
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 27 February 2023
Sec. Mucosal Immunity
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1093340
 Alexander Humberg1,2*
Alexander Humberg1,2* Lisa Neuenburg3
Lisa Neuenburg3 Hannah Boeckel3
Hannah Boeckel3 Mats Ingmar Fortmann3
Mats Ingmar Fortmann3 Christoph Härtel4Egbert Herting3Heilwig Hinrichs5Franziska Rademacher5
Christoph Härtel4Egbert Herting3Heilwig Hinrichs5Franziska Rademacher5 Jürgen Harder5
Jürgen Harder5Introduction: Preterm infants have an immature epidermis barrier function that may lead to an increased permeability to pathogens. On the surface of the human skin, antimicrobial peptides (AMPs) are important molecules of the innate immune system, have broad antimicrobial properties, and provide an essential role in integrity of the microbiome. Given the marked susceptibility of preterm infants to infection, we hypothesize a decreased expression of AMPs on the skin of preterm infants.
Materials and methods: In a prospective single-center study with 35 preterm and 20 term infants, we analyzed skin rinsing probes for the presence of the AMPs psoriasin (S100A7) and ribonuclease 7 (RNase 7) via enzyme-linked immunosorbent assay. Samples were taken from preterm infants < 34 0/7 weeks gestational age (mean ± SD gestational age, 28.8 ± 2.4 weeks) on days 0, 7, 14, and 28 after birth. Term infants (> 36 6/7 weeks) (controls) were washed on days 0 and 28.
Results: Psoriasin and RNase 7 were both expressed on skin of preterm and term infants and increased in concentration significantly over time. RNase 7 was more expressed in term infants on day 0 [preterm = 1.1 (0.7–2.9) vs. term = 2.0 (1.1–3.4) ng/ml, p = 0.017]. On day 28, premature infants showed higher values of psoriasin [preterm = 10.9 (5.6–14.2) vs. term = 6.3 (3.4–9.0) ng/ml, p < 0.001]. Notably, preterm infants with infectious or inflammatory context driven by histological proof of chorioamnionitis and early-onset or late-onset sepsis had higher concentrations of psoriasin as compared with non-affected preterm infants. After exclusion of infants with inflammatory hit, median concentrations of RNase 7 and psoriasin did not differ between preterm and full-term infants on days 0 and 28.
Discussion: Psoriasin and RNase 7 concentrations increase over time on the skin of newborn infants and seem to play a role in the first defense against infection. This is of particularly interest as the role of AMPs on a maturing skin microbiome and its possible new prevention strategies is unclear and needs to be determined.
Antimicrobial peptides (AMPs) on the surface of the skin have broad antimicrobial properties and contribute to immune protection as first line innate immune barrier (1, 2). In addition, AMPs play an essential role in establishing and maintaining a homogeneous colonization of the skin with microorganisms (3). AMPs are primarily released by cells of the immune system but are also produced by epithelial cells within the skin and at mucosal surfaces (4–7). Defensins [human beta-defensin (hBD)], cathelicidins (LL-37), psoriasin, ribonuclease 7 (RNAse 7), dermcidin, and adrenomedullin have been described on human skin. Because of their broad antimicrobial properties against bacteria, viruses, and fungi, these peptides enable effective immune protection. RNase 7 is able to prevent colonization by S. aureus and exhibit high antimicrobial activity in vitro against a variety of gram-positive and gram-negative bacteria and yeasts (8). The antimicrobial activity of psoriasin against pathogens is mediated by zinc deprivation. In healthy skin, psoriasin is continuously expressed and released in high concentrations and, therefore, effectively protects the skin from colonization and infection by E. coli. It shows increased expression after stimulation with bacteria or proinflammatory cytokines such as interleukin-1 (IL-1) or IL-17 (9).
Preterm infants born before completion of 36 weeks’ gestation are at increased risk for neonatal infections with significant effects on mortality and long-term morbidity (10, 11). Multiple factors and, mainly, a dysregulation of the immune system significantly contribute to an increased infection risk [for review, see (11)]. The skin of preterm infants has a reduced barrier function as the stratum corneum (SC) of the epidermis has fewer layers, vernix caseosa is not present, and reduced integrity function in the first weeks of life renders preterm skin permeable to pathogens. Furthermore, injuries of the premature skin through vascular access devices or adhesives play a key role for nosocomial infections (11).
It was found that, in relation to term infants, preterm infants demonstrate lower levels of AMPs in plasma, lung lavage, and feces (12–15). Furthermore, the expression of AMPs in the fetal skin depends on the maturity of the skin and the innate immune system (16). The deficiency of AMPs in early life may contribute to an increased risk for invasive infections. However, no data exist about AMP concentrations of the skin of premature infants and their role in immune protection of these vulnerable infants.
We here aim to characterize the expression and composition of skin AMPs in preterm and term infants in a longitudinal fashion within the first 28 days of life. We documented clinical factors that might influence the expression of AMPs, i.e., gestational age, histological chorioamnionitis, neonatal sepsis, and preterm skin care with sunflower oil. We addressed the expression of psoriasin (S100A7) and RNase 7 as most abundant AMPs on human skin surface (7).
All infants were born and recruited at the Department of Pediatrics, University Hospital Schleswig-Holstein, Campus Lübeck, between November 2019 and March 2021. Immediately after birth, infants were evaluated for inclusion criteria. To control for premature effects on AMP concentrations, we recruited infants with gestational age of < 34 0/7 weeks and as control infants > 36 6/7 weeks gestational age. Infants with diseases or injuries of the skin were not included. In term infants, any externals or cosmetics had to be left out. As most preterm infants received sunflower oil treatment of the skin, the left upper thigh was left out to control for effects of sunflower oil treatment. To reduce inter-observer variability, all patients are assessed only by two different researchers (LN and HB).
From all participants, non-invasive skin wash probes were taken from different sites (abdomen, left and right thigh, and upper arm). Skin rinsing procedure was performed in preterm infants immediately after birth on days 0, 7, 14, and 28 and in term infants on days 0 and 28. The skin was flushed with 1 ml of 37°C warmed buffer solution [10 mM sodium phosphate buffer (pH 7.2) with 0.1% Triton X-100] through sterile plastic tubes by pipetting 10 times up and down. The liquid was subsequently stored in tubes and immediately centrifuged (10 min, 10,000×g) and diluted 1:10 with buffer solution containing 10% (w/v) bovine serum albumin, aliquoted, and stored at −80°C. All materials are sent for laboratory analysis to the Department of Dermatology, University Hospital Schleswig-Holstein, Campus Kiel. After centrifugation, supernatants were analyzed for AMPs via “enzyme linked immunosorbent assay” (ELISA) as described elsewhere (17, 18). To analyze psoriasin expression, a monoclonal antibody from hybridoma mouse cells was used. For RNase 7 detection, the polyclonal antibody was derived from goat [23].
Placenta specimen from infants born < 34 0/7 weeks gestational age were analyzed for presence of histological chorioamnionitis at the Institute of Pathology, University Hospital Schleswig-Holstein, Campus Lübeck.
Gestational age was calculated from the best obstetric estimate based on early prenatal ultrasound and obstetric examination. Sepsis was defined as condition when neonatologists decided to treat the infant with antibiotics and to continue for at least 5 days due to the following reasons: ≥ 2 clinical signs of systemic inflammatory response: temperature > 38°C or < 36.5°C, tachycardia > 200/min, new onset or increased frequency of bradycardias or apneas, hyperglycemia > 140 mg/dl, base excess < −10 mval/l, changed skin color, and increased oxygen need; and 1 laboratory sign: C-reactive protein > 10mg/L, platelet count < 100/nl, immature/total neutrophil ratio > 0.2, and white blood cell count < 5/nl (NeoKISS) (19). Early-onset sepsis (EOS) was defined as signs of sepsis within the first 72 h after birth with or without proof of a causative agent in blood culture. Late-onset sepsis (LOS) was defined as signs of sepsis after the first 72 h after birth with or without proof of a causative agent in blood culture. Small for gestational age is defined as birth percentile < 10th according to gestational age (20).
Approval by the local ethics committee for research in human subjects of the University of Lübeck (file number 19-028) has been granted. The study protocol was registered on the German Clinical Trials Register portal (DRKS00021635) on 12 May 2020. The study is compliant with the Health Insurance Portability and Accountability Act of 1996 (HIPAA). The work has been carried out in accordance with The Code of Ethics of the World Medical Association (Declaration of Helsinki) for experiments involving humans. Immediately after birth and stabilization of the infants, oral consent was taken from parents or legal guardians prior the first skin washing. Within 48 h after birth, all parents or legal guardians were contacted for written informed consent. If then parents refused to participate, all materials and data were disposed.
Medical data were collected, pseudonymized, and entered into a data base. Clinical data concerning perinatal information, infections, antibiotic therapy duration, and duration of phototherapy were recorded.
Values of different probe sites (abdomen, thigh, and arm) were pooled and analyzed. Data were presented as means with standard deviation (SD), medians with interquartile ranges (IQRs), as well as numbers with frequency (n%) and their corresponding 95% confidence intervals (CIs). Differences of independent groups are tested via Mann–Whitney U-test and Kruskal–Wallis test. Wilcoxon signed-rank test is used to compare two related samples. Ordinary one-way ANOVA with Holm–Sidak’s multiple comparisons test was used for comparison of more than three related samples. The level of significance is set at p < 0.05.
Intra-assay coefficient of variation (CV) and inter-assay CV was calculated within each plate and the average CV across all plates. CV% was calculated by dividing the SDs by the means and converting to a percentage (×100).
All statistical analyses were performed with SPSS 26.0 software (IBM SPSS Statistics for Windows, Version 27.0. Munich, Germany) and with GraphPad Prism (Version 7.0, San Diego, CA).
This project was financed with funds from the Department of Pediatrics, University Hospital of Schleswig-Holstein, Campus Lübeck and the Department of Dermatology, Venerology and Allergology, Quincke Research Center, University-Hospital Schleswig-Holstein, Campus Kiel.
In this convenience sample study, a total of n = 62 infants had a first skin rinsing probe taken on day 0 of life (see Figure 1).
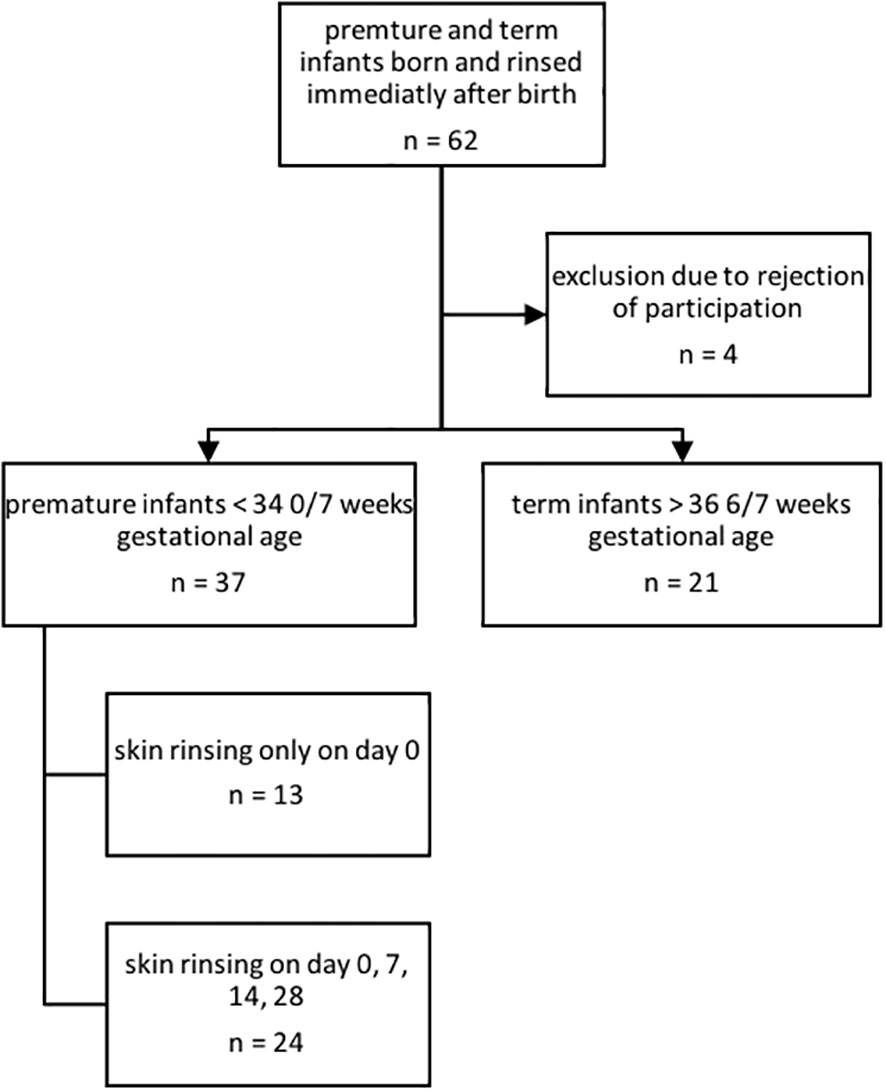
Figure 1 Inclusion and exclusion of premature and term infants for AMP characterization.
After refusal of participation by parents of four infants and exclusion due to unmet inclusion criteria (gestational age out of scope), n = 55 infants consisting of n = 35 premature infants and n = 20 term infants were included and followed up for 28 days. For further baseline cohort characteristics, please see Table 1. Until corrections of the study schedule, in n = 13 premature infants, rinsing solution was only collected on the first day of life.
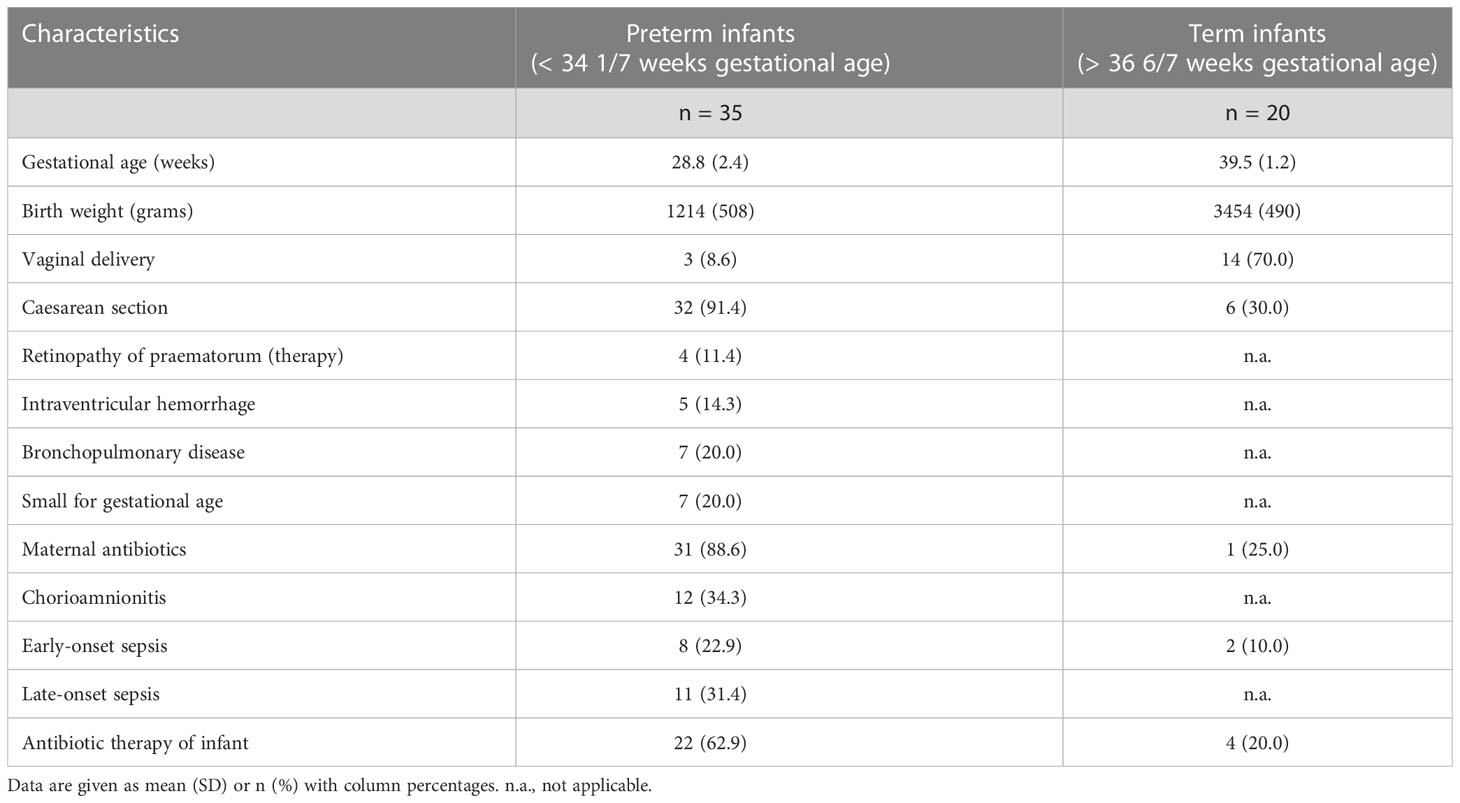
Table 1 Baseline characteristics of included preterm and term infants.
The inter-assay CV for the psoriasin ELISA was 8.1%, and the intra-assay CV was 8.5%. For RNase 7, the inter-assay CV was 6.9%, and the intra-assay CV was 8.1%.
On day 0, the median expression of psoriasin was similar in both preterm and term infants [preterm = 2.1 (IQR: 1.1–5.9) vs. term = 2.1 (1.3–4.4) ng/ml, p = 0.320], but RNase 7 was more expressed in term infants on day 0 [premature = 1.1 (0.7–2.9) vs. term = 2.0 (1.1–3.4) ng/ml, p = 0.017]. On day 28, premature infants showed higher values of psoriasin [preterm = 10.9 (5.6–14.2) vs. term = 6.3 (3.4–9.0) ng/ml, p < 0.001], but not for RNase 7 [preterm = 3.4 (2.4–4.9) vs. term = 3.5 (1.7–5.1) ng/ml, p = 0.574] (see Figure 2).
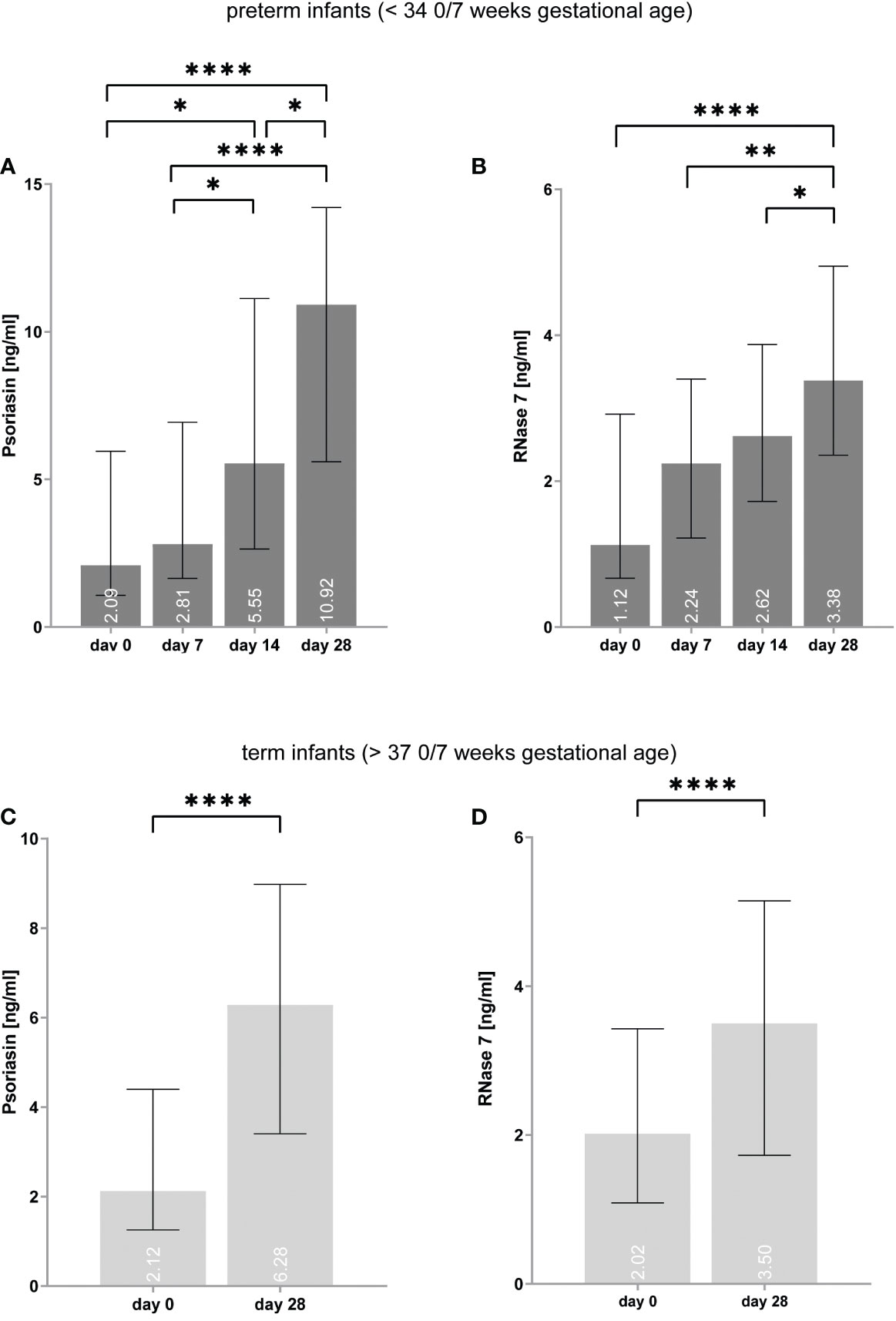
Figure 2 (A–D) Expression of psoriasin and RNase 7 in premature (A, B) and term (C, D) infants increases over time. *p < 0.05, **p < 0.01, and ****p < 0.0001 (ordinary one-way ANOVA with Holm–Sidak’s multiple comparisons test for preterm infants, Wilcoxon signed-rank test for term infants). Medians are given for each day in white numbers.
In preterm infants, psoriasin concentration was found to elevate from median values of 2.1 (1.1–5.9) ng/ml on day 0 to 10.9 (5.6–14.2) ng/ml on day 28 (p < 0.0001) (see Figure 2). In term infants, psoriasin increased from 2.1 (1.3–4.4) ng/ml to 6.3 (3.4–9.0) ng/ml (p < 0.0001). RNase 7 was also found to have higher concentrations on day 28 with 1.1 (0.7–2.9) ng/ml on day 0 and 3.4 (2.4–4.9) ng/ml on day 28 in premature infants (p < 0.0001) as well in term infants [2.0 (1.1–3.4) vs. 3.5 (1.7–5.1) ng/ml, p < 0.0001].
In utero exposure to placental inflammation, i.e., histological chorioamnionitis, increased the psoriasin [3.0 (1.4–7.7) vs. 9.9 (5.6–13.0) ng/ml, p < 0.0001] expression on the skin of preterm infants (see Figure 3), particularly for day 0 [1.4 (0.8–3.1) vs. 6.9 (3.4–12.3) ng/ml, p < 0.0001] and day 7 [2.7 (1.5–6.2) vs. 6.3 (3.1–13.2) ng/ml, p = 0.014] after birth. No significant effect was noted for the RNase 7 [2.1 (0.9–3.5) vs. 2.6 (1.2–4.0) ng/ml, p = 0.058] expression.
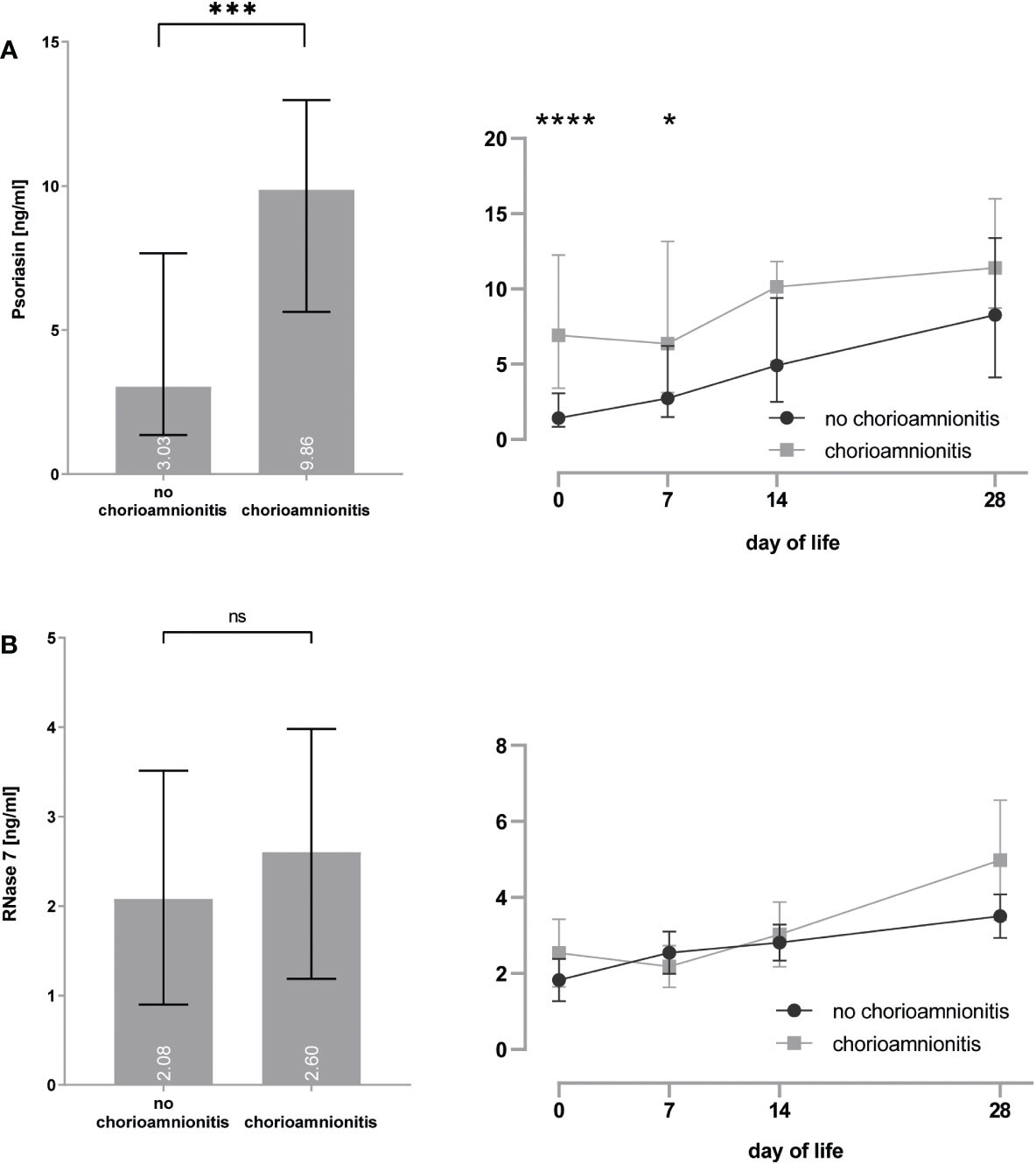
Figure 3 (A, B) Premature infants with histological proof of chorioamnionitis show increased concentrations of psoriasin (A), but no differences for RNase 7 (B) when compared to infants without signs of chorioamnionitis. *p < 0.05, ***p < 0.001, and ****p < 0.0001 (Mann–Whitney U-test for left bar graphs, Kruskal–Wallis Test for right line diagram). Medians are given for each day in white numbers. ns, not significant; p > 0.05.
Preterm infants with EOS showed increased psoriasin concentrations [4.0 (1.5–9.1) vs. 7.9 (4.7–12.1) ng/ml, p = 0.0002] particularly on day 0 [1.5 (1.0–4.5) vs. 5.3 (2.1–7.4) ng/ml, p = 0.014] and decreased RNase 7 concentrations [2.4 (1.2–4.0) vs. 1.6 (0.8–3.1), p = 0.01] (see Figure 4).
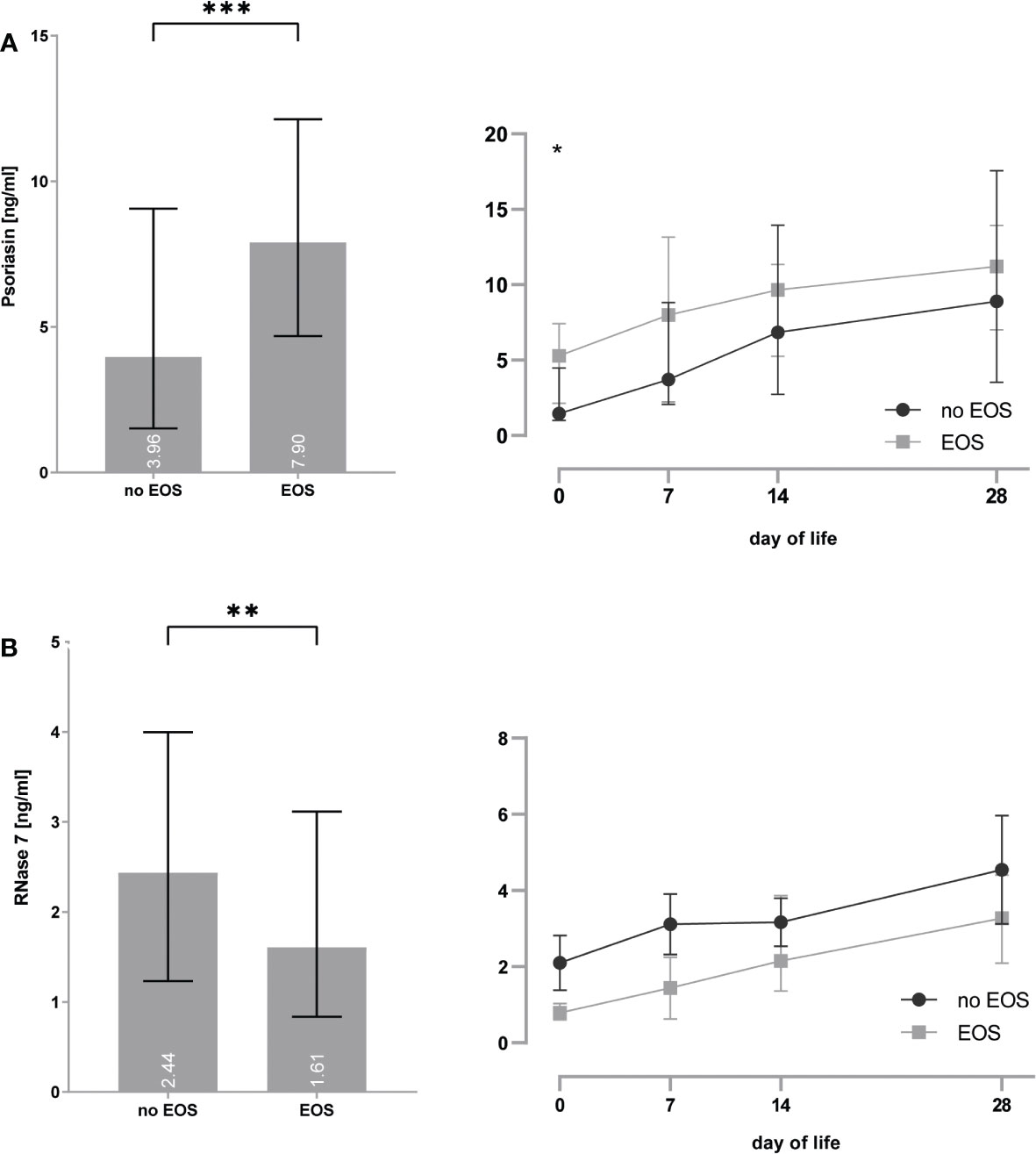
Figure 4 (A, B) Premature infants with clinical signs of early-onset sepsis (EOS) show increased concentrations of psoriasin (A) and decreased concentrations of RNase 7 (B) when compared to infants without signs of EOS. *p < 0.05, **p < 0.01, and ***p < 0.001 (Mann–Whitney U-test for left bar graphs, Kruskal–Wallis Test for right line diagram). Medians are given for each day in white numbers.Infants with LOS had increased expression of psoriasin [2.6 (1.5–6.5) vs. 7.4 (2.7–11.7), p < 0.0001], especially on day 7 [2.0 (1.3–3.6) vs. 7.6 (2.8–10.9), p = 0.008], but no differences in the concentrations of RNase 7 were found (see Figure 5).
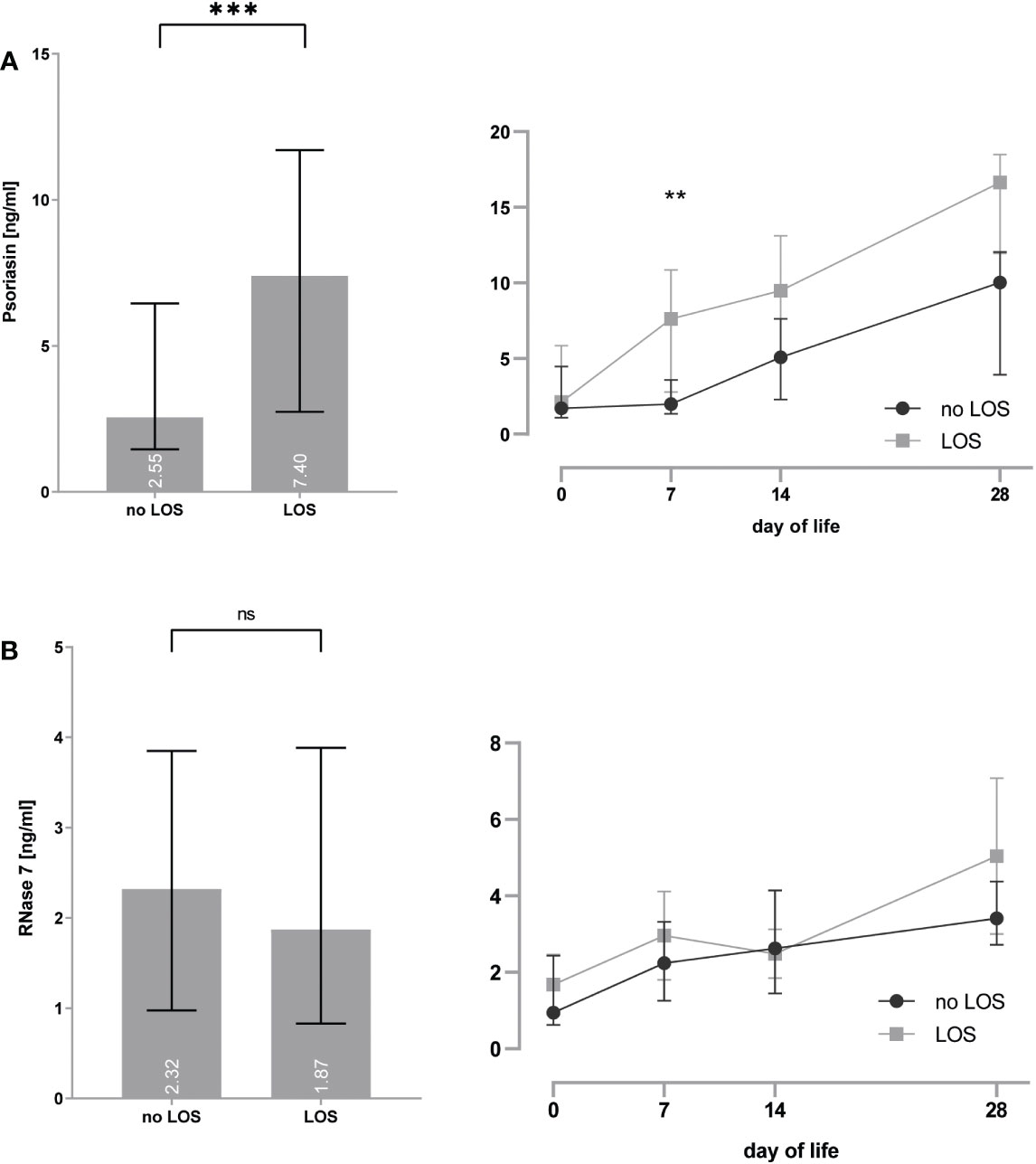
Figure 5 (A, B) Premature infants with clinical signs of late-onset sepsis (LOS) show increased concentrations of psoriasin (A), but no differences for RNase 7 (B) when compared to infants without signs of LOS. **p < 0.01 and ***p < 0.0001 (Mann–Whitney U-test for left bar graphs, Kruskal–Wallis Test for right line diagram). Medians are given for each day in white numbers. ns, not significant; p > 0.05.
Left and right upper thigh indicating use of sunflower oil for skin care in preterm infants with (right) and without (left upper thigh) care did not show significant differences for AMP expression (Table 2).
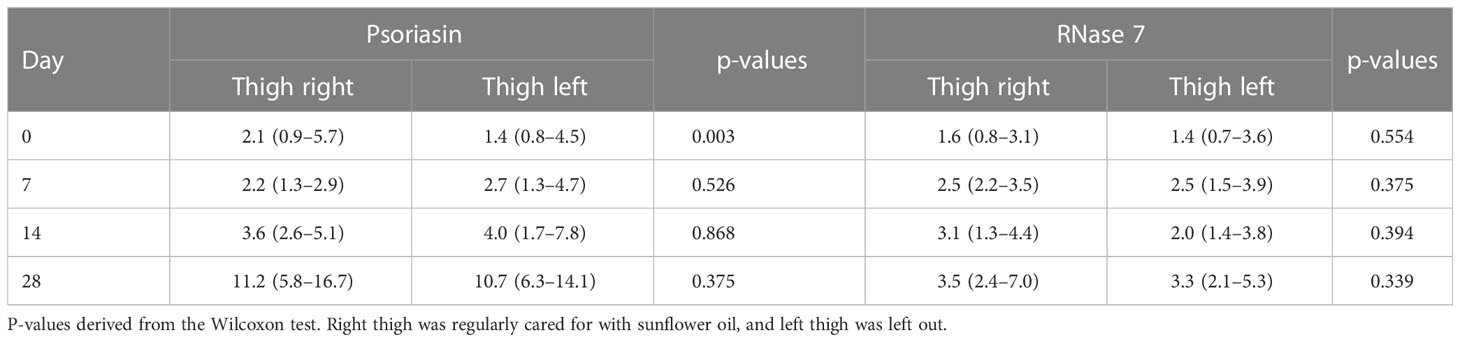
Table 2 Median (IQR) of psoriasin and RNase 7 concentrations of different probe sites and different days in preterm infants.
After exclusion of premature infants with proof of chorioamnionitis and sepsis (EOS and LOS), n=14 infants remained for analysis. This subcohort is characterized for later weeks of gestational age [30.1 (2.4) weeks] and increased birth weight [1,423 (643) g]. Here, concentrations of psoriasin and RNase 7 differ between first days of life and day 28 (see Table 3). We compared medians of psoriasin and RNase 7 in premature and term infants with exclusion of those with inflammatory complications. No further differences were found concerning the concentrations on days 0 and 28 in both groups (see Table 4).
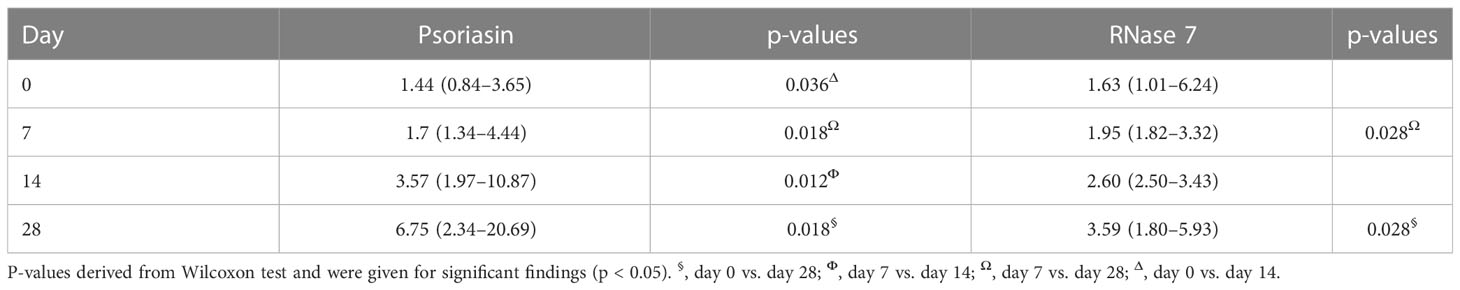
Table 3 Median (IQR) concentrations of psoriasin and RNase 7 in premature infants after exclusion of infants with chorioamnionitis and/or sepsis.
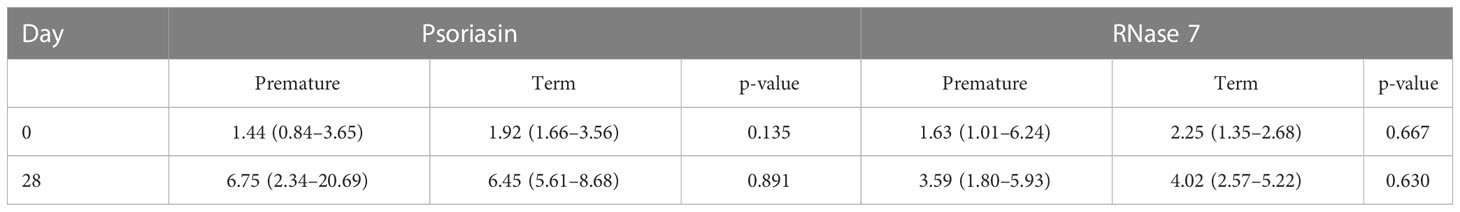
Table 4 Median (IQR) concentrations of psoriasin and RNase 7 in premature and term infants after exclusion of infants with chorioamnionitis and/or sepsis. P-values derived from Mann–Whitney U-test. No significant findings (p < 0.05).
In a convenience sample study, we demonstrated for the first time that psoriasin and RNAse 7 levels (a) accelerate expression over time and (b) correlate with chorioamnionitis (psoriasin) or sepsis. We further found that psoriasin and RNAse 7 levels do not differ between preterm and term infants with respect to day of life.
AMPs are essential molecules of the innate and adaptive defense and are expressed on sites with close contact to microorganisms such as the skin. Because of their broad antimicrobial properties against bacteria, viruses, and fungi, these peptides provide effective protection against pathogens but are also believed to shape the preferred microbiota (21). Typical AMP members expressed in human skin are the hBDs, cathelicidin (LL-37), psoriasin, RNase 7, dermcidin, and adrenomedullin (8, 22–26). In vernix caseosa, a substance covering the human fetus from the 28th week of gestation and in the epidermis of fetuses from 18 weeks, AMPs including psoriasin and RNase 7 are detectable (27–29). Because AMP measurement on the vulnerable skin surface of newborns requires the use of non-invasive methods, we used an established skin rinsing method to quantify AMPs on the newborn skin. Skin rinsing is an established method to quantify AMPs and comparable with the tape stripping method (30). Increase of AMP level on the skin surface of patients with inflamed skin (e.g., psoriasis and atopic dermatitis) measured by the skin rinsing method correlates well with increase of AMP immunostainings in biopsies (31). We also measured levels of hBD-2 and hBD-3, which were under the detection limit of the ELISA. In our prospective study, we noted that, with increasing postnatal age, concentrations of psoriasin und RNase 7 rise toward those found on healthy adult skin. The reason for this is unknown but could be assumed as an accompanying effect of AMP development with postnatal epithelial development as the skin structures mature within the first weeks of life (32, 33). Another reason for increasing concentrations after birth could be the contact of the skin to the outer world (29). AMPs play an essential role in maintaining a homogeneous colonization of the skin with microorganisms. For example, Staphylococcus epidermidis is the most frequently isolated species of the skin microbiome and a common pathogen of neonatal LOS (34). There is emerging evidence that the induction of RNase 7 and other skin AMPs is triggered by members of the microbiota such as Staphylococcus epidermidis to shape and balance skin colonization (21). Colonization with bacterial strains starts immediately after birth and could be an important component for increasing AMPs. The increase of AMP expression may prepare the skin to cope with or avoid future infections.
In our study, preterm infants showed higher concentrations of psoriasin than term controls. This observation can be explained by the high rate of inflammatory events such as chorioamnionitis, EOS, and LOS in this vulnerable cohort. Infants with these complications show increased levels of psoriasin. This effect is found at first days of life in infants with chorioamnionitis and EOS and at later days in infants with LOS, indicating a time-dependent inflammatory reaction. AMPs are important effector molecules in the regulation and communication of the innate and adaptive immune system through the control of cell migration, proliferation and differentiation, modulation of toll-like receptors, and the production of cytokines (35–40). Stimuli such as the expression of proinflammatory cytokines, mechanical injuries, or inflammatory processes lead to an upregulation of AMPs (32) and could explain our observation of higher psoriasin levels in premature infants. Total concentrations of RNase 7 were decreased in infants with EOS. This is of particular interest as, for example, in adult patients, reduced RNase 7 concentrations in urines are associated with a higher risk for urinary tract infections (41). This might explain and give a reason why certain populations are more susceptible to infections, which needs to be subject for large-scale prospective studies.
Neonatal sepsis is often the result of infections with bacterial, viral, or fungal microorganisms with a case fatality rate inversely related to gestational age (42). Transmission of pathogens in EOS occurs in utero transplacentar, via ascending bacteria entering the uterus or during the passage of the newborn through the birth canal. Late-onset bloodstream infections occur more frequently in neonates with central venous access or other medical devices. However, the intestines and lungs of especially preterm infants are also an important source of infections.
We could show that preterm and term infants show reduced concentrations of AMPs at the beginning of life but develop similar concentrations on the skin within the first weeks of life. This observation joins many studies on the development of the immune system that is constantly maturing in newborn infants (43). The ability to modulate the immune responses has been reported for several skin AMPs, and it could be argued that reduced concentrations of AMPs on the skin after birth increase the risk for neonatal infections. However, the concentrations of AMPs in preterm infants differ not from those derived from healthy full-term newborns. As preterm infants have a 3–10 times higher incidence of infection than full-term infants (44), our observation cannot fully explain the increased risk for preterm infants. Taking our findings into account, the risk for sepsis should be nearly similar in both cohorts. Therefore, other immune-specific factors will play a role in the increase of infection risk in neonates.
In our cohort, the skin of premature infants is regularly cared for with sunflower oil. We therefore let out skin care on the left thigh to control for skin care effects. We here found no differences in the expression of psoriasin or RNase 7 in dependence of use of sunflower oil. Phototherapy did not influence the expression of AMPs (data not shown).
Our study has some limitations. As this study was initially planned as explorative study, the first 13 participants were only analyzed on day one and missing on the later examination dates. Furthermore, the explorative design does not allow to adjust for all different clinical parameters that potentially affect AMP expression. For example, the role of chorioamnionitis could not be classified in more detail in our analysis, as we did not record detailed placental pathology findings, e.g., Blanc-defined stages. Further analyses should take this information into account.
We analyzed the amount of psoriasin and RNase 7 as these AMPs are the most abundant ones found on human skin that serve immunomodulatory roles in skin immunity. The methods of collecting and analyzing skin rinsing probes is well-established (45). Different techniques for AMP sampling exist, which may hamper direct comparison between studies. Most studies use analysis of skin biopsies, tape strips, and rinsing solution from the skin. Rinsing solution techniques have the advantage as a minimally invasive technique and its possibility for multiple testing of the same patient over time. The use of skin biopsies has been predominant but has the disadvantage of being invasive and contains the risk of catching deeper layers of the skin with AMPs from other cells rather than AMPs locally present (46).
In conclusion, this study shows that with increasing age of infants, concentrations of skin derived AMPs rise and that infectious complications modify the expression of psoriasin and RNase 7. Further studies are needed to clarify if varying concentrations of skin AMPs are dependent from maturing skin structures or developing microbial colonization and if AMP concentrations impact the risk for infections in newborns.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by the local ethics committee for research in human subjects of the University of Lübeck (file number 19-028). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Conceived and designed the experiments: AH, LN, HB, HH, JH, and CH. Performed the experiments: LN, HH, JH, and HB. Analyzed the data: AH, JH, MF, FR, and HH. Contributed reagents/materials/analysis tools: AH, HH, JH, MF, and EH. Wrote the paper: AH, JH, CH, and EH. The authors gave the final approval of the version to be published. Each author participated sufficiently in the work to take public responsibility for appropriate portions of the content and agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors contributed to the article and approved the submitted version.
This work was supported by the Department of Pediatrics, University Hospital of Schleswig-Holstein, Campus Lübeck; and by the Department of Dermatology, Venerology and Allergology, Quincke Research Center, University-Hospital Schleswig-Holstein, Campus Kiel. The study sponsors had no involvement on study design, the collection, analysis, and interpretation of data; the writing of the report; and the decision to submit the manuscript for publication. Individual participant data reported in this article are available on reasonable request. Proposals may be submitted up to 36 months following article publication.
The authors would like to thank all participating infants and their parents.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
EOS, early-onset sepsis; LOS, late-onset sepsis; AMP, antimicrobial peptide.
1. Battersby AJ, Khara J, Wright VJ, Levy O, Kampmann B. Antimicrobial proteins and peptides in early life: Ontogeny and translational opportunities. Front Immunol (2016) 7:309. doi: 10.3389/fimmu.2016.00309
2. Harder J, Schröder JM. Antimicrobial peptides in human skin. Chem Immunol Allergy (2005) 86:22–41. doi: 10.1159/000086650
3. Capone KA, Dowd SE, Stamatas GN, Nikolovski J. Diversity of the human skin microbiome early in life. J Invest Dermatol (2011) 131:2026–32. doi: 10.1038/jid.2011.168
4. Huttner KM, Bevins CL. Antimicrobial peptides as mediators of epithelial host defense. Pediatr Res (1999) 45:785–94. doi: 10.1203/00006450-199906000-00001
5. Simanski M, Köten B, Schröder JM, Gläser R, Harder J. Antimicrobial RNases in cutaneous defense. J Innate Immun (2012) 4(3):241–7. doi: 10.1159/000335029
6. Becknell B, Spencer JD. A review of ribonuclease 7’s structure, regulation, and contributions to host defense. Int J Mol Sci (2016) 17(3):423. doi: 10.3390/ijms17030423
7. Rademacher F, Dreyer S, Kopfnagel V, Gläser R, Werfel T, Harder J. The antimicrobial and immunomodulatory function of RNase 7 in skin. Front Immunol (2019) 10:2553. doi: 10.3389/fimmu.2019.02553
8. Harder J, Schröder J-M. RNase 7, a novel innate immune defense antimicrobial protein of healthy human skin. J Biol Chem (2002) 277:46779–84. doi: 10.1074/jbc.M207587200
9. Gläser R, Köten B, Wittersheim M, Harder J. Psoriasin: Key molecule of the cutaneous barrier? JDDG J der Dtsch Dermatologischen Gesellschaft (2011) 9(11):897–902. doi: 10.1111/j.1610-0387.2011.07683.x
10. Humberg A, Fortmann I, Siller B, Kopp MV, Herting E, Göpel W, et al. Preterm birth and sustained inflammation: consequences for the neonate. Semin Immunopathol (2020) 42:451–68. doi: 10.1007/s00281-020-00803-2
11. Collins A, Weitkamp JH, Wynn JL. Why are preterm newborns at increased risk of infection? Arch Dis Child Fetal Neonatal Ed (2018) 103(4):F391–F394. doi: 10.1136/archdischild-2017-313595
12. Jenke ACW, Zilbauer M, Postberg J, Wirth S. Human β-defensin 2 expression in ELBW infants with severe necrotizing enterocolitis. Pediatr Res (2012) 72:513–20. doi: 10.1038/pr.2012.110
13. Starner TD, Agerberth B, Gudmundsson GH, McCray PB. Expression and activity of beta-defensins and LL-37 in the developing human lung. J Immunol (2005) 174:1608–15. doi: 10.4049/jimmunol.174.3.1608
14. Faust K, Göpel W, Moser K, Temole G, Bartels M, Wieg C, et al. Differential expression of antimicrobial polypeptides in cord blood samples of preterm and term infants. Acta Paediatr (2014) 103:e143–7. doi: 10.1111/apa.12544
15. Schaller-Bals S, Schulze A, Bals R. Increased levels of antimicrobial peptides in tracheal aspirates of newborn infants during infection. Am J Respir Crit Care Med (2002) 165:992–5. doi: 10.1164/ajrccm.165.7.200110-020
16. Schuster C, Gläser R, Fiala C, Eppel W, Harder J, Schröder JM, et al. Prenatal human skin expresses the antimicrobial peptide RNase 7. Arch Dermatol Res (2013) 305:545–9. doi: 10.1007/s00403-013-1340-y
17. Wittersheim M, Cordes J, Meyer-Hoffert U, Harder J, Hedderich J, Gläser R. Differential expression and in vivo secretion of the antimicrobial peptides psoriasin (S100A7), RNase 7, human beta-defensin-2 and -3 in healthy human skin. Exp Dermatol (2013) 22(5):364–6. doi: 10.1111/exd.12133
18. Gläser R, Navid F, Schuller W, Jantschitsch C, Harder J, Schröder JM, et al. UV-B radiation induces the expression of antimicrobial peptides in human keratinocytes in vitro and in vivo. J Allergy Clin Immunol (2009) 123(5):1117-23. doi: 10.1016/j.jaci.2009.01.043
19. Leistner R, Piening B, Gastmeier P, Geffers C, Schwab F. Nosocomial infections in very low birthweight infants in Germany: Current data from the national surveillance system NEO-KISS. Klin Padiatr (2013) 225:75–80. doi: 10.1055/s-0033-1334886
20. Voigt M, Rochow N, Schneider K, Hagenah H-P, Scholz R, Hesse V, et al. New percentile values for the anthropometric Dimensions of twin neonates: analysis of perinatal survey data of 2007-2011 from all 16 states of Germany. Z Geburtshilfe Neonatol (2014) 218:210–7. doi: 10.1055/s-0034-1385857
21. Rademacher F, Gläser R, Harder J. Antimicrobial peptides and proteins: Interaction with the skin microbiota. Exp Dermatol (2021) 30:1496–508. doi: 10.1111/exd.14433
22. Braff MH, Bardan A, Nizet V, Gallo RL. Cutaneous defense mechanisms by antimicrobial peptides. J Invest Dermatol (2005) 125:9–13. doi: 10.1111/j.0022-202X.2004.23587.x
23. Braff MH, Zaiou M, Fierer J, Nizet V, Gallo RL. Keratinocyte production of cathelicidin provides direct activity against bacterial skin pathogens. Infect Immun (2005) 73:6771–81. doi: 10.1128/IAI.73.10.6771-6781.2005
24. Madsen P, Rasmussen HH, Leffers H, Honoré B, Dejgaard K, Olsen E, et al. Molecular cloning, occurrence, and expression of a novel partially secreted protein “Psoriasin” that is highly up-regulated in psoriatic skin. J Invest Dermatol (1991) 97:701–12. doi: 10.1111/1523-1747.ep12484041
25. Schittek B, Hipfel R, Sauer B, Bauer J, Kalbacher H, Stevanovic S, et al. Dermcidin: a novel human antibiotic peptide secreted by sweat glands. Nat Immunol (2001) 2:1133–7. doi: 10.1038/ni732
26. Allaker R. An investigation into the antimicrobial effects of adrenomedullin on members of the skin, oral, respiratory tract and gut microflora. FEMS Immunol Med Microbiol (1999) 23:289–93. doi: 10.1016/S0928-8244(98)00148-5
27. Marchini G, Lindow S, Brismar H, Stabi B, Berggren V, Ulfgren A-K, et al. The newborn infant is protected by an innate antimicrobial barrier: Peptide antibiotics are present in the skin and vernix caseosa. Br J Dermatol (2002) 147:1127–34. doi: 10.1046/j.1365-2133.2002.05014.x
28. Yoshio H, Tollin M, Gudmundsson GH, Lagercrantz H, Jörnvall H, Marchini G, et al. Antimicrobial polypeptides of human vernix caseosa and amniotic fluid: Implications for newborn innate defense. Pediatr Res (2003) 53:211–6. doi: 10.1203/01.PDR.0000047471.47777.B0
29. Visscher MO, Adam R, Brink S, Odio M. Newborn infant skin: Physiology, development, and care. Clin Dermatol (2015) 33:271–80. doi: 10.1016/j.clindermatol.2014.12.003
30. Ono S, Eda N, Mori T, Otsuka A, Nakamura N, Inai Y, et al. Tape stripping method is useful for the quantification of antimicrobial peptides on the human skin surface including the stratum corneum. Sci Rep (2020) 10:15259. doi: 10.1038/s41598-020-72111-6
31. Harder J, Dressel S, Wittersheim M, Cordes J, Meyer-Hoffert U, Mrowietz U, et al. Enhanced expression and secretion of antimicrobial peptides in atopic dermatitis and after superficial skin injury. J Invest Dermatol (2010) 130:1355–64. doi: 10.1038/jid.2009.432
32. Clausen M-L, Agner T. Antimicrobial peptides, infections and the skin barrier. Curr Probl Dermatol (2016) 49:38–46. doi: 10.1159/000441543
33. Niyonsaba F, Kiatsurayanon C, Chieosilapatham P, Ogawa H. Friends or foes? host defense (antimicrobial) peptides and proteins in human skin diseases. Exp Dermatol (2017) 26:989–98. doi: 10.1111/exd.13314
34. Dong Y, Speer CP, Glaser K. Beyond sepsis: Staphylococcus epidermidis is an underestimated but significant contributor to neonatal morbidity. Virulence (2018) 9:621–33. doi: 10.1080/21505594.2017.1419117
35. Niyonsaba F, Ushio H, Nagaoka I, Okumura K, Ogawa H. The human β-defensins (-1, -2, -3, -4) and cathelicidin LL-37 induce IL-18 secretion through p38 and ERK MAPK activation in primary human keratinocytes. J Immunol (2005) 175:1776–84. doi: 10.4049/jimmunol.175.3.1776
36. Niyonsaba F, Iwabuchi K, Matsuda H, Ogawa H, Nagaoka I. Epithelial cell-derived human β-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase c-dependent pathway. Int Immunol (2002) 14:421–6. doi: 10.1093/intimm/14.4.421
37. Chen X, Takai T, Xie Y, Niyonsaba F, Okumura K, Ogawa H. Human antimicrobial peptide LL-37 modulates proinflammatory responses induced by cytokine milieus and double-stranded RNA in human keratinocytes. Biochem Biophys Res Commun (2013) 433:532–7. doi: 10.1016/j.bbrc.2013.03.024
38. Di Nardo A, Braff MH, Taylor KR, Na C, Granstein RD, McInturff JE, et al. Cathelicidin antimicrobial peptides block dendritic cell TLR4 activation and allergic contact sensitization. J Immunol (2007) 178:1829–34. doi: 10.4049/jimmunol.178.3.1829
39. Yang D, Chen Q, Schmidt AP, Anderson GM, Wang JM, Wooters J, et al. Ll-37, the neutrophil granule–and epithelial cell–derived cathelicidin, utilizes formyl peptide receptor–like 1 (Fprl1) as a receptor to chemoattract human peripheral blood neutrophils, monocytes, and T cells. J Exp Med (2000) 192:1069–74. doi: 10.1084/jem.192.7.1069
40. Chessa C, Bodet C, Jousselin C, Wehbe M, Lévêque N, Garcia M. Antiviral and immunomodulatory properties of antimicrobial peptides produced by human keratinocytes. Front Microbiol (2020) 11:1155. doi: 10.3389/fmicb.2020.01155
41. Becknell B, Eichler TE, Beceiro S, Li B, Easterling RS, Carpenter AR, et al. Ribonucleases 6 and 7 have antimicrobial function in the human and murine urinary tract. Kidney Int (2015) 87:151–61. doi: 10.1038/ki.2014.268
42. Dijksteel GS, Ulrich MMW, Middelkoop E, Boekema BKHL. Review: Lessons learned from clinical trials using antimicrobial peptides (AMPs). Front Microbiol (2021) 12:616979. doi: 10.3389/fmicb.2021.616979
43. Ygberg S, Nilsson A. The developing immune system - from foetus to toddler. Acta Paediatr (2012) 101:120–7. doi: 10.1111/j.1651-2227.2011.02494.x
44. Shane AL, Sánchez PJ, Stoll BJ. Neonatal sepsis. Lancet (2017) 390:1770–80. doi: 10.1016/S0140-6736(17)31002-4
45. Hermes BM, Rademacher F, Chung C, Tiegs G, Bendix M-C, de Zwaan M, et al. Skin microbiota analysis in patients with anorexia nervosa and healthy-weight controls reveals microbial indicators of healthy weight and associations with the antimicrobial peptide psoriasin. Sci Rep (2022) 12:15515. doi: 10.1038/s41598-022-19676-6
Keywords: antimicrobial peptides, premature infants, sepsis, skin barrier, chorioamnionitis
Citation: Humberg A, Neuenburg L, Boeckel H, Fortmann MI, Härtel C, Herting E, Hinrichs H, Rademacher F and Harder J (2023) Antimicrobial skin peptides in premature infants: Comparison with term infants and impact of perinatal factors. Front. Immunol. 14:1093340. doi: 10.3389/fimmu.2023.1093340
Received: 15 December 2022; Accepted: 14 February 2023;
Published: 27 February 2023.
Edited by:
Kazunari Kaneko, Kansai Medical University, JapanReviewed by:
Hideaki Tanizaki, Kansai Medical University, JapanCopyright © 2023 Humberg, Neuenburg, Boeckel, Fortmann, Härtel, Herting, Hinrichs, Rademacher and Harder. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander Humberg, YWxleGFuZGVyLmh1bWJlcmdAdWttdWVuc3Rlci5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.