
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 18 August 2022
Sec. Cytokines and Soluble Mediators in Immunity
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.979749
This article is part of the Research Topic Gasdermins in the Defense Against Pathogens View all 8 articles
 Laura D. Manzanares-Meza1,2
Laura D. Manzanares-Meza1,2 Claudia I. Gutiérrez-Román1
Claudia I. Gutiérrez-Román1 Albertana Jiménez-Pineda1
Albertana Jiménez-Pineda1 Felipe Castro-Martínez1
Felipe Castro-Martínez1 Genaro Patiño-López3
Genaro Patiño-López3 Eunice Rodríguez-Arellano4
Eunice Rodríguez-Arellano4 Ricardo Valle-Rios3,5
Ricardo Valle-Rios3,5 Vianney F. Ortíz-Navarrete2
Vianney F. Ortíz-Navarrete2 Oscar Medina-Contreras1*
Oscar Medina-Contreras1*Mucosal innate immunity functions as the first line of defense against invading pathogens. Members of the IL-1 family are key cytokines upregulated in the inflamed mucosa. Inflammatory cytokines are regulated by limiting their function and availability through their activation and secretion mechanisms. IL-1 cytokines secretion is affected by the lack of a signal peptide on their sequence, which prevents them from accessing the conventional protein secretion pathway; thus, they use unconventional protein secretion pathways. Here we show in mouse macrophages that LPS/ATP stimulation induces cytokine relocalization to the plasma membrane, and conventional secretion blockade using monensin or Brefeldin A triggers no IL-36γ accumulation within the cell. In silico modeling indicates IL-36γ can pass through both the P2X7R and Gasdermin D pores, and both IL-36γ, P2X7R and Gasdermin D mRNA are upregulated in inflammation; further, experimental blockade of these receptors’ limits IL-36γ release. Our results demonstrate that IL-36γ is secreted mainly by an unconventional pathway through membrane pores formed by P2X7R and Gasdermin D.
Mucosal innate immunity functions as the first line of defense against invading pathogens. A key cellular component of these innate immune responses are macrophages (1). During infection and the subsequent inflammation, macrophages recognize Pathogen Associated Molecular Patterns (PAPs) and initiate downstream signaling that promotes the expression and secretion of inflammatory cytokines (2). Several members of the IL-1 family (IL-1α, IL-1β, IL-18, IL-33) are key cytokines upregulated in the inflamed mucosa (3).
This superfamily of cytokines is formed by eleven members, classified into three subfamilies: IL-1 subfamily, IL-18 subfamily and, IL-36 subfamily (4). The IL-36 subfamily has been recently described and involved in the development of several inflammatory diseases such as inflammatory bowel disease, rheumatoid arthritis, or psoriasis (5, 6).
The IL-36 subfamily consists of four members, a receptor antagonist (IL-36Ra), and three agonists, IL-36α, IL-36β and, IL-36γ (5). These agonists signal through the same receptor (IL-36R) (7) in a MyD88-dependent manner and activate NFκB and MAPK pathways; thus, leading to leukocyte recruitment and amplification of inflammation (8).
Inflammatory cytokines are regulated through their activation and secretion mechanisms to limit their function and availability (9), and cytokine processing is the first regulatory checkpoint (10). IL-1 cytokine superfamily requires processing to achieve full bioactivity (11). IL-36 cytokines are processed by neutrophil proteases (elastase, proteinase-3, and cathepsin G) (12) and macrophage- and epithelial cell-derived cathepsin S (13).
The second checkpoint corresponds to its secretion, and this mechanism is a fundamental response to damage and infection (14). IL-1 cytokines secretion is affected by the lack of a signal peptide on their sequence (10), which prevents them from accessing the conventional protein secretion pathway (CPS). Thus, they use unconventional protein secretion pathways (UPS) that involve membrane pores, vesicles, transporters, among others (15). IL-1β is associated with two types of unconventional secretory pathways (16), vesicles and membrane pores (17). The receptor P2X7R is involved in IL-1β secretion under ATP exposure, a stimulus that opens a pore and promotes vesicle shedding (18). Similarly, the membrane pore formed by Gasdermin D (GSDMD) has been involved in IL-1β secretion under inflammatory conditions (19). However, it remains unknown if members of the IL-36 family follow the same secretion mechanisms.
The activation and secretion checkpoints are of paramount importance as they have a direct impact on cytokine functionality, which could help in the development of new therapeutic targets. Since IL-36 cytokines participate in the development of several inflammatory and autoimmune diseases, and little is known about their function and signaling mechanisms, exploring the mechanisms that regulate the secretion of the IL-36γ cytokine is fundamental to further understand IL-36 cytokines biology.
We studied IL-36γ secretion, and we found that LPS induced expression of IL-36γ cytokine in IC21 macrophages and perinuclear localization, and LPS/ATP induced its secretion. We explored non-conventional secretion pathways and observed P2X7R or GSDMD pores blockade limits the cytokine release. Collectively, our results show that IL-36γ cytokine follows a non-conventional secretion pathway using P2X7R and GSDMD pores.
Murine macrophages cell line IC21 (TIB-186) was cultured according to the manufacturer’s instructions (ATCC, VA).
1x106 IC21 cells/well were seeded in a 48-well plate (Corning, MA) and grown in RPMI-1640 (Corning, MA) medium supplemented with 10% fetal bovine serum (PAN Biotech, Germany), antibiotic/antimycotic (Corning, MA), and L-glutamine (Corning, MA). 24h after cell seeding the cells were stimulated for 18h with 1µg/ml LPS (Sigma-Aldrich, MO), and protein lysates were purified.
LPS-stimulated IC21 cells were co-stimulated for 1h with 3mM ATP (In vivoGen, CA), in the presence or absence of 2 µM monensin (Biolegend, CA), 5 µg/ml brefeldin A (Biolegend, CA), 200nM A438079 hydrochloride (TOCRIS, UK) or 20µM Necrosulfonamide (NSA; TOCRIS, UK) (specific inhibitors specific for P2X7R and Gasdermin D respectively). Supernatants and protein lysates were evaluated after co-stimulation.
The proteins present in the supernatant were precipitated with a 1:4 water-acetone mix at -20°C for 24h. The solution was centrifuged at 4700 g (Thermo-Fisher, MA) for 30min, and the pellet was washed twice with water-acetone and resuspended in RIPA (Sigma-Aldrich, MO).
40µg total protein were probed by western blot using primary Ab anti-IL-36γ TA505994; Origene, MD; 1:4,000), anti-GAPDH (GTX100118; GeneTex, CA; 1:1,000), and secondary anti-mouse-HRP (7076P2; Cell Signaling, MA; 1:5,000).
3.5x105 IC21 cells were seeded in 25mm sterile coverslips (Corning, MA) in a six-well plate (Corning, MA) and were LPS-induced and ATP-stimulated as before. After stimulation, the cells were fixed with 4% paraformaldehyde, permeabilized with 0.2% Triton, blocked with 0.2% BSA in PBS, and stained with the rabbit primary antibody α-IL-36γ (TA326667; Origene, MD; 1:1000), followed by staining with the secondary Alexa Fluor 594 α-rabbit IgG antibody (8889S; Cell Signaling, MA; 1:2000) and DAPI (Sigma-Aldrich, MO). Images were acquired using a white light laser Leica TCS SP8 X confocal microscope at 60X (Leica, Wetzlar, Germany).
The amino acid sequence of mouse IL-36γ, P2X7R, and GSDMD were retrieved from UniProtKB (Q8R460, Q9D8T2, and Q9Z1M0, respectively). Models were built in the SWISS-MODEL server (20) using the crystals for human IL-36γ, rat P2X7R, and mouse GSDMD as templates, and the models with a Qmean score closest to 0 were accepted. The program MOE was used to refine the models using energy minimization and removal of water molecules, and the server SymmDock (21) was used to build the 16-subunit oligomer GSDMD pore.
GraphPad Prism v9.3 software was used for data analysis. Statistical significance was determined by Student t-test between two groups. Mean ± SD of the data is presented. p<0.05 was statistically significant.
We first explored whether IL-36γ expression and secretion is altered by ATP, as other IL-1 superfamily cytokines (18). We stimulated IC21 murine macrophages and evaluated IL-36γ expression at protein (Figures 1A, B) and mRNA levels (data not shown). Stimulation with LPS increased IL-36γ expression, similar to previous reports in response to bacterial components (22). This correlates with the expression other members of the IL-36 subfamily under similar inflammatory conditions (23). We further observed that LPS/ATP stimulation induces the liberation of IL-36γ to the extracellular milieu (Figures 1A, C). Taken together, these results suggests that bacterial components, like LPS and ATP, not only induce IL-36γ expression but also trigger its secretion.
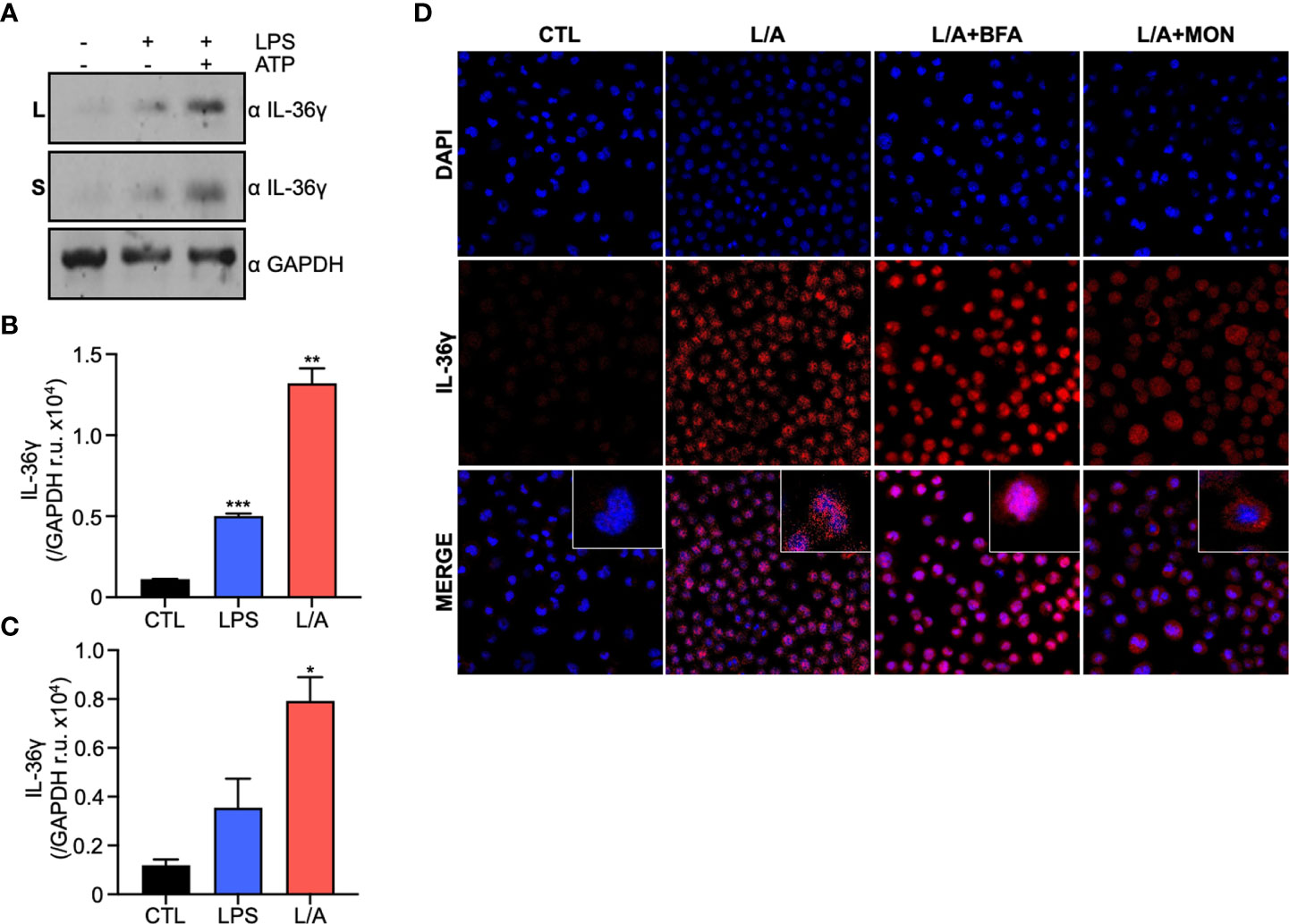
Figure 1 IL-36γ is secreted in response to ATP in a non-conventional pathway. (A) IL-36γ protein expression in lysates (L) and supernatants (S) of LPS or LPS/ATP activated macrophages. Densitometry of IL-36γ intensity in lysates (B) and supernatants (C) of cell stimulated with LPS or LPS/ATP (L/A). (D) IF of IC21 macrophages stimulated with LPS/ATP (L/A) and treated with monensin (Mon) or brefeldin A (BfA). *p<0.05, **p<0.01, ***p<0.001.
Cytokine secretion to the extracellular milieu is of paramount importance during inflammation to coordinate defense mechanisms in the host. Eucaryotic cells use the conventional and unconventional protein secretion pathways. The conventional secretory pathway involves the translocation of proteins into the endoplasmic reticulum (ER), transport to the Golgi apparatus and finally to the plasma membrane for their secretion. This pathway depends on a signal peptide at the N-terminal portion of the protein to follow this route (24). As it has been hypothesized that IL-36γ lacks a signal peptide (11), we used the server signal IP 5.0 to predict the presence of a signal peptide in silico and confirmed there is no signal peptide predicted on IL-36γ amino acid sequence (data not shown). We then explored if IL-36γ follows the conventional secretory pathway using monensin (MN) and brefeldin A (BfA) to block secretion. MN is a Na+ ionophore that affects regions of the Golgi apparatus that are linked to the final stages of secretory vesicle maturation. BfA is a macrocyclic lactone produced by fungi that inhibits protein secretion from the ER to the Golgi apparatus (25). We observed that when MN or BfA are added to the cell culture, stimulation with LPS/ATP triggers no IL-36γ accumulation within the cell (Figure 1D), demonstrating that IL-36γ does not follow the conventional secretory pathway for its secretion.
Lack of a signal peptide on IL-36γ sequence suggests it uses the unconventional secretory pathway. Of the four unconventional secretion pathways, membrane pores are the preferred mechanism for proteins that lack a signal peptide (15). In macrophages P2X7R (26) and GSDMD (19) form pores during inflammation or hyperactivation in an ATP-dependent manner. Both pores have been previously involved in IL-1β secretion (18, 19). Thus, we evaluated if IL-36γ, like IL-1β, uses P2X7R and/or GSDMD to exit the cell. We performed an in silico modeling of full-length and processed mouse IL-36γ, P2X7R, and GSDMD through three different servers, Swiss-Model, IntFold, and Robetta. The quality of the models was assessed through the Qmean score, Ramachandran score, and Molprobity score. The models built through the Swiss-Model server had the best scores and were used for further analysis. Full length (FL) and serin 42 processed (S42) IL-36γ were modeled based on the human IL-36γ crystal. The validation of the model gave a Qmean score of -0.53, Ramachandran score of 96.64% and Mol probity score of 1.29, indicating a good quality model. Processed IL-36γ model had the same scores as the FL variant, which suggests that the size reduction does not generate significant structural changes in the in silico model. Then, we analyzed the size and net charge, and we observed that both proteins have a positive net charge and an approximate diameter of 3.7nm (FL) and 3.5nm (S42) (Figure 2A). Next, we built the mouse homotrimer P2X7R in an open state, with bound ATP, based on the rat crystal due to the 85% identity. Validation of the model gave a Qmean score of -2.70, Ramachandran score of 94.79 and Molprobity score of 1.26. The pore diameter of P2X7R in an open state is 5nm in the in silico model, and has a negative net charge, particularly in the transmembrane and extracellular portions of the receptor (Figure 2B). Next, we built the NT-GSDMD model since it is the fraction that oligomerizes and forms the membrane pore. The model was generated based on the mouse crystal of full GSDMD due to the 97.97% identity. We obtained a medium-low quality model with a Q-mean score of-4.73, Ramachandran score of 86.67%, and Molprobity of 2.86. This low quality could be due to the low resolution of the crystal in the NT portion of the protein. We built the 16nm oligomer through the server SymmDock, which generates a 14nm diameter pore (Figure 2C). According to the data obtained by our in silico models, 3.7 and 3.5nm positively charged IL-36γ can theoretically pass through both the 5nm P2X7R pore and the 14nm GSDMD pore.
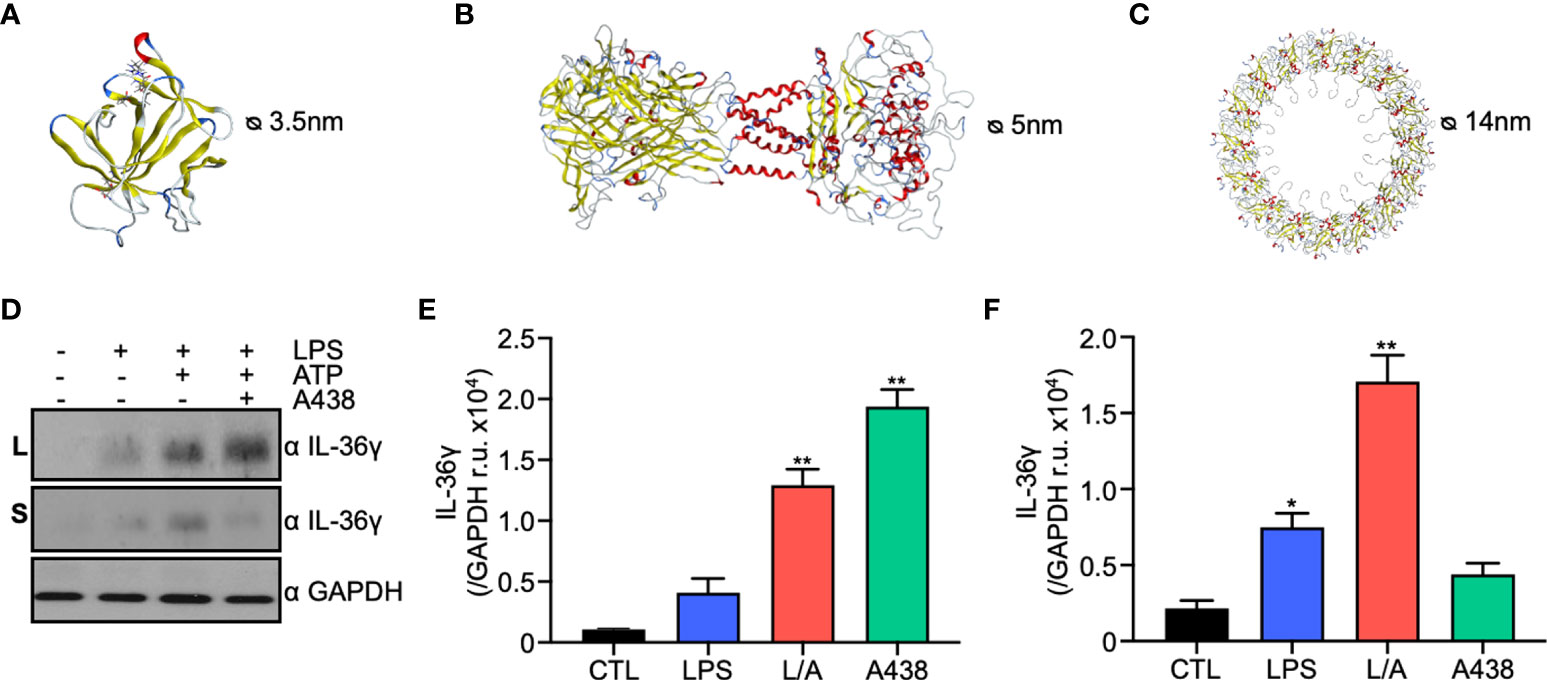
Figure 2 IL-36γ is secreted through the membrane pores P2X7R and GSDMD of the unconventional secretory pathway type I. (A) In silico model of mouse IL-36γ generated by SWISS-MODEL. (B) In silico model of P2X7R generated by SWISS-MODEL. (C) In silico model of GSDMD pore generated by SymmDock sever. (D) Western blot of lysates and supernatants under control, LPS, LPS/ATP (L/A), LPS/ATP+A438 conditions. Densitometry of IL-36γ in lysates (E) and supernatants (F). *p<0.05, **p<0.01.
Consistent with these results, we observed P2x7r and Gsdmd are upregulated in a microarray from pro-inflammatory intestinal mouse macrophages (Figure S1A) as well as LPS-activated IC21 macrophages (Figure S1B), suggesting an increased bioavailability in conditions where IL-36 γ is expressed.
To confirm the in silico data, we evaluated IL-36γ secretion through P2X7R and GSDMD experimentally. First, we used the P2X7R specific inhibitor A438 that binds to the inner face of the receptor blocking its opening, even in the presence of ATP. The presence of IL-36γ was evaluated by western blot in lysates and supernatants of IC21 macrophages under LPS or LPS/ATP and A438. We observed that in the presence of a P2X7R specific inhibitor, IL-36γ is accumulated within the cell, as observed in the lysates (Figures 2D, E). In contrast, when we evaluated the supernatants, we observed a clear reduction in secreted IL-36γ of approximately 75% (Figures 2D, F), suggesting that P2X7R participates in IL-36γ secretion. Then, we evaluated IL-36γ secretion by GSDMD pore. We used the specific inhibitor Necrosulfonamide (NSA), which binds to the NT domain of processed GSDMD and prevents its oligomerization and pore formation in the plasma membrane. Similar to P2X7R, we observed that in the presence of NSA, IL-36γ is accumulated within the cell (Figure S1C), suggesting that GSDMD also participates in IL-36γ secretion. Our data demonstrate that P2X7R and GSDMD have a role in IL-36γ secretion.
Finally, we evaluated IL-36γ localization within the cell in CTL, LPS, LPS/ATP, and LPS/ATP+A438/NSA conditions (Figure 3). We observed that IL-36γ localizes in the cytoplasm of IC21 macrophages, particularly in a perinuclear fashion, in CTL and LPS conditions. In the presence of LPS/ATP we observed that IL-36γ is accumulated in a scatter pattern in the cytoplasm and close to the plasma membrane, which falls in line with the induction of its secretion by the same stimulus, showing that LPS/ATP-induced secretion mobilizes IL-36γ towards the plasma membrane. In contrast, when we added the specific inhibitors A438 or NSA, IL-36γ changes its location and goes back to the perinuclear region as observed in CTL and LPS conditions. In contrast, when we added the specific inhibitor NSA, IL-36γ localizes close to the plasma membrane. Thus, our results show that the inhibition of IL-36γ secretion through P2X7R prevents the cytokine relocalization within the cell.
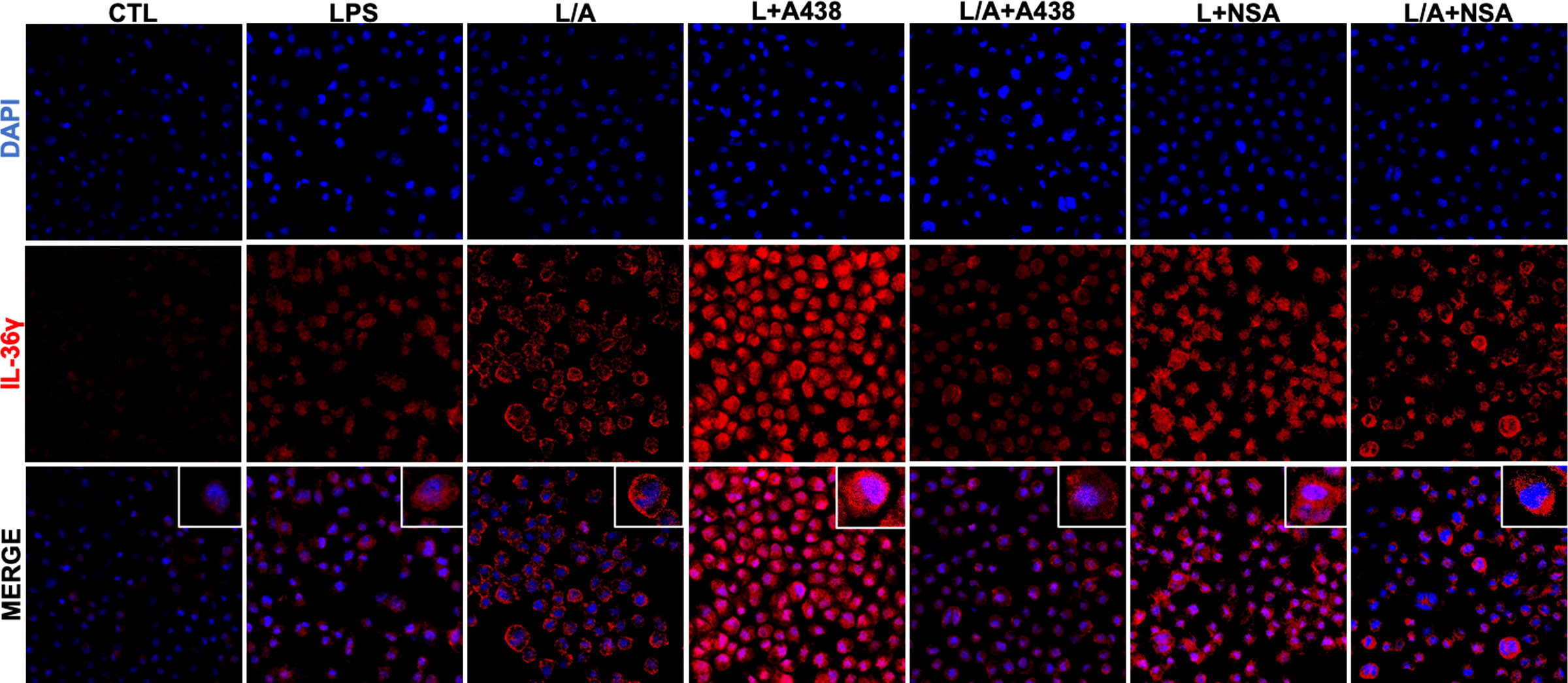
Figure 3 IL-36γ is localized in the cytoplasm of IC21 macrophages. IL-36γ localization in IC21 macrophages in control, LPS, LPS/ATP (L/A), LPS/ATP+A438, and LPS/ATP+NSA conditions. nuclei staining (DAPI), IL-36γ (Alexa Fluor594).
Together, our results demonstrate that IL-36γ is secreted mainly by an unconventional pathway through membrane pores formed by P2X7R and GSDMD.
In recent years there has been a growing interest in IL-36γ function and its involvement in the development of inflammatory diseases. However, a few mechanisms in this cytokine biology remain unknown. IL-36γ lack of a signal peptide (8, 22) suggests it could have access to unconventional secretory pathways (15, 27). Our data shows that the addition of LPS/ATP induces IL-36γ secretion. Our data agree with previous reports that have established that secretion of some members of the superfamily of IL-1, such as IL-1β, IL-18, and IL-36α, need two signals. LPS works as the first signal as it induces the expression of the cytokines; meanwhile, ATP works as the second signal triggering the secretion of the cytokines (23). It is interesting that such stimulation also activates P2X7R, an ATP receptor, as well as the inflammasome, which in turn activates GSDMD. GSDMD, once it is cleaved by caspase 1, oligomerizes and forms a pore on the plasma membrane (26, 28). Both P2X7R and GSDMD are members of the unconventional secretory pathway type I (15). Thus, we decided to evaluate if IL-36γ, like IL-1β and IL-18 (17, 29), is also secreted through these membrane pores. First, we evaluated in silico the IL-36γ secretion capabilities. According to our generated 3D models, the 3.7nm and 3.5nm size (full length and processed respectively), as well as the net positive charge of IL-36γ, allow its passage through the 5nm P2X7R and the 14nm GSDMD pores. Experimentally, we observed a significant intracellular accumulation and an extracellular reduction in IL-36γ levels when both P2X7R and GSDMD pores are blocked, confirming IL-36γ uses an unconventional secretion pathway. Our data suggest that secretion through membrane pores of the unconventional secretory pathway type I could be a common secretion mechanism for cytokines of the IL-1 superfamily. Consistent with our results, it has been observed that IL-36α is also secreted through P2X7R (23).
IL-36γ localizes preferentially to the cytoplasm of IC21 macrophages, particularly in a perinuclear region under homeostatic conditions. After LPS stimulation it remains localized to the perinuclear region, but LPS/ATP scattered IL-36γ in the cytoplasm closer to the plasma membrane, which agrees with its secretion in response to this stimulus. Other members of the superfamily like IL-1α, IL-1β, and IL-18 have polybasic regions in their sequence, which allows them to adhere to the plasma membrane in preparation for secretion (16); we found through in silico prediction that IL-36γ contains a small polybasic region that could allow its association to the plasma membrane through electrostatic interactions. Also, we observed that under the P27XR specific inhibition, IL-36γ appears to go back to the perinuclear area. In contrast, under GSDMD specific inhibitor IL-36γ remains close to the membrane. Suggesting that ATP is the stimuli that affects IL-36γ localization since we only observed this phenomenon when P2X7R is blocked. Previous studies have shown that IL-33, member of IL-1 superfamily, localizes to the nucleus under homeostatic conditions and it relocalizes to the cytoplasm in response to stress. This relocalization depends on the microtubule network and ATP (30). It has been shown that ATP participates in microtubule assembly (31) and it triggers vesicle movement towards the plus end for their secretion (32). Thus, IL-36γ localization dependance on ATP could be explained if IL-36γ travels along microtubules to be secreted. It’s worth exploring if IL-36γ is intracellularly localized in vesicles that could potentially travel along microtubules towards the plasma membrane for its exocytosis.
Here we have demonstrated that IL-36γ is secreted through an unconventional secretory pathway type I, specifically through the P2X7R and GSDMD membrane pores in a mechanism dependent on LPS/ATP stimulation. As P2X7R/GSDMD have an important role in the secretion of IL-36γ; thus, we believe that blocking these pores can be a novel therapeutic approach to limit IL-36 cytokines bioavailability in several autoimmune diseases, cancer, obesity, and chronic inflammatory pain, among others, and ameliorate symptoms, as has been proposed elsewhere (33, 34).
Publicly available datasets were analyzed in this study. This data can be found here: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token=olgvmwkanfgzlcd&acc=GSE68269.
OM-C and LM-M conceived and designed the analysis. LM-M, CG-R, AJP and FC-M collected the data. GP-L, ER-A, RV-R and VO-N contributed data or analysis tools. LM-M and OM-C performed the analysis. OM-C and LM-M wrote the paper. All authors contributed to the article and approved the submitted version.
This work was supported by CONACYT grant CB2016-01/280815 to O. Medina-Contreras. L Manzanares-Meza received the CONACYT scholarship 589088.
We thank D. Castro-Eguiluz for her scientific editing and writing support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.979749/full#supplementary-material
Supplementary Figure 1 | (A) Microarray analysis from DSS-treated mouse colonic macrophages. (B) qPCR expression for P2x7r and Gsdmd in IC21 macrophages stimulated with LPS or LPS/ATP. (C) Densitometry of IL-36γ in macrophages under CTL, LPS/ATP or LPS/ATP+NSA stimulation.
1. Ross EA, Devitt A, Johnson JR. Macrophages: The good, the bad, and the gluttony. Front Immunol (2021) 12:708186. doi: 10.3389/fimmu.2021.708186
2. Paludan SR, Pradeu T, Masters SL, Mogensen TH. Constitutive immune mechanisms: mediators of host defence and immune regulation. Nat Rev Immunol (2021) 21:137–50. doi: 10.1038/s41577-020-0391-5
3. Dinarello CA. Introduction to the interleukin-1 family of cytokines and receptors: Drivers of innate inflammation and acquired immunity. Immunol Rev (2018) 281:5–7. doi: 10.1111/imr.12624
4. Dinarello C, Arend W, Sims J, Smith D, Blumberg H, O’Neill L, et al. IL-1 family nomenclature. Nat Immunol (2010) 11:973–3. doi: 10.1038/ni1110-973
5. Debets R, Timans JC, Homey B, Zurawski S, Sana TR, Lo S, et al. Two novel IL-1 family members, IL-1δ and IL-1ϵ, function as an antagonist and agonist of NF-κB activation through the orphan IL-1 receptor-related protein 2. J Immunol (2001) 167:1440–6. doi: 10.4049/jimmunol.167.3.1440
6. Towne J, Sims J. IL-36 in psoriasis. Curr Opin Pharmacol (2012) 12:486–90. doi: 10.1016/j.coph.2012.02.009
7. Towne JE, Garka KE, Renshaw BR, Virca GD, Sims JE. Interleukin (IL)-1F6, IL-1F8, and IL-1F9 signal through IL-1Rrp2 and IL-1RAcP to activate the pathway leading to NF-κB and MAPKs. J Biol Chem (2004) 279:13677–88. doi: 10.1074/jbc.M400117200
8. Gabay C, Towne JE. Regulation and function of interleukin-36 cytokines in homeostasis and pathological conditions. J Leukoc Biol (2015) 97:645–52. doi: 10.1189/jlb.3RI1014-495R
9. Stenken JA, Poschenrieder AJ. Bioanalytical chemistry of cytokines – A review. Anal Chim Acta (2014) 853:95–115. doi: 10.1016/j.aca.2014.10.009
10. Dinarello CA. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol Rev (2017) 281:8–27. doi: 10.1111/imr.12621
11. Towne JE, Renshaw BR, Douangpanya J, Lipsky BP, Shen M, Gabel CA, et al. Interleukin-36 (IL-36) ligands require processing for full agonist (IL-36α, IL-36β, and IL-36γ) or antagonist (IL-36Ra) activity. J Biol Chem (2011) 286:42594–602. doi: 10.1074/jbc.M111.267922
12. Henry CM, Sullivan GP, Clancy DM, Afonina IS, Kulms D, Martin SJ. Neutrophil-derived proteases escalate inflammation through activation of IL-36 family cytokines. Cell Rep (2016) 14:708–22. doi: 10.1016/j.celrep.2015.12.072
13. Ainscough JS, Macleod T, McGonagle D, Brakefield R, Baron JM, Alase A, et al. Cathepsin s is the major activator of the psoriasis-associated proinflammatory cytokine IL-36γ. Proc Natl Acad Sci (2017) 114:E2748–57. doi: 10.1073/pnas.1620954114
14. Stanley AC, Lacy P. Pathways for cytokine secretion. Physiology (2010) 25:218–29. doi: 10.1152/physiol.00017.2010
15. Rabouille C. Pathways of unconventional protein secretion. Trends Cell Biol (2017) 27:230–40. doi: 10.1016/j.tcb.2016.11.007
16. Monteleone M, Stanley AC, Chen KW, Brown DL, Bezbradica JS, von Pein JB, et al. Interleukin-1β maturation triggers its relocation to the plasma membrane for gasdermin-D-Dependent and -independent secretion. Cell Rep (2018) 24:1425–33. doi: 10.1016/j.celrep.2018.07.027
17. Qu Y, Franchi L, Nunez G, Dubyak GR. Nonclassical IL-1β secretion stimulated by P2X7 receptors is dependent on inflammasome activation and correlated with exosome release in murine macrophages. J Immunol (2019) 179:1913–25. doi: 10.4049/jimmunol.179.3.1913
18. Giuliani AL, Sarti AC, Falzoni S, Virgilio FD. The P2X7 receptor-Interleukin-1 liaison. Front Pharmacol (2017) 8:123. doi: 10.3389/fphar.2017.00123
19. Evavold CL, Ruan J, Tan Y, Xia S, Wu H, Kagan JC. The pore-forming protein gasdermin d regulates interleukin-1 secretion from living macrophages. Immunity (2018) 48:35–44.e6. doi: 10.1016/j.immuni.2017.11.013
20. Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res (2018) 46:W296–303. doi: 10.1093/nar/gky427
21. Schneidman-Duhovny D, Inbar Y, Nussinov R, Wolfson HJ. Geometry-based flexible and symmetric protein docking. Proteins Struct Funct Bioinform (2005) 60:224–31. doi: 10.1002/prot.20562
22. Medina-Contreras O, Harusato A, Nishio H, Flannigan KL, Ngo V, Leoni G, et al. Cutting edge: IL-36 receptor promotes resolution of intestinal damage. J Immunol (2016) 196:34–8. doi: 10.4049/jimmunol.1501312
23. Martin U, Scholler J, Gurgel J, Renshaw B, Sims JE, Gabel CA. Externalization of the leaderless cytokine IL-1F6 occurs in response to Lipopolysaccharide/ATP activation of transduced bone marrow macrophages. J Immunol (2009) 183:4021–30. doi: 10.4049/jimmunol.0803301
24. Viotti C. Unconventional protein secretion, methods and protocols. Methods Mol Biol (2016) 1459:3–29. doi: 10.1007/978-1-4939-3804-9_1
25. O’Neil-Andersen NJ, Lawrence DA. Differential modulation of surface and intracellular protein expression by T cells after stimulation in the presence of monensin or brefeldin a. Clin Diagn Lab Immunol (2002) 9:243–50. doi: 10.1128/CDLI.9.2.243-250.2001
26. Alves LA, de Melo Reis RA, de Souza CAM, de Freitas MS, Teixeira PCN, Ferreira DNM, et al. The P2X7 receptor: Shifting from a low- to a high-conductance channel — an enigmatic phenomenon? Biochim Biophys Acta Biomembr (2014) 1838:2578–87. doi: 10.1016/j.bbamem.2014.05.015
27. Boutet M-A, Bart G, Penhoat M, Amiaud J, Brulin B, Charrier C, et al. Distinct expression of interleukin (IL)-36α, β and γ, their antagonist IL-36Ra and IL-38 in psoriasis, rheumatoid arthritis and crohn’s disease. Clin Exp Immunol (2016) 184:159–73. doi: 10.1111/cei.12761
28. Wiley JS, Sluyter R, Gu BJ, Stokes L, Fuller SJ. The human P2X7 receptor and its role in innate immunity. Tissue Antigens (2011) 78:321–32. doi: 10.1111/j.1399-0039.2011.01780.x
29. Gardella S, Andrei C, Poggi A, Zocchi MR, Rubartelli A. Control of interleukin-18 secretion by dendritic cells: role of calcium influxes. FEBS Lett (2000) 481:245–8. doi: 10.1016/S0014-5793(00)02015-9
30. Kakkar R, Hei H, Dobner S, Lee RT. Interleukin 33 as a mechanically responsive cytokine secreted by living cells. J Biol Chem (2012) 287:6941–8. doi: 10.1074/jbc.M111.298703
31. Murray JW, Bananis E, Wolkoff AW. Reconstitution of ATP-dependent movement of endocytic vesicles along microtubules In vitro: An oscillatory bidirectional process. Mol Biol Cell (2000) 11:419–33. doi: 10.1091/mbc.11.2.419
33. Ngo VL, Kuczma M, Maxim E, Denning TL. IL-36 cytokines and gut immunity. Immunology (2021) 163:145–54. doi: 10.1111/imm.13310
Keywords: IL-36γ, inflammation, macrophages, cytokin receptors, secretion
Citation: Manzanares-Meza LD, Gutiérrez-Román CI, Jiménez-Pineda A, Castro-Martínez F, Patiño-López G, Rodríguez-Arellano E, Valle-Rios R, Ortíz-Navarrete VF and Medina-Contreras O (2022) IL-36γ is secreted through an unconventional pathway using the Gasdermin D and P2X7R membrane pores. Front. Immunol. 13:979749. doi: 10.3389/fimmu.2022.979749
Received: 27 June 2022; Accepted: 04 August 2022;
Published: 18 August 2022.
Edited by:
Juan Bautista De Sanctis, Palacký University Olomouc, CzechiaReviewed by:
Rodolfo Fernández-Gómez, Independent Researcher, Caracas, VenezuelaCopyright © 2022 Manzanares-Meza, Gutiérrez-Román, Jiménez-Pineda, Castro-Martínez, Patiño-López, Rodríguez-Arellano, Valle-Rios, Ortíz-Navarrete and Medina-Contreras. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oscar Medina-Contreras, b21lZGluYUBoaW1mZy5lZHUubXg=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.