
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 24 November 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.933191
This article is part of the Research Topic Systemic Lupus Erythematosus - Predisposition Factors, Pathogenesis, Diagnosis, Treatment and Disease Models View all 21 articles
 Antoine N. Kraemer1,2
Antoine N. Kraemer1,2 Anna-Lena Schäfer1,2
Anna-Lena Schäfer1,2 Dalina T. L. Sprenger1,2
Dalina T. L. Sprenger1,2 Bettina Sehnert1,2
Bettina Sehnert1,2 Johanna P. Williams1,2Aileen Luo1,2Laura Riechert1,2Qusai Al-Kayyal1,2
Johanna P. Williams1,2Aileen Luo1,2Laura Riechert1,2Qusai Al-Kayyal1,2 Hélène Dumortier3Jean-Daniel Fauny3Zoltan Winter4Kathrin Heim5
Hélène Dumortier3Jean-Daniel Fauny3Zoltan Winter4Kathrin Heim5 Maike Hofmann5
Maike Hofmann5 Martin Herrmann6
Martin Herrmann6 Guido Heine7
Guido Heine7 Reinhard E. Voll1,2*
Reinhard E. Voll1,2* Nina Chevalier1,2*
Nina Chevalier1,2*Vitamin D (VD) deficiency is a highly prevalent worldwide phenomenon and is extensively discussed as a risk factor for the development of systemic lupus erythematosus (SLE) and other immune-mediated diseases. In addition, it is now appreciated that VD possesses multiple immunomodulatory effects. This study aims to explore the impact of dietary VD intake on lupus manifestation and pathology in lupus-prone NZB/W F1 mice and identify the underlying immunological mechanisms modulated by VD. Here, we show that low VD intake accelerates lupus progression, reflected in reduced overall survival and an earlier onset of proteinuria, as well higher concentrations of anti-double-stranded DNA autoantibodies. This unfavorable effect gained statistical significance with additional low maternal VD intake during the prenatal period. Among examined immunological effects, we found that low VD intake consistently hampered the adoption of a regulatory phenotype in lymphocytes, significantly reducing both IL-10-expressing and regulatory CD4+ T cells. This goes along with a mildly decreased frequency of IL-10-expressing B cells. We did not observe consistent effects on the phenotype and function of innate immune cells, including cytokine production, costimulatory molecule expression, and phagocytic capacity. Hence, our study reveals that low VD intake promotes lupus pathology, likely via the deviation of adaptive immunity, and suggests that the correction of VD deficiency might not only exert beneficial functions by preventing osteoporosis but also serve as an important module in prophylaxis and as an add-on in the treatment of lupus and possibly other immune-mediated diseases. Further research is required to determine the most appropriate dosage, as too-high VD serum levels may also induce adverse effects, possibly also on lupus pathology.
Systemic lupus erythematosus (SLE) is a chronic autoimmune disease affecting many organs and causing significant morbidity and mortality. SLE is characterized by immune dysregulation and a breakdown of tolerance to self-antigens. This results in the production of autoantibodies, which are most commonly directed against nuclear self-antigens, in particular double-stranded DNA (dsDNA). These autoantibodies may form immune complexes (ICs) with autoantigens, ultimately leading to inflammation and tissue damage (1, 2). In SLE, the kidneys are often afflicted by this inflammation, which can result in lupus nephritis (LN), which eventually leads to renal failure (3).
Generally, in SLE, there may be defects in various branches of the immune system, explaining the great heterogeneity of clinical presentations. Several dysregulations within the innate immune compartment have been reported in the context of SLE, including an impaired clearance of apoptotic debris by macrophages. For instance, in the germinal centers (GCs) of the lymph nodes, this results in an increased exposure of nuclear self-antigens and increases the risk of autoreactive B-cell activation by follicular dendritic cells and ultimately drives the production of autoantibodies (4–7). Furthermore, monocytes and macrophages in SLE display abnormal cytokine production, adopting a more pro-inflammatory cytokine profile (8–10). The adaptive immune system is also afflicted by dysregulation in patients with SLE. For instance, studies have reported an increased presence of B-cell survival factors that increase the activation and survival of autoreactive B cells (11, 12). In addition, an increase in follicular T helper (TFH) cells and a decrease of regulatory T cells (Tregs) has been reported in SLE (13–15).
The observation that autoimmune diseases, such as SLE, are usually rare among non-Westernized populations suggests a significant impact of the profound lifestyle and environmental changes in modern Western societies on the development of such diseases. Although the exact pathological mechanisms underlying SLE are still elusive, it is becoming increasingly clear that the pathogenesis of SLE and other immune-mediated diseases are the result of a complex interplay between genetic and environmental factors. The environmental factors that have been suggested to pose a risk for developing SLE include infectious agents, Western dietary habits, and ultraviolet (UV) radiation inducing apoptotic cell death. A highly discussed environmental risk factor for SLE and other autoimmune diseases is the deficiency of vitamin D (VD) (16–20). Multiple studies have shown a high prevalence of VD deficiency among SLE patients, likely, in part, due to the fact that the majority of SLE patients are hypersensitive to disease-triggering UV light and thus protect themselves from sunlight (21–29). However, the relevance of VD on the pathogenesis, whether it is a secondary phenomenon or involved in disease manifestation and progression, is not entirely clear.
VD is a secosteroid hormone and is biosynthesized in the epidermis upon exposure to UV-B radiation, or ingested from dietary sources, such as cod liver oil, and supplements, all in the form of cholecalciferol (vitamin D3). VD ultimately exerts its effects via the regulation of genes and the epigenome, through the actions of the active VD metabolite 1,25(OH)2D3 (calcitriol) binding the VD receptor, which is expressed by many immune cells (30–35). Although VD is most prominently known for its regulatory role in calcium and phosphate homeostasis, the immunomodulatory effects of VD have become increasingly appreciated. For instance, VD promotes the differentiation of monocytes into macrophages while inhibiting the maturation of differentiating dendritic cells (DCs) (36–39). In the latter, VD induces a tolerogenic phenotype by suppressing their antigen-presenting capacity, thus inhibiting the ability to activate T cells (39–41). In the adaptive immune system, VD is known to promote Treg differentiation while inhibiting TH1 and TH17 cell differentiation (42–46). There is also in vitro evidence showing that VD inhibits the differentiation of B cells into plasma cells, decreases antibody production, and promotes IL-10 expression (47–51).
The ability of VD to regulate antigen presentation and tip lymphocyte differentiation toward a more anti-inflammatory or immunoregulatory phenotype suggests therapeutic potential for SLE and other autoimmune diseases. Therefore, we investigated the effects of dietary VD intake on lupus pathology and associated immunological effects in NZB/W F1 mice, a spontaneous model of murine lupus (52). We found that low VD intake accelerates lupus progression, with this effect becoming significant when low VD intake commenced during the prenatal period. In our broad immune-phenotypic analysis, we found that low VD intake consistently hampered the adoption of a regulatory phenotype in lymphocytes. Hence, our data provide evidence of possible adverse effects of VD deficiency on lupus progression and encourage further studies to elucidate detailed mechanisms. These should also address optimal dosage, as excessively elevated VD serum levels may exert adverse effects, possibly also on lupus progression.
Lupus-prone NZB/W F1 mice were generated by crossing NZB/BlNJ with NZW/LacJ mice, which were purchased from The Jackson Laboratory. CD45.1 C57BL/6 animals (B6.SJL-Ptprca Pepcb/BoyJ) were bred in our own facility. For all experiments, the mice were housed in individually ventilated cages on a 12 h light/dark cycle, with food and water ad libitum. To test the influence of VD, randomized mice were fed the following diets: a low-VD diet (< 50 IU/kg cholecalciferol and Ca:P=2.5:1, Altromin C 1017 mod.), a normal VD diet (500 IU/kg cholecalciferol and Ca:P=1.2:1, Altromin C 1000 control diet) or a high-VD diet (76,500 or 38,000 IU/kg cholecalciferol and Ca:P=1.5:1, Altromin C 1017 mod.). All diets were purchased from Altromin. In a first VD-feeding approach, treatments were started after weaning at roughly 5 weeks of age and continued throughout the entire duration of the experiment. In a second VD-feeding approach, NZB/BlNJ and NZW/LacJ parent animals were also fed the different VD diets and NZB/W F1 offspring were weaned on the same food that was fed continuously throughout the entire duration of the experiment. Blood and urine were collected and mice were euthanized at defined time points for organ harvest and downstream experiments. Animals were regularly monitored and euthanized when reaching defined ethical endpoints [proteinuria in addition to a deteriorating general health condition and/or significant weight loss (≥ 20% over the course of 2 days)].
To determine grip strength, mice were placed individually on top of the center of a stainless-steel grid (50 cm × 30 cm) with a mesh size of 12 mm and a thickness of 1.6 mm. The grid was subsequently inverted by 180°, resulting in mice hanging upside down on all four limbs roughly 30 cm over a soft landing spot. The duration of hanging time was measured and normalized to bodyweight.
For the µCT scanning of the tibiae, a dedicated preclinical scanner (Siemens Inveon PET/CT/SPECT Multimodality System, Siemens Healthineers, Germany) was used at a tube voltage of 80 kV and a tube current of 500 μA. The samples were positioned, fixed, and images were acquired with an isotropic resolution of 17.04 μm.
Urine samples were collected by spontaneous urination. For a semiquantitative measurement of proteinuria, Albustix test strips (Siemens) were used. According to the color scale provided by the manufacturer, albuminuria was categorized as follows: 0–1 = trace, 1 = 30 mg/dl, 2 = 100 mg/dl, 3 = 300 mg/dl, and 4 > 2,000 mg/dl. Mice were deemed proteinuric after scoring a 2 on the color scale for at least two consecutive weeks.
Serum titers of IgG autoantibodies directed against dsDNA, as well as serum titers of total IgG were determined by enzyme-linked immunosorbent assay (ELISA). For anti-dsDNA IgG ELISAs, 384-well microtiter plates (Greiner Bio One) were precoated with 15 µl 20 µg/ml Poly-L-Lysin (Sigma-Aldrich) for 1 h at 37°C, followed by coating with 15 µl 20 µg/ml calf thymus DNA (Sigma-Aldrich) at 4°C overnight (o/n). Plates were blocked with 2% fetal calf serum (FCS) in Phosphate-buffered saline (PBS) for 2 h at RT. For total IgG ELISAs, 384-well microtiter plates were coated with 15 µl 1 µg/ml goat anti-mouse IgG (Jackson ImmunoResearch) at 4°C o/n. The plates were blocked with 2% FCS in PBS for 2 h at RT. Samples were diluted in 2% FCS in PBS and 40 µl incubated for 2 h at RT. Bound IgG was detected with 15 µl 160 ng/µl HRP-conjugated goat anti-mouse IgG secondary antibody (Jackson ImmunoResearch). Development was performed with a 3,3’5,5’-Tetramethylbenzidine (TMB) substrate (Thermo Fisher Scientific), according to the manufacturer’s protocol. The absorbance at 450 nm was measured using the Spark® 10 M multimode microplate reader (Tecan). To determine anti-dsDNA IgG titers, expressed as arbitrary units (A.U.), reference sera of nephritic female NZB/W F1 mice were used to create a standard curve. To determine total IgG titers, expressed as A.U., reference sera of wild-type C57BL/6 mice were used to create a standard curve.
Single-cell suspensions of the spleen were obtained by the mechanic dissociation of the spleen and of the bone marrow by flushing out the bone marrow from truncated tibiae and femurs via centrifugation. Peritoneal exudate cells (PECs) were obtained via peritoneal lavage; peripheral blood immune cells were retrieved from blood collected in Ethylenediaminetetraacetic acid (EDTA)-coated tubes. Red blood cell (RBC) lysis was performed for 5 min, for all cell suspensions, except PECs. Following incubation with anti-CD16/32 antibodies (101330, BioLegend) to block non-specific Fc receptor binding, single-cell suspensions were stained with Biotin- or fluorochrome-conjugated monoclonal antibodies diluted in 2% FCS/PBS for 30 min on ice. For intracellular and intranuclear staining, cells were fixed and permeabilized with BD Cytofix/Cytoperm (BD Biosciences) and the eBioscience FoxP3/Transcription Factor Staining Buffer Set (eBioscience), respectively. For intracellular cytokine staining, cells were stimulated with 50 ng/ml PMA (Sigma-Aldrich), 1 µg/ml ionomycin (Sigma-Aldrich), and brefeldin A (eBioscience) for 4 h at 37°C/5% CO2 prior to staining and fixation. To identify apoptotic cells, annexin V staining was performed using Annexin V Binding Buffer (BD Biosciences). The following anti-mouse antibodies were used: B220/CD45R Biotin (103204, BioLegend), B220/CD45R Pacific Blue (103230, BioLegend), B220/CD45R APC-Cy7 (103224, BioLegend), CD11b Biotin (13-0112-82, eBioscience), CD11b FITC (101206, BioLegend), CD11b PE-Cy7 (101215, BioLegend), CD11c APC (117309, BioLegend), CD11c Biotin (117303, BioLegend), CD11c PE-Cy7 (117318, BioLegend), CD127/IL-7Rα PE-Cy7 (135014, BioLegend), CD138 BV421 (142508, BioLegend), CD16/32 (101330, BioLegend), CD16/32 FITC (MCA2305F, Serotec), CD19 Biotin (115504, BioLegend), CD200R PE (123907, BioLegend), CD206 APC (141707, BioLegend), CD2 Biotin (100103, BioLegend), CD34 APC (128606, BioLegend), CD301 PE-Cy7 (145705, BioLegend), CD36 APC/Fire 750 (102617, BioLegend), CD3ϵ Biotin (13-0031-85, eBioscience), CD4 Biotin (100404, BioLegend), CD4 PE-Cy7 (100421, BioLegend), CD45 eFluor506 (69-0451-82, eBioscience), CD45.1 APC (110714, BioLegend), CD45.2 Pacific Blue (109819, BioLegend), CD80 APC/Fire 750 (104739, BioLegend), CD86 APC (105011, BioLegend), CD8α PerCP (100732, BioLegend), CD8β PerCP-Cy5.5 (140417, BioLegend), c-kit/CD117 PE (553355, BD Pharmingen), CXCR5 Biotin (551960, BD Pharmingen), F4/80 APC-Cy7 (123117, BioLegend), F4/80 FITC (123107, BioLegend), Fas/CD95 PE (554258, BD Pharmingen), FoxP3 APC (14-5773-82, eBioscience), GL7 FITC (553666, BD Pharmingen), I-A/I-E Pacific Blue (107619, BioLegend), ICOS-L PE (107405, BioLegend), IFN-γ APC (505809, BioLegend), IL-10 FITC (505006, BioLegend), IL-17A PE (559502, BD Pharmingen), Ly6C PE-Cy7 (560593, BD Pharmingen), Ly6G Biotin (127604 BioLegend), Ly6G Pacific Blue (127611, BioLegend), MerTK PE (151505, BioLegend), NK1.1 APC-Cy7 (108723, BioLegend), NK1.1 Biotin (108704, BioLegend), PD-1/CD279 PE (12-9985-83, eBioscience), PDCA-1/CD317 PE (12-3172-82 eBioscience), Sca-1/Ly6A/E Pacific Blue (108120, BioLegend), TACI/CD267 APC (17-5942-81, eBioscience), TCR-β APC-Cy7 (109220, BioLegend), TCR-β Biotin (109204, BioLegend), TCR-β PerCP-Cy5.5 (109227, BioLegend), TER-119 Biotin (116203, BioLegend), Tim-4 PE-Cy7 (130009, BioLegend).
As described previously (53), the following immune cell subsets were identified: B cells (% TCR-β-B220+/single cells), germinal center B cells (GC B cells) (% FashiGL7hi/B cells), CD4+ and CD8+ T cells (% TCR-β+B220-CD4+CD8- or TCR-β+B220-CD4-CD8+/single cells), expression of IL-10 in B cells, as well as IL-10, IL-17A and IFN-γ in CD4+ or CD8+ T cells, regulatory T cells (Tregs) (% FoxP3+/CD4+ T cells), follicular T helper (TFH) cells (% CXCR5hiPD-1hi/CD4+ T cells), conventional dendritic cells (cDCs) and plasmacytoid dendritic cells (pDCs) (% CD11chiPDCA-1-CD19-TCR-β-NK1.1- or PDCA-1hiCD19-TCR-β-NK1.1-/single cells), CD11b+ monocytic cells (% CD19-TCR-β-NK1.1-PDCA-1-CD11c-CD11b+Ly6G-/single cells), circulating CD11b+ monocytic cells (% Ly6G-CD11b+/CD45+ cells), neutrophils (%CD19-TCR-β-NK1.1-PDCA-1-CD11c-CD11b+Ly6Ghi/single cells), circulating neutrophils (% CD11b+ Ly6Ghi/CD45+ cells), natural killer (NK) cells (% CD19-TCR-β-NK1.1+/single cells) and expression of CD80 and CD86 on CD11b+ monocytic cells, cDCs, peritoneal macrophages, and B cells (Supplementary Figures 11, 12). Further immune cell subsets were also identified: plasma cells (PCs) (% CD138hi/single cells); peritoneal macrophages (F4/80+Ly6Gint/-); Ly6Chi peritoneal macrophages (% Ly6Chi/peritoneal macrophages); Ly6Chi CD11b+ monocytic cells (% Ly6Chi/CD11b+ monocytic cells); the expression of ICOS-L on CD11b+ monocytic cells, cDCs, peritoneal macrophages and B cells; and the expression of Tim-4, MerTK, CD206, I-A/I-E, CD36, CD301 and CD200R on peritoneal macrophages and bone marrow–derived macrophages (BMDMs). In peripheral blood, additional CD45-staining was included to identify leukocytes. Flow cytometric analysis was performed at the BD LSR Fortessa flow cytometer (Becton Dickinson) followed by data analysis using FlowJo™ Software (Becton Dickinson).
Single-cell suspensions of splenocytes were obtained by mechanic dissociation of the spleen, while the single-cell suspensions of PECs were obtained following peritoneal lavage. RBC lysis was performed for 5 min for the single-cell suspensions of splenocytes. Cells were subsequently stained with a cocktail of antibodies. The following anti-mouse antibodies were used: CD11b FITC (101206, BioLegend), CD11b PE (101207, BioLegend), CD11c APC (117309, BioLegend), CD19 FITC (115505, BioLegend), F4/80 PE (123109, BioLegend), Ly6G Pacific Blue (127611, BioLegend), NK1.1 FITC (108705, BioLegend), and TCR-β FITC (109205, BioLegend). Splenic CD11b+ innate immune cells were sorted as CD19-TCR-β-NK1.1-Ly6G-CD11b+, while peritoneal macrophages were sorted as F4/80+CD11c-Ly6G-CD11b+ (Supplementary Figure 13). Splenic CD11b+ innate immune cells and peritoneal macrophages were sorted by BD FACSMelody™ (Becton Dickinson) and stimulated with 50 ng/ml LPS for 2 h 45 min at 37°C/5% CO2 followed by RNA isolation.
0.4 × 106 murine bone marrow (BM) cells were seeded per well in a 48-well culture plate and cultured for 5 days in the presence of 20 ng/ml GM-CSF (Peprotech) or 20 ng/ml M-CSF (Peprotech) to generate BM-derived innate immune cells and BM-derived macrophages (BMDMs), respectively. The medium supplemented with GM-CSF or M-CSF was refreshed on day 3. On day 5 of incubation and immediately after an additional exchange of the culture medium, cells were stimulated with either 50 ng/ml LPS (Sigma-Aldrich) and 50 ng/ml Pam3CSK4 (In vivogen) or 1 µM CpG (In vivogen) and 1 µM R848 (Enzo Life Science) for 20 h. Cytokine concentrations in the culture supernatants were assessed by ELISA. Cytokine concentrations were determined using the IL-10, IL-1β, and TNF-α DuoSet kits (R&D Systems), according to the manufacturer’s protocol. The absorbance at 450 nm was measured using the Spark® 10 M multimode microplate reader (Tecan). Cytokine concentrations in supernatants were normalized to the total protein content of adherent cells, which was determined via BCA assay, according to the manufacturer’s protocol (Thermo Fisher Scientific).
The phagocytic capacity of both murine peritoneal macrophages and BMDMs (both CD45.2+) was investigated, by co-culturing said cells with CD45.1+ apoptotic thymocytes (ATs). Thymocytes were isolated from CD45.1+ C57BL/6 mice. After mechanical disruption to generate a single-cell suspension, thymocytes were treated with 1 µM dexamethasone for 6 h to induce apoptosis and subsequently labeled with CFDA-SE. 0.5 × 106 PECs were seeded per well in a 96-well U-bottom culture plate and rested for 2 h at 37°C/5% CO2. 2.5 × 106 CFDA-SE-labeled ATs were then added to PECs and coincubated for 1 h at 37°C/5% CO2. PECs were then placed on ice and washed and stained for flow cytometry in order to identify the percentage of peritoneal macrophages that had phagocytosed ATs. In a second phagocytosis assay, 0.4 × 106 murine BM cells were seeded per well in a 48-well culture plate and cultured for 5 days in the presence of 20 ng/ml M-CSF (Peprotech) to generate differentiated adherent BMDMs. The medium supplemented with M-CSF was refreshed on day 3. On day 5 of incubation, 2.5 × 106 ATs were added to BMDMs and coincubated for 1 h at 37°C/5% CO2. BMDMs were then washed, detached from the well bottom with trypsin 0.25% (Anprotec), and stained for flow cytometry in order to identify the percentage of BMDMs that had phagocytosed ATs.
Total RNA was extracted from homogenized kidney tissue or purified cells using TRIzol reagent (Invitrogen) or the RNeasy® Micro Kit (Qiagen), respectively. The QuantiTect Reverse Transcription Kit (Qiagen) was used for cDNA synthesis according to the manufacturer’s protocol. Transcripts were quantified by quantitative reverse transcription PCR (RT-qPCR) on a StepOnePlus™ Real-Time PCR System (Applied Biosystems) with predesigned TaqMan Gene Expression Assays and reagents according to the manufacturer’s instructions (Applied Biosystems), alternatively with predesigned SYBR Green master mixes (Thermo Fisher Scientific) and custom-designed primers (Supplementary Table 1). Probes with the following Applied Biosystems assay identification numbers were used: Mm99999915_g1 (GAPDH), Mm01288386_m1 (IL1B), Mm00446190_m1 (IL6), and Mm00443258_m1 (TNF). For each sample, mRNA abundance was normalized to the amount of GAPDH and is presented as fold gene expression (2(ΔΔCt)).
Kidney specimens were embedded in O.C.T. compound (Tissue-Tek), snap-frozen on dry ice, and cut into 8 µm sections using a cryotome (Zeiss, Germany). Dried kidney sections were immersed in pre-cooled acetone for 20 min on ice for fixation. Kidney sections were blocked with 10% FCS, 0.3% Triton in PBS for 2 h at RT and stained with FITC-conjugated polyclonal goat anti-mouse IgG (SouthernBiotech) overnight at 4°C. After washing with 0.05% Tween 20 in PBS, sections were fixed again in 4% PFA in PBS for 15 min at RT. After four rounds of washing, sections were incubated with DAPI (Roche) for 10 min at RT for the labeling of nuclei. Sections were mounted with a fluorescent Mounting Medium (Dako) and imaged with a fluorescence microscope (AxioImager 2, Zeiss), using a monochrome camera at ×20 magnification. For IgG quantification, the mean intensity of fluorescence in 30 glomeruli per section was determined.
Cell counts, including RBCs, platelets, white blood cells (WBCs), and WBC subtypes, in freshly isolated murine blood were determined by collecting blood in EDTA-coated tubes and analyzing them using a hematology cell counter (Scil Vet abc and Scil Vet abc Plus+).
For statistical analysis, InStat software Prism 9 (GraphPad software) was used. P-values less than or equal to 0.05 were considered significant. A statistical comparison between two experimental groups was performed using an unpaired t-test (normally distributed data), or a Mann–Whitney U-test (non-parametric data). A statistical comparison between three experimental groups was performed using an ordinary one-way ANOVA (normally distributed data) or a Kruskal–Wallis test (non-parametric data). The Kaplan–Meier method was used for estimating and displaying overall survival (OS) rates and onset of proteinuria. Outliers were determined via the Robust regression and outlier removal (ROUT) method and excluded in all data. To analyze the association between different parameters, Spearman’s rank correlation coefficients were calculated. In figures, asterisks denote statistical significance (*, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001; ****, p ≤ 0.0001).
Animal experiments were approved by the local governmental commission for animal protection in Freiburg (Regierungspräsidium Freiburg, approval nos. G15/164, G18/80 and G21/98).
VD deficiency is suggested to be an environmental risk factor for SLE, and the therapeutic potential of VD for SLE is being studied intensively. In light of this, we sought to explore the effects of long-term treatment with different dietary concentrations of cholecalciferol (VD) on lupus progression in female lupus-prone NZB/W F1 mice. Mice were continuously fed either a low-VD diet (<50 IU/kg; “LVD mice”), a normal-VD diet (500 IU/kg; “NVD mice”), or a high-VD diet (76,500 IU/kg; “HVD mice”) starting at 5 weeks of age. These dietary VD concentrations are based on previously published animal studies implementing comparable cholecalciferol concentrations (54–57), as well as human studies showing that high VD doses can be safely applied and may even be required to achieve VD sufficiency (58, 59). The measurement of serum concentrations of 25(OH)D3 (calcidiol), the main circulating VD metabolite, at 19–20 weeks of age revealed that the low-VD diet resulted in very low 25(OH)D3 concentrations (0–4 ng/ml; mean 2.2 ng/ml) indicative of severe VD deficiency (defined as <5 ng/ml in humans), while the normal-VD diet resulted in concentrations (22–45 ng/ml; mean: 32.9 ng/ml) that lie within the range of VD sufficiency for humans (30–60 ng/ml) and the high-VD diet produced high concentrations (90–171 ng/ml; mean: 121.2 ng/ml) in the upper range of tolerable VD levels and, in a few cases, in the lower end of the VD toxicity range for humans (>150 ng/ml) (60) (Figure 1A). Although no statistical significance was reached, our data point toward dose-dependent effects of VD on disease progression. LVD mice displayed the shortest OS and earliest onset of proteinuria, a clinical hallmark of LN (median survival: 33.0 weeks; median onset of proteinuria: 30.1 weeks), while NVD mice displayed intermediate OS and proteinuria onset (median survival: 39.9 weeks; median onset of proteinuria: 32.6 weeks) and HVD mice the longest OS and latest proteinuria onset (median survival: 41.4 weeks; median onset of proteinuria: 34.1 weeks) (Figures 1B, C). At 19–20 weeks of age, we detected significantly elevated serum anti-dsDNA IgG titers in LVD mice compared to HVD mice; in addition, there was a trend toward elevated total IgG concentrations in LVD mice. These differences were no longer observed at 28–30 weeks of age (Figures 1D, E).
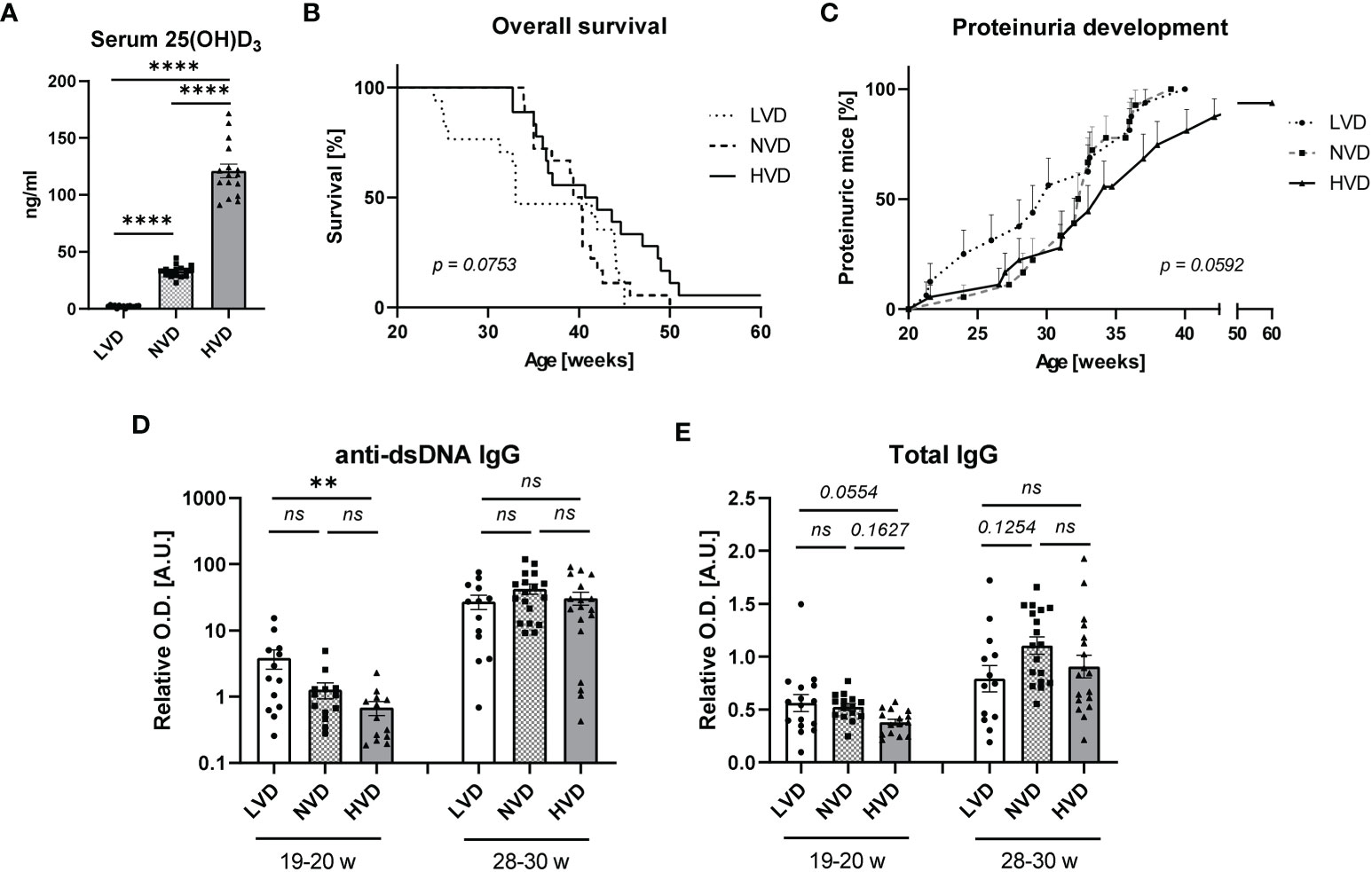
Figure 1 Low vitamin D (VD) intake, starting at 5 weeks of age, mildly accelerates disease progression in female lupus-prone NZB/W F1 mice. (A) Serum 25(OH)D3 concentrations were determined in 19–20-week-old female mice (n = 15–17), fed either a low-, normal-, or high-VD diet, with the latter containing 76,500 IU/kg. (B) Overall survival (OS) of LVD mice (n = 17), NVD mice (n = 18) and HVD mice (n = 18). The Kaplan–Meier method was used for estimating OS. Disease progression was monitored, including (C) proteinuria development, which was assessed semiquantitatively once a week, as well as serum (D) anti-dsDNA IgG and (E) total IgG titers, which were determined at an age of 19–20 weeks and 28–30 weeks in LVD mice (n = 13–15), NVD mice (n = 13–18), and (HVD mice (n = 13–18). Results are displayed as (B) survival curves, (C) time curves, and (D, E) scatter plots, with each data point representing an individual mouse in the latter. Data are expressed as (C) percents ± SEM or (D, E) mean ± SEM. P ≤ 0.05 was considered significant, p ≥ 0.2 is indicated as ns, not significant. O.D., optical density; A.U., arbitrary unit; LVD, low vitamin D; NVD, normal vitamin D; HVD, high vitamin D. ** = p < 0.01; **** = p < 0.0001
The maternal VD status during pregnancy and early-life VD exposure have been suggested to be important determinants of immune regulation and the development of immune-mediated disorders in later life (61–68). When differential VD feeding commenced at 5 weeks of age, an age at which mice can almost be considered “adolescent” (69), we observed only mild effects of VD. Therefore, we decided to additionally explore the effects of VD when already administered during gestation and infancy and continued into adulthood. Since we observed the strongest differences between LVD and HVD animals, all further experiments were performed with a comparison of low-VD versus high-VD treatments only. In experiments involving maternal VD feeding, we reduced the cholecalciferol dose of the high-VD diet to 38,000 IU/kg, to avoid VD toxicity, which still produced comparably high serum concentrations of 25(OH)D3 in HVD mice (70–155 ng/ml; mean: 104.3 ng/ml) (Supplementary Figure 1). Following this approach, we did indeed observe stronger effects, with LVD mice exhibiting significantly reduced OS and significantly earlier proteinuria onset (Figures 2A, B). At 20 weeks of age, anti-dsDNA IgG and total IgG concentrations were once again significantly reduced in LVD mice (Figures 2C, D).
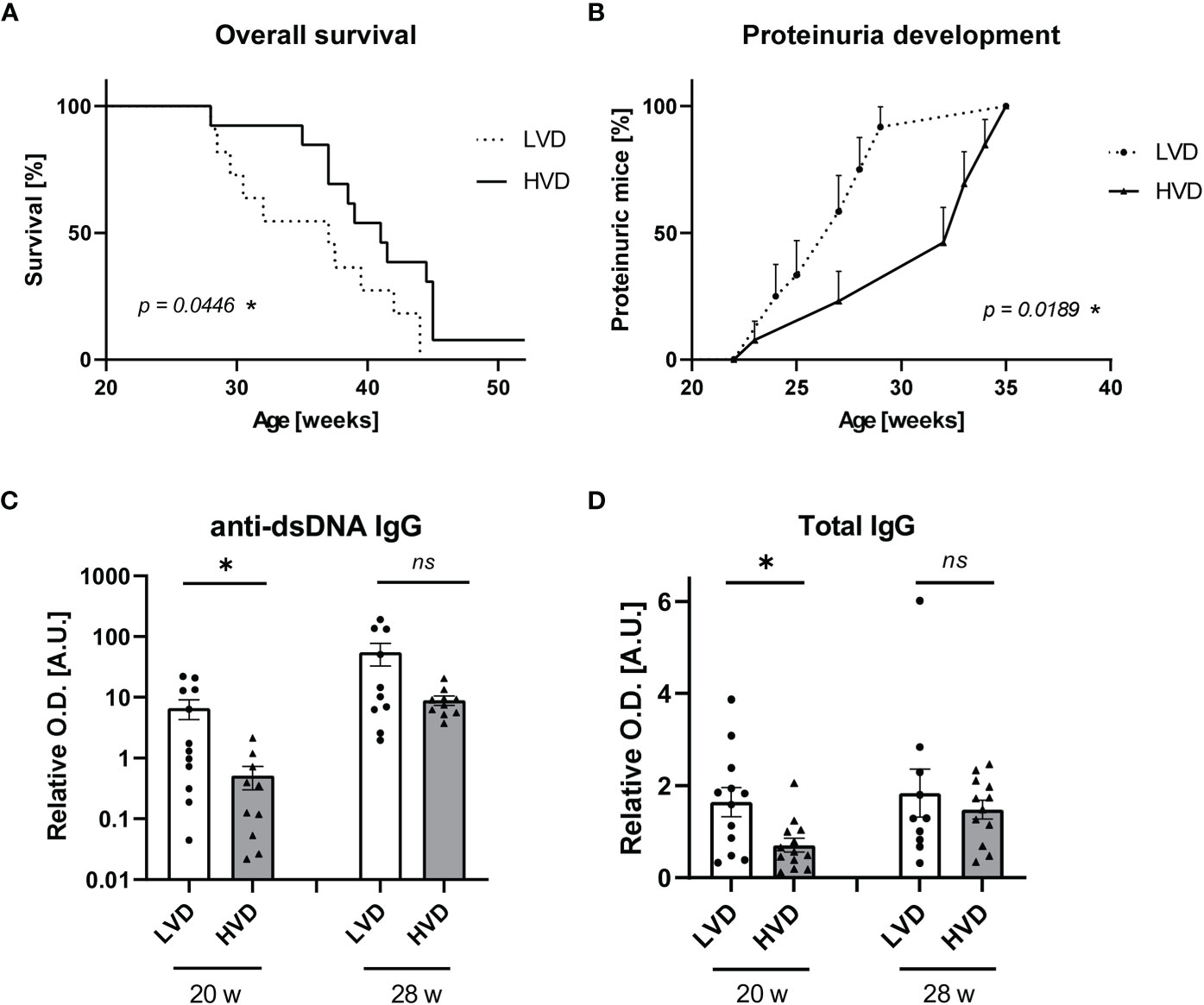
Figure 2 Low VD intake from the prenatal period onward accelerates disease progression in female lupus-prone NZB/W F1 mice. (A) OS of LVD mice (n = 11) and HVD mice (n = 13). The Kaplan–Meier method was used for estimating OS. Disease progression was monitored, including (B) proteinuria development, which was assessed semiquantitatively once a week, as well as serum (C) anti-dsDNA IgG and (D) total IgG titers, which were determined at an age of 20 and 28 weeks in LVD mice (n = 10–12), and HVD mice (n = 10–13). Results are displayed as (A) survival curves, (B) time curves, and (C, D) scatter plots, with each data point representing an individual mouse in the latter. Data are expressed as (B) percents ± SE or (C, D) mean ± SEM. P ≤ 0.05 was considered significant, p ≥ 0.2 is indicated as ns, not significant. O.D., optical density; A.U., arbitrary unit; LVD, low vitamin D; HVD, high vitamin D. * = p < 0.05
These differences in anti-dsDNA IgG were only significant at an early time point (20 weeks), when there was no relevant proteinuria detectable in the mice. At later time points, there was no difference in the concentrations of anti-dsDNA IgG and total IgG observed between the groups. This finding may indicate that beneficial effects of VD are more pronounced before the onset of disease or at the early stages of lupus. To obtain information on whether VD may also have prevented inflammation in the kidney, we performed immunofluorescence analysis for renal IgG deposits and determined TLR-7 and TLR-9 mRNA levels as a putative measure of downstream activation induced by the deposition of nucleic acid–containing immune complexes (ICs) in animals with beginning nephritis (25–26 weeks) (70, 71). We found that the concentrations of anti-dsDNA IgG in serum correlated with the amount of IgG deposits in kidneys, while no association was found between IC deposition or TLR-7/TLR-9 levels and proteinuria onset (Supplementary Figures 3, 4). Thus, it may be speculated that other, possibly VD-associated factors, than IC deposition influence the progression of nephritis and proteinuria.
To confirm that the reduced OS in LVD mice was not due to aberrant bone metabolism as a result of VD deficiency, bone density and grip strength were measured, revealing no significant differences between LVD and HVD mice (Supplementary Figure 5). Serum VD levels increased further upon longer exposure to the HVD diet, as shown when comparing the serum 25(OH)D3 levels of young (40–70 ng/ml; mean: 55.0 ng/ml) and older (70–155 ng/ml; mean: 104.3 ng/ml) HVD animals Supplementary Figure 1). These data suggest the accumulation of VD in the mice over time. However, HVD mice did not show signs of hypercalcemia due to VD intoxication, such as more frequent urination or increased water consumption, reduced/changed mobility indicative of fatigue or bone pain, weight loss, or reduced food intake indicative of reduced appetite or vomiting (data not shown).
Altogether, these results suggest detrimental effects of low VD intake on lupus progression in NZB/W F1 mice, with a stronger effect arising as a result of additional low maternal VD intake during gestation and lactation. The positive correlation between serum VD levels and age at proteinuria development as well as OS and the negative association with anti-dsDNA IgG support the dose-dependent beneficial effects of VD (Supplementary Figure 2). The fact that no such correlation was found within the individual groups indicates that rather global changes in the VD status matter, such as the presence of VD deficiency or not, while subtle changes within a high or low VD serum level range only have a minor impact (Supplementary Table 2).
In search of potential immunological effects of low VD intake that may contribute to the accelerated lupus progression in LVD mice, we compared the distribution, differentiation, and phenotype of adaptive and innate immune cell populations between LVD and HVD mice. This analysis was performed in the experiment where mice received either a low- or high-VD diet from the prenatal period onward, considering that under these conditions, effects on disease progression were most pronounced. We primarily explored these immunological parameters in 12–15-week-old male NZB/W F1 animals that do not display signs of lupus-like disease, since a skewed immune status, due to disease activity, is unlikely in these mice. This facilitates the identification of truly VD-mediated immunological effects. Considering that SLE patients have altered immune cell subset distribution, we additionally performed this immunological examination in 25–26-week-old female animals with established autoantibodies and beginning proteinuria, as these results might be of higher relevance for an active disease state.
Upon examination of circulating blood cell populations, we found a significantly higher frequency of eosinophils in young healthy LVD mice, while total circulating granulocytes in these mice were unaffected (Figure 3A). However, this effect had vanished in 25–26-week-old female animals with beginning proteinuria (Figure 3B). While LVD females displayed elevated RBC counts and a higher frequency of circulating lymphocytes, in yet-healthy LVD males, RBCs, total WBCs, and other WBC subtypes were unchanged in the blood compared to HVD mice. (Supplementary Figure 6; Supplementary Table 3)
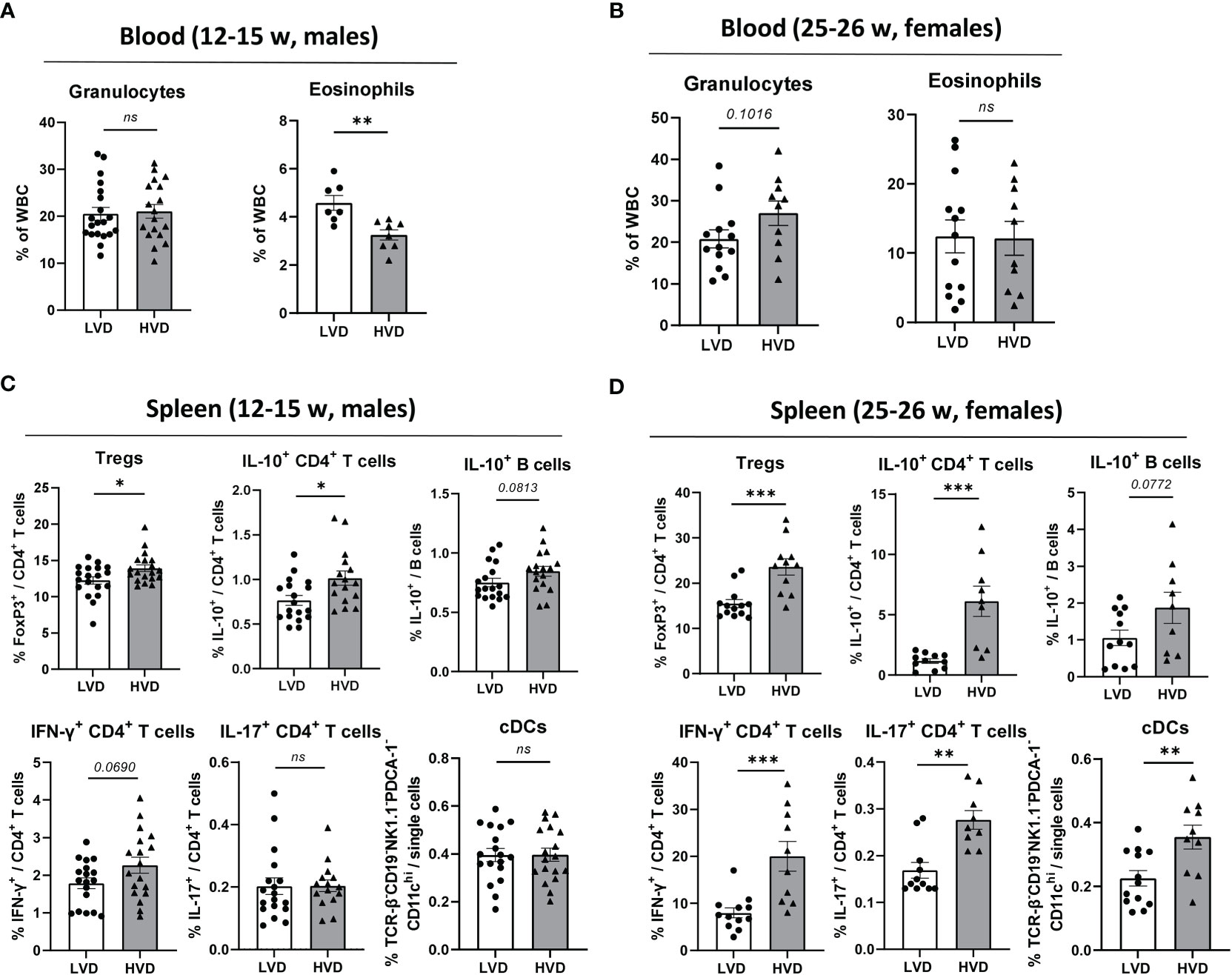
Figure 3 Low VD intake consistently hampers the adoption of a regulatory phenotype in lymphocytes. (A, B) Hematology analysis of granulocytes and eosinophils in the peripheral blood of (A) 12–15-week-old male and (B) 25–26-week-old female NZB/W F1 LVD and HVD mice. (C, D) Frequencies of Tregs, IL-10+ CD4+ T cells, IL-10+ B cells, IFN-γ+ CD4+ T cells, IL-17+ CD4+ T cells, and cDCs in the spleen of (C) 12–15-week-old LVD and HVD male NZB/W F1 mice, as well as (D) 25–26-week-old LVD and HVD female NZB/W F1 mice. Results are displayed as scatter plots, with each data point representing an individual mouse. Data are expressed as mean ± SEM. N/group: males = 7–20; females = 9–13. P ≤ 0.05 was considered significant, p ≥ 0.2 is indicated as ns, not significant. Tregs, regulatory T cells; cDCs, conventional dendritic cells; LVD, low vitamin D; HVD, high vitamin D. * = p < 0.05; ** = p < 0.01; **** = p < 0.0001.
As previously described (53), the frequencies of B cells, plasma cells, CD4+ and CD8+ T cells, CD4+ T cell differentiation into Tregs and TFH cells, IL-10 expression in B cells, and IL-10, IL-17A and IFN-γ expression in CD4+ and CD8+ T cells were explored in the spleen via flow cytometry. Within the compartment of innate immune cells, the frequencies of CD11b+ monocytic cells, conventional dendritic cells (cDCs), plasmacytoid dendritic cells (pDCs), NK cells and neutrophils were determined in the spleen (Table 1). The only consistent effects we could identify in both healthy male and in early-diseased female mice were the significantly decreased frequencies of Tregs and IL-10+ CD4+ T cells in LVD mice as well as a trend of lower frequencies of IL-10+ B cells, suggesting that low VD intake reduces the differentiation of these lymphocyte populations (Figures 3C, D). Low VD intake did not result in any shifts in the distribution of main innate immune cell populations in the spleen of male animals, and only LVD females displayed decreased frequencies of cDCs (Table 1; Supplementary Figure 7; Figure 3D). The frequencies of other effector T cell subsets, including IL-17+ CD4+ T cells, TFH cells, and IFN-γ+ CD4+ and CD8+ T cells, were unchanged between LVD and HVD males, which was also true for B-cell subsets such as germinal center (GC) B cells and plasma cells. Among females, however, HVD mice displayed significantly increased frequencies of IL-17+ CD4+ T cells, as well as IFN-γ+ CD4+, and CD8+ T cells (Table 1; Figure 3D).
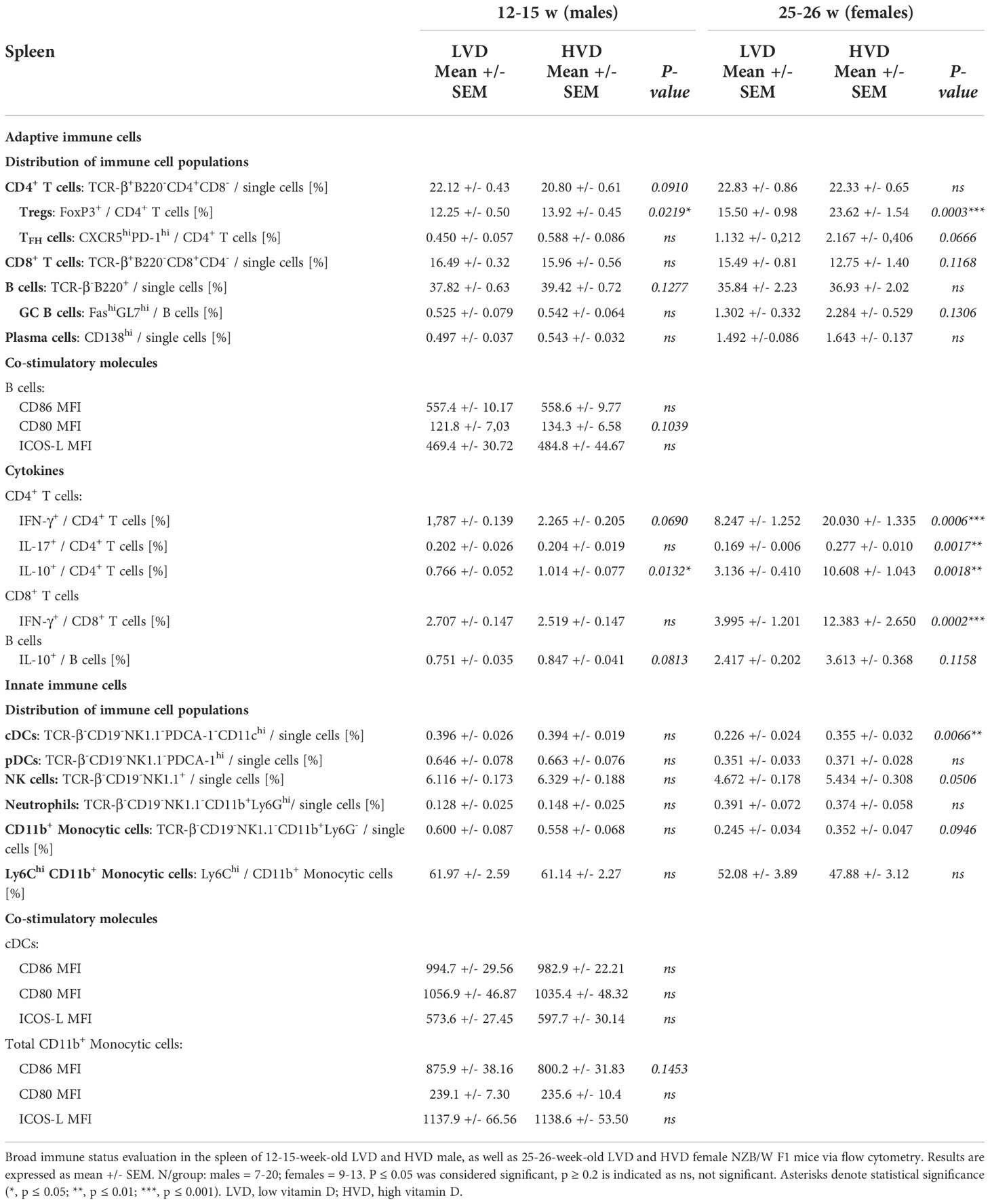
Table 1 Immunological changes in spleen of LVD and HVD mice.
To summarize, these data reveal the consistent inhibitory effects of low VD intake on the differentiation of regulatory lymphocyte populations and sporadic somewhat pro-inflammatory effects in early-diseased animals; the latter might have been provoked by the long-term intake of high VD doses or the inflammatory milieu.
Innate immune cells are reported to play a significant role in lupus pathogenesis, with their phagocytic capacity and cytokine production having been shown to be abnormal in SLE patients (4–10). An increased production of pro-inflammatory cytokines is also characteristic of innate immune cell reprogramming, termed “trained immunity,” induced by not only infectious but also sterile triggers such as Western dietary habits (72–76).
Having observed no differences in the distribution of main innate immune cells between LVD and HVD mice (Table 1; Supplementary Figure 7), we examined in more detail the impact of low VD intake on their functionality and the potential adoption of a trained immunity phenotype. To that end, as reported by Christ et al. (76), we investigated whether our dietary modification would affect the expression of costimulatory molecules and cytokine production in innate immune cells from different sites in young male mice. The production of the pro-inflammatory cytokines TNF-α, IL-1ß, and IL-6 in purified splenic CD11b+ innate immune cells and peritoneal macrophages was explored by quantitative reverse transcription PCR, after short-term stimulation with LPS (Supplementary Figure 13). In addition, we explored cytokine production in BM cells after 5-day differentiation into BM-derived innate immune cells, comprising a mix of dendritic cells and macrophages, and BM-derived macrophages (BMDMs), via the addition of GM-CSF or M-CSF, respectively (77, 78). Here, the secretion of TNF-α, IL-1ß, and IL-10 was determined by ELISA after 20 h stimulation with a panel of TLR ligands. Purified splenic CD11b+ innate immune cells displayed significantly increased TNF mRNA levels and a trend of elevated IL1B mRNA levels in LVD mice (Figure 4A), while this effect was not reflected in purified peritoneal macrophages (Figure 4B). In contrast, we found no significant differences in cytokine secretion by BM-derived innate immune cells or BMDMs between LVD and HVD mice (Figures 4C, D).
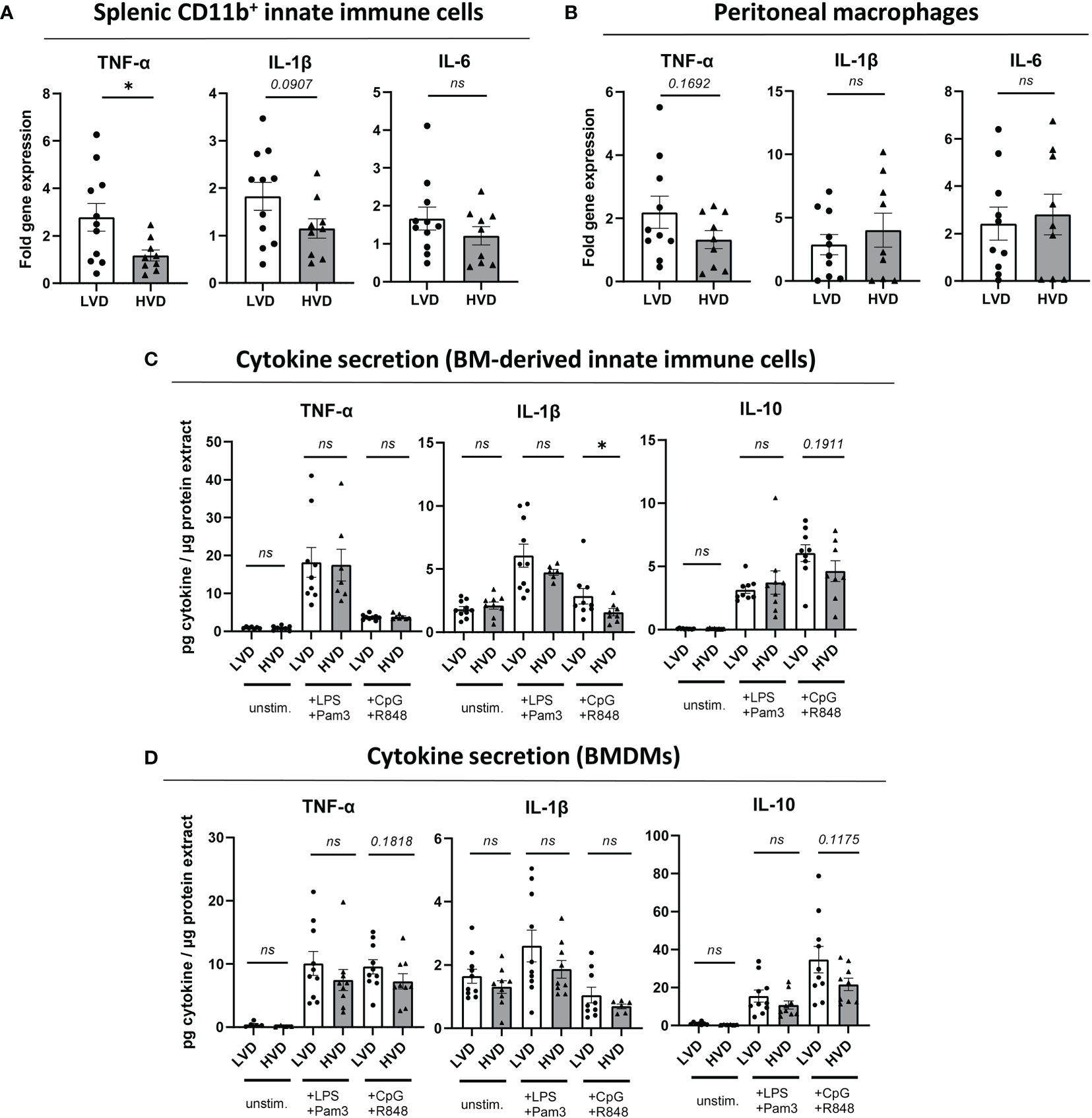
Figure 4 Low VD intake has no consistent effects on cytokine production in innate immune cells. (A, B) Fold gene expression of pro-inflammatory cytokines, following stimulation with 50 ng/ml LPS for 2 h 45 min, in (A) splenic CD11b+ innate immune cells and (B) peritoneal macrophages of 12–15-week-old male NZB/W F1 mice. (C, D) Cytokine levels in culture supernatants of (C) BM-derived innate immune cells (+ GM-CSF) and (D) BMDMs (+ M-CSF) from 12–15-week-old male NZB/W F1 mice, following stimulation with either 50 ng/ml LPS and 50 ng/ml Pam3CSK4, or 1 µM CpG and 1 µM R848, for 20 h, normalized to total protein content of adherent cells. Results are displayed as scatter plots, with each data point representing an individual mouse. Data are expressed as mean ± SEM. N/group: LVD = 9-11; HVD = 6-9. P ≤ 0.05 was considered significant, p ≥ 0.2 is indicated as ns, not significant. BM, bone marrow; BMDMs, BM-derived macrophages; Pam3, Pam3CSK4; unstim., unstimulated; LVD, low vitamin D; HVD, high vitamin D. * = p < 0.05
Trained immunity has also been reported to entail an increased expression of costimulatory molecules (75, 76, 79, 80). Thus, we measured the surface expression of CD86, CD80, and ICOS-L on splenic cDCs and CD11b+ monocytic cells, circulating CD11b+ monocytic cells and peritoneal macrophages. We observed no differences between LVD and HVD mice (Supplementary Figure 9). In support of this, we also found no increase in the frequencies of splenic or circulating Ly6ChiCD11b+ monocytic cells, which are considered pro-inflammatory and have been reported to be increased following the induction of trained immunity (Supplementary Figure 10A) (81–84).
Impaired phagocytosis of apoptotic cells by macrophages has been implicated in SLE pathogenesis. Therefore, we next determined the effect of low VD intake on the phagocytosis of apoptotic cells by both peritoneal macrophages and BMDMs, the latter being generated by a 5-day culture of BM cells with M-CSF. Phagocytosis was measured by co-culturing CD45.2+ macrophages with CD45.1+ CFDA-SE-labeled apoptotic thymocytes (ATs) and quantifying phagocytes that had phagocytosed ATs as CD45.2+CD45.1-CD11b+F4/80+CFDA-SE+ cells via flow cytometry (Supplementary Figure 14). As CD45.2+CD45.1+CFDA-SE+ cells represent cells with non-ingested ATs attached to their surface, these cells were not counted. In addition to measuring phagocytic capacity, we examined the expression of surface markers associated with phagocytosis. Between LVD and HVD mice, there were no differences in the phagocytic capacity of peritoneal macrophages or BMDMs, or the surface expression of phagocytosis-associated markers on these cells (Figure 5).
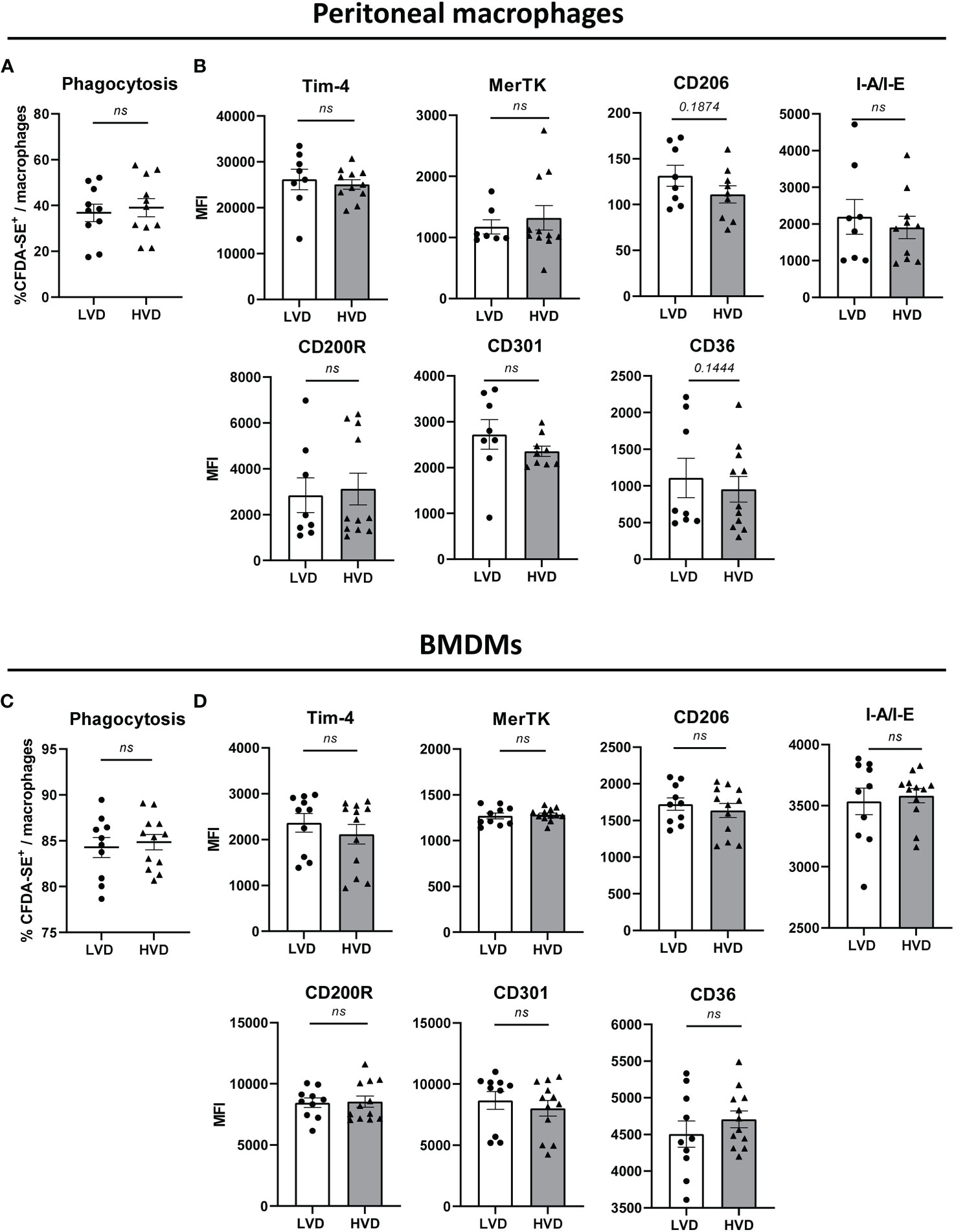
Figure 5 Low VD intake has no effect on phagocytic capacity and surface expression of phagocytosis-associated markers in macrophages. (A, C) Phagocytic uptake of apoptotic thymocytes by (A) peritoneal macrophages and (C) BM-derived macrophages (BMDMs) of 12–15-week-old LVD (n = 10) and HVD (n = 11-12) male animals. (B, D) Surface expression of phagocytosis-associated markers by (B) peritoneal macrophages and (D) BMDMs of 12–15-week-old LVD (n = 7-10) and HVD (n= 10-12) male animals. Results are displayed as scatter plots, with each data point representing an individual mouse. Data are expressed as mean ± SEM. P ≤ 0.05 was considered significant, p ≥ 0.2 is indicated as ns, not significant. MFI, mean fluorescence intensity; BMDMs, BM-derived macrophages; LVD, low vitamin D; HVD, high vitamin D.
Together, these data do not suggest stringent effects of low VD intake on the phenotype and function of innate immune cells, including cytokine production, costimulatory molecule expression, and phagocytic capacity.
Environmental factors are becoming increasingly acknowledged as triggers of the development of autoimmune diseases such as SLE. Since VD is known to exert immunomodulatory effects and there is a high prevalence of VD deficiency among SLE patients, VD deficiency has been thoroughly contemplated as a risk factor for SLE (21–27). In this study, we explored the effects of dietary VD on lupus-prone NZB/W F1 mice and attempted to identify the immunological effects of low VD intake that could promote lupus progression. We report that low VD intake accelerates lupus progression, with this effect becoming significant when low VD intake commenced during the prenatal period. Our data reveal that low VD intake reduces the differentiation of immunoregulatory lymphocyte populations. Thus, these data add to the understanding of how VD deficiency may play a role in promoting lupus development.
While many studies demonstrate an inverse relationship between VD levels and disease activity in SLE patients (85–87), the collective data from interventional studies investigating the effects of oral VD supplementation on SLE disease activity are inconsistent (88–91). Contradictory results may, in part, be due to variations in the administered cholecalciferol dose, study design, and placebo group, as well as different treatment durations. The age groups of cohorts also differ between studies, since some studies investigate the effect of oral VD supplementation in pediatric or juvenile patients, while most others involve adults. Age-dependent effects were also addressed in our study. Our data demonstrating the detrimental effects of low VD feeding on lupus progression did not achieve statistical significance when fed from an age of 5 weeks onward, in contrast to when low VD intake commenced during the prenatal period. This suggests that the maternal VD status during gestation may play a role in autoimmune development in offspring. Some, but not all studies, support the notion of a role of the maternal VD status during pregnancy in the risk of the development of immune-mediated diseases in offspring (61–66, 92–96). VD is known to be a potent epigenetic modulator, and epigenetic abnormalities have been reported in SLE (32, 33, 97, 98). Interestingly, both human and mouse studies have demonstrated that the maternal VD status during pregnancy affects DNA methylation in offspring, including genes involved in apoptosis regulation and immune function (99, 100). Furthermore, it has been suggested that the effectiveness of an individual’s response to VD can vary and that individuals can be categorized as high-, mid-, or low-VD responders (101, 102). Accordingly, there is some dispute on the desired VD status and the recommended daily intake. Our results support the recommendations of sufficient daily intake of VD and the relevance of avoiding VD deficiency. However, and as discussed below, our data and others also show that it needs to be considered that very high VD levels over a longer period may not only bear the risk of hypercalcemia but also, even below toxic levels, they might induce somewhat pro-inflammatory changes with potentially adverse effects on the progression of autoimmune pathology (54, 103). As this was not reported by other trials (58, 59), more work is required to elucidate the relevance and impact of VD dosage in health and disease.
To the best of our knowledge, there have been no studies investigating the effect of dietary cholecalciferol feeding on murine lupus pathology. As an alternative to oral cholecalciferol treatment, many preclinical studies employ the application of calcitriol, the active VD metabolite. Calcitriol application may generally induce stronger effects than oral cholecalciferol supplementation, since direct calcitriol application bypasses regulatory steps within physiological VD metabolism (104, 105). Two studies involving lupus-prone MRL/l mice receiving calcitriol treatment show effectiveness in ameliorating disease (106, 107). We also explored the effects of continuous calcitriol injections in female NZB/W F1 mice from 5 weeks of age onward and found that it neither improved OS and proteinuria, nor inhibited autoantibody production (data not shown). As calcitriol application resulted in slight weight loss and a mild deterioration of general health in some animals, interfering side effects by the continuous application were likely. Therefore, a long-term treatment of mice with calcitriol may entail difficulties, in contrast to cholecalciferol, which has a wider therapeutic index.
We observed that low VD intake resulted in elevated anti-dsDNA IgG and total IgG concentrations. This finding is in concordance with a study by Terrier et al. that reported a significant reduction in anti-dsDNA levels in VD-deficient SLE patients, following an oral cholecalciferol supplementation regimen (108). The fact that we observed no significant difference in autoantibody titers between the groups at 28 weeks of age may suggest that VD primarily delays the rise in autoantibody titers during early disease pathogenesis (20 weeks) and that later during pathogenesis (28 weeks), the moderate effects of VD are no longer sufficient to dampen the production of autoantibodies. Indeed, it has been observed in the context of a different experimental model that the immunoregulatory effects of VD are overridden by strong immune activity (109). As discussed below, it could also be speculated that increasingly high VD levels might, over time, weaken the initially preponderant immunoregulatory effects. Alternatively, increased proteinuria in 28-week-old LVD mice may result in the increased renal loss of immunoglobulins, thereby resulting in the decreased serum concentrations of anti-dsDNA as well as total IgG.
Alongside autoantibody-mediated IC deposition, there are other factors that contribute to the development of nephritis and proteinuria that may be regulated by VD. Indeed, VD and VD analogs have various antiproteinuric and anti-inflammatory effects on the kidneys (110–115). Thus, it is possible that the accelerated proteinuria development we observed in LVD mice was due to a combination of elevated autoantibody titers and a lack of VD-mediated nephroprotective effects. Some data exist on the effects of VD in the context of LN; a recent study put the nephroprotective effects of VD to the test and reported that in vitro calcitriol treatment protects podocytes from autoantibody-induced injury by reducing aberrant autophagy (116). Moreover, the treatment of MRL/l mice with a calcitriol analog inhibits the development of LN (117). Mechanistically, VD was reported to exert its antiproteinuric effect by inhibiting the expression of heparanase and by modulating the renin–angiotensin–aldosterone system (110, 111). In addition, immunological effects of VD and VD analogs have been reported in mouse models of renal fibrosis and obstructive nephropathy, such as a reduction of IL-6 and IL-1β in the kidney and reduced T cell and macrophage infiltration (113, 115). We also examined the immunological effects of low VD intake, starting at 5 weeks of age, in the kidneys of 27-week-old female animals with beginning nephritis, but observed no significant differences between LVD and HVD mice in the frequencies of immune cells (data not shown). In contrast to our data and numerous studies demonstrating the nephroprotective effects of VD, Vaisberg et al. reported a worsening of histopathological findings in the kidneys of female NZB/W F1 mice receiving long-term intraperitoneal (i.p.) cholecalciferol treatment (118). The possible explanations for these contradictory results remain elusive. We can only speculate that application route or dose might play a role.
Generally, our results suggest that VD could have beneficial effects on the early disease phase and delay its onset, while during a later stage, it may limit nephritis progression. The beneficial impact on the early disease phase is supported by reduced anti-dsDNA IgG in 20-week-old HVD female mice and the increased frequencies of regulatory lymphocyte subsets in yet-healthy HVD animals. Further, our data do not prove, but indirectly suggest, that other factors than IC deposition might influence the progression of nephritis and proteinuria, such as renal VD-mediated effects, since we found a correlation between anti-dsDNA IgG in serum and IgG deposition in the kidney but no association between IgG deposition or renal TLR-7/TLR-9 levels and proteinuria onset. However, some limitations need to be considered; immunofluorescence only allows a semiquantitative analysis of IC deposition. Moreover, the disease activity of LN may be influenced by other factors than IC deposition, such as IC composition, autoantibody subclass as well as glycosylation pattern, renal cellular composition, and cellular activation and differentiation status. The exact significance of the causative relationship between IC quantity versus other factors and nephritis progression is not clearly established. As these indications are only indirect, future studies should address in more detail the important question at which phase of disease pathogenesis exactly VD exerts its beneficial effects.
Given the effects of low VD intake on autoantibody production and lupus progression, we suspected immunological effects. We observed an increase in the frequency of circulating eosinophils in yet-healthy LVD mice, while the total circulating granulocytes in these mice were not elevated, suggesting increased eosinophil differentiation. This is supported by human studies demonstrating a negative correlation between serum VD and blood eosinophil levels (119–121). However, in SLE, the role of eosinophils has hardly been studied. While there are a small number of case studies of patients with SLE and various other autoimmune disorders displaying hypereosinophilic syndrome (HES) (122–126), it is not evident whether HES precedes SLE or vice versa. Similarly, the role of eosinophils in LN is not well explored. One study reported an increased frequency of eosinophiluria in LN patients compared to SLE patients without renal involvement (127). However, this does not necessarily implicate eosinophils in LN pathogenesis. On the other hand, in another study on patients with various immune and non-immune kidney diseases, peripheral eosinophilia was associated with increased renal eosinophil infiltration and progression to end-stage kidney disease (128). Since we no longer observed elevated levels of circulating eosinophils in females with beginning nephritis, it is possible that this could indeed be due to increased renal eosinophil infiltration, although this needs to be investigated. As eosinophils are potent producers of the plasma cell survival factor APRIL (129, 130), one could further speculate about them representing an important factor in sustaining lupus and nephritis progression.
As the most consistent immunological effect occurring in both healthy animals and those with beginning disease, we observed an inhibitory effect of low VD intake on the differentiation of regulatory lymphocyte populations, encompassing Tregs and IL-10-producing lymphocytes. This is in concordance with previous studies showing that VD promotes Treg differentiation and augments IL-10 production in CD4+ T cells and B cells (42–44, 51, 57, 131). This is relevant in the context of lupus since Tregs and IL-10 play an established and important immunoregulatory role in constraining the development of autoimmunity (132–139). Furthermore, IL-10-producing putative regulatory B cells have also been reported to exert protective effects in lupus-prone mice (140, 141). However, IL-10 possesses context-dependent dual functions that can also involve pro-inflammatory effects and may support antibody responses (142–144). Only in female mice with beginning disease did we note sporadic pro-inflammatory immunological effects in the HVD group, including the increased frequencies of cDCs, IL17+ and IFN-γ+ T cells. Considering that most studies have reported the ability of VD to lower the expression of pro-inflammatory cytokines, such as IFN-γ (145–147), these results were surprising at first. On the other hand, some studies report enhancing effects of VD on IFN-γ expression (148, 149). In our study, the significance and cause of the sporadic pro-inflammatory changes observed in diseased HVD mice remain unresolved. A possible explanation might be that inflammation and the disease context could have modulated the effects of VD on immune cells. In addition, the adverse effects of very high and long-term serum 25(OH)D3 concentrations in the upper tolerable range cannot be excluded. In support of such a scenario, Häusler et al. have shown that mice supplemented with high doses of cholecalciferol and displaying high serum VD levels developed fulminant experimental autoimmune encephalomyelitis (EAE) with a massive infiltration of the central nervous system by activated myeloid cells, TH1 and TH17 cells. In contrast, moderate supplementation reduced EAE severity along with an expansion of Tregs (54). Considering that VD seemed to successively accumulate in our mice fed the high-VD diet, there may be similar effects explaining our observations; in young HVD mice displaying serum VD levels in a healthy range, only an increase in putatively beneficial immunoregulatory lymphocyte subsets was found, while in older HVD mice with markedly elevated serum VD levels, additional and putatively pro-inflammatory immune effects comparable to those reported by Häusler et al. (54) were noted. As high VD intake still conferred beneficial effects on disease pathology in our study, we suspect that beneficial high VD effects may have outweighed pro-inflammatory effects occurring at later disease stages. This may also explain why serum anti-dsDNA IgG was only decreased in HVD mice at 20 but not 28 weeks of age. Further, and as discussed above, VD may exert additional nephroprotective effects limiting nephritis progression. This underlines the importance of exploring the influence of VD dosage, as still, it is not fully resolved whether normal or rather high-normal serum VD levels should be aimed for to reach optimal beneficial effects in inflammatory immune-mediated diseases and autoimmunity (103, 150–152). To address these questions, future studies should monitor more closely the development of disease and immunological changes in relation to serum VD levels and the intake of different VD doses. Thus, in retrospect, one limitation of our study is that immunological effects have not additionally been explored for the NVD group.
We also explored the effects of VD on the phenotype and function of innate immune cells in more detail. Dysregulated production of pro-inflammatory cytokines plays a central role in autoimmunity. An increased production of pro-inflammatory cytokines is characteristic of a trained immunity phenotype in innate immune cells, and it has been argued that monocytes and macrophages from patients with autoimmune disease display such features (8, 9, 72). Alongside infectious triggers, environmental factors can also induce such reprogramming of innate immune cells, as previously shown by a study on myeloid cells from mice fed a Western diet (76). We therefore sought to investigate whether low VD intake similarly affects cytokine production, since VD regulates the epigenome and epigenetic rewiring underlies the adoption of a trained immunity phenotype (153, 154). We examined cytokine production by various innate immune cells but only observed increased pro-inflammatory cytokine production in splenic CD11b+ innate immune cells of LVD mice. In accordance with the overall inconsistent effects of VD on the cytokine milieu, we did not observe any differences between LVD and HVD mice in the upregulation of costimulatory molecules in innate immune cells, which is another feature of trained immunity (75, 76, 79, 80). In summary, our data demonstrate the effects of VD on the cells of the adaptive immune system but do not suggest a major impact on the cytokine production, phagocytic capacity, or activation status of innate immune cells in the employed setting.
In conclusion, low VD intake has detrimental effects on the manifestation and progression of lupus-like disease in NZB/W F1 mice. Since SLE is a multifactorial disease with an important contribution of environmental factors to disease manifestation, it is likely that VD deficiency represents an important environmental factor that may tip the balance toward the development of manifest autoimmunity. Therefore, adequate VD intake remains important in SLE, especially for VD-deficient patients. Moreover, sufficient VD intake may delay or prevent the manifestations of SLE in individuals at risk, such as relatives of SLE patients, especially if they are positive for antinuclear antibodies. Further research is necessary to better understand the immunological mechanisms underlying the effects of VD in lupus and to determine optimal VD dosage and serum concentrations.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
The animal study was reviewed and approved by local governmental commission for animal protection of Freiburg (Regierungspräsidium Freiburg, approval nos. G15/164, G18/80 and G21/98).
NC, REV, ANK, A-LS, and DTLS designed research. NC, ANK, A-LS, DTLS, JPW, LR, QA, AL, ZW, and KH performed research. NC, ANK, A-LS, DTLS, J-DF, MHe, and BS analyzed data. HD and MHo contributed new reagents/analytical tools. NC, ANK, A-LS, DTLS, BS, JPW, LR, QA-K, HD, KH, MHo, MHe, GH, and REV contributed to manuscript editing. ANK and NC wrote the paper. All authors contributed to the article and approved the submitted version.
This work was supported by the Ministry of Science, Research, and Arts Baden-Wurttemberg (Margarete von Wrangell Programm to NC and MHo), Deutsche Forschungsgemeinschaft (DFG) (TRR 130, project 12 to RV and NC and project 19 to GH; SFB 1160 project 12 to RV and project A02 to MHo), and the Erika Bürgy Fundation (Stiftung für die Region – Sparkasse Pforzheim Calw Treuhandstiftung Erika Bürgy Stiftung) to RV, as well as the Medical Faculty of the University of Freiburg (MOTI-VATE fellowship to JW and AL). This project was cofinanced by the European Regional Development Fund (ERDF) of the European Union in the framework of the INTERREG V Upper Rhine programme (project PERSONALIS).
We thank all staff of the animal facility (CEMT) for help with animal care and husbandry and Rita Rzepka and Ingeborg Wünsche for organizational and infrastructural measures. The bone density measurement was performed in the Institute of Radiology, Preclinical Imaging Platform Erlangen (PIPE), Friedrich-Alexander Universität Erlangen and was supported by DFG SFB1181 project Z02 to Tobias Bäuerle.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.933191/full#supplementary-material
1. Tsokos GC, Lo MS, Reis PC, Sullivan KE. New insights into the immunopathogenesis of systemic lupus erythematosus. Nat Rev Rheumatol (2016) 12(12):716–30. doi: 10.1038/nrrheum.2016.186
2. Podolska MJ, Biermann MH, Maueröder C, Hahn J, Herrmann M. Inflammatory etiopathogenesis of systemic lupus erythematosus: An update. JIR (2015) 8:161–71. doi: 10.2147/JIR.S70325
3. Jaryal A, Vikrant S. Current status of lupus nephritis. Indian J Med Res (2017) 145(2):167–78. doi: 10.4103/ijmr.IJMR_163_16
4. Gaipl US, Voll RE, Sheriff A, Franz S, Kalden JR, Herrmann M. Impaired clearance of dying cells in systemic lupus erythematosus. Autoimmun Rev (2005) 4(4):189–94. doi: 10.1016/j.autrev.2004.10.007
5. Herrmann M, Voll RE, Zoller OM, Hagenhofer M, Ponner BB, Kalden JR. Impaired phagocytosis of apoptotic cell material by monocyte-derived macrophages from patients with systemic lupus erythematosus. Arthritis Rheumatism (1998) 41(7):1241–50. doi: 10.1002/1529-0131(199807)41:7<1241::AID-ART15>3.0.CO;2-H
6. Ren Y, Tang J, Mok MY, Chan AWK, Wu A, Lau CS. Increased apoptotic neutrophils and macrophages and impaired macrophage phagocytic clearance of apoptotic neutrophils in systemic lupus erythematosus. Arthritis Rheumatism (2003) 48(10):2888–97. doi: 10.1002/art.11237
7. Baumann I, Kolowos W, Voll RE, Manger B, Gaipl U, Neuhuber WL, et al. Impaired uptake of apoptotic cells into tingible body macrophages in germinal centers of patients with systemic lupus erythematosus. Arthritis Rheumatism (2002) 46(1):191–201. doi: 10.1002/1529-0131(200201)46:1<191::AID-ART10027>3.0.CO;2-K
8. Byrne JC, Ní Gabhann J, Lazzari E, Mahony R, Smith S, Stacey K, et al. Genetics of SLE: Functional relevance for Monocytes/Macrophages in disease. Clin Dev Immunol (2012) 2012:e582352. doi: 10.1155/2012/582352
9. Steinbach F, Henke F, Krause B, Thiele B, Burmester GR, Hiepe F. Monocytes from systemic lupus erythematous patients are severely altered in phenotype and lineage flexibility. Ann Rheumatic Diseases. (2000) 59(4):283–8. doi: 10.1136/ard.59.4.283
10. Sule S, Rosen A, Petri M, Akhter E, Andrade F. Abnormal production of pro- and anti-inflammatory cytokines by lupus monocytes in response to apoptotic cells. PloS One (2011) 6(3):e17495. doi: 10.1371/journal.pone.0017495
11. Stohl W, Metyas S, Tan SM, Cheema GS, Oamar B, Xu D, et al. B lymphocyte stimulator overexpression in patients with systemic lupus erythematosus: Longitudinal observations. Arthritis Rheumatism (2003) 48(12):3475–86. doi: 10.1002/art.11354
12. Kaul A, Gordon C, Crow MK, Touma Z, Urowitz MB, van Vollenhoven R, et al. Systemic lupus erythematosus. Nat Rev Dis Primers. (2016) 2(1):1–21. doi: 10.1038/nrdp.2016.39
13. Choi JY, Ho JH, Pasoto SG, Bunin V, Kim ST, Carrasco S, et al. Circulating follicular helper–like T cells in systemic lupus erythematosus: Association with disease activity. Arthritis Rheumatol (2015) 67(4):988–99. doi: 10.1002/art.39020
14. Miyara M, Amoura Z, Parizot C, Badoual C, Dorgham K, Trad S, et al. Global natural regulatory T cell depletion in active systemic lupus erythematosus. J Immunol (2005) 175(12):8392–400. doi: 10.4049/jimmunol.175.12.8392
15. Mellor-Pita S, Citores MJ, Castejon R, Tutor-Ureta P, Yebra-Bango M, Andreu JL, et al. Decrease of regulatory T cells in patients with systemic lupus erythematosus. Ann Rheumatic Diseases. (2006) 65(4):553–4. doi: 10.1136/ard.2005.044974
16. Kamen DL, Cooper GS, Bouali H, Shaftman SR, Hollis BW, Gilkeson GS. Vitamin d deficiency in systemic lupus erythematosus. Autoimmun Rev (2006) 5(2):114–7. doi: 10.1016/j.autrev.2005.05.009
17. Cutolo M, Otsa K. Review: Vitamin d, immunity and lupus. Lupus (2008) 17(1):6–10. doi: 10.1177/0961203307085879
18. Song GG, Bae SC, Lee YH. Association between vitamin d intake and the risk of rheumatoid arthritis: A meta-analysis. Clin Rheumatol (2012) 31(12):1733–9. doi: 10.1007/s10067-012-2080-7
19. Duan S, Lv Z, Fan X, Wang L, Han F, Wang H, et al. Vitamin d status and the risk of multiple sclerosis: A systematic review and meta-analysis. Neurosci Letters. (2014) 570:108–13. doi: 10.1016/j.neulet.2014.04.021
20. Del Pinto R, Pietropaoli D, Chandar AK, Ferri C, Cominelli F. Association between inflammatory bowel disease and vitamin d deficiency: A systematic review and meta-analysis. Inflamm Bowel Diseases. (2015) 21(11):2708–17. doi: 10.1097/MIB.0000000000000546
21. Müller K, Kriegbaum NJ, Baslund B, Sørensen OH, Thymann M, Bentzen K. Vitamin D3 metabolism in patients with rheumatic diseases: Low serum levels of 25-hydroxyvitamin D3 in patients with systemic lupus erythematosus. Clin Rheumatol (1995) 14(4):397–400. doi: 10.1007/BF02207671
22. Ruiz-Irastorza G, Egurbide MV, Olivares N, Martinez-Berriotxoa A, Aguirre C. Vitamin d deficiency in systemic lupus erythematosus: prevalence, predictors and clinical consequences. Rheumatology (2008) 47(6):920–3. doi: 10.1093/rheumatology/ken121
23. Heine G, Lahl A, Müller C, Worm M. Vitamin d deficiency in patients with cutaneous lupus erythematosus is prevalent throughout the year. Br J Dermatol (2010) 163(4):863–5. doi: 10.1111/j.1365-2133.2010.09948.x
24. Attar SM, Siddiqui AM. Vitamin d deficiency in patients with systemic lupus erythematosus. Oman Med J (2013) 28(1):42–7. doi: 10.5001/omj.2013.10
25. Toloza S, Cole D, Gladman D, Ibañez D, Urowitz M. Vitamin d insufficiency in a large female SLE cohort. Lupus (2010) 19(1):13–9. doi: 10.1177/0961203309345775
26. Bonakdar ZS, Jahanshahifar L, Jahanshahifar F, Gholamrezaei A. Vitamin d deficiency and its association with disease activity in new cases of systemic lupus erythematosus. Lupus (2011) 20(11):1155–60. doi: 10.1177/0961203311405703
27. Muñoz-Ortego J, Torrente-Segarra V, Prieto-Alhambra D, Salman-Monte T, Carbonell-Abello J. Prevalence and predictors of vitamin d deficiency in non-supplemented women with systemic lupus erythematosus in the Mediterranean region: A cohort study. Scandinavian J Rheumatol (2012) 41(6):472–5. doi: 10.3109/03009742.2012.697189
28. Foering K, Chang AY, Piette EW, Cucchiara A, Okawa J, Werth VP. Characterization of clinical photosensitivity in cutaneous lupus erythematosus. J Am Acad Dermatol (2013) 69(2):205–13. doi: 10.1016/j.jaad.2013.03.015
29. Sanders CJG, Weelden HV, Kazzaz G a. A, Sigurdsson V, Toonstra J, Bruijnzeel-Koomen CAFM. Photosensitivity in patients with lupus erythematosus: A clinical and photobiological study of 100 patients using a prolonged phototest protocol. Br J Dermatol (2003) 149(1):131–7. doi: 10.1046/j.1365-2133.2003.05379.x
30. Dankers W, Colin EM, van Hamburg JP, Lubberts E. Vitamin d in autoimmunity: Molecular mechanisms and therapeutic potential. Front Immunol (2017) 7:697. doi: 10.3389/fimmu.2016.00697
31. Pike JW, Meyer MB. The vitamin d receptor: New paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Endocrinol Metab Clinics North America. (2010) 39(2):255–69. doi: 10.1016/j.ecl.2010.02.007
32. Fetahu IS, Höbaus J, Kállay E. Vitamin d and the epigenome. Front Physiol (2014) 5:164. doi: 10.3389/fphys.2014.00164
33. Seuter S, Pehkonen P, Heikkinen S, Carlberg C. Dynamics of 1α,25-dihydroxyvitamin D3-dependent chromatin accessibility of early vitamin d receptor target genes. Biochim Biophys Acta (BBA) - Gene Regul Mechanisms. (2013) 1829(12):1266–75. doi: 10.1016/j.bbagrm.2013.10.003
34. Wang Y, Zhu J, DeLuca HF. Where is the vitamin d receptor? Arch Biochem Biophysics. (2012) 523(1):123–33. doi: 10.1016/j.abb.2012.04.001
35. Liu PT, Stenger S, Li H, Wenzel L, Tan BH, Krutzik SR, et al. Toll-like receptor triggering of a vitamin d-mediated human antimicrobial response. Science (2006) 311(5768):1770–3. doi: 10.1126/science.1123933
36. Tanaka H, Abe E, Miyaura C, Shiina Y, Suda T. 1 alpha,25-dihydroxyvitamin D3 induces differentiation of human promyelocytic leukemia cells (HL-60) into monocyte-macrophages, but not into granulocytes. Biochem Biophys Res Commun (1983) 117(1):86–92. doi: 10.1016/0006-291X(83)91544-9
37. Bar-Shavit Z, Teitelbaum SL, Reitsma P, Hall A, Pegg LE, Trial J, et al. Induction of monocytic differentiation and bone resorption by 1,25-dihydroxyvitamin D3. PNAS (1983) 80(19):5907–11. doi: 10.1073/pnas.80.19.5907
38. Griffin MD, Lutz W, Phan VA, Bachman LA, McKean DJ, Kumar R. Dendritic cell modulation by 1α,25 dihydroxyvitamin D3 and its analogs: A vitamin d receptor-dependent pathway that promotes a persistent state of immaturity in vitro and in vivo. PNAS (2001) 98(12):6800–5. doi: 10.1073/pnas.121172198
39. Penna G, Adorini L. 1α,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol (2000) 164(5):2405–11. doi: 10.4049/jimmunol.164.5.2405
40. Ferreira GB, Gysemans CA, Demengeot J, da CJPMCM, AS V, Overbergh L, et al. 1,25-dihydroxyvitamin D3 promotes tolerogenic dendritic cells with functional migratory properties in NOD mice. J Immunol (2014) 192(9):4210–20. doi: 10.4049/jimmunol.1302350
41. Ferreira GB, Vanherwegen AS, Eelen G, Gutiérrez ACF, Van Lommel L, Marchal K, et al. Vitamin D3 induces tolerance in human dendritic cells by activation of intracellular metabolic pathways. Cell Rep (2015) 10(5):711–25. doi: 10.1016/j.celrep.2015.01.013
42. Urry Z, Chambers ES, Xystrakis E, Dimeloe S, Richards DF, Gabryšová L, et al. The role of 1α,25-dihydroxyvitamin D3 and cytokines in the promotion of distinct Foxp3+and IL-10+ CD4+ T cells. Eur J Immunol (2012) 42(10):2697–708. doi: 10.1002/eji.201242370
43. Kang SW, Kim SH, Lee N, Lee WW, Hwang KA, Shin MS, et al. 1,25-dihyroxyvitamin D3 promotes FOXP3 expression via binding to vitamin d response elements in its conserved noncoding sequence region. J Immunol (2012) 188(11):5276–82. doi: 10.4049/jimmunol.1101211
44. Zhou Q, Qin S, Zhang J, Zhon L, Pen Z, Xing T. 1,25(OH)2D3 induces regulatory T cell differentiation by influencing the VDR/PLC-γ1/TGF-β1/pathway. Mol Immunol (2017) 91:156–64. doi: 10.1016/j.molimm.2017.09.006
45. Tian Y, Wang C, Ye Z, Xiao X, Kijlstra A, Yang P. Effect of 1,25-dihydroxyvitamin D3 on Th17 and Th1 response in patients with behçet’s disease. Invest Ophthalmol Vis Sci (2012) 53(10):6434–41. doi: 10.1167/iovs.12-10398
46. Chang JH, Cha HR, Lee DS, Seo KY, Kweon MN. 1,25-dihydroxyvitamin D3 inhibits the differentiation and migration of TH17 cells to protect against experimental autoimmune encephalomyelitis. PloS One (2010) 5(9):e12925. doi: 10.1371/journal.pone.0012925
47. Iho S, Takahashi T, Kura F, Sugiyama H, Hoshino T. The effect of 1,25-dihydroxyvitamin D3 on in vitro immunoglobulin production in human b cells. J Immunol (1986) 136(12):4427–31.
48. Chen S, Sims GP, Chen XX, Gu YY, Chen S, Lipsky PE. Modulatory effects of 1,25-dihydroxyvitamin D3 on human b cell differentiation. J Immunol (2007) 179(3):1634–47. doi: 10.4049/jimmunol.179.3.1634
49. Chen WC, Vayuvegula B, Gupta S. 1,25-dihydroxyvitamin D3-mediated inhibition of human b cell differentiation. Clin Exp Immunol (1987) 69(3):639–46.
50. Lemire JM, Adams JS, Sakai R, Jordan SC. 1 alpha,25-dihydroxyvitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest. (1984) 74(2):657–61. doi: 10.1172/JCI111465
51. Heine G, Niesner U, Chang HD, Steinmeyer A, Zügel U, Zuberbier T, et al. 1,25-dihydroxyvitamin D3 promotes IL-10 production in human b cells. Eur J Immunol (2008) 38(8):2210–8. doi: 10.1002/eji.200838216
52. Andrews BS, Eisenberg RA, Theofilopoulos AN, Izui S, Wilson CB, McConahey PJ, et al. Spontaneous murine lupus-like syndromes Clinical and immunopathological manifestations in several strains. J Exp Med (1978) 148(5):1198–215. doi: 10.1084/jem.148.5.1198
53. Schäfer AL, Eichhorst A, Hentze C, Kraemer AN, Amend A, Sprenger DTL, et al. Low dietary fiber intake links development of obesity and lupus pathogenesis. Front Immunol (2021) 12:696810. doi: 10.3389/fimmu.2021.696810
54. Häusler D, Torke S, Peelen E, Bertsch T, Djukic M, Nau R, et al. High dose vitamin d exacerbates central nervous system autoimmunity by raising T-cell excitatory calcium. Brain (2019) 142(9):2737–55. doi: 10.1093/brain/awz190
55. Mallya SM, Corrado KR, Saria EA, Yuan F ning F, Tran HQ, Saucier K, et al. Modeling vitamin d insufficiency and moderate deficiency in adult mice via dietary cholecalciferol restriction. Endocr Res (2016) 41(4):290–9. doi: 10.3109/07435800.2016.1141937
56. Williamson L, Hayes A, Hanson ED, Pivonka P, Sims NA, Gooi JH. High dose dietary vitamin D3 increases bone mass and strength in mice. Bone Rep (2017) 6:44–50. doi: 10.1016/j.bonr.2017.02.001
57. Martens PJ, Centelles-Lodeiro J, Ellis D, Cook DP, Sassi G, Verlinden L, et al. High serum vitamin d concentrations, induced via diet, trigger immune and intestinal microbiota alterations leading to type 1 diabetes protection in NOD mice. Front Immunol (2022) 9: doi: 10.3389/fimmu.2022.902678
58. Drozdenko G, Heine G, Worm M. Oral vitamin d increases the frequencies of CD38+ human b cells and ameliorates IL-17-producing T cells. Exp Dermatol (2014) 23(2):107–12. doi: 10.1111/exd.12300
59. Kimball SM, Ursell MR, O’Connor P, Vieth R. Safety of vitamin D3 in adults with multiple sclerosis. Am J Clin Nutr (2007) 86(3):645–51. doi: 10.1093/ajcn/86.3.645
60. Norman AW. Vitamin d nutrition is at a crossroads. Public Health Nutr (2011) 14(4):744–5. doi: 10.1017/S1368980011000280
61. Ismailova K, Poudel P, Parlesak A, Frederiksen P, Heitmann BL. Vitamin d in early life and later risk of multiple sclerosis–a systematic review, meta-analysis. PloS One (2019) 14(8):e0221645. doi: 10.1371/journal.pone.0221645
62. Fronczak CM, Barón AE, Chase HP, Ross C, Brady HL, Hoffman M, et al. In utero dietary exposures and risk of islet autoimmunity in children. Diabetes Care (2003) 26(12):3237–42. doi: 10.2337/diacare.26.12.3237
63. Song H, Yang L, Jia C. Maternal vitamin d status during pregnancy and risk of childhood asthma: A meta-analysis of prospective studies. Mol Nutr Food Res (2017) 61(5). doi: 10.1002/mnfr.201600657
64. Wolsk HM, Chawes BL, Litonjua AA, Hollis BW, Waage J, Stokholm J, et al. Prenatal vitamin d supplementation reduces risk of asthma/recurrent wheeze in early childhood: A combined analysis of two randomized controlled trials. PloS One (2017) 12(10):e0186657. doi: 10.1371/journal.pone.0186657
65. Grant CC, Crane J, Mitchell EA, Sinclair J, Stewart A, Milne T, et al. Vitamin d supplementation during pregnancy and infancy reduces aeroallergen sensitization: a randomized controlled trial. Allergy (2016) 71(9):1325–34. doi: 10.1111/all.12909
66. Petriashvili M. Impact of maternal vitamin d status on the formation of atopic dermatitis in young children. Global Pediatr Health (2021) 8. doi: 10.1177/2333794X211022916
67. Zerofsky MS, Jacoby BN, Pedersen TL, Stephensen CB. Daily cholecalciferol supplementation during pregnancy alters markers of regulatory immunity, inflammation, and clinical outcomes in a randomized controlled trial. J Nutr (2016) 146(11):2388–97. doi: 10.3945/jn.116.231480
68. Wagner CL, Hollis BW. The implications of vitamin d status during pregnancy on mother and her developing child. Front Endocrinol (Lausanne). (2018) 9:500. doi: 10.3389/fendo.2018.00500
69. Dutta S, Sengupta P. Men and mice: Relating their ages. Life Sci (2016) 152:244–8. doi: 10.1016/j.lfs.2015.10.025
70. Celhar T, Lu HK, Benso L, Rakhilina L, Lee HY, Tripathi S, et al. TLR7 protein expression in mild and severe lupus-prone models is regulated in a leukocyte, genetic, and IRAK4 dependent manner. Front Immunol (2019) 10:1546. doi: 10.3389/fimmu.2019.01546
71. Ewaschuk JB, Backer JL, Churchill TA, Obermeier F, Krause DO, Madsen KL. Surface expression of toll-like receptor 9 is upregulated on intestinal epithelial cells in response to pathogenic bacterial DNA. Infect Immun (2007) 75(5):2572–9. doi: 10.1128/IAI.01662-06
72. Arts RJW, Joosten LAB, Netea MG. The potential role of trained immunity in autoimmune and autoinflammatory disorders. Front Immunol (2018) 9:298. doi: 10.3389/fimmu.2018.00298
73. Netea MG, Domínguez-Andrés J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol (2020) 20(6):375–88. doi: 10.1038/s41577-020-0285-6
74. Kleinnijenhuis J, Quintin J, Preijers F, Benn CS, Joosten LAB, Jacobs C, et al. Long-lasting effects of BCG vaccination on both heterologous Th1/Th17 responses and innate trained immunity. JIN (2014) 6(2):152–8. doi: 10.1159/000355628
75. Quintin J, Saeed S, Martens JHA, Giamarellos-Bourboulis EJ, Ifrim DC, Logie C, et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe (2012) 12(2):223–32. doi: 10.1016/j.chom.2012.06.006
76. Christ A, Günther P, Lauterbach MAR, Duewell P, Biswas D, Pelka K, et al. Western Diet triggers NLRP3-dependent innate immune reprogramming. Cell (2018) 172(1):162-175.e14. doi: 10.1016/j.cell.2017.12.013
77. Helft J, Böttcher J, Chakravarty P, Zelenay S, Huotari J, Schraml BU, et al. GM-CSF mouse bone marrow cultures comprise a heterogeneous population of CD11c(+)MHCII(+) macrophages and dendritic cells. Immunity (2015) 42(6):1197–211. doi: 10.1016/j.immuni.2015.05.018
78. Feuerstein R, Gres V, Perdigó NE, Baasch S, Freudenhammer M, Elling R, et al. Macrophages are a potent source of streptococcus-induced IFN-β. J Immunol (2019) 203(12):3416–26. doi: 10.4049/jimmunol.1900542
79. Sohrabi Y, Dos Santos JC, Dorenkamp M, Findeisen H, Godfrey R, Netea MG, et al. Trained immunity as a novel approach against COVID-19 with a focus on bacillus calmette–guérin vaccine: mechanisms, challenges and perspectives. Clin Trans Immunol (2020) 9(12):e1228. doi: 10.1002/cti2.1228
80. Hong M, Sandalova E, Low D, Gehring AJ, Fieni S, Amadei B, et al. Trained immunity in newborn infants of HBV-infected mothers. Nat Commun (2015) 6:6588. doi: 10.1038/ncomms7588
81. Geissmann F, Jung S, Littman DR. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity (2003) 19(1):71–82. doi: 10.1016/S1074-7613(03)00174-2
82. Kratofil RM, Kubes P, Deniset JF. Monocyte conversion during inflammation and injury. Arteriosclerosis Thrombosis Vasc Biol (2017) 37(1):35–42. doi: 10.1161/ATVBAHA.116.308198
83. Olingy CE, San Emeterio CL, Ogle ME, Krieger JR, Bruce AC, Pfau DD, et al. Non-classical monocytes are biased progenitors of wound healing macrophages during soft tissue injury. Sci Rep (2017) 7(1):447. doi: 10.1038/s41598-017-00477-1
84. Théroude C, Reverte M, Heinonen T, Ciarlo E, Schrijver IT, Antonakos N, et al. Trained immunity confers prolonged protection from listeriosis. Front Immunol (2021) 12:723393. doi: 10.3389/fimmu.2021.723393
85. Sakthiswary R, Raymond AA. The clinical significance of vitamin d in systemic lupus erythematosus: A systematic review. PloS One (2013) 8(1):e55275. doi: 10.1371/journal.pone.0055275
86. Mok CC, Birmingham DJ, Ho LY, Hebert LA, Song H, Rovin BH. Vitamin d deficiency as marker for disease activity and damage in systemic lupus erythematosus: a comparison with anti-dsDNA and anti-C1q. Lupus (2012) 21(1):36–42. doi: 10.1177/0961203311422094
87. Szodoray P, Tarr T, Bazso A, Poor G, Szegedi G, Kiss E. The immunopathological role of vitamin d in patients with SLE: data from a single centre registry in Hungary. Scand J Rheumatol (2011) 40(2):122–6. doi: 10.3109/03009742.2010.507220
88. Andreoli L, Dall’Ara F, Piantoni S, Zanola A, Piva N, Cutolo M, et al. A 24-month prospective study on the efficacy and safety of two different monthly regimens of vitamin d supplementation in pre-menopausal women with systemic lupus erythematosus. Lupus (2015) 24(4–5):499–506. doi: 10.1177/0961203314559089
89. Karimzadeh H, Shirzadi M, Karimifar M. The effect of vitamin d supplementation in disease activity of systemic lupus erythematosus patients with vitamin d deficiency: A randomized clinical trial. J Res Med Sci (2017) 22(1):4. doi: 10.4103/1735-1995.199089
90. Lima GL, Paupitz J, Aikawa NE, Takayama L, Bonfa E, Pereira RMR. Vitamin d supplementation in adolescents and young adults with juvenile systemic lupus erythematosus for improvement in disease activity and fatigue scores: A randomized, double-blind, placebo-controlled trial. Arthritis Care Res (2016) 68(1):91–8. doi: 10.1002/acr.22621
91. Petri M, Bello KJ, Fang H, Magder LS. Vitamin d in SLE: Modest association with disease activity and urine Protein/Creatinine ratio. Arthritis Rheumatol (2013) 65(7):1865–71. doi: 10.1002/art.37953
92. Granfors M, Augustin H, Ludvigsson J, Brekke HK. No association between use of multivitamin supplement containing vitamin d during pregnancy and risk of type 1 diabetes in the child. Pediatr Diabetes. (2016) 17(7):525–30. doi: 10.1111/pedi.12334
93. Sørensen IM, Joner G, Jenum PA, Eskild A, Torjesen PA, Stene LC. Maternal serum levels of 25-hydroxy-vitamin d during pregnancy and risk of type 1 diabetes in the offspring. Diabetes (2012) 61(1):175–8. doi: 10.2337/db11-0875
94. Miettinen ME, Reinert L, Kinnunen L, Harjutsalo V, Koskela P, Surcel HM, et al. Serum 25-hydroxyvitamin d level during early pregnancy and type 1 diabetes risk in the offspring. Diabetologia (2012) 55(5):1291–4. doi: 10.1007/s00125-012-2458-8
95. Litonjua AA. Childhood asthma may be a consequence of vitamin d deficiency. Curr Opin Allergy Clin Immunol (2009) 9(3):202–7. doi: 10.1097/ACI.0b013e32832b36cd
96. Brustad N, Eliasen AU, Stokholm J, Bønnelykke K, Bisgaard H, Chawes BL. High-dose vitamin d supplementation during pregnancy and asthma in offspring at the age of 6 years. JAMA (2019) 321(10):1003–5. doi: 10.1001/jama.2019.0052
97. Richardson B, Scheinbart L, Strahler J, Gross L, Hanash S, Johnson M. Evidence for impaired t cell dna methylation in systemic lupus erythematosus and rheumatoid arthritis. Arthritis Rheumatism (1990) 33(11):1665–73. doi: 10.1002/art.1780331109
98. Coit P, Jeffries M, Altorok N, Dozmorov MG, Koelsch KA, Wren JD, et al. Genome-wide DNA methylation study suggests epigenetic accessibility and transcriptional poising of interferon-regulated genes in naïve CD4+ T cells from lupus patients. J Autoimmunity. (2013) 43:78–84. doi: 10.1016/j.jaut.2013.04.003
99. Anderson CM, Gillespie SL, Thiele DK, Ralph JL, Ohm JE. Effects of maternal vitamin d supplementation on the maternal and infant epigenome. Breastfeeding Med (2018) 13(5):371–80. doi: 10.1089/bfm.2017.0231
100. Xue J, Schoenrock SA, Valdar W, Tarantino LM, Ideraabdullah FY. Maternal vitamin d depletion alters DNA methylation at imprinted loci in multiple generations. Clin Epigenetics. (2016) 8:107. doi: 10.1186/s13148-016-0276-4
101. Carlberg C, Haq A. The concept of the personal vitamin d response index. J Steroid Biochem Mol Biol (2018) 175:12–7. doi: 10.1016/j.jsbmb.2016.12.011
102. Saksa N, Neme A, Ryynänen J, Uusitupa M, de Mello VDF, Voutilainen S, et al. Dissecting high from low responders in a vitamin D3 intervention study. J Steroid Biochem Mol Biol (2015) 148:275–82. doi: 10.1016/j.jsbmb.2014.11.012
103. Fragoso YD, Adoni T, Damasceno A, de Albuquerque Damasceno CA, Ferreira MLB, Finkelzstejn A, et al. Unfavorable outcomes during treatment of multiple sclerosis with high doses of vitamin d. J Neurological Sci (2014) 346(1):341–2. doi: 10.1016/j.jns.2014.08.019
104. Christakos S, Ajibade DV, Dhawan P, Fechner AJ, Mady LJ. Vitamin d: Metabolism. Endocrinol Metab Clinics North America. (2010) 39(2):243–53. doi: 10.1016/j.ecl.2010.02.002
105. Olmos-Ortiz A, Avila E, Durand-Carbajal M, Díaz L. Regulation of calcitriol biosynthesis and activity: Focus on gestational vitamin d deficiency and adverse pregnancy outcomes. Nutrients (2015) 7(1):443–80. doi: 10.3390/nu7010443
106. Lemire JM, Ince A, Takashima M. 1,25-dihydroxyvitamin D3 attenuates the expression of experimental murine lupus of MRL/l mice. Autoimmunity (1992) 12(2):143–8. doi: 10.3109/08916939209150321
107. Liu D, Fang YX, Wu X, Tan W, Zhou W, Zhang Y, et al. 1,25-(OH)2D3/Vitamin d receptor alleviates systemic lupus erythematosus by downregulating Skp2 and upregulating p27. Cell Comm Signaling (2019) 17(1):163. doi: 10.1186/s12964-019-0488-2
108. Terrier B, Derian N, Schoindre Y, Chaara W, Geri G, Zahr N, et al. Restoration of regulatory and effector T cell balance and b cell homeostasis in systemic lupus erythematosus patients through vitamin d supplementation. Arthritis Res Ther (2012) 14(5):R221. doi: 10.1186/ar4060
109. Heine G, Tabeling C, Hartmann B, Calera CRG, Kühl AA, Lindner J, et al. 25-hydroxvitamin D3 promotes the long-term effect of specific immunotherapy in a murine allergy model. J Immunol (2014) 193(3):1017–23. doi: 10.4049/jimmunol.1301656
110. Gembillo G, Siligato R, Amatruda M, Conti G, Santoro D. Vitamin d and glomerulonephritis. Medicina (Kaunas). (2021) 57(2):186. doi: 10.3390/medicina57020186
111. Garsen M, Sonneveld R, Rops ALWMM, Huntink S, van Kuppevelt TH, Rabelink TJ, et al. Vitamin d attenuates proteinuria by inhibition of heparanase expression in the podocyte. J Pathol (2015) 237(4):472–81. doi: 10.1002/path.4593
112. Masola V, Zaza G, Onisto M, Gambaro G. Heparanase: another renal player controlled by vitamin d. J Pathol (2016) 238(1):7–9. doi: 10.1002/path.4639
113. Bi J, Watanabe H, Fujimura R, Nishida K, Nakamura R, Oshiro S, et al. A downstream molecule of 1,25-dihydroxyvitamin D3, alpha-1-acid glycoprotein, protects against mouse model of renal fibrosis. Sci Rep (2018) 8(1):17329. doi: 10.1038/s41598-018-35339-x
114. Agarwal R, Acharya M, Tian J, Hippensteel RL, Melnick JZ, Qiu P, et al. Antiproteinuric effect of oral paricalcitol in chronic kidney disease. Kidney Int (2005) 68(6):2823–8. doi: 10.1111/j.1523-1755.2005.00755.x
115. Tan X, Wen X, Liu Y. Paricalcitol inhibits renal inflammation by promoting vitamin d receptor–mediated sequestration of NF-κB signaling. J Am Soc Nephrol (2008) 19(9):1741–52. doi: 10.1681/ASN.2007060666
116. Yu Q, Qiao Y, Liu D, Liu F, Gao C, Duan J, et al. Vitamin d protects podocytes from autoantibodies induced injury in lupus nephritis by reducing aberrant autophagy. Arthritis Res Ther (2019) 21(1):19. doi: 10.1136/annrheumdis-2019-eular.1556
117. Abe J, Nakamura K, Takita Y, Nakano T, Irie H, Nishii Y. Prevention of immunological disorders in MRL/l mice by a new synthetic analogue of vitamin D3: 22-Oxa-1α, 25-dihydroxyvitamin D3. J Nutr Sci Vitaminol (1990) 36(1):21–31. doi: 10.3177/jnsv.36.21
118. Vaisberg MW, Kaneno R, Franco MF, Mendes NF. Influence of cholecalciferol (vitamin D3) on the course of experimental systemic lupus erythematosus in F1 (NZBxW) mice. J Clin Lab Analysis. (2000) 14(3):91–6. doi: 10.1002/(SICI)1098-2825(2000)14:3<91::AID-JCLA2>3.0.CO;2-O
119. Souto Filho JTD, de Andrade AS, Ribeiro FM, Alves P de AS, Simonini VRF. Impact of vitamin d deficiency on increased blood eosinophil counts. Hematology/Oncol Stem Cell Ther (2018) 11(1):25–9. doi: 10.1016/j.hemonc.2017.06.003
120. Carriero V, Bullone M, Bertolini F, Ricciardolo FLM. Vitamin d negatively correlates with blood eosinophilia in stable adult asthmatics. Eur Respir J (2019) 54(suppl 63). doi: 10.1183/13993003.congress-2019.PA4305
121. Wu HY, Chen JX, Tian HQ, Zhang XL, Bian HY, Cheng L. Serum 25-hydroxyvitamin d inversely associated with blood eosinophils in patients with persistent allergic rhinitis. Asia Pacific Allergy (2017) 7(4):213–20. doi: 10.5415/apallergy.2017.7.4.213
122. Diny NL, Rose NR, Čiháková D. Eosinophils in autoimmune diseases. Front Immunol (2017) 8:484. doi: 10.3389/fimmu.2017.00484
123. Markusse HM, Schravenhoff R, Beerman H. Hypereosinophilic syndrome presenting with diarrhoea and anaemia in a patient with systemic lupus erythematosus. Netherlands J Med (1998) 52(2):79–81. doi: 10.1016/S0300-2977(97)00086-7
124. Thomeer M, Moerman P, Westhovens R, den EAV, Dequeker J, Demedts M. Systemic lupus erythematosus, eosinophilia and loffler’s endocarditis. unusual Assoc Eur Respir J (1999) 13(4):930–3. doi: 10.1034/j.1399-3003.1999.13d38.x
125. Habibagahi Z, Ali Nazarinia M, Aaflaki E, Ali Ostovan M, Hadi Bagheri M. Systemic lupus erythematosus and hyper-eosinophilic syndrome: an unusual association. West Indian Med J (2009) 58(1):69–71.
126. Aydoğdu S, Uçar O, Cetin M. A case of systemic lupus erythematosus presenting with hypereosinophilia and loeffler endocarditis. Acta Cardiol (2010) 65(5):571–3. doi: 10.2143/AC.65.5.2056245
127. Brito TNS, Vilar MJ, Almeida JB, Faria ALSB, Medeiros SDV, Medeiros MCC, et al. Measuring eosinophiluria, urinary eosinophil cationic protein and urinary interleukin-5 in patients with lupus nephritis. Allergy Asthma Clin Immunol (2014) 10(1):61. doi: 10.1186/s13223-014-0061-x
128. Tariq A, Okamato K, Tariq A, Rosenberg AZ, Soliman KM, Ploth DW, et al. Eosinophilia and risk of incident end stage kidney disease. BMC Nephrolog (2020) 21(1):14. doi: 10.1186/s12882-020-1685-3
129. Blosse A, Peru S, Levy M, Marteyn B, Floch P, Sifré E, et al. APRIL-producing eosinophils are involved in gastric MALT lymphomagenesis induced by helicobacter sp infection. Sci Rep (2020) 10:14858. doi: 10.1038/s41598-020-71792-3
130. Chu VT, Fröhlich A, Steinhauser G, Scheel T, Roch T, Fillatreau S, et al. Eosinophils are required for the maintenance of plasma cells in the bone marrow. Nat Immunol (2011) 12(2):151–9. doi: 10.1038/ni.1981
131. James J, Weaver V, Cantorna MT. Control of circulating IgE by the vitamin d receptor In vivo involves b cell intrinsic and extrinsic mechanisms. J Immunol (2017) 198(3):1164–71. doi: 10.4049/jimmunol.1601213
132. Horwitz DA. Regulatory T cells in systemic lupus erythematosus: past, present and future. Arthritis Res Ther (2008) 10(6):227. doi: 10.1186/ar2511
133. Hayashi T, Hasegawa K, Adachi C. Elimination of CD4+CD25+ T cell accelerates the development of glomerulonephritis during the preactive phase in autoimmune-prone female NZB × NZW F1 mice. Int J Exp Pathol (2005) 86(5):289–96. doi: 10.1111/j.0959-9673.2005.00438.x
134. Scalapino KJ, Tang Q, Bluestone JA, Bonyhadi ML, Daikh DI. Suppression of disease in new Zealand Black/New Zealand white lupus-prone mice by adoptive transfer of ex vivo expanded regulatory T cells. J Immunol (2006) 177(3):1451–9. doi: 10.4049/jimmunol.177.3.1451
135. Eggenhuizen PJ, Ng BH, Ooi JD. Treg enhancing therapies to treat autoimmune diseases. Int J Mol Sci (2020) 21(19):7015. doi: 10.3390/ijms21197015
136. Asano M, Toda M, Sakaguchi N, Sakaguchi S. Autoimmune disease as a consequence of developmental abnormality of a T cell subpopulation. J Exp Med (1996) 184(2):387–96. doi: 10.1084/jem.184.2.387
137. Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown single Mech self-tolerance causes various autoimmune diseases. J Immunol (1995) 155(3):1151–64.
138. Ng THS, Britton GJ, Hill EV, Verhagen J, Burton BR, Wraith DC. Regulation of adaptive immunity; the role of interleukin-10. Front Immunol (2013) 4:129. doi: 10.3389/fimmu.2013.00129
139. Groux H, Cottrez F. The complex role of interleukin-10 in autoimmunity. J Autoimmunity. (2003) 20(4):281–5. doi: 10.1016/S0896-8411(03)00044-1
140. Watanabe R, Ishiura N, Nakashima H, Kuwano Y, Okochi H, Tamaki K, et al. Regulatory b cells (B10 cells) have a suppressive role in murine lupus: CD19 and B10 cell deficiency exacerbates systemic autoimmunity. J Immunol (2010) 184(9):4801–9. doi: 10.4049/jimmunol.0902385
141. Baglaenko Y, Manion KP, Chang NH, Loh C, Lajoie G, Wither JE. Suppression of autoimmunity by CD5+ IL-10-producing b cells in lupus-prone mice. Genes Immun (2015) 16(5):311–20. doi: 10.1038/gene.2015.17
142. Amend A, Wickli N, Schäfer AL, Sprenger DTL, Manz RA, Voll RE, et al. Dual role of interleukin-10 in murine NZB/W F1 lupus. Int J Mol Sci (2021) 22(3):1347. doi: 10.3390/ijms22031347
143. Beebe AM, Cua DJ, de Waal Malefyt R. The role of interleukin-10 in autoimmune disease: systemic lupus erythematosus (SLE) and multiple sclerosis (MS). Cytokine Growth Factor Rev (2002) 13(4):403–12. doi: 10.1016/S1359-6101(02)00025-4
144. Saxena A, Khosraviani S, Noel S, Mohan D, Donner T, Hamad ARA. Interleukin-10 paradox: A potent immunoregulatory cytokine that has been difficult to harness for immunotherapy. Cytokine (2015) 74(1):27–34. doi: 10.1016/j.cyto.2014.10.031
145. Yi X, Yang P, Sun M, Yang Y, Li F. Decreased 1,25-dihydroxyvitamin D3 level is involved in the pathogenesis of vogt-Koyanagi-Harada (VKH) disease. Mol Vis (2011) 17:673–9.
146. Cippitelli M, Santoni A. Vitamin D3: a transcriptional modulator of the interferon-γ gene. Eur J Immunol (1998) 28(10):3017–30. doi: 10.1002/(SICI)1521-4141(199810)28:10<3017::AID-IMMU3017>3.0.CO;2-6
147. Joshi S, Pantalena LC, Liu XK, Gaffen SL, Liu H, Rohowsky-Kochan C, et al. 1,25-dihydroxyvitamin D3 ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A ▿. Mol Cell Biol (2011) 31(17):3653–69. doi: 10.1128/MCB.05020-11
148. Sommer A, Fabri M. Vitamin d regulates cytokine patterns secreted by dendritic cells to promote differentiation of IL-22-Producing T cells. PloS One (2015) 10(6):e0130395. doi: 10.1371/journal.pone.0130395
149. Yu S, Cantorna MT. The vitamin d receptor is required for iNKT cell development. Proc Natl Acad Sci U S A. (2008) 105(13):5207–12. doi: 10.1073/pnas.0711558105
150. Antico A, Tampoia M, Tozzoli R, Bizzaro N. Can supplementation with vitamin d reduce the risk or modify the course of autoimmune diseases? a systematic review of the literature. Autoimmun Rev (2012) 12(2):127–36. doi: 10.1016/j.autrev.2012.07.007
151. Lemke D, Klement RJ, Schweiger F, Schweiger B, Spitz J. Vitamin d resistance as a possible cause of autoimmune diseases: A hypothesis confirmed by a therapeutic high-dose vitamin d protocol. Front Immunol (2021) 12:655739. doi: 10.3389/fimmu.2021.655739
153. Arts RJW, Novakovic B, ter Horst R, Carvalho A, Bekkering S, Lachmandas E, et al. Glutaminolysis and fumarate accumulation integrate immunometabolic and epigenetic programs in trained immunity. Cell Metab (2016) 24(6):807–19. doi: 10.1016/j.cmet.2016.10.008
Keywords: vitamin D, autoimmunity, lupus, SLE, diet, choleacliferol, mice, autoimmune
Citation: Kraemer AN, Schäfer A-L, Sprenger DTL, Sehnert B, Williams JP, Luo A, Riechert L, Al-Kayyal Q, Dumortier H, Fauny J-D, Winter Z, Heim K, Hofmann M, Herrmann M, Heine G, Voll RE and Chevalier N (2022) Impact of dietary vitamin D on immunoregulation and disease pathology in lupus-prone NZB/W F1 mice. Front. Immunol. 13:933191. doi: 10.3389/fimmu.2022.933191
Received: 30 April 2022; Accepted: 17 October 2022;
Published: 24 November 2022.
Edited by:
Kutty Selva Nandakumar, Halmstad University, SwedenReviewed by:
Trine N. Jorgensen, Case Western Reserve University, United StatesCopyright © 2022 Kraemer, Schäfer, Sprenger, Sehnert, Williams, Luo, Riechert, Al-Kayyal, Dumortier, Fauny, Winter, Heim, Hofmann, Herrmann, Heine, Voll and Chevalier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nina Chevalier, bmluYS5jaGV2YWxpZXJAdW5pa2xpbmlrLWZyZWlidXJnLmRl; Reinhard E. Voll, cmVpbmhhcmQudm9sbEB1bmlrbGluaWstZnJlaWJ1cmcuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.