 Yiran Yang
Yiran Yang Wanning Wang
Wanning Wang Jing Weng
Jing Weng Huifang Li3
Huifang Li3 Yanmin Ma
Yanmin Ma Lingyan Liu
Lingyan Liu Wei Ma
Wei Ma
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 29 August 2022
Sec. Immunological Tolerance and Regulation
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.976289
This article is part of the Research Topic Immune Responses and Immunotolerance in the Process of Reproduction View all 16 articles
The HLA class Ib molecule is an alloantigen that causes transplant rejection on behalf of individual human and plays an important role in maternal-fetal immune tolerance. Early studies on HLA class Ib focused on the mechanism of HLA-G-induced immune escape, but in recent years, studies on the mechanism of HLA-G have deepened and gradually explored the mechanism of HLA-E and HLA-F, which are also HLA class Ib molecules. In the maternal-fetal interface, trophoblast cells express HLA class Ib molecules to protect the fetus from maternal immune cells by binding to inhibitory receptors of decidual immune cells (DICs) and shifting Th1/Th2 balance toward Th2 bias. Further studies on the molecular mechanism of HLA class Ib molecules provide a reference for its application in the field of clinical assisted reproduction.
Major histocompatibility complex (MHC) is a group of genes closely linked on the same chromosome, which is closely related to immune response and transplantation rejection. Human MHC is called Human Leukocyte Antigen (HLA) complex. HLA complex is located on the short arm of chromosome 6, with a total of 224 loci. It is divided into three regions according to the structure and function of each point gene and its coding product, namely class I, class II and class III gene region. HLA class I gene region contains non-classical HLA-E, F, G and other loci (1), the genes on which are called HLA class Ib genes (2). The probability of HLA being identical between two unrelated individuals is extremely small, and this variability leads to allograft rejection. As diploid organisms, humans have two different HLA inherited from both parents (3).
The embryo implantation process can be regarded as a semi-allogeneic transplant process. The embryo with paternal antigen will theoretically cause maternal transplantation rejection, which is contrary to the fact that it is not attacked by the maternal immune system before delivery (4). Therefore, it can be inferred that there is a special tolerance effect on the maternal-fetal interface to ensure the normal progress of pregnancy.
Maternal-fetal immune tolerance occurs at the maternal-fetal interface, which is composed of maternal decidua and chorion developed from the trophoblast of blastocyst. According to previous studies, it is generally believed that the immune tolerance of the maternal immune system to the fetus is related to the interaction between the immunotolerant microenvironment of the extravillous trophoblast and the deciduous layer. There are a large number of DICs in the immunotolerant microenvironment of the decidual layer to ensure the normal process of embryo implantation and spiral artery reconstruction in early pregnancy (5). DICs exist throughout pregnancy, and the population frequency varies with the stages of pregnancy. Decidual natural killer cells (dNK) account for the majority of DICs, followed by decidual macrophages and decidual T cells and so on. However, when maternal blood is in direct contact with syncytiotrophoblasts (STBs) in late pregnancy, DICs can enter the maternal-fetal interface and carry out immune rejection to the fetus (6–8).
In recent years, relevant studies have shown that HLA class Ib play an important role in maternal-fetal immune tolerance (9). In order to prevent the fetus from being attacked by the maternal immune system, extravillous trophoblasts (EVTs) express HLA class Ib molecules such as HLA-E, HLA-F and HLA-G to change the function of DICs and regulate its subtypes (7). Therefore, this review will elaborate on the comprehensive mechanisms and frontier applications in HLA class Ib, in the hope of offering new ideas for the diagnosis and treatment of pregnancy-related diseases.
The HLA class Ib genes include HLA-E, HLA-F and HLA-G, and are characterized as being non-polymorphic compared to their classical counterparts and encode molecules involved in immune regulation and immune suppression (2). The HLA class Ib genes expressed on the blastocyst, which developed from the fertilized egg and further differentiates into inner cell mass (ICM) and trophectoderm (TE) (7). TE further differentiates into epithelial trophoblasts (10). As one type of epithelial trophoblasts, cytotrophoblasts (CTBs) have two distinct differentiation pathways, generating syncytiotrophoblasts (STBs) and extravillous trophoblasts (EVTs) (11).
The expression of HLA-E on the cell surface is regulated by the acquisition of peptides derived from the leader sequences of HLA-C and HLA-G molecules (12), and can be discovered in all stages of pregnancy. HLA-E was found to be expressed on the surface of EVTs (13), and it was weakly expressed in CTBs and STBs at 5 weeks of gestation (14). The expressions of HLA-F are less well understood, based on existing research results, it is still controversial whether HLA-F is expressed in cells or on the surface of cells (12). In fact, studies have shown that HLA-F was observed to be expressed on the surface of EVTs (14), and in the cytoplasm of CTBs, STBs and EVTs (15, 16). HLA-G is most abundantly expressed at immune-privileged sites (2), it can be found extensively expressed on the surface of EVTs during the entire pregnancy (12), and its soluble isoforms can be secreted by CTBs and STBs (Figure 1) (15, 17).
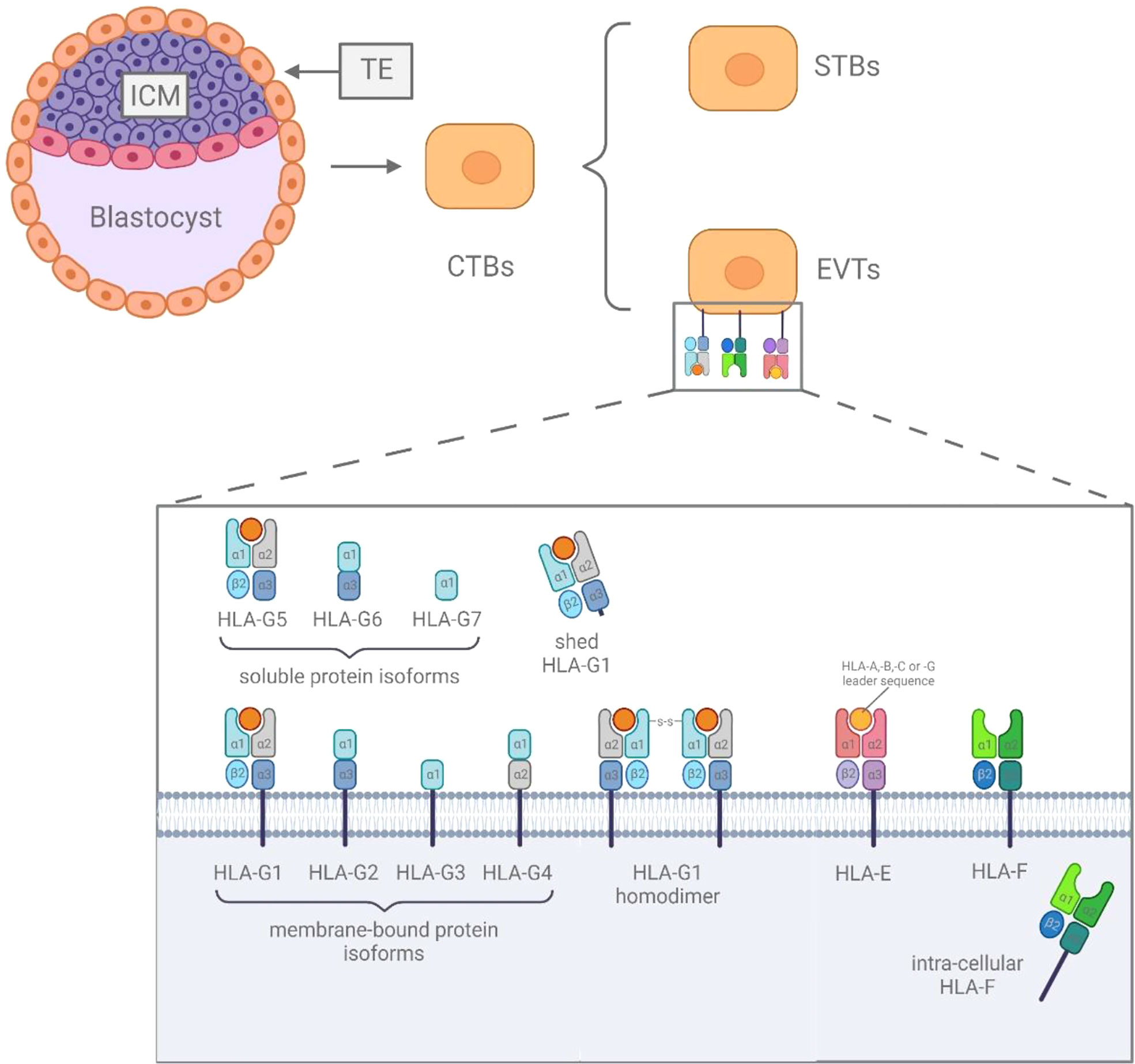
Figure 1 The difference among HLA class Ib.
HLA-G is exclusively expressed in extravillous trophoblast cells (18, 19). It can be present in seven isoforms, four membrane-bound (m) isoforms (mHLA-G: HLA-G1, -G2, -G3 and -G4) and three soluble (s) isoforms (sHLA-G: HLA-G5, -G6 and -G7). In addition, the membrane-bound HLA-G1 can also be available in soluble form, named shed HLA-G1, which is mediated by metalloproteinase cleavage (20). Furthermore, there are HLA dimers linked by disulfide bonds, which have shown higher receptor affinity and slower dissociation rates in several studies (21, 22). In 1997, HLA-G was first proved to have a protective effect on the fetus after semi-allogeneic transplantation in the maternal immune system, which was also supported in pathological research by Yie et al (23). Nowadays, HLA-G is considered to be a major immune checkpoint molecule and plays a crucial role in maternal-fetal immune tolerance (24, 25). HLA-G can increase the activation threshold of immune cells before immune response by up-regulating the expression of inhibitory receptors on immune cells such as dNK cells and decidual T cells (26–28).
sHLA-G and mHLA-G recognize and bind killer cell immunoglobulin-like receptor KIR2DL4, immunoglobulin-like transcript 2 (ILT2), immunoglobulin-like transcript 4 (ILT4) and inhibitory receptor CD94/NKG2A of C-type lectin superfamily (Figure 2) on dNK cells, inactivating NK cell effectors (29–36). KIR2DL4 transduces inhibitory signals and is a killing inhibitory receptor on the surface of NK cells (37). HLA-G is the only known ligand of KIR2DL4 (38). The combination can transmit immunosuppressive signals, hence, to promote the development of a beneficial immune tolerance environment to the trophoblast/fetus, by protecting trophoblast cells from the killing of maternal dNK cells (39–44). In addition, the combination of HLA-G and KIR2DL4 can also promote dNK cells to secrete pro-inflammatory and pro-angiogenic factors (i.e., IL-1B, IL-6, IL-8, TNF-α, MMP, IFN- γ, VEGF, ANG). These cytokines ensure sufficient blood supply for the developing fetus and promote embryo implantation by invading the decidua and participating in the vascular remodeling of uterine spiral artery (Table 1) (45). ILT2 is expressed on the surface of T cells, B cells, monocytes/macrophages, dendritic cells (DC) and NK cells, can recognize the expression of HLA-G β-2 microglobulin. HLA-G inhibits the cytotoxicity of dNK cells by up-regulating the expression of ILT2 (26, 28, 46, 47). At the same time, the interaction between ILT2 and HLA-G can also inhibit the formation of NK-cell synapse and significantly reduce the ability of NK cells to kill target cells (Table 1) (28). ILT4 also binds HLA-G and plays a similar function to ILT2 (Table 1) (44, 45). CD94/NKG2A is another inhibitory receptor on the surface of dNK cells. It can recognize HLA-G1 (and HLA-E) expressed on trophoblasts, and negatively regulate the cytotoxicity of dNK cells (Table 1) (41, 48, 49).
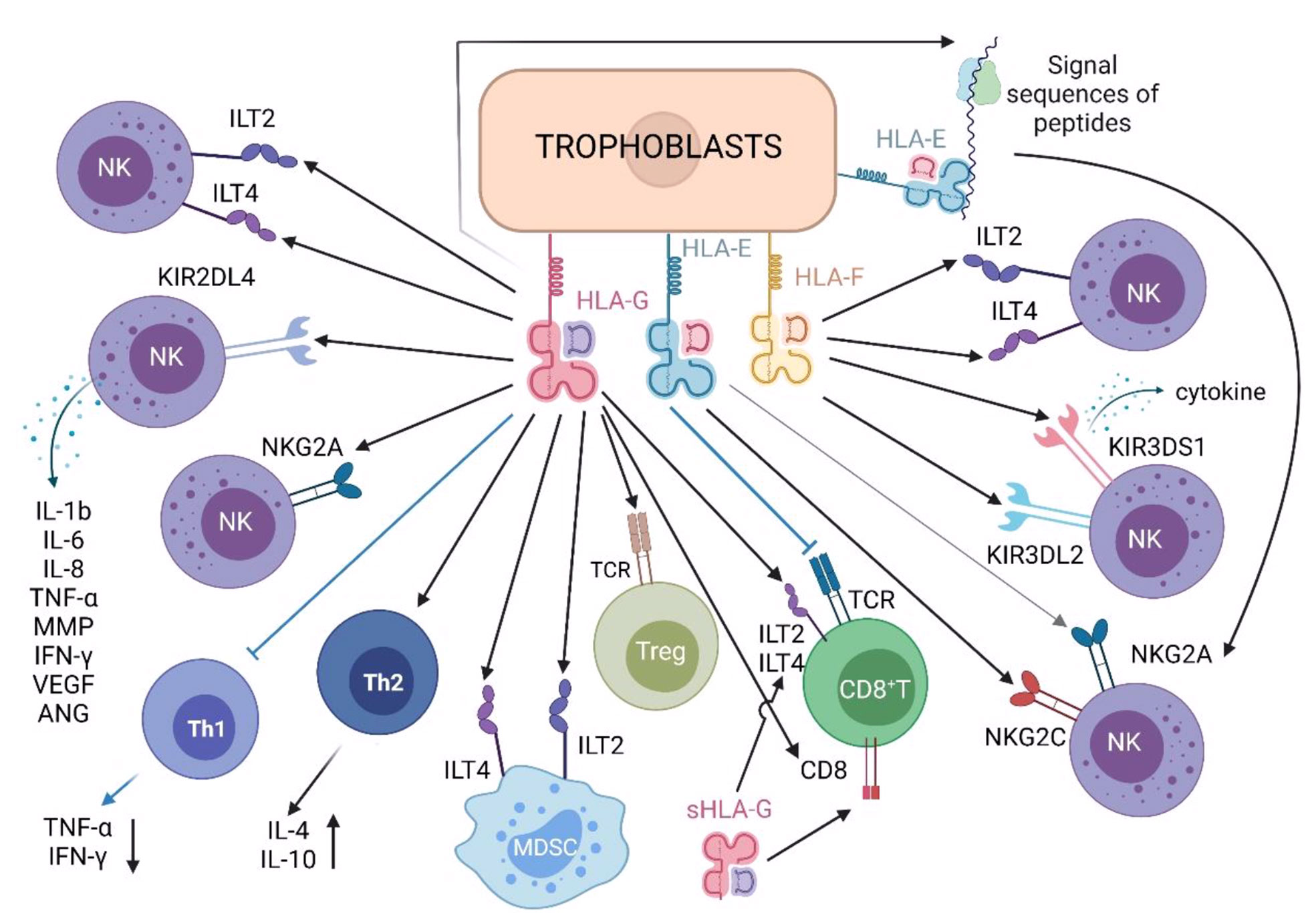
Figure 2 Roles of HLA class Ib molecules in immune cells in maternal decidua.

Table 1 The recent studies of HLA class Ib in Reproductive immunology.
mHLA-G can even inhibit the cytotoxicity of NK cells by binding to the suppressive subset of NK cells (NK-ireg) and secreting inhibitory molecules (75).
Trogocytosis (76) is a direct transfer of membrane and membrane-related molecules between cells in a contact manner (39). CD4+ T cells, CD8+ T cells, dNK cells and monocytes have been found that can obtain HLA-G through trogocytosis (45, 77). For example, CD4+ T cells can obtain HLA-G protein from decidual DC through trogocytosis and become the trogocytosis-based generation of temporary regulatory CD4+ HLA-Gacq+ T cells (78–80). In addition, trogocytosis prolongs the effect of HLA-G on KIR2DL4-mediated signaling in dNK cells, increases the secretion of cytokines and other small proteins, and plays an important role in placental and fetal development, as well as the establishment of immune tolerance. Therefore, trogocytosis contributes to HLA-G-mediated extensive immunosuppression at the maternal-fetal interface (39).
Studies have shown that both membrane-bound and soluble HLA-G proteins inhibited T cells alloproliferation (50). This inhibition involved engagement of ILT2 and ILT4 receptors by HLA-G (Table 1) (50, 51). Moreover, HLA-G can interaction with CD8 to enhance FasL expression, leading to apoptosis of CD8+ T cells (52), and can mediate cell-cycle inhibition of alloreactive T cells through Fas/FasL interaction (Table 1) (50), effectively inhibit the immune killing activity of cytotoxic T cells(CTL).
Treg cells protect the fetus after semi-allogeneic transplantation by binding with HLA-G (Figure 2), achieve maternal-fetal immune tolerance (53). Both mice and human cell experiments have shown that CD4+ CD25high regulatory T cell population increases during pregnancy, and this trend can be observed as early as early pregnancy (54). At the same time, HLA-G has been found to have a direct induction effect on the enrichment of Treg cells (Table 1) (6, 40).
Normal pregnancy process tends to participate in humoral immunity with Th2-type cytokines and avoid cellular immunity with Th1-type cytokines. The imbalance of Th1/Th2 ratio can lead to maternal rejection of the fetus. HLA-G can promote the production of Th2-type cytokine interleukin-4 (IL-4) and up-regulate Th2-type cytokine interleukin-10 (IL-10) (55), and inhibit the synthesis of Th1-type cytokine interferon- γ(IFN- γ). HLA-G can also inhibit the tumor necrosis factor-α(TNF-α), which belongs to Th1-type cytokine. Thus, it can block the activation of NK cells, regulate placental growth and then maintain Th1/Th2 balance (Table 1) (56).
MDSCs are innate immune cells, which increase during pregnancy. MDSCs simultaneously express ILT2 and ILT4 and inhibit the immune killing function of T cells (44). Natascha Köstlin et al. first described the direct effect of human sHLA-G on MDSCs (Figure 2). The study showed that sHLA-G induced MDSCs functionally and quantitatively through the signal transduction of ILT4 and the activation of transcription activator 3 (STAT3), reducing the positive rates of CD4+ T cells and CD8+ T cells (Table 1) (27).
In the process of reproductive immunity, HLA-E plays a protective role in the earliest events of implantation but not in active EVT invasion (19). HLA E mainly down-regulate the immune response at the maternal-fetal interface by cooperating with classical HLA class I molecules to ensure the success of pregnancy.
The expression of HLA-E is induced by many costimulators regulating classical HLA-I. One of HLA-E’s key functions is to regulate the activity of NK cells (81). In other words, the synergistic up-regulation of HLA-E and classical HLA-I is to protect target cells from NK mediated cytotoxicity. In an inflammatory environment in which many immune cells are recruited, this mechanism will protect bystander cells from immune system attacks while still allowing targeted destruction of some dysfunctional/virus infected cells (82). In the process of reproductive immunity, the immunological mechanism that determines the success of pregnancy mainly depends on the interaction between placental trophoblast and decidual immune microenvironment. HLA-E is an immunosuppressive factor, so the decrease of its expression level will weaken the down-regulation of immune response mechanism, leading to the termination of pregnancy (83).
HLA-E is of vital importance in the maternal immune process, because it can down regulate the maternal immune response, so as to protect the fetus from the attack of the maternal immune system. Among them, the interaction between HLA-E and NK cell CD94/NKG2A receptor is the most typical (Table 1) (57–59).
NK cells can express a series of regulatory receptors related to their activation and inhibition, and selectively kill “non-self” components (84). HLA class I molecules are key regulators of NK cell activation. They regulate NK cell activity through interaction with inhibitory receptors and activated receptors. Therefore, HLA molecules are the key immune checkpoint of NK cells (85).
CD94/NKG2A heterodimer is an inhibitory receptor. It interacts with the trimer ligand which consists of HLA-E, β2m and a nonameric peptide. The receptor protein NKG2A can regulate the activity of NK cells, and CD94 plays a corresponding role after binding with the ligands (60).
HLA-E can interact with TCR on CD8+ T cells and regulatory T cells (61, 62), and with receptors NKG2A and NKG2C expressed on NK cells and some T cell subsets (Figure 2).Under physiological state, the engagement of CD94/NKG2A and HLA-E induced inhibitory signals that prevents NK cell activation (63). In other words, HLA-E can stimulate immune activation and inhibitory effects. When both NKG2A and NKG2C form heterodimers with CD94, NKG2A family members expressing inhibitory effects have more affinity for HLA-E than NKG2C receptors expressing activating effects (Table 1) (64, 65). For these two receptors, the results of receptor ligand interaction mainly depend on the level of HLA-E expression. In addition, P. Tripathi et al. investigated the HLA-E gene polymorphism of normal pregnant women and patients with recurrent spontaneous abortion. They found that the expression product of HLA-EG allele had high affinity with the receptor and was highly associated with successful pregnancy (57).
In addition, HLA-E single chain trimer contains an additional (G4S)3 linker, which is fused with the peptide from HLA-G signal sequence into a non-polymorphic peptide, and presented by HLA-E. It can also inhibit NK cells dependent lysis by binding to CD94/NKG2A (81).
Norman Shreeve et al. obtained a relatively complete conclusion that maternal HLA-B/HLA-E/NKG2A pathway was conducive to healthy pregnancy and might have an impact on the health of offspring (59). By regulating this pathway, the occurrence of preeclampsia syndrome could be reduced, and abnormal pregnancy could be avoided.
In 2021, Eva Prašnikar and his team identified NKG2C/HLA-Eα2 domains and nonameric peptide were key elements involved in the molecular mechanism of signal transduction through intertwined hydrogen bond networks (86). Some studies also found that Ly49 gene family was involved in the synergistic regulation of the above process (87).
Because HLA-E can only be expressed in cells after binding to the signal peptide sequence derived from HLA-G (88), the expression of HLA-G was detected in all HLA-E expressing trophoblasts. Therefore, we speculate that HLA-G and E cooperate to inhibit the activity of dNK cells by binding inhibitory receptors, so as to protect trophoblasts from dNK cells killing, enable the maternal-fetal immune tolerance to embryonic alloantigens, and regulate the infiltration process of trophoblasts (88).
HLA-F was first discovered in 1990 (89), and there are few studies on the mechanism of HLA-F. The latest research showed that HLA-F could be expressed in an open conformation and bind to many Killer Cell Immunoglobulin Like Receptors (KIR receptors) on the surface of NK cells. They could also be expressed as HLA-F tetramers, binding to ILT2 and ILT4 (Figure 2) (69), which played a role in endometrial specific immune regulation during blastocyst implantation (90).
HLA-F protein is expressed by extravillous trophoblast cells with immunomodulatory properties and plays a role at the maternal-fetal interface (91). It was found that the expression of HLA-F protein could be detected on the cell surface of the extravillous trophoblast invading the maternal decidua (15, 92). HLA-F protein is related to the maternal-fetal immune tolerance to placental tissue. The EVT cell surface HLA-F in it interacts with the maternal immune cells of the decidua and protect the invading EVT from immune attack (14). HLA-F is expressed throughout pregnancy, and the expression increases with the passage of pregnancy time (93, 94). Its abnormally low expression (compared with healthy pregnant women) will lead to many diseases, such as Intrahepatic Cholestasis of Pregnancy (ICP), gestational hypertension, preeclampsia and gestational diabetes mellitus (19). These diseases are also closely related to low expression of HLA-G and HLA-E (95–99).
HLA-F is essential in the normal function of decidual trophoblast cells, as HLA-F is the third necessary partner of HLA-E and HLA-G in cell communication, while cell communication is the basis of pregnancy immunology. On this basis, studies showed that CD4+ CD25+ Treg mediated maternal tolerance to fetus (72, 100). These findings confirm that HLA-F is involved in the interaction between placental derived extravillous trophoblast and regulatory cells.
The variation of HLA-F and TAP2 genes leaded to shorter pregnancy time, indicating their role in endometrial specific immune regulation during implantation. The study also showed that HLA-F was necessary in maternal fetal immune regulation, which laid a foundation for the later study of the role of HLA-F in reproductive immunology. Moreover, any gene expression disorder in endometrium during pregnancy might lead to pregnancy failure. A variety of HLA molecules played an important and independent role at the maternal-fetal interface (101).
Aura Burian et al. found that HLA-F and MHC-I open conformers(OCs) regulated the reactivity and specificity of KIR3DL2 to the target cells and effectors (66). KIR3DL2 receptor was an inhibitory receptor on the surface of NK cells (67). It can be activated after binding with HLA-F to produce immunosuppression, resulting in the weakening of Uterine nature killer (uNK) cells’ killing effect and the establishment of maternal immune tolerance to placental tissue (68). At the same time, because MHC-I OC and HLA-F are co-expressed, and the affinity between HLA-F and different MHC-I alleles is different, the expression level of HLA-F may be modified by allele MHC-I OC, and vice versa.
HLA-F has direct molecular interaction with ILT2 and ILT4 (69, 70). HLA-F might be a peptide binding molecule that can reach the cell surface and bind to the target peptide. On the cell surface, it can interact with LIR1 (ILT2) and LIR2 (ILT4) receptors to change the activation threshold of immune effector cells (69). During pregnancy, if CD4+ Th1 cells release Th1 cytokines, the immune response mainly mediated by cellular immune response will lead to pregnancy failure. Therefore, the expression of HLA-F in endometrial system, the expression of ILT2 and ILT4 on decidual T cells (Table 1) (71), and the interaction between HLA-F tetramer and ILT2 and ILT4 receptors, will reduce the immune rejection and protect the fetus from the damage caused by maternal immune rejection, so as to achieve the purpose of successful pregnancy.
The researchers experimentally confirmed the receptor ligand relationship between KIR3DS1 and MHC-I OCs - the activation of KIR might be mainly in the inflammatory response of up-regulated HLA-F and MHC-I OCs (Table 1) (73, 74). Then, some researchers proposed that HLA-F interacts with KIR3DS1 to activate NK cells (72). uNK cells are the main immune cells in uterine decidua (102). The contact between uNK cells and extravillous trophoblast cells is the first step of maternal immune recognition of placental tissue. Despite the strong lethality of uNK cells, the activation of uNK cells during pregnancy may be a necessary condition for the secretion of cytokines and growth factors, which are essential for the blood supply of the placenta.
All three SNPs in the HLA-F locus regulate the expression level of HLA-F in the secretory endometrium of patients with recurrent abortion (RPL), especially the direction of the A allele of rs2523393 SNP is associated with a better chance of pregnancy. At the same time, the functional basis of specific HLA-F single nucleotide polymorphism genotypes and diploids may lead to the increase of HLA-F mRNA and protein levels in some secretory endometrium, which may have an increased impact on embryo implantation and pregnancy development (91).
The expression of HLA-F protein suggests that immune cells are activated (91). Takanori Shobu et al. suggested that the expression of HLA-E was similar to that of HLA-F. Their expression increased from the second trimester of pregnancy to full term, which was consistent with the time of rapid fetal growth. Both HLA-E and HLA-F might work together to prepare an environment that supports fetal growth (103). However, this was only confirmed by finding that the expression trend of HLA-E and HLA-F was similar and consistent with the time of rapid fetal growth.
In conclusion, HLA-Ib can be regarded as an immunosuppressive factor. When the expression of HLA increases, the maternal immune effect on embryos will be weakened, so as to achieve the purpose of successful pregnancy. However, how HLA-Ib expression is regulated has not been completely elucidated, and the polymorphism of the HLA Class Ib genes and proteins are very low. Therefore, when HLA-Ib expressed by the fetal trophoblast cells, they are unlikely to be seen as foreign by the maternal immune system. Furthermore, it is certain that when the body has an inflammatory reaction, the level of HLA in maternal serum will be affected (19). This indicates that the expression of HLA Ib is similar to other classic HLA molecules and will be affected by the inflammatory reaction. HLA-E, F and G not only play an independent role in the establishment of maternal-fetal immune tolerance, but also the synergistic effect of the three plays an important role. Abnormal expression of anyone will lead to pregnancy failure. In future, new discoveries may rise from the interaction mechanism between HLA-F and other MHC molecules, so as to understand the mechanism of maternal-fetal immune tolerance more deeply and lay a theoretical foundation for solving more reproductive immune diseases.
Maternal-fetal immune tolerance has clinical significance for pathological conditions of pregnancy, which include recurrent spontaneous abortion (RSA), Pre-eclampsia (PE), repeated implantation failure (RIF), etc.
Several studies have reported that the differences of expression levels of HLA-G and alternative splice patterns are based on the 14-bp ins/del HLA-G polymorphism in exon 8 (the 3′UTR) (104). And the 14-bp ins HLA-G genotype associated with the low expression of HLA-G, and risk of RSA (104). However, recent studies have shown that HLA-G 14 bp insertion allele shows no significant association with RSA (45, 105, 106), deserving further research in the future.
PE is a multisystemic pregnancy disorder, which associated with the level of HLA class Ib (104). Several studies have reported significant reduced expression of HLA-G and sHLA-G in PE comparison with placentas from uncomplicated pregnancies. However, some studies have shown that the expression of HLA-G in the placenta did not observe any significant differences in expression intensity between cases of pre-eclampsia and controls (103, 104). The conflicting experimental results may arise from differences in the size, and the origin of the studied cohort, and may also be interfered by concurrent multiple diseases. In conclusion, the results remain controversial, and still need more research to figure out.
RIF is determined when transferred embryos fail to implant after several in vitro fertilization (IVF) treatment attempts (107). RIF may be due to the couple’s similarity in HLA components (107). If such a similarity is found, high-dose IV immunoglobulin should be offered before embryo transfer (108).
Based on the mechanism study of HLA class Ib molecules–HLA-E, HLA-F and HLA-G related to maternal-fetal immune tolerance, it can provide a new idea for the detection methods of in vitro fertilization-embryo transfer. At present, the research on the significance of HLA class Ib molecules for clinical embryo transfer mainly focuses on HLA-G (109, 110).
The latest research of Izabela Nowak et al. has made a great inspiration for HLA-G in the field of assisted reproduction. Their study found that the concentration of 59.73 IU/ml of sHLA-G had the best sensitivity (58.82%) and specificity (66.10%) to distinguish the successful pregnancy from the patients without pregnancy or abortion. At the same time, they concluded that the average and/or median concentration of sHLA-G after embryo transfer increased or at least remained at the level before embryo transfer (111). It suggested that the prediction of embryo transfer results could be accelerated by detecting the content of sHLA-G in maternal plasma samples, though the standard values of sHLA-G concentrations in plasma need to be supported by more experimental data. Venkatappa Vani et al. measured sHLA-G levels in embryo-spent medium (E-SM) samples and noticed that there was a positive correlation between sHLA-G levels and blastocysts’ grade scores, 60% of top-quality embryos have high level (3.86 ± 0.26ng/mL) of sHLA-G (112). Moreover, they showed a highly significant (P <.0001) association of sHLA-G with pregnancy outcome with live births, and the level of sHLA-G in E-SMs is a significant (P <.001) predictor of pregnancy outcomes (112).
Progesterone is known to be an immunomodulatory steroid hormone secreted by corpus luteum and placenta, which can maintain the endometrium and embryo implantation (113). Izabela Nowak et al. proposed that progesterone can induce the expression of HLA-G through progesterone response elements (111). Since the level of HLA-G was positively correlated with progesterone supplementation, exogenous progesterone might increase the expression of HLA-G (111). The Cochrane systematic review and the Practice Committee of the American Society of reproductive medicine also confirmed the important role of progesterone supplementation in luteal support for patients undergoing assisted reproductive technology surgery (114–116). Izabela Nowak et al. also mentioned that HLA-G was positively correlated with the supplement of corticosteroid and negatively correlated with estradiol, suggesting that the active and appropriate supplement of progesterone and corticosteroids during pregnancy, as well as the control of estradiol intake were conducive to the induction of HLA-G expression and beneficial in the maintenance and success of pregnancy (111).
Christina Bailey-Hytholt et al. innovatively invented a non-invasive cell enrichment technology. On a 50 degree inclined acrylic plate surface, they enhanced the adhesion of trophoblasts to the surface and limited the adhesion of cervical cells. They used gravity to make the cells move along the inclined plane, collecting trophoblasts in heterogeneous cervical cell populations, and identifying JEG-3 trophoblasts by HLA-G antibody staining and fluorescence imaging. It was confirmed that the purity of trophoblast in clinical cervical samples can be increased by 396 ± 52% (117). The design of the enrichment device can be used by non-professionals with low cost and high speed and can be directly integrated into the automatic cell picker instrument on the surface of the slide. The application of this technology in the detection of HLA-G content can speed up the detection of the success of embryo implantation and has a great application prospect in clinical detection.
As immune tolerance molecules expressed on the surface of trophoblast cells, HLA class Ib molecules has made a great contribution in the process of pregnancy, protecting the fetus from immune cells and maintaining normal growth. At present, the research on HLA-E and HLA-G has made the mechanisms and physiological functions clearer, but the research on HLA-F is still very few. Studies in recent years have basically confirmed its crucial role in maternal-fetal immune tolerance. However, the research on HLA-F is far from enough compared with HLA-E and HLA-G, and future experimental studies can focus on the role of HLA-F in physiological and pathological states. At the same time, some studies have proposed the synergy effects among HLA class Ib molecules, which can also be a new direction of future research. In addition, a deeper understanding of the mechanism of HLA class Ib molecules, especially HLA-G, and their expression in the extravillous trophoblast at the maternal-fetal interface can be used as a non-invasive biological index for rapid detection of embryo implantation. It can provide clues for maintaining normal pregnancy and further help to improve the success rate of embryo transplantation.
Both YY and WW contributed to the study design and manuscript preparation and revision; JW and LL supervised the manuscript and provided financial support; HL and YM helped on literature collection; WM contributed to the overall supervision. All authors read and approved the manuscript.
This work was financially supported by National Natural Science Foundation of China (No. 81901554), and Beijing Natural Science Foundation (No. 7222002).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
DICs, decidual immune cells; MHC, Major histocompatibility complex; HLA, Human leukocyte antigen; NK, nature killer; dNK, Decidual natural killer; uNK, Uterine natural killer; ICM, inner cell mass; TE, trophectoderm; CTB, cytotrophoblasts; STBs, syncytiotrophoblasts; EVTs, extravillous trophoblasts; mHLA-G, membrane-bound isoforms; sHLA-G, soluble isoforms; ILT, immunoglobulin-like transcript; DC, dendritic cells; CTL, cytotoxic T cells; IL-4 (10), interleukin-4 (10); IFN- γ, interferon- γ; TNF-α, tumor necrosis factor-α; MDSCs, Myeloid-derived suppressor cells; TCR,T cell receptor; MHC-I OC, MHC-I open conformers; RSA, recurrent spontaneous abortion; PE, Pre-eclampsia; RIF, repeated implantation failure; IVF, in vitro fertilization.
1. Heinrichs H, Orr HT. HLA non-A,B,C class I genes: Their structure and expression. Immunol Res (1990) 9(4):265–74. doi: 10.1007/BF02935526
2. Nilsson LL, Hviid TVF. HLA class ib-receptor interactions during embryo implantation and early pregnancy. Hum Reprod Update (2022) 28(3):435–54. doi: 10.1093/humupd/dmac007
3. Juch H, Blaschitz A, Dohr G, Hutter H. HLA class I expression in the human placenta. Wien Med Wochenschr (2012) 162(9-10):196–200. doi: 10.1007/s10354-012-0070-7
4. Tafuri A, Alferink J, Moller P, Hammerling GJ, Arnold B. T Cell awareness of paternal alloantigens during pregnancy. Science (1995) 270(5236):630–3. doi: 10.1126/science.270.5236.630
5. Diaz-Hernandez I, Alecsandru D, Garcia-Velasco JA, Dominguez F. Uterine natural killer cells: From foe to friend in reproduction. Hum Reprod Update (2021) 27(4):720–46. doi: 10.1093/humupd/dmaa062
6. Al-Khunaizi NR, Tabbara KS, Farid EM. Is there a role for HLA-G in the induction of regulatory T cells during the maintenance of a healthy pregnancy? Am J Reprod Immunol (2020) 84(2):e13259. doi: 10.1111/aji.13259
7. Xu L, Li Y, Sang Y, Li DJ, Du M. Crosstalk between trophoblasts and decidual immune cells: The cornerstone of maternal-fetal immunotolerance. Front Immunol (2021) 12:642392. doi: 10.3389/fimmu.2021.642392
8. Ander SE, Diamond MS, Coyne CB. Immune responses at the maternal-fetal interface. Sci Immunol (2019) 4(31):eaat6114. doi: 10.1126/sciimmunol.aat6114
9. Bakleicheva MO, Bespalova ON, Ivashchenko TE. Role of class I HLA (G, e, and c) expression in early reproductive losses. Akusherstvo i ginekologiya (2020) 2020(2):30–6. doi: 10.18565/aig.2020.2.30-36
10. Liu Y, Fan X, Wang R, Lu X, Dang YL, Wang H, et al. Single-cell RNA-seq reveals the diversity of trophoblast subtypes and patterns of differentiation in the human placenta. Cell Res (2018) 28(8):819–32. doi: 10.1038/s41422-018-0066-y
11. Vento-Tormo R, Efremova M, Botting RA, Turco MY, Vento-Tormo M, Meyer KB, et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature (2018) 563(7731):347–53. doi: 10.1038/s41586-018-0698-6
12. Lin XX, Xie YM, Zhao SJ, Liu CY, Mor G, Liao AH. Human leukocyte antigens: The unique expression in trophoblasts and their crosstalk with local immune cells. Int J Biol Sci (2022) 18(10):4043–52. doi: 10.7150/ijbs.73616
13. Bhalla A, Stone PR, Liddell HS, Zanderigo A, Chamley LW. Comparison of the expression of human leukocyte antigen (HLA)-G and HLA-e in women with normal pregnancy and those with recurrent miscarriage. Reproduction (2006) 131(3):583–9. doi: 10.1530/rep.1.00892
14. Hackmon R, Pinnaduwage L, Zhang J, Lye SJ, Geraghty DE, Dunk CE. Definitive class I human leukocyte antigen expression in gestational placentation: HLA-f, HLA-e, HLA-c, and HLA-G in extravillous trophoblast invasion on placentation, pregnancy, and parturition. Am J Reprod Immunol (2017) 77(6):e12643. doi: 10.1111/aji.12643
15. Ishitani A, Sageshima N, Lee N, Dorofeeva N, Hatake K, Marquardt H, et al. Protein expression and peptide binding suggest unique and interacting functional roles for HLA-e, f, and G in maternal-placental immune recognition. J Immunol (2003) 171(3):1376–84. doi: 10.4049/jimmunol.171.3.1376
16. Nagamatsu T, Fujii T, Matsumoto J, Yamashita T, Kozuma S, Taketani Y. Human leukocyte antigen f protein is expressed in the extra-villous trophoblasts but not on the cell surface of them. Am J Reprod Immunol (2006) 56(3):172–7. doi: 10.1111/j.1600-0897.2006.00414.x
17. Dahl M, Hviid TV. Human leucocyte antigen class ib molecules in pregnancy success and early pregnancy loss. Hum Reprod Update (2012) 18(1):92–109. doi: 10.1093/humupd/dmr043
18. Proll J, Bensussan A, Goffin F, Foidart JM, Berrebi A, Le Bouteiller P. Tubal versus uterine placentation: Similar HLA-G expressing extravillous cytotrophoblast invasion but different maternal leukocyte recruitment. Tissue Antigens (2000) 56(6):479–91. doi: 10.1034/j.1399-0039.2000.560601.x
19. Dunk CE, Bucher M, Zhang J, Hayder H, Geraghty DE, Lye SJ, et al. Human leukocyte antigen HLA-c, HLA-G, HLA-f and HLA-e placental profiles are altered in early severe preeclampsia and preterm birth with chorioamnionitis. Am J Obstet Gynecol (2022). doi: 10.1016/j.ajog.2022.07.021
20. Park GM, Lee S, Park B, Kim E, Shin J, Cho K, et al. Soluble HLA-G generated by proteolytic shedding inhibits NK-mediated cell lysis. Biochem Biophys Res Commun (2004) 313(3):606–11. doi: 10.1016/j.bbrc.2003.11.153
21. Gonen-Gross T, Achdout H, Arnon TI, Gazit R, Stern N, Horejsí V, et al. The CD85J/leukocyte inhibitory receptor-1 distinguishes between conformed and beta 2-microglobulin-free HLA-G molecules. J Immunol (2005) 175(8):4866–74. doi: 10.4049/jimmunol.175.8.4866
22. Nardi Fda S, König L, Wagner B, Giebel B, Santos Manvailer LF, Rebmann V. Soluble monomers, dimers and HLA-g-expressing extracellular vesicles: The three dimensions of structural complexity to use HLA-G as a clinical biomarker. Hla (2016) 88(3):77–86. doi: 10.1111/tan.12844
23. Yie SM, Li LH, Li YM, Librach C. HLA-G protein concentrations in maternal serum and placental tissue are decreased in preeclampsia. Am J Obstet Gynecol (2004) 191(2):525–9. doi: 10.1016/j.ajog.2004.01.033
24. Persson G, Staehr CS, Klok FS, Lebech M, Hviid TVF. Evidence for a shift in placental HLA-G allelic dominance and the HLA-G isoform profile during a healthy pregnancy and preeclampsiadagger. Biol Reprod (2021) 105(4):846–58. doi: 10.1093/biolre/ioab121
25. Gu C, Park S, Seok J, Jang HY, Bang YJ, Kim GIJ. Altered expression of ADM and ADM2 by hypoxia regulates migration of trophoblast and HLA-G expressiondagger. Biol Reprod (2021) 104(1):159–69. doi: 10.1093/biolre/ioaa178
26. LeMaoult J, Zafaranloo K, Le Danff C, Carosella ED. HLA-G up-regulates ILT2, ILT3, ILT4, and KIR2DL4 in antigen presenting cells, NK cells, and T cells. FASEB J (2005) 19(6):662–4. doi: 10.1096/fj.04-1617fje
27. Kostlin N, Ostermeir AL, Spring B, Schwarz J, Marme A, Walter CB, et al. HLA-G promotes myeloid-derived suppressor cell accumulation and suppressive activity during human pregnancy through engagement of the receptor ILT4. Eur J Immunol (2017) 47(2):374–84. doi: 10.1002/eji.201646564
28. Favier B, Lemaoult J, Lesport E, Carosella ED. ILT2/HLA-G interaction impairs NK-cell functions through the inhibition of the late but not the early events of the NK-cell activating synapse. FASEB J (2010) 24(3):689–99. doi: 10.1096/fj.09-135194
29. Kniotek M, Roszczyk A, Zych M, Szafarowska M, Jerzak M. Differences in the expression of KIR, ILT inhibitory receptors, and VEGF production in the induced decidual NK cell cultures of fertile and RPL women. BioMed Res Int (2021) 2021:6673427. doi: 10.1155/2021/6673427
30. Clements CS, Kjer-Nielsen L, Kostenko L, Hoare HL, Dunstone MA, Moses E, et al. Crystal structure of HLA-G: A nonclassical MHC class I molecule expressed at the fetal-maternal interface. Proc Natl Acad Sci U S A (2005) 102(9):3360–5. doi: 10.1073/pnas.0409676102
31. Pende D, Sivori S, Accame L, Pareti L, Falco M, Geraghty D, et al. HLA-G recognition by human natural killer cells. Involvement of CD94 both as inhibitory and as activating receptor complex. Eur J Immunol (1997) 27(8):1875–80. doi: 10.1002/eji.1830270809
32. Zhuang B, Shang J, Yao Y. HLA-G: An important mediator of maternal-fetal immune-tolerance. Front Immunol (2021) 12:744324. doi: 10.3389/fimmu.2021.744324
33. Wang P, Greenland JR. HLA-G/ILT2 signaling on the path to tolerance. J Heart Lung Transplant (2022) 41(6):852–3. doi: 10.1016/j.healun.2022.03.004
34. Yan WH, Fan LA. Residues Met76 and Gln79 in HLA-G alpha1 domain involve in KIR2DL4 recognition. Cell Res (2005) 15(3):176–82. doi: 10.1038/sj.cr.7290283
35. Morandi F, Ferretti E, Castriconi R, Dondero A, Petretto A, Bottino C, et al. Soluble HLA-G dampens CD94/NKG2A expression and function and differentially modulates chemotaxis and cytokine and chemokine secretion in CD56bright and CD56dim NK cells. Blood (2011) 118(22):5840–50. doi: 10.1182/blood-2011-05-352393
36. Navarro F, Llano M, Bellón T, Colonna M, Geraghty DE, López-Botet M. The ILT2(LIR1) and CD94/NKG2A NK cell receptors respectively recognize HLA-G1 and HLA-e molecules co-expressed on target cells. Eur J Immunol (1999) 29(1):277–83. doi: 10.1002/(SICI)1521-4141(199901)29:01<277::AID-IMMU277>3.0.CO;2-4
37. Faure M, Long EO. KIR2DL4 (CD158d), an NK cell-activating receptor with inhibitory potential. J Immunol (2002) 168(12):6208–14. doi: 10.4049/jimmunol.168.12.6208
38. Zheng G, Guo Z, Li W, Xi W, Zuo B, Zhang R, et al. Interaction between HLA-G and NK cell receptor KIR2DL4 orchestrates HER2-positive breast cancer resistance to trastuzumab. Signal Transduct Target Ther (2021) 6(1):236. doi: 10.1038/s41392-021-00629-w
39. Reed J, Reichelt M, Wetzel SA. Lymphocytes and trogocytosis-mediated signaling. Cells (2021) 10(6):1478. doi: 10.3390/cells10061478
40. Djurisic S, Skibsted L, Hviid TV. A phenotypic analysis of regulatory T cells and uterine NK cells from first trimester pregnancies and associations with HLA-G. Am J Reprod Immunol (2015) 74(5):427–44. doi: 10.1111/aji.12421
41. Koopman LA, Kopcow HD, Rybalov B, Boyson JE, Orange JS, Schatz F, et al. Human decidual natural killer cells are a unique NK cell subset with immunomodulatory potential. J Exp Med (2003) 198(8):1201–12. doi: 10.1084/jem.20030305
42. Weiss E, Andrade HS, Lara JR, Souza AS, Paz MA, Lima THA, et al. KIR2DL4 genetic diversity in a Brazilian population sample: implications for transcription regulation and protein diversity in samples with different ancestry backgrounds. Immunogenetics (2021) 73(3):227–41. doi: 10.1007/s00251-021-01206-9
43. Sun JY, Wu R, Xu J, Xue HY, Lu XJ, Ji J. Placental immune tolerance and organ transplantation: Underlying interconnections and clinical implications. Front Immunol (2021) 12:705950. doi: 10.3389/fimmu.2021.705950
44. Bespalova O, Bakleicheva M, Ivashchenko T, Tral T, Tolibova G, Kogan I. Expression of HLA-G and KIR2DL4 receptor in chorionic villous in missed abortion. Gynecol Endocrinol (2020) 36(sup1):43–7. doi: 10.1080/09513590.2020.1816716
45. Rouas-Freiss N, Moreau P, LeMaoult J, Papp B, Tronik-Le Roux D, Carosella ED. Role of the HLA-G immune checkpoint molecule in pregnancy. Hum Immunol (2021) 82(5):353–61. doi: 10.1016/j.humimm.2021.01.003
46. Narang K, Cheek EH, Enninga EAL, Theiler RN. Placental immune responses to viruses: Molecular and histo-pathologic perspectives. Int J Mol Sci (2021) 22(6):2921. doi: 10.3390/ijms22062921
47. Chen QY, Chen YX, Han QY, Zhang JG, Zhou WJ, Zhang X, et al. Prognostic significance of immune checkpoints HLA-G/ILT-2/4 and PD-L1 in colorectal cancer. Front Immunol (2021) 12:679090. doi: 10.3389/fimmu.2021.679090
48. Jabrane-Ferrat N, Siewiera J. The up side of decidual natural killer cells: New developments in immunology of pregnancy. Immunology (2014) 141(4):490–7. doi: 10.1111/imm.12218
49. Ponte M, Cantoni C, Biassoni R, Tradori-Cappai A, Bentivoglio G, Vitale C, et al. Inhibitory receptors sensing HLA-G1 molecules in pregnancy: Decidua-associated natural killer cells express LIR-1 and CD94/NKG2A and acquire p49, an HLA-G1-specific receptor. Proc Natl Acad Sci U S A (1999) 96(10):5674–9. doi: 10.1073/pnas.96.10.5674
50. Naji A, Durrbach A, Carosella ED, Rouas-Freiss N. Soluble HLA-G and HLA-G1 expressing antigen-presenting cells inhibit T-cell alloproliferation through ILT-2/ILT-4/FasL-mediated pathways. Hum Immunol (2007) 68(4):233–9. doi: 10.1016/j.humimm.2006.10.017
51. Shiroishi M, Tsumoto K, Amano K, Shirakihara Y, Colonna M, Braud VM, et al. Human inhibitory receptors ig-like transcript 2 (ILT2) and ILT4 compete with CD8 for MHC class I binding and bind preferentially to HLA-G. Proc Natl Acad Sci U S A (2003) 100(15):8856–61. doi: 10.1073/pnas.1431057100
52. Fournel S, Aguerre-Girr M, Huc X, Lenfant F, Alam A, Toubert A, et al. Cutting edge: soluble HLA-G1 triggers CD95/CD95 ligand-mediated apoptosis in activated CD8+ cells by interacting with CD8. J Immunol (2000) 164(12):6100–4. doi: 10.4049/jimmunol.164.12.6100
53. Wang W, Zhao Y, Zhou X, Sung N, Chen L, Zhang X, et al. Dynamic changes in regulatory T cells during normal pregnancy, recurrent pregnancy loss, and gestational diabetes. J Reprod Immunol (2022) 150:103492. doi: 10.1016/j.jri.2022.103492
54. Guerin LR, Prins JR, Robertson SA. Regulatory T-cells and immune tolerance in pregnancy: A new target for infertility treatment? Hum Reprod Update (2009) 15(5):517–35. doi: 10.1093/humupd/dmp004
55. Carosella ED, Moreau P, Aractingi S, Rouas-Freiss N. HLA-G: A shield against inflammatory aggression. Trends Immunol (2001) 22(10):553–5. doi: 10.1016/S1471-4906(01)02007-5
56. Yousefzadeh Y, Soltani-Zangbar MS, Hemmatzadeh M, Shomali N, Mahmoodpoor A, Ahmadian Heris J, et al. Fetomaternal immune tolerance: Crucial mechanisms of tolerance for successful pregnancy in humans. Immunol Invest (2022) 51(4):1108–25. doi: 10.1080/08820139.2021.1909061
57. Tripathi P, Naik S, Agrawal S. HLA-e and immunobiology of pregnancy. Tissue Antigens (2006) 67(3):207–13. doi: 10.1111/j.1399-0039.2005.00550.x
58. Petrie EJ, Clements CS, Lin J, Sullivan LC, Johnson D, Huyton T, et al. CD94-NKG2A recognition of human leukocyte antigen (HLA)-e bound to an HLA class I leader sequence. J Exp Med (2008) 205(3):725–35. doi: 10.1084/jem.20072525
59. Shreeve N, Depierreux D, Hawkes D, Traherne JA, Sovio U, Huhn O, et al. The CD94/NKG2A inhibitory receptor educates uterine NK cells to optimize pregnancy outcomes in humans and mice. Immunity (2021) 54(6):1231–44 e4. doi: 10.1016/j.immuni.2021.03.021
60. Prasnikar E, Perdih A, Borisek J. All-atom simulations reveal a key interaction network in the HLA-E/NKG2A/CD94 immune complex fine-tuned by the nonameric peptide. J Chem Inf Model (2021) 61(7):3593–603. doi: 10.1021/acs.jcim.1c00414
61. Pietra G, Romagnani C, Moretta L, Mingari MC. HLA-e and HLA-e-bound peptides: recognition by subsets of NK and T cells. Curr Pharm Des (2009) 15(28):3336–44. doi: 10.2174/138161209789105207
62. Pereira BI, Devine OP, Vukmanovic-Stejic M, Chambers ES, Subramanian P, Patel N, et al. Senescent cells evade immune clearance via HLA-e-mediated NK and CD8(+) T cell inhibition. Nat Commun (2019) 10(1):2387. doi: 10.1038/s41467-019-10335-5
63. Masilamani M, Nguyen C, Kabat J, Borrego F, Coligan JE. CD94/NKG2A inhibits NK cell activation by disrupting the actin network at the immunological synapse. J Immunol (2006) 177(6):3590–6. doi: 10.4049/jimmunol.177.6.3590
64. Vales-Gomez M, Reyburn HT, Erskine RA, Lopez-Botet M, Strominger JL. Kinetics and peptide dependency of the binding of the inhibitory NK receptor CD94/NKG2-a and the activating receptor CD94/NKG2-c to HLA-e. EMBO J (1999) 18(15):4250–60. doi: 10.1093/emboj/18.15.4250
65. Braud VM, Allan DS, O'Callaghan CA, Soderstrom K, D'Andrea A, Ogg GS, et al. HLA-e binds to natural killer cell receptors CD94/NKG2A, b and c. Nature (1998) 391(6669):795–9. doi: 10.1038/35869
66. Goodridge JP, Burian A, Lee N, Geraghty DE. HLA-f and MHC class I open conformers are ligands for NK cell ig-like receptors. J Immunol (2013) 191(7):3553–62. doi: 10.4049/jimmunol.1300081
67. Schmitt C, Marie-Cardine A, Bensussan A. Therapeutic antibodies to KIR3DL2 and other target antigens on cutaneous T-cell lymphomas. Front Immunol (2017) 8:1010. doi: 10.3389/fimmu.2017.01010
68. Geraghty D, Goodridge J, Burian A, Lee N. HLA-f and MHC-I open conformers are ligands for natural killer cell receptor KIR3DL2. J Immunol (2013) 190:3553–62. doi: 10.4049/jimmunol.1300081
69. Lepin EJ, Bastin JM, Allan DS, Roncador G, Braud VM, Mason DY, et al. Functional characterization of HLA-f and binding of HLA-f tetramers to ILT2 and ILT4 receptors. Eur J Immunol (2000) 30(12):3552–61. doi: 10.1002/1521-4141(200012)30:12<3552::AID-IMMU3552>3.0.CO;2-L
70. Colonna M, Samaridis J, Cella M, Angman L, Allen RL, O'Callaghan CA, et al. Human myelomonocytic cells express an inhibitory receptor for classical and nonclassical MHC class I molecules. J Immunol (1998) 160(7):3096–100.
71. Lombardelli L, Aguerre-Girr M, Logiodice F, Kullolli O, Casart Y, Polgar B, et al. HLA-G5 induces IL-4 secretion critical for successful pregnancy through differential expression of ILT2 receptor on decidual CD4(+) T cells and macrophages. J Immunol (2013) 191(7):3651–62. doi: 10.4049/jimmunol.1300567
72. Lunemann S, Schobel A, Kah J, Fittje P, Holzemer A, Langeneckert AE, et al. Interactions between KIR3DS1 and HLA-f activate natural killer cells to control HCV replication in cell culture. Gastroenterology (2018) 155(5):1366–71.e3. doi: 10.1053/j.gastro.2018.07.019
73. Burian A, Wang KL, Finton KA, Lee N, Ishitani A, Strong RK, et al. HLA-f and MHC-I open conformers bind natural killer cell ig-like receptor KIR3DS1. PloS One (2016) 11(9):e0163297. doi: 10.1371/journal.pone.0163297
74. Garcia-Beltran WF, Holzemer A, Martrus G, Chung AW, Pacheco Y, Simoneau CR, et al. Open conformers of HLA-f are high-affinity ligands of the activating NK-cell receptor KIR3DS1. Nat Immunol (2016) 17(9):1067–74. doi: 10.1038/ni.3513
75. Giuliani M, Giron-Michel J, Negrini S, Vacca P, Durali D, Caignard A, et al. Generation of a novel regulatory NK cell subset from peripheral blood CD34+ progenitors promoted by membrane-bound IL-15. PloS One (2008) 3(5):e2241. doi: 10.1371/journal.pone.0002241
76. Brown T. Observations by immunofluorescence microscopy and electron microscopy on the cytopathogenicity of naegleria fowleri in mouse embryo-cell cultures. J Med Microbiol (1979) 12(3):363–71. doi: 10.1099/00222615-12-3-363
77. Tilburgs T, Evans JH, Crespo ÂC, Strominger JL. The HLA-G cycle provides for both NK tolerance and immunity at the maternal-fetal interface. Proc Natl Acad Sci U S A (2015) 112(43):13312–7. doi: 10.1073/pnas.1517724112
78. Hsu P, Santner-Nanan B, Joung S, Peek MJ, Nanan R. Expansion of CD4(+) HLA-g(+) T cell in human pregnancy is impaired in pre-eclampsia. Am J Reprod Immunol (2014) 71(3):217–28. doi: 10.1111/aji.12195
79. LeMaoult J, Caumartin J, Daouya M, Favier B, Le Rond S, Gonzalez A, et al. Immune regulation by pretenders: Cell-to-cell transfers of HLA-G make effector T cells act as regulatory cells. Blood (2007) 109(5):2040–8. doi: 10.1182/blood-2006-05-024547
80. HoWangYin KY, Alegre E, Daouya M, Favier B, Carosella ED, LeMaoult J. Different functional outcomes of intercellular membrane transfers to monocytes and T cells. Cell Mol Life Sci (2010) 67(7):1133–45. doi: 10.1007/s00018-009-0239-4
81. Gornalusse GG, Hirata RK, Funk SE, Riolobos L, Lopes VS, Manske G, et al. HLA-e-expressing pluripotent stem cells escape allogeneic responses and lysis by NK cells. Nat Biotechnol (2017) 35(8):765–72. doi: 10.1038/nbt.3860
82. Wyatt RC, Lanzoni G, Russell MA, Gerling I, Richardson SJ. What the HLA-I!-Classical and non-classical HLA class I and their potential roles in type 1 diabetes. Curr Diabetes Rep (2019) 19(12):159. doi: 10.1007/s11892-019-1245-z
83. Xu QH, Liu H, Wang LL, Zhu Q, Zhang YJ, Muyayalo KP, et al. Roles of gammadeltaT cells in pregnancy and pregnancy-related complications. Am J Reprod Immunol (2021) 86(5):e13487. doi: 10.1111/aji.13487
84. Moretta A, Marcenaro E, Parolini S, Ferlazzo G, Moretta L. NK cells at the interface between innate and adaptive immunity. Cell Death Differ (2008) 15(2):226–33. doi: 10.1038/sj.cdd.4402170
85. Duygu B, Olieslagers TI, Groeneweg M, Voorter CEM, Wieten L. HLA class I molecules as immune checkpoints for NK cell alloreactivity and anti-viral immunity in kidney transplantation. Front Immunol (2021) 12:680480. doi: 10.3389/fimmu.2021.680480
86. Prasnikar E, Perdih A, Borisek J. Nonameric peptide orchestrates signal transduction in the activating HLA-E/NKG2C/CD94 immune complex as revealed by all-atom simulations. Int J Mol Sci (2021) 22(13):6670. doi: 10.3390/ijms22136670
87. Zhang X, Feng J, Chen S, Yang H, Dong Z. Synergized regulation of NK cell education by NKG2A and specific Ly49 family members. Nat Commun (2019) 10(1):5010. doi: 10.1038/s41467-019-13032-5
88. Morandi F, Pistoia V. Interactions between HLA-G and HLA-e in physiological and pathological conditions. Front Immunol (2014) 5:394. doi: 10.3389/fimmu.2014.00394
89. Geraghty DE, Wei XH, Orr HT, Koller BH. Human leukocyte antigen f (HLA-f). an expressed HLA gene composed of a class I coding sequence linked to a novel transcribed repetitive element. J Exp Med (1990) 171(1):1–18. doi: 10.1084/jem.171.1.1
90. Papuchova H, Saxtorph MH, Hallager T, Jepsen IE, Eriksen JO, Persson G, et al. Endometrial HLA-f expression is influenced by genotypes and correlates differently with immune cell infiltration in IVF and recurrent implantation failure patients. Hum Reprod (2022). doi: 10.1093/humrep/deac118
91. Langkilde CH, Nilsson LL, Jorgensen N, Funck T, Perin TL, Hornstrup MB, et al. Variation in the HLA-f gene locus with functional impact is associated with pregnancy success and time-to-pregnancy after fertility treatment. Hum Reprod (2020) 35(3):705–17. doi: 10.1093/humrep/dez276
92. Lee N, Ishitani A, Geraghty DE. HLA-f is a surface marker on activated lymphocytes. Eur J Immunol (2010) 40(8):2308–18. doi: 10.1002/eji.201040348
93. Djurisic S, Hviid TVF. HLA class lb molecules and immune cells in pregnancy and preeclampsia. Front Immunol (2014) 5:652. doi: 10.3389/fimmu.2014.00652
94. Yip L, McCluskey J, Sinclair R. Immunological aspects of pregnancy. Clinics Dermatol (2006) 24(2):84–7. doi: 10.1016/j.clindermatol.2005.10.022
95. Chen M, Yan J, Han Q, Luo J, Zhang Q. Identification of hub-methylated differentially expressed genes in patients with gestational diabetes mellitus by multi-omic WGCNA basing epigenome-wide and transcriptome-wide profiling. J Cell Biochem (2020) 121(5-6):3173–84. doi: 10.1002/jcb.29584
96. Lundberg M, Krogvold L, Kuric E, Dahl-Jorgensen K, Skog O. Expression of interferon-stimulated genes in insulitic pancreatic islets of patients recently diagnosed with type 1 diabetes. Diabetes (2016) 65(10):3104–10. doi: 10.2337/db16-0616
97. Mando C, Pileri P, Mazzocco MI, Lattuada D, Zolin A, Plebani M, et al. Maternal and fetal HLA-G 14 bp gene polymorphism in pregnancy-induced hypertension, preeclampsia, intrauterine growth restricted and normal pregnancies. J Matern Fetal Neonatal Med (2016) 29(9):1509–14. doi: 10.3109/14767058.2015.1052398
98. Nishizawa A, Kumada K, Tateno K, Wagata M, Saito S, Katsuoka F, et al. Analysis of HLA-G long-read genomic sequences in mother-offspring pairs with preeclampsia. Sci Rep (2020) 10(1):20027. doi: 10.1038/s41598-020-77081-3
99. Sommese L, Benincasa G, Schiano C, Marfella R, Grimaldi V, Sorriento A, et al. Genetic and epigenetic-sensitive regulatory network in immune response: A putative link between HLA-G and diabetes. Expert Rev Endocrinol Metab (2019) 14(4):233–41. doi: 10.1080/17446651.2019.1620103
100. Aluvihare VR, Kallikourdis M, Betz AG. Regulatory T cells mediate maternal tolerance to the fetus. Nat Immunol (2004) 5(3):266–71. doi: 10.1038/ni1037
101. Burrows CK, Kosova G, Herman C, Patterson K, Hartmann KE, Velez Edwards DR, et al. Expression quantitative trait locus mapping studies in mid-secretory phase endometrial cells identifies HLA-f and TAP2 as fecundability-associated genes. PloS Genet (2016) 12(7):e1005858. doi: 10.1371/journal.pgen.1005858
102. Bulmer JN, Williams PJ, Lash GE. Immune cells in the placental bed. Int J Dev Biol (2010) 54(2-3):281–94. doi: 10.1387/ijdb.082763jb
103. Shobu T, Sageshima N, Tokui H, Omura M, Saito K, Nagatsuka Y, et al. The surface expression of HLA-f on decidual trophoblasts increases from mid to term gestation. J Reprod Immunol (2006) 72(1-2):18–32. doi: 10.1016/j.jri.2006.02.001
104. Persson G, Melsted WN, Nilsson LL, Hviid TVF. HLA class ib in pregnancy and pregnancy-related disorders. Immunogenetics (2017) 69(8-9):581–95. doi: 10.1007/s00251-017-0988-4
105. de Almeida BS, Muniz YCN, Prompt AH, Castelli EC, Mendes-Junior CT, Donadi EA. Genetic association between HLA-G 14-bp polymorphism and diseases: A systematic review and meta-analysis. Hum Immunol (2018) 79(10):724–35. doi: 10.1016/j.humimm.2018.08.003
106. Meuleman T, Lashley LE, Dekkers OM, van Lith JM, Claas FH, Bloemenkamp KW. HLA associations and HLA sharing in recurrent miscarriage: A systematic review and meta-analysis. Hum Immunol (2015) 76(5):362–73. doi: 10.1016/j.humimm.2015.02.004
107. Simon A, Laufer N. Repeated implantation failure: clinical approach. Fertil Steril (2012) 97(5):1039–43. doi: 10.1016/j.fertnstert.2012.03.010
108. Elram T, Simon A, Israel S, Revel A, Shveiky D, Laufer N. Treatment of recurrent IVF failure and human leukocyte antigen similarity by intravenous immunoglobulin. Reprod BioMed Online (2005) 11(6):745–9. doi: 10.1016/S1472-6483(10)61694-X
109. Kofod L, Lindhard A, Bzorek M, Eriksen JO, Larsen LG, Hviid TVF. Endometrial immune markers are potential predictors of normal fertility and pregnancy after in vitro fertilization. Am J Reprod Immunol (2017) 78(3):e12684. doi: 10.1111/aji.12684
110. Rizzo R, Andersen AS, Lassen MR, Sorensen HC, Bergholt T, Larsen MH, et al. Soluble human leukocyte antigen-G isoforms in maternal plasma in early and late pregnancy. Am J Reprod Immunol (2009) 62(5):320–38. doi: 10.1111/j.1600-0897.2009.00742.x
111. Nowak I, Wilczynska K, Radwan P, Wisniewski A, Krasinski R, Radwan M, et al. Association of soluble HLA-G plasma level and HLA-G genetic polymorphism with pregnancy outcome of patients undergoing in vitro fertilization embryo transfer. Front Immunol (2019) 10:2982. doi: 10.3389/fimmu.2019.02982
112. Vani V, Vasan SS, Adiga SK, Varsha SR, Sachdeva G, Kumar P, et al. Soluble human leukocyte antigen-G is a potential embryo viability biomarker and a positive predictor of live-births in humans. Am J Reprod Immunol (2021) 86(6):e13499. doi: 10.1111/aji.13499
113. Mesiano S. Progesterone - historical perspective. J Steroid Biochem Mol Biol (2022) 25:106157. doi: 10.1016/j.jsbmb.2022.106157
114. Czyzyk A, Podfigurna A, Genazzani AR, Meczekalski B. The role of progesterone therapy in early pregnancy: From physiological role to therapeutic utility. Gynecol Endocrinol (2017) 33(6):421–4. doi: 10.1080/09513590.2017.1291615
115. Practice Committee of the American Society for Reproductive M. Current clinical irrelevance of luteal phase deficiency: A committee opinion. Fertil Steril (2015) 103(4):e27–32. doi: 10.1016/j.fertnstert.2014.12.128
116. Di Renzo GC, Giardina I, Clerici G, Brillo E, Gerli S. Progesterone in normal and pathological pregnancy. Horm Mol Biol Clin Investig (2016) 27(1):35–48. doi: 10.1515/hmbci-2016-0038
Keywords: HLA class Ib, HLA-G, HLA-E, HLA-F, pregnancy, maternal-fetal immune tolerance, decidual immune cells
Citation: Yang Y, Wang W, Weng J, Li H, Ma Y, Liu L and Ma W (2022) Advances in the study of HLA class Ib in maternal-fetal immune tolerance. Front. Immunol. 13:976289. doi: 10.3389/fimmu.2022.976289
Received: 23 June 2022; Accepted: 09 August 2022;
Published: 29 August 2022.
Edited by:
Yin Tailang, Faculty of Medical Sciences, Wuhan University, ChinaReviewed by:
Xiuxiu Xu, University of Science and Technology of China, ChinaCopyright © 2022 Yang, Wang, Weng, Li, Ma, Liu and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Weng, d2VuZ2ppbmdAY2NtdS5lZHUuY24=; Lingyan Liu, TGluZ3lhbkBjY211LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.