
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Immunol. , 19 July 2022
Sec. Vaccines and Molecular Therapeutics
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.946318
This article is part of the Research Topic Advancing the Understanding of Emergence of SARS-CoV-2 Genetic Variants and COVID-19 Vaccine Efficacy: Essential Clinical and Molecular Insights and Breakthroughs View all 39 articles
 Iris Medits1
Iris Medits1 David N. Springer1
David N. Springer1 Marianne Graninger1
Marianne Graninger1 Jeremy V. Camp1Eva Höltl2
Jeremy V. Camp1Eva Höltl2 Stephan W. Aberle1
Stephan W. Aberle1 Marianna T. Traugott3Wolfgang Hoepler3Josef Deutsch4Oliver Lammel5Christian Borsodi1
Marianna T. Traugott3Wolfgang Hoepler3Josef Deutsch4Oliver Lammel5Christian Borsodi1 Elisabeth Puchhammer-Stöckl1Alexander Zoufaly3
Elisabeth Puchhammer-Stöckl1Alexander Zoufaly3 Lukas Weseslindtner1
Lukas Weseslindtner1 Judith H. Aberle1†
Judith H. Aberle1† Karin Stiasny1*†
Karin Stiasny1*†Background and Methods: The SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2) Omicron (B.1.1.529) variant is the antigenically most distinct variant to date. As the heavily mutated spike protein enables neutralization escape, we studied serum-neutralizing activities of naïve and vaccinated individuals after Omicron BA.1 or BA.2 sub-lineage infections in live virus neutralization tests with Omicron BA.1, Omicron BA.2, wildtype (WT, B1.1), and Delta (B.1.617.2) strains. Serum samples obtained after WT infections and three-dose mRNA vaccinations with and without prior infection were included as controls.
Results: Primary BA.1 infections yielded reduced neutralizing antibody levels against WT, Delta, and Omicron BA.2, while samples from BA.2-infected individuals showed almost no cross-neutralization against the other variants. Serum neutralization of Omicron BA.1 and BA.2 variants was detectable after three-dose mRNA vaccinations, but with reduced titers. Vaccination-breakthrough infections with either Omicron BA.1 or BA.2, however, generated equal cross-neutralizing antibody levels against all SARS-CoV-2 variants tested.
Conclusions: Our study demonstrates that although Omicron variants are able to enhance cross-neutralizing antibody levels in pre-immune individuals, primary infections with BA.1 or BA.2 induced mostly variant-specific neutralizing antibodies, emphasizing the differently shaped humoral immunity induced by the two Omicron variants. These data thus contribute substantially to the understanding of antibody responses induced by primary Omicron infections or multiple exposures to different SARS-CoV-2 variants and are of particular importance for developing vaccination strategies in the light of future emerging variants.
The SARS-CoV-2 (severe acute respiratory syndrome coronavirus) Omicron (B.1.1.529) variant of concern (VOC) is now prevalent in large parts of the world. It has been divided into several lineages [BA.1 to BA.5, their descendants and BA.1/BA.2 recombinant forms1 (1)], which are characterized by a heavily mutated spike protein, leading to substantial escape from antibodies induced by previous infections and/or vaccinations [reviewed in (2–5)]. The rapid increase in Omicron BA.2 infections, which has recently replaced BA.1 as the dominant variant2 (6), indicates that it is more transmissible than BA.1 (7) and/or may escape antibody-mediated immunity, potentially including the protection gained from Omicron BA.1 infections (8, 9).
The data reported so far yielded a heterogeneous picture with respect to the degree of cross-neutralization between Omicron and pre-Omicron variants, with most studies using samples from pre-immune, i.e. vaccinated and/or convalescent, individuals (10–17). Here, we determined neutralizing antibody titers in serum samples collected after primary as well as vaccination-breakthrough infections with Omicron variants BA.1 or BA.2. Using live virus assays with an ancestral wildtype (WT) strain and three VOCs (Delta, Omicron BA.1, Omicron BA.2), we found that infections with Omicron variants boostered cross-neutralizing antibodies in pre-immune individuals. Primary infections with one of the Omicron sub-lineages, however, induced mainly variant-specific neutralizing antibodies; particularly BA.2 infections generated a sub-lineage-specific neutralization pattern.
Serum samples were collected from non-vaccinated patients hospitalized after a SARS-CoV-2 WT infection between March and November 2020 (before the emergence of VOCs) (Table S1). Samples from non-vaccinated individuals with primary SARS-CoV-2 Omicron infections were collected at primary health-care centers between January and April 2022, and identification of SARS-CoV-2 infection was based on PCR testing. Omicron-infected patients developed various symptoms including fever, cough, headache, rhinitis, sore throat, muscle or body aches, fatigue, diarrhea, and hoarseness. According to the family physicians, none of the patients suffered from an underlying immune deficiency, and no particular pre-existing conditions were reported. Samples from vaccinated and/or SARS-CoV-2 infected individuals were sent to the diagnostic laboratory of the Center for Virology, Medical University of Vienna, Austria. Individuals were regularly tested by SARS-CoV-2 PCR and/or lateral flow assays under the COVID-19-mass-testing program in Austria, which excluded additional SARS-CoV-2 infections.
Vero E6 cells (ECACC #85020206) were from the European Collection of Authenticated Cell Cultures (ECACC), and VeroE6-TMPRSS2 cells were kindly provided by Anna Ohradanova-Repic. Both cell lines were cultured in Dulbecco’s Modified Eagle Medium (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) containing 10% Fetal Bovine Serum (Capricorn Scientific GmbH, Ebsdorfergrund, Germany) and 1% Penicillin-Streptomycin-Glutamine (Gibco, Thermo Fisher Scientific) at 37°C and 5% CO2. The cell lines were tested negative for mycoplasma contamination by the MycoAlertTM Mycoplasma Detection Kit (Lonza Group Ltd, Basel, Switzerland) in regular intervals.
SARS-CoV-2 strains were isolated from nasopharyngeal swabs from COVID-19 patients using either Vero E6 cells (WT D614G and Delta) or VeroE6-TMPRSS2 cells (both Omicron variants) (18–20). The sequences of the strains were determined by next generation sequencing and uploaded to the GISAID database (WT, B.1.1 with the D614G mutation: EPI_ISL_438123; Delta, B.1.617.2-like, sub-lineage AY.122: EPI_ISL_4172121; Omicron, B.1.1.529+BA.*, sub-lineage BA.1.17: EPI_ISL_9110894; Omicron, B.1.1.529+BA.*, sub-lineage BA.2.9: EPI_ISL_11110193. Pango lineages were determined with Pango v.4.0.6, Pango-data v1.6.) (18–20).
Identification of Omicron BA.1 and BA.2 variants was performed with nasopharyngeal swabs obtained from 19 patients (n=8, primary Omicron infection; n=11, Omicron vaccination breakthrough infections) using the mutation assay VirSNiP SARS-CoV-2 Spike S371L S373P (TIB MOLBIOL, Berlin, Germany). Characteristic melting peaks for the mutations S371LS373P and S371FS373P indicated an infection with Omicron BA.1 and BA.2, respectively.
The live virus neutralization test (NT) was performed as described previously (18–20). Two-fold serial dilutions of heat-inactivated serum samples were incubated with 50–100 TCID50 SARS-CoV-2 for one hour at 37°C before the mixtures were added to Vero E6 cells. After three to five days at 37°C, NT titers were expressed as the reciprocal of the serum dilution required for prevention of virus-induced cytopathic effects (CPE), which was assessed by microscopy and validated by two different operators. At least one negative and two positive human polyclonal samples were included as controls in each assay. NT titers of serum samples ≥10 were considered positive.
Statistical analysis was performed with GraphPad Prism 9.3.1. The Mann-Whitney test was used for pairwise comparisons. The Kruskal-Wallis test followed by Dunn´s multiple comparison was used for analyzing four groups. P values < 0.05 were considered significant.
We analyzed the neutralizing capacity of serum samples obtained after primary Omicron BA.1 and BA.2 infections as well as after mRNA-vaccination-breakthrough infections with both Omicron variants against a WT strain (isolated early in the pandemic with the D614G mutation) and the three variants of concern Delta, Omicron BA.1, and Omicron BA.2 in live virus neutralization assays (see Methods). As controls, we included samples from hospitalized patients obtained after primary WT infections as well as from individuals after three doses of an mRNA vaccine with and without prior SARS-CoV-2 infection. The characteristics of these cohorts are summarized in Table 1 and Tables S1, S2.
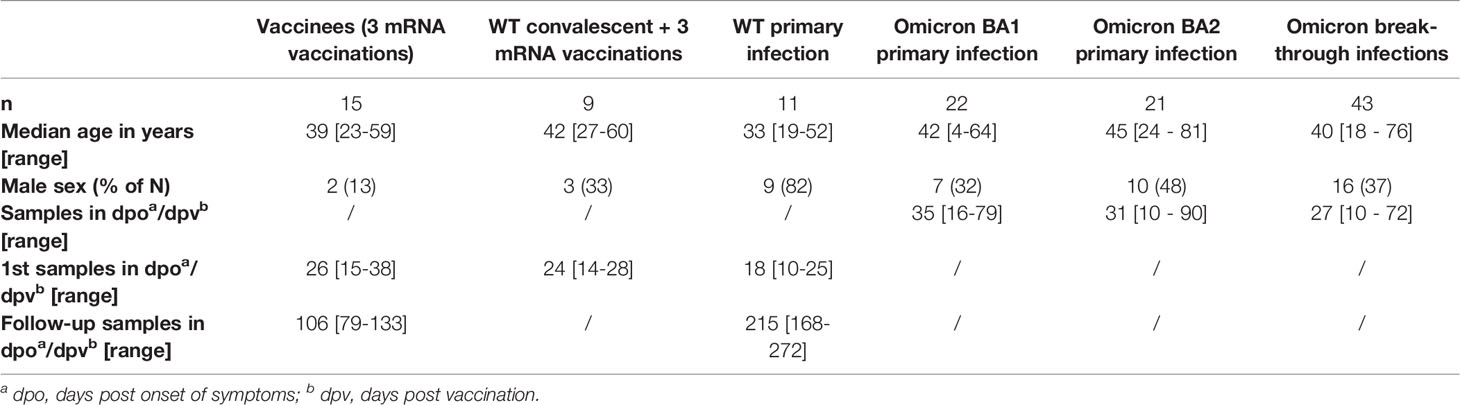
Table 1 Demographics of study cohorts and sampling time points.
Serum samples from vaccinees collected one and three months after the third dose of an mRNA vaccine efficiently cross-neutralized the two Omicron variants (Figure 1A and Table 2). Neutralization titers were significantly lower than for WT (Figure 1A and Table 2), but there was no significant difference between BA.1 and BA.2 neutralization (Mann-Whitney test, p > 0.05). We also detected Omicron cross-neutralization in serum samples from individuals who had a WT infection before being vaccinated three times (Figure 1B and Table 2).
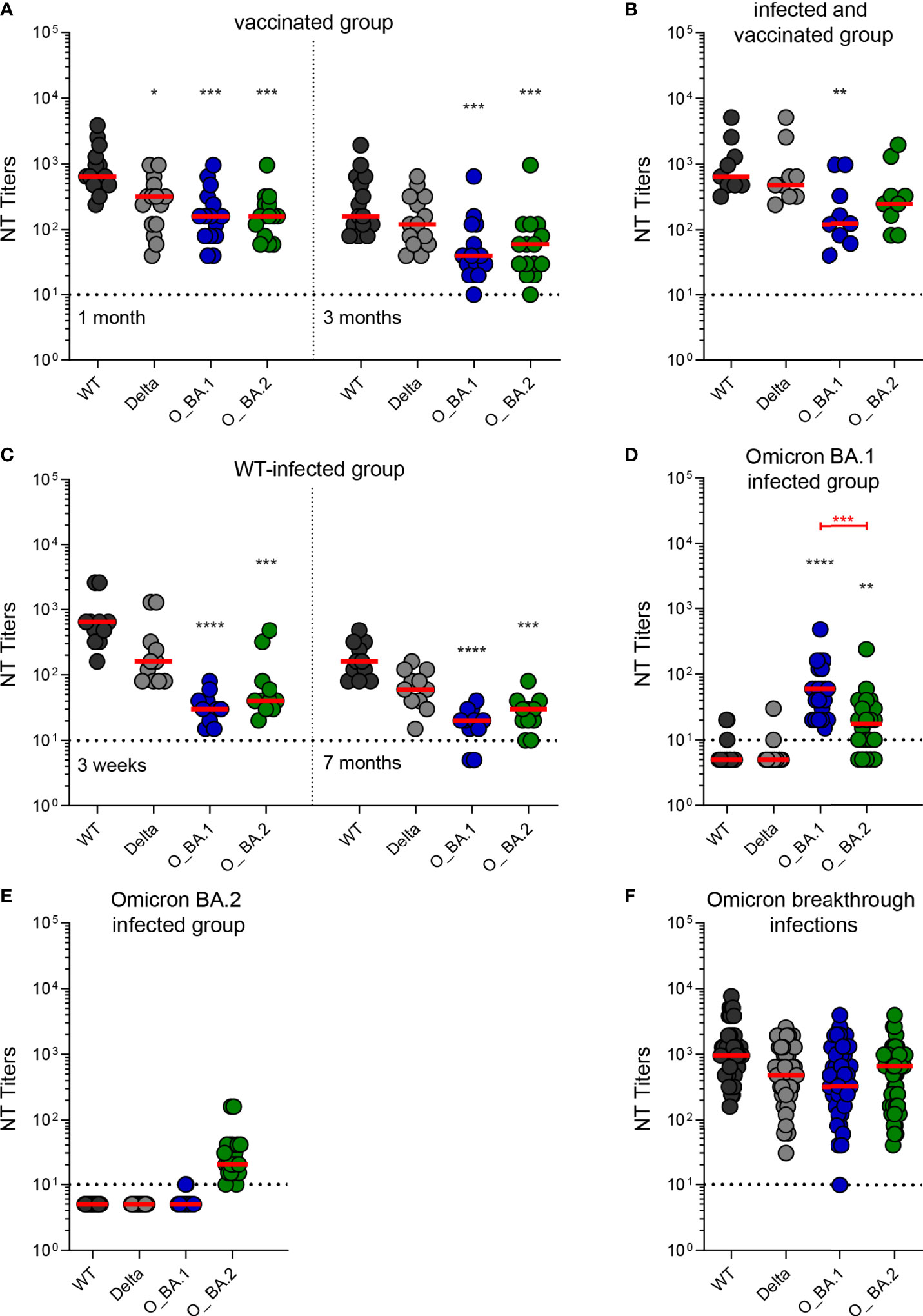
Figure 1 Neutralizing antibody titers against an ancestral wildtype (WT) strain and three variants of concern (Delta, Omicron BA.1, and Omicron BA.2) in post-vaccination and post-infection serum samples. (A) Serum samples from individuals who had received three doses of an mRNA vaccine collected one month and three months after the last vaccination (n=15). (B) Serum samples from individuals who were infected with a WT strain, followed by three doses of an mRNA vaccine collected three weeks after the last vaccination (n=9). (C) Serum samples from individuals who were infected with a WT strain collected three weeks and seven months after infection (n=11). (D) Serum samples obtained from individuals who had a primary infection with Omicron BA.1 collected one month after infection (n=22). (E) Serum samples obtained from individuals who had a primary infection with Omicron BA.2 collected one month after infection (n=21). (F) Serum samples from individuals who were infected with an Omicron variant and had previously been vaccinated collected one month after infection (Omicron breakthrough infections, n=43). Horizontal dotted lines show the cut-off, dots individual sera, and red lines median titers. Black asterisks indicate significant differences to WT (D614G). The Kruskal-Wallis test with Dunn’s post-hoc test was used for significance testing (*, p < 0.05; **, p < 0.01; ***, p <0.001; ****, p < 0.0001). Significant differences between Omicron BA.1 and BA.2 neutralization titers are indicated by red asterisks (Mann-Whitney test; ***, p < 0.001). WT, wildtype strain (B.1.1) with the D614G substitution; Delta, Delta VOC (B.1.617.2-like, sub-lineage AY.122); O_BA.1, Omicron (B.1.1.529) sub-lineage BA.1 VOC; O_BA.2, Omicron sub-lineage BA.2 VOC; NT, neutralization test.
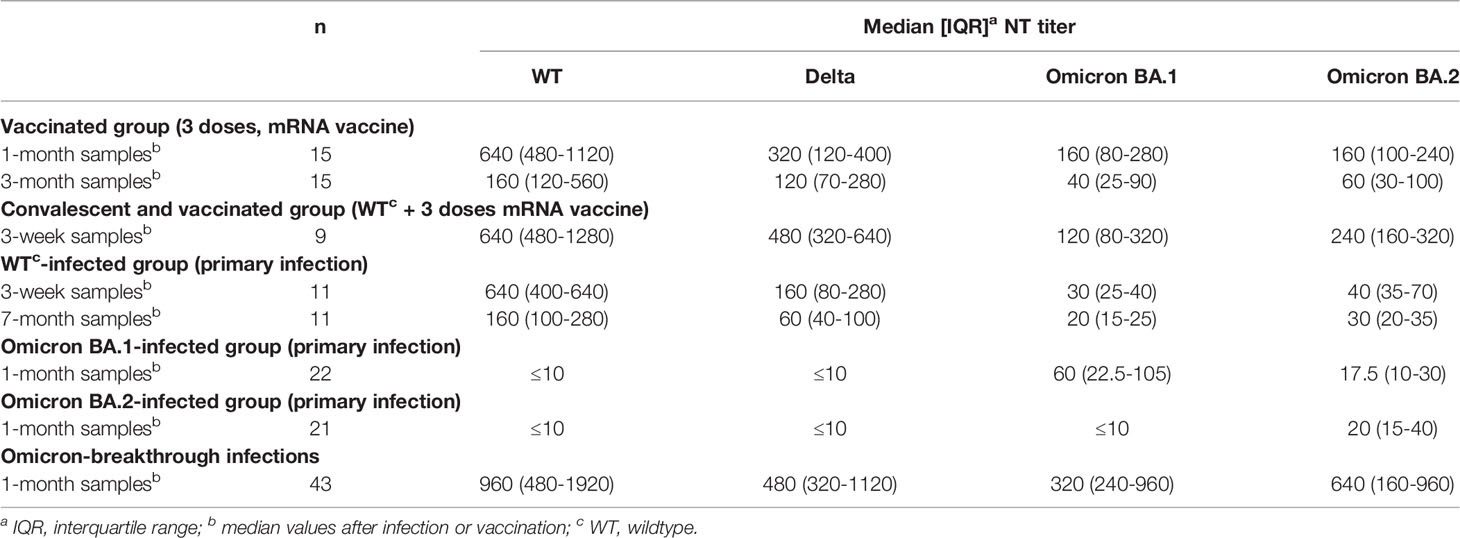
Table 2 Serum-neutralization titers of cohorts analyzed in this study.
Samples from individuals after primary infections with WT, Omicron BA.1, or Omicron BA.2 neutralized the heterologous strains to much lesser extents than the homologous virus (Figures 1C–E and Table 2). Neutralization titers against Omicron BA.1 and BA.2 variants were significantly reduced in samples obtained three weeks and seven months after infection with an ancestral WT strain (Figure 1C). Omicron BA.1-convalescents developed only very low levels of WT- and Delta VOC-neutralizing antibodies; 18/22 samples were below the cut-off (Figure 1D). Even BA.2 neutralization titers were significantly lower in sera of this group (Figure 1D). In contrast, samples from Omicron BA.2 convalescents exhibited almost no cross-neutralization of any other virus strain tested (Figure 1E). However, all samples from individuals after an Omicron infection who had been previously vaccinated (Omicron-breakthrough infection) were able to neutralize the Omicron BA.1 and BA.2 sub-lineages as efficiently as the WT and Delta viruses (Figure 1F and Table 2). Stratification of the Omicron-breakthrough infections according to the infecting sub-lineage revealed similar neutralization patterns of sera after Omicron BA.1 or BA.2 infection (Figure S1).
In this study, we provide a direct comparison of (cross)-neutralizing antibody responses after primary Omicron BA.1 and BA.2 infections. Consistent with the antigenic distance between Omicron BA.1 and BA.2 VOCs from pre-Omicron strains as well as from each other (8, 9, 16), we show that neutralizing antibodies present in serum samples from patients with primary Omicron infection are highly variant specific. While samples from individuals after primary BA.1 infection exhibited some cross-neutralization of BA.2 (Figure 1D), as reported by others (17, 21), samples from primary BA.2-infected individuals showed almost no cross-neutralizing activities against any other variant tested (Figure 1E). Similar results were obtained in neutralization assays performed with Omicron BA.1 and BA.2 post-infection serum samples from hamsters, with the highest levels of neutralizing antibodies against the homologous strain (22). In agreement with our results, BA.1 post-infection samples yielded stronger cross-neutralization of BA.2 than did BA.2 post-infection sera with BA.1.
The mutations in the Omicron spike proteins strongly affect their antigenicity in comparison to pre-Omicron variants (23, 24), as indicated by a substantial resistance to neutralizing antibodies induced by prior SARS-CoV-2 infections and/or vaccinations (Figures 1A–C) [reviewed in (2–5)]. The considerable antigenic differences between the Omicron BA.1 and BA.2 spikes in the receptor-binding domain (RBD) and the N-terminal domain (23, 24) (as illustrated in Figure 2), the major targets of neutralizing antibodies, offer an explanation for the sub-lineage-specific neutralization profiles observed after primary BA.1 or BA.2 infections (Figures 1D, E). Consistent with our data, primary Omicron infections elicited RBD-specific B cells with only narrow specificity for these variants (14).
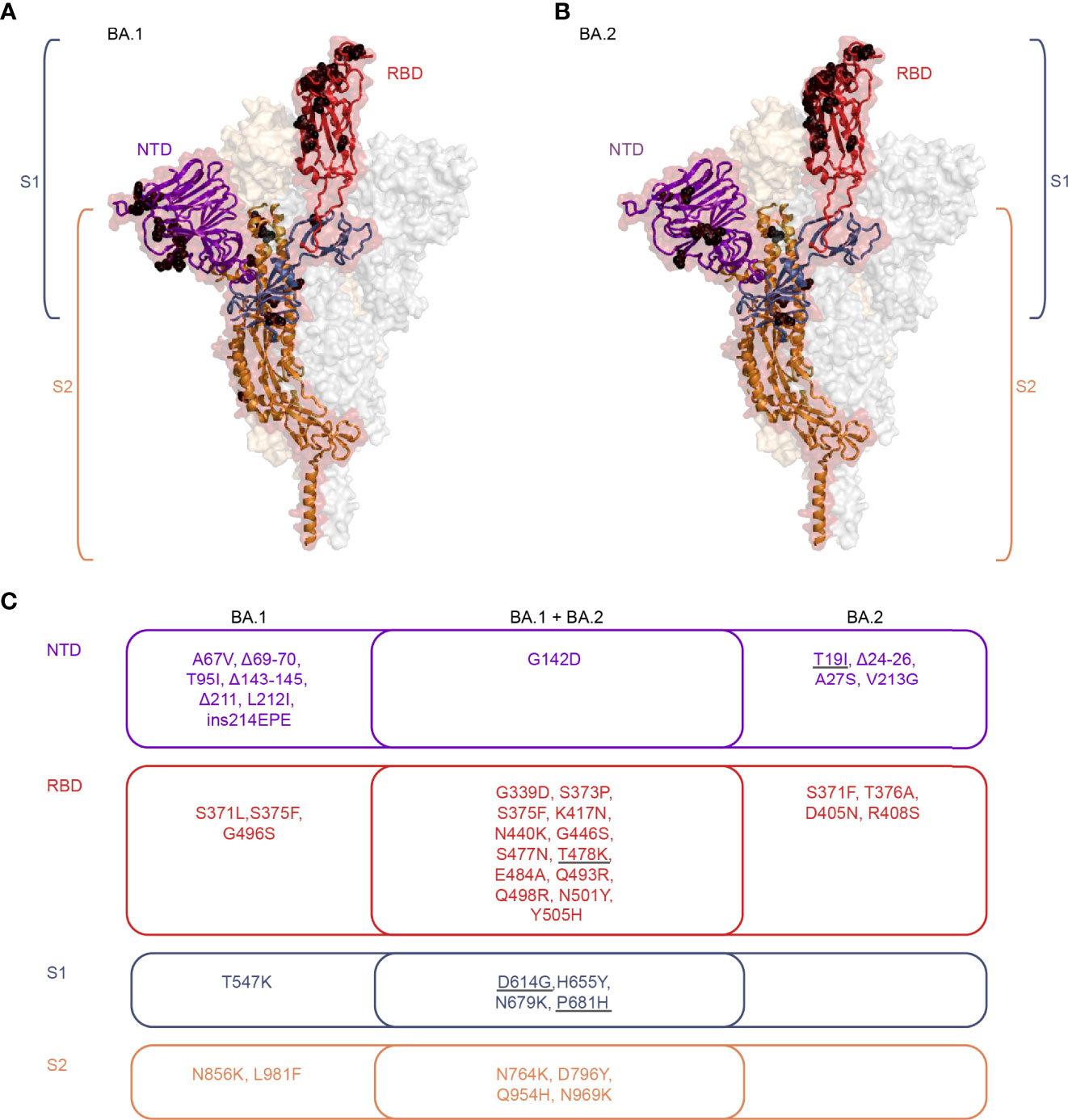
Figure 2 Structural representation of mutations present in Omicron BA.1 and BA.2 sub-lineages. (A, B) Cartoon representation of one monomer of the spike protein (side view) combined with a semi-transparent surface representation of the trimeric spike in its RBD-up-conformation [PDB 7KRR, (25)]. The three protomers are colored in red, grey, and bright orange. The S1 subunit is colored blue in the single protomer, with the RBD highlighted in red and the NTD highlighted in purple. S2 is colored in orange. The black spheres indicate the mutations present in sub-lineages BA.1 (A) and BA.2 (B). The structures were generated with PyMol (https://pymol.org). (C) Venn diagram illustrating the BA.1 and BA.2 sub-lineage-specific and shared mutations, colored according to the regions highlighted in panels A and B. The grey underlines indicate mutations present in the Delta variant. RBD, Receptor-binding domain; NTD, N-terminal domain; S1, Subunit 1; S2, Subunit 2.
Multiple exposures to pre-Omicron-SARS-CoV-2, via three mRNA vaccinations (Figure 1A) or mixed infection-vaccination scenarios (Figure 1B), led to high titers with efficient neutralization of both Omicron variants. These data are in agreement with other studies that suggested an increase in the magnitude as well as the breadth of neutralizing antibody responses by repeated exposure to the original antigen (26–35). Previous data (36) suggest that a three-dose vaccination regimen enhances the breadth of antibody responses in that memory B cells are also formed against conserved epitopes. These are typically of lower immunogenicity than the more strain-specific and immunodominant epitopes, which is why repeated exposures may be needed for broader variant neutralization.
The Omicron NT titers, however, were lower in these cohorts than against pre-Omicron variants (Figures 1A, B) and a fast waning of neutralizing antibodies was observed (Figure 1A). In contrast, vaccination-breakthrough infections with Omicron induced similarly high levels of neutralizing antibodies against all variants tested (Figure 1F) (9, 13, 14, 37), thus leading to a strong expansion of neutralization breadth. Apparently, an adequate number of conserved epitopes for neutralizing antibodies are present in the spike proteins of Omicron and pre-Omicron variants, suggesting that Omicron-adapted vaccines might increase the effectiveness of booster immunizations.
A limitation of our study is the relatively small sample size. However, due to increasing SARS-CoV-2 immunity in the human population, it is difficult to find unvaccinated persons who have only been exposed to a single Omicron variant, which is why previous studies were performed with hamster post-Omicron infection samples (8, 22, 38). In addition, we focused on the neutralizing activities of antibodies, but also interactions with Fc receptors might be involved in protection independent of the neutralizing potency of antibodies (39).
In summary, the highly variant-specific neutralization profiles obtained with the two Omicron sub-lineages BA.1 and BA.2 in naïve individuals underscore the antigenic distance between the two variants. The rapid emergence of further immune-escape variants (Omicron BA.4 and BA.5), which have been associated with a resurgence of SARS-CoV-2 infections in South Africa (40, 41), highlights the importance of global variant surveillance and antigenic characterization to estimate the impact of the continuing SARS-CoV-2 evolution on the ongoing pandemic (42).
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The analyses were approved by the Ethics Committee of the Medical University of Vienna (EK 1291/2021, EK 1926/2020, EK 2156/2019, EK 1035/2016, EK 1513/2016). Hospitalized patients provided written informed consent to participate in this study. Written informed consent was not required for the analysis of anonymized leftover samples from routine laboratory diagnosis in accordance with the national legislation and the institutional requirements.
Conceptualization, KS and JA. Investigation, KS, JA, IM, JC, and SA. Formal analysis, KS, JA, and IM. Resources, DS, MG, EH, MT, JD, OL, CB, EP-S, LW, and AZ. Writing - original draft, KS, JA, and IM. Writing - review & editing, all authors. Visualization, IM. Supervision, KS. Funding acquisition, JA and IM. All authors contributed to the article and approved the submitted version.
IM acknowledges funding by the “Hochschuljubiläumsfonds der Stadt Wien” (Project H-334533/2021) and JA by the Medical-scientific fund of the Mayor of the federal capital Vienna [Grant Covid003].
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Jutta Hutecek, Silvia Schwödiauer, Cornelia Hell, Sylvia Malik, Barbara Dalmatiner and Michaela Binder for excellent technical assistance. A preliminary version of this manuscript has been posted at Research Square (https://doi.org/10.21203/rs.3.rs-1536794/v1) (43).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.946318/full#supplementary-material
1. World Health Organization. Tracking Sars-Cov-2 Variants (2022). Available at: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants (Accessed May 6, 2022).
2. Fan Y, Li X, Zhang L, Wan S, Zhang L, Zhou F. Sars-Cov-2 Omicron Variant: Recent Progress and Future Perspectives. Signal Transduct Target Ther (2022) 7(1):141. doi: 10.1038/s41392-022-00997-x
3. Flemming A. Omicron, the Great Escape Artist. Nat Rev Immunol (2022) 22(2):75. doi: 10.1038/s41577-022-00676-6
4. Jung C, Kmiec D, Koepke L, Zech F, Jacob T, Sparrer KMJ, et al. Omicron: What Makes the Latest Sars-Cov-2 Variant of Concern So Concerning? J Virol (2022) 96(6):e0207721. doi: 10.1128/jvi.02077-21
5. Simon-Loriere E, Schwartz O. Towards Sars-Cov-2 Serotypes? Nat Rev Microbiol (2022) 20(4):187–8. doi: 10.1038/s41579-022-00708-x
6. World Health Organization. Weekly Epidemiological Update on Covid-19 - 22 March 2022 (2022). Available at: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19—22-march-2022 (Accessed May 6, 2022).
7. Lyngse FP, Kirkeby CT, Denwood M, Christiansen LE, Mølbak K, Møller CH, et al. Transmission of Sars-Cov-2 Omicron Voc Subvariants Ba.1 and Ba.2: Evidence From Danish Households. medRxiv (2022), 2022.01.28.22270044. doi: 10.1101/2022.01.28.22270044
8. Mykytyn AZ, Rissmann M, Kok A, Rosu ME, Schipper D, Breugem TI, et al. Omicron Ba.1 and Ba.2 Are Antigenically Distinct Sars-Cov-2 Variants. bioRxiv (2022) eabq4450. doi: 10.1126/sciimmunol.abq4450
9. Yamasoba D, Kimura I, Nasser H, Morioka Y, Nao N, Ito J, et al. Virological Characteristics of the Sars-Cov-2 Omicron Ba.2 Spike. Cell (2022) 185(12):2103–15 e19. doi: 10.1016/j.cell.2022.04.035
10. Arora P, Zhang L, Rocha C, Sidarovich A, Kempf A, Schulz S, et al. Comparable Neutralisation Evasion of Sars-Cov-2 Omicron Subvariants Ba.1, Ba.2, and Ba.3. Lancet Infect Dis (2022) 22(6):766–7. doi: 10.1016/S1473-3099(22)00224-9
11. Chen LL, Chu AW, Zhang RR, Hung IF, To KK. Serum Neutralisation of the Sars-Cov-2 Omicron Sublineage Ba.2. Lancet Microbe (2022) 3(6):e404. doi: 10.1016/S2666-5247(22)00060-X
12. Cheng SM, Mok CKP, Chan KC, Ng SS, Lam BH, Luk LL, et al. Sars-Cov-2 Omicron Variant Ba.2 Neutralisation in Sera of People With Comirnaty or Coronavac Vaccination, Infection or Breakthrough Infection, Hong Kong, 2020 to 2022. Euro Surveill (2022) 27(18):2200178. doi: 10.2807/1560-7917.ES.2022.27.18.2200178
13. Evans JP, Zeng C, Qu P, Faraone J, Zheng YM, Carlin C, et al. Neutralization of Sars-Cov-2 Omicron Sub-Lineages Ba.1, Ba.1.1, and Ba.2. Cell Host Microbe (2022). doi: 10.1016/j.chom.2022.04.014
14. Park Y-J, Pinto D, Walls AC, Liu Z, De Marco A, Benigni F, et al. Imprinted Antibody Responses Against Sars-Cov-2 Omicron Sublineages. bioRxiv (2022), 2022.05.08.491108. doi: 10.1101/2022.05.08.491108
15. Rossler A, Knabl L, von Laer D, Kimpel J. Neutralization Profile After Recovery From Sars-Cov-2 Omicron Infection. N Engl J Med (2022) 386(18):1764–6. doi: 10.1056/NEJMc2201607
16. Rössler A, Netzl A, Knabl L, Schäfer H, Wilks SH, Bante D, et al. Ba.2 Omicron Differs Immunologically From Both Ba.1 Omicron and Pre-Omicron Variants. medRxiv (2022), 2022.05.10.22274906. doi: 10.1101/2022.05.10.22274906
17. Zou J, Kurhade C, Xia H, Liu M, Xie X, Ren P, et al. Cross-Neutralization of Omicron Ba.1 Against Ba.2 and Ba.3 Sars-Cov-2. Nat Commun (2022) 13(1):2956. doi: 10.1038/s41467-022-30580-5
18. Graninger M, Camp JV, Aberle SW, Traugott MT, Hoepler W, Puchhammer-Stöckl E, et al. Heterogeneous Sars-Cov-2-Neutralizing Activities After Infection and Vaccination. Front Immunol (2022) 13:888794. doi: 10.3389/fimmu.2022.888794
19. Koblischke M, Traugott MT, Medits I, Spitzer FS, Zoufaly A, Weseslindtner L, et al. Dynamics of Cd4 T Cell and Antibody Responses in Covid-19 Patients With Different Disease Severity. Front Med (Lausanne) (2020) 7:592629(771). doi: 10.3389/fmed.2020.592629
20. Sieber J, Mayer M, Schmidthaler K, Kopanja S, Camp JV, Popovitsch A, et al. Long-Lived Immunity in Sars-Cov-2-Recovered Children and Its Neutralizing Capacity Against Omicron. Front Immunol (2022) 13:882456. doi: 10.3389/fimmu.2022.882456
21. Richardson SI, Madzorera VS, Spencer H, Manamela NP, van der Mescht MA, Lambson BE, et al. Sars-Cov-2 Omicron Triggers Cross-Reactive Neutralization and Fc Effector Functions in Previously Vaccinated, But Not Unvaccinated, Individuals. Cell Host Microbe (2022) 30(6):880–6 e4. doi: 10.1016/j.chom.2022.03.029
22. Uraki R, Kiso M, Iida S, Imai M, Takashita E, Kuroda M, et al. Characterization and Antiviral Susceptibility of Sars-Cov-2 Omicron/Ba.2. Nature (2022). doi: 10.1038/s41586-022-04856-1
23. Javanmardi K, Segall-Shapiro TH, Chou C-W, Boutz DR, Olsen RJ, Xie X, et al. Antibody Escape and Cryptic Cross-Domain Stabilization in the Sars-Cov-2 Omicron Spike Protein. bioRxiv (2022), 2022.04.18.488614. doi: 10.1101/2022.04.18.488614
24. Zhang J, Tang W, Gao H, Lavine CL, Shi W, Peng H, et al. Structural and Functional Characteristics of Sars-Cov-2 Omicron Subvariant Ba.2 Spike. bioRxiv (2022), 2022.04.28.489772. doi: 10.1101/2022.04.28.489772
25. Zhang J, Cai Y, Xiao T, Lu J, Peng H, Sterling SM, et al. Structural Impact on Sars-Cov-2 Spike Protein by D614g Substitution. Science (2021) 372(6541):525–30. doi: 10.1126/science.abf2303
26. Belik M, Jalkanen P, Lundberg R, Reinholm A, Laine L, Vaisanen E, et al. Comparative Analysis of Covid-19 Vaccine Responses and Third Booster Dose-Induced Neutralizing Antibodies Against Delta and Omicron Variants. Nat Commun (2022) 13(1):2476. doi: 10.1038/s41467-022-30162-5
27. Bowen JE, Sprouse KR, Walls AC, Mazzitelli IG, Logue JK, Franko NM, et al. Omicron Ba.1 and Ba.2 Neutralizing Activity Elicited by a Comprehensive Panel of Human Vaccines. bioRxiv (2022). doi: 10.1101/2022.03.15.484542
28. Chen Y, Tong P, Whiteman N, Moghaddam AS, Zarghami M, Zuiani A, et al. Immune Recall Improves Antibody Durability and Breadth to Sars-Cov-2 Variants. Sci Immunol (2022):eabp8328. doi: 10.1126/sciimmunol.abp8328
29. Gruell H, Vanshylla K, Tober-Lau P, Hillus D, Schommers P, Lehmann C, et al. mRNA Booster Immunization Elicits Potent Neutralizing Serum Activity Against the Sars-Cov-2 Omicron Variant. Nat Med (2022) 28(3):477–80. doi: 10.1038/s41591-021-01676-0
30. Lusvarghi S, Pollett SD, Neerukonda SN, Wang W, Wang R, Vassell R, et al. Sars-Cov-2 Ba.1 Variant Is Neutralized by Vaccine Booster-Elicited Serum But Evades Most Convalescent Serum and Therapeutic Antibodies. Sci Transl Med (2022) 14(645):eabn8543. doi: 10.1126/scitranslmed.abn8543
31. Pajon R, Doria-Rose NA, Shen X, Schmidt SD, O'Dell S, McDanal C, et al. Sars-Cov-2 Omicron Variant Neutralization After mRNA-1273 Booster Vaccination. N Engl J Med (2022) 386(11):1088–91. doi: 10.1056/NEJMc2119912
32. Rossler A, Riepler L, Bante D, von Laer D, Kimpel J. Sars-Cov-2 Omicron Variant Neutralization in Serum From Vaccinated and Convalescent Persons. N Engl J Med (2022) 386(7):698–700. doi: 10.1056/NEJMc2119236
33. Wratil PR, Stern M, Priller A, Willmann A, Almanzar G, Vogel E, et al. Three Exposures to the Spike Protein of Sars-Cov-2 by Either Infection or Vaccination Elicit Superior Neutralizing Immunity to All Variants of Concern. Nat Med (2022) 28(3):496–503. doi: 10.1038/s41591-022-01715-4
34. Yu J, Collier AY, Rowe M, Mardas F, Ventura JD, Wan H, et al. Neutralization of the Sars-Cov-2 Omicron Ba.1 and Ba.2 Variants. N Engl J Med (2022) 386(16):1579–80. doi: 10.1056/NEJMc2201849
35. Walls AC, Sprouse KR, Bowen JE, Joshi A, Franko N, Navarro MJ, et al. Sars-Cov-2 Breakthrough Infections Elicit Potent, Broad, and Durable Neutralizing Antibody Responses. Cell (2022) 185(5):872–80.e3. doi: 10.1016/j.cell.2022.01.011
36. Muecksch F, Wang Z, Cho A, Gaebler C, Ben Tanfous T, DaSilva J, et al. Increased Memory B Cell Potency and Breadth After a Sars-Cov-2 mRNA Boost. Nature (2022). doi: 10.1038/s41586-022-04778-y
37. Ai J, Wang X, He X, Zhao X, Zhang Y, Jiang Y, et al. Antibody Evasion of Sars-Cov-2 Omicron Ba.1, Ba.1.1, Ba.2 and Ba.3 Sub-Lineages. Cell Host & Microbe (2022) doi: 10.1016/j.chom.2022.05.001
38. Willett BJ, Kurshan A, Thakur N, Newman J, Manali M, Tyson G, et al. Distinct Antigenic Properties of the Sars-Cov-2 Omicron Lineages Ba.4 and Ba.5. bioRxiv (2022), 2022.05.25.493397. doi: 10.1101/2022.05.25.493397
39. Bartsch YC, Tong X, Kang J, Avendano MJ, Serrano EF, Garcia-Salum T, et al. Omicron Variant Spike-Specific Antibody Binding and Fc Activity Are Preserved in Recipients of mRNA or Inactivated Covid-19 Vaccines. Sci Transl Med (2022) 14(642):eabn9243. doi: 10.1126/scitranslmed.abn9243
40. Khan K, Karim F, Ganga Y, Bernstein M, Jule Z, Reedoy K, et al. Omicron Sub-Lineages Ba.4/Ba.5 Escape Ba.1 Infection Elicited Neutralizing Immunity. medRxiv (2022), 2022.04.29.22274477. doi: 10.1101/2022.04.29.22274477
41. Tegally H, Moir M, Everatt J, Giovanetti M, Scheepers C, Wilkinson E, et al. Continued Emergence and Evolution of Omicron in South Africa: New Ba.4 and Ba.5 Lineages. medRxiv (2022), 2022.05.01.22274406. doi: 10.1101/2022.05.01.22274406
42. DeGrace MM, Ghedin E, Frieman MB, Krammer F, Grifoni A, Alisoltani A, et al. Defining the Risk of Sars-Cov-2 Variants on Immune Protection. Nature (2022) 605(7911):640–52. doi: 10.1038/s41586-022-04690-5
Keywords: SARS-CoV-2, SARS-CoV-2 variant of concern, Omicron sub-lineages, SARS-CoV-2 neutralization, immune escape
Citation: Medits I, Springer DN, Graninger M, Camp JV, Höltl E, Aberle SW, Traugott MT, Hoepler W, Deutsch J, Lammel O, Borsodi C, Puchhammer-Stöckl E, Zoufaly A, Weseslindtner L, Aberle JH and Stiasny K (2022) Different Neutralization Profiles After Primary SARS-CoV-2 Omicron BA.1 and BA.2 Infections. Front. Immunol. 13:946318. doi: 10.3389/fimmu.2022.946318
Received: 17 May 2022; Accepted: 16 June 2022;
Published: 19 July 2022.
Edited by:
Madhusudhanan Narasimhan, University of Texas Southwestern Medical Center, United StatesReviewed by:
Gunnveig Grødeland, University of Oslo, NorwayCopyright © 2022 Medits, Springer, Graninger, Camp, Höltl, Aberle, Traugott, Hoepler, Deutsch, Lammel, Borsodi, Puchhammer-Stöckl, Zoufaly, Weseslindtner, Aberle and Stiasny. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karin Stiasny, a2FyaW4uc3RpYXNueUBtZWR1bml3aWVuLmFjLmF0
†These authors have contributed equally to this work and share last authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.