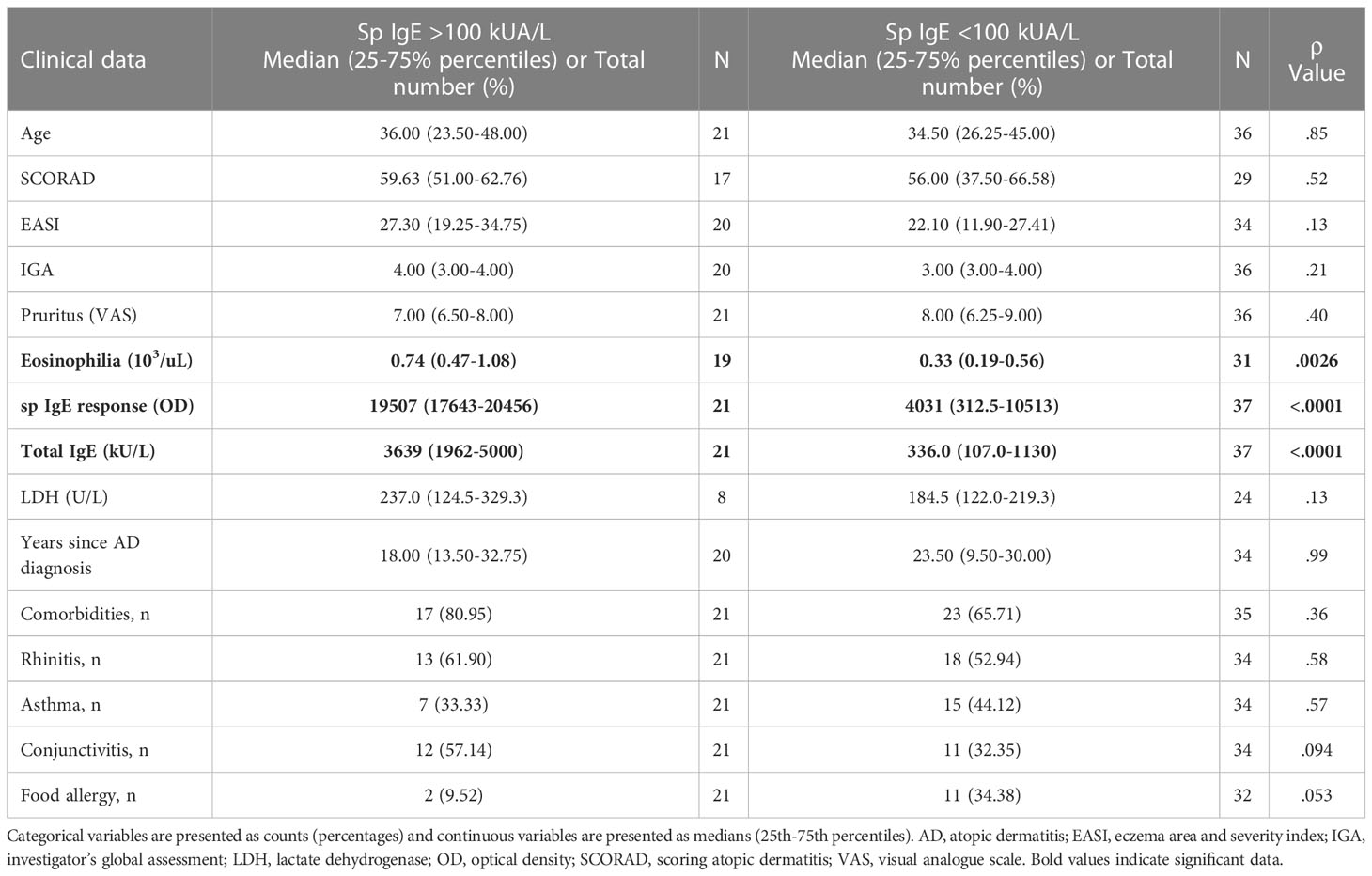
Table 2 Differences in clinical characteristics between patients with sp IgE >100 kUA/L and patients with sp IgE <100 kUA/L.
In line with the previous observation, the IL-31 mRNA expression in cutaneous lesions was greater in the high than in the low sp IgE groups (Figure S8A). Furthermore, plasma levels of CCL18, CCL22, CCL27, periostin and sIL-2R were elevated in the high compared to the low sp IgE groups (Figures 3C; S8B). Although results were significant (p <.05) there was clear overlap between both groups and increased number of patients would be required.
The IL-31 response by memory T cells activated with SEB showed no significant differences between high sp IgE and low sp IgE groups (Figure S8C). Additionally, no correlation between the HDM- and SEB-induced IL-31 production was observed in the high sp IgE and the low sp IgE groups (Figure S8D).
In moderate-to-severe AD the role of allergen in T-cell mediated IL-31 production and its possible relationship with the clinical status of the patients is presently uncharacterized. Our results suggest that the degree of allergen sensitization allows stratifying patients for IL-31 production by T cells with different clinical features.
In lesional AD skin CD4+ T cells are the most abundant infiltrating lymphocytes and they are mainly CLA+ T cells (39, 40). Until now, T-cell-derived IL-31 production has been mainly analyzed by qRT-PCR and intracellular flow cytometry of polyclonal-activated PBMC in a low number of patients and without any association with the clinic (8, 21, 23, 35, 41). In our study, HDM-induced IL-31 production by memory T cells defined two groups of AD patients: the IL-31 producing group (group 1) and the IL-31 non-producing group (group 2). In group 1, compared with group 2, HDM-induced IL-31 in CLA+ T cells positively correlated with patient´s pruritus intensity, and plasma levels of CCL27 and periostin. In addition, plasma levels of HDM-specific IgE and total IgE were increased in group1.
IL-31 is one of the main drivers of pruritus (2). For the first time, a direct correlation between memory T-cell-derived IL-31 response to HDM in vitro and patient’s pruritus was demonstrated, which underlines the clinical relevance of this mechanism in AD. This translational observation stablishes a relationship between allergen and patient’s pruritus through a mechanism restricted to a subset of circulating skin-homing memory T cells. The lack of difference in pruritus intensity between group 1 and group 2 of patients may be due to the broad range of pruritogens in AD, such as IL-13 or IL-4 (1), which could promote the itch signaling pathway in patients that do not produce IL-31 upon HDM stimulation. Since IL-31RA blockade improves symptoms in AD (25, 27, 42), further studies should explore the influence of allergen sensitization and the clinical response to anti-IL-31RA therapies in pruritus.
In group 1 of patients, IL-31 produced by CLA+ T cells positively correlates with CCL27, being CLA+ T cells and CCL27 mechanistically closely related, since CLA+ T cells express the CCR10 receptor that specifically binds CCL27 (43, 44). The relationship between IL-31 producers and plasma CCL27 suggests that lesional keratinocytes produce the CLA-attracting chemokine CCL27 to facilitate further recruitment of CLA+ T cells to the inflamed skin where, upon activation with HDM, would produce IL-31. CCL27 in the stratum corneum has been proposed as a biomarker of clinical response to nemolizumab, an anti-IL-31RA, treatment in AD (42), supporting the significant correlation between CCL27 and SCORAD in group 1 of patients. Besides this, not all patients respond to nemolizumab treatment, suggesting that allergen sensitization may be of help to identify anti-IL-31RA responder patients.
Periostin is an extracellular matrix protein closely related to Th2 immune response with an emerging role in pruritus and barrier dysfunction (45–48). In canine models, epicutaneous application of HDM to sensitized dogs induced upregulation of Th2 signature, including IL-31 and periostin (49). Nevertheless, the association between periostin plasma levels and IL-31, for HDM-triggered memory T cells, has not previously been reported.
Group 1 of patients showed increased levels of IL-13, IL-4, IL-5, IL-17A and IL-22, all of them commonly found in AD lesional skin (17), in response to HDM by the CLA+ T-cell cocultures, suggesting a strong inflammatory response in this group. Moreover, the direct correlation of IL-31 with IL-13 and IL-4 underlines the allergen-specific and Th2-phenotype of IL-31, and it is also supported by the IL-4-dependent IL-31 production reported in polyclonal stimulated CD4+ T-cell clones (5). The skin transcriptome confirmed the inflammatory signature in group 1 of patients, enriched in functions such as “response to external stimulus”, “inflammatory response”, “cellular response to chemokines” and “response to IL-1”. Among the up-regulated genes in this group, IL-20 is associated with IL-31-induced barrier disruption (18), and has also been related with pruritus in murine AD models (50). On the contrary, group 2 of patients lacked a transcriptomic dysregulation of inflammatory genes.
Group 2 of patients responded in most of the endpoints evaluated in this study (e.g. severity, pruritus), which is given by the nature that they are AD patients, so they share some clinical characteristics with group 1 of patients. The most significant clinical difference between both groups was the degree of IgE sensitization to HDM, along with total IgE levels, which were related with the differential IL-31 response by skin-derived memory T cells upon stimulation with allergen. Thereby, two groups that apparently would be the same population may show distinct molecular mechanisms driving disease immunopathogenesis.
AD is associated with elevated IgE, sensitization to aeroallergens and eosinophilia (51). Total IgE levels correlate with patients’ severity and are increased in patients sensitized to aeroallergens (33, 52). IgE levels are not a diagnostic requirement, but they are useful for determining prognosis, long-term outcome prediction or choosing therapy (53, 54). When stratifying patients according to their sp IgE levels, the increased IL-31 in vitro response in patients with high levels of sp IgE (over 100 kUA/L) supported the HDM contribution to disease activity in AD (32). Noteworthy, the increased IL-31 mRNA expression in lesional skin from high sp IgE patients may explain why topical exposure to HDM induced IL-31 only in a subset of HDM sensitized patients (21). The high sp IgE group also showed enhanced plasma levels of periostin, CCL27, CCL18, CCL22 and sIL-2R, thus associating the allergen-exposure and IL-31 response with Th2-immune response and general inflammation (55). To evaluate signs of HDM clinical symptoms and to compare the reactivity of CLA+ memory T cells to HDM with other allergens may be considered to complement these current findings.
A recent research described an increased IL-31RA expression on memory B cells from AD patients with total IgE levels > 1000 kU/L compared to controls, pointing to an association between IL-31-mediated mechanisms and atopic IgE-producing phenotype (56). For this reason, we classified patients according to their total IgE levels into high (> 1000 kU/L) and low (< 1000 kU/L) groups and found an increased HDM-induced memory T-cell IL-31, IL-13 and IL-4 response in the former group (Figure S9A), along with enhanced CCL26 mRNA expression in cutaneous lesions (Figure S9B) and plasma levels of CCL27, periostin, CCL17, CCL18, CCL22 and sIL-2R (Figure S9C). These data suggest that in allergen sensitized patients, IL-31 may play a role not only as T-cell-derived mediator by CLA+ T cells but also influencing IL-31RA-expressing B cells. This hypothesis is currently being investigated.
The knowledge generated from the present study is translationally relevant, since the allergen-induced IL-31 production by memory T cells was confined to the CLA+ T-cell subset, which reflects the skin-related mechanisms taking place in AD lesions and other inflammatory cutaneous diseases (57–61). Interestingly it has recently been reported that an early-term effect of dupilumab treatment on IL-13, IL-4, IL-5 and IL-22 expression is preferentially found in circulating CD4+ CLA+ T cells from AD patients (62, 63). Besides this, circulating allergen-specific and CLA+ T cells share the same TCRB CDR3 regions as lesional T cells from AD skin (64), supporting the relevance of our results for IL-31 produced by peripheral skin-homing T cells.
Circulating CLA+ T cells have been described to be the main memory T-cell subset producing IL-31 (23). When we complemented the analysis of T-cell mediated IL-31 response in the same patients by using SEB (65), IL-31 levels produced by CLA+ memory T cells were lower than those observed after HDM stimulation, without significant differences between groups 1 and 2 of patients or sp IgE levels. Additionally, a correlation between HDM- and SEB-induced IL-31 response was not detected, suggesting different roles of these stimuli in T-cell-mediated IL-31 production in AD. The molecular mechanisms by which SEB and HDM activate T cells are different. SEB directly binds to HLA class II molecules on the surface of antigen presenting cells and stimulates T cells expressing particular TCR. SEB can also induce skin inflammation by the following mechanisms: the direct binding to antigen presenting cells in the skin, direct binding to MHC class II keratinocytes, and induction of IgE-mediated immune responses (66). On the other hand, HDM requires uptake, processing and presentation by antigen presenting cells, a process that can be favored by the presence of allergen-specific IgE on the surface of antigen presenting cells (67). Therefore, the IgE-mediated uptake and antigen presentation may be favored in group 1 of patients, since they present higher levels of HDM-specific IgE.
Other inflammatory cells have been shown to produce IL-31: group 2 innate lymphoid cells (ILC2), basophils, eosinophils, dendritic cells, macrophages and mast cells (9–14) A single study has shown IL-31 expression by ILC2 (12), but the clinical relevance of alarmins, which are needed for ILC2 activation, in AD is not clear since all targeted therapies against them failed in clinical trials (68). Regarding basophils, IL-31 has been shown in chronic spontaneous urticaria and healthy donors (10), and no study using AD-derived basophils has shown IL-31 production. In eosinophils, IL-3 induces IL-31 production, but clinical relevance of eosinophils in AD has not been confirmed by anti-IL-5 therapy (14, 68). Dendritic cells express a hundred times less IL-31 than Th2 cells and whether dendritic cells produce physiologically relevant IL-31 quantities is underexplored (9). Recently, a new network comprising IL-31+ M2 macrophages, basophils, periostin and TSLP has been described (13); nonetheless, the clinical relevance of these results are unknown due to lack of efficacy of TSLP blockade for AD treatment (68, 69). Mast cells from psoriatic skin and healthy donors express IL-31, but this expression has not been shown for AD (11). Altogether, although IL-31 can be produced by different cell sources besides CLA+ memory T cells, no association with clinical pruritus has been reported, therefore their translational relevance is still under investigation. Additional studies on the involvement of IL-31 production by these non-CLA+ memory T cells on epithelial cells in coculture assays may be of help to further understand more mechanisms of IL-31 in AD.
This study has some limitations. We tried to detect IL-31 by intracellular flow cytometry but it was a complex issue due to the lack of good tools (5), and plasma or serum IL-31 levels were difficult to detect. Additionally, sample size for gene-array analysis as well as qRT-PCR was limited and more detailed investigations on the interactions between lesional epidermal cells and CLA+ memory T cells are needed.
Although the role of allergens in AD is still controversial and at the moment the stratification of response to anti-IL-31RA depending on IgE status has not been performed, it can be hypothesized that assessing specific IgE status to HDM in candidates for anti-IL-31RA therapy may eventually be helpful for identifying patients more prone to good clinical response to anti-IL-31RA and with a more favorable response in comparison to dupilumab or JAK inhibitors.
In summary, our findings bring new data on the mechanisms of allergen sensitization and IL-31 production by memory T cells in moderate-to-severe AD patients. The biphasic IL-31 response to HDM demonstrated with this translational ex vivo model may explain the partial IL-31 expression after HDM topical exposure in HDM-sensitized patients (21) and the heterogeneous response to anti-IL-31-directed therapies.
The original contributions presented in the study are publicly available in the Gene Expression Omnibus repository, accession number GSE226073.
The study involved human participants and was reviewed and approved by the Ethics Committee in Hospital de Bellvitge, Hospital del Granollers, Hospital del Mar and Hospital de la Santa Creu I Sant Pau (Spain). Participants provided written informed consent to participate in this study.
LS-dSN, IF-N, RP and LS-B contributed to the design of the study. IF-N, MB-O, AG, LC-B, MB-C, MF and ES-B included participants. LS-dSN, IF-N and IG-J participated in the investigation. LS-dSN analyzed the data and prepared figures. LS-dSN and LS-B wrote the manuscript. MF, RP and LS-B contributed to the funding acquisition. RP and LS-B supervised the study. All authors contributed to the article and approved the submitted version.
The study was funded by FIS/ISCIII 2021 (Ministerio de Economía y Competitividad e Instituto de Salud Carlos III; PI21/01179 and PI21/00335) and Fondo Europeo de Desarrollo Regional (FEDER). Additionally, Sans-de San Nicolàs L was granted by a PhD fellowship from the Agency for Management of University and Research Grants of the Generalitat de Catalunya (FI-SDUR 2020); García-Jiménez I was granted by a PhD fellowship from the Universitat de Barcelona (PREDOCS-UB 2020).
AG has received grants from AbbVie, Janssen, Lilly, Novartis and Sanofi, and lecture fees from AbbVie, Almirall, Leo Pharma and Sanofi. ES-B is a consultant for AbbVie, Almirall, Galderma, Genentec, Leo Pharma, Lilly, Novartis, Pfizer, Pierre Fabre and Sanofi. LS-B is a consultant for AbbVie, Janssen, Leo Pharma, Novartis and Sanofi.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1124018/full#supplementary-material
1. Steinhoff M, Ahmad F, Pandey A, Datsi A, AlHammadi A, Al-Khawaga S, et al. Neuroimmune communication regulating pruritus in atopic dermatitis. J Allergy Clin Immunol (2022) 149(6):1875–98. doi: 10.1016/j.jaci.2022.03.010
2. Datsi A, Steinhoff M, Ahmad F, Alam M, Buddenkotte J. Interleukin-31: The “itchy” cytokine in inflammation and therapy. Allergy Eur J Allergy Clin Immunol (2021) 76(10):2982–97. doi: 10.1111/all.14791
3. Nemmer JM, Kuchner M, Datsi A, Oláh P, Julia V, Raap U, et al. Interleukin-31 signaling bridges the gap between immune cells, the nervous system and epithelial tissues. Front Med (2021) 8:639097. doi: 10.3389/fmed.2021.639097
4. Raap U, Wichmann K, Bruder M, Ständer S, Wedi B, Kapp A, et al. Correlation of IL-31 serum levels with severity of atopic dermatitis. J Allergy Clin Immunol (2008) 122(2):421–3. doi: 10.1016/j.jaci.2008.05.047
5. Stott B, Lavender P, Lehmann S, Pennino D, Durham S, Schmidt-Weber CB. Human IL-31 is induced by IL-4 and promotes TH2-driven inflammation. J Allergy Clin Immunol (2013) 132(2):446–54. doi: 10.1016/j.jaci.2013.03.050
6. Dillon SR, Sprecher C, Hammond A, Bilsborough J, Rosenfeld-Franklin M, Presnell SR, et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol (2004) 5(7):752–60. doi: 10.1038/ni1084
7. McCandless EE, Rugg CA, Fici GJ, Messamore JE, Aleo MM, Gonzales AJ. Allergen-induced production of IL-31 by canine Th2 cells and identification of immune, skin, and neuronal target cells. Vet Immunol Immunopathol (2014) 157(1–2):42–8. doi: 10.1016/j.vetimm.2013.10.017
8. Datsi A, Raba K, Kellermann S, van Lierop A, Olah P, Sorg R, et al. 029 the IL-31-producing circulating T cells subset represents a unique population of CLA+ CRTH2+ CCR4+ effector memory T cells. J Invest Dermatol (2019) 139(9):S219. doi: 10.1016/j.jid.2019.07.032
9. Cevikbas F, Wang X, Akiyama T, Kempkes C, Savinko T, Antal A, et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J Allergy Clin Immunol (2014) 133(2):448–60. doi: 10.1016/j.jaci.2013.10.048
10. Raap U, Gehring M, Kleiner S, Rüdrich U, Eiz-Vesper B, Haas H, et al. Human basophils are a source of - and are differentially activated by - IL-31. Clin Exp Allergy (2017) 47(4):499–508. doi: 10.1111/cea.12875
11. Niyonsaba F, Ushio H, Hara M, Yokoi H, Tominaga M, Takamori K, et al. Antimicrobial peptides human β-defensins and cathelicidin LL-37 induce the secretion of a pruritogenic cytokine IL-31 by human mast cells. J Immunol (2010) 184(7):3526–34. doi: 10.4049/jimmunol.0900712
12. Alkon N, Bauer WM, Krausgruber T, Goh I. Single-cell analysis reveals innate lymphoid cell lineage infidelity in atopic dermatitis. J Allergy Clin Immunol (2022) 149(2):624–39. doi: 10.1016/j.jaci.2021.07.025
13. Hashimoto T, Yokozeki H, Karasuyama H, Satoh T. IL-31-generating network in atopic dermatitis comprising macrophages, basophils, thymic stromal lymphopoietin, and periostin. J Allergy Clin Immunol Am Acad Allergy Asthma Immunol (2022) 18:S0091–6749(22)01567-6. doi: 10.1016/j.jaci.2022.11.009
14. Kunsleben N, Rüdrich U, Gehring M, Novak N, Kapp A, Raap U. IL-31 induces chemotaxis, calcium mobilization, release of reactive oxygen species, and CCL26 in eosinophils, which are capable to release IL-31. J Invest Dermatol (2015) 135(7):1908–11. doi: 10.1038/jid.2015.106
15. Kasraie S, Niebuhr M, Baumert K, Werfel T. Functional effects of interleukin 31 in human primary keratinocytes. Allergy Eur J Allergy Clin Immunol (2011) 66(7):845–52. doi: 10.1111/j.1398-9995.2011.02545.x
16. Nakashima C, Otsuka A, Kabashima K. Interleukin-31 and interleukin-31 receptor: New therapeutic targets for atopic dermatitis. Exp Dermatol (2018) 27(4):327–31. doi: 10.1111/exd.13533
17. Möbus L, Rodriguez E, Harder I, Stölzl D, Boraczynski N, Gerdes S, et al. Atopic dermatitis displays stable and dynamic skin transcriptome signatures. J Allergy Clin Immunol (2021) 147(1):213–23. doi: 10.1016/j.jaci.2020.06.012
18. Cornelissen C, Marquardt Y, Czaja K, Wenzel J, Frank J, Lüscher-Firzlaff J, et al. IL-31 regulates differentiation and filaggrin expression in human organotypic skin models. J Allergy Clin Immunol (2012) 129(2):426–33. doi: 10.1016/j.jaci.2011.10.042
19. Hänel KH, Pfaff CM, Cornelissen C, Amann PM, Marquardt Y, Czaja K, et al. Control of the physical and antimicrobial skin barrier by an IL-31–IL-1 signaling network. J Immunol (2016) 196(8):3233–44. doi: 10.4049/jimmunol.1402943
20. Dai X, Shiraishi K, Muto J, Utsunomiya R, Mori H, Murakami M, et al. Nuclear IL-33 plays an important role in IL-31−Mediated downregulation of FLG, keratin 1, and keratin 10 by regulating signal transducer and activator of transcription 3 activation in human keratinocytes. J Invest Dermatol (2022) 142(1):136–44. doi: 10.1016/j.jid.2021.05.033
21. Sonkoly E, Muller A, Lauerma AI, Pivarcsi A, Soto H, Kemeny L, et al. IL-31: A new link between T cells and pruritus in atopic skin inflammation. J Allergy Clin Immunol (2006) 117(2):411–7. doi: 10.1016/j.jaci.2005.10.033
22. Singh B, Jegga AG, Shanmukhappa KS, Edukulla R, Khurana GH, Medvedovic M, et al. IL-31-driven skin remodeling involves epidermal cell proliferation and thickening that lead to impaired skin-barrier function. PloS One (2016) 11(8):e0161877. doi: 10.1371/journal.pone.0161877
23. Bilsborough J, Leung DYM, Maurer M, Howell M, Boguniewcz M, Yao L, et al. IL-31 is associated with cutaneous lymphocyte antigen-positive skin homing T cells in patients with atopic dermatitis. J Allergy Clin Immunol (2006) 117(2):418–25. doi: 10.1016/j.jaci.2005.10.046
24. Feld M, Garcia R, Buddenkotte J, Katayama S, Lewis K, Muirhead G, et al. The pruritus- and TH2-associated cytokine IL-31 promotes growth of sensory nerves. J Allergy Clin Immunol (2016) 138(2):500–8. doi: 10.1016/j.jaci.2016.02.020
25. Silverberg JI, Pinter A, Alavi A, Lynde C, Bouaziz JD, Wollenberg A, et al. Nemolizumab is associated with a rapid improvement in atopic dermatitis signs and symptoms: subpopulation (EASI ≥ 16) analysis of randomized phase 2B study. J Eur Acad Dermatol Venereol (2021) 35(7):1562–8. doi: 10.1111/jdv.17218
26. Serra-Baldrich E, Santamaria-Babí LF, Silvestre JF. Nemolizumab: un innovador tratamiento biológico para el control de la interleuquina 31 (IL-31) clave en la dermatitis atópica y el prurigo nodular. Actas Dermosifiliogr (2022) 113(7):674–84. doi: 10.1016/j.ad.2021.12.014
27. Silverberg JI, Pinter A, Pulka G, Poulin Y, Bouaziz JD, Wollenberg A, et al. Phase 2B randomized study of nemolizumab in adults with moderate-to-severe atopic dermatitis and severe pruritus. J Allergy Clin Immunol (2020) 145(1):173–82. doi: 10.1016/j.jaci.2019.08.013
28. Horejs-Hoeck J, Schwarz H, Lamprecht S, Maier E, Hainzl S, Schmittner M, et al. Dendritic cells activated by IFN-γ/STAT1 express IL-31 receptor and release proinflammatory mediators upon IL-31 treatment. J Immunol (2012) 188(11):5319–26. doi: 10.4049/jimmunol.1101044
29. Kasraie S, Niebuhr M, Werfel T. Interleukin (IL)-31 induces pro-inflammatory cytokines in human monocytes and macrophages following stimulation with staphylococcal exotoxins. Allergy Eur J Allergy Clin Immunol (2010) 65(6):712–21. doi: 10.1111/j.1398-9995.2009.02255.x
30. Cheung PFY, Wong CK, Ho AWY, Hu S, Chen DP, Lam CWK. Activation of human eosinophils and epidermal keratinocytes by Th2 cytokine IL-31: Implication for the immunopathogenesis of atopic dermatitis. Int Immunol (2010) 22(6):453–67. doi: 10.1093/intimm/dxq027
31. Wong CK, Leung KML, Qiu HN, Chow JYS, Choi AOK, Lam CWK. Activation of eosinophils interacting with dermal fibroblasts by pruritogenic cytokine IL-31 and alarmin IL-33: Implications in atopic dermatitis. PloS One (2012) 7(1):e29815. doi: 10.1371/journal.pone.0029815
32. Mittermann I, Wikberg G, Johansson C, Lupinek C, Lundeberg L, Crameri R, et al. IgE sensitization profiles differ between adult patients with severe and moderate atopic dermatitis. PloS One (2016) 11(5):e0156077. doi: 10.1371/journal.pone.0156077
33. Sonesson A, Bartosik J, Christiansen J, Roscher I, Nilsson F, Schmidtchen A, et al. Sensitization to skin-associated microorganisms in adult patients with atopic dermatitis is of importance for disease severity. Acta Derm Venereol (2013) 93(3):340–5. doi: 10.2340/00015555-1465
34. Tamamoto-Mochizuki C, Olivry T. IL-31 and IL-31 receptor expression in acute experimental canine atopic dermatitis skin lesions. Vet Dermatol (2021) 32(6):631–e169. doi: 10.1111/vde.13034
35. Szegedi K, van Lier A, Res PC, Chielie S, Bos JD, Kezic S, et al. House dust mite allergens der f and der p induce IL-31 production by blood-derived T cells from atopic dermatitis patients. Exp Dermatol (2018) 27(4):393–5. doi: 10.1111/exd.13438
36. Ferran M, Galván AB, Rincón C, Romeu ER, Sacrista M, Barboza E, et al. Streptococcus induces circulating CLA+ memory T-cell-dependent epidermal cell activation in psoriasis. J Invest Dermatol (2013) 133(4):999–1007. doi: 10.1038/jid.2012.418
37. Leung DYM, Gately M, Trumble A, Ferguson-Darnell B, Schlievert PM, Picker LJ. Bacterial superantigens induce T cell expression of the skin-selective homing receptor, the cutaneous lymphocyte-associated antigen, via stimulation of interleukin 12 production. J Exp Med (1995) 181(2):747–53. doi: 10.1084/jem.181.2.747
38. Chan JR, Blumenschein W, Murphy E, Diveu C, Wiekowski M, Abbondanzo S, et al. IL-23 stimulates epidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasis pathogenesis. J Exp Med (2006) 203(12):2577–87. doi: 10.1084/jem.20060244
39. Leung DYM, Bhan AK, Schneeberger EE, Geha RS. Characterization of the mononuclear cell infiltrate in atopic dermatitis using monoclonal antibodies. J Allergy Clin Immunol (1983) 71(1 PART 1):47–56. doi: 10.1016/0091-6749(83)90546-8
40. Picker LJ, Michie SA, Rott LS, Butcher EC. A unique phenotype of skin-associated lymphocytes in humans: Preferential expression of the HECA-452 epitope by benign and malignant T cells at cutaneous sites. Am J Pathol (1990) 136(5):1053–68.
41. Szegedi K, Kremer AE, Kezic S, Teunissen MBM, Bos JD, Luiten RM, et al. Increased frequencies of IL-31-producing T cells are found in chronic atopic dermatitis skin. Exp Dermatol (2012) 21(6):431–6. doi: 10.1111/j.1600-0625.2012.01487.x
42. Sidbury R, Alpizar S, Laquer V, Dhawan S, Abramovits W, Loprete L, et al. Pharmacokinetics, safety, efficacy, and biomarker profiles during nemolizumab treatment of atopic dermatitis in adolescents. Dermatol Ther (2022) 12(3):631–42. doi: 10.1007/s13555-021-00678-7
43. Homey B, Wang W, Soto H, Buchanan ME, Wiesenborn A, Catron D, et al. Cutting edge: The orphan chemokine receptor G protein-coupled receptor-2 (GPR-2, CCR10) binds the skin-associated chemokine CCL27 (CTACK/ALP/ILC). J Immunol (2000) 164(7):3465–70. doi: 10.4049/jimmunol.164.7.3465
44. Homey B, Alenius H, Müller A, Soto H, Bowman EP, Yuan W, et al. CCL27-CCR10 interactions regulate T cell-mediated skin inflammation. Nat Med (2002) 8(2):157–65. doi: 10.1038/nm0202-157
45. Masouka M, Shiraishi H, Ochta S, Suzuki S, Arima K, Aoki S, et al. Periostin promotes chronic allergic inflammation in response to Th2 cytokines. Jorunal Clin Investig (2012) 122(7):2590–600. doi: 10.1172/JCI58978
46. Guttman-Yassky E, Bissonnette R, Ungar B, Suárez-Fariñas M, Ardeleanu M, Esaki H, et al. Dupilumab progressively improves systemic and cutaneous abnormalities in patients with atopic dermatitis. J Allergy Clin Immunol (2019) 143(1):155–72. doi: 10.1016/j.jaci.2018.08.022
47. Hashimoto T, Mishra SK, Olivry T, Yosipovitch G. Periostin, an emerging player in itch sensation. J Invest Dermatol (2021) 141(10):2338–43. doi: 10.1016/j.jid.2021.03.009
48. Mitamura Y, Nunomura S, Nanri Y, Ogawa M, Yoshihara T, Masuoka M, et al. The IL-13/periostin/IL-24 pathway causes epidermal barrier dysfunction in allergic skin inflammation. Allergy Eur J Allergy Clin Immunol (2018) 73(9):1881–91. doi: 10.1111/all.13437
49. Olivry T, Mayhew D, Paps JS, Linder KE, Peredo C, Rajpal D, et al. Early activation of Th2/Th22 inflammatory and pruritogenic pathways in acute canine atopic dermatitis skin lesions. J Invest Dermatol (2016) 136(10):1961–9. doi: 10.1016/j.jid.2016.05.117
50. Zhiping L, Xiao S, Chen W, Re Z, Yang H, Steinhoff M, et al. IL- 20 promotes cutaneous inflammation and peripheral itch sensation in atopic dermatitis. FASEB J (2022) 36(6):e22334. doi: 10.1096/fj.202101800R
51. Leung DYM. Role of IgE in atopic dermatitis. Curr Opin Immunol (1993) 5(6):956–62. doi: 10.1016/0952-7915(93)90112-6
52. Laske N, Niggemann B. Does the severity of atopic dermatitis correlate with serum IgE levels? Pediatr Allergy Immunol (2004) 15(1):86–8. doi: 10.1046/j.0905-6157.2003.00106.x
53. Chow S, Seow CS, Dizon MV, Godse K, Foong H, Chan V, et al. A clinician’s reference guide for the management of atopic dermatitis in asians. Asia Pac Allergy (2018) 8(4):e41. doi: 10.5415/apallergy.2018.8.e41
54. Kiiski V, Karlsson O, Remitz A, Reitamo S. High serum total IgE predicts poor long-term outcome in atopic dermatitis. Acta Derm Venereol (2015) 95(8):943–7. doi: 10.2340/00015555-2126
55. Leonard A, Wang J, Yu L, Liu H, Estrada Y, Greenlees L, et al. Atopic dermatitis endotypes based on allergen sensitization, reactivity to staphylococcus aureus antigens, and underlying systemic inflammation. J Allergy Clin Immunol Pract (2020) 8(1):236–247.e3. doi: 10.1016/j.jaip.2019.08.013
56. Unger Z, Gomez-Casado C, van Lierop A, Pongratz G, Bleck D, Homey B. 027 memory b cells of atopic individuals preferentially express IL-31RA: A putative role of IL-31 in b cell biology. J Invest Dermatol (2022) 142(12):S185. doi: 10.1016/j.jid.2022.09.036
57. Ferran M, Santamaria-Babi LF. Pathological mechanisms of skin homing T cells in atopic dermatitis. World Allergy Organ J (2010) 3(3):44–7. doi: 10.1097/WOX.0b013e3181d675f8
58. Czarnowicki T, Santamaria-Babí LF, Guttman-Yassky E. Circulating CLA+ T cells in atopic dermatitis and their possible role as peripheral biomarkers. J Allergy Clin Immunol (2017) 72(3):366–72. doi: 10.1111/all.13080
59. DeJesús-Gil C, Sans-de San Nicolàs L, García-Jiménez I, Ferran M, Celada A, Chiriac A, et al. The translational relevance of human circulating memory cutaneous lymphocyte-associated antigen positive T cells in inflammatory skin disorders. Front Immunol (2021) 12:652613. doi: 10.3389/fimmu.2021.652613
60. Langan SM, Irvine AD, Weidinger S. Atopic dermatitis. Lancet (2020) 396(10247):345–60. doi: 10.1016/S0140-6736(20)31286-1
61. Santamaria-Babi LF. Atopic dermatitis Pathogenesis :Lessons from immunology. Dermatol Pr Concept (2022) 12(1):e2022152. doi: 10.5826/dpc.1201a152
62. Bakker DS, van der Wal MM, Heeb LEM, Giovannone B, Asamoah M, Delemarre EM, et al. Early and long-term effects of dupilumab treatment on circulating T-cell functions in patients with moderate-to-Severe atopic dermatitis. J Invest Dermatol (2021) 141(8):1943–53. doi: 10.1016/j.jid.2021.01.022
63. Roesner LM, Bridgewood C, McGonagle D, Wittmann M. Dupilumab: An opportunity to unravel In vivo actions of IL-4 and IL-13 in humans. J Invest Dermatol (2021) 141(8):1879–81. doi: 10.1016/j.jid.2021.02.007
64. Roesner LM, Farag AK, Pospich R, Traidl S, Werfel T. T Cell receptor sequencing specifies psoriasis as a systemic and atopic dermatitis as a skin-focused, allergen-driven disease. Allergy (2022) 77(9):2737–47. doi: 10.1111/all.15272
65. Sans-De San Nicolàs L, Figueras-Nart I, Bonfill-Ortí M, De Jesús-Gil C, García-Jiménez I, Guilabert A, et al. SEB-induced IL-13 production in CLA+ memory T cells defines Th2 high and Th2 low responders in atopic dermatitis. Allergy (2022) 77(11):3448–51. doi: 10.1111/all.15424
66. Skov L, Olsen JV, Giorno R, Schlievert PM, Baadsgaard O, Leung DYM. Application of staphylococcal enterotoxin b on normal and atopic skin induces up-regulation of T cells by a superantigen-mediated mechanism. J Allergy Clin Immunol (2000) 105(4):820–6. doi: 10.1067/mai.2000.105524
67. Novak N, Valenta R, Bohle B, Laffer S, Haberstok J, Kraft S, et al. FcϵRI engagement of langerhans cell-like dendritic cells and inflammatory dendritic epidermal cell-like dendritic cells induces chemotactic signals and different T-cell phenotypes in vitro. J Allergy Clin Immunol (2004) 113(5):949–57. doi: 10.1016/j.jaci.2004.02.005
68. Trier AM, Kim BS. Insights into atopic dermatitis pathogenesis lead to newly approved systemic therapies. Br J Dermatol (2022) ljac016. doi: 10.1093/bjd/ljac016
69. Simpson EL, Parnes JR, She D, Crouch S, Rees W, Mo M, et al. Tezepelumab, an anti–thymic stromal lymphopoietin monoclonal antibody, in the treatment of moderate to severe atopic dermatitis: A randomized phase 2a clinical trial. J Am Acad Dermatol (2019) 80(4):1013–21. doi: 10.1016/j.jaad.2018.11.059
Keywords: atopic dermatitis, CCL27, CLA+ memory T cells, house dust mite, IgE, IL-31, periostin, pruritus
Citation: Sans-de San Nicolàs L, Figueras-Nart I, García-Jiménez I, Bonfill-Ortí M, Guilabert A, Curto-Barredo L, Bertolín-Colilla M, Ferran M, Serra-Baldrich E, Pujol RM and Santamaria-Babí LF (2023) Allergen sensitization stratifies IL-31 production by memory T cells in atopic dermatitis patients. Front. Immunol. 14:1124018. doi: 10.3389/fimmu.2023.1124018
Received: 14 December 2022; Accepted: 28 February 2023;
Published: 13 March 2023.
Edited by:
Jongdae Lee, Guangzhou Medical University, ChinaReviewed by:
Hans David Brightbill, Genentech Inc., United StatesCopyright © 2023 Sans-de San Nicolàs, Figueras-Nart, García-Jiménez, Bonfill-Ortí, Guilabert, Curto-Barredo, Bertolín-Colilla, Ferran, Serra-Baldrich, Pujol and Santamaria-Babí. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis F. Santamaria-Babí, bHVpcy5zYW50YW1hcmlhQHViLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.