 Marton Kaique de Andrade Cavalcante1,2
Marton Kaique de Andrade Cavalcante1,2 Rafael de Freitas e Silva1,3
Rafael de Freitas e Silva1,3 Valéria Rêgo Alves Pereira1
Valéria Rêgo Alves Pereira1 Maria Carolina Accioly Brelaz-de-Castro1,2*
Maria Carolina Accioly Brelaz-de-Castro1,2*- 1Department of Immunology, Aggeu Magalhães Institute - Oswaldo Cruz Foundation, Recife, Brazil
- 2Parasitology Laboratory, Federal University of Pernambuco, Vitoria de Santo Antão, Brazil
- 3Department of Natural Sciences, University of Pernambuco, Garanhuns, Brazil
Introduction: CL and the Early Immune Response
Cutaneous Leishmaniasis (CL) is considered a neglected disease, mainly linked to low socioeconomic conditions (1, 2). Currently, estimates suggest 900,000 to 1.5 million new cases per year with 95% of the cases in the Americas, Mediterranean and Asia (3).
The disease clinical manifestation can range from a single to multiple lesions, being observed in 90% of cases a nodular ulcerative squamous lesion. After inoculation of the metacyclic promastigote of Leishmania into the host skin by the sandfly bite, a papule forms at the site of bite, and later it turns into an ulcerated lesion with delineated borders, a reddish background, and an intense inflammatory infiltration (lymphocytes, phagocytes, and plasma cells) (4–9).. Factors such as the species of Leishmania involved in the infection and the host immune response directly influence the lesion type and clinical outcome in the patient (10, 11).
After dermis inoculation the promastigotes cause activation of the complement system and factor C3b deposition on the parasite surface. However it has been demonstrated that Leishmania protease GP63 is able to inactivate C3b (12). These promastigotes are opsonized, by the C3 molecule of the complement system, to be phagocytized by phagocytic immune cells. Among these phagocytic cells are neutrophils, dendritic cells, and macrophages (13–15).
Neutrophils are the first cells to arrive at the infection site, being attracted by complement proteins, cytokines (e.g., IL-8), and chemokines (e.g., CXCL1 and CXCL2). These cells can eliminate the parasites through the action of nitric oxide (NO) and other reactive oxygen species (ROS), and also produce high levels of chemokines such as CXCL8 and CXCL9, responsible for the recruitment of more neutrophils and Th1 cells (16–18). In addition, because they are short-lived phagocytic cells, they promote the entry of more promastigotes in macrophages phagocyting dead neutrophils. Thus, neutrophils serve as a “trojan horses”, but the macrophages also produce cytokines which will in turn activate other immune cells (19, 20).
Resident dendritic cells (DCs) seems to play a key role in the immunopathogenesis of CL (21, 22). Once these cells interact with Leishmania there is an increase in the expression of co-stimulatory molecules, such as CD40, CD80 and CD86 which are essential for T cell activation (23, 24). Thus, it is hypothesized that resident DCs recruit monocytes, which differentiate into monocyte-derived dendritic cells (moDCs). These cells have an intense phagocytic activity, and evidences suggests that they favor the growth and survival of parasites (25, 26).
When Leishmania spp. are phagocytized, they are trapped in vesicles called phagosomes, which then fuse with lysosomes and turn into acidic phagolysosomes. From this point on, cellular processes such as oxidative stress and nitric oxide (NO) production take place, thus making the phagolysosome very acidic and hydrolytic. Promastigotes, which are sensitive to acidic and hydrolytic environments, begin to be eliminated. However, as a survival strategy, the promastigotes begin to transform into the form of amastigotes, since the latter are more resistant (13, 27).
The infected macrophages stimulate the production of pro-inflammatory cytokines (IL-1, TNF, IL-18 and IL-12) and chemokines (CXCL10, CCL4, CCL8, CCL11 and CXCL8), which also act to kill the parasite. At the same time the adaptive immune response induce activation of T cells which produce a cellular immune response, and the B-lymphocytes will be involved in the production of antibodies (28, 29).
Natural Killer Cells
The NK cells are innate immune cells which correspond to 5-20% of circulating lymphocytes in the blood (30–32). These cells can be found in various locations in the body such as liver, bone marrow, and thymus (33) and can directly kill infected or modified tumor cells, in addition to their cytokine-producing function (30, 34).
NK cells are phenotypically known to express CD56 and CD16 and lack CD3 expression in humans. In mouse they express NK1.1 (NKR-P1C), NCR1 (NKp45/CD335) and CD49b, and there is no expression of CD56 by rodent NK cells (35–38). Natural Killer T cells (NKT) are a less frequent subpopulation (0.1-0.5% of peripheral blood leukocytes) of NKs characterized by CD3 and CD56 expression (38–41). NKT cells are T lymphocytes that possess characteristics of both these cells and NK cells, thus adding function of both cell types. There are two types of NKT cells: type I NKT or invariant NKT (iNKT), which express and invariant TCR with the Vα24-Jα18 segment associated with the Vβ11 chain, and type II NKT or non-invariant NKT cells, which express the TCR with varying TCRs on their cell surface (42–44). Among the characteristics of an NKT cell are the presence of the invariant TCR, expression of CD1d, and high production of cytokines, especially IFN-γ, TNF, IL-4, IL-10, and IL-13 (45–49).
Why Does the Study of NK and NKT Cells Are Important in Cutaneous Leishmaniasis?
NK cells are part of the innate immune response and are known for their ability to kill infected cells, as well as to produce cytokines that act in the activation of other immune cells (Figure 1). For some time, studies have been trying to dissect the role of NK cells in CL, as well as their contributions to the cure or progression of the disease.
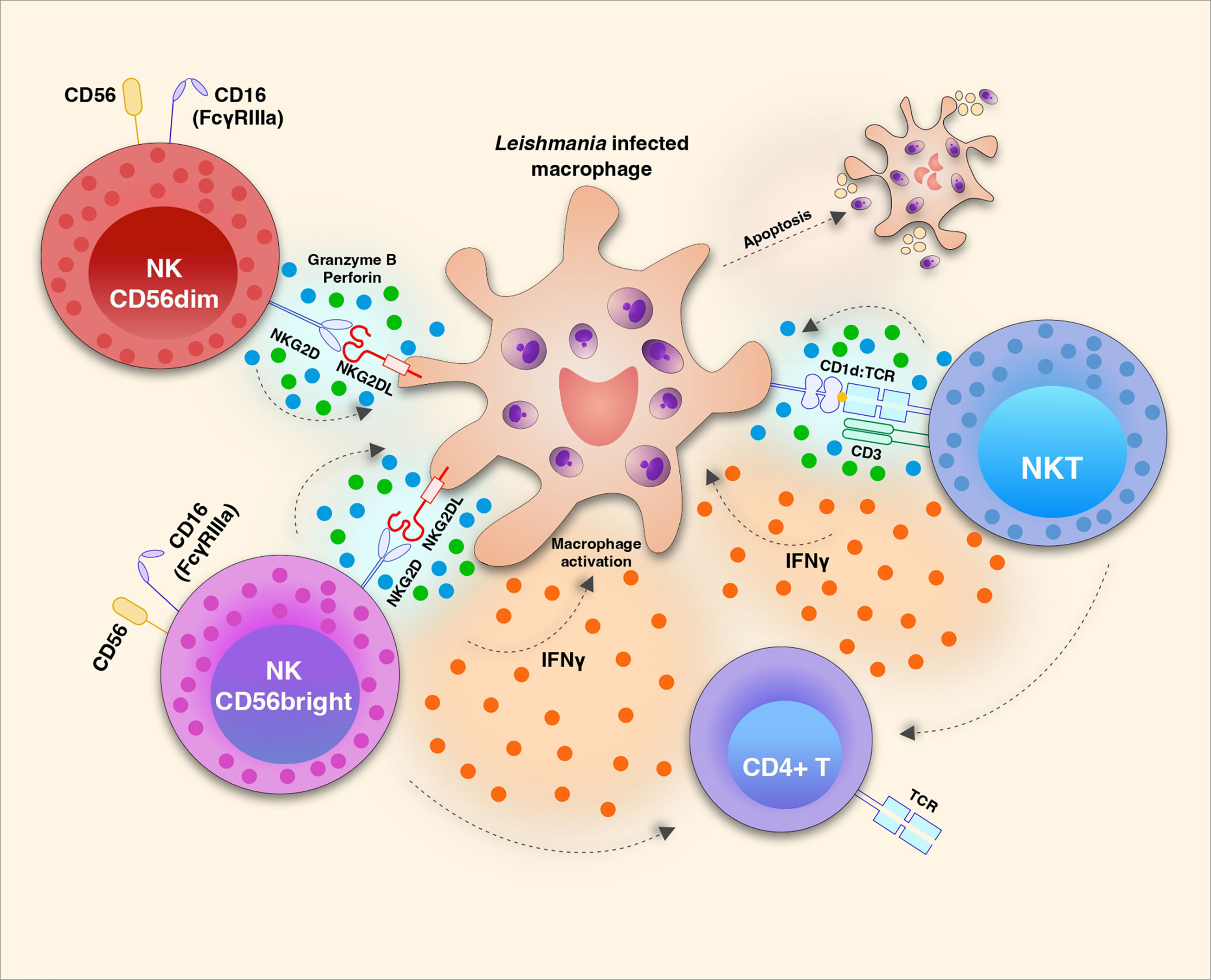
Figure 1 Natural killer (NK) cell subsets play distinct roles during Leishmania infection. NKG2D/NKG2D ligand (L) interaction triggers the effector function of CD56dim (also CD16+ in humans) NK cells to release cytotoxic granules with Granzyme b and Perforin which induces apoptosis of Leishmania infected macrophages. It is not yet clear tough if this mechanism restricts parasite growth or amplify amastigote proliferation. The same receptor-ligand interaction induces the activation of CD56bright NK cells which are strong producers of cytokines such as IFN-γ and TNF which in turn can act in the activation of macrophages and T cells. The potential recognition of glycolipid antigens present by CD1d to the invariant TCR of NKT cells may induce the production and release of IFN-γ and also apoptotic factors Granzyme B and Perforin.
Some authors suggest that these cells act to fight infection due to their cytotoxic role, however, other authors suggest that these cells are also responsible for more collateral damage contributing to lesion exacerbation (50). To tackle this problem, a 5-year cohort study observed that even after clinical cure of CL, the patients’ NK cells continue to produce Interferon-γ (IFN-γ), and this was associated with protection against CL (51). IFN-γ is key-cytokine for macrophage activation and consequent parasite clearance (52).
It has also been shown that the in vitro stimulation of peripheral blood mononuclear cells from healthy individuals with Leishmania antigen/peptides induces an increase in the frequency of NK cells specially (53, 54). On the other hand, Lieke and colleagues observed a decrease in NK cell proliferation in animal and human models using L. major and L. aethiopica where the promastigote form of Leishmania inhibited proliferation of isolated naive NK cells (55, 56).
NK cells are strong producers of cytokines and cytotoxic granules such as granzyme and perforin and surface markers such as NKG2D, triggering activation of other cells, increasing phagocytosis and parasite elimination (57). However, paradoxically, some works have shown evidence that these NK-derived effector molecules play a key role in the immunopathogenesis of CL, in addition to the inflammation developed. It was observed that patients with active lesion present a higher frequency of these cells, which in turn is directly associated with high levels of IFN-γ, Tumor Necrosis Factor (TNF), as well as granzyme A, granzyme B, granulysin and perforin. For this reason, it was found that most of the cytotoxic activity generated in CL is related to NK and not only to CD8+ T cells (58–60). Another study showed that there is no change in the frequency of NK cells according to the period of infection, but in patients who are on treatment as well as in clinically cured patients there is a reduction in their degranulation potential (61).
There seems to be a divergent role of NK cells according to the clinical form that the patient displays. It has been seen that patients with the diffuse form of the disease have a lower frequency of NK cells, also exhibiting lower levels of cytokines such as IFN-γ and TLR expression such as TLR1, TLR2 and TLR6 (62, 63). This fact seems to be related to the severity of the disease. Unlike patients who present the localized form, who exhibit high levels of cytokines, besides a higher frequency of NK cells (63).
Experimental models have demonstrated that depleting NK cells in C57BL/6 mice through the administration of anti-asialo-GM1 or NK1.1 antibodies induces lesion exacerbation in the first weeks of infection. In addition, these animals showed swelling in the local tissue and higher parasite numbers when compared to normal animals (64). NK cell depletion has also been shown to lead to a reduction in IFN-γ levels, which may compromise Th1 response development (64, 65).
Studies on the role of NKT in Leishmania infection are still scarce (48). An in vitro study used monocyte-derived DCs generated from healthy donors’ buffy coats observed that Leishmania infantum infected DCs increase the expression of CD1d, causing NKT cells to recognize these cells. In addition, the researchers observed that the percentage of NKT cells producing IFN-γ was twice as high as that of IL-4-producing cells. Thus, it can be seen that NKT cells can act both in the production of cytokines for the activation of other immune cells, such as T lymphocytes, as well as act in a cytotoxic way on infected cells that cannot be lysed by conventional NK cells (66).
Studies have observed subpopulations of NKT cells that express CD4 and CD8 markers, where these subpopulations have distinct functions. While CD4+ NKT cells are strong and potent producers of cytokines such as IL-2, IL-4 and IL-13, CD8+ NKT cells act more aggressively in fighting infection, through their cytotoxic activity (67–69). Studies by Gumperz et al. (67) and Carvalho et al. (70) observed higher numbers of CD8+ NKT cells in healthy individuals (67, 70).
Recently it has been shown that antigen from Leishmania braziliensis is able to induce activation of NKT cells in peripheral blood mononuclear cells (PBMC) from patients with CL in addition to CD107a+ NKT cells, thus suggesting that these cells may be involved in inflammation and consequently lesion formation (59). This is confirmed by Ferraz and colleagues (2017) who observed the presence of these cells at the site of injury (71). CD107a is a surface marker found on NK cells as well as CD8+ T cells and is used to assess the degranulation of these cells, allowing one to investigate whether or not there is granule release and consequent lysis of infected cells (72, 73).
Currently, there is no consensus on the true function of NK and NKT cells in CL. The data presented by the studies suggest that NK cells appear to contribute strongly to a toxic environment and consequently to the development of lesions. At the same time, these cells seem to be crucial for the development of a cellular immune response, in view of their high production of IFN-γ, which helps in the activation of cells of the adaptive immune response. Studies to understand the factors that lead NK cells to generate a balance at the site of injury are strongly recommended.
Thus, more comprehensive research on these cells and their interaction with Leishmania is urgent. For example, single-cell RNA sequencing studies are necessary and can help understand the contribution of NK cells to lesion formation or resolution in CL. Moreover, those studies would foster the development of new treatments, diagnostics and vaccines to effectively combat CL.
Author Contributions
MC, RS and MB-D-C wrote the manuscript. VP contributed to the discussion of the draft and made final corrections. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by UFPE-Propesqi nº 12/2021. MC is the recipient of a Ph.D. fellowship from FACEPE.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Kedzierski L, Evans KJ. Immune Responses During Cutaneous and Visceral Leishmaniasis. Parasitology (2014) 141:1544–62. doi: 10.1017/S003118201400095X
2. Savoia D. Recent Updates and Perspectives on Leishmaniasis. J Infect Dev Ctries (2015) 9:588–96. doi: 10.3855/jidc.6833
3. Organización Panamericana de la Salud. Manual De Procedimientos Para Vigilancia Y Control De Las Leishmaniasis En Las Américas. 1st ed. OPS: Washington (2019). p. 166.
4. Brasil. Manual De Vigilância Da Leishmaniose Tegumentar. In: D de V das DT, editor, 2nd ed. Brasília: Ministério da Saúde; Secretaria de Vigilândia em Saúde. p. 4:189.
5. Sandoval Pacheco CM, Araujo Flores GV, Favero Ferreira A, Sosa Ochoa W, Ribeiro da Matta VL, Zúniga Valeriano C, et al. Histopathological Features of Skin Lesions in Patients Affected by non-Ulcerated or Atypical Cutaneous Leishmaniasis in Honduras, Central America. Int J Exp Pathol (2018) 99:249–57. doi: 10.1111/iep.12295
6. Barral-Netto M, Barral A, Brodskyn C, Carvalho Em, Reed Sg. Cytotoxicity in Human Mucosal and Cutaneous Leishmaniasis. Parasite Immunol (1995) 17:21–8. doi: 10.1111/j.1365-3024.1995.tb00962.x
7. Pirmez C, Yamamura M, Uyemura K, Paes-Oliveira M, Conceicao-Silva F, Modlin RL. Cytokine Patterns in the Pathogenesis of Human Leishmaniasis. J Clin Invest (1993) 91:1390–5. doi: 10.1172/JCI116341
8. Da-Cruz AM, Conceicao-Silva F, Bertho AL, Coutinho SG. Leishmania-Reactive CD4+ and CD8+ T Cells Associated With Cure of Human Cutaneous Leishmaniasis. Infect Immun (1994) 62:2614–8. doi: 10.1128/iai.62.6.2614-2618.1994
9. Antonelli LRV, Dutra WO, Almeida RP, Bacellar O, Carvalho EM, Gollob KJ. Activated Inflammatory T Cells Correlate With Lesion Size in Human Cutaneous Leishmaniasis. Immunol Lett (2005) 101:226–30. doi: 10.1016/j.imlet.2005.06.004
10. Mokni M. Cutaneous Leishmaniasis. Ann Dermatol Venereol (2019) 146:232–46. doi: 10.1016/j.annder.2019.02.002
11. Gurel MS, Tekin B, Uzun S. Cutaneous Leishmaniasis: A Great Imitator. Clin Dermatol (2020) 38:140–51. doi: 10.1016/j.clindermatol.2019.10.008
12. Filho AAP, Nascimento AA de S, Saab NAA, Fugiwara RT, D’Ávila Pessoa GC, Koerich LB, et al. Evasion of the Complement System by Leishmania Through the Uptake of Factor H, a Complement Regulatory Protein. Acta Trop (2021) 224:1–10. doi: 10.1016/j.actatropica.2021.106152
13. Rossi M, Fasel N. How to Master the Host Immune System ? Leishmania Parasites Have the Solutions! Int Immunol (2017) 30:103–11. doi: 10.1093/intimm/dxx075
14. Bahrami F, Harandi AM, Rafati S. Biomarkers of Cutaneous Leishmaniasis. Front Cell Infect Microbiol (2018) 8:222. doi: 10.3389/fcimb.2018.00222
15. Jacobs T, Andrä J, Gaworski I, Graefe S, Mellenthin K, Krömer M, et al. Complement C3 is Required for the Progression of Cutaneous Lesions and Neutrophil Attraction in Leishmania Major Infection. Med Microbiol Immunol (2005) 194:143–9. doi: 10.1007/s00430-004-0229-y
16. Conceição J, Davis R, Carneiro PP, Giudice A, Muniz AC, Wilson ME, et al. Characterization of Neutrophil Function in Human Cutaneous Leishmaniasis Caused by Leishmania Braziliensis. PLoS Negl Trop Dis (2016) 10:1–19. doi: 10.1371/journal.pntd.0004715
17. Cardoso T, Bezerra C, Medina LS, Ramasawmy R, Scheriefer A, Bacellar O, et al. Leishmania Braziliensis Isolated From Disseminated Leishmaniasis Patients Downmodulate Neutrophil Function. Parasite Immunol (2019) 41:1–8. doi: 10.1111/pim.12620
18. Ronet C, Passelli K, Charmoy M, Scarpellino L, Myburgh E, La TYH, et al. TLR2 Signaling in Skin Nonhematopoietic Cells Induces Early Neutrophil Recruitment in Response to Leishmania Major Infection. J Invest Dermatol (2019) 139:1318–28. doi: 10.1016/j.jid.2018.12.012
19. Hurrell BP, Regli IB, Tacchini-Cottier F. Different Leishmania Species Drive Distinct Neutrophil Functions. Trends Parasitol (2016) 32:392–401. doi: 10.1016/j.pt.2016.02.003
20. Santos-Mateus D, Passero F, Rodrigues A, Valério-Bolas A, SIlva-Pedrosa R, Pereira M, et al. The Battle Between Leishmania and the Host Immune System at a Glance. Int Trends Immun (2015) 4:28–34. doi: 10.18281/iti.2016.1.3
21. Liu D, Uzonna JE. The Early Interaction of Leishmania With Macrophages and Dendritic Cells and its Influence on the Host Immune Response. Front Cell Infect Microbiol (2012) 2:83. doi: 10.3389/fcimb.2012.00083
22. Contreras I, Estrada JA, Guak H, Martel C, Borjian A, Ralph B, et al. Impact of Leishmania Mexicana Infection on Dendritic Cell Signaling and Functions. PLoS Negl Trop Dis (2014) 8:1–13. doi: 10.1371/journal.pntd.0003202
23. Qi H, Popov V, Soong L. Leishmania Amazonensis -Dendritic Cell Interactions In Vitro and the Priming of Parasite-Specific CD4 + T Cells In Vivo. J Immunol (2001) 167:4534–42. doi: 10.4049/jimmunol.167.8.4534
24. Xin L, Li K, Soong L. Down-Regulation of Dendritic Cell Signaling Pathways by Leishmania Amazonensis Amastigotes. Mol Immunol (2008) 45:3371–82. doi: 10.1016/j.molimm.2008.04.018
25. Charmoy M, Brunner-Agten S, Aebischer D, Auderset F, Launois P, Milon G, et al. Neutrophil-Derived CCL3 Is Essential for the Rapid Recruitment of Dendritic Cells to the Site of Leishmania Major Inoculation in Resistant Mice. PLoS Pathog (2010) 6:e1000755. doi: 10.1371/journal.ppat.1000755
26. León B, López-Bravo M, Ardavín C. Monocyte-Derived Dendritic Cells Formed at the Infection Site Control the Induction of Protective T Helper 1 Responses Against Leishmania. Immunity (2007) 26:519–31. doi: 10.1016/j.immuni.2007.01.017
27. Carneiro MBH, Roma EH, Ranson AJ, Doria NA, Debrabant A, Sacks DL, et al. NOX2-Derived Reactive Oxygen Species Control Inflammation During Leishmania Amazonensis Infection by Mediating Infection-Induced Neutrophil Apoptosis. J Immunol (2018) 200:196–208. doi: 10.4049/jimmunol.1700899
28. Díaz NL, Zerpa O, Tapia FJ. Chemokines and Chemokine Receptors Expression in the Lesions of Patients With American Cutaneous Leishmaniasis. Mem Inst Oswaldo Cruz (2013) 108:446–52. doi: 10.1590/S0074-0276108042013008
29. Basu MK, Ray M. Macrophage and Leishmania: An Unacceptable Coexistence. Crit Rev Microbiol (2005) 31:145–54. doi: 10.1080/10408410591005101
30. Campbell KS, Hasegawa J. Natural Killer Cell Biology: An Update and Future Directions. J Allergy Clin Immunol (2013) 132:536–44. doi: 10.1016/j.jaci.2013.07.006
31. Lopez-Vergés S, Milush JM, Pandey S, York VA, Arakawa-hoyt J, Pircher H, et al. CD57 Defines a Functionally Distinct Population of Mature NK Cells in the Human CD56 Dim CD16+ NK-Cell Subset. Blood (2010) 116:3865–74. doi: 10.1182/blood-2010-04-282301.An
32. Cooper MA, Fehniger TA, Caligiuri MA. The Biology of Human Natural Killer-Cell Subsets. Trends Immunol (2001) 22:633–40. doi: 10.1016/S1471-4906(01)02060-9
33. Wu Y, Tian Z, Wei H. Developmental and Functional Control of Natural Killer Cells by Cytokines. Front Immunol (2017) 8:930. doi: 10.3389/fimmu.2017.00930
34. Bogdan C. Natural Killer Cells in Experimental and Human Leishmaniasis. Front Cell Infect Microbiol (2012) 2:69. doi: 10.3389/fcimb.2012.00069
35. Lanier LL, Testi R, Bindl J, Phillips JH. Identity of Leu-19 (CD56) Leukocyte Differentiation Antigen and Neural Cell Adhesion Molecule. J Exp Med (1989) 169:2233–8. doi: 10.1084/jem.169.6.2233
36. Tsukerman P, Stern-Ginossar N, Yamin R, Ophir Y, Stanietsky AMN, Mandelboim O. Expansion of CD16 Positive and Negative Human NK Cells in Response to Tumor Stimulation. Eur J Immunol (2014) 44:1517–25. doi: 10.1002/eji.201344170
37. Walzer T, Bléry M, Chaix J, Fuseri N, Chasson L, Robbins SH, et al. Identification, Activation, and Selective In Vivo Ablation of Mouse NK Cells via Nkp46. Proc Natl Acad Sci U.S.A. (2007) 104:3384–9. doi: 10.1073/pnas.0609692104
38. Inngjerdingen M, Kveberg L, Naper C, Vaage JT. Natural Killer Cell Subsets in Man and Rodents. Tissue Antigens (2011) 78:81–8. doi: 10.1111/j.1399-0039.2011.01714.x
39. Papamichail M, Perez SA, Gritzapis AD, Baxevanis CN. Natural Killer Lymphocytes : Biology , Development , and Function. Cancer Immunol Immunother (2004) 53:176–86. doi: 10.1007/s00262-003-0478-4
40. Cichocki F, Sitnicka E, Bryceson YT. NK Cell Development and Function - Plasticity and Redundancy Unleashed. Semin Immunol (2014) 26:114–26. doi: 10.1016/j.smim.2014.02.003
41. Shimasaki N, Jain A, Campana D. NK Cells for Cancer Immunotherapy. Nat Rev Drug Discovery (2020) 19:200–18. doi: 10.1038/s41573-019-0052-1
42. Lantz O, Bendelac A. An Invariant T Cell Receptor α Chain is Used by a Unique Subset of Major Histocompatibility Complex Class I-Specific CD4+ and CD4-8- T Cells in Mice and Humans. J Exp Med (1994) 180:1077–106. doi: 10.1084/jem.180.3.1097
43. Godfrey DI, Hammond KJL, Poulton LD, Smyth MJ, Baxter AG. NKT Cells: Facts, Functions and Fallacies. Immunol Today (2000) 21:573–83. doi: 10.1016/S0167-5699(00)01735-7
44. Miko E, Barakonyi A, Meggyes M, Szereday L. The Role of Type I and Type II NKT Cells in Materno-Fetal Immunity. Biomedicines (2021) 9:1–12. doi: 10.3390/biomedicines9121901
45. Exley MA, Koziel MJ. To be or Not to be NKT: Natural Killer T Cells in the Liver. Hepatology (2004) 40:1033–40. doi: 10.1002/hep.20433
46. Slauenwhite D, Johnston B. Regulation of NKT Cell Localization in Homeostasis and Infection. Front Immunol (2015) 6:255. doi: 10.3389/fimmu.2015.00255
47. Zamora-Chimal J, Fernández-Figueroa EA, Ruiz-Remigio A, Wilkins-Rodríguez AA, Delgado-Domínguez J, Salaiza-Suazo N, et al. NKT Cell Activation by Leishmania Mexicana LPG: Description of a Novel Pathway. Immunobiology (2017) 222:454–62. doi: 10.1016/j.imbio.2016.08.003
48. Zamora-Chimal J, Hernández-Ruiz J, Becker I. NKT Cells in Leishmaniasis. Immunobiology (2017) 222:641–6. doi: 10.1016/j.imbio.2016.11.014
49. Borg NA, Wun KS, Kjer-Nielsen L, Wilce MCJ, Pellicci DG, Koh R, et al. CD1d-Lipid-Antigen Recognition by the Semi-Invariant NKT T-Cell Receptor. Nature (2007) 448:44–9. doi: 10.1038/nature05907
50. Aranha FCS, Ribeiro U, Basse P, Corbett CEP, Laurenti MD. Interleukin-2-Activated Natural Killer Cells may Have a Direct Role in the Control of Leishmania (Leishmania) Amazonensis Promastigote and Macrophage Infection. Scand J Immunol (2005) 62:334–41. doi: 10.1111/j.1365-3083.2005.01681.x
51. Muniz AC, Bacellar O, Lima Lago E, Carvalho AM, Carneiro PP, Guimarães LH, et al. Immunologic Markers of Protection in Leishmania (Viannia) Braziliensis Infection: A 5-Year Cohort Study. J Infect Dis (2016) 214:570–6. doi: 10.1093/infdis/jiw196
52. Toepp AJ, Petersen CA. The Balancing Act: Immunology of Leishmaniosis. Res Vet Sci (2020) 130:19–25. doi: 10.1016/j.rvsc.2020.02.004
53. Maasho K, Sanchez F, Schurr E, Hailu A, Akuffo H. Indications of the Protective Role of Natural Killer Cells in Human Cutaneous Leishmaniasis in an Area of Endemicity. Infect Immun (1998) 66:2698–704. doi: 10.1128/iai.66.6.2698-2704.1998
54. Nylén S, Maasho K, McMahon-Pratt D, Akuffo H. Leishmanial Amastigote Antigen P-2 Induces Major Histocompatibility Complex Class II-Dependent Natural Killer-Cell Reactivity in Cells From Healthy Donors. Scand J Immunol (2004) 59:294–304. doi: 10.1111/j.0300-9475.2004.01388.x
55. Lieke T, Nyln S, Eidsmo L, Schmetz C, Berg L, Akuffo H. The Interplay Between Leishmania Promastigotes and Human Natural Killer Cells In Vitro Leads to Direct Lysis of Leishmania by NK Cells and Modulation of NK Cell Activity by Leishmania Promastigotes. Parasitology (2011) 138:1898–909. doi: 10.1017/S0031182011001363
56. Lieke T, Nylén S, Eidsmo L, McMaster WR, Mohammadi AM, Khamesipour A, et al. Leishmania Surface Protein Gp63 Binds Directly to Human Natural Killer Cells and Inhibits Proliferation. Clin Exp Immunol (2008) 153:221–30. doi: 10.1111/j.1365-2249.2008.03687.x
57. Rasolzadeh S, Haji Fatahaliha M, Hosseini M, Jafari R, Miahipour A, Movassaghpour AA, et al. Recombinant LPG3 Stimulates IFN-Γ and TNF-A Secretion by Human NK Cells. Iran J Parasitol (2015) 10:554–60.
58. Campos TM, Novais FO, Saldanha M, Costa R, Lordelo M, Celestino D, et al. Granzyme B Produced by Natural Killer Cells Enhances Inflammatory Response and Contributes to the Immunopathology of Cutaneous Leishmaniasis. J Infect Dis (2020) 221:973–82. doi: 10.1093/infdis/jiz538
59. Cunha CF, Ferraz-Nogueira R, Costa VFA, Pimentel MIF, Chometon TQ, Lyra MR, et al. Contribution of Leishmania Braziliensis Antigen-Specific CD4+ T, CD8+ T, NK and CD3+CD56+NKT Cells in the Immunopathogenesis of Cutaneous Leishmaniasis Patients: Cytotoxic, Activation and Exhaustion Profiles. PLoS One (2020) 15:1–15. doi: 10.1371/journal.pone.0229400
60. Novais FO, Carvalho LP, Graff JW, Beiting DP, Ruthel G, Roos DS, et al. Cytotoxic T Cells Mediate Pathology and Metastasis in Cutaneous Leishmaniasis. PLoS Pathog (2013) 9:1–11. doi: 10.1371/journal.ppat.1003504
61. Cunha CF, Ferraz R, Pimentel MIF, Lyra MR, Schubach AO, Da-Cruz AM, et al. Cytotoxic Cell Involvement in Human Cutaneous Leishmaniasis: Assessments in Active Disease, Under Therapy and After Clinical Cure. Parasite Immunol (2016) 38:244–54. doi: 10.1111/pim.12312
62. Salaiza-Suazo N, Volkow P, Pérez Tamayo R, Moll H, Gillitzer R, Pérez-Torres A, et al. Treatment of Two Patients With Diffuse Cutaneous Leishmaniasis Caused by Leishmania Mexicana Modifies the Immunohistological Profile But Not the Disease Outcome. Trop Med Int Heal (1999) 4:801–11. doi: 10.1046/j.1365-3156.1999.00491.x
63. Salaiza-suazo N, Ferna EA, Can IC, Becker I, Carrada-figueroa G, Aguirre-garcı M. NK Cell Activity Differs Between Patients With Localized and Diffuse Cutaneous Leishmaniasis Infected With Leishmania Mexicana : A Comparative Study of TLRs and Cytokines. PLoS One (2014) 9:1–17. doi: 10.1371/journal.pone.0112410
64. Laskay T, Röllinghoff M, Solbach W. Natural Killer Cells Participate in the Early Defense Against Leishmania Major Infection in Mice. Eur J Immunol (1993) 23:2237–41. doi: 10.1002/eji.1830230928
65. Scharton TM, Scott P. Natural Killer Cells are a Source of Interferon γ That Drives Differentiation of CD4+ T Cell Subsets and Induces Early Resistance to Leishmania Major in Mice. J Exp Med (1993) 178:567–78. doi: 10.1084/jem.178.2.567
66. Campos-martín Y, Colmenares M, López-núñez M, Savage PB, Martínez-naves E, Campos-martı Y, et al. Immature Human Dendritic Cells Infected With Leishmania Infantum Are Resistant to NK-Mediated Cytolysis But Are Efficiently Recognized by NKT Cells. J Immunol (2006) 176:6172–9. doi: 10.4049/jimmunol.176.10.6172
67. Gumperz JE, Miyake S, Yamamura T, Brenner MB. Functionally Distinct Subsets of CD1d-Restricted Natural Killer T Cells Revealed by CD1d Tetramer Staining. J Exp Med (2002) 195:625–36. doi: 10.1084/jem.20011786
68. O’Reilly V, Zeng SG, Bricard G, Atzberger A, Hogan AE, Jackson J, et al. Distinct and Overlapping Effector Functions of Expanded Human CD4 +, Cd8α + and CD4 -Cd8α - Invariant Natural Killer T Cells. PLoS One (2011) 6:2–12. doi: 10.1371/journal.pone.0028648
69. Montoya CJ, Pollard D, Martinson J, Kumari K, Wasserfall C, Mulder CB, et al. Characterization of Human Invariant Natural Killer T Subsets in Health and Disease Using a Novel Invariant Natural Killer T Cell-Clonotypic Monoclonal Antibody, 6B11. Immunology (2007) 122:1–14. doi: 10.1111/j.1365-2567.2007.02647.x
70. Carvalho KI, Melo KM, Bruno FR, Snyder-Cappione JE, Nixon DF, Costa-Carvalho BT, et al. Skewed Distribution of Circulating Activated Natural Killer T (NKT) Cells in Patients With Common Variable Immunodeficiency Disorders (CVID). PLoS One (2010) 5:1–8. doi: 10.1371/journal.pone.0012652
71. Ferraz R, Cunha CF, Pimentel MIF, Lyra MR, Pereira-Da-Silva T, Schubach AO, et al. CD3+CD4negCD8neg (Double Negative) T Lymphocytes and NKT Cells as the Main Cytotoxic-Related-CD107a+ Cells in Lesions of Cutaneous Leishmaniasis Caused by Leishmania (Viannia) Braziliensis. Parasites Vectors (2017) 10:1–12. doi: 10.1186/s13071-017-2152-2
72. Aktas E, Kucuksezer UC, Bilgic S, Erten G, Deniz G. Relationship Between CD107a Expression and Cytotoxic Activity. Cell Immunol (2009) 254:149–54. doi: 10.1016/j.cellimm.2008.08.007
Keywords: natural killer, cutaneous leishmaniasis, immune response, tegumentary leishmaniasis, leishmania
Citation: Cavalcante MKA, de Freitas e Silva R, Pereira VRA and Brelaz-de-Castro MCA (2022) Opinion Article: NK Cells in Cutaneous Leishmaniasis: Protection or Damage? Front. Immunol. 13:933490. doi: 10.3389/fimmu.2022.933490
Received: 01 May 2022; Accepted: 01 June 2022;
Published: 01 July 2022.
Edited by:
Pedro H Gazzinelli-Guimaraes, National Institute of Allergy and Infectious Diseases (NIH), United StatesReviewed by:
Tiago Rodrigues Ferreira, National Institute of Allergy and Infectious Diseases (NIH), United StatesCopyright © 2022 Cavalcante, de Freitas e Silva, Pereira and Brelaz-de-Castro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Carolina Accioly Brelaz-de-Castro, Y2Fyb2xpbmEuYnJlbGF6QGdtYWlsLmNvbQ==