 Maud Deny1,2
Maud Deny1,2 Alexandros Popotas1
Alexandros Popotas1 Laurence Hanssens3
Laurence Hanssens3 Nicolas Lefèvre1,3
Nicolas Lefèvre1,3 Luis Alexis Arroba Nuñez1,2
Luis Alexis Arroba Nuñez1,2 Ghislaine Simo Ouafo1,2
Ghislaine Simo Ouafo1,2 Francis Corazza4
Francis Corazza4 Georges Casimir1,3
Georges Casimir1,3 Mustapha Chamekh1,2*
Mustapha Chamekh1,2*- 1Inflammation Unit, Laboratory of Pediatric Research, Faculty of Medicine, Université Libre de Bruxelles (ULB), Brussels, Belgium
- 2Université Libre de Bruxelles (ULB) Center for Research in Immunology (U-CRI), Brussels, Belgium
- 3Institut de Mucoviscidose – Unité Pédiatrique, Hôpital Universitaire des Enfants Reine Fabiola, Université Libre de Bruxelles (ULB), Brussels, Belgium
- 4Laboratoire de Médecine Translationnelle, Centre Hospitalier Universitaire Brugmann, Université Libre de Bruxelles (ULB), Brussels, Belgium
Previous studies have reported sex disparity in cystic fibrosis (CF) disease, with females experiencing more pulmonary exacerbations and frequent microbial infections resulting in shorter survival expectancy. This concerns both pubertal and prepubertal females, which is in support to the prominent role of gene dosage rather than the hormonal status. The underlying mechanisms are still poorly understood. The X chromosome codes for a large number of micro-RNAs (miRNAs) that play a crucial role in the post-transcriptional regulation of several genes involved in various biological processes, including inflammation. However, their level of expression in CF males and females has not been sufficiently explored. In this study, we compared in male and female CF patients the expression of selected X-linked miRNAs involved in inflammatory processes. Cytokine and chemokine profiles were also evaluated at both protein and transcript levels and cross-analyzed with the miRNA expression levels. We observed increased expression of miR-223-3p, miR-106a-5p, miR-221-3p and miR-502-5p in CF patients compared to healthy controls. Interestingly, the overexpression of miR-221-3p was found to be significantly higher in CF girls than in CF boys and this correlates positively with IL-1β. Moreover, we found a trend toward lower expression in CF girls than in CF boys of suppressor of cytokine signaling 1 (SOCS1) and the ubiquitin-editing enzyme PDLIM2, two mRNA targets of miR-221-3p that are known to inhibit the NF-κB pathway. Collectively, this clinical study highlights a sex-bias in X-linked miR-221-3p expression in blood cells and its potential contribution to sustaining a higher inflammatory response in CF girls.
Introduction
Cystic fibrosis (CF) is a monogenic disease caused by the mutation of cystic fibrosis transmembrane conductance regulator (CFTR) gene, an anion channel-protein coding sequence that plays a crucial role in the airway surface liquid homeostasis. CFTR dysfunction leads to an imbalance between Cl- secretion and Na+ uptake, resulting in airway surface dehydration and impaired mucociliary clearance (1, 2). A chronic inflammation within the lung is the hallmark of the disease, with exacerbation episodes linked to bacterial infection by opportunistic pathogens such as Staphylococcus aureus and Pseudomonas aeruginosa, leading to progressive airflow obstruction, tissue destruction and ultimately decreased respiratory function (1, 2).
We and others have reported sex-differences in the course of CF, with females experiencing more severe inflammation and exhibiting poorer prognosis compared to males (3–6). This sex disparity in CF exists despite the progress made in the early diagnosis and in the close therapeutic monitoring over the last years (6–10). Currently, the underlying mechanisms are poorly understood. Sex hormones and X-linked sexual genetic architecture have been proposed as potential factors (11–13). The essential role of the X chromosome is supported by the finding that such sex-differences in CF outcomes are also observed in young prepubertal children. Indeed, it has been shown that prepubertal girls have a significantly higher recruitment of neutrophils within the lungs and develop more episodes of infection than boys (3).
Interestingly, genomic distribution analysis revealed a high density of micro-RNAs (miRNAs) on the X chromosome (14). miRNAs are small non-coding RNA sequences of ±22 nucleotides that regulate gene expression at post-transcriptional level by repressing translation of specific mRNA targets through complementary pairing with their 3’ untranslated regions. Growing evidence points to their potential role in chronic inflammatory diseases impacting lung tissues, such as in CF (15–21). Indeed, studies have shown that CF patients have a dysregulation in the expression of number of miRNAs, including those targeting the mutated CFTR transporter or its derivatives, such as miR-145 or miR-494 (18, 22), or those involved in inflammatory pathology, such as the miR-126, miR-17 or miR-155 (23–25). Other studies showed a significant difference in the expression of X-linked miRNAs in monocytes from CF patients compared to non-CF patients (26). However, few reports investigated whether biological sex could impact miRNAs expression profiling in CF patients. Using miRNA array analysis of the plasma from six CF male and six CF female patients, Mooney et al. reported differential expression of two circulating miRNAs, miR-885-5p and miR-193a-5p, between girls and boys (27). Both miRNAs are not localized on the X chromosome and have no apparent role in the inflammatory process. Of note, females display mosaic cells expressing two X-linked gene alleles and one of the X chromosomes is randomly inactivated early in embryogenesis to ensure the compensation dosage. Yet, about 15-23% of human X-chromosome-linked genes escape the inactivation with some variability between tissues and individuals (28, 29). This may result in differential expression of X-linked genes between males and females. In this study, we asked whether blood leukocytes from male and female CF patients exhibit a differential expression profile of X-linked miRNAs with potent impact on the inflammatory process. We focused on miRNAs that have been shown to be dysregulated in the airways of CF patients compared to healthy individuals (23) and for which there is accumulating evidence of their direct or indirect role in the regulation of inflammation.
Materials and methods
Ethics statement
All blood samples were obtained from control subjects and CF patients at the Queen Fabiola University Children’s Hospital (HUDERF) in Brussels, Belgium. Written consent was obtained from the children’s parents or legal guardians. The protocols were approved by the hospital ethics committee of the Brugmann University Hospital (CE 2016/162).
Characteristics of patients
29 CF patients (15 males and 14 females) and 20 healthy controls (10 males and 10 females) were recruited. The patients were diagnosed using a sweat test and checked for CFTR gene mutation. The different characteristics of the patients are presented in Table 1.
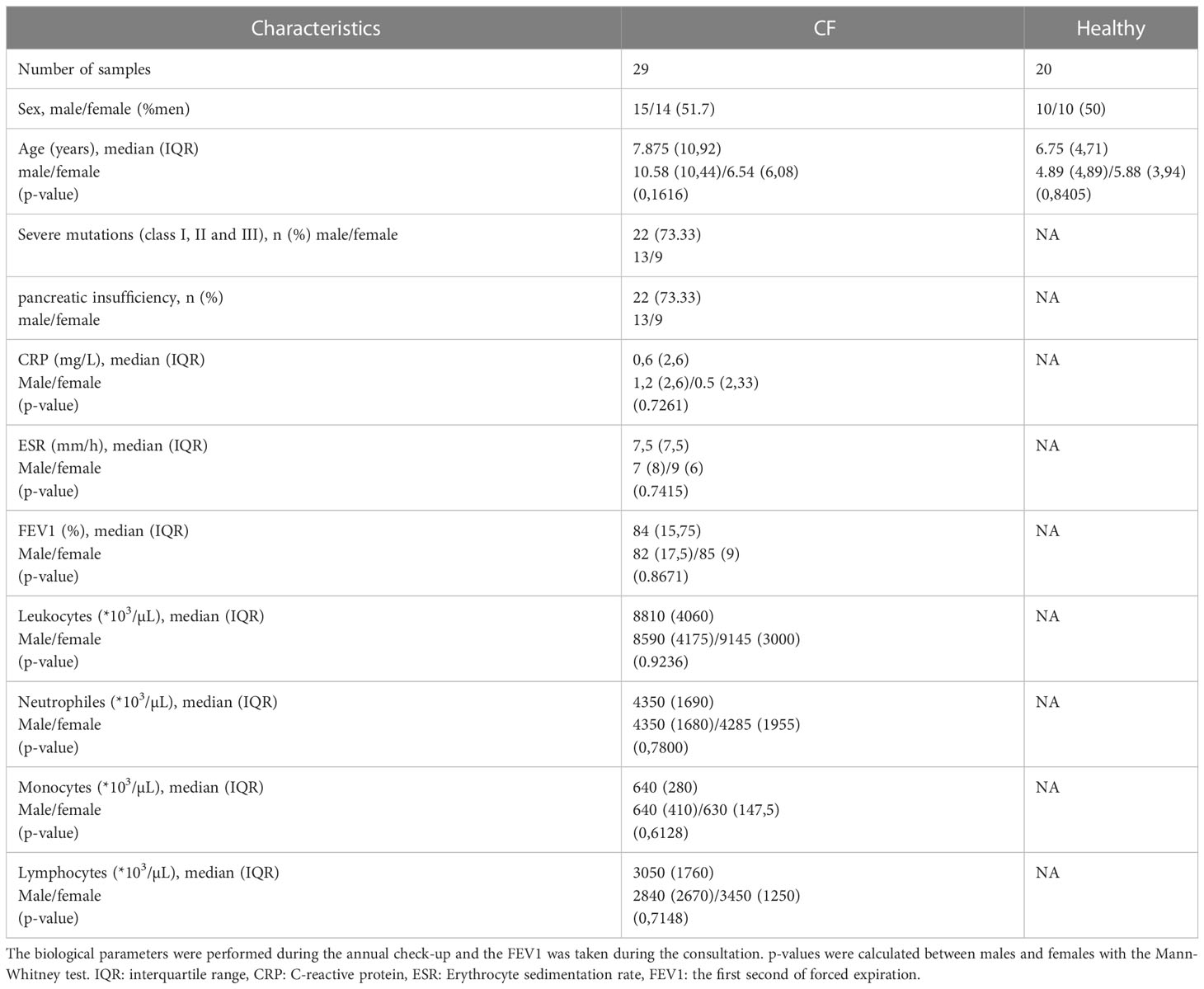
Table 1 Characteristics of CF patients and healthy controls enrolled in the study.
Sample processing
Blood samples were collected in EDTA tubes. Blood from CF patients and healthy controls was processed within 4 hours of collection to extract plasma and leukocytes. After extraction from plasma, blood cells were treated with the erythrolysis buffer and leucocytes were resuspended in Trizol (Qiagen). The plasma and leucocytes were stored at -80°C until use.
Detection of inflammatory cytokines in plasma
The quantification of inflammatory mediator levels in the plasma was performed by Luminex Multiplex Assays (ThermoFischer, EPX210-1585 0-901) according to the manufacturer’s instructions. Plate reading and quantification were performed using MAGPIX™ Instrument System (Invitrogen), and the results were analyzed using the ProcartaPlex™ program (ThermoFischer). In most samples, cytokine levels were above detection limits. The extrapolation was made for samples that were below the detection limit.
Total RNA extraction and miRNAs and mRNAs expression analysis
Total RNA was isolated using the miRNeasy mini kit (Qiagen) following the manufacturer’s instructions from leukocytes samples. RNA concentrations were assessed using the NanoVue Plus spectrophotometer (Biochrom). The reverse transcription of total RNA (including miRNAs) was performed with the miScript II RT Kit (Qiagen) following the manufacturer’s instructions. qRT-PCR was performed with the miScript SYBR Green PCR Kit (Qiagen) according to standard protocols. We used the small-nucleolar RNA C/D box 95 (SNORD95) as miRNA internal control and two house-keeping genes (GAPDH and beta-actin) for mRNA expression analysis. Reactions with no-template and no-reverse transcriptase were performed to unsure negative controls. The specificity of each primer pairs was further verified by controlling the melt curve profile. Relative quantification of miRNAs expression was calculated using 2−ΔΔCt method; the fold change differences were relative to sex-matched healthy controls. The primers used in this study are listed in Table 2.
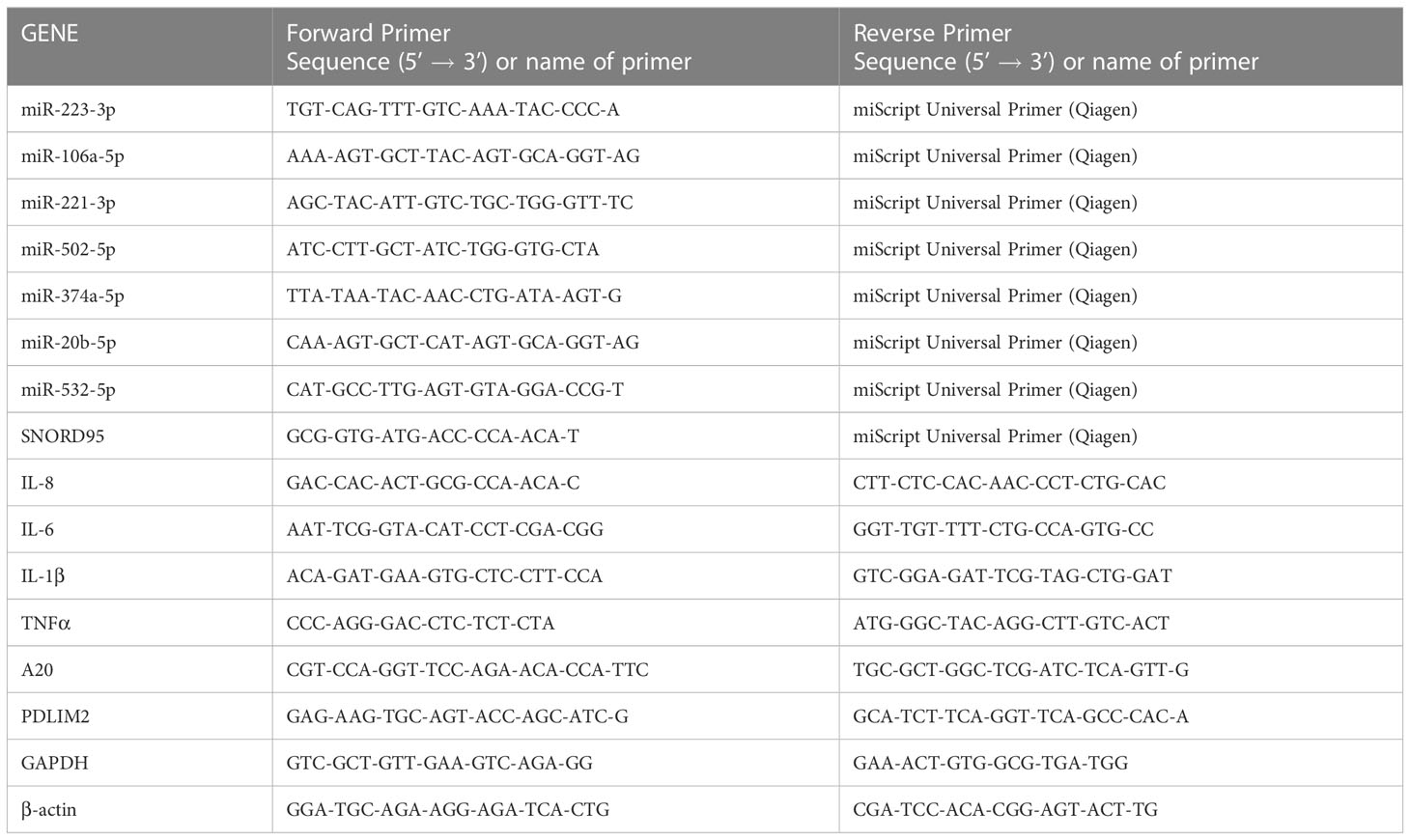
Table 2 Oligonucleotide primers used for miRNA and mRNA qRT-PCR analysis.
Statistical analysis
We used the non-parametric Mann-Whitney test for independent groups to compare males and females. Correlations were tested using the non-parametric Spearman test. Bonferroni correction was applied when needed. Differences were considered statistically significant with p (two-tailed) <0.05 using Prism 8.0 statistical software (GraphPad, San Diego, California), with *, **, and *** symbols respectively signify p <0.05, p <0.01, and p <0.001.
Results
Comparative analysis of the expression of inflammatory mediators
A persistent and damaging chronic inflammation is the hallmark of CF pathology. To evaluate the clinical status of the patients, we analyzed various inflammatory parameters. As shown in Table 1, patients included in this study have a relatively stable clinical situation with low levels of C-reactive protein (CRP), erythrocyte sedimentation rate (ESR), normal blood cell composition, and a mild to moderate pulmonary phenotype as measured by first second of forced expiration (FEV1). Of note, no sex differences were observed for these parameters (Table 1).
To further analyze the inflammatory state of CF patients, we also evaluated the balance of circulating pro- and anti-inflammatory mediators (TNFα, IL-1β, IL-6, IL-8, IL-10, IL-12p70, IL-17A, IL-18, CCL2, CCL3, CXCL10 and CCL4) in plasma. Compared with healthy controls, CF patients had higher amount of most cytokine and chemokine mediators, with the exception of CCL4 which was significantly lower. We found no obvious difference between CF boys and girls, although we noticed a marked production of TNFα, IL-1β, IL-8, IL-10, IL-12p70, IL-17A, and CXCL10 in CF females than CF males when compared to their matched controls (Figure 1).
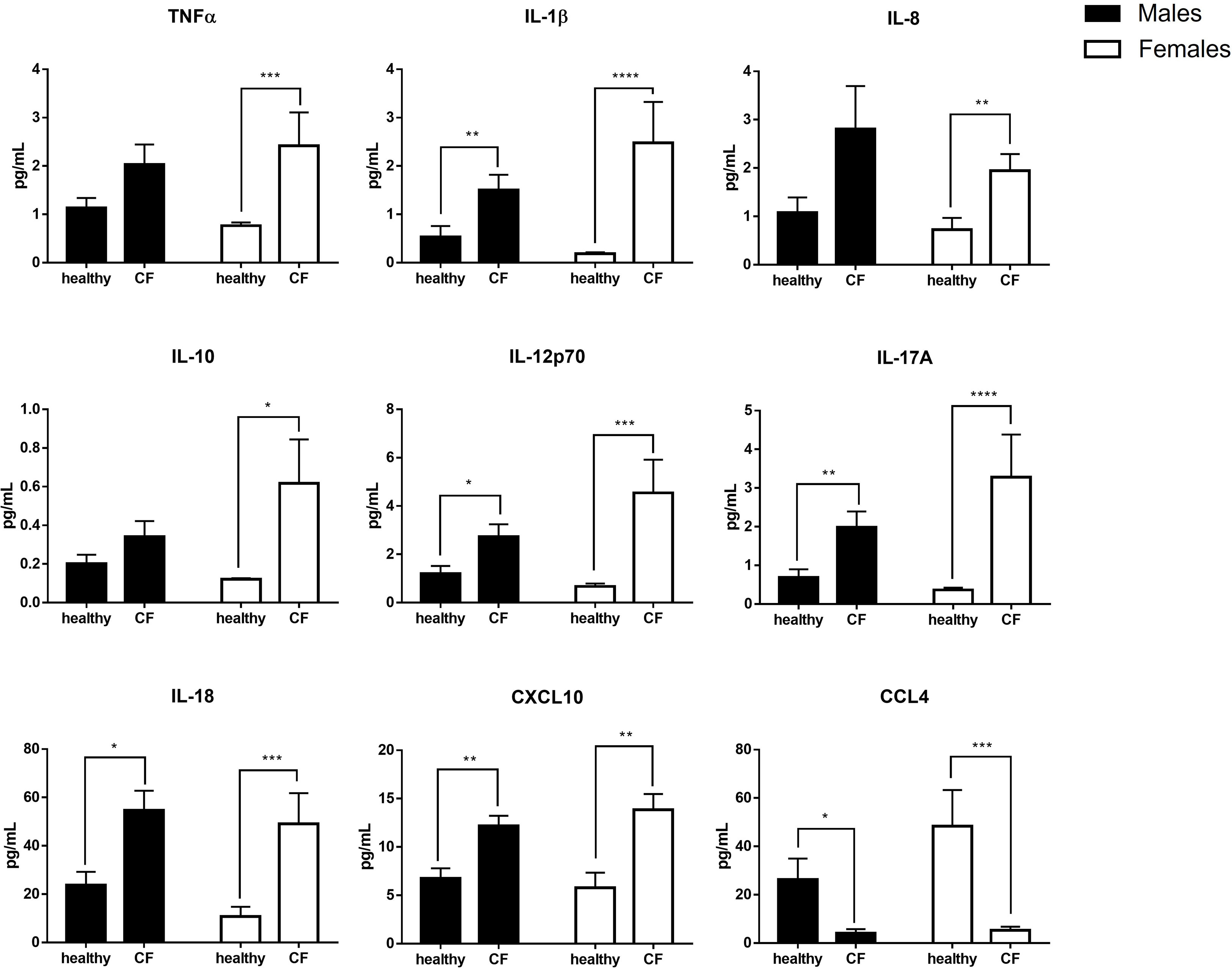
Figure 1 Sex-based analysis of the production of pro- and anti-inflammatory mediators in the plasma of CF patients (n=29; 15 males and 14 females) and healthy controls (n=20; 10males and 10 females). Graphs show the mean ± SEM. *p<0,05; **p<0,01; ***p<0,001: ****p<0,0001 (Mann Whitney U test).
In parallel, we evaluated by qRT-PCR the expression of major inflammatory cytokine transcripts. As shown in Figure 2, the level of expression of IL-1β is significantly higher only in CF females compared to control, while the expression IL-8 increased in both CF males and females compared to sex-matched controls. We found, however, no significant difference for IL-6 and TNFα. Overall, although CF patients enrolled in this study exhibited a relatively stable clinical state, these results indicate an upregulation of several inflammatory mediators that might impact the physiopathology.
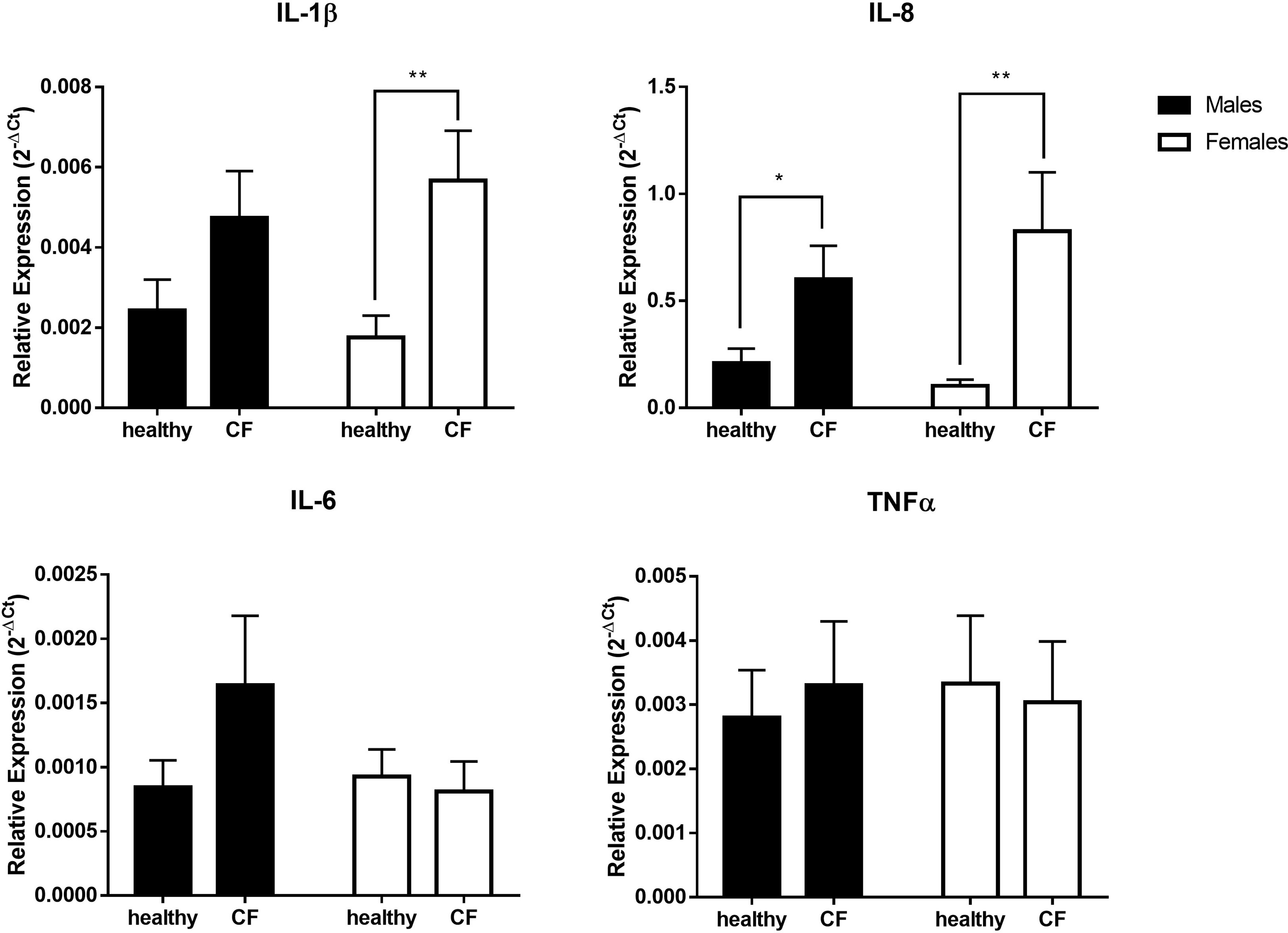
Figure 2 Sex-based assessment of inflammatory cytokines expression in blood leukocytes from male healthy controls (n=10) versus male CF patients (n=15) and female healthy controls (n=10) versus female CF patients (n=14). The values were normalized with two internal controls, GAPDH and β-actin. Graphs show the mean ± SEM. *p<0,05; **p<0,01 (Mann Whitney U test).
X-linked miRNA expression profile in blood leukocytes from CF male and female patients
Next, we sought to investigate by qRT-PCR the relative expression of a panel of X-linked miRNAs in blood leucocytes. Specifically, we wished to explore the expression profile of miR-223-3p, miR-106a-5p, miR-221-3p, miR-502-5p, miR-374a-5p, miR-20b-5p and miR-532-5p given their modulatory role, shown previously in different inflammatory settings (30–36), and also because their expression was found to be dysregulated in CF airway epithelial cells (23). We first analyzed all the patients regardless of sex. The expression level of miR-374a-5p, miR-20b-5p and miR-532-5p did not significantly change in CF patients compared to controls in either boys or girls (data not shown). However, different profile was obtained for the other tested miRNAs. We found a significant increase in the expression of miR-223-3p, miR-106a-5p, and miR-221-3p in both CF males and females compared to their sex-matched controls (Figure 3A). The expression of miR-502-5p is significantly higher only in CF males compared to controls (Figure 3A). We also evaluated the magnitude of the relative expression of these miRNAs in males and females, by considering the fold changes to their matched controls. As shown Figure 3B, we found no significant difference for miR-502 between males and females, whereas the overexpression of miR-221-3p is significantly higher in CF females than in CF males.
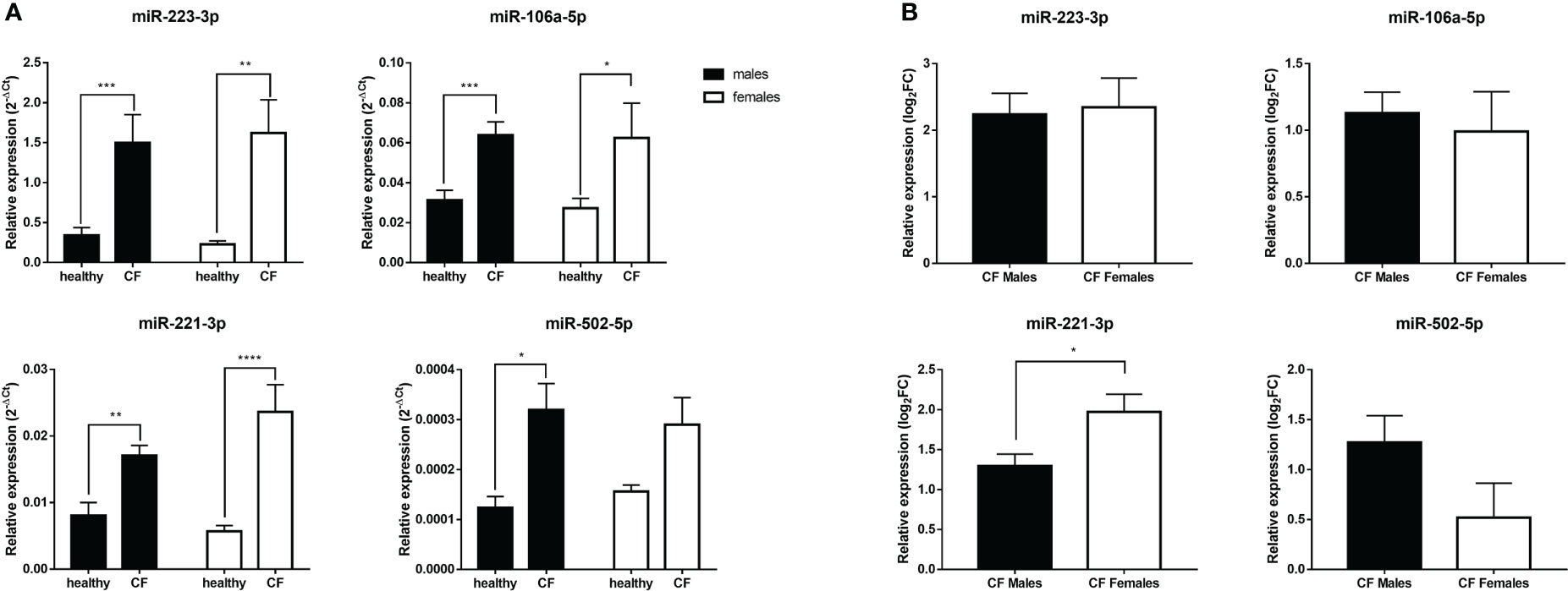
Figure 3 (A) Sex-based comparative study of the expression of X-linked miR-221-3p, miR-502-5p, miR-223-3p and miR-106a-5p in blood leukocytes of male healthy controls (n=10) versus CF males (n=15) and female healthy controls (n=10) versus CF female patients (n=14). The values (2-ΔCT) were normalized with the internal control SNORD95. (B) Sex-based comparative study of fold change values (log2(2-ΔΔCT)) of X-linked miR-221-3p, miR-502-5p, miR-223-3p and miR-106a-5p expression in blood leukocytes between CF males (n=15) and CF females (n=14). The fold change values (log2(2-ΔΔCT)) are relative to those from healthy male or female controls (n=10/group). Graphs show the mean ± SEM. *p<0,05; **p<0,01; ***p<0,001; ****p<0,0001 (Mann Whitney U test).
Next, we tested whether the overexpression of miR-221-3p in females could impact the expression of its known mRNA targets: A20 and PDLIM2, two ubiquitin-editing enzymes, and SOCS1 and SOCS3 that are inhibitors of JAK/STAT and NF-kB signaling pathways (32, 37–39). We also tested JAK3, a newly identified miR-221-3p target (40). The data of PDLIM2 and SOCS1 are shown in Figure 4. There is a substantial difference with a trend towards lower expression of SOCS1 (p=0,05) in CF females compared to CF males. Similar trend is observed for PDLIM2, while no obvious difference could be seen for A20, SOCS3 and JAK3 (data not shown)
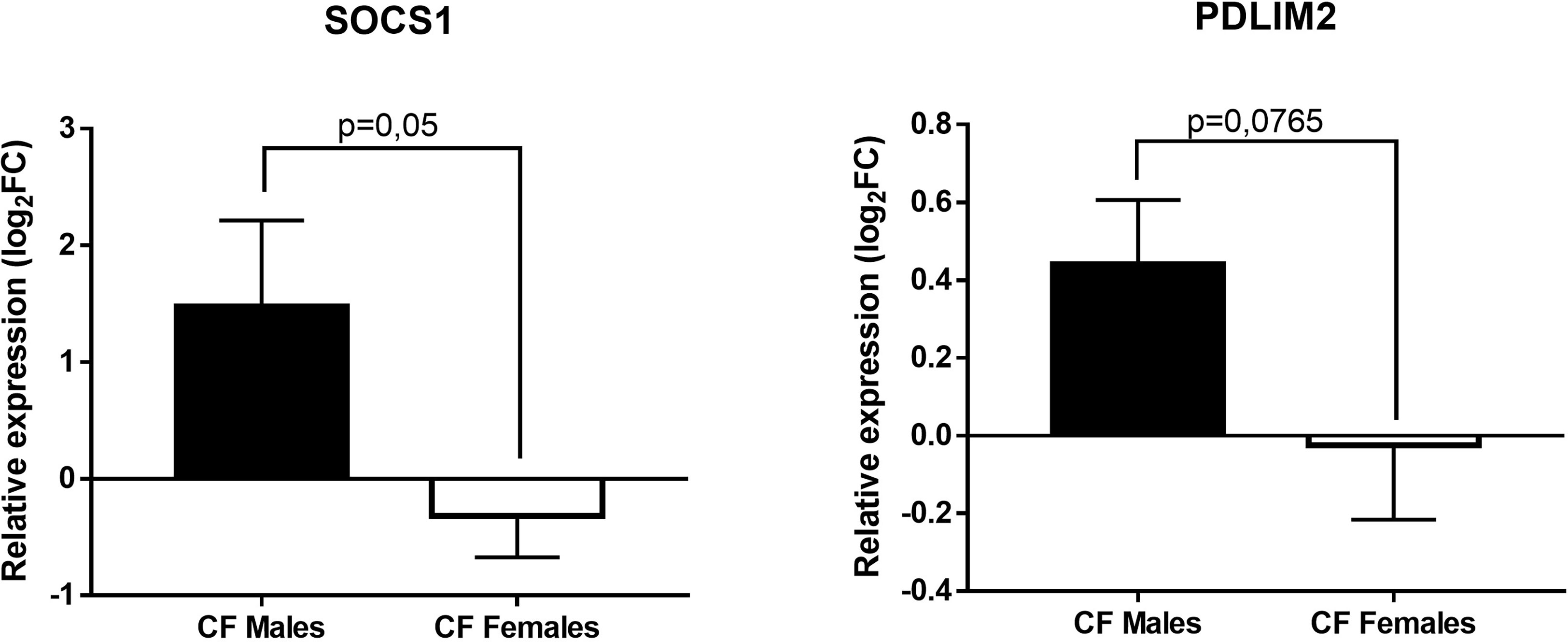
Figure 4 Sex-based comparative study of fold change values (log2(2-ΔΔCT)) of SOCS1 and PDLIM2 expression in blood leukocytes between CF males (n=15) and CF females (n=14). The fold change values (2-ΔΔCT) are relative to those from sex-matched healthy controls (n=10/group). Graphs show the mean ± SEM.
It has been reported that sex difference in inflammatory response in CF can be observed in infants under 8 years of age where the level of sex hormones is usually very low (3). We analyzed data in the subgroup of prepubertal infants (age <8 years) to see if there were any gender differences. As for the entire cohort, we observed that miR-221-3p is overexpressed in CF prepubertal females compared to males with the same trends of lower expression of the PDLIM2 and SOCS1 mRNA targets (Figure S1). Although the number of patients remains small, these results suggest that sex hormones are less likely to play a major role.
The overexpression of miR-221-3p in CF patients correlates positively with the upregulation of IL-1β transcript
To see whether the expression profile of miRNAs correlates with the inflammatory state of CF patients, we cross-analyzed the expression level of each miRNA with all tested cytokines and chemokines, both at protein and transcript levels. The only significant positive correlation was observed between miR-221-3p and IL-1β transcript (Figure 5), a finding consistent with the pro-inflammatory role of this microRNA. This seems also true when considering only prepubertal (age <8 years) boys and girls (Figure S2), suggesting that sex hormones may not have a great impact.
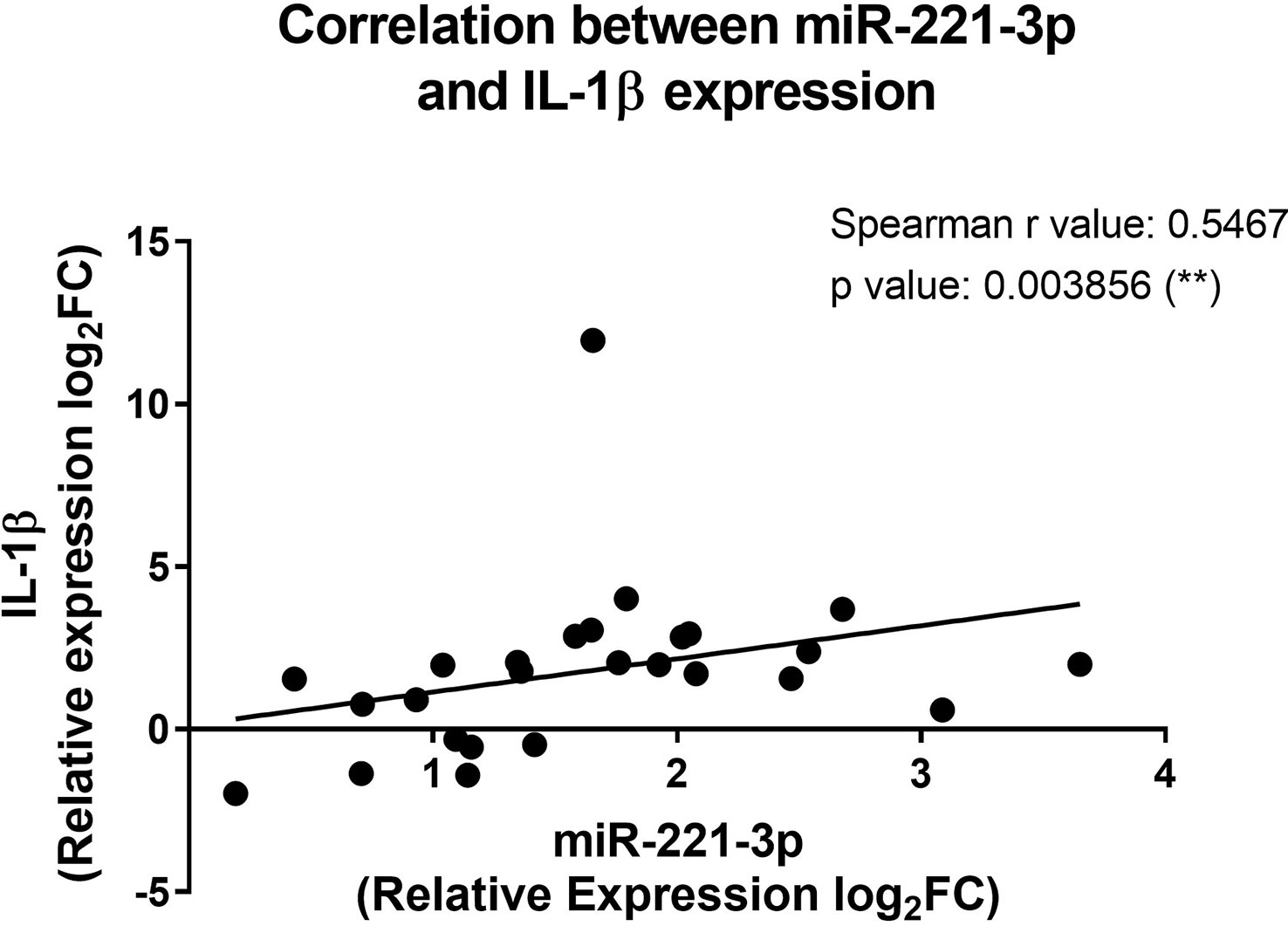
Figure 5 Correlation between the miR-221-3p and the IL-1β expression in blood leukocytes from CF patients. The fold change values (2-ΔΔCT) are relative to those from healthy controls (n=10/group). **p<0,01 (Spearman test).
Discussion
It is well documented that the biological sex is a contributing factor in the susceptibility to CF, with a better prognosis for boys compared to girls. However, there is paucity of data related to molecular markers that could be relevant to the mechanisms at work. In this study, we examined the potential contribution of selected X-linked miRNAs that are involved in the regulation of the inflammatory process and whose aberrant expression has been described in CF patients.
Despite the overall higher production of pro-inflammatory cytokines in CF compared with controls, patients enrolled in this study exhibited a relatively stable clinical state with respect to clinical parameters routinely recorded during medical monitoring. Similar observations have been previously reported in human and experimental CF (41–45). Currently, better therapeutic care that significantly improves the outcome of the disease is progressing thanks to the newborn screening allowing an early diagnosis. While gender difference still exists in CF, significant advances in timely treatment and patient follow-up, have largely contributed to reduce the difference observed in the pathophysiological evolution and inflammatory phenotype between boys and girls. However, substantial differences between males and females could be detected at the transcript level, in particular using signal amplification methods such as qRT-PCR.
Regarding miRNAs, we observed a significant increase in the expression of miR-223-3p, miR-106a-5p, miR-502-5p and miR-221-3p in blood cells from CF patients compared with healthy controls. The miR-223-3p is highly expressed by myeloid cells and is particularly involved in the control of granulopoiesis (46). In addition, we and others have previously shown that miR-223-3p is an essential modulator of the inflammation through the regulation of neutrophil activity and macrophage polarization (47–52). The overexpression of miR-223-3p observed in blood leukocytes of CF patients is in line with previous studies describing its upregulation in CF endobronchial brush samples (23) as well as in a CFTR gene-mutated bronchial epithelial cell line (53). Further studies are needed to depict the functional role of miR-223-3p in the inflammatory pathogenesis of CF. On the other hand, the miR-106a-5p is known to be involved in the control of monocytopoiesis (54) and different studies suggested its anti-inflammatory potential (31, 55). In CF, the expression of miR-106a was shown to be downregulated in endobronchial brush samples (23). In contrast, we detected a significant expression of miR-106a in blood leukocytes in our CF cohort. This discrepancy could be explained by differences in the samples used (endobronchial brush versus blood leukocytes), in patient’s clinical status or in the experimental design related to data normalization. It is likely that the increase in the expression level of miR-106a in the blood cells of patients with cystic fibrosis can be considered as one of the compensatory mechanisms to limit excessive inflammation. On the other hand, few studies addressed the role of miR-502-5p in inflammation. Zhang G et al. reported it potential role in inhibiting the IL-1β-induced NF-κB pathway in chondrocytes by targeting TRAF2 (33). The contribution of miR-502-5p in the inflammatory signaling pathways in CF remains unclear.
Notably, this study revealed that miR-221-3p is significantly overexpressed in blood cells from CF females compared to CF males. Similar trends were obtained when comparing prepubertal CF patients, suggesting a non-prominent role of sex hormones. However, this requires a large cohort and hormone assay for a firm conclusion. Since no significant difference were observed in leukocyte count and blood cell composition between genders, the overexpression of miR-221-3p seen in females would most likely be attributed to a differential cell activation and/or gene inactivation escape occurring naturally in one of the X chromosomes. The miR-221 was mainly investigated in cancer where studies have shown its potential contribution to tumor progression through modulation of cell proliferation, invasion, and apoptosis (56). Moreover, its aberrant expression was reported in inflammatory diseases as well, like in rheumatoid arthritis, atherosclerosis and asthma (57, 58). In CF, miR-221-3p was shown to be overexpressed in CF airway epithelial cells (59). Interestingly, we found that the overexpression of miR-221-3p in CF blood cells is positively correlated with the expression of IL-1β. We also observed a trend towards lower expression in CF females of SOCS1 and PDLIM2, two mRNA targets of miR-221-3p. Of note, it would be interesting to investigate how miR-221-3p could impact the expression of its main targets at protein level as well. Although the causative link between miR-221 and IL-1β-related inflammatory profile within blood cells derived from CF patients remains unclair, this observational clinical study is in line with previous experimental studies supporting the pro-inflammatory role for miR-221-3p by promoting the activation of the NF-κB signaling pathway (32, 37–39, 60).
Our investigation addresses for the first time the potential contribution of X-linked miRNAs derived from CF blood leukocytes in the sex-bias of the inflammatory response. The study has some limitations. We cannot rule out the role of CFTR gene mutation type that may have different functional consequences, ranging from mild to a severe disease phenotype (61). As mentioned elsewhere, although our results do not appear to be affected when considering only prepubertal patients, the cohort remains too small to rule out a potential impact of sex hormones on miRNA expression (62, 63). It would be interesting to investigate, in future studies, a large homogenous cohort that includes not only low to moderate clinical phenotypes but also cases with pulmonary exacerbation to examine whether the gender difference in miRNA expression may have an important impact on the inflammatory process. Collectively, this pilot study suggests the potential contribution of X-linked pro-inflammatory miR-221-3p in the sex-bias of the inflammatory process and highlights the need to unravel its role in the pathophysiology of CF and determine how this may help optimize the personal follow-up of patients.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by the hospital ethics committee of the Brugmann University Hospital (CE 2016/162). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
MD performed experiments, analyzed data, and wrote the first draft. AP patient recruitment and reviewed the manuscript. LH patient recruitment and reviewed the manuscript. NL patient recruitment and reviewed the manuscript. LAAN performed experiments. GSO performed experiments. FC reviewed the manuscript. GC reviewed the manuscript. MC supervised the work, analyzed data, and reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
We have received financial support from “IRIS research”.
Acknowledgments
We would like to thank “IRIS research” for their valuable support. MD is a PhD fellow of the Belgian Kids’ Fund. We also thank the David and Alice Van Burren Fund, the Jaumotte-Demoulin Foundation and the Héger-Masson Foundation. In addition, we thank the Réseau Mère-Enfant de la Francophonie (RMEF). We would also like to thank all the children and their parents who agreed to participate in the study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1114239/full#supplementary-material
References
1. Lyczak JB, Cannon CL, Pier GB. Lung infections associated with cystic fibrosis. Clin Microbiol Rev (2002) 15(2):194–222. doi: 10.1128/CMR.15.2.194-222.2002
2. Turcios NL. Cystic fibrosis lung disease: An overview. Respir Care (2020) 65(2):233–51. doi: 10.4187/respcare.06697
3. Casimir GJA, Mulier S, Hanssens L, Knoop C, Ferster A, Hofman B, et al. Chronic inflammatory diseases in children are more severe in girls. Shock (2010) 34(1):23–6. doi: 10.1097/SHK.0b013e3181ce2c3d
5. Demko CA, Byard PJ, Davis PB. Gender differences in cystic fibrosis: Pseudomonas aeruginosa infection. J Clin Epidemiol (1995) 48(8):1041–9. doi: 10.1016/0895-4356(94)00230-N
6. Harness-Brumley CL, Elliott AC, Rosenbluth DB, Raghavan D, Jain R. Gender differences in outcomes of patients with cystic fibrosis. J Women’s Health (2014) 23(12):1012–20. doi: 10.1089/jwh.2014.4985
9. Olesen HV, Pressler T, Hjelte L, Mared L, Lindblad A, Knudsen PK, et al. Gender differences in the Scandinavian cystic fibrosis population. Pediatr Pulmonol (2010) 45(10):959–65. doi: 10.1002/ppul.21265
10. Reid DW, Leigh Blizzard C, Shugg DM, Flowers C, Cash C, Greville HM. Changes in cystic fibrosis mortality in australia, 1979-2005. Med J Aust (2011) 195(7):392–5. doi: 10.5694/mja10.11229
11. Saint-Criq V, Harvey BJ. Estrogen and the cystic fibrosis gender gap. Steroids. (2014) 81:4–8. doi: 10.1016/j.steroids.2013.11.023
12. Lam GY, Goodwin J, Wilcox PG, Quon BS. Sex disparities in cystic fibrosis: Review on the effect of female sex hormones on lung pathophysiology and outcomes. ERJ Open Res (2021) 7(1):00475–2020. doi: 10.1183/23120541.00475-2020
13. Ober C, Loisel DA, Gilad Y. Sex-specific genetic architecture of human disease. Nat Rev Genet (2009) 9(12):911–22. doi: 10.1038/nrg2415
14. Di Palo A, Siniscalchi C, Salerno M, Russo A, Gravholt CH, Potenza N. What microRNAs could tell us about the human X chromosome. Cell Mol Life Sci (2020) 77(20):4069–80. doi: 10.1007/s00018-020-03526-7
15. Finotti A, Fabbri E, Lampronti I, Gasparello J, Borgatti M, Gambari R. MicroRNAs and long non-coding RNAs in genetic diseases. Mol Diagnosis Ther (2019) 23(2):155–71. doi: 10.1007/s40291-018-0380-6
16. Narożna B, Langwiński W, Szczepankiewicz A. Non-coding RNAs in pediatric airway diseases. Genes (Basel) (2017) 8(12):348. doi: 10.3390/genes8120348
17. Mckiernan PJ, Greene CM. MicroRNA dysregulation in cystic fibrosis. Mediators Inflamm (2015) 2015:1–7. doi: 10.1155/2015/529642
18. Greene CM. MicroRNA expression in cystic fibrosis airway epithelium. Biomolecules (2013) 3(1):157–67. doi: 10.3390/biom3010157
19. Soni DK, Biswas R. Role of non-coding RNAs in post-transcriptional regulation of lung diseases. Front Genet (2021) 12:1–18. doi: 10.3389/fgene.2021.767348
20. Sonneville F, Ruffin M, Guillot L, Rousselet N, Le Rouzic P, Corvol H, et al. New insights about miRNAs in cystic fibrosis. Am J Pathol (2015) 185(4):897–908. doi: 10.1016/j.ajpath.2014.12.022
21. Mitash N, Donovan JE, Swiatecka-Urban A. The role of microRNA in the airway surface liquid homeostasis. Int J Mol Sci (2020) 21(11):3848. doi: 10.3390/ijms21113848
22. Ramachandran S, Karp PH, Osterhaus SR, Jiang P, Wohlford-Lenane C, Lennox KA, et al. Post-transcriptional regulation of cystic fibrosis transmembrane conductance regulator expression and function by MicroRNAs. Am J Respir Cell Mol Biol (2013) 49(4):544–51. doi: 10.1165/rcmb.2012-0430OC
23. Oglesby IK, Bray IM, Chotirmall SH, Stallings RL, Neill SJO, Mcelvaney NG, et al. miR-126 is downregulated in cystic fibrosis airway epithelial cells and regulates TOM1 expression. J Immunol (2010) 184:1702–9. doi: 10.4049/jimmunol.0902669
24. Oglesby IK, Vencken SF, Agrawal R, Gaughan K, Molloy K, Higgins G, et al. MIR-17 overexpression in cystic fibrosis airway epithelial cells decreases interleukin-8 production. Eur Respir J (2015) 46(5):1350–60. doi: 10.1183/09031936.00163414
25. Bhattacharyya S, Balakathiresan NS, Dalgard C, Gutti U, Armistead D, Jozwik C, et al. Elevated miR-155 promotes inflammation in cystic fibrosis by driving hyperexpression of interleukin-8. J Biol Chem (2011) 286(13):11604–15. doi: 10.1074/jbc.M110.198390
26. McKiernan PJ, Molloy KP, Cryan SA, McElvaney NG, Greene CM. X Chromosome–encoded microRNAs are functionally increased in cystic fibrosis monocytes. Am J Respir Crit Care Med (2018) 197(5):668–70. doi: 10.1164/rccm.201707-1417LE
27. Mooney C, McKiernan PJ, Raoof R, Henshall DC, Linnane B, McNally P, et al. Plasma microRNA levels in male and female children with cystic fibrosis. Sci Rep (2020) 10(1):1–8. doi: 10.1038/s41598-020-57964-1
28. Carrel L, Willard HF. X-Inactivation profile reveals extensive variability in X-linked gene expression in females. Nat Publ Gr (2005) 434:400–4. doi: 10.1038/nature03479
29. Tukiainen T, Villani AC, Yen A, Rivas MA, Marshall JL, Satija R, et al. Landscape of X chromosome inactivation across human tissues. Nature (2017) 550(7675):244–8. doi: 10.1038/nature24265
30. Roffel MP, Bracke KR, Heijink IH, Maes T. miR-223: A key regulator in the innate immune response in asthma and COPD. Front Med (2020) 7(May). doi: 10.3389/fmed.2020.00196
31. Yang J, Chen Y, Jiang K, Yang Y, Zhao G, Guo S, et al. MicroRNA-106a provides negative feedback regulation in lipopolysaccharide-induced inflammation by targeting TLR4. Int J Biol Sci (2019) 15(11):2308–19. doi: 10.7150/ijbs.33432
32. Liu S, Sun X, Jiang Y, Liu Z, Cao X, Hou Y, et al. A miR-221/222-mediated feedback loop maintains constitutive activation of NF-κB and STAT3 signaling in human colorectal cancers. Gastroenterology (2014) 147(4):847–859.e11. doi: 10.1053/j.gastro.2014.06.006
33. Zhang G, Sun Y, Wang Y, Liu R, Bao Y, Li Q. MiR-502-5p inhibits IL-1β-induced chondrocyte injury by targeting TRAF2. Cell Immunol (2016) 302:50–7. doi: 10.1016/j.cellimm.2016.01.007
34. Yang Z, Guo Z, Dong J, Sheng S, Wang Y, Yu L, et al. miR-374a regulates inflammatory response in diabetic nephropathy by targeting MCP-1 expression. Front Pharmacol (2018) 9(AUG):1–8. doi: 10.3389/fphar.2018.00900
35. Lou J, Wang Y, Zhang Z, Qiu W. MiR-20b inhibits mycobacterium tuberculosis induced inflammation in the lung of mice through targeting NLRP3. Exp Cell Res (2017) 358(2):120–8. doi: 10.1016/j.yexcr.2017.06.007
36. Yan X, Zeng D, Yijing HZ, Shi ZY, Wu Y, Tang H, et al. Mirna-532-5p regulates cums-induced depression-like behaviors and modulates lps-induced proinflammatory cytokine signaling by targeting stat3. Neuropsychiatr Dis Treat (2020) 16:2753–64. doi: 10.2147/NDT.S251152
37. Zhao D, Zhuang N, Ding Y, Kang Y, Shi L. MiR-221 activates the NF-κB pathway by targeting A20. Biochem Biophys Res Commun (2016) 472(1):11–8. doi: 10.1016/j.bbrc.2015.11.009
38. Xu G, Yang F, Ding CL, Wang J, Zhao P, Wang W, et al. MiR-221 accentuates IFN’s anti-HCV effect by downregulating SOCS1 and SOCS3. Virology (2014) 462–463(1):343–50. doi: 10.1016/j.virol.2014.06.024
39. Wang T, Jiang L, Wei X, Dong Z, Liu B, Zhao J, et al. Inhibition of miR-221 alleviates LPS-induced acute lung injury via inactivation of SOCS1/NF-κB signaling pathway. Cell Cycle (2019) 18(16):1893–907. doi: 10.1080/15384101.2019.1632136
40. Quero L, Tiaden AN, Hanser E, Roux J, Laski A, Hall J, et al. miR-221-3p drives the shift of M2-macrophages to a pro-inflammatory function by suppressing JAK3/STAT3 activation. Front Immunol (2020) 10(1). doi: 10.3389/fimmu.2019.03087
41. Tang A, Sharma A, Jen R, Hirschfeld AF, Chilvers MA, Lavoie PM, et al. Inflammasome-mediated IL-1β production in humans with cystic fibrosis. PloS One (2012) 7(5):1–12. doi: 10.1371/journal.pone.0037689
42. Paats MS, Bergen IM, Bakker M, Hoek RAS, Nietzman-Lammering KJ, Hoogsteden HC, et al. Cytokines in nasal lavages and plasma and their correlation with clinical parameters in cystic fibrosis. J Cyst Fibros (2013) 12(6):623–9. doi: 10.1016/j.jcf.2013.05.002
43. Litman PM, Day A, Kelley TJ, Darrah RJ. Serum inflammatory profiles in cystic fibrosis mice with and without bordetella pseudohinzii infection. Sci Rep (2021) 11(1):1–11. doi: 10.1038/s41598-021-97033-9
44. Bonfield TL, Panuska JR, Konstan MW, Hilliard KA, Hilliard JB, Ghnaim H, et al. Inflammatory cytokines in cystic fibrosis lungs. Am J Respir Crit Care Med (1995) 152:2111–8. doi: 10.1164/ajrccm.152.6.8520783
45. Eickmeier O, Huebner M, Herrmann E, Zissler U, Rosewich M, Baer PC, et al. Sputum biomarker profiles in cystic fibrosis (CF) and chronic obstructive pulmonary disease (COPD) and association between pulmonary function. Cytokine (2010) 50(2):152–7. doi: 10.1016/j.cyto.2010.02.004
46. Johnnidis JB, Harris MH, Wheeler RT, Stehling-sun S, Lam MH, Kirak O, et al. Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature (2008) 451(7182):1125–9. doi: 10.1038/nature06607
47. Dorhoi A, Iannaccone M, Farinacci M, Faé KC, Schreiber J, Moura-Alves P, et al. MicroRNA-223 controls susceptibility to tuberculosis by regulating lung neutrophil recruitment. J Clin Invest (2013) 123(11):4836–48. doi: 10.1172/JCI67604
48. Deny M, Romano M, Denis O, Casimir G, Chamekh M. Progressive control of streptococcus agalactiae-induced innate inflammatory response is associated with time course expression of MicroRNA-223 by neutrophils. Infect Immun (2020) 88(12):1–12. doi: 10.1128/IAI.00563-20
49. Feng Z, Qi S, Zhang Y, Qi Z, Yan L, Zhou J, et al. Ly6G+ neutrophil-derived miR-223 inhibits the NLRP3 inflammasome in mitochondrial DAMP-induced acute lung injury. Cell Death Dis (2017) 8(11):e3170. doi: 10.1038/cddis.2017.549
50. Zhuang G, Meng C, Guo X, Cheruku PS, Shi L. A novel regulator of macrophage activation miR-223 in obesity-associated adipose tissue inflammation. Circulation (2012) 125(23):2892–903. doi: 10.1161/CIRCULATIONAHA.111.087817
51. Ying W, Tseng A, Chang RCA, Morin A, Brehm T, Triff K, et al. MicroRNA-223 is a crucial mediator of PPARγ-regulated alternative macrophage activation. J Clin Invest (2015) 125(11):4149–59. doi: 10.1172/JCI81656
52. Deny M, Arroba Nuñez LA, Romano M, Denis O, Casimir G, Chamekh M. Sex difference in innate inflammatory response and macrophage polarization in streptococcus agalactiae-induced pneumonia and potential role of microRNA-223-3p. Sci Rep (2022) 12(1):1–14. doi: 10.1038/s41598-022-21587-5
53. Oglesby IK, Chotirmall SH, McElvaney NG, Greene CM. Regulation of cystic fibrosis transmembrane conductance regulator by MicroRNA-145, -223, and -494 is altered in ΔF508 cystic fibrosis airway epithelium. J Immunol (2013) 190(7):3354–62. doi: 10.4049/jimmunol.1202960
54. Fontana L, Pelosi E, Greco P, Racanicchi S, Testa U, Liuzzi F, et al. MicroRNAs 17-5p-20a-106a control monocytopoiesis through AML1 targeting and m-CSF receptor upregulation. Nat Cell Biol (2007) 9(7):775–87. doi: 10.1038/ncb1613
55. Hong Z, Hong H, Liu J, Zheng X, Huang M, Li C, et al. MIR-106a is downregulated in peripheral blood mononuclear cells of chronic hepatitis b and associated with enhanced levels of interleukin-8. Mediators Inflamm (2015) 2015:629862. doi: 10.1155/2015/629862
56. Garofalo M, Quintavalle C, Romano G, M. Croce C, Condorelli G. miR221/222 in cancer: Their role in tumor progression and response to therapy. Curr Mol Med (2011) 12(1):27–33. doi: 10.2174/156652412798376170
57. Song J, Ouyang Y, Che J, Li X, Zhao Y, Yang K, et al. Potential value of miR-221/222 as diagnostic, prognostic, and therapeutic biomarkers for diseases. Front Immunol (2017) 8(FEB):1–9. doi: 10.3389/fimmu.2017.00056
58. Qin HB, Xu B, Mei JJ, Li D, Liu JJ, Zhao DY, et al. Inhibition of miRNA-221 suppresses the airway inflammation in asthma. Inflammation (2012) 35(4):1595–9. doi: 10.1007/s10753-012-9474-1
59. Oglesby IK, Agrawal R, Mall MA, McElvaney NG, Greene CM. miRNA-221 is elevated in cystic fibrosis airway epithelial cells and regulates expression of ATF6. Mol Cell Pediatr (2015) 2(1):1–8. doi: 10.1186/s40348-014-0012-0
60. Nejad C, Stunden HJ, Gantier MP. A guide to miRNAs in inflammation and innate immune responses. FEBS J (2018) 285(20):3695–716. doi: 10.1111/febs.14482
61. Marson FAL, Bertuzzo CS, Ribeiro JD. Classification of CFTR mutation classes. Lancet Respir Med (2016) 4(8):e37–8. doi: 10.1016/S2213-2600(16)30188-6
62. Sharma S, Eghbali M. Influence of sex differences on microRNA gene regulation in disease. Biol Sex Differ (2014) 5(1):1–8. doi: 10.1186/2042-6410-5-3
Keywords: cystic fibrosis, miRNA- microRNA, sex, inflammation, X chromosome (human)
Citation: Deny M, Popotas A, Hanssens L, Lefèvre N, Arroba Nuñez LA, Ouafo GS, Corazza F, Casimir G and Chamekh M (2023) Sex-biased expression of selected chromosome x-linked microRNAs with potent regulatory effect on the inflammatory response in children with cystic fibrosis: A preliminary pilot investigation. Front. Immunol. 14:1114239. doi: 10.3389/fimmu.2023.1114239
Received: 02 December 2022; Accepted: 22 March 2023;
Published: 03 April 2023.
Edited by:
Guoshun Wang, Louisiana State University Health Sciences Center, United StatesReviewed by:
Roopa Biswas, Uniformed Services University of the Health Sciences, United StatesValeria Rachela Villella, University of Naples Federico II, Italy
Copyright © 2023 Deny, Popotas, Hanssens, Lefèvre, Arroba Nuñez, Ouafo, Corazza, Casimir and Chamekh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mustapha Chamekh, TW9zdGFmYS5jaGFtZWtoQHVsYi5iZQ==