
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 29 July 2022
Sec. Alloimmunity and Transplantation
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.929995
 Natalia Redondo1,2*
Natalia Redondo1,2* Isabel Rodríguez-Goncer1,2Patricia Parra1,2Tamara Ruiz-Merlo1,2Francisco López-Medrano1,2,3Esther González4
Isabel Rodríguez-Goncer1,2Patricia Parra1,2Tamara Ruiz-Merlo1,2Francisco López-Medrano1,2,3Esther González4 Natalia Polanco4
Natalia Polanco4 Hernando Trujillo4Ana Hernández4Rafael San Juan1,2,3Amado Andrés3,4
Hernando Trujillo4Ana Hernández4Rafael San Juan1,2,3Amado Andrés3,4 José María Aguado1,2,3
José María Aguado1,2,3 Mario Fernández-Ruiz1,2,3
Mario Fernández-Ruiz1,2,3Risk stratification for cytomegalovirus (CMV) infection after kidney transplantation (KT) remains to be determined. Since endosomal toll-like receptors (TLRs) are involved in viral sensing, we investigated the impact of common single-nucleotide polymorphisms (SNPs) located within TLR3 and TLR9 genes on the occurrence of overall and high-level (≥1,000 IU/ml) CMV infection in a cohort of 197 KT recipients. Homozygous carriers of the minor allele of TLR3 (rs3775291) had higher infection-free survival compared with reference allele carriers (60.0% for TT versus 42.3% for CC/CT genotypes; P-value = 0.050). Decreased infection-free survival was observed with the minor allele of TLR9 (rs352139) (38.2% for TC/CC versus 59.3% for TT genotypes; P-value = 0.004). After multivariable adjustment, the recessive protective effect of the TLR3 (rs3775291) TT genotype was confirmed (adjusted hazard ratio [aHR]: 0.327; 95% CI: 0.167–0.642; P-value = 0.001), as was the dominant risk-conferring effect of TLR9 (rs352139) TC/CC genotypes (aHR: 1.865; 95% CI: 1.170–2.972; P-value = 0.009). Carriers of the TLR9 (rs352139) TC/CC genotypes showed lower CMV-specific interferon-γ-producing CD4+ T-cell counts measured by intracellular cytokine staining compared with the TT genotype (median of 0.2 versus 0.7 cells/μl; P-value = 0.003). In conclusion, TLR3/TLR9 genotyping may inform CMV infection risk after KT.
Compared to non-transplant subjects, solid organ transplant (SOT) recipients are at an increased risk of infectious complications due to the sustained effect of immunosuppressive therapies (1, 2). Current regimens are mainly aimed at abolishing adaptive cell-mediated responses, rendering the innate immune system a relevant player in the host defense against opportunistic pathogens (2). Cytomegalovirus (CMV) constitutes a major cause of morbidity and mortality in the SOT population (3). Following primary infection and similar to other members of the Herpesviridae family, CMV establishes life-long latency, which is usually controlled by the immune system. In certain circumstances—such as iatrogenic immunosuppression or biological stress—CMV may reactivate and cause clinically evident disease (4).
Despite advances in the understanding of the role of CMV-specific immunity over recent years, the choice of the optimal prevention strategy against CMV after SOT remains essentially informed by two factors only: donor and recipient (D/R) serostatus and previous use of T-cell-depleting agents (5, 6). The burden of post-transplant CMV infection and disease observed in large contemporary cohorts (7, 8), however, suggests that there is still room for improvement in risk stratification strategies (9). Increasing evidence supports the impact of single-nucleotide polymorphisms (SNPs) in genes that orchestrate innate and adaptive responses on the risk of CMV infection and disease (3, 10). Most of these studies have focused on pattern recognition receptors (PRRs). The major family of PRRs are toll-like receptors (TLRs), which may be classified according to their cellular location and ligand recognition specificity. Membrane-bound TLRs (such as TLR2, TLR4, or TLR5) primarily recognize pathogen-associated molecular patterns (PAMPs) of bacterial origin, whereas TLRs located within endosomal compartments (TLR3, TLR7, TLR8, and TLR9) predominantly bind viral nucleic acids (11, 12).
TLR3 recognizes extracellular double-stranded RNA (dsRNA), a by-product generated during replication and transcription of most viruses (13). It has been well-established the effect of genetic defects in the TLR3 pathway on the development of central nervous system (CNS) infection due to herpes simplex virus (HSV) and varicella-zoster virus (VZV) among otherwise immunocompetent individuals (14–16). Comparable data for the susceptibility to CMV in SOT recipients is still lacking. On the other hand, TLR9 senses and is activated by unmethylated cytosine-phosphate-guanine (CpG) dinucleotides. Various studies have reported that newborns carrying certain SNPs in the TLR9 gene (e.g., rs187084, rs352140) are at an increased risk of congenital CMV infection (17–19). By using a large multicenter cohort of seropositive kidney transplant (KT) recipients (OPERA Study), we first reported that the presence of the TT genotype in the TLR9 (rs5743836) SNPs provided protection against CMV viremia (20). However, mo further studies have investigated whether genetic variations in TLR9 are associated with the risk of developing post-transplant CMV infection or disease (10).
Given this paucity of data and the critical involvement of endosomal TLR-mediated signaling pathways in viral sensing, we aimed to investigate the influence of common SNPs in the TLR3 (rs3775291) and TLR9 (rs352139, rs5743836) genes on the incidence of CMV events in a cohort of KT recipients. The rs3775291 SNP is located within exon 4 of the TLR3 gene and implies a variation of 1234C>T, which changes a conserved leucine by a phenylalanine at position 412 (21). In relation to SNPs located in TLR9, rs352139 is located in intron 1 of exon 2 (1174T>C), whereas rs5743836 SNP is situated in the promoter region of the gene at position −1237A>G. In both cases, the polymorphism impacts TLR9 expression (22, 23). We also attempted to correlate the presence of genetic determinants of CMV infection with functional markers of CMV-specific T-cell-mediated immunity (CMV-CMI).
This observational study was based on a prospectively maintained database that included all consecutive adult patients undergoing KT at the University Hospital “12 de Octubre” between November 2014 and December 2016. Double organ recipients and those suffering from graft loss within the first post-transplant week were excluded. For this study, we also excluded patients at low risk for CMV infection (D−/R−). Participants were enrolled at the time of transplantation and followed up for at least 12 months, unless graft loss (i.e., retransplantation or return to dialysis) or death occurred earlier. Scheduled follow-up visits were carried out at baseline, every 2 weeks during the first 3 months, and monthly thereafter, as well as whenever clinically indicated. Pre-transplant, peri-operative, and post-transplant variables were prospectively recorded by means of a standardized case report form, and pseudo-anonymized data were entered into a secure REDCap database. Descriptions of immunosuppression and prophylaxis regimens are detailed in Supplementary Methods. The study was performed in accordance with the ethical standards outlined in the Declarations of Helsinki and Istanbul. All the patients provided informed consent and the local Clinical Research Ethics Committee approved the protocol (number 14/030). This study was prepared in accordance with the methodological recommendations drawn by the STREGA initiative.
The primary study outcomes were the occurrence of overall CMV infection and high-level CMV infection (defined by a viral load ≥1,000 IU/ml) during the first year after transplantation. This cut-off value of ≥1,000 IU/ml is the threshold usually applied in our institution to initiate preemptive antiviral therapy in KT recipients with asymptomatic CMV viremia. The development of CMV disease during that period was considered a secondary outcome. The diagnosis of CMV infection required polymerase chain reaction (PCR)-confirmed CMV replication regardless of the presence of attributable symptoms. The CMV viral load was quantified as previously described (24). Briefly, 200 μl of whole blood was used for DNA extraction using a NucliSENS® easyMag® instrument (bioMérieux Diagnostics, Marcy l’Etoile, France), according to the instructions of the manufacturer. A commercial real-time PCR assay (RealStar® CMV PCR kit 1.0, Altona Diagnostics GmbH, Hamburg, Germany) was used for viral DNA quantification. CMV viremia was assessed per protocol fortnightly during the first 2 months and then monthly thereafter during the first year. Additionally, real-time PCR was performed whenever CMV disease was suspected. CMV disease included viral syndrome or end-organ disease and required the documentation of CMV viremia along with attributable symptoms (25), as detailed in Supplementary Methods.
Whole blood specimens were collected in EDTA tubes at patient inclusion and stored at −80°C until analysis. DNA was extracted with the KingFisher™ Duo Prime system (Thermo Fisher Scientific, Waltham, MA) using the MagMax™ DNA Multi-Sample Ultra 2.0 kit, following the instructions of the manufacturer. TLR3 (rs3775291) and TLR9 (rs5743836, rs352139) genotyping was performed by TaqMan technology (Thermo Fisher Scientific) in a QuantStudio 3 system (Applied Biosystems, Foster City, CA). Sequences of TaqMan probes were as follows: ACTTGCTCATTCTCCCTTACACATA[T/C]TCAACCTAACCAAGAATAAAATCTC for TLR3 (rs3775291), TGGGATGTGCTGTTCCCTCTGCCTG[A/G]AAACTCCCCCAAGTCTCATATGACC for TLR9 (rs5743836) and TGTGTGAGTGGCCGGCCCCCAGCTC[C/T]ACCTCCACCCACTCCACTTCATGGG for TLR9 (rs352139). SNP and allele (genotype) calling was carried out by a standard end-point analysis with the aid of commercial genotype-calling software (TaqMan™ Genotyper Software v1.0.1) and the QuantStudio Design and Analysis Software v1.5.1 (both from Applied Biosystems).
The magnitude and functionality of the CMV-CMI were investigated by two different methods. We used a commercial enzyme-linked immunosorbent assay (ELISA)-based interferon (IFN)-γ release assay (QuantiFERON®-CMV [QTF-CMV], Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instructions and as described elsewhere (26). Additionally, we performed in a second subgroup the intracellular cytokine staining (ICS) method with flow cytometry to enumerate CMV pp65 and immediate-early (IE)-1-specific IFN-γ-producing CD8+ and CD4+ T cells, as previously described (27). Details on both techniques are available in Supplementary Methods.
Quantitative data are shown as the mean ± standard deviation (SD) or the median with interquartile range (IQR). The qualitative variables are expressed as absolute and relative frequencies. The normality of the distributions was tested with the Kolgomorov–Smirnov test. The deviation from the Hardy–Weinberg equilibrium (HWE) for each SNP was evaluated by the χ2 test with one degree of freedom. Comparisons of the cumulative incidence of study outcomes (overall and high-level CMV infection and CMV disease) according to different genotypes for the SNPs investigated were performed by the χ2 test or the Fisher’s exact test, as appropriate. Additional pairwise comparisons were conducted between different SNP genotype groups, either individually or in combination. Survival probabilities were estimated by the Kaplan–Meier method with study outcomes as events, and differences between groups were compared using the log-rank test. Multivariable Cox regression models were constructed with study outcomes as the dependent variables and using the entered method for entering the explicative variables. Results were expressed as hazard ratios (HRs) and 95% confidence intervals (CIs). Models were adjusted by those variables with a P-value <0.05 at the univariate level. Multicollinearity was analyzed with the variance inflation factor, with values of <3 being considered acceptable. We also performed a haplotype analysis by creating a score based on the SNP genotypes found to exert an independent impact on the primary outcomes. All the significance tests were two-tailed and considered significant at a P-value <0.05. Statistical analysis was performed using SPSS v21 (Statistical Package for Social Sciences, Chicago, IL) and graphs were generated with Prism v6.0 (GraphPad Software Inc., La Jolla, CA).
Overall, we included 197 KT recipients. As detailed in Table 1, 111 patients (56.3%) received valganciclovir prophylaxis for a median of 103 days (IQR: 91–148). The median follow-up period was 1,122 days (IQR: 956–1,297), and all but six patients (97.1%) completed the one-year follow-up with a functioning graft. The only relevant modification in the immunosuppression regimen was the conversion to a mammalian target of rapamycin (mTOR) inhibitor (target trough level of 4–6 ng/ml), which was performed on an individual basis beyond the third month in 19 patients (9.6%) due to the occurrence of tacroIimus-related adverse effects, difficult-to-treat CMV or BK infection, or post-transplant malignancy. Ten patients (5.1%) died at a median interval of 917.5 days (IQR: 600.3–1,390) from transplantation, and eight patients (4.1%) experienced graft loss. One- and two-year survival rates were 97.9% and 96.6%, whereas death-censored graft survival was 98.5% and 96.5%, respectively.
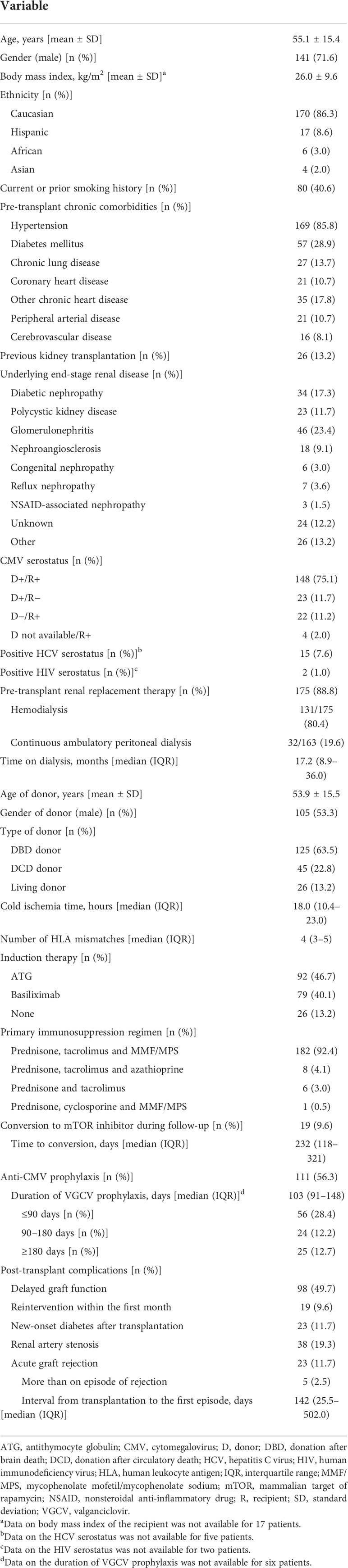
Table 1 Demographics and clinical characteristics of the study cohort (n = 197).
By month 12, 109 patients had developed at least one episode of CMV infection, resulting in a one-year cumulative incidence of 55.3% (95% CI: 48.1–62.4). The median interval from the first episode of infection was 73 days (IQR: 38.5–149.5). The one-year cumulative incidence of high-level CMV infection was 36.5% (29.8–43.7). Regarding the secondary outcome, 22 patients experienced CMV disease, which comprised viral syndrome (18 patients [81.8%]), colitis (3 patients [13.6%]) and hepatitis (one patient [4.5%]), yielding a one-year cumulative incidence of 11.2% (95% CI: 7.1–16.4).
All the samples were successfully genotyped. The genotype frequency distribution of each SNP is detailed in Table S1 of the Supporting Material. The TLR9 (rs5743836) SNP significantly deviated from the HWE. First, we investigated whether the presence of specific alleles within these SNPs was correlated with the cumulative incidence of CMV infection in the study population. As observed in Table 2, carriers of the minor TLR3 allele in the homozygous state (TT) had a lower 12-month incidence of CMV infection (40.0%) compared than homozygotes (CC) or heterozygotes (CT) for the reference allele (51.0% and 66.2%, respectively) (P-value = 0.036). A numerical difference was also observed for TLR9 (rs5743836), where the alternative G allele both in heterozygous (AG) or homozygous states (GG) was apparently associated with CMV infection (cumulative incidence rates of 64.4% and 66.7%, respectively) compared to the AA genotype (51.7%), although the difference did not reach statistical significance (P-value = 0.256). On the other hand, the minor C allele of TLR9 (rs352139) was also associated with an increased incidence of CMV infection, either in the heterozygous (TC) or homozygous state (CC) (56.8% and 66.7%, respectively) compared with homozygous patients (TT) for the reference allele (40.7%) (P-value = 0.017).
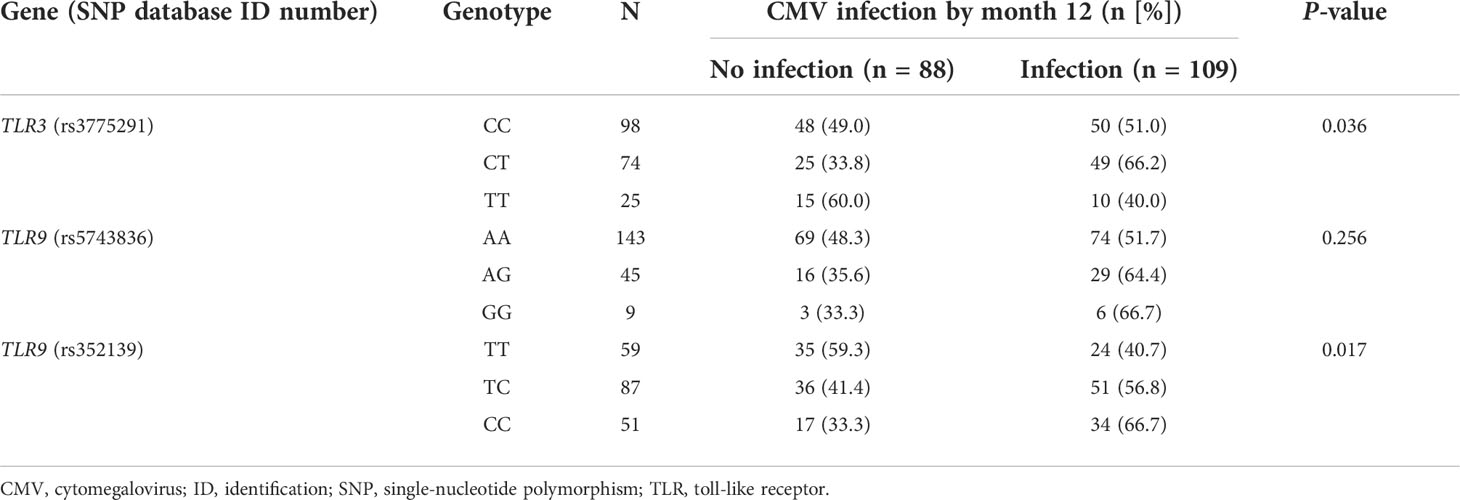
Table 2 One-year cumulative incidence of CMV infection according to the genotypic distribution of the candidate SNPs.
Next, we evaluated the impact of minor alleles in rs3775291, rs5743836, and rs352139 on the susceptibility to CMV in dominant (homozygous or heterozygous states) and recessive models (homozygous state only). In the case of the TLR3 (rs3775291) SNP, we found a trend toward a lower incidence of CMV infection among carriers of the T allele in the homozygous state (40.0% for TT versus 57.6% for CC/CT; P-value = 0.099), suggestive of a recessive protective effect. Regarding the TLR9 gene, the presence of the minor allele in homozygous or heterozygous states was associated with the occurrence of CMV infection, both for rs5743836 (64.8% for AG/GG versus 51.7% for AA; P-value = 0.1) and rs352139 (61.6% for TC/CC versus 40.7% for TT; P-value = 0.007), indicative of a dominant risk conferring effect (Table 3).
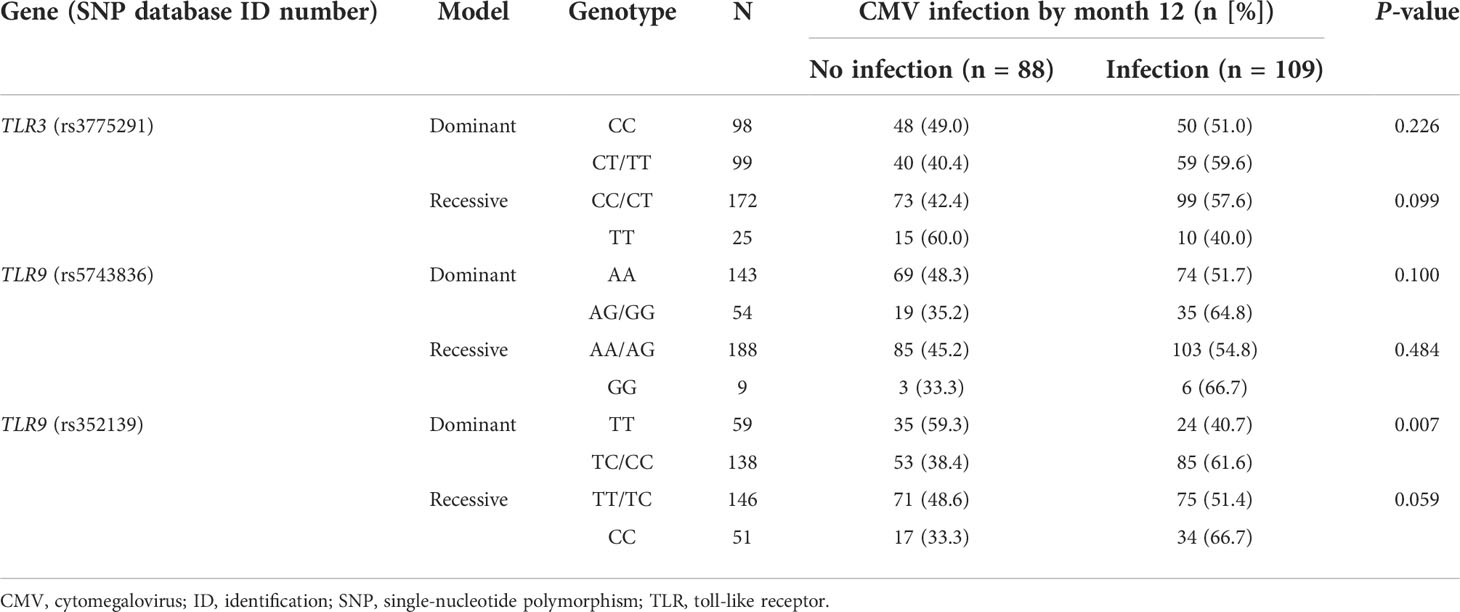
Table 3 One-year cumulative incidence of CMV infection according to recessive and dominant models for the minor alleles of candidate SNPs.
We also explored the effect of candidate SNPs on the development of high-level CMV infection during the first post-transplant year (Table S2). Consistent with the associations observed for CMV viremia at all levels, the incidence of this outcome was lower among carriers of the TLR3 (rs3775291) TT genotype, although the difference did not reach statistical significance (20.0% [5/25] versus 39.0% [67/172]; P-value = 0.066). The opposite dominant effect was confirmed for both TLR9 SNPs, with an increased incidence of high-level infection among homozygous or heterozygous carriers of the minor alleles of rs5743836 (48.1% [26/54] versus 32.2% [46/143]; P-value = 0.038) and rs352139 (41.3% [57/138] versus 25.4% [15/59]; P-value = 0.034).
In view of the associations observed between candidate SNPs in the TLR3 and TLR9 genes and the one-year cumulative incidence of overall and high-level CMV infection, we plotted event free-survival curves according to the selected genotype combinations (Figure 1). Homozygous carriers of the minor T allele of TLR3 (rs3775291) were more likely to remain free from CMV infection compared with carriers of the reference allele in homozygous or heterozygous states (CC/CT) (one-year survival rates: 60.0% versus 42.3%; log-rank test P-value = 0.050). In contrast, patients carrying the minor G allele (AG/GG) of TLR9 (rs5743836) had lower CMV infection-free survival than homozygous carriers of the reference allele (AA) (one-year survival rates: 35.2% versus 48.1%; log-rank test P-value = 0.063). Similar findings were found for the minor C allele of TLR9 (rs352139) (one-year survival rates: 38.2% versus 59.3% for TC/CC and TT genotypes, respectively; log-rank test P-value = 0.004).
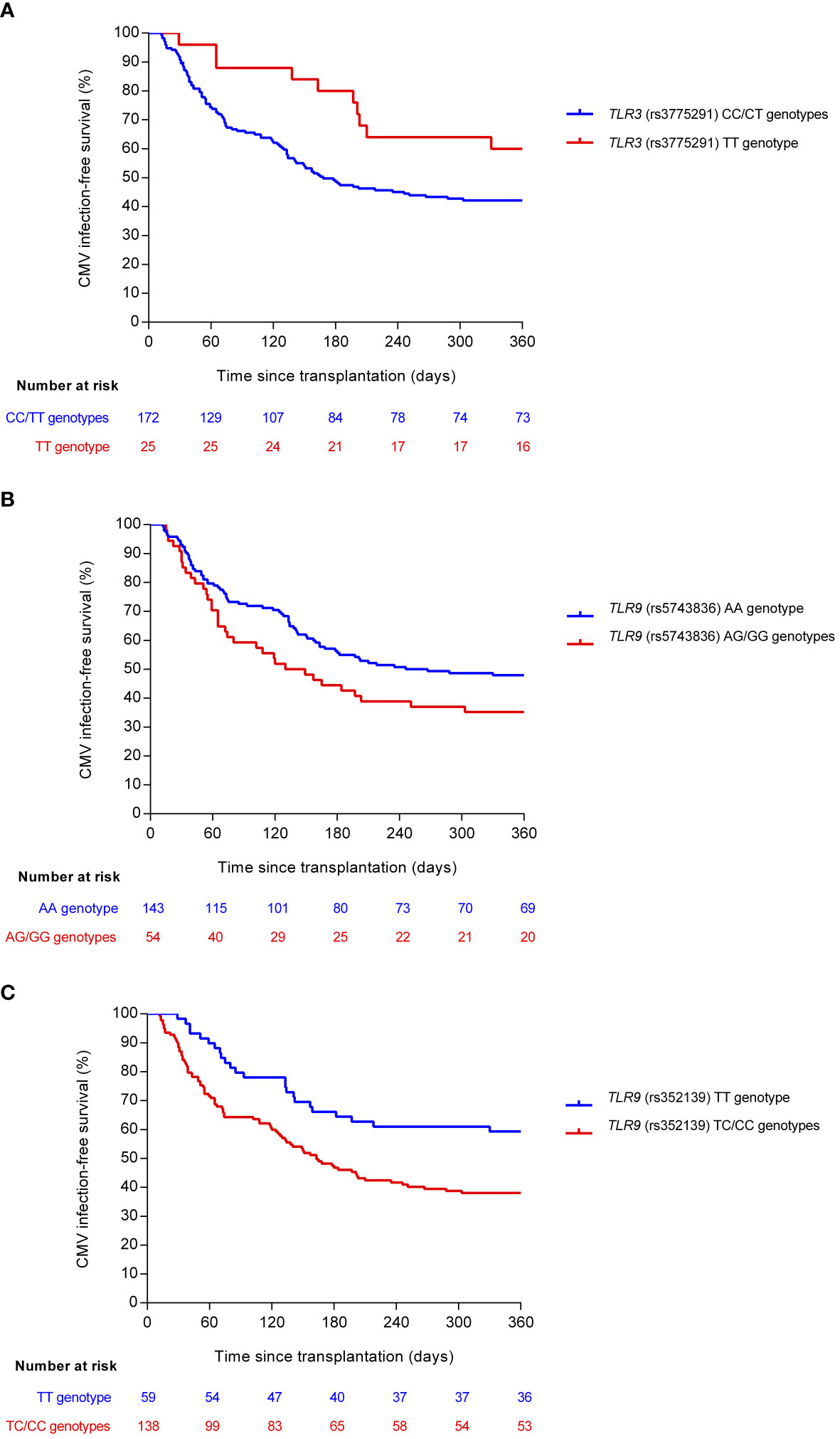
Figure 1 Comparison of CMV infection-free survival according to selected genotypes of candidate SNPs: (A) TLR3 (rs3775291) (log-rank P-value = 0.050), (B) TLR9 (rs5743836) (log-rank P-value = 0.063), and (C) TLR9 (rs352139) (log-rank P-value = 0.004). Reference and minor alleles are shown as blue and red curves, respectively. CMV, cytomegalovirus; SNP, single-nucleotide polymorphism.
Comparable results were obtained for high-level (≥1,000 IU/ml) CMV infection. Indeed, there was a non-significant trend toward an increased CMV infection-free survival among the homozygous carriers of the TLR3 (rs3775291) protective T allele (log-rank test P-value = 0.066). On the other hand, the presence of the minor risk alleles within the TLR9 SNPs—AG/GG for rs5743836 and TC/CC for rs352139—was associated with a decreased probability of remaining free from high-level infection during the first year (log-rank test P-values = 0.031 and 0.024, respectively) (Figure S1).
We subsequently tested by multivariable analysis the impact of these SNPs on study outcomes. To this end, Cox regression models were adjusted by clinical variables found to be associated at the univariable level with the occurrence of overall (Table S3) and high-level CMV infection (Table S4). After controlling for recipient and donor age, D+/R− CMV serostatus, use of antiviral prophylaxis, and graft function by month 1, the protective effect of the minor allele of the TLR3 (rs3775291) SNP in the homozygous state (TT) was confirmed (adjusted HR [aHR]: 0.327; 95% CI: 0.167–0.642; P-value = 0.001). No independent effect was observed for TLR9 (rs5743836) (aHR: 1.260; 95% CI: 0.836–1.899; P-value = 0.269), whereas the presence of the minor allele of TLR9 (rs352139) in the dominant model (TC/CC) had a significant risk effect (aHR: 1.865; 95% CI: 1.170–2.972; P-value = 0.009) (Table 4). Regarding the high-level CMV infection, the TLR9 (rs352139) SNP was found to act as an independent risk factor (aHR: 1.859; 95% CI: 1.039–3.328; P-value = 0.037) and the TLR3 (rs3775291) SNP to play a protective role (aHR: 0.409; 95% CI: 0.162–1.036; P-value = 0.059) (Table S5).

Table 4 Multivariable Cox regression models assessing the impact of selected SNPs on the incidence of CMV infection during the first post-transplant year.
We explored the additive impact on the risk of CMV infection of the number of risk genotypes in candidate SNPs. Genotypes were categorized as unfavorable as follows: the presence of the major C allele of TLR3 (rs3775291) in either a homozygous or heterozygous state (concordant with a recessive protective effect for the minor T allele); the presence of the minor G allele of TLR9 (rs5743836) in a homozygous or heterozygous state; and the presence of the minor C allele of TLR9 (rs352139) also in a homozygous or heterozygous state (concordant in both latter cases with a dominant risk model). Patients were accordingly divided: no unfavorable genotypes (seven patients [3.6%]), one genotype (59 [29.9%]), two genotypes (88 [44.7%]) and three genotypes (43 [21.8%]). We observed that the CMV infection-free survival progressively decreased with the increasing number of unfavorable genotypes (one-year survival rates: 57.1% [no unfavorable genotypes], 62.7% [one genotype], 37.1% [two genotypes] and 32.5% [three genotypes]; log-rank test P-value = 0.002) (Figure 2). Similar results were observed for high-level CMV infection (Figure S2). After multivariable adjustment, the number of unfavorable genotypes in candidate SNPs remained significantly associated with the risk of CMV infection (aHR [per additional genotype]: 1.528; 95% CI: 1.212–1.927; P-value = 0.017).
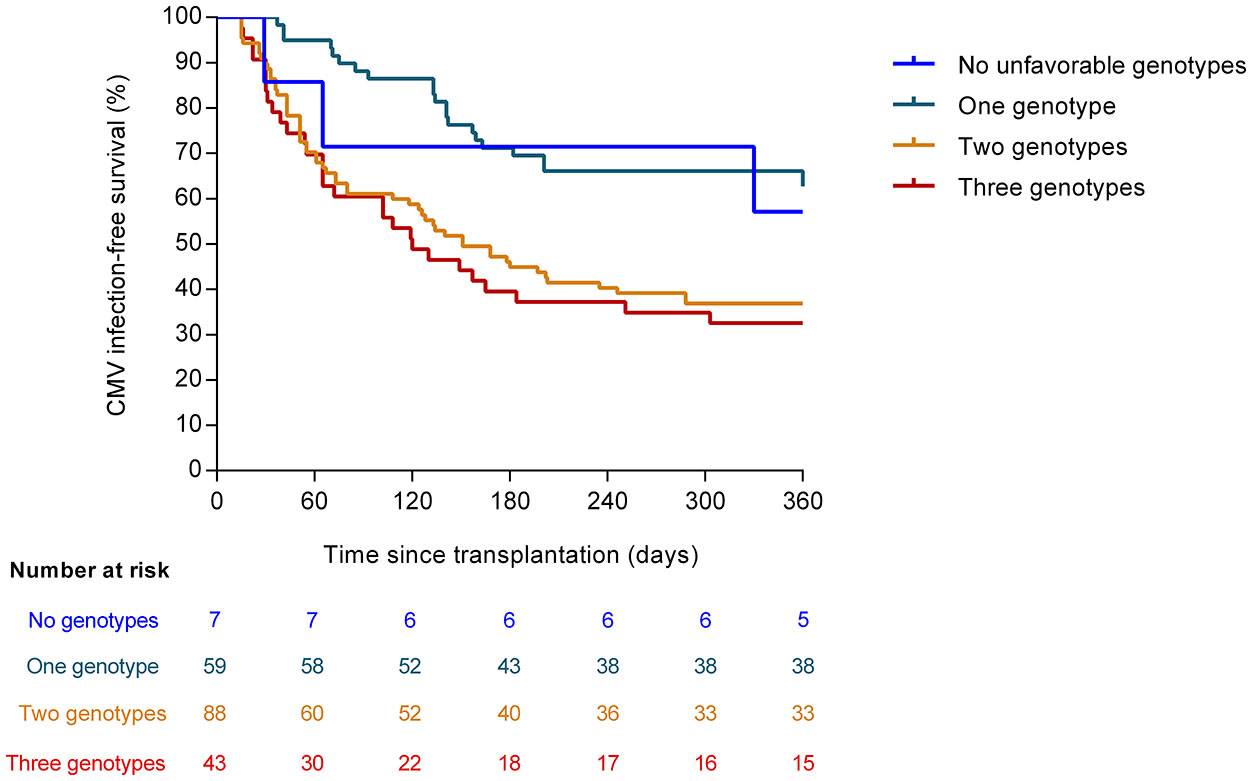
Figure 2 CMV infection-free survival according to the number of unfavorable genotypes in candidate SNPs (log-rank P-value = 0.002). The unfavorable genotypes were as follows: major C allele of TLR3 (rs3775291) in homozygous or heterozygous state; minor G allele of TLR9 (rs5743836) in homozygous or heterozygous state; and minor C allele of TLR9 (rs352139) in homozygous or heterozygous state. CMV: cytomegalovirus.
We also explored the association between different allelic variants in candidate SNPs and CMV disease (secondary outcome). Concordant with the results obtained for overall and high-level CMV infection, carriers of the G allele of TLR9 (rs5743836) in the homozygous or heterozygous state showed a trend toward a lower disease-free survival than homozygous carriers of the reference A allele (one-year survival rates: 83.2% versus 90.8%, respectively; log-rank test P-value = 0.149). The presence of the minor C allele of TLR9 (rs352139) was also associated with a lower disease-free survival (one-year survival rates: 86.8% versus 93.2% for TC/CC and TT genotypes, respectively; log-rank test P-value = 0.194) (Figure S3). None of these differences, however, achieved statistical significance. The small number of events of CMV disease (n = 22) prevented us from performing multivariable adjustment.
Finally, we investigated whether genetic polymorphisms in the TLR3 and TLR9 genes—which are essentially involved in activating the innate response—were correlated with the magnitude and functionality of CMV-CMI as the major effective components of the adaptive immunity against viruses. To this aim, we used two approaches: the ELISA-based QTF-CMV assay, which mainly detects CD8+ T-cell responses; and the reference ICS method (5, 28). In detail, the QTF-CMV assay was performed in 78 patients (39.6% of the overall cohort) at months 3, 4, and 5 (± one week), resulting in a total of 232 individual monitoring points. The enumeration of CMV pp65 and IE-1-specific IFN-γ-producing CD8+ and CD4+ T cells by ICS, on the other hand, was performed in a subgroup of 31 patients (15.7% of the cohort) at months 3 and 4, totaling 44 individual assessments. As depicted in Figure S4, there were no differences in the results of the QTF-CMV assay according to the presence of unfavorable genotypes in candidate SNPs, either by considering reactive responses (IFN-γ production [CMV minus nil] ≥0.2 IU/ml and ≥25% of nil) or by analyzing IFN-γ levels as a continuous variable. Additionally, no differences were observed in CMV-specific IFN-γ-producing CD8+ T-cell counts enumerated by ICS. Of note, carriers of the minor C allele of the TLR9 (rs352139) SNP (TC/CC genotypes) had lower CMV-specific IFN-γ-producing CD4+ T-cell counts compared with those bearing the TT genotype (median of 0.2 [IQR: 0.1–0.3] versus 0.7 [IQR: 0.7–1.0] cells/μl, respectively; P-value = 0.003) (Figure 3). Comparison was not feasible for TLR3 since only one patient with ICS data was homozygous for the minor T allele.
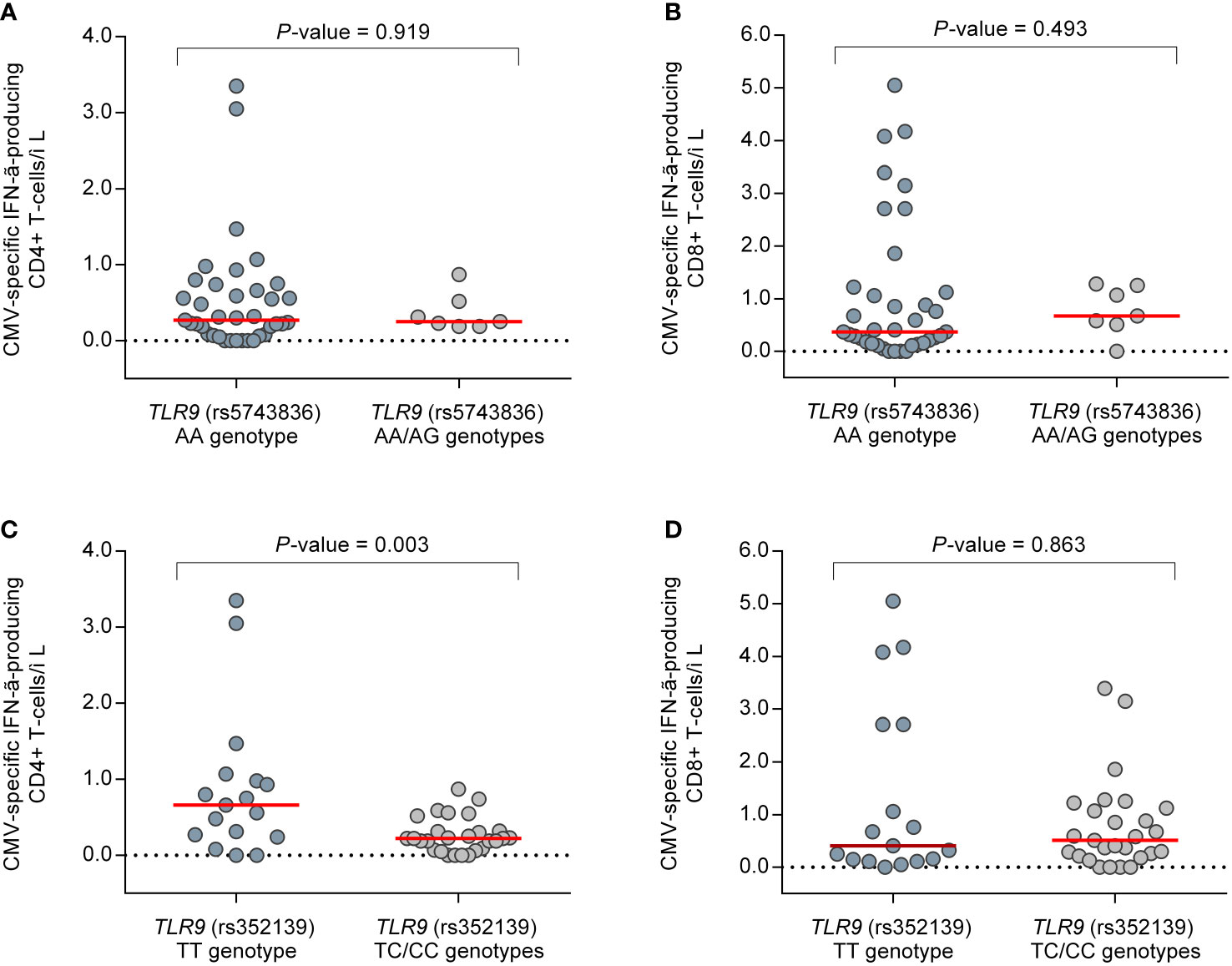
Figure 3 Comparison of CMV pp65 and IE-1-specific IFN-γ-producing CD4+ and CD8+ T-cell counts enumerated by intracellular cytokine stating in a subgroup of 31 patients (44 individual monitoring points) between selected genotypes of candidate SNPs: (A) CD4+ and (B) CD8+ T cells according to TLR9 (rs5743836); (C) CD4+ and (D) CD8+ T cells according to TLR9 (rs352139). Horizontal bars and whiskers represent median values and interquartile ranges, respectively. CMV, cytomegalovirus; IFN-γ, interferon-γ.
The contributing effect to the individual susceptibility to CMV of genetic variability in genes coding for immune mediators and receptors has been extended to the specific setting of SOT over the past years (10). This work contributes to this emerging field of research with two SNPs that have not been assessed in that population, such as TLR3 (rs3775291) and TLR9 (rs352139). Additionally, we have analyzed the TLR9 (rs5743836) SNP in an attempt to validate the association with CMV infection previously reported in a multicenter cohort of seropositive KT recipients (20). Both TLR3 and TLR9 encode for endosomal PRRs that act cooperatively to orchestrate immune responses against viral pathogens, thus providing biological plausibility to our findings.
First, we have found that the presence in a homozygous state of the alternative allele of TLR3 (rs3775291) (TT genotype) exerts a protective role on the risk of CMV viremia at any level and, with borderline significance, at high-level (≥1,000 IU/ml) infection. This polymorphism implies a C → T substitution which leads to the replacement of a conserved leucine with phenylalanine at position 412 (L412F). This missense mutation is mapped within the 14 leucine rich repeat domain of the TLR3 ectodomain and affects the binding capacity to dsRNA, resulting in a decreased signaling cascade compared to the wild-type form (21, 29). The TLR3 (rs3775291) SNP has been linked with a protective effect against other viral infections. Kindberg et al. investigated the role of this polymorphism on the risk of tick-borne encephalitis (TBE), and reported that the functional form of TLR3—reflecting a homozygous state for the reference allele (CC genotype)—was overrepresented among patients with TBE virus infection as compared to healthy individuals. The authors suggested that homozygous carriers of the minor T allele would mount a reduced inflammatory response, attenuating the severity of TBE infection (30). Other authors found that the presence of the minor allele of TLR3 (rs3775291) both in heterozygous or homozygous states conferred natural resistance against human immunodeficiency virus (HIV) infection in a cohort of 102 at-risk individuals. Interestingly, in vitro assays showed that HIV replication in peripheral mononuclear cells (PBMCs) carrying the L412F variant was significantly reduced compared with that of homozygous samples for the reference allele (31). The minor allelic variant at rs3775291 has been found to be more common among healthy HSV-2-seronegative individuals than in patients with genital HSV-2 infection, pointing to a protective phenotype, although no differences were observed in disease severity. Homozygous carriers of minor TLR3 alleles expressed higher TLR3 mRNA levels in response to HSV-2 stimulation than those homozygous for the reference alleles, suggesting an impaired expression (32).
The inverse association observed in our cohort between the rs3775291 T allele in a recessive model and the occurrence of CMV events not only applied to the overall infection but was also shown for episodes of high-level viremia, although this protective effect could not be proven for CMV disease. Beyond the insufficient statistical power due to the small number of cases of disease, this lack of correlation may be explained by the fact that the allelic variation in TLR3 acts at the innate level rather than shaping adaptive immunity, as supported by the observation that fibroblasts from TLR3-deficient patients show impaired IFN responses, although PBMCs from the same individuals adequately respond upon stimulation (32–34).
Taken together, the available evidence suggests that the protective role exerted by the TLR3 (rs3775291) TT genotype would be due to a dampened inflammatory response mediated by the impaired TLR3 form, since it has been shown that the pro-inflammatory environment may act as a trigger for CMV reactivation (35, 36). Of note, the L412F substitution is only partially defective in terms of TLR function (21), which would be concordant with a recessive rather than dominant inheritance model.
Secondly, we have analyzed the TLR9 (rs5743836) SNP investigated by our group in a previous multicenter cohort that included 315 CMV-seropositive KT recipients (20). In line with the results obtained from the OPERA Study, in actual experience we found that patients bearing the alternative G allele in a heterozygous or homozygous state (AG/GG genotypes) had a higher incidence of CMV infection compared to AA carriers, with this increased risk being even more evident for high-level viremia. This association, however, was not maintained after adjustment for clinical covariables (such as recipient and donor age, D+/R− serostatus, or use of valganciclovir prophylaxis). The TLR9 (rs5743836) SNP—which is mapped in the promoter region of the gene affecting its transcription—has been related to increased susceptibility to other viral infections, such as dengue (37). Carvalho et al. reported that PBMCs heterozygous for the minor allelic variant of rs5743836 show a loop of TLR9/interleukin-6 signaling amplification upon CpG stimuli, resulting in increased B-cell activation and production of pro-inflammatory cytokines (22). It may be hypothesized that this milieu would in turn promote CMV reactivation.
Another relevant finding of our study is the risk conferred by the minor allelic variant of TLR9 (rs352139) in the dominant model (TC/CC genotypes) for overall and high-level CMV infection and—without reaching statistical significance—CMV disease. This polymorphism (+1174 T>C) is located within intron 1 and has been associated with differences in TLR9 expression at the transcriptional level (38). The combined presence of the minor SNP alleles at positions +1,174 and −1,486 can downregulate TLR9 expression. Since rs352139 is an intronic polymorphism, it has been speculated that the C allele would generate an alternative splicing site that influences mRNA levels and therefore protein synthesis (39). Associations in the same direction between the TLR9 (rs352139) SNP and infection have been reported, including maternal CMV infection in a cohort of Zimbabwean pregnant women in which homozygous carriers of the minor C allele were at higher risk than homozygous for the reference allele (40). Interestingly, the CT genotype of rs352139 has been recently shown to be associated with an increased risk of Epstein–Barr virus-related infectious mononucleosis, which points to a role in the susceptibility to herpesviruses different from CMV (41). Further studies on tuberculosis (42, 43) and malaria (44) have reached comparable conclusions.
Similar to TLR3, TLR9 is an endosomal PRR that is activated upon recognition of CpG motifs (12). This triggers the signaling cascade via the adaptor molecule MyD88 that stimulates the pro-inflammatory nuclear transcription factor NF-κβ and IFN production (12, 45). MyD88 is also an important mediator, inducing T-cell activation (46, 47). Taking this into consideration, it is plausible that polymorphisms in the TLR9 gene lead to an impaired signaling cascade that would influence both innate and adaptive responses against CMV. This notion is supported by the novel observation that carriers of TC/CC genotypes of rs352139 exhibited lower counts of CMV-specific IFN-γ-producing CD4+ T cells by ICS than those bearing the TT genotype. The lack of differences in the QTF-CMV results between favorable and unfavorable genotypes may be explained by the fact that this assay essentially measures CD8+ T-cell responses (48). Indeed, CMV-specific CD8+ T-cell counts in the ICS assay did not differ either. These results would point to a specific impact of TLR9 SNPs in CD4+ T-cell activation, which is consistent with the role shown for CD4+ T-cell-mediated responses in the long-term control with latent infection (49, 50).
Our study has some noteworthy limitations. The relatively small sample size has prevented us from performing a sensitivity analysis limited to patients not receiving antiviral prophylaxis. This limitation particularly applies to the analysis of CMV disease. Nevertheless, the type of prevention strategy was controlled for in the Cox models for overall and high-level CMV infection. Additionally, data on CMV-CMI by the QTF-CMV assay and ICS were retrospectively derived from previous studies (26, 27), which meant that this information was lacking in most genotyped patients. A significant HWE deviation was observed in the genotypic frequency of TLR9 (rs5743836) SNP, which may have introduced some bias. Moreover, since the frequency of the minor allele G is very low in Caucasian ethnicities (0.14), only nine individuals in this cohort were carriers of the GG genotype. Both situations would have been overcome with a larger sample size. Finally, we did not genotype SNPs in the donors, although the clinical relevance is unclear due to the small amount of donor lymphoid tissue in the renal graft compared to other types of SOT such as the small bowel.
Nevertheless, our results are original since no evidence on the effect of TLR3 (rs3775291) and TLR9 (rs352139) SNPs on the risk of CMV events was available for the specific SOT population. Additionally, one of the main barriers to the implementation of genotyping into clinical practice is that most studies usually include a discovery cohort only and lack independent validation (10). In this line, we have reinforced our previous results concerning the significance of TLR9 (rs5743836) on the occurrence of CMV viremia among KT recipients (20). Future studies should confirm the associations observed between these SNPs and the susceptibility of an individual to CMV infection and disease, as well as their potential implications for risk stratification and minimization. Finally, functional analysis should attempt to elucidate the biological mechanism underlying these associations.
The data that support the findings of this study are available upon reasonable request to the corresponding author.
The studies involving human participants were reviewed and approved by the Clinical Research Ethics Committee Hospital 12 de Octubre (Study protocol number 14/030). The patients/participants provided their written informed consent to participate in this study.
NR and MF-R designed the study, performed statistical calculations, wrote the manuscript and supervised all aspects of the study. PP performed wet-lab analyses. TR-M collected the samples. IR-G, FL-M, EG, NP, AH, HT, RS, and AA involved in patients’ recruitment and data registration. IR-G, FL-M, RS, AA, and JA critically reviewed the manuscript and provided significant input and feedback on the draft manuscript. All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This work was supported by the Instituto de Salud Carlos III (ISCIII), Spanish Ministry of Science and Innovation (PIE13/00045, PI17/01120, PI20/01084)—co-financed by the European Development Regional Fund “A way to achieve Europe” and by the European Social Fund (ESF) “The ESF invests in your future”. IR-G. holds a research training contract “Río Hortega” (CM19/00163) and MF-R holds a research contract “Miguel Servet” (CP18/00073), both from the ISCIII, Spanish Ministry of Science and Innovation.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.929995/full#supplementary-material
1. Sanclemente G, Moreno A, Navasa M, Lozano F, Cervera C. Genetic variants of innate immune receptors and infections after liver transplantation. World J Gastroenterol (2014) 20(32):11116–30. doi: 10.3748/wjg.v20.i32.11116
2. Meneghini M, Bestard O, Grinyo JM. Immunosuppressive drugs modes of action. Best Pract Res Clin Gastroenterol (2021) 54-55:101757. doi: 10.1016/j.bpg.2021.101757
3. Sezgin E, An P, Winkler CA. Host genetics of cytomegalovirus pathogenesis. Front Genet (2019) 10:616. doi: 10.3389/fgene.2019.00616
4. Tyl MD, Betsinger CN, Cristea IM. Virus-host protein interactions as footprints of human cytomegalovirus replication. Curr Opin Virol (2021) 52:135–47. doi: 10.1016/j.coviro.2021.11.016
5. Torre-Cisneros J, Aguado JM, Caston JJ, Almenar L, Alonso A, Cantisan S, et al. Management of cytomegalovirus infection in solid organ transplant recipients: SET/GESITRA-SEIMC/REIPI recommendations. Transplant Rev (Orlando) (2016) 30(3):119–43. doi: 10.1016/j.trre.2016.04.001
6. Kotton CN, Kumar D, Caliendo AM, Huprikar S, Chou S, Danziger-Isakov L, et al. The third international consensus guidelines on the management of cytomegalovirus in solid-organ transplantation. Transplantation (2018) 102(6):900–31. doi: 10.1097/TP.0000000000002191
7. Manuel O, Kralidis G, Mueller NJ, Hirsch HH, Garzoni C, van Delden C, et al. Impact of antiviral preventive strategies on the incidence and outcomes of cytomegalovirus disease in solid organ transplant recipients. Am J Transpl (2013) 13(9):2402–10. doi: 10.1111/ajt.12388
8. Fernandez-Ruiz M, Arias M, Campistol JM, Navarro D, Gomez-Huertas E, Gomez-Marquez G, et al. Cytomegalovirus prevention strategies in seropositive kidney transplant recipients: an insight into current clinical practice. Transpl Int (2015) 28(9):1042–54. doi: 10.1111/tri.12586
9. Navarro D, Fernandez-Ruiz M, Aguado JM, Sandonis V, Perez-Romero P. Going beyond serology for stratifying the risk of CMV infection in transplant recipients. Rev Med Virol (2019) 29(1):e2017. doi: 10.1002/rmv.2017
10. Redondo N, Navarro D, Aguado JM, Fernandez-Ruiz M. Human genetic polymorphisms and risk of viral infection after solid organ transplantation. Transplant Rev (Orlando) (2021) 36(1):100669.doi: 10.1016/j.trre.2021.100669
11. Carty M, Guy C, Bowie AG. Detection of viral infections by innate immunity. Biochem Pharmacol (2021) 183:114316. doi: 10.1016/j.bcp.2020.114316
12. Fitzgerald KA, Kagan JC. Toll-like receptors and the control of immunity. Cell (2020) 180(6):1044–66. doi: 10.1016/j.cell.2020.02.041
13. Jacobs BL, Langland JO. When two strands are better than one: the mediators and modulators of the cellular responses to double-stranded RNA. Virology (1996) 219(2):339–49. doi: 10.1006/viro.1996.0259
14. Zhang SY. Herpes simplex virus encephalitis of childhood: inborn errors of central nervous system cell-intrinsic immunity. Hum Genet (2020) 139(6-7):911–8. doi: 10.1007/s00439-020-02127-5
15. Mielcarska MB, Bossowska-Nowicka M, Toka FN. Functional failure of TLR3 and its signaling components contribute to herpes simplex encephalitis. J Neuroimmunol (2018) 316:65–73. doi: 10.1016/j.jneuroim.2017.12.011
16. Sironi M, Peri AM, Cagliani R, Forni D, Riva S, Biasin M, et al. TLR3 mutations in adult patients with herpes simplex virus and varicella-zoster virus encephalitis. J Infect Dis (2017) 215(9):1430–4. doi: 10.1093/infdis/jix166
17. Paradowska E, Jablonska A, Studzinska M, Skowronska K, Suski P, Wisniewska-Ligier M, et al. TLR9 -1486T/C and 2848C/T SNPs are associated with human cytomegalovirus infection in infants. PloS One (2016) 11(4):e0154100. doi: 10.1371/journal.pone.0154100
18. Wujcicka W, Paradowska E, Studzinska M, Wilczynski J, Nowakowska D. Toll-like receptors genes polymorphisms and the occurrence of HCMV infection among pregnant women. Virol J (2017) 14(1):64. doi: 10.1186/s12985-017-0730-8
19. Wujcicka W, Paradowska E, Studzinska M, Gaj Z, Wilczynski J, Lesnikowski Z, et al. TLR9 2848 GA heterozygotic status possibly predisposes fetuses and newborns to congenital infection with human cytomegalovirus. PloS One (2015) 10(4):e0122831. doi: 10.1371/journal.pone.0122831
20. Fernandez-Ruiz M, Corrales I, Arias M, Campistol JM, Gimenez E, Crespo J, et al. Association between individual and combined SNPs in genes related to innate immunity and incidence of CMV infection in seropositive kidney transplant recipients. Am J Transpl (2015) 15(5):1323–35. doi: 10.1111/ajt.13107
21. Ranjith-Kumar CT, Miller W, Sun J, Xiong J, Santos J, Yarbrough I, et al. Effects of single nucleotide polymorphisms on toll-like receptor 3 activity and expression in cultured cells. J Biol Chem (2007) 282(24):17696–705. doi: 10.1074/jbc.M700209200
22. Carvalho A, Osorio NS, Saraiva M, Cunha C, Almeida AJ, Teixeira-Coelho M, et al. The c allele of rs5743836 polymorphism in the human TLR9 promoter links IL-6 and TLR9 up-regulation and confers increased b-cell proliferation. PloS One (2011) 6(11):e28256. doi: 10.1371/journal.pone.0028256
23. Omar AH, Yasunami M, Yamazaki A, Shibata H, Ofori MF, Akanmori BD, et al. Toll-like receptor 9 (TLR9) polymorphism associated with symptomatic malaria: a cohort study. Malar J (2012) 11:168. doi: 10.1186/1475-2875-11-168
24. Rodriguez-Goncer I, Ruiz-Ruigomez M, Lopez-Medrano F, Corbella L, Polanco N, Gonzalez Monte E, et al. CMV infection, valganciclovir exposure, and the risk of BK viremia and associated nephropathy after kidney transplantation: Is there a link? Transpl Infect Dis (2021) 23(4):e13597. doi: 10.1111/tid.13597
25. Ljungman P, Boeckh M, Hirsch HH, Josephson F, Lundgren J, Nichols G, et al. Definitions of cytomegalovirus infection and disease in transplant patients for use in clinical trials. Clin Infect Dis (2017) 64(1):87–91. doi: 10.1093/cid/ciw668
26. Fernandez-Ruiz M, Rodriguez-Goncer I, Parra P, Ruiz-Merlo T, Corbella L, Lopez-Medrano F, et al. Monitoring of CMV-specific cell-mediated immunity with a commercial ELISA-based interferon-gamma release assay in kidney transplant recipients treated with antithymocyte globulin. Am J Transpl (2020) 20(8):2070–80. doi: 10.1111/ajt.15793
27. Fernandez-Ruiz M, Gimenez E, Vinuesa V, Ruiz-Merlo T, Parra P, Amat P, et al. Regular monitoring of cytomegalovirus-specific cell-mediated immunity in intermediate-risk kidney transplant recipients: predictive value of the immediate post-transplant assessment. Clin Microbiol Infect (2019) 25(3):381.e1– e10. doi: 10.1016/j.cmi.2018.05.010
28. Fernandez-Ruiz M, Kumar D, Humar A. Clinical immune-monitoring strategies for predicting infection risk in solid organ transplantation. Clin Transl Immunol (2014) 3(2):e12. doi: 10.1038/cti.2014.3
29. Enkhbayar P, Kamiya M, Osaki M, Matsumoto T, Matsushima N. Structural principles of leucine-rich repeat (LRR) proteins. Proteins (2004) 54(3):394–403. doi: 10.1002/prot.10605
30. Kindberg E, Vene S, Mickiene A, Lundkvist A, Lindquist L, Svensson L. A functional toll-like receptor 3 gene (TLR3) may be a risk factor for tick-borne encephalitis virus (TBEV) infection. J Infect Dis (2011) 203(4):523–8. doi: 10.1093/infdis/jiq082
31. Sironi M, Biasin M, Cagliani R, Forni D, De Luca M, Saulle I, et al. A common polymorphism in TLR3 confers natural resistance to HIV-1 infection. J Immunol (2012) 188(2):818–23. doi: 10.4049/jimmunol.1102179
32. Svensson A, Tunback P, Nordstrom I, Padyukov L, Eriksson K. Polymorphisms in toll-like receptor 3 confer natural resistance to human herpes simplex virus type 2 infection. J Gen Virol (2012) 93(Pt 8):1717–24. doi: 10.1099/vir.0.042572-0
33. Zhang SY, Jouanguy E, Ugolini S, Smahi A, Elain G, Romero P, et al. TLR3 deficiency in patients with herpes simplex encephalitis. Science (2007) 317(5844):1522–7. doi: 10.1126/science.1139522
34. Guo Y, Audry M, Ciancanelli M, Alsina L, Azevedo J, Herman M, et al. Herpes simplex virus encephalitis in a patient with complete TLR3 deficiency: TLR3 is otherwise redundant in protective immunity. J Exp Med (2011) 208(10):2083–98. doi: 10.1084/jem.20101568
35. Cook CH, Trgovcich J, Zimmerman PD, Zhang Y, Sedmak DD. Lipopolysaccharide, tumor necrosis factor alpha, or interleukin-1beta triggers reactivation of latent cytomegalovirus in immunocompetent mice. J Virol (2006) 80(18):9151–8. doi: 10.1128/JVI.00216-06
36. Traub S, Demaria O, Chasson L, Serra F, Desnues B, Alexopoulou L. Sex bias in susceptibility to MCMV infection: implication of TLR9. PloS One (2012) 7(9):e45171. doi: 10.1371/journal.pone.0045171
37. Singh AK, Prakash S, Garg RK, Jain P, Kumar R, Jain A. Study of single nucleotide polymorphisms in endosomal toll-like receptors-3, 7, and 9 genes in patients with dengue: A case-control study. Cureus (2021) 13(5):e14883. doi: 10.7759/cureus.14883
38. Fang J, Hu R, Hou S, Ye Z, Xiang Q, Qi J, et al. Association of TLR2 gene polymorphisms with ocular behcet's disease in a Chinese han population. Invest Ophthalmol Vis Sci (2013) 54(13):8384–92. doi: 10.1167/iovs.13-12878
39. Tao K, Fujii M, Tsukumo S, Maekawa Y, Kishihara K, Kimoto Y, et al. Genetic variations of toll-like receptor 9 predispose to systemic lupus erythematosus in Japanese population. Ann Rheum Dis (2007) 66(7):905–9. doi: 10.1136/ard.2006.065961
40. Mhandire DZ, Mhandire K, Magadze M, Wonkam A, Kengne AP, Dandara C. Genetic variation in toll like receptors 2, 7, 9 and interleukin-6 is associated with cytomegalovirus infection in late pregnancy. BMC Med Genet (2020) 21(1):113. doi: 10.1186/s12881-020-01044-8
41. Jablonska A, Studzinska M, Szenborn L, Wisniewska-Ligier M, Karlikowska-Skwarnik M, Gesicki T, et al. TLR4 896A/G and TLR9 1174G/A polymorphisms are associated with the risk of infectious mononucleosis. Sci Rep (2020) 10(1):13154. doi: 10.1038/s41598-020-70129-4
42. Varshney D, Singh S, Sinha E, Mohanty KK, Kumar S, Kumar Barik S, et al. Systematic review and meta-analysis of human toll-like receptors genetic polymorphisms for susceptibility to tuberculosis infection. Cytokine (2022) 152:155791. doi: 10.1016/j.cyto.2021.155791
43. Zhao L, Liu K, Kong X, Tao Z, Wang Y, Liu Y. Association of polymorphisms in toll-like receptors 4 and 9 with risk of pulmonary tuberculosis: a meta-analysis. Med Sci Monit (2015) 21:1097–106. doi: 10.12659/MSM.893755
44. Mario-Vasquez JE, Naranjo-Gonzalez CA, Montiel J, Zuluaga LM, Vasquez AM, Tobon-Castano A, et al. Association of variants in IL1B, TLR9, TREM1, IL10RA, and CD3G and native American ancestry on malaria susceptibility in Colombian populations. Infect Genet Evol (2021) 87:104675. doi: 10.12659/MSM.893755
45. Kawasaki T, Kawai T. Toll-like receptor signaling pathways. Front Immunol (2014) 5:461. doi: 10.3389/fimmu.2014.00461
46. Rahman AH, Taylor DK, Turka LA. The contribution of direct TLR signaling to T cell responses. Immunol Res (2009) 45(1):25–36. doi: 10.1007/s12026-009-8113-x
47. Gelman AE, Zhang J, Choi Y, Turka LA. Toll-like receptor ligands directly promote activated CD4+ T cell survival. J Immunol (2004) 172(10):6065–73. doi: 10.4049/jimmunol.172.10.6065
48. Giulieri S, Manuel O. QuantiFERON®-CMV assay for the assessment of cytomegalovirus cell-mediated immunity. Expert Rev Mol Diagn (2011) 11(1):17–25. doi: 10.1586/erm.10.109
49. Gerna G, Lilleri D, Chiesa A, Zelini P, Furione M, Comolli G, et al. Virologic and immunologic monitoring of cytomegalovirus to guide preemptive therapy in solid-organ transplantation. Am J Transpl (2011) 11(11):2463–71. doi: 10.1111/j.1600-6143.2011.03636.x
Keywords: cytomegalovirus, kidney transplantation, single-nucleotide polymorphism (SNP), toll-like receptor 3 (TLR3), toll-like receptor 9 (TLR9)
Citation: Redondo N, Rodríguez-Goncer I, Parra P, Ruiz-Merlo T, López-Medrano F, González E, Polanco N, Trujillo H, Hernández A, San Juan R, Andrés A, Aguado JM and Fernández-Ruiz M (2022) Influence of single-nucleotide polymorphisms in TLR3 (rs3775291) and TLR9 (rs352139) on the risk of CMV infection in kidney transplant recipients. Front. Immunol. 13:929995. doi: 10.3389/fimmu.2022.929995
Received: 27 April 2022; Accepted: 05 July 2022;
Published: 29 July 2022.
Edited by:
Brian Duncan Tait, The University of Melbourne, AustraliaReviewed by:
A. Raj Kumar Patro, Kalinga Institute of Medical Sciences (KIMS), IndiaCopyright © 2022 Redondo, Rodríguez-Goncer, Parra, Ruiz-Merlo, López-Medrano, González, Polanco, Trujillo, Hernández, San Juan, Andrés, Aguado and Fernández-Ruiz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natalia Redondo, bmF0YWxpYS5yZWRvbmRvLmltYXMxMkBoMTJvLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.