
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 06 February 2023
Sec. Vaccines and Molecular Therapeutics
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1101366
 Véronique Corbière1*
Véronique Corbière1* Eleonora E. Lambert2
Eleonora E. Lambert2 Marine Rodesch3
Marine Rodesch3 Jacqueline A. M. van Gaans-van den Brink2
Jacqueline A. M. van Gaans-van den Brink2 Alicja Misiak4Elles Simonetti5,6Anne Van Praet1Audrey Godefroid1
Alicja Misiak4Elles Simonetti5,6Anne Van Praet1Audrey Godefroid1 Dimitri A. Diavatopoulos5,6
Dimitri A. Diavatopoulos5,6 Cécile A. C. M. van Els2,7Françoise Mascart1 and PERISCOPE WP5 Task 7 working group
Cécile A. C. M. van Els2,7Françoise Mascart1 and PERISCOPE WP5 Task 7 working groupIntroduction: The characterization of B. pertussis (Bp) antigen-specific CD4+ T cell cytokine responses should be included in the evaluation of immunogenicity of pertussis vaccines but is often hindered by the lack of standardized robust assays.
Methods: To overcome this limitation, we developed a two-step assay comprising a short-term stimulation of fresh whole blood with Bp antigens and cryopreservation of the stimulated cells, followed later on by batch-wise intracellular cytokine analysis by flow cytometry. Blood samples collected from recently acellular (aP) vaccine boosted subjects with a whole-cell- or aP-primed background was incubated for 24 hrs with Pertussis toxin, Filamentous hemagglutinin or a Bp lysate (400µl per stimulation). Antigen-specific IFN-γ-, IL-4/IL-5/IL-13-, IL-17A/IL-17F- and/or IL-22-producing CD4+ T cells were quantified by flow cytometry to reveal Th1, Th2, and Th17-type responses, respectively. The frequencies of IFN-γ-producing CD8+ T cells were also analyzed.
Results: We demonstrate high reproducibility of the Bp-specific whole blood intracellular staining assay. The results obtained after cryopreservation of the stimulated and fixed cells were very well correlated to those obtained without cryopreservation, an approach used in our previously published assay. Optimization resulted in high sensitivity thanks to very low non-specific backgrounds, with reliable detection of Bp antigen-specific Th1, Th2 and Th17-type CD4+ T cells, in the lowest range frequency of 0.01-0.03%. Bp antigen-specific IFN-γ+ CD8+ T lymphocytes were also detected. This test is easy to perform, analyse and interpret with the establishment of strict criteria defining Bp antigen responses.
Discussion: Thus, this assay appears as a promising test for evaluation of Bp antigen-specific CD4+ T cells induced by current and next generation pertussis vaccines.
Pertussis, a bacterial respiratory disease caused by Bordetella pertussis (Bp) (1), remains one of the most uncontrolled vaccine-preventable disease despite high vaccine coverage (2). Therefore, new vaccine strategies are urgently required. Whole-cell pertussis (wP) vaccines, first implemented in the 1940s, were replaced in most industrialized countries in the 1990s-2000s by less reactogenic, acellular pertussis (aP) vaccines containing highly purified Bp antigens adsorbed to aluminum hydroxide as adjuvant. Despite clinical efficacy of aP vaccines against disease (3), pertussis outbreaks have been reported in countries where aP vaccines were implemented (4–6), with increases in pertussis cases in aP-primed adolescents (7). Fast waning of immunity as well as a suboptimal quality of immune responses induced by aP vaccines have been suggested to contribute to pertussis resurgence (8). In addition, while aP vaccines protect against symptomatic disease, baboon studies have demonstrated that they do not prevent asymptomatic carriage and transmission of pertussis (9). In contrast, wP-vaccinated non-human primates cleared the infection more rapidly than control animals (9), stressing the importance of the development of next generation vaccines with detailed characterization of the immune responses they induce in addition to demonstrating their protective efficacy.
While serum antibodies are the most currently used biomarker for new vaccine evaluation, the role of CD4+ T lymphocytes in protection against pertussis is now largely accepted (10). First described in mouse models (10, 11), the role of antigen-specific IFN-γ-producing CD4+ T lymphocytes (Th1) in protection was further demonstrated in a non-human primate model (12). Whereas Th1 responses are also induced in humans following infection and immunization with wP vaccines, aP vaccines induce a Th2-dominated CD4+ T cell response that is characterized by production of IL-4, IL-5 and/or IL-13 (13–17). In addition, the induction and protective role of IL-17-producing CD4+ T cells (Th17) have been demonstrated in several animal models after wP vaccination and pertussis infection (18–21). Consequently, not only Th1 and Th2-type but also Th17-type CD4+ T cell responses need to be evaluated during vaccine studies. Finally, in addition to Bp-specific CD4+ T cells, CD8+ T lymphocytes were also reported to participate in the IFN-γ response to Bp antigens (15, 22–25). Even though the significance of Bp-specific CD8+ T cell response is to our knowledge still unknown, their characterization may merit attention.
Currently available T cell immunoassays comprise proliferation assays, measurement of released cytokine concentrations by ELISA, enumeration of the cytokine-secreting cells by ELISPOT or detection of cytokine-containing T cells by multiparameter flow cytometry (FC). These assays are most often performed on cryopreserved peripheral blood mononuclear cells (PBMC) with inherent potential artefacts induced by cell isolation and freezing/thawing procedures, which may affect the proportion, phenotype and functions of cells, especially of effector cells (26–29). Moreover, using frozen cells for stimulation may compromise the optimal presentation of proteins by antigen-presenting cells to CD4+ T lymphocytes (30). Additionally, some of these techniques require long in vitro stimulation times with the antigens (15, 16, 31), potentially inducing further artefacts, and, except for FC, they do not allow a characterization of the cytokine-producing cells at the cellular level.
To overcome these limitations, we recently developed a whole blood assay (32), which includes a short in vitro stimulation step of whole blood (WB) with Bp antigens, followed by intracellular cytokine staining (ICS) to assess Th1, Th2 and Th17-type cytokine-producing and activated T lymphocyte populations specific for Bp antigens. This technique was successfully used for the characterization of human antigen-specific CD4+ T cell responses in a booster vaccination study (33). However, in a clinical study setting, antigen stimulation of fresh blood samples followed by antibody staining of cells and FC analysis is logistically challenging and does not allow for simultaneous measurements of longitudinal samples from the same individual.
To address these limitations, we introduced a cryopreservation step after stimulation of whole blood with Bp antigens, based on an approach that was successfully developed to characterize mycobacteria-specific CD4+ T cell responses (34, 35). We further optimized and validated this assay on fresh blood samples from recently vaccinated subjects to demonstrate feasibility and accuracy to detect and characterize low Bp antigen-specific Th1/Th2/Th17 CD4+ T cell responses.
This study was approved by the Ethics Committee ULB-Hôpital Erasme (aggregation number OMO21, study protocol P2018/515), and all the subjects gave their written informed consent.
Blood samples were collected from 12 wP-primed adults and two 5-6-years-old aP-primed children after the administration of an aP vaccine booster. Nine adults were recently boosted (between D13 and D37) whereas the three others received the boost some years before the blood sampling, with a maximum of 6 years (Boostrix®, GSK Biologicals, Belgium). The two children were recently boosted (between 1-2 months; Tetravac™ -Sanofi Pasteur, Lyon, France). Mean age of the adults was 33 years (21-52 years in the range), whereas the children were 5-6 years-old. Venous blood samples were collected in sodium heparin tubes (BD Biosciences, Erembodegem, Belgium) and transported directly to the laboratory.
Pertussis toxin (PT) and filamentous hemagglutinin (FHA), the two Bp antigens present in all aP vaccines (36), were selected for in vitro stimulation of blood cells in the Bp-specific whole blood intracellular staining (BpWB-ICS) assay to expand vaccine-induced Bp-specific precursor T lymphocytes. A Bp lysate (BPL) was also used to detect cellular immune responses to Bp antigens that are not included in aP vaccines. A genetically detoxified PT mutant, with an inactivated S1 subunit (R9K, E129A), was chosen for the in vitro T cell assay (LIST Biological laboratories Inc., Campbell, CA, USA, #184) to avoid the toxic activity on target cells (37). PT was resuspended following the manufacturer’s instructions and stored in aliquots at 500 µg/mL at 4°C. The endotoxin content determined by a kinetic chromogenic LAL assay was 0.02 EU/µg. Prior to use, each aliquot of PT was heat-inactivated at 80°C during 10 minutes using a Polymerase Chain reaction block, as recommended to abolish in vitro mitogenic activity of the B oligomer on T lymphocytes (16, 31, 37). Heat-inactivated aliquots were kept at 4°C until use. FHA was kindly provided by Sanofi Pasteur (Marcy-l’Étoile, France) and stored at 700 µg/mL at 4°C. The endotoxin content was below the detection limit of 0.005 EU/µg FHA. The BPL of strain B1917 (38) was kindly provided by Q Biologicals (Ghent, Belgium) and stored at 3.4 mg/mL at -80°C. A Bp sonicate, initially used for assay optimization before the availability of the BPL, was kindly provided by K. Mills (TCD, Dublin, Ireland) and stored at 596 µg/mL at -80°C. BPL and BP sonicate were heat-inactivated as described above for PT. All Bp antigens were aliquoted in low-binding tubes (Maxymum recovery from Axygen, VWR, Leuven, Belgium) for long-term storage. TT used to assess the intra-assay reproducibility, was purchased from Calbiochem (Sigma-Aldrich, Bornem, Belgium) and stored at 200 µg/mL at -20°C. As a positive control, Staphylococcus aureus enterotoxin B (SEB) was purchased from Sigma (Sigma-Aldrich) and stored at 1 mg/mL at -20°C.
Blood samples were processed within 3 hrs maximum of their collection (median of 30 min, 25th percentile - 75th percentile (P25-P75): 18-60 min, range: 15-180 min), consistent with other studies that recommended a short delay between blood collection and processing (34, 39). Freshly drawn whole blood samples were diluted 1:1 with RPMI 1640 medium (Invitrogen/ThermoFisher, Merelbeke, Belgium), supplemented with 40 µg/ml gentamycin (Invitrogen/ThermoFisher). Costimulatory anti-CD28 and anti-CD49d antibodies (clones L293 and L25.3, respectively, BD Biosciences, Mountain View, CA) were added to the diluted blood, each at a final concentration of 1 µg/mL to increase cytokine expression in antigen-specific T cells (34). Two-fold diluted blood was distributed in 15 ml round-bottom polypropylene tubes, each of which containing 800 µl of diluted blood (corresponding to 400 µl original blood volume) and incubated in the absence (negative control) or presence of antigen, i.e. 5 µg/mL PT, 5 µg/mL FHA, 10 µg/mL BPL, 10 µg/mL TT, 10 µg/mL Bp sonicate or 1 µg/mL SEB. The optimal antigen concentrations were either based on the literature (BP sonicate, TT) or on preliminary experiments from the PERISCOPE Consortium (PT, FHA, BPL). To assess intra-assay variability, some stimulations were performed in duplicate. Tubes were loosely covered by caps and incubated for 24 hrs at 37°C and 5% CO2. During the last 4 to 5 hrs of incubation, 10 µg/ml Brefeldin-A (Sigma-Aldrich) together with 1/1,000 Monensin (BD Biosciences, Erembodegem, Belgium) was added to each tube to inhibit cellular protein transport (26). Cells were subsequently pelleted at 500 x g for 10 min.
The protocol was adapted from the BD protocol (Alternative Protocol, Activation and Intracellular Staining of Whole Blood, BD Biosciences). Briefly, erythrocytes were lysed by adding 10 mL BD PharmLyse (BD Biosciences) to the pelleted cells for 10 min at room temperature (RT) in the dark. After washing with phosphate buffered saline (PBS, Westburg – Lonza, The Netherlands), cells were centrifuged at 500 x g for 10 min, the supernatants were discarded, and the white blood cells were fixed by incubation with 1 mL of the Fixation/Permeabilisation solution (from the BD Cytofix/Cytoperm Fixation/Permeabilisation kit, BD biosciences) during 20 min at RT in the dark. After centrifugation at 500 x g for 5 min, the white blood cells were cryopreserved in 500 µl Recovery Cell Culture Freezing medium (ThermoFisher), a formulation based on Dulbecco’s Modified Eagle Medium (High Glucose) with optimized levels of foetal bovine serum, bovine serum and 10% DMSO per condition. Cells were transferred into cryovials transferred into cryovial, before storage at -80°C for delayed analysis by batches. In case of direct staining and flow cytometry acquisition, the white blood cells were directly permeabilised by incubation with 2 mL of the Perm/Wash solution (from the BD Cytofix/Cytoperm Fixation/Permeabilisation kit, BD biosciences) for 10 min at RT in the dark.
Cryovials containing the stimulated, fixed and frozen white blood cells were retrieved from the -80°C freezer and thawed in a water bath at 37°C for 2 min. Thawed cells were transferred from cryovials to 15 mL conical tubes containing 5 mL PBS. After centrifugation at 500 x g for 5 min, the cells were permeabilized by adding 2 mL of the Perm/Wash solution (BD Biosciences) and incubated at RT for 10 min.
After permeabilization, either directly on freshly stimulated cells or after cryopreservation of the stimulated and fixed cells, surface and intracellular antibody staining of the cells was combined and performed in the presence of Perm/Wash buffer during 30 min at RT in the dark. Cells were stained with one of the two monoclonal antibody panels; panel 1: CD3-PcP, CD4-APC H7, CD8-PE, CD14/CD19-V500, IFN-γ-BV421, IL-5/IL-13-APC, IL-17A-FITC (Details see Table 1, panel 1); or panel 2: CD3-APC-H7, CD4-BB515, CD8-PerCP-Cy5.5, IFN-γ-BV421, IL-4-APC, IL-5-APC, IL-13-APC, IL-17A-PE, IL-17F-PE, IL-22-PE-Cy7 (Details see Table 1, panel 2). The antibody panel 1 was optimised in order to improve the detection of Th1 and Th17-type responses. After staining, cells were washed with 2 mL Perm/Wash buffer, transferred in FACS tubes (BD Biosciences), and washed again with PBS before they were recovered in FACS Flow buffer (BD Biosciences) for acquisition.
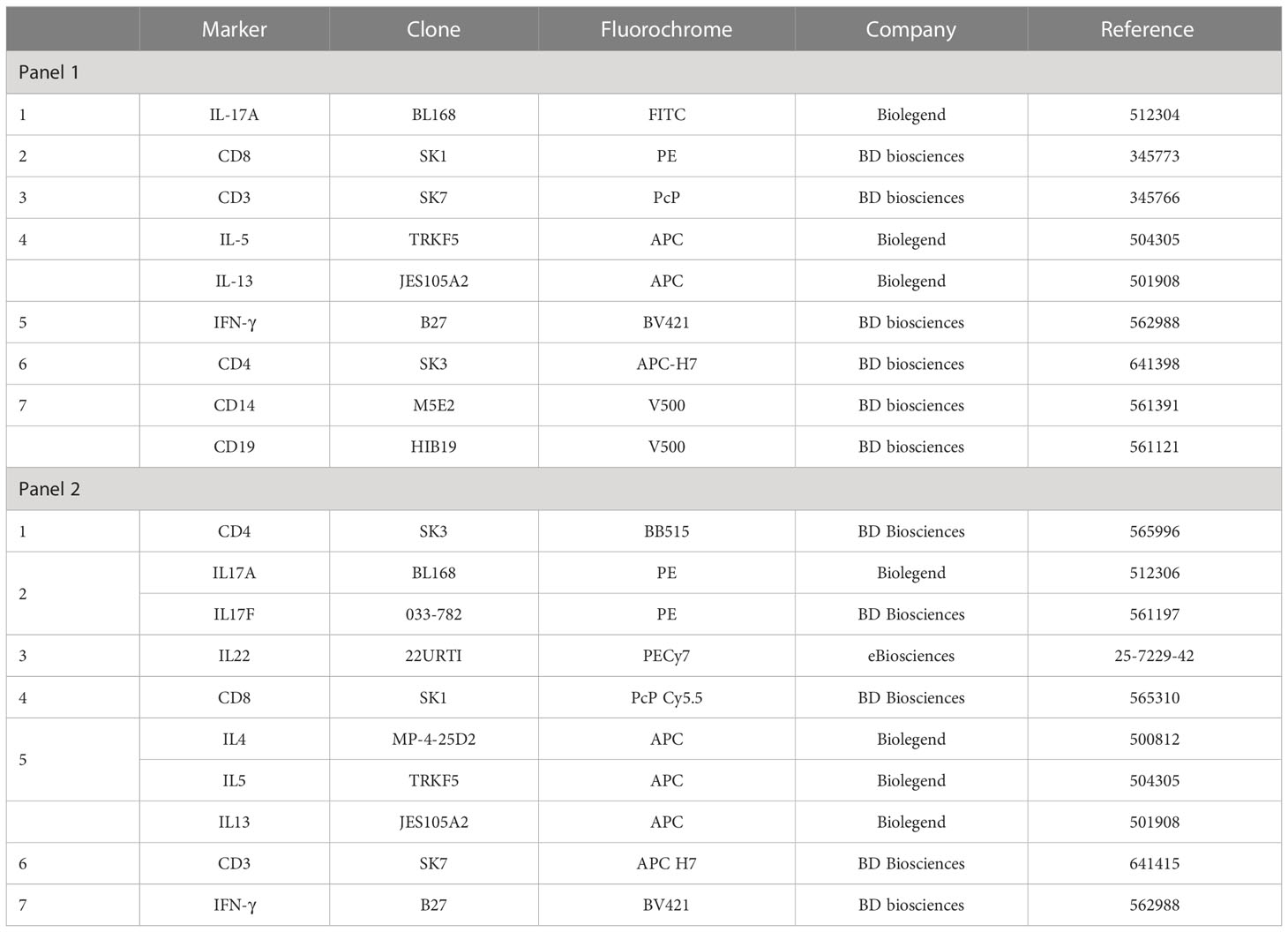
Table 1 Antibody clones and fluorochromes.
Acquisition was initially performed on a FACSCanto II flow cytometer (BD Biosciences) to assess the intra-assay reproducibility and a BD-LSR Fortessa flow cytometer (BD Biosciences) was used subsequently for the next experiments. The content of the tube was completely acquired. Compensations were performed using Comp Beads (BD Bioscience) tubes individually stained with each fluorophore, and compensation matrices were calculated with FACSDiva. Cytometer Setting and Tracking (CST) beads (BD Biosciences) were acquired before each experiment to ensure that cytometer parameters remained consistent across all experiments.
All flow cytometry data analyses were performed with FlowJo software (version 9.5.3, Tree Star, Ashland, OR USA) by using sequential gating. Preliminary experiments using a live/dead dye demonstrated that most cells were alive after the stimulation, and successive gates were applied to exclude dead cells. The gating strategy shown in Supplementary Figure 1 is an example of cryopreserved cells stained with antibody panel 2. Briefly, after selection of a time of homogenous acquisition, lymphocytes were determined according to their size and granularity. After doublet exclusions and CD3+ T cells selection, CD4+ CD8- T cells were analysed for their content in cytokines. The numbers of acquired events and percentages of CD4+ T cells that produce IFN-γ, IL-4/IL-5/IL-13, IL-17A/IL-17F or IL-22 were reported. The numbers of events recorded in the CD3 gate were 264,500 (median with range: 97,396-533,000) and 344,000 (median with range: 96,432-577,000) for the samples stained freshly and after cryopreservation, respectively. For CD4+ T lymphocytes, these numbers were 119,000 (median with range: 43,536-346,000) and 144,500 (median with range: 53,800-377,000), respectively. CD8+ T lymphocytes were analysed for their content in IFN-γ, and the numbers of CD8 events were 72,973 (median, range: 17,647-149,000) and 89,034 (median, range: 23,288-164,000), respectively. The numbers of acquired events were thus higher after cryopreservation within the CD3, CD4 and CD8 gates (p=0.0001, p=0.0003, p=0.0004, respectively).
The non-specific backgrounds were defined as the percentage of cytokine-producing CD4+ or CD8+ T lymphocytes among cells incubated in the absence of antigen. Percentages of antigen-specific cytokine-producing CD4+ and IFN-γ-producing CD8+ T cells were determined after the non-specific background values were subtracted.
To determine whether a Bp antigen-specific T cell response was relevant, we first compared the results obtained for the stimulated conditions to those from unstimulated condition by checking carefully the Flowjo analyses. The percentages of cytokine-containing cells in a diluted whole blood sample after Bp antigen stimulation were arbitrarily defined as significant i.e. different from the non-specific background, based on two criteria: a stimulation index (SI, percentage of cytokine positive cells in Bp antigen-stimulated condition/percentage of cytokine positive cells in the absence of antigen) ≥ 2, and a percentage of Bp antigen-specific cells (i.e. after subtraction of the non-specific background) ≥ 0.030% (Table 2). Both a high number of events acquired in the parent gate and a low background in the absence of antigen allowed us to consider low percentages of positive cells in stimulated conditions to be antigen-specific. Results were considered doubtful in two cases (1): a SI > 2 with a percentage of positive cells between 0.010–0.030%, and (2) a SI between 1.5 and 2, with a percentage of positive cells ≥ 0.010%. Results were always considered negative in case of a percentage of positive cells lower than 0.010%. These criteria were validated by checking back the Flowjo analyses.
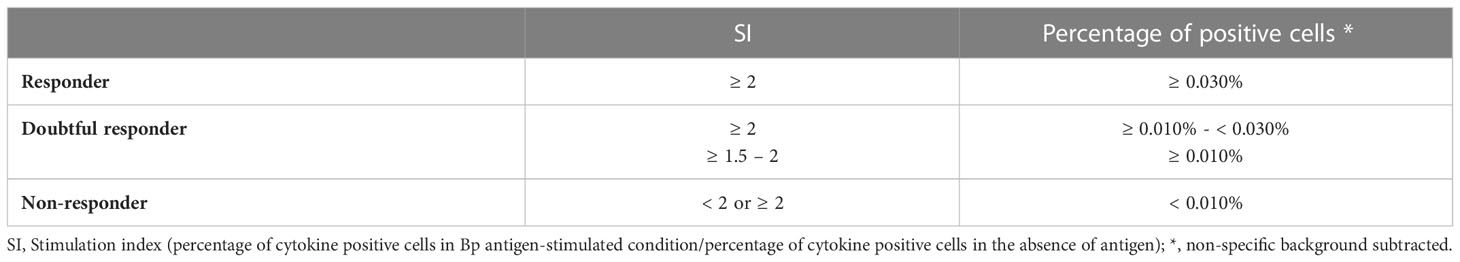
Table 2 Criteria to define responders and doubtful responders to Bp antigens.
Subjects with clearly detectable or doubtful percentages of Bp antigen-specific T cells were identified as responders or doubtful responders to that antigen, respectively. Subjects were considered as non-responders when a negative result was recorded.
Graphpad Prism 7.03 for Windows (Graphpad software, La Jolla, CA, USA) was used for statistical analysis. Correlations were evaluated by a non-parametric Spearman test. Wilcoxon matched-pairs signed rank test was applied to compare the results obtained for paired samples. Friedman’s test was used to compared three or more paired groups, with Dunn’s test as post hoc analysis to compare two conditions. A value of p<0.05 was considered significant. *, p<0.05; **, p<0.01.
In order to examine the intra-assay reproducibility of the BpWB-ICS assay, blood samples from five adults (n°1 to n°5) was divided into two aliquots and processed separately from the incubation step until the direct staining of the cells, to detect intracellular cytokines within CD4+ T cells (Figure 1, Table 1, panel 1). The percentages of IFN-γ- or IL-17A-producing CD4+ T cells obtained for the duplicates were measured under unstimulated and stimulated conditions with antigens (PT, FHA, Bp lysate, Bp sonicate, TT or SEB).
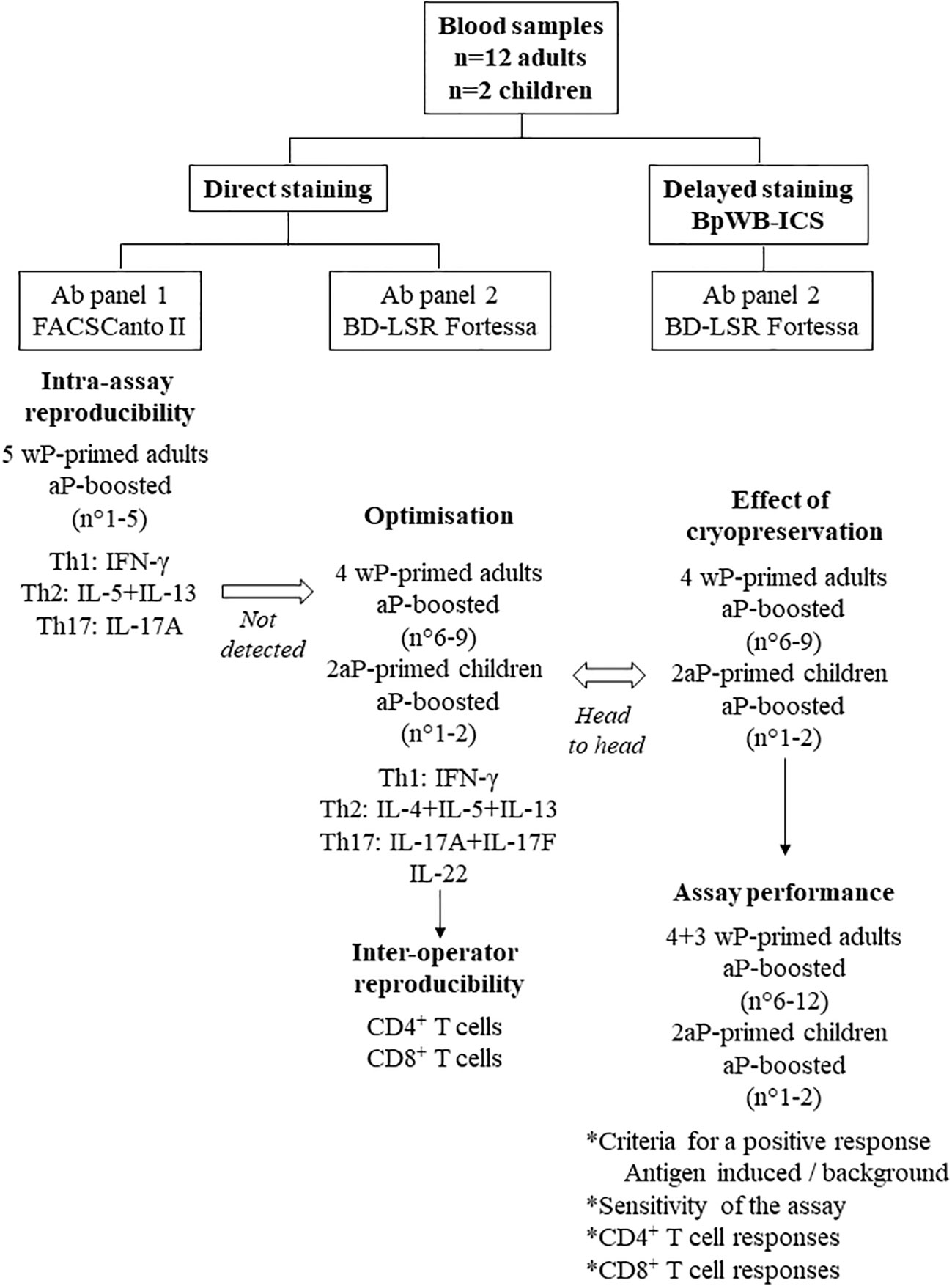
Figure 1 Flow chart of the study with the different steps of development of the BpWB-ICS. Blood samples were collected from 12 adults and from two children. The adults were all primed with a whole-cell pertussis vaccine (wP), whereas the children were primed with an acellular pertussis vaccine (aP). All the subjects were aP boosted. The intra-assay reproducibility was first evaluated on blood samples from five adults, using a direct staining of the Bp-antigen-stimulated cells. As antigen-specific Th2 cells were not detected, further optimisation was performed on blood samples from four adults and from two children. The antibody panel 2 was determined, a BD-LSR Fortessa replaced the FACSCanto II, and the inter-operator reproducibility was evaluated. Results obtained for the same blood samples were analysed head to head without and after a cryopreservation step to develop the BpWB-ICS with a delayed staining of the cells. This allowed us to further analyse the assay performance of the BpWB-ICS.
No Th2-containing CD4+ T cells were detected whereas the percentages of antigen-induced IFN-γ- or IL-17A-producing CD4+ T reached 0.604% and 0.034% in the median, respectively (P25-P75: 0.097%-3.18% for IFN-γ and 0.010%-0.118% for IL17-A). Results obtained in the absence of antigen were very low (0.011% and 0.004% in the median, for IFN-γ and IL-17A, respectively). The results obtained for the two different aliquots of blood were very well correlated (r=0.980, p<0.0001 for IFN-γ and r=0.958, p<0.0001 for IL-17A (Figure 2). The coefficients of variation (CVs) of the results obtained for the duplicates were 16% for IFN-γ and 22% for IL-17A, indicating that the precision of the assay may be considered as good (40, 41).
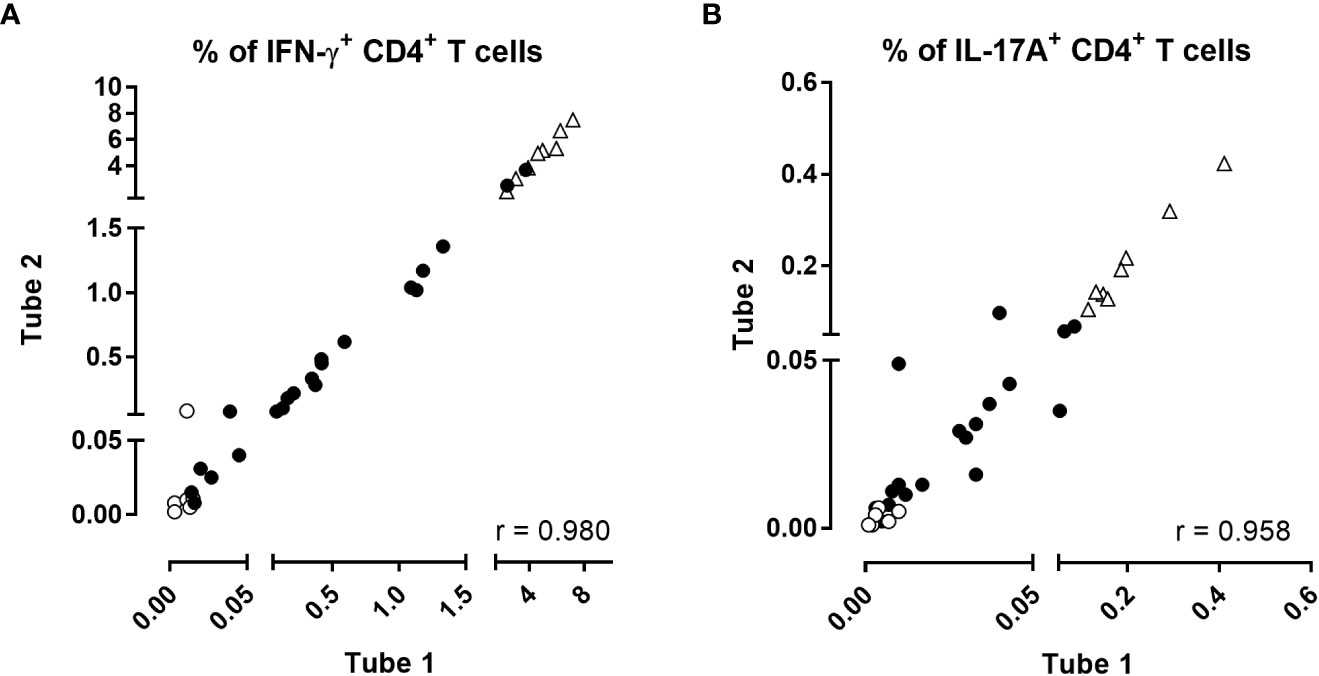
Figure 2 Intra-assay reproducibility of the BpWB-ICS assay. The percentages of (A) IFN-γ- and (B) IL-17A-producing CD4+ T cells obtained after direct staining of two separate aliquots of blood were compared two by two for each stimulation condition. Two different aliquots (400µl/aliquot) from single blood samples were assayed separately from stimulation until the direct staining of the cells. Negative (no antigen, open circles) and positive (SEB, open triangles) controls were used in parallel to blood incubated during 24 hrs with 5 µg/ml PT, 5 µg/ml FHA, 10 µg/ml BPL, 10 µg/ml Bp sonicate or 10 µg/ml TT (black circles). Thirty-five comparisons were performed on five adults (n°1-5), two of which recently aP boosted, two aP boosted less than three years before, and one, six years before. For the two subjects recently boosted, blood was collected before and after aP booster vaccination. After 24 hrs incubation including a 5 hrs protein transport arrest, the cells were fixed, directly permeabilized and stained with antibody panel 1 (Table 1). The acquisition was performed on a Canto II flow cytometer, and FlowJo software (version 9.5.3) was used for the analysis. Correlations were evaluated by a non-parametric Spearman test with Graphpad Prism 7.03 software (Graphpad software, La Jolla, CA, USA).
To assess a potential operator effect on the flow cytometry analyses, we compared results of acquired data obtained for the same samples and analysed with the FlowJo software by two different operators, one experienced and one novice operator.
Blood samples from four adults (n°6 to n°9) and from two children (n°1, n°2), were included here and they were processed with an optimized antibody panel (panel 2, Table 1) to further expand the detection of Th2 and Th17-type CD4+ T cells (Figure 1). As a flow cytometer LSR Fortessa became available, datasets were analysed on this cytometer, and we extent the analysis to CD8+ T lymphocytes in addition to CD4+ T lymphocytes.
The percentages of total CD3+, CD4+ and CD8+ T lymphocytes obtained by the two operators were strongly correlated (Figure 3A, r=0.992, p<0.0001). Similarly, the percentages of IFN-γ-producing-CD4+ T cells, IL-4/IL-5/IL-13 and IL-17A/IL-17F were highly correlated (Figures 3B–D, p<0.0001). The percentages of IL-22-producing CD4+ T cells were low but also well correlated Supplementary Figure 2A). Similar to CD4+ T cells, the percentages of IFN-γ-producing CD8+ T cells were also well correlated Supplementary Figure 2B). The percentages of other cytokines produced by CD8+ T cells were insignificant.
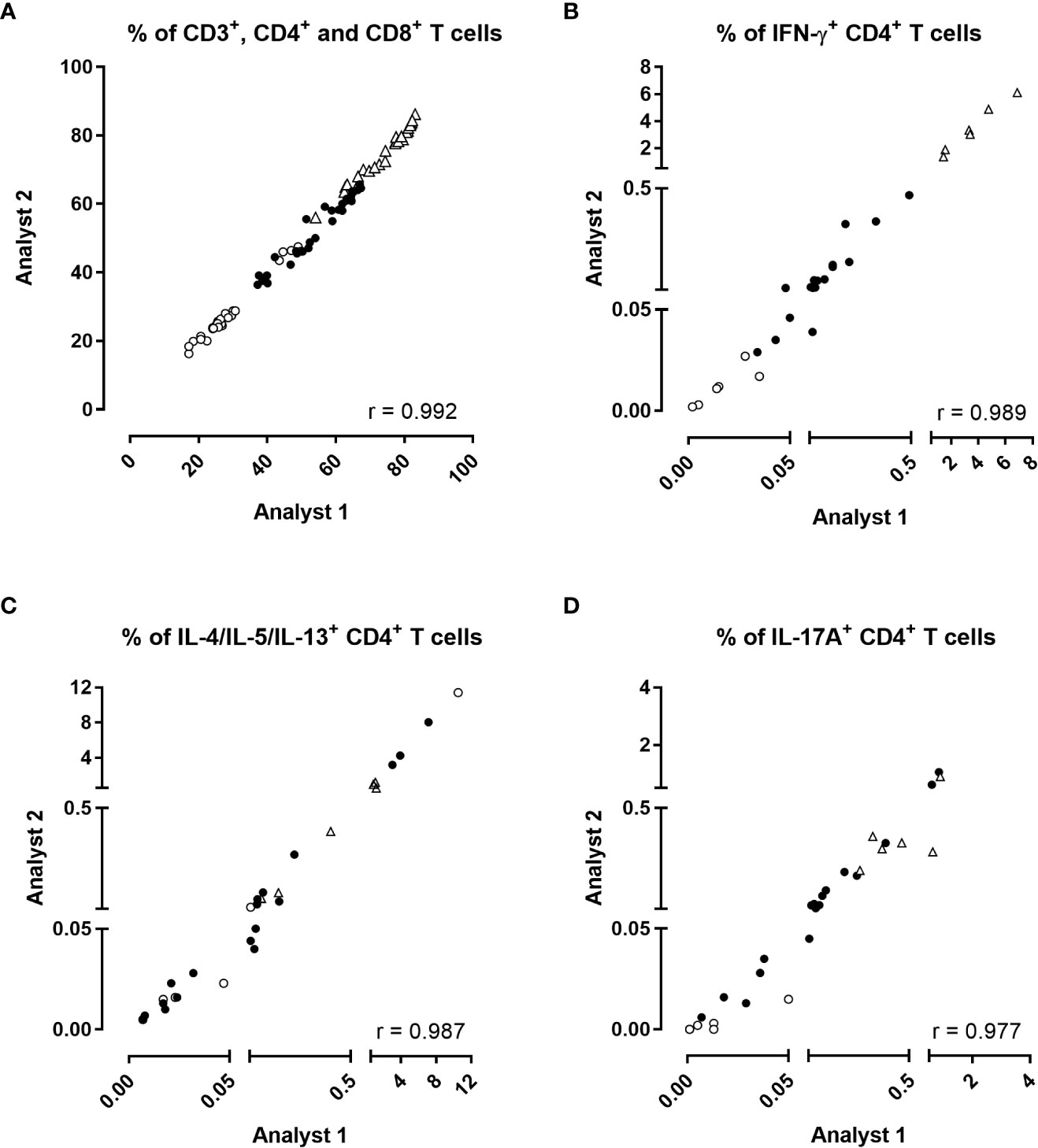
Figure 3 Inter-operator reproducibility of the analysis. The percentages of (A) CD3+ (open triangles), CD4+ (black circles), CD8+ (open circles) T cells, and the percentages of (B) IFN-γ, (C) IL-4/IL-5/IL-13 or (D) IL-17A/IL17-F-producing CD4+ T cells obtained by two different operators were compared two by two for each stimulation condition after direct staining of the cells. Diluted blood was incubated during 24 hrs with 5 µg/ml PT, 5 µg/ml FHA or 10 µg/ml BPL (black circles). Negative (no antigen, open circles) and positive (SEB, open triangles) controls were used in parallel. Thirty comparisons were performed on samples from four adults (n°6-9), and two 5-6 years-old children, all recently aP vaccine boosted. The cells were fixed, directly permeabilized, and stained with antibody panel 2, optimized for the detection of Th2 and Th17-type responses (Table 1). The acquisition was performed on a LSR Fortessa flow cytometer, and FlowJo software (version 9.5.3) was used for the analysis. Correlations were evaluated by a non-parametric Spearman test with Graphpad Prism 7.03 software (Graphpad software, La Jolla, CA, USA).
To evaluate the effect of cryopreservation of stimulated and fixed cells, blood samples from four adults (n°6 to n°9) and two 5-6 years old-children (n°1, n°2), were divided into two series of aliquots. The first series was stained directly, while the second one was stained after cryopreservation of the stimulated and fixed cells at -80°C during a median of 12 months (range: 1 month – 22 months), for a head to head comparison (Figure 1).
The percentages of total CD3+, CD4+ and CD8+ T lymphocytes obtained from fresh versus frozen samples were highly correlated (Figure 4A, r=0.956, p<0.0001), as were the percentages of IFN-γ-producing CD4+ T cells (Figure 4B, r=0.951 and p<0.0001). The correlation between the percentages of CD4+ T cells-producing IL-4/IL-5/IL-13 obtained by the two procedures was slightly lower albeit still satisfactory (Figure 4C, r=0.766, p<0.0001). Eight discrepancies out of 30 results were noticed between results of Th2-producing cells obtained by the two procedures, with slightly higher percentages noticed for the direct staining than for the procedure performed after cryopreservation (Figure 4C). These discrepancies originated from two adults for several stimulating conditions, and from one child for the unstimulated condition. The percentages of IL-17A/IL-17F-producing CD4+ T cells obtained by the two procedures were perfectly correlated (Figure 4D, r=0.974 and p<0.0001). Good correlations were also found for the very low percentages of IL-22-producing CD4+ T cells (r=0.860, p<0.0001, Supplementary Figure 3A), and for IFN-γ-producing CD8+ T cells (r=0.883, p<0.0001, Supplementary Figure 3B). The non-specific backgrounds were similar for direct staining compared to staining after cryopreservation (Supplementary Figure 4). Figure 5 summarized the final workflow of the BpWB-ICS assay.
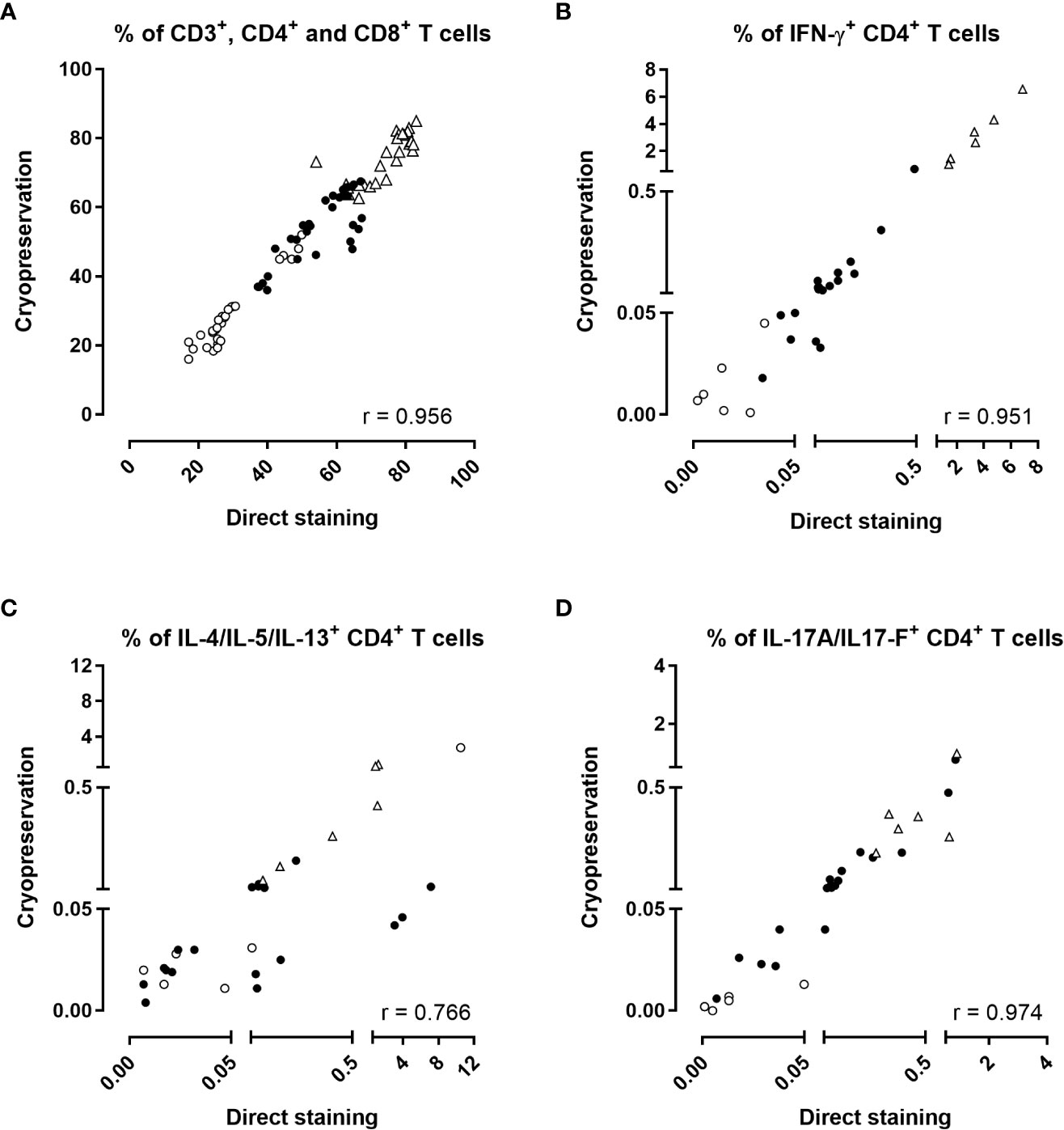
Figure 4 Effect of cryopreservation in the BpWB-ICS assay to detect Bp antigen-specific cytokine-producing CD4+ T cells. The percentages of (A) CD3+ (open triangles), CD4+ (black circles), CD8+ (open circles), and the percentages of (B) IFN-γ, (C) IL-4/IL-5/IL-13 or (D) IL-17A/IL-17F-producing CD4+ T cells obtained by a direct staining procedure were compared to those obtained after cryopreservation of the stimulated and fixed cells. Results were compared two by two for each stimulation condition. Diluted blood was incubated during 24 hrs with 5 µg/ml PT, 5 µg/ml FHA or 10 µg/ml BPL (black circles). Negative (no antigen, open circles) and positive (SEB, open triangles) controls were used in parallel. Thirty comparisons were performed on samples from four adults (n°6-9) and two 5-6 years-old children, all recently aP vaccine boosted. The cells were fixed and then, directly permeabilized for the direct staining, or cryopreserved in the Recovery Freezing Medium (ThermoFisher) for storage at -80°C. Frozen samples were thawed in PBS before permeabilization. The cells were stained with antibody panel 2 (Table 1). The acquisition was performed on a LSR Fortessa flow cytometer and FlowJo software (version 9.5.3) was used for the analysis. Correlations were evaluated by a non-parametric Spearman test with Graphpad Prism 7.03 software (Graphpad software, La Jolla, CA, USA).
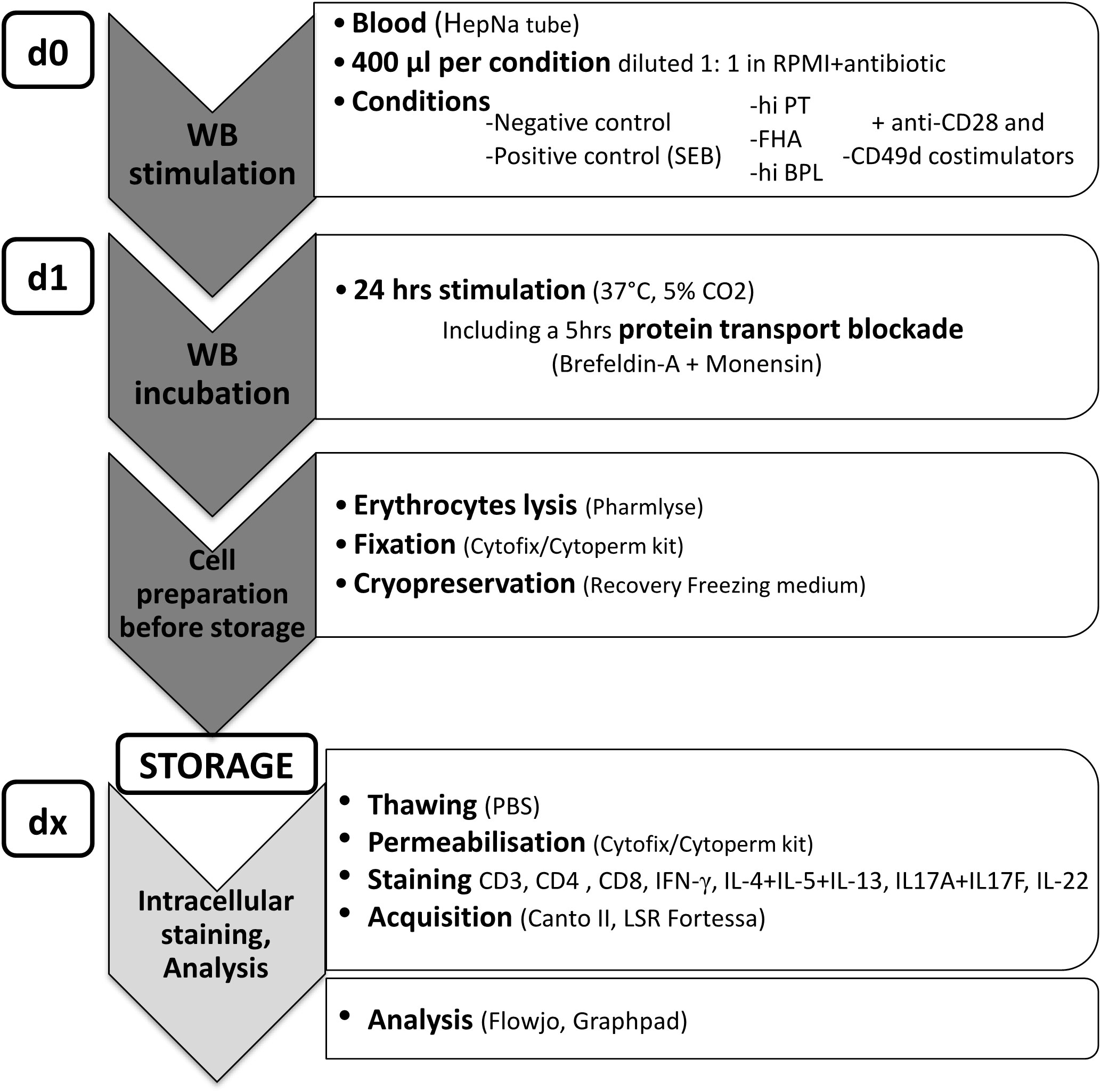
Figure 5 Workflow for the BpWB-ICS assay. Blood was collected in sodium heparin tubes and processed within three hours, maximum. Per stimulation, 400 µl of blood was diluted 1:1 with RPMI supplemented with gentamycin before incubation for 24 hrs in the presence of 1 µg/ml anti-CD28 and anti-CD49 co-stimulants in 15 ml round-bottom polypropylene tube (day 0, d0). Diluted blood was stimulated with 5 µg/ml heat-inactivated (hi) PT, 5 µg/ml FHA or 10 µg/ml hi Bp lysate. Negative (no antigen) and positive (SEB) controls were used in parallel. Protein transport arrest reagents (10 µg/ml Brefeldin-A and 1/1,000 Monensin) were added during the last 5 hrs of the incubation. After 24 hrs incubation (d1), red blood cells were lysed with the Pharmlyse buffer (BD Biosciences), and the white blood cells were fixed with the Fixation/Permeabilisation solution (BD biosciences), before freezing in the Recovery Freezing medium (ThermoFisher) and storage at -80°C for delayed analysis in batches (dx). After thawing in PBS, the cells were permeabilized with the BD Perm/Wash buffer (BD Biosciences), and stained with antibody panel 2 (Table 1) for analysis of IFN-γ-, IL-4/IL-5/IL-13-, IL-17A/IL-17F- or IL-22-producing CD4+ T cells to reveal Th1, Th2, and Th17-type responses, respectively.
To determine the assay performance and the capacity of the assay to detect Bp antigen-specific Th1, Th2, and Th17-type responses, we compared the percentages of cytokine-producing CD4+ T cells of cryopreserved samples stimulated with PT, FHA, BPL or SEB, to the non-specific background, after staining with antibody panel 2 (Figure 1). Samples from seven adults (n°6 to n° 12) and from two children (n°1, n°2), all recently aP vaccine boosted, were analysed (Figure 1).
Except for Th2-producing cells, background percentages were remarkably low, with median values of 0.012%, 0.007%, and 0.009% for IFN-γ, IL-17A/IL-17F, and IL-22-producing CD4+ T cells, respectively (Supplementary Figure 5). This background was higher for the percentage of IL-4/IL-5/IL-13-producing CD4+ T lymphocytes with a median of 0.031%, although they were still in an acceptable range (Supplementary Figure 5).
In contrast, the percentages of IFN-γ-producing CD4+ T cells obtained after stimulation with PT, FHA, BPL or SEB were higher and significantly different from the background percentages (Figure 6A). Overall, the percentages of antigen-induced-IL-4/IL-5/IL-13-producing CD4+ T cells were not significantly different from the background percentages, due to one subject with a very high IL-4/IL-5/IL-13 background (Figure 6B). Excluding this outlier subject resulted in significant differences between PT, FHA or SEB-stimulated compared to unstimulated cells (p=0.016 for PT and FHA and 0.008 for SEB). The percentages of IL-17A/IL-17F-producing CD4+ T cells obtained after stimulation were higher and significantly different than the background percentages (Figure 6C). Significant percentages of IL-22-producing CD4+ T cells, different from the background, were only noticed after stimulation with BPL or SEB (Figure 6D).
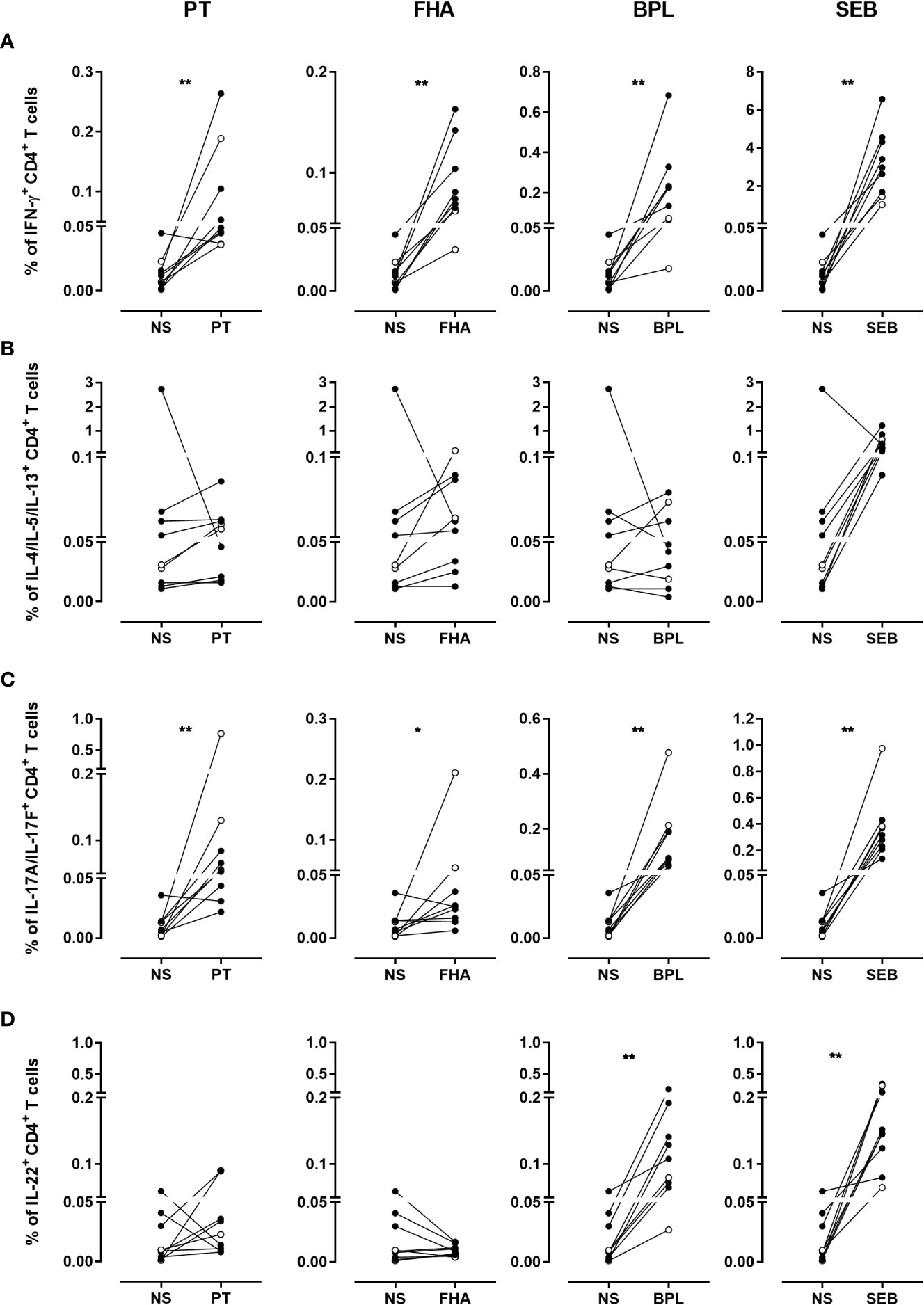
Figure 6 Frequencies of cytokine-producing CD4+ T cells in response to PT, FHA, BPL and SEB. The percentages of (A) IFN-γ-, (B) IL-4/IL-5/IL-13-, (C) IL-17A/IL-17F- and (D) IL-22- producing CD4+ T cells in cryopreserved samples after stimulation with PT, FHA, BPL or SEB are compared to those obtained in the absence of antigen (NS). Diluted blood was incubated during 24 hrs with 5 µg/ml PT, 5 µg/ml FHA or 10µg/ml BPL. Negative and SEB positive controls were used in parallel. The comparison is performed for seven adults (black circles) (n°6-12), and two 5-6 years-old children (open circles), all recently aP vaccine boosted. The cryopreserved cells were processed for intracellular staining as described in Figure 5, and stained with antibody panel 2 (Table 1). The acquisition was performed on a LSR Fortessa flow cytometer and FlowJo software (version 9.5.3) was used for the analysis. Comparisons were performed by using the Wilcoxon matched-pairs signed rank test with Graphpad Prism 7.03 software. *p<0.05; **p<0.01.
The sensitivity of the assay was assessed after staining cryopreserved samples previously stimulated with PT, FHA, BPL, with antibody panel 2. Samples were from seven adults (n°6 to n°12) and from two children (n°1, n°2), all recently aP vaccine boosted (Figure 1).
Based on defined criteria of positivity (Table 2), PT-specific IFN-γ-producing CD4+ T cells were detected for 6/7 adults and 1/2 children (Figure 7A), with 0.047% of positive cells (median) and a SI of 8 (median) (Supplementary Table 1). The second child had a doubtful response. All the adults (7/7) and one child had FHA- and BPL-specific IFN-γ-producing CD4+ T cells, with both high percentages of positive cells and SI (Supplementary Table 1). The second child had a doubtful percentage of positive cells (Figure 7A and Supplementary Table 2). Overall, IFN-γ-producing CD4+ T cells were detected to at least one Bp antigen in all subjects.
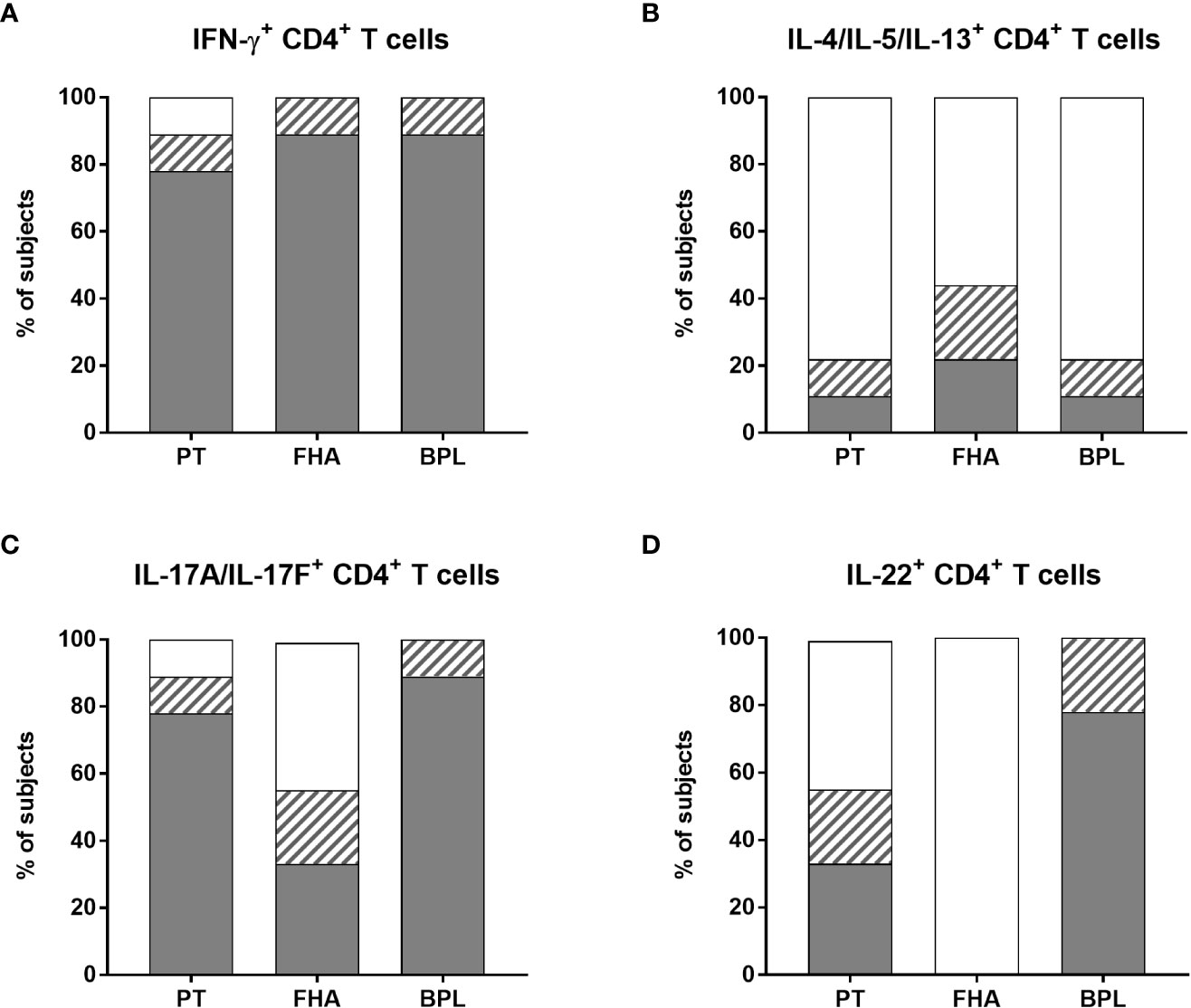
Figure 7 Distribution of non-responders and responders in the BpWB-ICS assay. The percentages of responders (grey areas), doubtful responders (hatched areas) and non-responders (white areas) to PT, FHA and BPL, based on their specific (A) IFN-γ-, (B) IL-4/IL-5/IL-13-, (C) IL-17A/IL-17F-, (D) and IL-22-positive CD4+ T cells were determined. The analysis was performed for seven adults (n°6-12), and two 5-6 years-old children, all recently aP vaccine boosted. The results obtained after staining of the Bp antigen-stimulated cryopreserved blood cells were considered after subtraction of the non-specific background. The criteria shown in Table 2 were applied for the classification in responders, doubtful responders and non-responders.
Bp antigen induced-Th2-type CD4+ T cells were clearly detected in the children only. Both children had FHA-specific Th2-type CD4+ T cells, one of them had also a PT-specific response, whereas the other one had also a BPL-specific response (Figure 7B and Supplementary Table 1). The apparent Th2-type CD4+ T cell response detected for one adult was non-specific, as it resulted from very high background (Figure 6). Some additional Th2-type responses were detected but considered doubtful, and induced by FHA (2/7 adults), PT (the second child), and BPL (1/7 adult) (Figure 7B and supplementary Table 2).
As for IFN-γ, most subjects had PT-specific IL-17A/IL-17F-producing CD4+ T cells (5/7 adults and 2/2 children) (Figure 7C), with 0.064% of positive cells (median) and a SI of 41 (median) (Supplementary Table 1). Almost all the subjects were responders to BPL (6/7 adults and 2/2 children) (Figure 7C and Supplementary Table 1). In contrast, only three subjects had detectable FHA-specific IL-17A/IL-17F-producing CD4+ T cells (1/7 adults and 2/2 children) (Figure 7C and supplementary Table 1). In addition, doubtful Th17-type responses were noticed for three adults (one induced by both PT and FHA, a second by FHA, and a third one by BPL) (Figure 7C and Supplementary Table 2). Of note, even if most subjects had both PT-specific IFN-γ- and IL-17A/IL-17F CD4+ T cells, these cytokines were rarely co-expressed by the same cells (Supplementary Figure 6), so that Boolean analysis was not further performed.
Finally, PT-specific IL-22-producing CD4+ T cells were detected only in three subjects (2/7 adults and 1/2 children), while no FHA-specific IL-22-producing CD4+ T cells were noticed (Figure 7D and supplementary Table 1). In contrast, seven subjects had BPL-induced IL-22-producing CD4+ T cells (6/7 adults and 1/2 children) (Figure 7D and supplementary Table 1). In addition, doubtful IL-22 responses were noticed for four subjects, and induced by PT (1/7 adults and 1/2 children) and BPL (1/7 adults and 1/2 children) (Figure 7D and supplementary Table 2). Subjects with specific IL-22-producing CD4+ T lymphocytes also had IL-17-producing cells, but the cells were different (Supplementary Figure 6).
The presence of IFN-γ-producing CD8+ T cells was investigated in adults n°6 to n°12 and in the two children, following the same procedure as described in Figure 5.
The median background for IFN-γ-producing CD8+ T cells was 0.014% (P25-P75:0.010%-0.049%, Figure 8A), which was significantly higher than the background of IFN-γ-producing CD4+ T lymphocytes (median: 0.012%, P25-P75:0.004%-0.020% (Supplementary Figure 5), and p=0.039, data not shown). Overall, the percentages of IFN-γ-producing CD8+ T cells after stimulation were significantly higher than the background for PT, BPL and SEB, but not for FHA (Figure 8B). The same criteria used to determine Bp antigen-specific CD4+ T cell responses were applied for IFN-γ-producing CD8+ T cells. Even if less frequent than for the CD4+ T cells, PT- and FHA-specific IFN-γ-producing CD8+ T lymphocytes were detected in 3 and 2 adults, respectively, while BPL induced IFN-γ-producing CD8+ T cells in all subjects. In addition, one adult and 2 children had a doubtful response to PT, and three adults to FHA (Figure 8C).
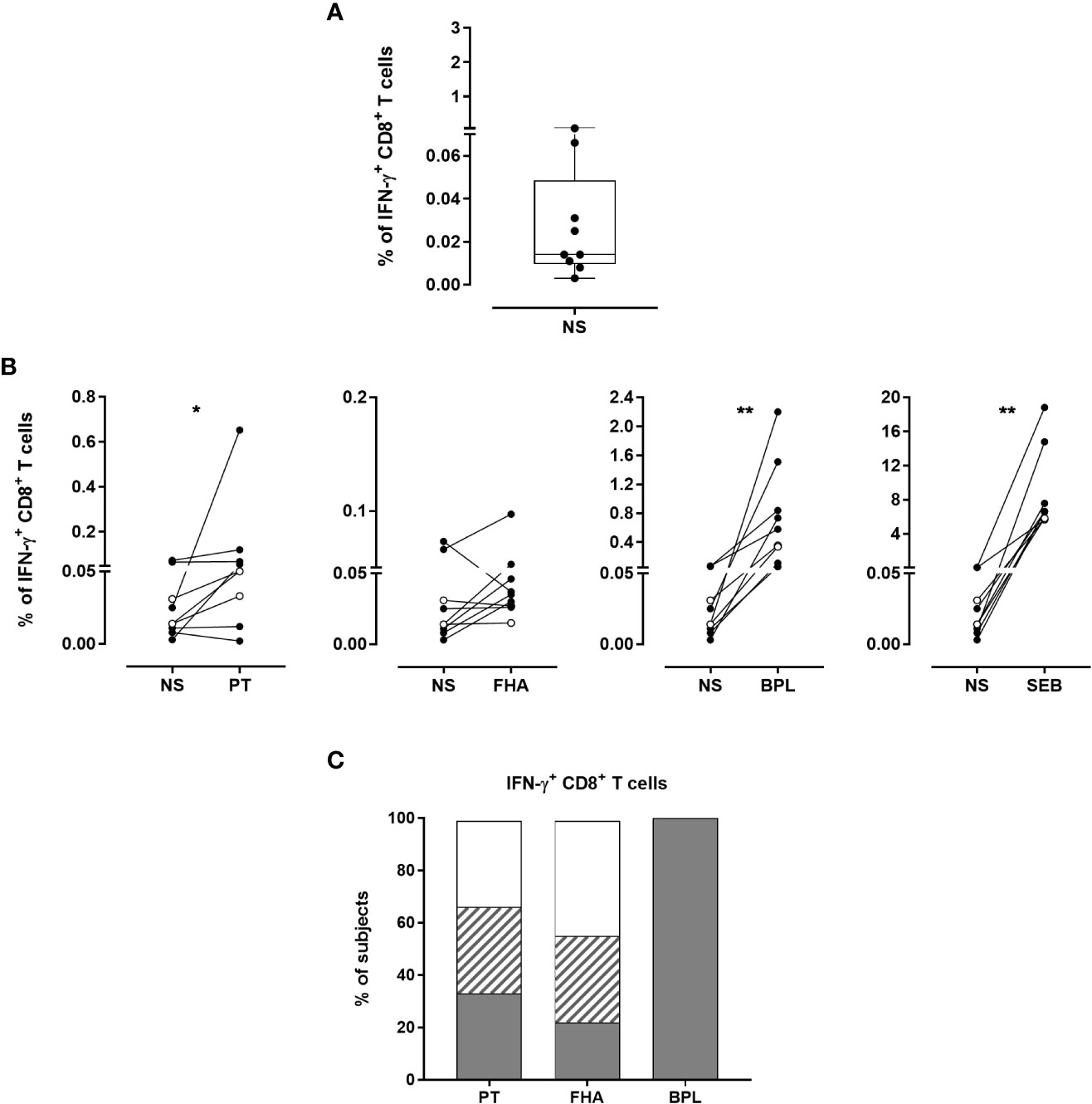
Figure 8 Detection of Bp antigen-specific IFN-γ-producing CD8+ T lymphocytes. The presence of IFN-γ-producing CD8+ T cells was investigated following the same procedure as described in Figure 5. The analysis was performed for seven adults (n°6-12) and two 5-6 years-old children, all recently aP vaccine boosted. (A) Non-specific background of IFN-γ-producing CD8+ T cells was determined after 24 hrs incubation in the absence of antigen (NS). The cryopreserved cells were processed for intracellular staining as described in Figure 5, and were stained with antibody panel 2 (Table 1). Horizontal line within the box, box and whisker represent the median, the P25-P75 and the range, respectively. (B) The percentages of IFN-γ-producing CD8+ T cells obtained after staining of PT-, FHA-, BPL- or SEB-stimulated cryopreserved blood cells were compared to those obtained in absence of antigen (NS). Black and open circles represent the results obtained from adult and children samples, respectively. The Wilcoxon matched-pairs signed rank test with Graphpad Prism 7.03 software was used for statistical analysis. *p<0.05; **p<0.01. (C) The percentages of responders (grey areas), doubtful responders (hatched areas) and non-responders (white areas) to PT, FHA and BPL are based on IFN-γ-producing CD8+ T cells after background subtraction. The criteria shown in Table 2 were applied for the classification in responders, doubtful responders and non-responders.
The availability of standardized robust assays for the quantification of cytokine-producing CD4+ T cells specific for Bp antigens, as described here, is essential for the evaluation of immunological responses to pertussis vaccines. Similar assays, performed on WB, have already been developed and validated in the field of tuberculosis (34, 35) and dengue vaccination study (42), but are, to our knowledge, not yet in use in the field of pertussis vaccines. Previously, we reported on the development of an initial version of a Bp antigen-specific WB FC assay (33). Here we further refined the assay and validated the method for reproducibility. We included a convenience step of cryopreservation of the stimulated cells to enable batch-wise analysis of longitudinal samples from the same individual. This BpWB-ICS assay allows for simultaneous detection of Bp antigen-specific Th1/Th2/Th17-type CD4+ and Bp antigen-specific IFN-γ CD8+ T cell responses on a total of 2 mL WB, and is thus useful for adult as well as pediatric clinical studies. It is easy to perform, includes negative and positive controls, and is feasible using an eight-color FC.
A main advantage of the BpWB-ICS assay over PBMC ICS assays is that, being performed on WB, it closely reflects the in vivo blood immune status, as all cells and components of the blood are present during the stimulation time with antigens (26). PBMC isolation and freezing/thawing procedures that are most often used, require larger blood volumes, are source of pre-analytic variabilities, and have proven difficult to standardize (26, 27). Additionally, such procedures introduce technical bias affecting cellular proportions, phenotypes and functions (26, 27), especially when recall responses are investigated (28, 29), which may lead to potentially biased conclusions (28, 43). In contrast to long in vitro stimulation time with antigens, classically used in the pertussis field for the evaluation of lymphocyte proliferation and for measurement of secreted cytokine concentrations (15, 16, 31), we used a short in vitro stimulation period, which is more suitable to detect effector memory T cells induced by recent vaccination.
Another major advantage of the BpWB-ICS assay refined here is that staining of cytokine-containing cells and their analysis by FC may be performed later on by batches of selected stored samples, thanks to the addition of a cryopreservation step after stimulation. Here we show that the cryopreservation of the stimulated and fixed cells had no impact on the results, as they were very well correlated to those obtained without cryopreservation. This procedure allows thus for the simultaneous analysis of samples collected at different time points for the same subject in clinical studies, reducing the variability inherent to the cell processing and analysis by FC. It also offers the possibility to select for analysis only samples of interest and for instance to focus only on samples from subjects with a complete longitudinal follow-up. Workflows may thus be easier to organize, and workload can be significantly reduced. In addition, this approach avoids a potential detrimental influence of cryopreservation on antigen presentation of Bp proteins by antigen-presenting cells (30).
We demonstrate here high reproducibility of the results, as well as high sensitivity for detection of rare specific events due to very low non-specific backgrounds. This was obtained thanks to technical improvements resulting in very high numbers of acquired events by FC, to careful optimization of the panel of antibodies used for cell staining, and to the FC analysis strategy as recommended (26, 44). To raise the sensitivity of detection of specific Th2 responses, that are often difficult to evaluate, combined labelling of three different Th2 cytokines (IL-4, IL-5, IL-13) within the same fluorescence channel was performed (15–17, 45). Similarly, labelling for IL-17A and IL-17F was combined to investigate Th17 responses (46). In addition to sensitivity improvement, this approach avoided the need for more than an eight-color FC. Finally, reproducibility of the data analysis via FlowJo software by operators with different levels of experience was high. The assay is thus well standardized from the step of blood collection until the FC data analysis provided that the standard operational procedure (Figure 5) is strictly applied.
ELISPOT is often reported to be more sensitive than FC to detect antigen-specific T cells among PBMC (47, 48), especially for the detection of low-frequent memory T cells before vaccine booster (49). However, to detect and visualize the cellular IFN-γ recall responses to a protein antigen such as tetanus toxoid after booster vaccine administration, higher sensitivity of FC compared to ELISPOT was demonstrated (49). The superiority of ELISPOT over FC to measure rare events before vaccine booster was suggested to be explained by low cytokine production per memory T cell per time unit, favoring measurement of accumulated cytokines during the in vitro culture period (47), as is the case for ELISPOT and not for FC. However, since the BpWB-ICS assay described here was adapted for the detection of very low percentages of positive cells, FC now become the method of choice to evaluate T cell responses induced by pertussis vaccines in clinical studies. Compared to ELISPOT, FC also presents the advantage to be feasible on WB on very small volumes, and to allow for the characterization of the phenotype of the cytokine-producing cells, thereby excluding potential contribution of other cell types, such as NK cells, to cytokine production (50).
This refined version of the BpWB-ICS assay comprises well-defined criteria for positivity of responses. Based on these criteria, we confirmed the presence in blood from recently boosted subjects of Bp-specific Th1-type CD4+ T cells, associated with Bp-specific Th2-type CD4+ T cells in children who, in contrast to adults, were aP-primed during infancy. In addition, we detected significant Bp antigen-specific Th17 lymphocytes, as most wP-primed adults and the two aP-primed children had PT- and BPL-specific IL-17A/IL-17F-producing CD4+ T cells. The induction of IL-17 by pertussis vaccines was only rarely reported in humans until now. They were detected either at very low levels in supernatants of Bp-stimulated PBMC from aP-primed children (51–53), or within CD4+ T cells from recently aP boosted individuals with a wP or aP-primed background (33). Considering the contribution of IL-17 in protection against Bp in non-human primates and other animal models (10), optimal detection of IL-17 production, as feasible with the BpWB-ICS assay, is thus a clear added value of this test to be used in human vaccine studies. Interestingly, we observed FHA-specific Th1/Th2/Th17 mixed responses in children, as described in mice immunized with aP vaccines (20). The protective role of Th17 responses against Bp in these cases remains questionable, as Th17 function may be inhibited in a Th2 environment (54, 55). Low percentages of PT-specific IL-22-producing CD4+ T cells were also detected, in agreement with previous results in aP boosted subjects (33). The potential role of IL-22 in the defence against B. pertussis has to our knowledge not yet been investigated, even if IL-22 has recently attracted great interest being considered as a regulator of host defence in the lung (56).
Finally, the BpWB-ICS assay also detected PT- and FHA-specific IFN-γ-producing CD8+ T lymphocytes in most subjects and in response to BPL in all of them. Such Bp antigen-induced cellular immune responses were previously reported during Bp infection and after vaccination in infants and young adults (15, 22–25). Their detection here sustains their involvement in immunity against Bp, but their potential role in protection still requires investigation.
A limitation of the study is that the validation of the refined version of the BpWB-ICS assay was only performed on blood samples from a small number of recently aP vaccine boosted subjects. However, the previous version of the assay was validated by testing blood samples from a large clinical booster vaccination study (33). The major difference of the refined version was the addition of a cryopreservation step and we demonstrated here high correlations of the results obtained head-to-head without and with a cryopreservation step. One might thus reasonably assume that the performance of the refined BpWB-ICS assay will be equally satisfactory in large clinical studies.
In summary, the BpWB-ICS assay described here is an optimal test to characterize both the quantity and the quality of Bp antigen-specific CD4+ and CD8+ T cells in a very small blood volume, during clinical studies with a longitudinal design. It appears therefore as a promising test to assess the immunogenicity of current and next generation pertussis vaccines.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by the Ethics Committee ULB-Hôpital Erasme (aggregation number OMO21, study protocol P2018/515). The patients/participants provided their written informed consent to participate in this study.
We acknowledge all the participants of WP5 Task 7 within the European PERISCOPE project/consortium:
Prof. Jacques van Dongen (Leiden University Medical Center, Netherlands); CACMvE, JAMvGvdB, EEL (National Institute for Public Health and the Environment RIVM, Netherlands); DAD, ES (Radboud University Medical Center RUMC, Netherlands); Prof. Kingston Mills, AM (Trinity College Dublin, Ireland); Prof. Beate Kampmann, Dr. Thomas Rice (Imperial College of London, United Kingdom); Dr Sophie Roetynck (Medical Research Council-Unit The Gambia at London School of hygiene and tropical medicine, The Gambia); Dr Vanessa Contreras (Commissariat à l'énergie atomique et aux énergies alternatives, INSERM U1184, France); FM, VC (Université Libre de Bruxelles U.L.B., Belgium).
FM, VC conceived the project and designed the study. VC, MR recruited patients, collected blood samples and clinical data. VC, EEL, JAMvGvdB, AM, ES and DAD participate to the development of the assay. VC optimized the experimental protocol, designed the database, analyzed the data, interpreted the data, and drafted the manuscript. VC, AP, AG performed the experiments for the optimization of the assay. AG analyzed the data. FM interpreted the data, and substantially revised the manuscript. CACMvE, DAD and EEL substantially revised the manuscript. All authors contributed to the article and approved the submitted version.
This research was conducted within the framework of the European PERISCOPE project/consortium. This PERISCOPE (Pertussis Correlates of Protection Europe) project has received funding from the Innovative Medicines Initiative 2 Joint Undertaking under grant agreement No 115910. This Joint Undertaking receives support from the European Union’s Horizon 2020 research and innovation programme and EFPIA and BMGF. http://www.imi.europa.eu
We are grateful to all the subjects who participated in this study. We kindly acknowledge Prof. Françoise Vermeulen and the nurses Sarah Van Hoey and Lieve Bachely for the recruitment of children. We thank Violette Dirix for the initiation of this project during its development in the TB field and Sarah Islane for her contribution in samples preparation. We are also grateful to Prof. Julia Almeida (University of Salamanca, Spain) for her support and advices during the development of the assay, and Prof Alberto Orfao (University of Salamanca, Spain) for his interest regarding this assay. We also thank Alison Hill (University of Southampton, UK) for helpful discussion concerning the implementation of the assay for human Challenge study. The authors thank Camille Locht for critically reading the manuscript, resulting in significant improvement.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1101366/full#supplementary-material
1. Mattoo S, Cherry JD. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to bordetella pertussis and other bordetella subspecies. Clin Microbiol Rev (2005) 18:57. doi: 10.1128/CMR.18.2.326-382.2005
2. Havers FP, Moro PL, Hariri S, Skoff T. Chapter 16: Pertussis. In: Centers for disease control and prevention. Epidemiology and prevention of vaccine-preventable diseases. Halls E, Wodi AP, Hamborsky J, Morelli V, Schillie S, eds. 14th ed. (Washington, D.C.: Public Health Foundation) (2021) 2021:239–54. Available at: https://www.cdc.gov/vaccines/pubs/pinkbook/index.html.
3. Gustafsson L, Reizenstein E, Storsaeter J. A controlled trial of a two-component acellular, a five-component acellular, and a whole-cell pertussis vaccine. N Engl J Med (1996) 334(6):7. doi: 10.1056/NEJM199602083340602
4. Misegades LK, Winter K, Harriman K, Talarico J, Messonnier NE, Clark TA, et al. Association of childhood pertussis with receipt of 5 doses of pertussis vaccine by time since last vaccine dose, California, 2010. JAMA (2012) 308(20):2126. doi: 10.1001/jama.2012.14939
5. Witt MA, Katz PH, Witt DJ. Unexpectedly limited durability of immunity following acellular pertussis vaccination in preadolescents in a north American outbreak. Clin Infect Dis (2012) 54(12):1730–5. doi: 10.1093/cid/cis287
6. Klein NP, Bartlett J, Fireman B, Rowhani-Rahbar A, Baxter R. Comparative effectiveness of acellular versus whole-cell pertussis vaccines in teenagers. Pediatrics (2013) 131(6):e1716–22. doi: 10.1542/peds.2012-3836
7. Clark TA. Changing pertussis epidemiology: Everything old is new again. J Infect Dis (2014) 209(7):978–81. doi: 10.1093/infdis/jiu001
8. Brummelman J, Wilk MM, Han WGH, van Els CACM, Mills KHG. Roads to the development of improved pertussis vaccines paved by immunology. Pathog Dis (2015) 73(8):ftv067. doi: 10.1093/femspd/ftv067
9. Warfel JM, Zimmerman LI, Merkel TJ. Acellular pertussis vaccines protect against disease but fail to prevent infection and transmission in a nonhuman primate model. Proc Natl Acad Sci (2014) 111(2):787–92. doi: 10.1073/pnas.1314688110
10. Fedele G, Cassone A, Ausiello CM. T-Cell immune responses to bordetella pertussis infection and vaccination. Pathog Dis (2015) 73(7):ftv051. doi: 10.1093/femspd/ftv051
11. Mills KHG. Cell-mediated immunity to bordetella pertussis: Role of Thl cells in bacterial clearance in a murine respiratory infection model. Infect Immun (1993) 61:12. doi: 10.1128/iai.61.2.399-410.1993
12. Warfel JM, Beren J, Kelly VK, Lee G, Merkel TJ. Nonhuman primate model of pertussis. Infect Immun (2012) 80(4):1530–6. doi: 10.1128/IAI.06310-11
13. Ausiello CM, Urbani F, la Sala A, Lande R, Cassone A. Vaccine- and antigen-dependent type 1 and type 2 cytokine induction after primary vaccination of infants with whole-cell or acellular pertussis vaccines. Infect Immun (1997) 65(6):2168–74. doi: 10.1128/iai.65.6.2168-2174.1997
14. Ausiello CM, Lande R, Urbani F, Di Carlo B, Stefanelli P, Salmaso S, et al. Cell-mediated immunity and antibody responses to bordetella pertussis antigens in children with a history of pertussis infection and in recipients of an acellular pertussis vaccine. J Infect Dis (2000) 181(6):1989–95. doi: 10.1086/315509
15. Mascart F, Verscheure V, Malfroot A, Hainaut M, Piérard D, Temerman S, et al. Bordetella pertussis infection in 2-Month-Old infants promotes type 1 T cell responses. J Immunol (2003) 170(3):1504–9. doi: 10.4049/jimmunol.170.3.1504
16. Mascart F, Hainaut M, Peltier A, Verscheure V, Levy J, Locht C. Modulation of the infant immune responses by the first pertussis vaccine administrations. Vaccine (2007) 25(2):391–8. doi: 10.1016/j.vaccine.2006.06.046
17. Dirix V, Verscheure V, Goetghebuer T, Hainaut M, Debrie AS, Locht C, et al. Cytokine and antibody profiles in 1-year-old children vaccinated with either acellular or whole-cell pertussis vaccine during infancy. Vaccine (2009) 27(43):6042–7. doi: 10.1016/j.vaccine.2009.07.075
18. Higgins SC, Jarnicki AG, Lavelle EC, Mills KHG. TLR4 mediates vaccine-induced protective cellular immunity to bordetella pertussis: Role of IL-17-producing T cells. J Immunol Baltim Md (1950) 177(11):7980–9. doi: 10.4049/jimmunol.177.11.7980
19. Feunou PF, Bertout J, Locht C. T- and B-cell-Mediated protection induced by novel, live attenuated pertussis vaccine in mice. cross protection against parapertussis. PloS One (2010) 5(4):10. doi: 10.1371/journal.pone.0010178
20. Ross PJ, Sutton CE, Higgins S, Allen AC, Walsh K, Misiak A, et al. Relative contribution of Th1 and Th17 cells in adaptive immunity to bordetella pertussis: Towards the rational design of an improved acellular pertussis vaccine. PloS Pathog (2013) 9(4):e1003264. doi: 10.1371/journal.ppat.1003264
21. Warfel JM, Merkel TJ. Bordetella pertussis infection induces a mucosal IL-17 response and long-lived Th17 and Th1 immune memory cells in nonhuman primates. Mucosal Immunol (2013) 6(4):787–96. doi: 10.1038/mi.2012.117
22. Ryan M, Murphy G, Ryan E, Nilsson L, Shackley F, Gothefors L, et al. Distinct T-cell subtypes induced with whole cell and acellular pertussis vaccines in children. Immunology (1998) 93(1):1–10. doi: 10.1046/j.1365-2567.1998.00401.x
23. Rieber N, Graf A, Hartl D, Urschel S, Belohradsky BH, Liese J. Acellular pertussis booster in adolescents induces Th1 and memory CD8+ T cell immune response. PloS One (2011) 6(3):e17271. doi: 10.1371/journal.pone.0017271
24. Dirix V, Verscheure V, Vermeulen F, De Schutter I, Goetghebuer T, Locht C, et al. Both CD4 + and CD8 + lymphocytes participate in the IFN-γ response to filamentous hemagglutinin from Bordetella pertussis in infants, children, and adults. Clin Dev Immunol (2012) 2012:1–9. doi: 10.1155/2012/795958
25. de Rond L, Schure RM, Öztürk K, Berbers G, Sanders E, van Twillert I, et al. Identification of pertussis-specific effector memory T cells in preschool children. Clin Vaccine Immunol (2015) 22(5):561–9. doi: 10.1128/CVI.00695-14
26. Smith SG, Smits K, Joosten SA, van Meijgaarden KE, Satti I, Fletcher HA, et al. Intracellular cytokine staining and flow cytometry: Considerations for application in clinical trials of novel tuberculosis vaccines. PloS One (2015) 10(9):e0138042. doi: 10.1371/journal.pone.0138042
27. Duffy D, Rouilly V, Braudeau C, Corbière V, Djebali R, Ungeheuer MN, et al. Standardized whole blood stimulation improves immunomonitoring of induced immune responses in multi-center study. Clin Immunol (2017) 183:325–35. doi: 10.1016/j.clim.2017.09.019
28. Ford T, Wenden C, Mbekeani A, Dally L, Cox JH, Morin M, et al. Cryopreservation-related loss of antigen-specific IFNc producing CD4+ T-cells can skew immunogenicity data in vaccine trials: Lessons from a malaria vaccine trial substudy. Vaccine (2017) 35(15):1898–906. doi: 10.1016/j.vaccine.2017.02.038
29. Li Y, Mateu E, Díaz I. Impact of cryopreservation on viability, phenotype, and functionality of porcine PBMC. Front Immunol (2021) 12:765667. doi: 10.3389/fimmu.2021.765667
30. De Rosa SC, Lu FX, Yu J, Perfetto SP, Falloon J, Moser S, et al. Vaccination in humans generates broad T cell cytokine responses. J Immunol Baltim Md (2004) 173(9):5372–80. doi: 10.4049/jimmunol.173.9.5372
31. Carollo M, Palazzo R, Bianco M, Smits K, Mascart F, Ausiello CM. Antigen-specific responses assessment for the evaluation of bordetella pertussis T cell immunity in humans. Vaccine (2012) 30(9):1667–74. doi: 10.1016/j.vaccine.2011.12.104
32. Diavatopoulos DA, Mills KHG, Kester KE, Kampmann B, Silerova M, Heininger U, et al. PERISCOPE: road towards effective control of pertussis. Lancet Infect Dis (2019) 19(5):e179–86. doi: 10.1016/S1473-3099(18)30646-7
33. Lambert EE, Corbière V, van Gaans-van den Brink JAM, Duijst M, Venkatasubramanian PB, Simonetti E, et al. Uncovering distinct primary vaccination-dependent profiles in human bordetella pertussis specific CD4+ T-cell responses using a novel whole blood assay. Vaccines (2020) 8(2):225. doi: 10.3390/vaccines8020225
34. Hanekom WA, Hughes J, Mavinkurve M, Mendillo M, Watkins M, Gamieldien H, et al. Novel application of a whole blood intracellular cytokine detection assay to quantitate specific T-cell frequency in field studies. J Immunol Methods (2004) 291(1–2):185–95. doi: 10.1016/j.jim.2004.06.010
35. Kagina BM, Mansoor N, Kpamegan EP, Penn-Nicholson A, Nemes E, Smit E, et al. Qualification of a whole blood intracellular cytokine staining assay to measure mycobacteria-specific CD4 and CD8 T cell immunity by flow cytometry. J Immunol Methods (2015) 417:22–33. doi: 10.1016/j.jim.2014.12.003
36. Mascart F, Dirix V, Locht C. The human immune responses to pertussis and pertussis vaccines. In: Rohani P, Scarpino S, editors. Pertussis: Epidemiology, immunology, and evolution. Oxford University Press (2018) 112–132. doi: 10.1093/oso/9780198811879.003.0007
37. Locht C, Coutte L, Mielcarek N. The ins and outs of pertussis toxin. FEBS J (2011) 278(23):4668–82. doi: 10.1111/j.1742-4658.2011.08237.x
38. Bart MJ, Zeddeman A, van der Heide HGJ, Heuvelman K, van Gent M, Mooi FR. Complete genome sequences of bordetella pertussis isolates B1917 and B1920, representing two predominant global lineages. Genome Announc (2014) 2(6):e01301–14. doi: 10.1128/genomeA.01301-14
39. Smith SG, Joosten SA, Verscheure V, Pathan AA, McShane H, Ottenhoff THM, et al. Identification of major factors influencing ELISpot-based monitoring of cellular responses to antigens from mycobacterium tuberculosis. PloS One (2009) 4(11):e7972. doi: 10.1371/journal.pone.0007972
40. Weinberg A, Zhang L, Brown D, Erice A, Polsky B, Hirsch MS, et al. Viability and functional activity of cryopreserved mononuclear cells. Clin Diagn Lab Immunol (2000) 7(4):714–6. doi: 10.1128/CDLI.7.4.714-716.2000
41. Nomura LE, Walker JM, Maecker HT. Optimization of whole blood antigen-specific cytokine assays for CD4(+) T cells. Cytometry (2000) 40(1):60–8. doi: 10.1002/(SICI)1097-0320(20000501)40:1<60::AID-CYTO8>3.0.CO;2-J
42. Harenberg A, Begue S, Mamessier A, Gimenez-Fourage S, Ching Seah C, Wei Liang A, et al. Persistence of Th1/Tc1 responses one year after tetravalent dengue vaccination in adults and adolescents in Singapore. Hum Vaccines Immunother (2013) 9(11):2317–25. doi: 10.4161/hv.25562
43. Kvarnström M, Jenmalm MC, Ekerfelt C. Effect of cryopreservation on expression of Th1 and Th2 cytokines in blood mononuclear cells from patients with different cytokine profiles, analysed with three common assays: An overall decrease of interleukin-4. Cryobiology (2004) 49(2):157–68. doi: 10.1016/j.cryobiol.2004.06.003
44. Bacher P, Scheffold A. Flow-cytometric analysis of rare antigen-specific T cells. Cytom Part J Int Soc Anal Cytol (2013) 83(8):692–701. doi: 10.1002/cyto.a.22317
45. Rowe J, Macaubas C, Monger TM, Holt BJ, Harvey J, Poolman JT, et al. Antigen-specific responses to diphtheria-tetanus-acellular pertussis vaccine in human infants are initially Th2 polarized. Infect Immun (2000) 68(7):3873–7. doi: 10.1128/IAI.68.7.3873-3877.2000
46. Wright JF, Guo Y, Quazi A, Luxenberg DP, Bennett F, Ross JF, et al. Identification of an interleukin 17F/17A heterodimer in activated human CD4+ T cells. J Biol Chem (2007) 282(18):13447–55. doi: 10.1074/jbc.M700499200
47. Helms T, Boehm BO, Asaad RJ, Trezza RP, Lehmann PV, Tary-Lehmann M. Direct visualization of cytokine-producing recall antigen-specific CD4 memory T cells in healthy individuals and HIV patients. J Immunol (2000) 164(7):3723–32. doi: 10.4049/jimmunol.164.7.3723
48. Asemissen AM, Nagorsen D, Keilholz U, Letsch A, Schmittel A, Thiel E, et al. Flow cytometric determination of intracellular or secreted IFNg for the quantification of antigen reactive T cells. J Immunol Methods (2001) 251(1‐2):101–8. doi: 10.1016/S0022-1759(01)00312-X
49. Tassignon J, Burny W, Dahmani S, Zhou L, Stordeur P, Byl B, et al. Monitoring of cellular responses after vaccination against tetanus toxoid: Comparison of the measurement of IFN-γ production by ELISA, ELISPOT, flow cytometry and real-time PCR. J Immunol Methods (2005) 305(2):188–98. doi: 10.1016/j.jim.2005.07.014
50. Desombere I, Meuleman P, Rigole H, Willems A, Irsch J, Leroux-Roels G. The interferon gamma secretion assay: A reliable tool to study interferon gamma production at the single cell level. J Immunol Methods (2004) 286(1–2):167–85. doi: 10.1016/j.jim.2004.01.001
51. Schure RM, de Rond L, Öztürk K, Hendrikx L, Sanders E, Berbers G, et al. Pertussis circulation has increased T-cell immunity during childhood more than a second acellular booster vaccination in Dutch children 9 years of age. PloS One (2012) 7(7):e41928. doi: 10.1371/journal.pone.0041928
52. Schure RM, Hendrikx LH, de Rond LGH, Öztürk K, Sanders EAM, Berbers GAM, et al. T-Cell responses before and after the fifth consecutive acellular pertussis vaccination in 4-Year-Old Dutch children. Clin Vaccine Immunol (2012) 19(11):1879–86. doi: 10.1128/CVI.00277-12
53. Schure RM, Hendrikx LH, de Rond LGH, Öztürk K, Sanders EAM, Berbers GAM, et al. Differential T- and b-cell responses to pertussis in acellular vaccine-primed versus whole-cell vaccine-primed children 2 years after preschool acellular booster vaccination. Clin Vaccine Immunol (2013) 20(9):1388–95. doi: 10.1128/CVI.00270-13
54. Eyerich K, Pennino D, Scarponi C, Foerster S, Nasorri F, Behrendt H, et al. IL-17 in atopic eczema: Linking allergen-specific adaptive and microbial-triggered innate immune response. J Allergy Clin Immunol (2009) 123(1):59–66.e4. doi: 10.1016/j.jaci.2008.10.031
55. Dubois V, Chatagnon J, Thiriard A, Bauderlique-Le Roy H, Debrie AS, Coutte L, et al. Suppression of mucosal Th17 memory responses by acellular pertussis vaccines enhances nasal bordetella pertussis carriage. NPJ Vaccines (2021) 6(1):6. doi: 10.1038/s41541-020-00270-8
Keywords: Bordetella pertussis, vaccine, whole blood assay, Th1/Th2/Th17, intracellular cytokine staining, flow cytometry
Citation: Corbière V, Lambert EE, Rodesch M, van Gaans-van den Brink JAM, Misiak A, Simonetti E, Van Praet A, Godefroid A, Diavatopoulos DA, van Els CACM, Mascart F and PERISCOPE WP5 Task 7 working group (2023) A semi high-throughput whole blood-based flow cytometry assay to detect and monitor Bordetella pertussis-specific Th1, Th2 and Th17 responses. Front. Immunol. 14:1101366. doi: 10.3389/fimmu.2023.1101366
Received: 17 November 2022; Accepted: 24 January 2023;
Published: 06 February 2023.
Edited by:
Simone A. Joosten, Leiden University Medical Center (LUMC), NetherlandsReviewed by:
Julie G. Burel, La Jolla Institute for Immunology (LJI), United StatesCopyright © 2023 Corbière, Lambert, Rodesch, van Gaans-van den Brink, Misiak, Simonetti, Van Praet, Godefroid, Diavatopoulos, van Els, Mascart and PERISCOPE WP5 Task 7 working group. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Véronique Corbière, dmVyb25pcXVlLmNvcmJpZXJlQHVsYi5iZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.