 Arnolda Jakovija
Arnolda Jakovija Tatyana Chtanova
Tatyana Chtanova
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 16 June 2023
Sec. Cytokines and Soluble Mediators in Immunity
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1060258
This article is part of the Research TopicInsights into the immunological microenvironment within neoplastic and inflammatory skin conditionsView all 7 articles
The skin is the body’s largest organ. It serves as a barrier to pathogen entry and the first site of immune defense. In the event of a skin injury, a cascade of events including inflammation, new tissue formation and tissue remodeling contributes to wound repair. Skin-resident and recruited immune cells work together with non-immune cells to clear invading pathogens and debris, and guide the regeneration of damaged host tissues. Disruption to the wound repair process can lead to chronic inflammation and non-healing wounds. This, in turn, can promote skin tumorigenesis. Tumors appropriate the wound healing response as a way of enhancing their survival and growth. Here we review the role of resident and skin-infiltrating immune cells in wound repair and discuss their functions in regulating both inflammation and development of skin cancers.
The skin is not only a physical barrier protecting us from infection but also an important immunological site, which in humans contains an estimated 20 billion T cells as well as a range of cells with innate and innate-like roles. Among them are Langerhans cells, dermal dendritic cells (DCs), macrophages, neutrophils, mast cells and innate lymphoid cells. These immune cells, together with keratinocytes and neurons, interact with the skin microbiota, to maintain skin homeostasis while protecting against pathogen invasion. Several recent reviews (1–3) explain how immune subsets and specialized immunological sites (such as hair follicles and sweat glands) interact with the skin microbiome. This review will focus specifically on how skin immune cells mediate wound repair and how this process can be co-opted by tumors.
Wound healing is a natural physiological reaction to tissue injury designed to prevent the onset of infection and restore tissue integrity (4). It follows a finely coordinated multistep process that includes hemostasis, inflammation, proliferation (new tissue formation), and tissue remodeling (5) (Figure 1).
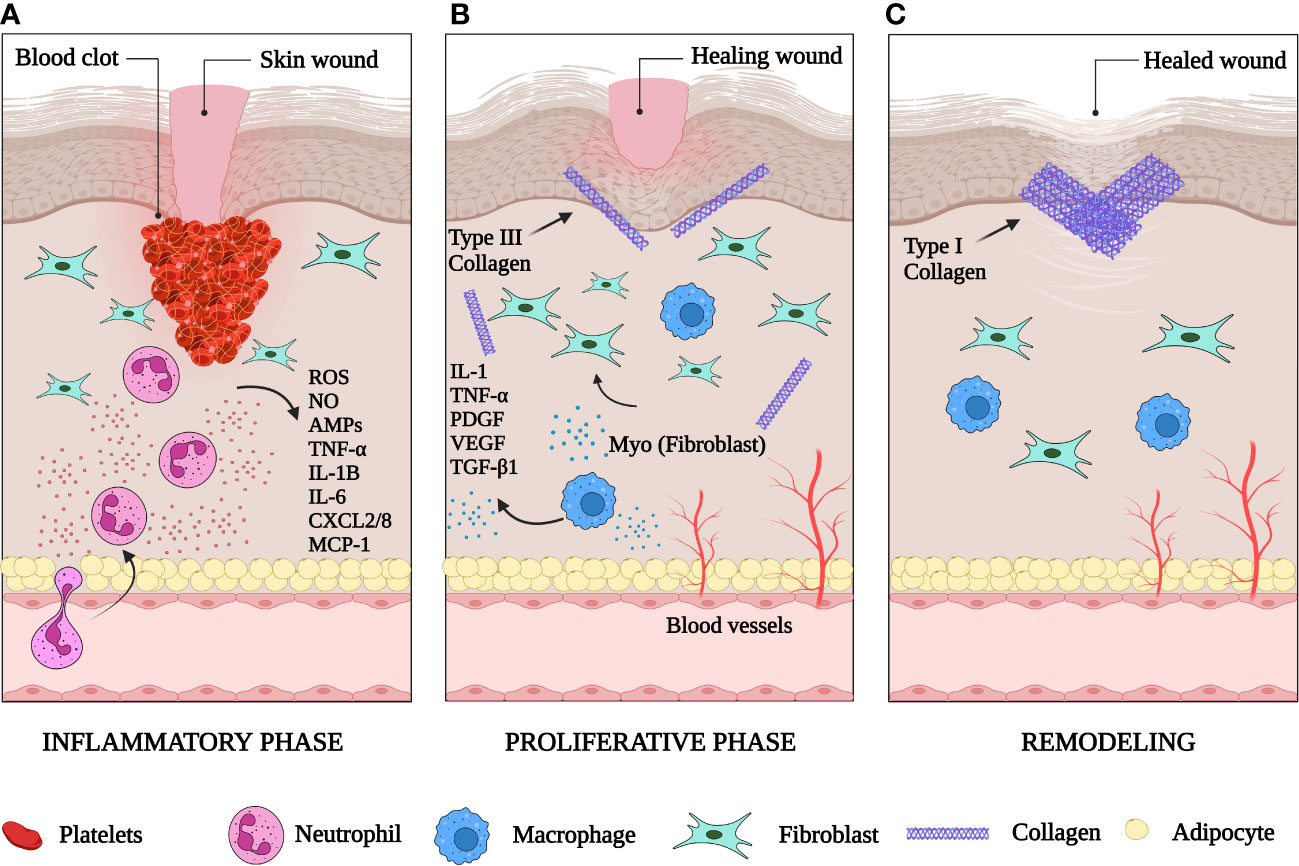
Figure 1 The phases of skin wound healing. (A) The inflammatory phase: one to three days after injury the wound is filled with a clot. Inflammatory cells have been recruited to the wound site. Neutrophils release reactive oxygen species (ROS), nitric oxide (NO), antimicrobial proteins (AMPs), TNFα, IL-1B, IL-6, CXCL2/8 and monocyte attracting protein-1 (MCP-1). (B) The proliferative phase: macrophages are recruited to clear dead tissue and debris. They secrete IL-1, TNFα, PDGF, VEGF and TGF-β1. New blood vessels form in the wound bed. Fibroblasts are activated in the wound and begin to deposit collagen. (C) The remodeling phase: wound contraction occurs, collagen III is replaced by collagen I, and the extracellular matrix is remodeled by proteases and other enzymes. Created with BioRender.com.
Vascular damage with resultant local hemorrhage is a universal characteristic of tissue injury (6). A few minutes after injury, platelets in the circulation begin to stick to the injured site and promote formation of blood clots (7), made up predominantly of crosslinked fibrin, plasma fibronectin and other extracellular matrix (ECM) proteins, such as vitronectin and thrombospondins (8).
Inflammatory cells enter damaged tissues through diapedesis by way of venules within minutes after injury (9, 10). Neutrophils are the first immune subset to respond to cutaneous damage (11). They deploy their antimicrobial arsenal to phagocytose and kill contaminating microorganisms and secrete an array of cytokines that recruit macrophages, T cells and additional neutrophils (12).
Mast cells are abundant in the skin and orchestrate the early stages of wound healing (13). They recognize interleukin (IL)-33 released by necrotic cells via ST2 receptor and secrete histamine and other cytokines that stimulate the immune response (13, 14). This is critical for attracting other immune cells to the wound and promoting inflammation (15).
Monocytes and macrophages follow neutrophils in wounds to remove dead cells and cellular debris and recruit T cells and natural killer (NK) cells to stimulate the proinflammatory response (16). Removal of dead neutrophils by macrophages heralds the end of the inflammatory period and the transition of macrophages to an M2 (anti-inflammatory) phenotype (17) [section 3.2]. This conversion from an M1 pro-inflammatory to M2 anti-inflammatory phenotype is a crucial step in the initiation of the proliferative and resolution phase (18). At the end of this stage, these macrophages either die at the wound or migrate to draining lymph nodes. These events promote subsequent wound healing phases (16) [section 2.2].
This stage includes angiogenesis, fibroplasia, and re-epithelialization which stimulate the closure of the lesion. Angiogenesis (formation of new microvasculature) enables transport of fluid, oxygen, nutrients, and immune-competent cells into the stroma (19). Fibroplasia commences with the formation of granulation tissue (20) and is characterized by the proliferation of fibroblasts, which deposit the collagen matrix required for adhesion and migration (21). Myofibroblasts, specialized fibroblasts with contractile properties, are responsible for the production of the ECM components that replace the temporary matrix in the wound within the granulation tissue (22). These cells have contractile abilities due to the presence of α-smooth muscle actin (α-SMA) in their microfilament bundles, making them a significant contributor to the contraction and maturation of the granulation tissue (23). The transition from the inflammatory to the proliferative phase occurs two to four weeks after injury as epithelial cells start the process of re-epithelialization that involves their proliferation and migration from the borders of the wound.
This phase marks the transition from granulation tissue to scar. It starts one to two weeks after wounding and continues for up to two years (24). At this stage, wound tissue is mainly dominated by collagen type I, which has replaced collagen type III (19). This results in the formation of a scar that contains dense connective tissue of reduced tensile strength and elasticity compared with normal skin (25). Granulation tissue is replaced by acellular scar after the completion of wound repair and myofibroblast apoptosis (26).
Acute wound healing is a highly regulated process that leads to the restoration of tissue integrity and resolution of inflammation. However, chronic wounds (like diabetic ulcers) can develop if the inflammatory process is not succeeded by the repair phase (27, 28). Many inflammatory skin conditions (such as atopic dermatitis, psoriasis, discoid lupus erythematosus) involve disruptions in immune function and signaling (29). This can result in persistent activation and increased production of pro-inflammatory molecules such as chemokines and cytokines, which exacerbate inflammation and cause abnormal cell growth (30). Diseases such as rheumatoid arthritis and psoriasis also show characteristics of aberrant wound healing (31, 32). Notably, chronic wound state is a risk factor for cancer development (33) and can promote malignant transformation (29, 34).
The granulation tissue of healing skin wounds contains a mixture of cells, including fibroblasts, blood vessels, and inflammatory cells. This strongly resembles the tumor stroma suggesting that epithelial tumors promote the formation of their stroma by activating the wound healing response of the host, which leads to the formation of new blood vessels and fibroblasts. This suggests that tumors hijack the proliferative program of wound repair to support their proliferation. The tumor microenvironment (TME) also shapes immune cell function to enhance an immunosuppressive and pro-angiogenic state, aiding tumor immune evasion and promoting metastasis (Figure 2). Unlike a wound, the tumor continues to grow uncontrollably, without the resolution of inflammation and proper tissue repair. This evidence has led to the suggestion that tumors represent ‘wounds that never heal’ (35). This idea is supported by the fact that many of the same signaling pathways and cellular players involved in wound healing are also activated in tumor development (36). But while wound healing involves the migration and proliferation of healthy cells to repair the damaged tissue, tumor cells acquire genetic changes that allow them to invade the surrounding tissues and metastasize (36).
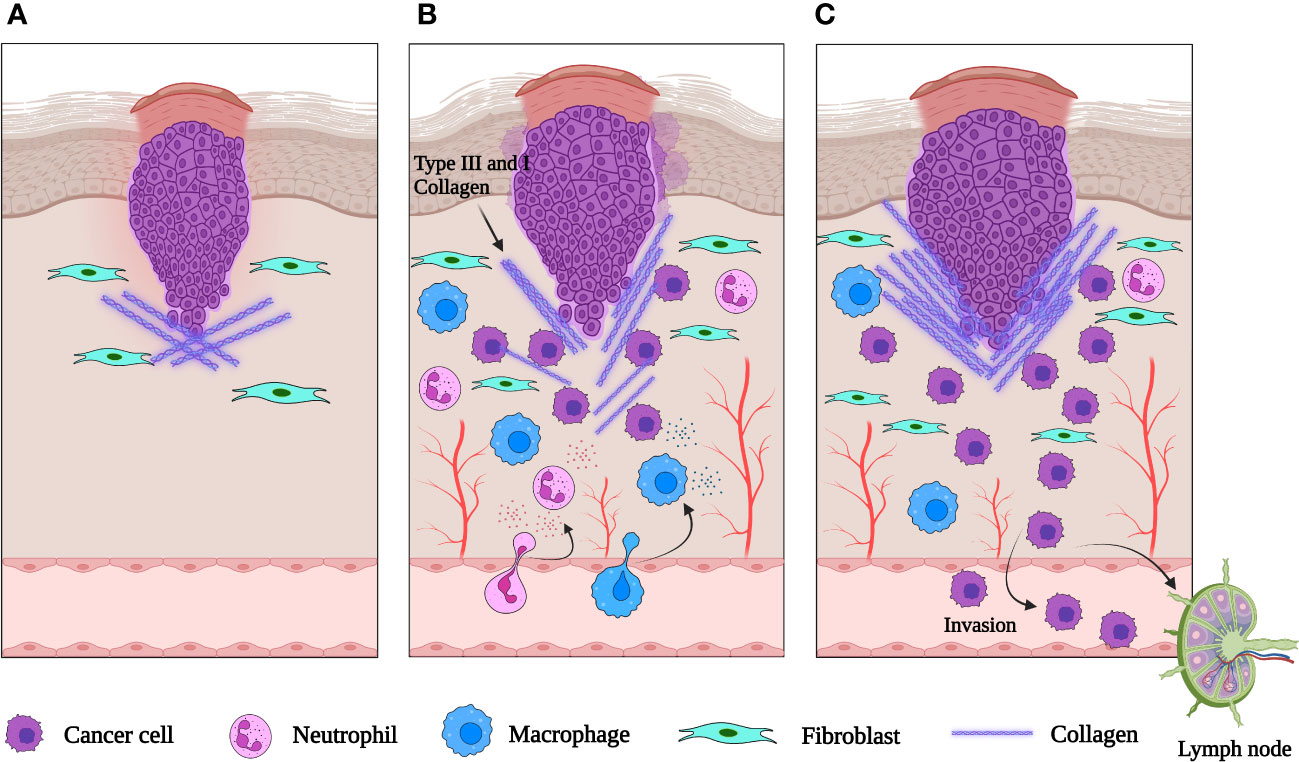
Figure 2 Schematic representation of an epithelial tumor. (A) When neoplasia is first initiated, fibroblasts are recruited to the tumor site and activated. (B) As the tumor grows, inflammatory cells are recruited to the tumor and release cytokines. VEGF and other signaling molecules induce neovascularization. (C) The abnormal extracellular matrix is pro-tumorigenic, pro-angiogenic and increases the invasiveness of the tumor. Created with BioRender.com.
In both wound healing and cancer, the initial inflammatory response is necessary to recruit immune cells to the site of injury or to the TME (36). But in chronic wounds or cancer, the inflammatory response becomes dysregulated and promotes further tissue damage, leading to impaired healing or tumor progression (37). Moreover, several studies have demonstrated an association between chronic wounds and skin cancer (38). The specific functions of immune cells can vary depending on the type of cancer and the stage of the disease, and more research is needed to fully understand the role of immune cells in skin repair and skin cancer. Next, we will review in detail the functions of immune cell subsets and inflammatory mediators in cutaneous wound healing and cancers.
Several molecules attract neutrophils to wounded skin: damage-associated molecular patterns (DAMPs); proinflammatory cytokines, including TNF-α; chemoattractants, such as CXCL1–3 and CXCL8(IL-8); anaphylatoxins C3a and C5a and macrophage inflammatory protein-1α (39, 40). In addition, damaged mitochondria from necrotic cells release other early signals, such as fMet-Leu-Phe (fMLP), derived either from translocated commensal organisms or from necrotic host cell mitochondria (41). At the injury site, neutrophils destroy pathogens via phagocytosis and degranulation, release of highly concentrated reactive oxygen species (ROS), antimicrobial proteins (AMPs) and neutrophil extracellular traps (NETs) (42). They amplify inflammation by secreting cytokines and chemokines, such as TNF-α, IL-1β, IL-6, CXCL8 and CXCL2 (43). Neutrophils also recruit macrophages and T cells via monocyte attracting protein-1 (MCP-1) (44) and play an important role in modulating adaptive immunity in response to infectious wounds (45–47).
LTB4 released from early recruited neutrophils acts as a chemoattractant and mediates an effect known as “neutrophil swarming” (48, 49), a dynamic response to inflammation first observed in neutrophils using two-photon microscopy. Intravital imaging has provided important insight into neutrophil function in skin infection and injury (47, 50, 51). For instance, Lammerman et al. used it to show that the lipid leukotriene B4 was a critical mediator of intercellular signaling among swarming neutrophils after cutaneous thermal injury (48). As neutrophils rearranged the collagen fiber network to create a collagen-free zone at the center of the wound, their clusters were maintained via integrin receptors (48). Real time observation of neutrophil dynamics in zebrafish demonstrated that neutrophil migration to the wound was due to the production of hydrogen peroxide (52).
Neutrophils are not only essential for eradicating pathogens and inhibiting their propagation when the skin barrier is compromised, but also play a beneficial role in the restoration of epithelial tissues. After sterilizing the wound, neutrophils initiate an apoptotic cell-death pathway which leads to efferocytosis by macrophages (53). However, if this process is impaired, neutrophils persist in the wound microenvironment and their associated inflammatory mediators contribute to the formation of chronic wounds (54). Once activated, neutrophils then initiate wound closure, re-epithelialization and formation of new vessels by expressing cytokines and growth factors, including TNF-α and VEGF (40, 55, 56). Neutrophil-derived VEGF plays an important role, for example, in neovascularization of injured murine cornea (57), highlighting neutrophil contributions to restoring tissue architecture.
The importance of neutrophils in tissue repair has been demonstrated in several studies. For instance, mice lacking fMLP receptors 1 and 2 show delays in neutrophil accumulation during the acute stage of injury, resulting in delayed wound closure (58). Likewise, mice deficient in CXCR2, a chemokine receptor important for neutrophil recruitment to the wound site, exhibit delayed re-epithelialization of skin wounds and delayed wound healing (59). In addition, neutropenic patients and mice deficient in the neutrophil protease matrix metalloproteinase 8 (MMP-8) display reduced skin wound repair (60, 61). Interestingly, a new role for neutrophils in wound repair has recently been demonstrated in an internal injury model where neutrophils were shown to carry pre-existing matrix into wounds, promoting fibroblast activation and scar formation (62). Whether such a mechanism also exists in skin injuries is of considerable interest.
In cancer, the release of DAMPS caused by hypoxia, nutrient starvation, cellular proliferation, and necrosis in the TME can recruit and activate neutrophils (63). Tumor and stromal cells can also secrete CXCR2 ligands, such as CXCL1, CXCL2 and CXCL5 to attract neutrophils (64, 65). Within tumors, neutrophils can be located in either the peripheral region or within the tumor core (66). Neutrophils infiltrating the tumor core are less motile compared to the peritumoral neutrophils. The reduction in motility may allow neutrophils to accumulate and promote inflammation (66).
Neutrophil anti-microbial and wound repair functions can be coopted by tumors to mediate immunosuppression and metastasis (67). Neutrophil-derived ROS can suppress T and NK cell responses in tumors (68–70) and activate cellular proliferation or survival signaling pathways, such as the NF-κB pathway and the synthesis of transcription factors like STAT3 (71), which are constitutively activated in skin cancer (72). Oxidative stress regulates the expression of intercellular adhesion protein-1 (ICAM-1), which together with IL-8, controls the transendothelial migration of neutrophils and may contribute to tumor metastasis (73). Consistent with an important role for neutrophils in metastasis, intravital imaging showed that neutrophils are among the first immune cells to arrive at metastatic tumor sites (74), where neutrophil derived NETs act as an adhesion substrate for cancer cells and degrade the extracellular matrix (75–77). Other neutrophil-derived factors, such as granules containing neutrophil elastase (NE), neutrophil collagenase (or MMP8), and gelatinase B (or MMP9) can remodel the ECM in the TME or act directly on tumor cells themselves to boost tumor proliferation and invasion (78). Consistent with this, MMP9 stimulates keratinocyte proliferation and invasion in skin cancer models (79, 80). Tumor neutrophils can release cytokines like oncostatin M, which induces VEGF and increases angiogenesis and tumor cell invasion (81).
Notably, neutrophil-derived mediators of wound repair and pathogen control can also act to eradicate cancerous cells and restrict metastatic dissemination (82, 83). For instance, neutrophils can mediate direct tumor killing by releasing ROS and cytotoxic enzymes, or by recruiting and activating other immune cells, such as cytotoxic T cells (84, 85). This points to a complex role for neutrophils in tumor immunity, where wound repair and pathogen killing mechanisms are applied within the TME in a context and co-stimulation dependent manner which is not yet fully understood.
Skin has two distinct macrophage populations: tissue resident, which derive from the extraembryonic yolk sack, and monocyte-derived, which originate from the bone marrow-derived monocytes recruited to the skin (86). Tissue resident macrophages monitor the skin microenvironment for signals that indicate cell stress, tissue injury or infection (87, 88). After acute injury they recognize DAMPs and release hydrogen peroxide (89), recruiting neutrophils and monocytes from the blood to further amplify the inflammatory response (90). Tissue resident macrophages are particularly important for the immediate response to injury, while the long-term response is dependent on the bone marrow-derived monocytes which differentiate into macrophages in situ (54).
At least three functional subsets of macrophages contribute to the different stages of wound healing and tissue repair: (i) pro‐inflammatory (traditionally referred to as “M1”), (ii) tissue repair or pro-wound healing, and (iii) anti‐inflammatory or pro-resolving macrophages (91, 92). Subsets (ii) and (iii) are collectively referred to as “M2” macrophages. Pro-inflammatory macrophages infiltrate the injury site shortly after the wound is formed to phagocytose and kill bacteria, remove cell debris, toxic metabolites and dead cells (93). They produce inflammatory mediators, such as nitric oxide, ROS, IL-1, IL-6 and TNF-α and secrete MMP-2 and MMP-9 to break down the ECM (94, 95). Macrophage-derived cytokines IL-12/23 and IFN-γ recruit T cells and natural killer cells to amplify the proinflammatory response (16). Pro-wound healing macrophages then release elevated levels of PDGF, insulin-like growth factor 1 (IGF-1), VEGF and TGF-β1 to promote cellular growth and proliferation (96). The function of pro-resolving macrophages is to restore homeostasis, minimize fibrosis via apoptosis of myofibroblasts, and to suppress further T cell proliferation (94). In acute wounds, these macrophages are responsible for tissue repair and neovascularization (97, 98). They also suppress the inflammatory response via secretion of IL-10, arginase 1, resistin-like molecule-α (RELMα) programmed death ligand 2 (PDL2) and TGF-β1, while promoting collagen reorganization and maturation (96, 99, 100). However, macrophages activated through RELMα can also orchestrate pro-fibrotic collagen crosslinking, which is essential for the formation of granulation tissue and progression to a persistent scar (101).
Macrophages are also prominent in the TME, where tumor cells can exploit the macrophage wound repair response (102). In cancer, M1 macrophages inhibit tumor growth, while the M2 phenotype (also known as tumor‐associated macrophages or TAMs) promotes tumor progression (94). TAMs can contribute to different stages of carcinogenesis: initiation, growth, invasion, and metastasis through production of cytokines, growth factors, pro-angiogenic factors, and MMPs (103, 104). For example, the presence of TAMs correlates with increased invasion, micro-vessel density, and COX-2 expression, which are characteristic of more aggressive cancers (105). In squamous cell carcinoma, TAMs have both pro-tumor and anti-tumor activities and appear to be responsible for VEGF-C-induced lymphangiogenesis (106, 107). The macrophage chemoattractant CCL2 is expressed on melanoma cells and regulates macrophage function in melanoma in a concentration-dependent manner (108). Like neutrophils, macrophages can also play a role in preventing skin cancer as intermittent deletion of macrophages can lead to the development of basal cell carcinomas (109).
Langerhans Cells (LCs) are epidermal immune cells of embryonic origin responsible for antigen presentation and the maintenance of tolerance in the skin (110). In severe injuries, which lead to LC loss in the epidermis, cytokines such as MCP1 can facilitate the recruitment of monocytes from the bone marrow which can then differentiate into LCs in the skin (111, 112). In response to trauma, LCs extend their dendrites through epidermal tight junctions and engulf foreign antigens via dendrite tips (113). The presence of antigens can trigger LC activation and migration to the lymph nodes (114), where they can shape T cell responses (115). A subset of skin LCs has been shown to induce the proliferation of resident memory T cells with a regulatory phenotype and their ability to suppress autologous skin resident Tem cell responses (116). This study suggests that the interaction between epidermal LCs and skin resident memory T regulatory cells is important for tolerance to self-antigens and memory response (116).
Dermal DCs are composed of conventional and non-conventional (plasmacytoid) DCs that differ in ontology and functions (117). Conventional DCs are derived from myeloid progenitor cells and are responsible for presenting antigens to T cells, while plasmacytoid DCs which are derived from lymphoid progenitor cells, produce type I interferons (IFNs) in response to viral infections (117). Following skin injury, dermal DCs rapidly migrate toward the site of the injury and surround it (118). Once close to the wound site, these cells can capture cutaneous antigens and deliver them via lymphatic vessels to naive T cells in the draining lymph nodes (119).
The precise contribution of LCs and DCs to skin wound healing is still under investigation. A recent study showed that depletion of langerin+ cells (LCs and a small sub-population of dermal DCs) led to faster wound closure in mice (120). The accelerated wound repair was due to enhanced keratinocyte proliferation in the epidermis and granulation tissue formation, suggesting that langerin+ cells inhibit keratinocyte proliferation during wound healing (120). On the other hand, in another study, loss of CD11c+ cells (LCs and DCs) resulted in failure of wound closure (121). In particular, re-epithelization did not occur, and the wounds remained completely open. Since depletion of langerin+ cells removes LCs, as well as a small sub-population of langerin-expressing dermal DCs, while leaving the majority of dermal DCs unaffected, these studies suggest that dermal DCs may have a pro-reparative role, whereas LCs may hinder tissue repair. This is supported by a study showing that LCs can produce TNF which can contribute to tissue damage (122). It is worth noting that mice lacking TNF exhibit improved wound healing (123). These studies point to the important roles of LCs and DCs in wound healing, but the precise contribution of each subset may depend on the type of injury and other cells in the microenvironment.
LCs and dermal DCs are often the first immune cells to encounter antigens from cutaneous cancers (124). The effectiveness of the immune response against tumors may depend on the ability of LCs and DCs to present antigens and activate anti-tumor T cells (125). For example, in squamous cell carcinoma (SCC), there is a reduction in the number of LCs and dermal DCs, which can disrupt the generation of adaptive immunity. In SCC, DCs are poor stimulators of T cell proliferation compared to their peritumoral or healthy skin counterparts (125). In contrast, LCs harvested from SCC lesions have been found to have an increased ability to stimulate CD4+ and CD8+ T cells in vitro, compared to LCs from healthy skin (126). LCs are potent stimulators of T cell responses making them optimal targets for immunization strategies against melanoma (127), especially since the spontaneous regression of melanoma in humans is often linked to a T cell predominant infiltrate into the lesion (128). LCs may also play a role in the epithelial–mesenchymal transition (EMT) in cutaneous cancers, due to the involvement of molecules that regulate LC migration and EMT (129).
Chemokines produced in the wound including CCL3, CCL4 and CCL5 (130) attract conventional T cells to the wound site. Recruited T cells can be found in murine wounds within 24 hours of injury and persist for 30 days (131). This long timeframe suggests that they may have important roles not only during inflammation but during the proliferative and remodeling phases. For example, cytotoxic T cells release substances that kill microorganisms and clear the infection (20). T cells can also participate in the later stages of wound healing, where they exert several functions: clearance of damaged cells and debris, regulation of immune response and prevention of excessive inflammation, promotion of angiogenesis and ECM remodeling (131).
Lymphocytes differentiate into various subsets to create specialized immune responses, such as helper T cells (Th1, Th2, Th17), innate lymphoid cells (ILC1, ILC2, ILC3), and unconventional T cells (γδ T cells, iNKT cells, MAIT cells) (132). These responses can be classified by the cytokines they produce e.g., IFN−γ for type 1 immunity, IL-4, IL-5, and IL-13 for type 2, and IL-17 and IL-22 for type 3 (133). Type 2 responses play an important role in maintaining homeostasis and repairing tissue damage, and are coordinated by tissue-resident cells like ILC2s, which expand after injury (134). For instance, healthy skin of naïve C57BL/6 mice contains a population of resident ILC2s that expand after wounding (135). The importance of ILC2s has been demonstrated in mice lacking IL-33 (which contributes to the expansion of ILC2s in both humans and mice) (136). Impaired re-epithelialization in these mice is associated with diminished numbers of activated ILC2s at the site of injury (135).
In addition, mouse epidermis is enriched for γδ T cells and CD8+ resident memory T (TRM) cells. CD8 TRM cells are sessile non-circulating cells and can appear after the resolution of skin inflammation caused by infection (137). γδ T cells have a T cell receptor (TCR) composed of γ and δ subunits and demonstrate characteristics normally associated with both innate and adaptive lymphoid cells. They are abundant in mouse but not in human epidermis and play a major role in wound healing. For instance, the lack of skin γδ T cells is associated with decreased inflammation and delayed wound resolution (138).
T cells are also a crucial component of the immune system’s response to cancer. They recognize cancer cell antigens to generate an anti-tumor immune response and can control certain infections and cancers including those located in the skin. The presence of CD8+ T cells in melanomas as well as in other cancers is associated with better clinical outcomes (139, 140). However, the TME can impair CD8+ T cell ability to respond to tumor antigens as a result of activation of checkpoint proteins, such as PD-1 and CTLA-4 (141, 142). The combination of immune checkpoint inhibitors, specifically anti-CTLA-4 and anti-PD-1 antibodies, is now providing an effective therapeutic strategy in many cancers, including advanced melanoma, for which tumor regression and long-term durable cancer control is possible in nearly 50% of patients (143, 144). Multiple studies have demonstrated that NK cells can also exert significant anti-tumor effects (145, 146). In particular, they have been shown to recognize and destroy melanoma cells in vitro and in vivo (147).
The regulatory T cell subset (Tregs) plays a balancing role in inflammation by suppressing the underlying immune response. However, increased number of Tregs in sites of chronic skin inflammation did not resolve the injury, but actively delayed wound healing (148). In tumors, e.g., melanoma, Treg infiltration is a poor prognostic indicator (149, 150). Intravital analysis of Treg behavior in vivo revealed that Tregs in the TEM are migratory, in contrast to the surrounding CD8 T cells, and form unstable contacts with CD11c+ APCs. This leads to a reduction in the levels of costimulatory molecules and the activation of inhibitory receptors, such as PD-1 and TIM-3, on CD8+ T cells (151).
Impaired responses to injury result in the development of chronic wounds, which have a major impact on the quality of life (152, 153). Yet there are few treatments available once the processes leading to non-healing chronic wounds, aberrant scarring and fibrosis have begun. This makes regulation of inflammatory pathways, and especially the switches between acute and chronic inflammation, attractive targets for intervention with treatments that could be relevant to non-healing wounds. One potential new approach to achieve resolution of inflammation in non-healing wounds or cancer is to target specific inflammatory pathways (18, 30). There are a number of therapies under investigation, such as immunomodulatory agents, which may reduce inflammation and promote healing (154). For instance, animal studies have shown that cytokines, such as IL-10, which dampen inflammation, can enhance wound healing (155, 156). Furthermore, a recent study demonstrated the efficacy of IL-10 in reducing inflammation, accelerating wound healing and reducing scarring in two preclinical murine models (157). In the same study, a phase II randomized controlled trial demonstrated the translation of this therapeutic effect from animals into humans (157). Stem cells or regenerative therapies can also be used to promote tissue repair (158, 159). Several clinical trials have utilized various types of adult stem cells to improve wound healing (160, 161). Although none of these treatments have been officially approved as of yet due to major limitations such as stem cell immunogenicity and their reduced survival in vivo (162), this research highlights how understanding of the mechanisms of wound repair can lead to the development of novel therapies for large or non-healing wounds.
Since the cellular and molecular players involved in generating wound stroma can be co-opted in cancer to build tumor stroma, understanding the mechanisms of stroma generation in wounds may suggest approaches that prevent tumor stroma generation. For example, the use of anti-angiogenic therapies, which target the formation of new blood vessels, has proven successful in cancer treatment, with anti-VEGFA antibodies currently being used to treat patients with metastatic colorectal cancer (163, 164). Likewise, analysis of how inflammation is subdued once the wound is repaired may aid the development of immunotherapeutic strategies for cancer treatment.
The growth and spread of cancer cells depend on the establishment of a microenvironment, which shares a lot of commonalities with the wound healing processes. Nuanced understanding of the immune system’s role in wound repair over the whole process, including not only angiogenesis and immunosuppression, but also its potential contributions to rebuilding structural integrity of the wound and re-establishing immune networks, is essential for the development of better approaches for promoting wound healing, and the advancement of novel antitumour therapies.
TC conceived and developed the manuscript. TC and AJ contributed to writing and editing this manuscript. All authors contributed to the article and approved the submitted version.
This research was supported by funding to TC from the National Breast Cancer Foundation (IIRS-22-053), UNSW Cellular Genomics Futures Institute, UNSW Sydney, inter-disciplinary funding scheme grants and ARC Discovery Project Grant DP220102278.
We thank Prof. Anthony Basten for critical reading of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Kabashima K, Honda T, Ginhoux F, Egawa G. The immunological anatomy of the skin. Nat Rev Immunol (2019) 19(1):19–30. doi: 10.1038/s41577-018-0084-5
2. Lunjani N, Ahearn-Ford S, Dube FS, Hlela C, O'Mahony L. Mechanisms of microbe-immune system dialogue within the skin. Genes Immun (2021) 22(5-6):276–88. doi: 10.1038/s41435-021-00133-9
3. Nguyen AV, Soulika AM. The dynamics of the skin's immune system. Int J Mol Sci (2019) 20(8):1811. doi: 10.3390/ijms20081811
4. Larouche J, Sheoran S, Maruyama K, Martino MM. Immune regulation of skin wound healing: mechanisms and novel therapeutic targets. Adv Wound Care (New Rochelle) (2018) 7(7):209–31. doi: 10.1089/wound.2017.0761
5. Schafer M, Werner S. Cancer as an overhealing wound: an old hypothesis revisited. Nat Rev Mol Cell Biol (2008) 9(8):628–38. doi: 10.1038/nrm2455
6. Luyendyk JP, Schoenecker JG, Flick MJ. The multifaceted role of fibrinogen in tissue injury and inflammation. Blood (2019) 133(6):511–20. doi: 10.1182/blood-2018-07-818211
7. Eisinger F, Patzelt J, Langer HF. The platelet response to tissue injury. Front Med (Lausanne) (2018) 5:317. doi: 10.3389/fmed.2018.00317
8. Bergmeier W, Hynes RO. Extracellular matrix proteins in hemostasis and thrombosis. Cold Spring Harb Perspect Biol (2012) 4(2). doi: 10.1101/cshperspect.a005132
9. Muller WA. How endothelial cells regulate transmigration of leukocytes in the inflammatory response. Am J Pathol (2014) 184(4):886–96. doi: 10.1016/j.ajpath.2013.12.033
10. Vestweber D. How leukocytes cross the vascular endothelium. Nat Rev Immunol (2015) 15(11):692–704. doi: 10.1038/nri3908
11. Wang J. Neutrophils in tissue injury and repair. Cell Tissue Res (2018) 371(3):531–9. doi: 10.1007/s00441-017-2785-7
12. Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol (2013) 13(3):159–75. doi: 10.1038/nri3399
13. Ud-Din S, Wilgus TA, Bayat A. Mast cells in skin scarring: a review of animal and human research. Front Immunol (2020) 11:552205. doi: 10.3389/fimmu.2020.552205
14. Lunderius-Andersson C, Enoksson M, Nilsson G. Mast cells respond to cell injury through the recognition of IL-33. Front Immunol (2012) 3:82. doi: 10.3389/fimmu.2012.00082
15. Bacci S. Fine regulation during wound healing by mast cells, a physiological role not yet clarified. Int J Mol Sci (2022) 23(3). doi: 10.3390/ijms23031820
16. Brancato SK, Albina JE. Wound macrophages as key regulators of repair: origin, phenotype, and function. Am J Pathol (2011) 178(1):19–25. doi: 10.1016/j.ajpath.2010.08.003
17. Oishi Y, Manabe I. Macrophages in inflammation, repair and regeneration. Int Immunol (2018) 30(11):511–28. doi: 10.1093/intimm/dxy054
18. Landen NX, Li D, Stahle M. Transition from inflammation to proliferation: a critical step during wound healing. Cell Mol Life Sci (2016) 73(20):3861–85. doi: 10.1007/s00018-016-2268-0
19. Gonzalez AC, Costa TF, Andrade ZA, Medrado AR. Wound healing - a literature review. Bras Dermatol (2016) 91(5):614–20. doi: 10.1590/abd1806-4841.20164741
20. Canedo-Dorantes L, Canedo-Ayala M. Skin acute wound healing: a comprehensive review. Int J Inflam (2019) 2019:3706315. doi: 10.1155/2019/3706315
21. Tracy LE, Minasian RA, Caterson EJ. Extracellular matrix and dermal fibroblast function in the healing wound. Adv Wound Care (New Rochelle) (2016) 5(3):119–36. doi: 10.1089/wound.2014.0561
22. Darby IA, Laverdet B, Bonte F, Desmouliere A. Fibroblasts and myofibroblasts in wound healing. Clin Cosmet Investig Dermatol (2014) 7:301–11. doi: 10.2147/CCID.S50046
23. Hinz B, Gabbiani G. Cell-matrix and cell-cell contacts of myofibroblasts: role in connective tissue remodeling. Thromb Haemost (2003) 90(6):993–1002. doi: 10.1160/TH03-05-0328
24. Broughton G 2nd, Janis JE, Attinger CE. Wound healing: an overview. Plast Reconstr Surg (2006) 117(7 Suppl):1e–S-32e-S. doi: 10.1097/01.prs.0000222562.60260.f9
25. Xue M, Jackson CJ. Extracellular matrix reorganization during wound healing and its impact on abnormal scarring. Adv Wound Care (New Rochelle) (2015) 4(3):119–36. doi: 10.1089/wound.2013.0485
26. Gurtner GC, Werner S, Barrandon Y, Longaker MT. Wound repair and regeneration. Nature (2008) 453(7193):314–21. doi: 10.1038/nature07039
27. Edmonds M. Body of knowledge around the diabetic foot and limb salvage. J Cardiovasc Surg (Torino) (2012) 53(5):605–16.
28. Nunan R, Harding KG, Martin P. Clinical challenges of chronic wounds: searching for an optimal animal model to recapitulate their complexity. Dis Model Mech (2014) 7(11):1205–13. doi: 10.1242/dmm.016782
29. Ju T, Hernandez L, Mohsin N, Labib A, Frech F, Nouri K. Evaluation of risk in chronic cutaneous inflammatory conditions for malignant transformation. J Eur Acad Dermatol Venereol (2023) 37(2):231–42. doi: 10.1111/jdv.18663
30. Coussens LM, Werb Z. Inflammation and cancer. Nature (2002) 420(6917):860–7. doi: 10.1038/nature01322
31. Morhenn VB, Nelson TE, Gruol DL. The rate of wound healing is increased in psoriasis. J Dermatol Sci (2013) 72(2):87–92. doi: 10.1016/j.jdermsci.2013.06.001
32. Schuster R, Rockel JS, Kapoor M, Hinz B. The inflammatory speech of fibroblasts. Immunol Rev (2021) 302(1):126–46. doi: 10.1111/imr.12971
33. Wu S, Rhee KJ, Albesiano E, Rabizadeh S, Wu X, Yen HR, et al. A human colonic commensal promotes colon tumorigenesis via activation of T helper type 17 T cell responses. Nat Med (2009) 15(9):1016–22. doi: 10.1038/nm.2015
34. Huber S, Gagliani N, Zenewicz LA, Huber FJ, Bosurgi L, Hu B, et al. IL-22BP is regulated by the inflammasome and modulates tumorigenesis in the intestine. Nature (2012) 491(7423):259–63. doi: 10.1038/nature11535
35. Dvorak HF. Tumors: wounds that do not heal. similarities between tumor stroma generation and wound healing. N Engl J Med (1986) 315(26):1650–9. doi: 10.1056/NEJM198612253152606
36. Dvorak HF. Tumors: wounds that do not heal-redux. Cancer Immunol Res (2015) 3(1):1–11. doi: 10.1158/2326-6066.CIR-14-0209
37. Zhao H, Wu L, Yan G, Chen Y, Zhou M, Wu Y, et al. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal Transduct Target Ther (2021) 6(1):263. doi: 10.1038/s41392-021-00658-5
38. Sundaram GM, Quah S, Sampath P. Cancer: the dark side of wound healing. FEBS J (2018) 285(24):4516–34. doi: 10.1111/febs.14586
39. Rungelrath V, Kobayashi SD, DeLeo FR. Neutrophils in innate immunity and systems biology-level approaches. Wiley Interdiscip Rev Syst Biol Med (2020) 12(1):e1458. doi: 10.1002/wsbm.1458
40. Kovtun A, Messerer DAC, Scharffetter-Kochanek K, Huber-Lang M, Ignatius A. Neutrophils in tissue trauma of the skin, bone, and lung: two sides of the same coin. J Immunol Res (2018) 2018:8173983. doi: 10.1155/2018/8173983
41. Zhang Q, Raoof M, Chen Y, Sumi Y, Sursal T, Junger W, et al. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature (2010) 464(7285):104–7. doi: 10.1038/nature08780
42. Rosales C. Neutrophil: a cell with many roles in inflammation or several cell types? Front Physiol (2018) 9:113. doi: 10.3389/fphys.2018.00113
43. Phillipson M, Kubes P. The healing power of neutrophils. Trends Immunol (2019) 40(7):635–47. doi: 10.1016/j.it.2019.05.001
44. Yoshimura T, Galligan C, Takahashi M, Chen K, Liu M, Tessarollo L, et al. Non-myeloid cells are major contributors to innate immune responses via production of monocyte chemoattractant protein-1/CCL2. Front Immunol (2014) 4:482. doi: 10.3389/fimmu.2013.00482
45. Hampton HR, Bailey J, Tomura M, Brink R, Chtanova T. Microbe-dependent lymphatic migration of neutrophils modulates lymphocyte proliferation in lymph nodes. Nat Commun (2015) 6:7139. doi: 10.1038/ncomms8139
46. Hampton HR, Chtanova T. The lymph node neutrophil. Semin Immunol (2016) 28(2):129–36. doi: 10.1016/j.smim.2016.03.008
47. Yam AO, Chtanova T. Imaging the neutrophil: intravital microscopy provides a dynamic view of neutrophil functions in host immunity. Cell Immunol (2020) 350:103898. doi: 10.1016/j.cellimm.2019.01.003
48. Lammermann T, Afonso PV, Angermann BR, Wang JM, Kastenmuller W, Parent CA, et al. Neutrophil swarms require LTB4 and integrins at sites of cell death in vivo. Nature (2013) 498(7454):371–5. doi: 10.1038/nature12175
49. Chtanova T, Schaeffer M, Han SJ, van Dooren GG, Nollmann M, Herzmark P, et al. Dynamics of neutrophil migration in lymph nodes during infection. Immunity (2008) 29(3):487–96. doi: 10.1016/j.immuni.2008.07.012
50. De Filippo K, Rankin SM. The secretive life of neutrophils revealed by intravital microscopy. Front Cell Dev Biol (2020) 8:603230. doi: 10.3389/fcell.2020.603230
51. Jakovija A, Chtanova T. Neutrophil interactions with the lymphatic system. Cells (2021) 10(8):2106. doi: 10.3390/cells10082106
52. Niethammer P, Grabher C, Look AT, Mitchison TJ. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature (2009) 459(7249):996–9. doi: 10.1038/nature08119
53. Ellis S, Lin EJ, Tartar D. Immunology of wound healing. Curr Dermatol Rep (2018) 7(4):350–8. doi: 10.1007/s13671-018-0234-9
54. Raziyeva K, Kim Y, Zharkinbekov Z, Kassymbek K, Jimi S, Saparov A. Immunology of acute and chronic wound healing. Biomolecules (2021) 11(5):700. doi: 10.3390/biom11050700
55. Natrus L, Ryzhko I, Blazquez-Navarro A, Panova T, Zaychenko G, Klymenko O, et al. Correlational analysis of the regulatory interplay between molecules and cellular components mediating angiogenesis in wound healing under normal and hyperglycemic conditions. Clin Hemorheol Microcirc (2021) 78(4):379–90. doi: 10.3233/CH-201077
56. Theilgaard-Monch K, Knudsen S, Follin P, Borregaard N. The transcriptional activation program of human neutrophils in skin lesions supports their important role in wound healing. J Immunol (2004) 172(12):7684–93. doi: 10.4049/jimmunol.172.12.7684
57. Gong Y, Koh DR. Neutrophils promote inflammatory angiogenesis via release of preformed VEGF in an in vivo corneal model. Cell Tissue Res (2010) 339(2):437–48. doi: 10.1007/s00441-009-0908-5
58. Liu M, Chen K, Yoshimura T, Liu Y, Gong W, Le Y, et al. Formylpeptide receptors mediate rapid neutrophil mobilization to accelerate wound healing. PloS One (2014) 9(6):e90613. doi: 10.1371/journal.pone.0090613
59. Devalaraja RM, Nanney LB, Du J, Qian Q, Yu Y, Devalaraja MN, et al. Delayed wound healing in CXCR2 knockout mice. J Invest Dermatol (2000) 115(2):234–44. doi: 10.1046/j.1523-1747.2000.00034.x
60. Wilgus TA, Roy S, McDaniel JC. Neutrophils and wound repair: positive actions and negative reactions. Adv Wound Care (New Rochelle) (2013) 2(7):379–88. doi: 10.1089/wound.2012.0383
61. Gutierrez-Fernandez A, Inada M, Balbin M, Fueyo A, Pitiot AS, Astudillo A, et al. Increased inflammation delays wound healing in mice deficient in collagenase-2 (MMP-8). FASEB J (2007) 21(10):2580–91. doi: 10.1096/fj.06-7860com
62. Ng LG. Neutrophils guide pre-existing matrix into injured organs to initiate tissue repair. Nat Immunol (2022) 23(4):472–3. doi: 10.1038/s41590-022-01173-7
63. Singel KL, Segal BH. Neutrophils in the tumor microenvironment: trying to heal the wound that cannot heal. Immunol Rev (2016) 273(1):329–43. doi: 10.1111/imr.12459
64. Jamieson T, Clarke M, Steele CW, Samuel MS, Neumann J, Jung A, et al. Inhibition of CXCR2 profoundly suppresses inflammation-driven and spontaneous tumorigenesis. J Clin Invest (2012) 122(9):3127–44. doi: 10.1172/JCI61067
65. Katoh H, Wang D, Daikoku T, Sun H, Dey SK, Dubois RN. CXCR2-expressing myeloid-derived suppressor cells are essential to promote colitis-associated tumorigenesis. Cancer Cell (2013) 24(5):631–44. doi: 10.1016/j.ccr.2013.10.009
66. Sody S, Uddin M, Gruneboom A, Gorgens A, Giebel B, Gunzer M, et al. Distinct spatio-temporal dynamics of tumor-associated neutrophils in small tumor lesions. Front Immunol (2019) 10:1419. doi: 10.3389/fimmu.2019.01419
67. Nolan E, Malanchi I. Connecting the dots: neutrophils at the interface of tissue regeneration and cancer. Semin Immunol (2021) 57:101598. doi: 10.1016/j.smim.2022.101598
68. Kusmartsev S, Nefedova Y, Yoder D, Gabrilovich DI. Antigen-specific inhibition of CD8+ T cell response by immature myeloid cells in cancer is mediated by reactive oxygen species. J Immunol (2004) 172(2):989–99. doi: 10.4049/jimmunol.172.2.989
69. Li L, Chen K, Xiang Y, Yoshimura T, Su S, Zhu J, et al. New development in studies of formyl-peptide receptors: critical roles in host defense. J Leukoc Biol (2016) 99(3):425–35. doi: 10.1189/jlb.2RI0815-354RR
70. Rice CM, Davies LC, Subleski JJ, Maio N, Gonzalez-Cotto M, Andrews C, et al. Tumour-elicited neutrophils engage mitochondrial metabolism to circumvent nutrient limitations and maintain immune suppression. Nat Commun (2018) 9(1):5099. doi: 10.1038/s41467-018-07505-2
71. Reuter S, Gupta SC, Chaturvedi MM, Aggarwal BB. Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med (2010) 49(11):1603–16. doi: 10.1016/j.freeradbiomed.2010.09.006
72. Maru GB, Gandhi K, Ramchandani A, Kumar G. The role of inflammation in skin cancer. Adv Exp Med Biol (2014) 816:437–69. doi: 10.1007/978-3-0348-0837-8_17
73. Taftaf R, Liu X, Singh S, Jia Y, Dashzeveg NK, Hoffmann AD, et al. ICAM1 initiates CTC cluster formation and trans-endothelial migration in lung metastasis of breast cancer. Nat Commun (2021) 12(1):4867. doi: 10.1038/s41467-021-25189-z
74. Headley MB, Bins A, Nip A, Roberts EW, Looney MR, Gerard A, et al. Visualization of immediate immune responses to pioneer metastatic cells in the lung. Nature (2016) 531(7595):513–7. doi: 10.1038/nature16985
75. Inoue M, Nakashima R, Enomoto M, Koike Y, Zhao X, Yip K, et al. Plasma redox imbalance caused by albumin oxidation promotes lung-predominant NETosis and pulmonary cancer metastasis. Nat Commun (2018) 9(1):5116. doi: 10.1038/s41467-018-07550-x
76. Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell (2010) 141(1):52–67. doi: 10.1016/j.cell.2010.03.015
77. Monti M, De Rosa V, Iommelli F, Carriero MV, Terlizzi C, Camerlingo R, et al. Neutrophil extracellular traps as an adhesion substrate for different tumor cells expressing RGD-binding integrins. Int J Mol Sci (2018) 19(8):2350. doi: 10.3390/ijms19082350
78. Bekes EM, Schweighofer B, Kupriyanova TA, Zajac E, Ardi VC, Quigley JP, et al. Tumor-recruited neutrophils and neutrophil TIMP-free MMP-9 regulate coordinately the levels of tumor angiogenesis and efficiency of malignant cell intravasation. Am J Pathol (2011) 179(3):1455–70. doi: 10.1016/j.ajpath.2011.05.031
79. Barillari G. The impact of matrix metalloproteinase-9 on the sequential steps of the metastatic process. Int J Mol Sci (2020) 21(12):4526. doi: 10.3390/ijms21124526
80. Stanciu AE, Zamfir-Chiru-Anton A, Stanciu MM, Popescu CR, Gheorghe DC. Imbalance between matrix metalloproteinases and tissue inhibitors of metalloproteinases promotes invasion and metastasis of head and neck squamous cell carcinoma. Clin Lab (2017) 63(10):1613–20. doi: 10.7754/Clin.Lab.2017.170339
81. Queen MM, Ryan RE, Holzer RG, Keller-Peck CR, Jorcyk CL. Breast cancer cells stimulate neutrophils to produce oncostatin m: potential implications for tumor progression. Cancer Res (2005) 65(19):8896–904. doi: 10.1158/0008-5472.CAN-05-1734
82. Furumaya C, Martinez-Sanz P, Bouti P, Kuijpers TW, Matlung HL. Plasticity in pro- and anti-tumor activity of neutrophils: shifting the balance. Front Immunol (2020) 11:2100. doi: 10.3389/fimmu.2020.02100
83. Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L, et al. Polarization of tumor-associated neutrophil phenotype by TGF-beta: "N1" versus "N2" TAN. Cancer Cell (2009) 16(3):183–94. doi: 10.1016/j.ccr.2009.06.017
84. Linde IL, Prestwood TR, Qiu J, Pilarowski G, Linde MH, Zhang X, et al. Neutrophil-activating therapy for the treatment of cancer. Cancer Cell (2023) 41(2):356–72 e10. doi: 10.1016/j.ccell.2023.01.002
85. Yam AO, Bailey J, Lin F, Jakovija A, Youlten SE, Counoupas C, et al. Neutrophil conversion to a tumor-killing phenotype underpins effective microbial therapy. Cancer Res (2023) 83(8):1315–28. doi: 10.1158/0008-5472.c.6599657
86. Gautier EL, Shay T, Miller J, Greter M, Jakubzick C, Ivanov S, et al. Gene-expression profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse tissue macrophages. Nat Immunol (2012) 13(11):1118–28. doi: 10.1038/ni.2419
87. Okabe Y, Medzhitov R. Tissue-specific signals control reversible program of localization and functional polarization of macrophages. Cell (2014) 157(4):832–44. doi: 10.1016/j.cell.2014.04.016
88. Okabe Y, Medzhitov R. Tissue biology perspective on macrophages. Nat Immunol (2016) 17(1):9–17. doi: 10.1038/ni.3320
89. Minutti CM, Knipper JA, Allen JE, Zaiss DM. Tissue-specific contribution of macrophages to wound healing. Semin Cell Dev Biol (2017) 61:3–11. doi: 10.1016/j.semcdb.2016.08.006
90. Peiseler M, Kubes P. More friend than foe: the emerging role of neutrophils in tissue repair. J Clin Invest (2019) 129(7):2629–39. doi: 10.1172/JCI124616
91. Wynn TA, Vannella KM. Macrophages in tissue repair, regeneration, and fibrosis. Immunity (2016) 44(3):450–62. doi: 10.1016/j.immuni.2016.02.015
92. Lucas T, Waisman A, Ranjan R, Roes J, Krieg T, Muller W, et al. Differential roles of macrophages in diverse phases of skin repair. J Immunol (2010) 184(7):3964–77. doi: 10.4049/jimmunol.0903356
93. Hirayama D, Iida T, Nakase H. The phagocytic function of macrophage-enforcing innate immunity and tissue homeostasis. Int J Mol Sci (2017) 19(1):92. doi: 10.3390/ijms19010092
94. Murray PJ, Wynn TA. Protective and pathogenic functions of macrophage subsets. Nat Rev Immunol (2011) 11(11):723–37. doi: 10.1038/nri3073
95. Madsen DH, Leonard D, Masedunskas A, Moyer A, Jurgensen HJ, Peters DE, et al. M2-like macrophages are responsible for collagen degradation through a mannose receptor-mediated pathway. J Cell Biol (2013) 202(6):951–66. doi: 10.1083/jcb.201301081
96. Krzyszczyk P, Schloss R, Palmer A, Berthiaume F. The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front Physiol (2018) 9:419. doi: 10.3389/fphys.2018.00419
97. Willenborg S, Lucas T, van Loo G, Knipper JA, Krieg T, Haase I, et al. CCR2 recruits an inflammatory macrophage subpopulation critical for angiogenesis in tissue repair. Blood (2012) 120(3):613–25. doi: 10.1182/blood-2012-01-403386
98. Mirza R, DiPietro LA, Koh TJ. Selective and specific macrophage ablation is detrimental to wound healing in mice. Am J Pathol (2009) 175(6):2454–62. doi: 10.2353/ajpath.2009.090248
99. Herold S, Mayer K, Lohmeyer J. Acute lung injury: how macrophages orchestrate resolution of inflammation and tissue repair. Front Immunol (2011) 2:65. doi: 10.3389/fimmu.2011.00065
100. Zhao Y, Xiong Z, Lechner EJ, Klenotic PA, Hamburg BJ, Hulver M, et al. Thrombospondin-1 triggers macrophage IL-10 production and promotes resolution of experimental lung injury. Mucosal Immunol (2014) 7(2):440–8. doi: 10.1038/mi.2013.63
101. Knipper JA, Willenborg S, Brinckmann J, Bloch W, Maass T, Wagener R, et al. Interleukin-4 receptor alpha signaling in myeloid cells controls collagen fibril assembly in skin repair. Immunity (2015) 43(4):803–16. doi: 10.1016/j.immuni.2015.09.005
102. Muliaditan T, Caron J, Okesola M, Opzoomer JW, Kosti P, Georgouli M, et al. Macrophages are exploited from an innate wound healing response to facilitate cancer metastasis. Nat Commun (2018) 9(1):2951. doi: 10.1038/s41467-018-05346-7
103. Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol (2005) 5(10):749–59. doi: 10.1038/nri1703
104. Qian BZ, Pollard JW. Macrophage diversity enhances tumor progression and metastasis. Cell (2010) 141(1):39–51. doi: 10.1016/j.cell.2010.03.014
105. Tjiu JW, Chen JS, Shun CT, Lin SJ, Liao YH, Chu CY, et al. Tumor-associated macrophage-induced invasion and angiogenesis of human basal cell carcinoma cells by cyclooxygenase-2 induction. J Invest Dermatol (2009) 129(4):1016–25. doi: 10.1038/jid.2008.310
106. Moussai D, Mitsui H, Pettersen JS, Pierson KC, Shah KR, Suarez-Farinas M, et al. The human cutaneous squamous cell carcinoma microenvironment is characterized by increased lymphatic density and enhanced expression of macrophage-derived VEGF-c. J Invest Dermatol (2011) 131(1):229–36. doi: 10.1038/jid.2010.266
107. Pettersen JS, Fuentes-Duculan J, Suarez-Farinas M, Pierson KC, Pitts-Kiefer A, Fan L, et al. Tumor-associated macrophages in the cutaneous SCC microenvironment are heterogeneously activated. J Invest Dermatol (2011) 131(6):1322–30. doi: 10.1038/jid.2011.9
108. Nesbit M, Schaider H, Miller TH, Herlyn M. Low-level monocyte chemoattractant protein-1 stimulation of monocytes leads to tumor formation in nontumorigenic melanoma cells. J Immunol (2001) 166(11):6483–90. doi: 10.4049/jimmunol.166.11.6483
109. Afaq F, Ahmad N, Mukhtar H. Suppression of UVB-induced phosphorylation of mitogen-activated protein kinases and nuclear factor kappa b by green tea polyphenol in SKH-1 hairless mice. Oncogene (2003) 22(58):9254–64. doi: 10.1038/sj.onc.1207035
110. Romani N, Clausen BE, Stoitzner P. Langerhans cells and more: langerin-expressing dendritic cell subsets in the skin. Immunol Rev (2010) 234(1):120–41. doi: 10.1111/j.0105-2896.2009.00886.x
111. Chopin M, Nutt SL. Establishing and maintaining the langerhans cell network. Semin Cell Dev Biol (2015) 41:23–9. doi: 10.1016/j.semcdb.2014.02.001
112. Ginhoux F, Merad M. Ontogeny and homeostasis of langerhans cells. Immunol Cell Biol (2010) 88(4):387–92. doi: 10.1038/icb.2010.38
113. Kubo A, Nagao K, Yokouchi M, Sasaki H, Amagai M. External antigen uptake by langerhans cells with reorganization of epidermal tight junction barriers. J Exp Med (2009) 206(13):2937–46. doi: 10.1084/jem.20091527
114. West HC, Bennett CL. Redefining the role of langerhans cells as immune regulators within the skin. Front Immunol (2017) 8:1941. doi: 10.3389/fimmu.2017.01941
115. Kitashima DY, Kobayashi T, Woodring T, Idouchi K, Doebel T, Voisin B, et al. Langerhans cells prevent autoimmunity via expansion of keratinocyte antigen-specific regulatory T cells. EBioMedicine (2018) 27:293–303. doi: 10.1016/j.ebiom.2017.12.022
116. Seneschal J, Clark RA, Gehad A, Baecher-Allan CM, Kupper TS. Human epidermal langerhans cells maintain immune homeostasis in skin by activating skin resident regulatory T cells. Immunity (2012) 36(5):873–84. doi: 10.1016/j.immuni.2012.03.018
117. Balan S, Saxena M, Bhardwaj N. Dendritic cell subsets and locations. Int Rev Cell Mol Biol (2019) 348:1–68. doi: 10.1016/bs.ircmb.2019.07.004
118. Goh CC, Li JL, Devi S, Bakocevic N, See P, Larbi A, et al. Real-time imaging of dendritic cell responses to sterile tissue injury. J Invest Dermatol (2015) 135(4):1181–4. doi: 10.1038/jid.2014.506
119. McLachlan JB, Catron DM, Moon JJ, Jenkins MK. Dendritic cell antigen presentation drives simultaneous cytokine production by effector and regulatory T cells in inflamed skin. Immunity (2009) 30(2):277–88. doi: 10.1016/j.immuni.2008.11.013
120. Rajesh A, Stuart G, Real N, Ahn J, Tschirley A, Wise L, et al. Depletion of langerin(+) cells enhances cutaneous wound healing. Immunology (2020) 160(4):366–81. doi: 10.1111/imm.13202
121. Li Z, Lamb R, Coles MC, Bennett CL, Ambler CA. Inducible ablation of CD11c(+) cells to determine their role in skin wound repair. Immunology (2021) 163(1):105–11. doi: 10.1111/imm.13312
122. Anderton H, Chopin M, Dawson CA, Nutt SL, Whitehead L, Silke N, et al. Langerhans cells are an essential cellular intermediary in chronic dermatitis. Cell Rep (2022) 39(10):110922. doi: 10.1016/j.celrep.2022.110922
123. Mori R, Kondo T, Ohshima T, Ishida Y, Mukaida N. Accelerated wound healing in tumor necrosis factor receptor p55-deficient mice with reduced leukocyte infiltration. FASEB J (2002) 16(9):963–74. doi: 10.1096/fj.01-0776com
124. Yanofsky VR, Mitsui H, Felsen D, Carucci JA. Understanding dendritic cells and their role in cutaneous carcinoma and cancer immunotherapy. Clin Dev Immunol (2013) 2013:624123. doi: 10.1155/2013/624123
125. Bluth MJ, Zaba LC, Moussai D, Suarez-Farinas M, Kaporis H, Fan L, et al. Myeloid dendritic cells from human cutaneous squamous cell carcinoma are poor stimulators of T-cell proliferation. J Invest Dermatol (2009) 129(10):2451–62. doi: 10.1038/jid.2009.96
126. Adams JL, Smothers J, Srinivasan R, Hoos A. Big opportunities for small molecules in immuno-oncology. Nat Rev Drug Discov (2015) 14(9):603–22. doi: 10.1038/nrd4596
127. Stoitzner P, Sparber F, Tripp CH. Langerhans cells as targets for immunotherapy against skin cancer. Immunol Cell Biol (2010) 88(4):431–7. doi: 10.1038/icb.2010.31
128. McGovern VJ. Spontaneous regression of melanoma. Pathology (1975) 7(2):91–9. doi: 10.3109/00313027509092702
129. Hieronymus T, Zenke M, Baek JH, Sere K. The clash of langerhans cell homeostasis in skin: should I stay or should I go? Semin Cell Dev Biol (2015) 41:30–8. doi: 10.1016/j.semcdb.2014.02.009
130. Balaji S, Watson CL, Ranjan R, King A, Bollyky PL, Keswani SG. Chemokine involvement in fetal and adult wound healing. Adv Wound Care (New Rochelle) (2015) 4(11):660–72. doi: 10.1089/wound.2014.0564
131. Wang X, Balaji S, Steen EH, Li H, Rae MM, Blum AJ, et al. T Lymphocytes attenuate dermal scarring by regulating inflammation, neovascularization, and extracellular matrix remodeling. Adv Wound Care (New Rochelle) (2019) 8(11):527–37. doi: 10.1089/wound.2019.0981
132. Eberl G. Immunity by equilibrium. Nat Rev Immunol (2016) 16(8):524–32. doi: 10.1038/nri.2016.75
133. Boothby IC, Cohen JN, Rosenblum MD. Regulatory T cells in skin injury: At the crossroads of tolerance and tissue repair. Sci Immunol (2020) 5(47):eaaz9631. doi: 10.1126/sciimmunol.aaz9631
134. Allen JE, Wynn TA. Evolution of Th2 immunity: a rapid repair response to tissue destructive pathogens. PloS Pathog (2011) 7(5):e1002003. doi: 10.1371/journal.ppat.1002003
135. Rak GD, Osborne LC, Siracusa MC, Kim BS, Wang K, Bayat A, et al. IL-33-Dependent group 2 innate lymphoid cells promote cutaneous wound healing. J Invest Dermatol (2016) 136(2):487–96. doi: 10.1038/JID.2015.406
136. Miller JE, Lingegowda H, Symons LK, Bougie O, Young SL, Lessey BA, et al. IL-33 activates group 2 innate lymphoid cell expansion and modulates endometriosis. JCI Insight (2021) 6(23):e149699. doi: 10.1172/jci.insight.149699
137. Jiang X, Clark RA, Liu L, Wagers AJ, Fuhlbrigge RC, Kupper TS. Skin infection generates non-migratory memory CD8+ T(RM) cells providing global skin immunity. Nature (2012) 483(7388):227–31. doi: 10.1038/nature10851
138. Munoz LD, Sweeney MJ, Jameson JM. Skin resident gammadelta T cell function and regulation in wound repair. Int J Mol Sci (2020) 21(23):9286. doi: 10.3390/ijms21239286
139. Ladanyi A, Somlai B, Gilde K, Fejos Z, Gaudi I, Timar J. T-Cell activation marker expression on tumor-infiltrating lymphocytes as prognostic factor in cutaneous malignant melanoma. Clin Cancer Res (2004) 10(2):521–30. doi: 10.1158/1078-0432.CCR-1161-03
140. Piersma SJ, Jordanova ES, van Poelgeest MI, Kwappenberg KM, van der Hulst JM, Drijfhout JW, et al. High number of intraepithelial CD8+ tumor-infiltrating lymphocytes is associated with the absence of lymph node metastases in patients with large early-stage cervical cancer. Cancer Res (2007) 67(1):354–61. doi: 10.1158/0008-5472.CAN-06-3388
141. Ahmadzadeh M, Johnson LA, Heemskerk B, Wunderlich JR, Dudley ME, White DE, et al. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood (2009) 114(8):1537–44. doi: 10.1182/blood-2008-12-195792
142. He X, Xu C. Immune checkpoint signaling and cancer immunotherapy. Cell Res (2020) 30(8):660–9. doi: 10.1038/s41422-020-0343-4
143. Carlino MS, Larkin J, Long GV. Immune checkpoint inhibitors in melanoma. Lancet (2021) 398(10304):1002–14. doi: 10.1016/S0140-6736(21)01206-X
144. Wei SC, Anang NAS, Sharma R, Andrews MC, Reuben A, Levine JH, et al. Combination anti-CTLA-4 plus anti-PD-1 checkpoint blockade utilizes cellular mechanisms partially distinct from monotherapies. Proc Natl Acad Sci USA (2019) 116(45):22699–709. doi: 10.1073/pnas.1821218116
145. Chiossone L, Dumas PY, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol (2018) 18(11):671–88. doi: 10.1038/s41577-018-0061-z
146. Lopez-Soto A, Gonzalez S, Smyth MJ, Galluzzi L. Control of metastasis by NK cells. Cancer Cell (2017) 32(2):135–54. doi: 10.1016/j.ccell.2017.06.009
147. Lakshmikanth T, Burke S, Ali TH, Kimpfler S, Ursini F, Ruggeri L, et al. NCRs and DNAM-1 mediate NK cell recognition and lysis of human and mouse melanoma cell lines in vitro and in vivo. J Clin Invest (2009) 119(5):1251–63. doi: 10.1172/JCI36022
148. Ameri AH, Moradi Tuchayi S, Zaalberg A, Park JH, Ngo KH, Li T, et al. IL-33/regulatory T cell axis triggers the development of a tumor-promoting immune environment in chronic inflammation. Proc Natl Acad Sci USA (2019) 116(7):2646–51. doi: 10.1073/pnas.1815016116
149. Huang L, Guo Y, Liu S, Wang H, Zhu J, Ou L, et al. Targeting regulatory T cells for immunotherapy in melanoma. Mol Biomed (2021) 2(1):11. doi: 10.1186/s43556-021-00038-z
150. Whiteside TL. FOXP3+ treg as a therapeutic target for promoting anti-tumor immunity. Expert Opin Ther Targets (2018) 22(4):353–63. doi: 10.1080/14728222.2018.1451514
151. Bauer CA, Kim EY, Marangoni F, Carrizosa E, Claudio NM, Mempel TR. Dynamic treg interactions with intratumoral APCs promote local CTL dysfunction. J Clin Invest (2014) 124(6):2425–40. doi: 10.1172/JCI66375
152. Zhu X, Olsson MM, Bajpai R, Jarbrink K, Tang WE, Car J. Health-related quality of life and chronic wound characteristics among patients with chronic wounds treated in primary care: a cross-sectional study in Singapore. Int Wound J (2022) 19(5):1121–32. doi: 10.1111/iwj.13708
153. Frykberg RG, Banks J. Challenges in the treatment of chronic wounds. Adv Wound Care (New Rochelle) (2015) 4(9):560–82. doi: 10.1089/wound.2015.0635
154. Petkovic M, Mouritzen MV, Mojsoska B, Jenssen H. Immunomodulatory properties of host defence peptides in skin wound healing. Biomolecules (2021) 11(7):952. doi: 10.3390/biom11070952
155. King A, Balaji S, Le LD, Crombleholme TM, Keswani SG. Regenerative wound healing: the role of interleukin-10. Adv Wound Care (New Rochelle) (2014) 3(4):315–23. doi: 10.1089/wound.2013.0461
156. Gordon A, Kozin ED, Keswani SG, Vaikunth SS, Katz AB, Zoltick PW, et al. Permissive environment in postnatal wounds induced by adenoviral-mediated overexpression of the anti-inflammatory cytokine interleukin-10 prevents scar formation. Wound Repair Regen (2008) 16(1):70–9. doi: 10.1111/j.1524-475X.2007.00326.x
157. Kieran I, Knock A, Bush J, So K, Metcalfe A, Hobson R, et al. Interleukin-10 reduces scar formation in both animal and human cutaneous wounds: results of two preclinical and phase II randomized control studies. Wound Repair Regen (2013) 21(3):428–36. doi: 10.1111/wrr.12043
158. Rinkevich Y, Lindau P, Ueno H, Longaker MT, Weissman IL. Germ-layer and lineage-restricted stem/progenitors regenerate the mouse digit tip. Nature (2011) 476(7361):409–13. doi: 10.1038/nature10346
159. Wong VW, Sorkin M, Gurtner GC. Enabling stem cell therapies for tissue repair: current and future challenges. Biotechnol Adv (2013) 31(5):744–51. doi: 10.1016/j.biotechadv.2012.11.006
160. Kirby GT, Mills SJ, Cowin AJ, Smith LE. Stem cells for cutaneous wound healing. BioMed Res Int (2015) 2015:285869. doi: 10.1155/2015/285869
161. Rosemann A. Why regenerative stem cell medicine progresses slower than expected. J Cell Biochem (2014) 115(12):2073–6. doi: 10.1002/jcb.24894
162. Dash BC, Xu Z, Lin L, Koo A, Ndon S, Berthiaume F, et al. Stem cells and engineered scaffolds for regenerative wound healing. Bioengineering (Basel) (2018) 5(1):23. doi: 10.3390/bioengineering5010023
163. Xu M, Zhang T, Xia R, Wei Y, Wei X. Targeting the tumor stroma for cancer therapy. Mol Cancer (2022) 21(1):208. doi: 10.1186/s12943-022-01670-1
Keywords: skin wound healing, skin immunity, innate response, skin cancer immunity, skin adaptive immunity
Citation: Jakovija A and Chtanova T (2023) Skin immunity in wound healing and cancer. Front. Immunol. 14:1060258. doi: 10.3389/fimmu.2023.1060258
Received: 03 October 2022; Accepted: 24 May 2023;
Published: 16 June 2023.
Edited by:
Snehlata Kumari, The University of Queensland, AustraliaReviewed by:
Sebastian Willenborg, University Hospital of Cologne, GermanyCopyright © 2023 Jakovija and Chtanova. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tatyana Chtanova, dC5jaHRhbm92YUB1bnN3LmVkdS5hdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.