 Zhidong Hu
Zhidong Hu Shui-Hua Lu
Shui-Hua Lu Douglas B. Lowrie
Douglas B. Lowrie Xiao-Yong Fan
Xiao-Yong Fan- 1Shanghai Public Health Clinical Center, Key Laboratory of Medical Molecular Virology of Ministry of Education (MOE)/Ministry of Health (MOH), Fudan University, Shanghai, China
- 2National Medical Center for Infectious Diseases of China, Shenzhen Third People Hospital, South Science & Technology University, Shenzhen, China
Tuberculosis (TB), caused by respiratory infection with Mycobacterium tuberculosis, remains a major global health threat. The only licensed TB vaccine, the one-hundred-year-old Bacille Calmette-Guérin has variable efficacy and often provides poor protection against adult pulmonary TB, the transmissible form of the disease. Thus, the lack of an optimal TB vaccine is one of the key barriers to TB control. Recently, the development of highly efficacious COVID-19 vaccines within one year accelerated the vaccine development process in human use, with the notable example of mRNA vaccines and adenovirus-vectored vaccines, and increased the public acceptance of the concept of the controlled human challenge model. In the TB vaccine field, recent progress also facilitated the deployment of an effective TB vaccine. In this review, we provide an update on the current virus-vectored TB vaccine pipeline and summarize the latest findings that might facilitate TB vaccine development. In detail, on the one hand, we provide a systematic literature review of the virus-vectored TB vaccines are in clinical trials, and other promising candidate vaccines at an earlier stage of development are being evaluated in preclinical animal models. These research sharply increase the likelihood of finding a more effective TB vaccine in the near future. On the other hand, we provide an update on the latest tools and concept that facilitating TB vaccine research development. We propose that a pre-requisite for successful development may be a better understanding of both the lung-resident memory T cell-mediated mucosal immunity and the trained immunity of phagocytic cells. Such knowledge could reveal novel targets and result in the innovative vaccine designs that may be needed for a quantum leap forward in vaccine efficacy. We also summarized the research on controlled human infection and ultra-low-dose aerosol infection murine models, which may provide more realistic assessments of vaccine utility at earlier stages. In addition, we believe that the success in the ongoing efforts to identify correlates of protection would be a game-changer for streamlining the triage of multiple next-generation TB vaccine candidates. Thus, with more advanced knowledge of TB vaccine research, we remain hopeful that a more effective TB vaccine will eventually be developed in the near future.
1 Introduction
Among the top 10 leading causes of death worldwide and the leading cause of death by a bacterial infection, tuberculosis (TB) caused 1.5 million deaths in 2020 (1). According to the World Health Organization (WHO), a quarter of the world population is infected with Mycobacterium tuberculosis (Mtb), the cause of TB (1). The Coronavirus Disease 2019 (COVID-19) pandemic reduced access to TB diagnosis and treatment and led to an increase in TB deaths in 2020, the first time during the past decade (1). Ambitious targets for TB control have been set by WHO. The “End TB Strategy” defined milestones and targets that aim to reduce TB incidence by 90% and deaths by 95% by 2035 compared with 2015 (2). However, progress has been slow and the 2020 milestone was far from reached (1). The lack of an optimal TB vaccine is regarded as one of the key barriers for TB control, thus, WHO timelines to control the global TB epidemic require a vaccine that is more effective, particularly in adolescents and adults. In this review, we focus on the history of research and the developmental progress of virus-vectored TB vaccines that are in clinical trials, those that are in pre-clinical animal research, and the latest ancillary findings facilitating TB vaccine research and development.
2 One-Hundred-Year-Old BCG: Successes and Failures
Mycobacterium bovis Bacille Calmette-Guérin (BCG), a vaccine based on attenuation of a bacterium naturally causing TB in cattle, is the only licensed TB vaccine up to now. BCG was first administered to the newborn infant of a woman with TB in France in 1921, neither adverse effects nor disease developed in the subsequent 5.5 years (3). In the next several years, thousands of children in families with a history of TB received BCG at the Pasteur Institute (4). In 1928, the intradermal route was found to be more reliable compared with the oral route, and this route continues to be used today (5). In the same year, the League of Nations (the predecessor of the United Nations) declared BCG to be safe for use. Although the “Lübeck disaster”, in which 72 newborn babies died from TB after BCG vaccination due to contamination with a virulent strain of Mtb (6), hindered the public acceptance of BCG for a long time, a resurgence of TB during World War II led to widespread BCG inoculation and public confidence in its safety was regained (7). In 1974, the WHO created the “Expanded Programme on Immunization”, to ensure that mothers and children have universal access to routinely recommended neonatal vaccines, and this resulted in more than 4 billion BCG vaccinations being administered to date (8). The vaccine has probably been administered to more humans than any other vaccine. Two-thirds of those countries giving BCG vaccination are estimated to have more than 90% coverage (9) and the vaccine is still the gold standard against which new candidates are compared.
The widespread use of the BCG vaccine in infants continues, primarily because it offers protection against the aggressive childhood forms of the disease: meningeal and miliary TB (10). However, for pulmonary TB prevention in adults, clinical trials have estimated its vaccine efficacy to range from 0% in south India to 80% in the UK (11). There are many hypotheses to explain this wide variation, including age at vaccination (12, 13), exposure to environmental mycobacteria (14), gender (15), risk of TB in the study population (16), etc. However, the proposed causes of variation often remain speculative and the basis is likely to be multifactorial.
The failure of TB control indicates that BCG is insufficient. Strategies to improve TB vaccination mainly address one of two approaches: optimization of the current BCG vaccine or development of novel vaccines such as subunit, vectored, and live attenuated vaccines. In the optimization of BCG we include BCG re-vaccination, change of inoculation route, and recombinant BCG construction. Although WHO does not recommend BCG re-vaccination due to a lack of proven efficacy of repeat doses for protection against TB, the most recently completed clinical trials showed BCG re-vaccination had an efficacy of 45.4% in primary Mtb infection prevention, which was defined immunologically by QuantiFERON-TB Gold In-tube assay conversion (17). However, the side effects of BCG re-vaccination hinder its application in humans with immune disorders. Although intravenous BCG immunization was consigned to the history books, this approach was recently re-evaluated in the non-human primate model of TB, in which nine out of ten intravenous BCG-vaccinated macaques showed slight or even no signs of TB disease post Mtb infection (18). However, safety concerns will impede application in humans. One of the most promising TB vaccines may provide an alternative. The genetically modified BCG-based vaccine VPM1002, in which the gene encoding urease C was replaced by the listeriolysin encoding gene from Listeria monocytogenes, showed the potential to replace the current BCG vaccine and is now undergoing three phase III clinical efficacy trials (19).
Besides the optimization of the current BCG vaccine, another approach is to utilize novel TB vaccines as a booster of the BCG vaccine, since most adults who acquire TB worldwide today were BCG-vaccinated as neonates. In the past decades, several viral vector-based vaccines and protein-adjuvant vaccines were designed to enhance BCG-primed immune protection. In this review, we focus on the research and development of viral vector-based TB vaccines.
3 Recombinant Virus-vectored TB Vaccines
Viruses provide some of the most widely used vaccine vectors. Recombinant virus-vectored vaccines are capable of inducing robust immune responses by mimicking the processes of pathogens invading the organism and resulting in the formation of long-lasting immune memory. Basically, most viral vaccine vectors have the following advantages: 1) they can accommodate genes encoding large antigenic fragments; 2) they have stable exogenous gene expression efficiency; 3) they can induce high levels of both cellular and humoral immune responses; 4) the immune responses induced by the vector itself have the potential to augment the antigen-specific immune memory to some extent; 5) they do not always require the use of adjuvants; 6) they are easy to manipulate and culture; 7) the use of attenuated or replication-deficient viruses with a clear mechanism of infection provides a strong safety profile; 8) strong immune memory can generally be induced by a single immunization, and repeated vaccinations might not be required (20–24). The major disadvantages of viral-vectored vaccines includes: 1) the pre-existing neutralizing antibodies against the vector might limiting its application in humans; 2) the host-induced anti-vector immunity might limit the booster vaccination strategies; 3) some viral vectors are not appropriate for use in immunocompromised individuals.
Although it was well illustrated that Th1 CD4+ T cell responses dominated anti-TB immune protection, Mtb can survive intracellularly for a long time after primary infection, the induction of immune responses that include high levels of cytotoxic T lymphocyte is also crucial to the clearance of intracellular Mtb (25–27). Cytotoxic T cells are prominent in the immune response to viruses and viruses accordingly provide one of the most widely used vector formats in the field of TB vaccines.
3.1 Mechanisms of Immune Protection Afforded by Virus-vectored Vaccines
The mechanisms of immune protection vary depending on the nature of the induction stimulus. Overall, recombinant viral-vectored vaccines carrying exogenous antigen fragments are able to invade host cells by using intrinsic viral mechanisms and undergo massive intracellular replication. The intracellular products and those secreted extracellularly induce cellular and humoral immune responses, respectively. The commonly-used attenuated or replication-deficient viruses are rapidly cleared after the host’s immune response is activated, while the antigen-specific immune cells are gradually transformed into memory cells that can remain for a long time. In addition, recombinant viral-vectored vaccines are able to induce a strong co-stimulatory molecular signaling and the formation of an inflammatory microenvironment, which together act as signals 2 and 3 of the T-cell/B-cell response pathways to enhance the host’s antigen-specific adaptive immunity.
3.2 Brief Introduction of the Widely Used Viral Vectors
The poxviruses are among the most widely studied viral vectors. They constitute a group of double-stranded DNA viruses that is divided into 2 subfamilies and 12 genera, among which, Orthopoxvirus, Molluscipoxvirus, Parapoxvirus, and Yatapoxvirus are known to infect humans (28, 29). Poxviruses of different genera infect different animals to cause different diseases and vaccinia virus, belonging to the genus Orthopoxvirus, although not fully non-pathogenic in human, has been a highly effective “live” vaccine against the smallpox epidemics that once ravaged humans (30). For safety reasons, most studies have chosen to use replication-deficient versions of poxviruses in vaccine vector development. The reduced immunogenicity consequent upon reduced replication can be offset by genetic modifications to knock-down molecules used by the virus to attenuate immune responses and by expression of immunostimulatory molecules in addition to the target antigens (31, 32). The types of genes encoded to enhance immune responses include: type I and type II interferons, genes regulating cytokines and chemokines, apoptosis and immunosuppression related molecules, antigen presentation signaling pathway molecules, etc. (33–35) Notable among poxvirus vaccine vectors are four strains of replication-deficient poxviruses, including modified vaccinia virus Ankara (MVA) (36), NYVAC derived from Copenhagen strain (37), ALVAC modified from canary poxvirus (38), avian poxvirus TROVAC (38), and another attenuated vaccinia strain, namely Chinese Tiantan strain poxvirus (39). The recombinant vaccine MVA85A, also known as AERAS-485, expresses Mtb immunodominant antigen Ag85A and was the first new TB vaccine to complete phase IIb clinical trials (40). It induced strong immune responses among Th1 and Th17 CD4+ T cells, in addition to moderate CD8+ T cell responses (41).
Adenovirus (Ad) is another widely used vaccine vector. About 50 human adenovirus (AdHu) serotypes have been identified, of which AdHu5 and AdHu35 are the two most widely used subtypes. Two human adenovirus-based recombinant vaccines against TB, AdAg85A (also known as AdHu5Ag85A) and AERAS-402 (also known as Crucell Ad35), in addition to ChAdOx1.85A, based on a chimpanzee adenovirus vector, are capable of inducing a strong CD8+ T-cell immune response in addition to high levels of Th1-type CD4+ T-cell immune response.
Other viral vectors including influenza virus, cytomegalovirus (CMV), Sendai virus (SeV), lentivirus, vesicular stomatitis virus (VSV), have also been applied to TB vaccine studies. Although these viruses infect cells by different mechanisms, most of them can induce high levels of antigen-specific Th1 CD4+ and CD8+ T cell immune responses as TB vaccine vectors. Among them, SeV85AB, a recombinant SeV-vectored vaccine expressing Ag85A and Ag85B, was the first viral-vectored TB vaccine found able to induce high levels of lung tissue-resident memory T cells (TRM)-mediated immune protection (42) and provided a new research direction for TB vaccines.
3.3 Brief History of Research and Development of Virus-vectored TB Vaccines
The recombinant poxvirus-vectored vaccine MVA85A was the first of the new TB vaccines to complete phase IIb clinical trials (40), but there are several other recombinant viral-vectored vaccines against TB in clinical and preclinical phases of evaluation. Herein, we give a brief overview of the history and the latest discoveries in the field.
3.3.1 MVA85A/AERAS-485
The MVA85A vaccine, which expresses the Mtb immunodominant antigen Ag85A, was developed by the University of Oxford in 2001, researchers found that intradermal (i.d.) or intramuscular (i.m.) immunization with this vaccine was able to induce a high level of antigen-specific immune response and protective immunity against Mtb challenge by decreasing the bacterial loads in organs in mouse models (43–45); subsequently, the vaccine-induced protection was further validated in other animal models such as guinea pigs (46), cattle (47) and rhesus monkeys (48). In the first phase I clinical trial, published in 2004, the vaccine was inoculated i.d. and induced a strong specific T-cell immune response in adults with or without a BCG immunization history (49). In 2010, the immunogenicity and safety of this i.d. vaccine were further confirmed in children and adolescents (50). Two phase I clinical trials completed in 2012 and 2013, were conducted to optimize the immunization dose (51) and route of administration (i.m. and i.d.) (52). During this period, from 2009 to 2011, researchers recruited 2,797 BCG-immunized infants between 4 and 6 months of age and i.d. administered either MVA85A or placebo and followed them for 19 to 28 months to complete the first phase IIb clinical trial (40). Although the safety of the vaccine was strongly demonstrated, the vaccine only induced weak antigen-specific immune responses and was not protective against TB disease (40), similar with the results that MVA85A did not reduce the bacterial burden of BCG-prime mice (53). Thus, the first viral-vectored TB vaccine clinical trial was declared a failure. In a proof-of-concept phase II trial, the number of immunization times was increased to two shots in a population of HIV-infected patients, but the efficacy, which was defined by QuantiFERON-TB Gold In-Tube conversion, remained poor (54). Nonetheless, researchers have not given up their efforts. A phase I clinical trial study in 2014 reported validation of the safety of mucosal immunization with MVA85A (55), and in 2019 a phase I clinical trial showed that an aerosol prime-intradermal boost regime was well-tolerated and induced potent antigen-specific mucosal and systemic immune responses (56). Additionally, i.m. inoculation was tested in a phase II clinical trial (NCT02178748) that indicated that a change in the route of administration may be a way to improve the vaccine’s protective efficacy. In 2021 a phase I trial showed that MVA85A delivered by aerosol was safe in UK adults with latent TB infection (57). Utility in a potential niche application was indicated in a phase II clinical trial that showed MVA85A vaccination in HIV-exposed newborns might be used to avoid the potential risk of BCG disease in this population (58).
Pre-clinical studies had indicated that the vaccine might work best as a booster in combination with other vaccines. To test this clinically, the vaccine was combined with the recombinant adenovirus-vectored vaccine AERAS-402 (59) with ChAdOx1.85A (currently in a phase II trial, NCT03681860) (60), with the recombinant avian poxvirus-vectored vaccine FP85A (61), or the protein adjuvant vaccine IMX313 (62), all of which had all been validated in phase I clinical trials.
Besides MVA85A, the potential of several other MVA-based recombinant TB vaccines has been indicated. For example, a multiphasic vaccine expressing 14 antigens representative of the three phases of TB infection (active, latent, and resuscitation) was subcutaneously (s.c.) immunized and induced potent multifunctional cell-mediated immunity in mice and rhesus macaque models (63). A recombinant MVA expressing α-crystallin by using i.d. route enhanced BCG-induced protection against Mtb infection in guinea pigs (64).
3.3.2 AdAg85A
Recombinant adenovirus vectors are widely used in the field of TB vaccine research. At least three vaccines are currently moving forward in clinical trials: AdAg85A based on AdHu5 (65, 66), AERAS-402 based on AdHu35 (67–73), and ChAdOx1.85A (60) based on a chimpanzee adenovirus vector, which would be discussed in detail below.
Recombinant AdHu5 vectored vaccine AdAg85A, was developed by McMaster University and published in 2004 (74). Similar to the MVA85A vaccine study, the safety, immunogenicity, and protective efficacy of the AdAg85A vaccine by using the aerosol and i.m. route were validated in animal models including mice (75–77), guinea pigs (78), cattle (79), goats (80), and rhesus macaques (81) before entering clinical trials. In the first clinical trial in 2013, AdAg85A was i.m. administrated in BCG-naïve and previously BCG-immunized healthy adults, and strong antigen-specific CD4+ and CD8+ T cell responses were observed (65). Most recently, in 2022, a phase Ib trial showed that aerosol delivery of AdAg85A was also safe and well-tolerated in previously BCG-vaccinated adults (66). A potential disadvantage of the vaccine is that substantial levels of anti-AdHu5 antibodies tend to be preexisting in humans, although the inventors of the vaccine demonstrated that AdHu5 antibodies do not affect the safety and immunogenicity of AdAg85A (65). However, a clinical trial of an AdHu5-based HIV vaccine was terminated due to the discovery that vaccinated subjects who had high titers of antibodies against adenovirus tended to have a higher incidence of HIV acquisition than those without anti-adenovirus antibodies in 2007 (82). Consequently, the role of preexisting vector-specific antibody responses remains controversial and there is currently an international preference for the use of AdHu35, the antibodies of which are largely absent from human serum. In addition, considering low sero-reactivity was observed in chimpanzee- and simian-derived adenoviral vectors compared with human-derived vectors in humans (83), several recombinant chimpanzee adenovirus-vectored TB vaccines were constructed, which will be described below. However, through a mouse model, the magnitude, quality and protective capacity of CD8+ T cells elicited using simian immunodeficiency virus Gag as the target antigen were compared, AdHu5 and AdCh3 vectors conferred the best efficacy (83, 84). These studies added a layer of complexity to balancing safety and vaccine efficacy in choosing adenovirus vectors.
3.3.3 AERAS-402/Crucell Ad35
The AdHu35-based recombinant vaccine, AERAS-402, expressing Ag85A, Ag85B, and TB10.4 was developed jointly by Crucell and the Aeras organization. In 2007, it was shown that this vaccine was i.m. immunized and was able to induce a strong T-cell immune response and a strong immune-protective effect against Mtb in a mouse model (85). The protection afforded by AERAS-402 singly or in combination with other vaccines was also validated in rhesus macaques (86–88). In 2010, the i.m. vaccine’s safety and immunogenicity were confirmed in healthy adults in a phase I clinical trial (67). In subsequent years, through several phase I and phase II clinical trials, researchers have expanded the potential target population to include healthy infants previously vaccinated with BCG (68), healthy adults immunized with BCG (69), adults with active or previous TB (70), latently infected populations (71), and HIV-infected patients (72). A two-dose i.m. regimen was also evaluated in BCG-vaccinated adults in phase I clinical trial in 2021 (73). Several AERAS-402-based phase II clinical trials targeting different populations, including adults treated for pulmonary TB, HIV-infected/BCG-vaccinated adults, and BCG-vaccinated healthy infants, have been completed (NCT02414828, NCT01017536, and NCT01198366). The safety of i.m. AERAS-402 was confirmed through these trials.
3.3.4 ChAdOx1.85A
To minimize any effects of preexisting anti-adenovirus antibodies in humans, researchers have developed the recombinant replication-deficient chimpanzee adenovirus-vectored vaccine ChAdOx1.85A. Its i.m. immunization is capable of inducing high levels of cellular immune response in BCG-primed mice and showing protective efficacy against Mtb infection in combination with MVA85A (89, 90). In 2020, a phase I clinical trial demonstrated that a ChAdOx1.85A i.m.-MVA85A i.m. vaccination regimen was well tolerated and immunogenic in healthy UK adults (60). As mentioned above, this vaccine strategy is now in a phase II clinical trial (NCT03681860).
Intranasally (i.n.) immunization with a recombinant ChAdOx1 vaccine expressing Rv1039c (PPE15) instead of Ag85A conferred better protection than ChAdOx1.85A in a murine model, meriting further evaluation in clinical trials (91). Similarly, a recombinant chimpanzee adenovirus-68-vectored vaccine expressing Ag85A, namely AdCh68Ag85A, was i.n. immunized and found to be superior to AdAg85A in the induction of T-cell responses and protection against Mtb infection in mice that had previously been exposed to human adenovirus (92). In addition, there is evidence that this vaccine could be used as a therapeutic vaccine: Immunotherapy with a single-dose respiratory mucosal but not parenteral application of AdCh68Ag85A as an adjunct to antibiotic therapy accelerated pulmonary Mtb clearance, limited lung pathology, and restricted disease in mice (93).
3.3.5 TB/FLU-04L
In 2006, a recombinant influenza virus-vectored vaccine expressing ESAT-6 was shown to be able to induce a high level of Th1 CD4+ T cell immune response with two i.n. injections in a mouse model (94). The protective efficacy of the vaccine was confirmed in mice and guinea pigs (95). This vaccine, named TB/FLU-04L, was aerosol immunized and completed a phase I clinical trial in 2015, but no study results have been published (NCT02501421). According to WHO reports in 2017 (96), a phase IIa clinical trial is being conducted in patients with latent TB infection. Besides TB/FLU-04L, another recombinant influenza virus-based vaccine expressing the dominant peptides of Ag85B was constructed, and robust TRM responses and protective efficacy were observed in a murine model by using i.n. route (97).
3.3.6 MCMV85A and RhCMV/TB
In 2014, a recombinant murine CMV-vectored vaccine MCMV85A expressing Ag85A was developed by the University of Oxford. This vaccine was inoculated intraperitoneal (i.p.) or intravascular (i.v.), and activated NK cells to provide early nonspecific protection against Mtb infection, which was further potentiated by a weak 85A-specific T cell response in a murine model (98). In 2018, a rhesus monkey CMV vector vaccine RhCMV/TB was described that encoded nine proteins from three phases of Mtb infection: acute (Ag85A, Ag85B, ESAT-6), latency (Rv1733, Rv2626, Rv3407), and resuscitation (RpfA, RpfC, RpfD). Two doses of s.c. RhCMV/TB induced high levels of specific CD4+ and CD8+ T cell immune responses, and provided long-lasting vaccine-mediated immune control after highly pathogenic Mtb strains challenge one year after immunization in rhesus macaques, in which 41% animals showed no TB disease evaluated by computed tomography scans or necropsy (99). However, although human CMV infection only causes asymptomatic infection in the immunocompetent population, CMV is highly species-specific and systemic disease with severe complications and high mortality rate might be occurred in immunocompromised individuals (100, 101). Moreover, epidemiological studies have identified the increased human CMV infection is an important risk factor for active TB disease and latent TB infection, which was found to be associated with the magnitude of IgG, enhanced CMV-driven T-cell activation, systemic inflammation, and immune dysregulation (102, 103). Thus, more animal and clinical studies are warranted to better understand CMV-vectored immunity, to ensure its safe translation to humans, especially in active TB patients and individuals with latent TB infection.
3.3.7 SeV85AB
SeV85AB, a recombinant SeV-vectored vaccine, is the first application of a SeV vector to the TB vaccine development and it expresses Mtb immunodominant antigens Ag85A and Ag85B and inherently has a high safety profile. Being based upon an RNA virus, the SeV vector has no risk of integration with the human genome. Furthermore, in contrast to respiratory pathogens such as the influenza virus, the SeV does not cause human disease and there are very low antibody levels present. In 2017, using a mouse model, we validated its immunogenicity and protective efficacy against Mtb infection in mice and demonstrated the establishment of a high level of TRM-mediated immune response in mucosal tissues by using i.n. route (42). Such memory cells can establish the first line of defense in the lung against Mtb invasion in the early phase of infection. In contrast, BCG vaccination usually produces a response of memory T cells in the circulatory system only after several weeks of infection. Therefore, this vaccine may be used to optimize the systemic BCG-induced immune protection against Mtb infection (104). This immunization strategy was further optimized in combination with recombinant DNA vaccines for improved protective efficacy (105).
3.3.8 Other Viral-vectored Vaccines
Several other promising viral vectors are being explored as candidates for TB vaccine construction. In 2008, Hamamatsu University School of Medicine constructed a lentiviral vector vaccine expressing MPT51 of Mtb. This construct enhanced the antigen presentation efficiency of dendritic cells, and intratracheal (i.t.) immunization of mice was able to induce a CD8+ T-cell immune response at the lung site and protection against Mtb infection (106). Recently, several other lentiviral vector-based TB vaccines have been developed, but they are at early animal model phases of investigation (107–110). Lentiviral vectors have been successfully used in the clinical trials of patients with advanced leukemia and other gene immunotherapy research (111, 112). In most of these TB vaccine studies described above, self-inactivation or non-integrating vector systems were chosen to get safe vaccines.
In 2008, researchers at McMaster University constructed a recombinant VSV-vectored vaccine VSVAg85A that expressed Ag85A and was able to induce an antigen-specific immune response and protection against Mtb infection but only for a short duration by using i.n. or i.m. routes. Combining the vaccination with AdAg85A in a prime-boost immunization strategy improved the protective efficacy (113); VSV-based boosting resulted in inferior protection compared with adenovirus-based boosting, and this was associated with differentially imprinted innate phagocytes at the mucosal site of immunization (114). Besides VSVAg85A, another VSV-based TB vaccine expressing Rv2660c, Rv3615c, and Mtb10.4 has generated antigen-specific T cell responses and immune protection in a BCG challenge murine model by using i.n. route (115, 116).
The use of a combination of vectors expressing the same antigen in order to enhance responses is a recurring theme. As mentioned in Section 3.3.1 above, a recombinant fowlpox virus FP9 that expressed Ag85A (FP9.Ag85A or FP85A) and boosted BCG/MVA85A-induced protective immunity in guinea pigs (46) also boosted immune responses to MVA85A in a clinical phase I trial in 2013 (61).
In 2014, a recombinant human parainfluenza type 2 virus-vectored vaccine expressing Ag85B, rhPIV2-Ag85B, was developed by the National Institute for Biomedical Innovation in Japan (117). This i.n. vaccine was able to induce a T-cell immune response and immune protection in a murine model that was subsequently found to be associated with induction of bronchus-associated lymphoid tissue (118). Similarly, a parainfluenza virus 5 vector expressing Ag85A and Ag85B has also shown immunogenicity and protective efficacy in a murine infection model by using i.n. route (119).
In 2020, different prime-boost strategies using the chimpanzee Ad3 (ChAd3) and MVA vectors expressing Ag85B, ESAT-6, Rv1733, Rv2626, and RpfD, were evaluated for immunogenicity and protective efficacy in highly susceptible rhesus macaques through different inoculation routes such as i.m., i.d., and aerosol. However, although specific immune responses were induced, none of these vaccine strategies conferred a protective effect compared to non-vaccinated controls (120).
To be noted, most of these studies used Mtb immunodominant antigens such as Ag85A, Ag85B, TB10.4, ESAT-6, etc., which were chosen based on their expression levels in Mtb and IFN-γ-inducing ability. However, the failure of MVA85A in its first phase IIb clinical trial suggest other antigens should be selected to construct a more effective vaccine. Recently, an unbiased immunopeptidomics pipeline for identifying novel antigens presented by MHC was developed, in which MHC I and MHC II complexes from BCG-infected THP-1 macrophages were immunoprecipitated and analyzed by liquid chromatography tandem mass spectrometry (121). Thus, identifying more efficient antigens by novel assays is also important in virus-vectored TB vaccine development.
Table 1 summarizes the viral vectored TB vaccine candidates that are currently in clinical trials, and Table 2 summarizes the candidates that are currently in preclinical animal model phases. These studies of novel virus-vectored TB vaccines have been successful in developing a number of candidates that have entered the TB vaccine pipeline (https://www.tbvi.eu/what-we-do/pipeline-of-vaccines/) and are at different stages of clinical trials in humans. This may lead to newly licensed vaccines capable of replacing/supplementing the current BCG vaccine and even conferring therapeutic benefit in patients with active/latent TB.
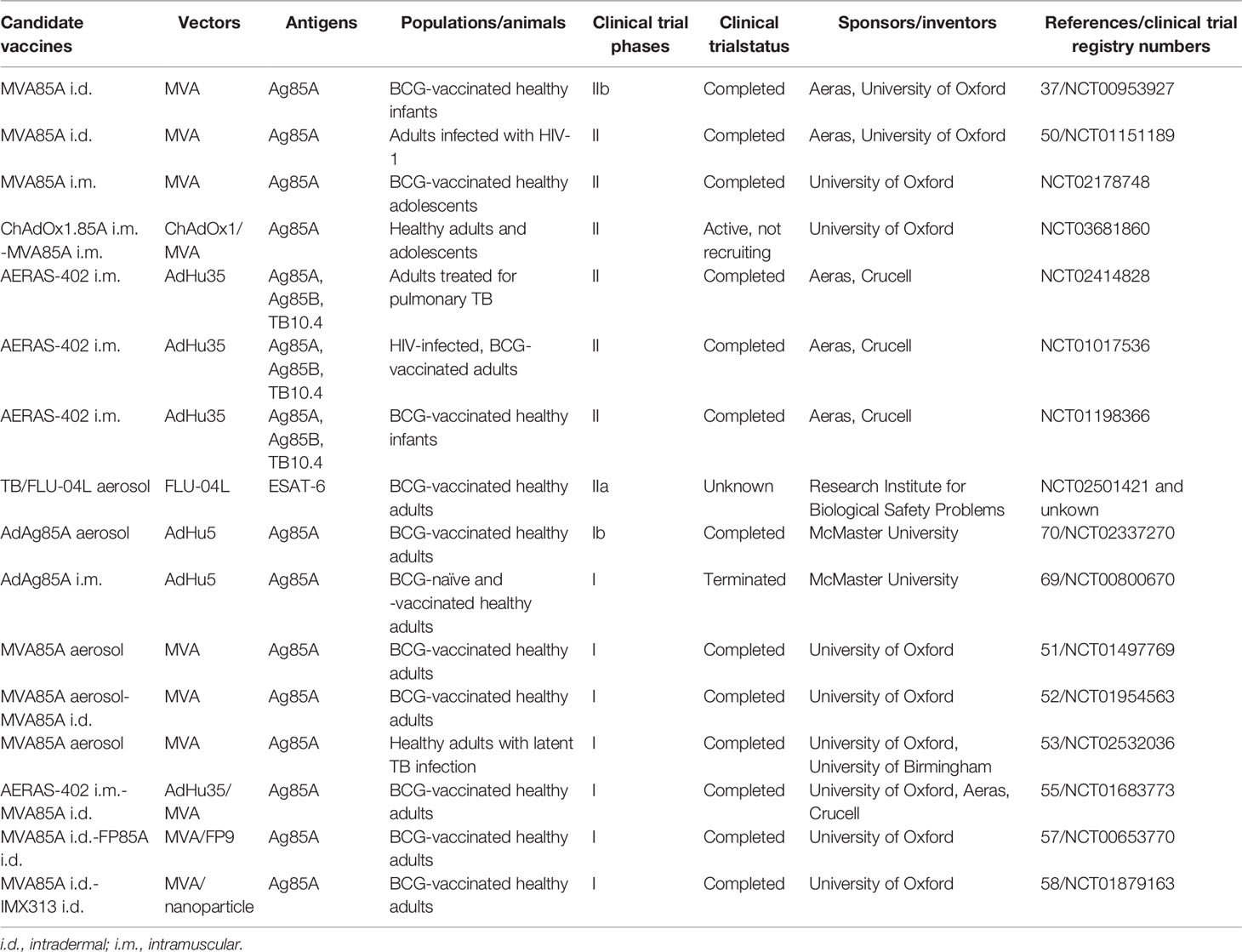
Table 1 Viral vectored TB vaccine candidates that are currently in clinical trials.
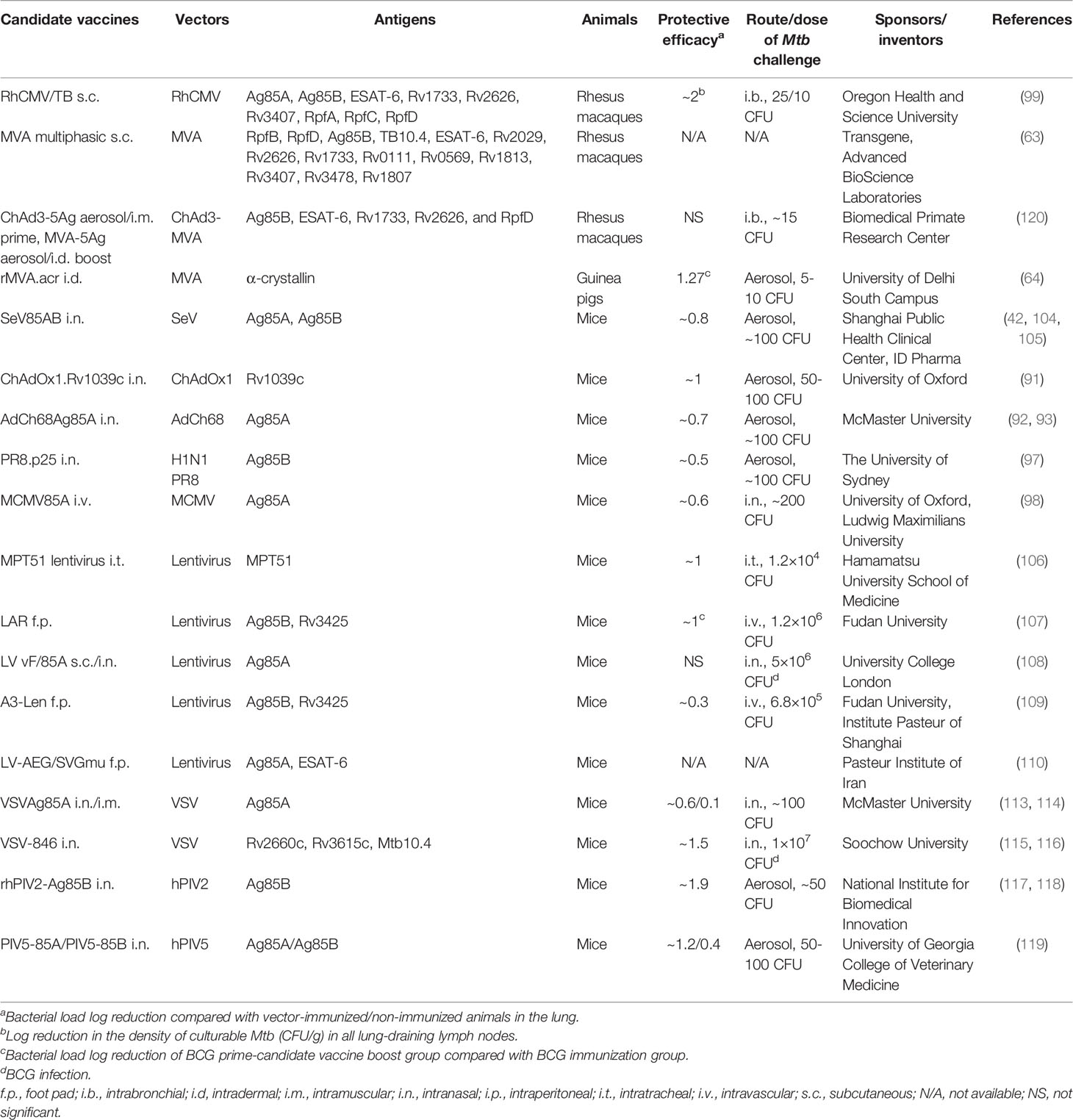
Table 2 Viral vectored TB vaccine candidates that are currently in preclinical animal model phases.
4 Latest Findings That Facilitate TB Vaccine Research Development
Over the past two decades, huge progress has been achieved in the field of TB vaccine development and more than a dozen candidate vaccines including viral-vectored vaccines are in clinical trials now. However, several knowledge gaps and challenges to the successful development of a universally effective TB vaccine remain. Here we summarize the latest findings facilitating TB vaccine research development.
4.1 Controlled Human Infection Model
The immunogenicity and protective efficacy of the TB vaccines that have entered clinical trials were first repeatedly validated in animal models such as mice, guinea pigs, and rhesus macaques before trial commencement. Validation in these animal models is not only time-consuming and costly but there is also a high technical barrier in undertaking Mtb challenge experiments in animals (requiring prolonged use of ABSL-3 level laboratories). In addition, the failure of the phase IIb clinical trial of the MVA85A vaccine showed that the currently available animal models do not predict human immunity well. Moreover, in the clinical trials evaluating TB vaccines, the assessment of protective efficacy relies on natural exposure to Mtb infection and requires enrollment and follow-up of tens of thousands of people for multiple years. The limited availability of suitable human populations in which this can be undertaken and the enormous cost of clinical trials further hinder the progress of TB vaccine development.
In recent years, the concept of a controlled human infection model has come under consideration to facilitate vaccine research progress. In this model, healthy volunteers are vaccinated with candidate vaccines and then are deliberately infected with the corresponding pathogen. The efficacy of the candidate vaccine is then assessed by either presence or absence of established infection or by disease progress. Recently, Vaxchora, a Cholera vaccine, was approved based on a human challenge study (122). In the field of TB, deliberately infecting healthy volunteers with virulent Mtb would not be ethical, since there is currently no method of anti-TB treatment that could reliably completely eradicate infection. As an alternative, Helen McShane, a co-inventor of MVA85A, described in 2012 a model in which humans are challenged with BCG (123). In this model, BCG-naive and BCG-vaccinated healthy volunteers in the UK were challenged with intradermal BCG, and the bacterial load was quantified from punch biopsies by PCR and bacterial culture (123). This model was used to assess the protective effect of MVA85A on BCG-vaccinated healthy adults (124), the data support the contention that this intradermal BCG challenge model is able to detect differences in anti-mycobacterial immunity induced by vaccination. In addition, an aerosol BCG challenge study is now underway in healthy UK adults to mimic the natural route of exposure (NCT02709278, NCT03912207).
Recently, Sarah Fortune and Eric Rubin described the development of an Mtb human challenge model, in which, Mtb’s growth is controlled by dependence on the availability of selection compounds; the bacteria are no longer viable once those compounds are removed. The study has not yet been published but was described in a commentary paper (125). This human Mtb infection model could substantially reduce the numbers of participants, study duration, and economic costs in TB vaccine studies.
4.2 Ultra-Low-Dose Aerosol Murine and Non-human Primate Infection Models
In general, in pre-clinical TB vaccine efficacy evaluation, animals are always infected using a single large bolus of Mtb that is delivered to the lungs by intratracheal or aerosol installation. However, in natural infection most people are infected by repeated inhalation of low doses. In animal models, the evidence for the efficacy of a TB vaccine is usually accepted as significant by the demonstration of 0.5-1 log10 lower numbers of Mtb in vaccinated compared to control animals at some point after challenge infection. A wide range of experimental and cost constraints dictate that the demonstration of larger and more meaningful effects with smaller infection challenges is difficult to achieve. Consequently, vaccine development often moves forward on the basis of modest vaccine impact compared to the potency required in human clinical efficacy testing, in which at least a 60% improvement in efficacy in prevention of the disease compared to BCG alone is required (126). These divergences between laboratory models and clinical scenarios hinder TB vaccine development.
Optimization of Mtb challenge doses in pre-clinical TB vaccine evaluation was undertaken in non-human primate models. Recent studies tended to use a lower dose than prior vaccine studies. For instance, in 2018, in the RhCMV/TB vaccine study, the monkeys received 10 or 25 bacteria (99). In 2019, a repeated limiting-dose challenge model used an average of 1.3 bacteria implanted weekly for 8 consecutive weeks by endobronchial installation (127). In 2020, in the BCG immunization route optimization study, the monkeys were challenged by bronchoscope with 4-36 bacteria (18). These low-dose-infection non-human primate models better mimicked the natural course of TB infection in humans and allowed investigators to observe vaccine-mediated sterilizing immunity.
In 2021, a model of ultra-low-dose aerosol infection in mice was established, in which infection was initiated by only 1-3 founding bacteria, instead of the conventional ~100 CFU dose. As in human TB, highly heterogeneous bacterial burdens, immune responses, and disease manifestations were observed in this model (128). In addition, the well-circumscribed granulomas shared features with human granulomas. Thus, this ultra-low dose infection murine model more closely replicates human disease. It is also much cheaper and easier to handle than the low-dose non-human primate models, thus, it might facilitate preclinical testing of vaccine and immunotherapeutic candidates and act as a gatekeeper to determine which vaccines show promise and warrant further testing.
4.3 TRM-Mediated Anti-TB Immune Protection
TRM represents a distinct subset of memory T cells that was found in the past decade. Unlike other memory T cells such as central memory and effector memory T cells, TRM cells colonize local tissues infected by pathogens and remain there for a long time after the pathogen has been eliminated without participating in blood circulation (129). They have been demonstrated at sites that include the intestines, skin, urogenital tract, and lung mucosa (130–133). When the pathogen invades again, TRM cells immediately sense and initiate immune responses so that the pathogen can be controlled or be eliminated at the early stage of infection. This process does not depend on memory cells in the peripheral circulation and is in the first line of defense of the body as an adaptive immune response to pathogen infection (134). In 2014, Daniel L. Barber’s group reported for the first time the role of lung TRM in anti-TB infection in a murine model. The Mtb-specific CD4+ T cells in lung tissues could be divided into two populations, namely, a subpopulation of TRM colonizing the lung parenchyma and other memory cell subsets circulating in the vasculature; the former were identified with molecular markers as KLRG1-CXCR3+ and the latter as KLRG1+CX3CR1+ (135). The adoptive transfer of lung Mtb-specific TRM between mice resulted in potent immune protection (135–137). Based on these observations, our investigation of the properties of circulating CD4+ T cells in patients with active TB led us to suggest that inhibition of KLRG1+ expression through the incorporation of a specific inhibitor of the Akt signaling pathway in a vaccine could enhance the protective responses in immunotherapeutic and perhaps prophylactic vaccination regimens (138).
In 2016, Stefan H. E. Kaufmann’s group reported for the first time TRM-mediated immune protection against TB infection, which was induced by mucosal delivery of BCG (139) and we reported that SeV85AB induced high levels of lung CD8+ TRM by using a comprehensive intravascular staining method (42). In the same year, several TB vaccines were also reported to be able to induce TRM (140–142). In fact, as early as 2010, researchers had found that treatment with FTY720 (an immunosuppressant that blocks memory cells in circulation (143)) partially counteracted the immune protection induced by the BCG vaccine (144), indirectly demonstrating that BCG possesses the ability to induce a certain level of TRM-mediated immune protection. In 2020, by using an intravascular staining method in non-human primates, intravenous BCG was shown to induce higher levels of lung parenchymal CD4+ T cells compared with intradermal vaccination, and this was associated with sterilizing immunity against Mtb challenge (18), indicating that vaccine-induced TRM also conferred Mtb resistance in this model.
By mimicking the route of infection in vaccination, a mucosal or intravenous vaccination might be an optimal vaccination strategy, targeting the induction of immune responses at the point of entry of the bacteria. However, the role of TRM in TB protection awaits further experimental confirmation; a better understanding of vaccine-induced lung TRM would facilitate novel TRM-targeting vaccine designs.
4.4 Role of Trained Immunity in TB Vaccine Development
Traditionally, vaccine development is mainly focused on the induction of the adaptive immune response that elicits antigen-specific long-term immune memory against infection. However, recently it has been shown that innate immunity also plays an important role in immune memory against homologous or even heterologous challenges (145, 146). Trained immunity, a de facto innate immune memory, has been defined as a long-term functional reprogramming of the innate immune cells that is evoked by endogenous or exogenous insults, with the cells then returning to a non-activated state and showing altered inflammatory responses against a second challenge (147, 148).
In 2020, a randomized clinical trial of BCG vaccination in the elderly showed immune protection against heterologous infections and improved survival (149). In 2021, another investigator-blind randomized controlled trial showed that BCG vaccination at birth significantly reduced all-cause infectious disease morbidity during the neonatal period (150). Trained immunity was proposed to be implicated in such BCG-induced heterologous protection. The first report of BCG-induced trained immunity showed that BCG vaccination in healthy volunteers enhanced the release of monocyte-derived cytokines in response to unrelated bacterial and fungal pathogens, and induced lymphocyte–independent protection of immunodeficiency SCID mice against disseminated candidiasis (151). In a study of BCG-induced trained immunity against Mtb infection in people, a global DNA methylation analysis revealed a stable and robust differential DNA methylation pattern among the promoters of genes belonging to immune pathways in “responders” to BCG vaccination but not in non-responders. Responders were defined as having an enhanced macrophage capacity to restrict the growth of Mtb associated with higher levels of IL-1β production (152). In rhesus macaques, mucosal or intravenous BCG inoculation conferred better protection against Mtb infection and TB disease than standard intradermal vaccination, and this was associated with the induction of enhanced trained immunity (153). β-glucan-induced trained immunity also afforded protection against Mtb infection (154). In contrast to BCG, Mtb infection impairs the development of protective trained immunity through impacting IFN-I signaling (155). These data suggest that vaccines that are aimed at enhancing trained immunity might give better protection against Mtb infection.
In 2018, the inventors of AdAg85A reported that respiratory infection with adenovirus could induce alveolar macrophages (AMs) that had a long-lasting memory that was sustained by an enhanced trained immunity phenotype in the local mucosal sites (156). This study suggested that non-specific trained immunity induced by the virus-vectored TB vaccine might contribute to the immune protection against Mtb infection. In 2020, they used a murine model of TB vaccination to investigate the role of AMs in host defense against Mtb and showed that respiratory mucosal immunization with AdA85A provided a type of trained immunity capable of potent protection against Mtb in the early stage of infection (157). In 2022, they further showed that mucosal immunization is superior to intramuscular immunization for the induction of trained immunity in AMs in a murine model of the SARS-CoV-2 vaccine (158), further adding to the evidence for the importance of local induction of trained immunity.
Cumulatively, the evidence suggests that the deciding battleground is the apoptosis of Mtb-infected macrophages in early infection, which is mediated by AMs and is enhanced by trained immunity. However, our understanding of the relative contribution of trained immunity to viral-vectored vaccine induction of T cell-mediated immune protection against TB remains limited. In addition, it remains to be answered that whether the anti-TB immune protection induced by BCG-prime-viral vector boost strategy is associated with the trained immunity, and the effective of live viral vector boost on innate immune training by BCG prime. Unraveling the elaborate molecular mechanisms of trained immunity will be critical for devising novel approaches to optimize the exploitation of trained immunity by TB vaccines.
4.5 Lack of Validated Immune Correlates of Protection by TB Vaccines
COPs are defined as laboratory biomarkers that are associated with protection from clinical disease. In particular, vaccine-inducible COPs are expected to be transformative in developing novel vaccines as they will de-risk the selection of candidate vaccines for human efficacy studies at an early stage. They might also substantially reduce the costs of large-scale clinical trials by helping to tailor the selection of participants being enrolled and by measuring vaccine immunogenicity and potential efficacy as a supplement, or even sometimes an alternative, to assessments of disease burden (159). Once validated in efficacy trials, COPs could potentially facilitate the development and licensure of vaccines. The absence of reliable parameters that could be used as COPs for TB vaccines represents one of the greatest challenges in TB vaccine development.
The complexities of Mtb infection create challenges in finding predictive markers of protective efficacy. The Th1 cytokines IFN-γ, IL-2, TNF-α, the Th17 cytokine IL-17, and other cytokines are active in the immune response against Mtb and are used as biomarkers to determine the antigen-specific T cell responses in TB vaccine evaluation research. However, studies have found that IFN-γ accounted for only about 30% of the CD4+ T cell-mediated immune protection against Mtb infection; and its overexpression even accelerated death in infected mice (137). More recently, close contacts of active TB patients who were persistently negative by IFN-γ release assay and tuberculin skin tests were defined as “resisters” of Mtb (160) and in 2019, a cohort study showed that “resisters” possess IgM and class-switched IgG antibody responses and non-IFN-γ T cell responses to Mtb-specific proteins (161), challenging the rationality of focus on assessing IFN-γ-based immunogenicity in TB vaccine design. In 2020, we found that T-cell activation status marker CD69 is associated with Mtb infection and may have the potential to distinguish latent TB infection (positive IFN-γ responses) and “resisters” (negative IFN-γ responses) (162). Based on these and similar studies, there is an urgent need to find novel molecular markers that are more correlated with immunogenicity/protective efficacy to be able to more accurately predict the protective efficacy of vaccines and to accelerate vaccine evaluation.
As mentioned above, TRM showed a potential for application among novel COPs in TB vaccine research. However, validation of vaccine-induced COPs is possible only when successful placebo-controlled efficacy trials become available. Only then can compelling comparisons be made. In 2019, a phase IIb clinical trial of the subunit TB vaccine M72/AS01E showed 49.7% efficacy against progression to TB compared with placebo control. This is the first novel TB vaccine to almost reach 50% protection in the past century (163). These vaccine cohorts offered an opportunity to identify COPs of vaccine-induced immune protection against Mtb infection and some strategies are in place. Bill & Melinda Gates Medical Research Institute, vaccine manufacturers, sponsors of clinical trials, and trial investigators have launched an international “TB Immune Correlate Program” consortium to identify immunological COPs for TB. The first priority is informed by existing knowledge and recent findings from animal models and clinical studies, including the magnitude of mycobacteria-specific Th1/Th17 CD4 T cell responses, magnitude/subclass/avidity of mycobacteria-specific mucosal IgA or IgG antibody responses, Fc-mediated, functional antibody activities, and trained immunity (27). However, the relatively small number of participants that reached clinical endpoints in the M72/AS01E clinical trial might restrict the statistical power of COPs discovery. Thus, larger clinical trials or the human infection model study are needed to validate the COPs that might be identified in the ongoing efforts.
5 Conclusion
Identified at the end of 2019, COVID-19 became a global public health threat within 3 months, it spread over the globe so rapidly that it was declared to be a “pandemic” by the WHO in March, 2020. Vaccinologists worked on the challenge immediately, leading to the development and deployment of novel vaccines within one year. Up to February of 2022, 144 and 195 COVID-19 vaccine candidates based on diverse platform technologies are being evaluated in clinical and preclinical stages, respectively, and dozens of vaccines have already been licensed to human use (164). The rapid COVID-19 vaccine development and deployment is critical for the world to return to pre-pandemic normalcy. Ironically, the only licensed TB vaccine is still the one-century-old BCG, which is inadequate. Thus, TB remains a leading cause of mortality from an infectious disease, only now surpassed by SARS-CoV-2 causing COVID-19. Considering the morbidity and mortality that is suffered from TB globally, it is time to accelerate commitment, investment, and implementation to stop the infectious disease agent that has killed more human beings than any other.
We believe that the success of COVID-19 vaccines and recent progress in TB vaccine research illustrate that the deployment of an effective TB vaccine is likely in the near future. The highly efficacious COVID-19 vaccines accelerated the vaccine development process in human use, with the notable example of mRNA vaccines and adenovirus-vectored vaccines, and increased the public acceptance of the concept of the controlled human challenge model, which might provide valuable experience on the development of TB vaccines. In this review, we have provided an update on the current viral vectored TB vaccine pipeline and summarized the latest findings that might facilitate TB vaccine developments. On the one hand, several viral vectored TB vaccines are in clinical trials, and other promising candidate vaccines at an earlier stage of development are being evaluated in preclinical animal models, and this sharply increases the likelihood of developing an effective TB vaccine in the near future, although this is far from certain. On the other hand, we propose that a better understanding of the lung-resident TRM-mediated mucosal immunity, and the unique trained immunity of phagocytic cells against intracellular Mtb infection, could help provide novel targets for innovative and superior TB vaccine designs. Moreover, new tools, such as controlled human infection and ultra-low-dose aerosol infection murine infection models, should facilitate TB vaccine development and selection in the preclinical phase of the investigation. In addition, identification of COPs in the M72/AS01E trial and other ongoing clinical trials could be valuable in streamlining triage and evaluation of next-generation TB vaccine candidates. Allocation of resources must include the discovery and development of early pipeline candidates to increase clinical trial capacity. With more advanced knowledge, we remain hopeful that a more effective TB vaccine will be developed sooner rather than later.
Author Contributions
ZH and X-YF conceived, designed and wrote the manuscript, X-YF, S-HL and DL edited the manuscript with conceptual advice. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by National Key Research and Development Program of China (2021YFC2301503), Grants from the National Natural and Science Foundation of China (81873884, 82171815, 82171739), Shanghai Science and Technology Commission (19XD1403100, 20Y11903400), and Shanghai Municipal Medical and Health Excellent Young Talents Training Program (GWV-10.2-YQ01).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. WHO. WHO Globlal TB Report. (2021). https://www.who.int/teams/global-tuberculosis-programme/tbreports/global-tuberculosis-report-2021
2. Uplekar M, Weil D, Lonnroth K, Jaramillo E, Lienhardt C, Dias HM, et al. WHO's New End TB Strategy. Lancet (2015) 385(9979):1799–801. doi: 10.1016/S0140-6736(15)60570-0
3. Calmette A, Guérin C, Boquet A, Nègre L. La Vaccination Préventive Contre La Tuberculose Par Le" BCG," [M]. Masson et cie (1927).
4. Calmette A. Preventive Vaccination Against Tuberculosis With BCG. Proc R Soc Med (1931) 24(11):1481–90. doi: 10.1177/003591573102401109
5. Wallgren A. Intradermal Vaccinations With BCG Virus: Preliminary Note. J Am Med Assoc (1928) 91(24):1876–81. doi: 10.1001/jama.1928.02700240030008
6. Lange L, Pescatore H. Bakteriologische Untersuchungen Zur Lübecker Säuglingstuberkulose. Die Säuglingstuberkulose in Lübeck. Springer (1935) 205–305. Available at: https://link.springer.com/chapter/10.1007/978-3-642-92013-4_3
7. Fine PE, Carneiro IA, Milstien JB, Clements CJ, Organization WH. Issues Relating to the Use of BCG in Immunization Programmes: A Discussion Document. World Health Organization (1999). Available at: https://apps.who.int/iris/bitstream/handle/10665/66120/WHO_V_B_99.23.pdf
8. Lobo N, Brooks NA, Zlotta AR, Cirillo JD, Boorjian S, Black PC, et al. 100 Years of Bacillus Calmette-Guerin Immunotherapy: From Cattle to COVID-19. Nat Rev Urol (2021) 18(10):611–22. doi: 10.1038/s41585-021-00481-1
9. Organization WH. WHO-UNICEF Estimates of BCG Coverage. (2020). https://apps.who.int/immunization_monitoring/globalsummary/timeseries/tswucoveragebcg.html
10. Trunz BB, Fine P, Dye C. Effect of BCG Vaccination on Childhood Tuberculous Meningitis and Miliary Tuberculosis Worldwide: A Meta-Analysis and Assessment of Cost-Effectiveness. Lancet (2006) 367(9517):1173–80. doi: 10.1016/S0140-6736(06)68507-3
11. Mangtani P, Abubakar I, Ariti C, Beynon R, Pimpin L, Fine PE, et al. Protection by BCG Vaccine Against Tuberculosis: A Systematic Review of Randomized Controlled Trials. Clin Infect Dis (2014) 58(4):470–80. doi: 10.1093/cid/cit790
12. Whittaker E, Nicol MP, Zar HJ, Tena-Coki NG, Kampmann B. Age-Related Waning of Immune Responses to BCG in Healthy Children Supports the Need for a Booster Dose of BCG in TB Endemic Countries. Sci Rep (2018) 8(1):15309. doi: 10.1038/s41598-018-33499-4
13. Nguipdop-Djomo P, Heldal E, Rodrigues LC, Abubakar I, Mangtani P. Duration of BCG Protection Against Tuberculosis and Change in Effectiveness With Time Since Vaccination in Norway: A Retrospective Population-Based Cohort Study. Lancet Infect Dis (2016) 16(2):219–26. doi: 10.1016/S1473-3099(15)00400-4
14. Abubakar I, Pimpin L, Ariti C, Beynon R, Mangtani P, Sterne JA, et al. Systematic Review and Meta-Analysis of the Current Evidence on the Duration of Protection by Bacillus Calmette-Guerin Vaccination Against Tuberculosis. Health Technol Assess (2013) 17(37):1–372v–vi. doi: 10.3310/hta17370
15. Rhodes SJ, Knight GM, Fielding K, Scriba TJ, Pathan AA, McShane H, et al. Individual-Level Factors Associated With Variation in Mycobacterial-Specific Immune Response: Gender and Previous BCG Vaccination Status. Tuberculosis (Edinb) (2016) 96:37–43. doi: 10.1016/j.tube.2015.10.002
16. Mangtani P, Nguipdop-Djomo P, Keogh RH, Trinder L, Smith PG, Fine PE, et al. Observational Study to Estimate the Changes in the Effectiveness of Bacillus Calmette-Guerin (BCG) Vaccination With Time Since Vaccination for Preventing Tuberculosis in the UK. Health Technol Assess (2017) 21(39):1–54. doi: 10.3310/hta21390
17. Nemes E, Geldenhuys H, Rozot V, Rutkowski KT, Ratangee F, Bilek N, et al. Prevention of M. Tuberculosis Infection With H4:IC31 Vaccine or BCG Revaccination. N Engl J Med (2018) 379(2):138–49. doi: 10.1056/NEJMoa1714021
18. Darrah PA, Zeppa JJ, Maiello P, Hackney JA, Wadsworth MH 2nd, Hughes TK, et al. Prevention of Tuberculosis in Macaques After Intravenous BCG Immunization. Nature (2020) 577(7788):95–102. doi: 10.1038/s41586-019-1817-8
19. Kaufmann SHE. Vaccine Development Against Tuberculosis Over the Last 140 Years: Failure as Part of Success. Front Microbiol (2021) 12:750124. doi: 10.3389/fmicb.2021.750124
20. Ura T, Okuda K, Shimada M. Developments in Viral Vector-Based Vaccines. Vaccines (Basel) (2014) 2(3):624–41. doi: 10.3390/vaccines2030624
21. Dalmia N, Ramsay AJ. Prime-Boost Approaches to Tuberculosis Vaccine Development. Expert Rev Vaccines (2012) 11(10):1221–33. doi: 10.1586/erv.12.94
22. Rollier CS, Reyes-Sandoval A, Cottingham MG, Ewer K, Hill AV. Viral Vectors as Vaccine Platforms: Deployment in Sight. Curr Opin Immunol (2011) 23(3):377–82. doi: 10.1016/j.coi.2011.03.006
23. Sasso E, D'Alise AM, Zambrano N, Scarselli E, Folgori A, Nicosia A. New Viral Vectors for Infectious Diseases and Cancer. Semin Immunol (2020) 50:101430. doi: 10.1016/j.smim.2020.101430
24. Ertl HC. Viral Vectors as Vaccine Carriers. Curr Opin Virol (2016) 21:1–8. doi: 10.1016/j.coviro.2016.06.001
25. Chen CY, Huang D, Wang RC, Shen L, Zeng G, Yao S, et al. A Critical Role for CD8 T Cells in a Nonhuman Primate Model of Tuberculosis. PloS Pathog (2009) 5(4):e1000392. doi: 10.1371/journal.ppat.1000392
26. Prezzemolo T, Guggino G, La Manna MP, Di Liberto D, Dieli F, Caccamo N. Functional Signatures of Human CD4 and CD8 T Cell Responses to Mycobacterium Tuberculosis. Front Immunol (2014) 5:180. doi: 10.3389/fimmu.2014.00180
27. Scriba TJ, Netea MG, Ginsberg AM. Key Recent Advances in TB Vaccine Development and Understanding of Protective Immune Responses Against Mycobacterium Tuberculosis. Semin Immunol (2020) 50:101431. doi: 10.1016/j.smim.2020.101431
28. Paoletti E. Poxvirus Recombinant Vaccines. Ann N Y Acad Sci (1990) 590:309–25. doi: 10.1111/j.1749-6632.1990.tb42239.x
29. Diven DG. An Overview of Poxviruses. J Am Acad Dermatol (2001) 44(1):1–16. doi: 10.1067/mjd.2001.109302
30. Copeman SM. Pathology of Vaccinia and Variola. Br Med J (1896) 1(1827):7–10. doi: 10.1136/bmj.1.1827.7
31. Moss B. Genetically Engineered Poxviruses for Recombinant Gene Expression, Vaccination, and Safety. Proc Natl Acad Sci U S A (1996) 93(21):11341–8. doi: 10.1073/pnas.93.21.11341
32. Kaynarcalidan O, Moreno Mascaraque S, Drexler I. Vaccinia Virus: From Crude Smallpox Vaccines to Elaborate Viral Vector Vaccine Design. Biomedicines (2021) 9(12):1780. doi: 10.3390/biomedicines9121780
33. Pantaleo G, Esteban M, Jacobs B, Tartaglia J. Poxvirus Vector-Based HIV Vaccines. Curr Opin HIV AIDS (2010) 5(5):391–6. doi: 10.1097/COH.0b013e32833d1e87
34. Gomez CE, Perdiguero B, Najera JL, Sorzano CO, Jimenez V, Gonzalez-Sanz R, et al. Removal of Vaccinia Virus Genes That Block Interferon Type I and II Pathways Improves Adaptive and Memory Responses of the HIV/AIDS Vaccine Candidate NYVAC-C in Mice. J Virol (2012) 86(9):5026–38. doi: 10.1128/JVI.06684-11
35. Garcia-Arriaza J, Esteban M. Enhancing Poxvirus Vectors Vaccine Immunogenicity. Hum Vaccin Immunother (2014) 10(8):2235–44. doi: 10.4161/hv.28974
36. Sutter G, Wyatt LS, Foley PL, Bennink JR, Moss B. A Recombinant Vector Derived From the Host Range-Restricted and Highly Attenuated MVA Strain of Vaccinia Virus Stimulates Protective Immunity in Mice to Influenza Virus. Vaccine (1994) 12(11):1032–40. doi: 10.1016/0264-410X(94)90341-7
37. Tartaglia J, Perkus ME, Taylor J, Norton EK, Audonnet JC, Cox WI, et al. NYVAC: A Highly Attenuated Strain of Vaccinia Virus. Virology (1992) 188(1):217–32. doi: 10.1016/0042-6822(92)90752-B
38. Paoletti E, Taylor J, Meignier B, Meric C, Tartaglia J. Highly Attenuated Poxvirus Vectors: NYVAC, ALVAC and TROVAC. Dev Biol Stand (1995) 84:159–63.
39. Hu Z, Wang J, Wan Y, Zhu L, Ren X, Qiu S, et al. Boosting Functional Avidity of CD8+ T Cells by Vaccinia Virus Vaccination Depends on Intrinsic T-Cell MyD88 Expression But Not the Inflammatory Milieu. J Virol (2014) 88(10):5356–68. doi: 10.1128/JVI.03664-13
40. Tameris MD, Hatherill M, Landry BS, Scriba TJ, Snowden MA, Lockhart S, et al. Safety and Efficacy of MVA85A, a New Tuberculosis Vaccine, in Infants Previously Vaccinated With BCG: A Randomised, Placebo-Controlled Phase 2b Trial. Lancet (2013) 381(9871):1021–8. doi: 10.1016/S0140-6736(13)60177-4
41. Ibanga HB, Brookes RH, Hill PC, Owiafe PK, Fletcher HA, Lienhardt C, et al. Early Clinical Trials With a New Tuberculosis Vaccine, MVA85A, in Tuberculosis-Endemic Countries: Issues in Study Design. Lancet Infect Dis (2006) 6(8):522–8. doi: 10.1016/S1473-3099(06)70552-7
42. Hu Z, Wong KW, Zhao HM, Wen HL, Ji P, Ma H, et al. Sendai Virus Mucosal Vaccination Establishes Lung-Resident Memory CD8 T Cell Immunity and Boosts BCG-Primed Protection Against TB in Mice. Mol Ther (2017) 25(5):1222–33. doi: 10.1016/j.ymthe.2017.02.018
43. McShane H, Brookes R, Gilbert SC, Hill AV. Enhanced Immunogenicity of CD4(+) T-Cell Responses and Protective Efficacy of a DNA-Modified Vaccinia Virus Ankara Prime-Boost Vaccination Regimen for Murine Tuberculosis. Infect Immun (2001) 69(2):681–6. doi: 10.1128/IAI.69.2.681-686.2001
44. McShane H, Behboudi S, Goonetilleke N, Brookes R, Hill AV. Protective Immunity Against Mycobacterium Tuberculosis Induced by Dendritic Cells Pulsed With Both CD8(+)- and CD4(+)-T-Cell Epitopes From Antigen 85A. Infect Immun (2002) 70(3):1623–6. doi: 10.1128/IAI.70.3.1623-1626.2002
45. Goonetilleke NP, McShane H, Hannan CM, Anderson RJ, Brookes RH, Hill AV. Enhanced Immunogenicity and Protective Efficacy Against Mycobacterium Tuberculosis of Bacille Calmette-Guerin Vaccine Using Mucosal Administration and Boosting With a Recombinant Modified Vaccinia Virus Ankara. J Immunol (2003) 171(3):1602–9. doi: 10.4049/jimmunol.171.3.1602
46. Williams A, Goonetilleke NP, McShane H, Clark SO, Hatch G, Gilbert SC, et al. Boosting With Poxviruses Enhances Mycobacterium Bovis BCG Efficacy Against Tuberculosis in Guinea Pigs. Infect Immun (2005) 73(6):3814–6. doi: 10.1128/IAI.73.6.3814-3816.2005
47. Vordermeier HM, Rhodes SG, Dean G, Goonetilleke N, Huygen K, Hill AV, et al. Cellular Immune Responses Induced in Cattle by Heterologous Prime-Boost Vaccination Using Recombinant Viruses and Bacille Calmette-Guerin. Immunology (2004) 112(3):461–70. doi: 10.1111/j.1365-2567.2004.01903.x
48. Verreck FA, Vervenne RA, Kondova I, van Kralingen KW, Remarque EJ, Braskamp G, et al. MVA.85A Boosting of BCG and an Attenuated, phoP Deficient M. Tuberculosis Vaccine Both Show Protective Efficacy Against Tuberculosis in Rhesus Macaques. PloS One (2009) 4(4):e5264. doi: 10.1371/journal.pone.0005264
49. McShane H, Pathan AA, Sander CR, Keating SM, Gilbert SC, Huygen K, et al. Recombinant Modified Vaccinia Virus Ankara Expressing Antigen 85A Boosts BCG-Primed and Naturally Acquired Antimycobacterial Immunity in Humans. Nat Med (2004) 10(11):1240–4. doi: 10.1038/nm1128
50. Scriba TJ, Tameris M, Mansoor N, Smit E, van der Merwe L, Isaacs F, et al. Modified Vaccinia Ankara-Expressing Ag85A, a Novel Tuberculosis Vaccine, is Safe in Adolescents and Children, and Induces Polyfunctional CD4+ T Cells. Eur J Immunol (2010) 40(1):279–90. doi: 10.1002/eji.200939754
51. Pathan AA, Minassian AM, Sander CR, Rowland R, Porter DW, Poulton ID, et al. Effect of Vaccine Dose on the Safety and Immunogenicity of a Candidate TB Vaccine, MVA85A, in BCG Vaccinated UK Adults. Vaccine (2012) 30(38):5616–24. doi: 10.1016/j.vaccine.2012.06.084
52. Meyer J, Harris SA, Satti I, Poulton ID, Poyntz HC, Tanner R, et al. Comparing the Safety and Immunogenicity of a Candidate TB Vaccine MVA85A Administered by Intramuscular and Intradermal Delivery. Vaccine (2013) 31(7):1026–33. doi: 10.1016/j.vaccine.2012.12.042
53. Tchilian EZ, Desel C, Forbes EK, Bandermann S, Sander CR, Hill AV, et al. Immunogenicity and Protective Efficacy of Prime-Boost Regimens With Recombinant (Delta)ureC Hly+ Mycobacterium Bovis BCG and Modified Vaccinia Virus Ankara Expressing M. Tuberculosis Antigen 85A Against Murine Tuberculosis. Infect Immun (2009) 77(2):622–31. doi: 10.1128/IAI.00685-08
54. Ndiaye BP, Thienemann F, Ota M, Landry BS, Camara M, Dieye S, et al. Safety, Immunogenicity, and Efficacy of the Candidate Tuberculosis Vaccine MVA85A in Healthy Adults Infected With HIV-1: A Randomised, Placebo-Controlled, Phase 2 Trial. Lancet Respir Med (2015) 3(3):190–200. doi: 10.1016/S2213-2600(15)00037-5
55. Satti I, Meyer J, Harris SA, Manjaly Thomas ZR, Griffiths K, Antrobus RD, et al. Safety and Immunogenicity of a Candidate Tuberculosis Vaccine MVA85A Delivered by Aerosol in BCG-Vaccinated Healthy Adults: A Phase 1, Double-Blind, Randomised Controlled Trial. Lancet Infect Dis (2014) 14(10):939–46. doi: 10.1016/S1473-3099(14)70845-X
56. Manjaly Thomas ZR, Satti I, Marshall JL, Harris SA, Lopez Ramon R, Hamidi A, et al. Alternate Aerosol and Systemic Immunisation With a Recombinant Viral Vector for Tuberculosis, MVA85A: A Phase I Randomised Controlled Trial. PloS Med (2019) 16(4):e1002790. doi: 10.1371/journal.pmed.1002790
57. Riste M, Marshall JL, Satti I, Harris SA, Wilkie M, Lopez Ramon R, et al. Phase I Trial Evaluating the Safety and Immunogenicity of Candidate TB Vaccine MVA85A, Delivered by Aerosol to Healthy M.tb-Infected Adults. Vaccines (Basel) (2021) 9(4):396. doi: 10.3390/vaccines9040396
58. Nemes E, Hesseling AC, Tameris M, Mauff K, Downing K, Mulenga H, et al. Safety and Immunogenicity of Newborn MVA85A Vaccination and Selective, Delayed Bacille Calmette-Guerin for Infants of Human Immunodeficiency Virus-Infected Mothers: A Phase 2 Randomized, Controlled Trial. Clin Infect Dis (2018) 66(4):554–63. doi: 10.1093/cid/cix834
59. Sheehan S, Harris SA, Satti I, Hokey DA, Dheenadhayalan V, Stockdale L, et al. A Phase I, Open-Label Trial, Evaluating the Safety and Immunogenicity of Candidate Tuberculosis Vaccines AERAS-402 and MVA85A, Administered by Prime-Boost Regime in BCG-Vaccinated Healthy Adults. PloS One (2015) 10(11):e0141687. doi: 10.1371/journal.pone.0141687
60. Wilkie M, Satti I, Minhinnick A, Harris S, Riste M, Ramon RL, et al. A Phase I Trial Evaluating the Safety and Immunogenicity of a Candidate Tuberculosis Vaccination Regimen, ChAdOx1 85A Prime - MVA85A Boost in Healthy UK Adults. Vaccine (2020) 38(4):779–89. doi: 10.1016/j.vaccine.2019.10.102
61. Rowland R, Pathan AA, Satti I, Poulton ID, Matsumiya MM, Whittaker M, et al. Safety and Immunogenicity of an FP9-Vectored Candidate Tuberculosis Vaccine (FP85A), Alone and With Candidate Vaccine MVA85A in BCG-Vaccinated Healthy Adults: A Phase I Clinical Trial. Hum Vaccin Immunother (2013) 9(1):50–62. doi: 10.4161/hv.22464
62. Minhinnick A, Satti I, Harris S, Wilkie M, Sheehan S, Stockdale L, et al. A First-in-Human Phase 1 Trial to Evaluate the Safety and Immunogenicity of the Candidate Tuberculosis Vaccine MVA85A-IMX313, Administered to BCG-Vaccinated Adults. Vaccine (2016) 34(11):1412–21. doi: 10.1016/j.vaccine.2016.01.062
63. Leung-Theung-Long S, Gouanvic M, Coupet CA, Ray A, Tupin E, Silvestre N, et al. A Novel MVA-Based Multiphasic Vaccine for Prevention or Treatment of Tuberculosis Induces Broad and Multifunctional Cell-Mediated Immunity in Mice and Primates. PloS One (2015) 10(11):e0143552. doi: 10.1371/journal.pone.0143552
64. Nangpal P, Bahal RK, Tyagi AK. Boosting With Recombinant MVA Expressing M. Tuberculosis Alpha-Crystallin Antigen Augments the Protection Imparted by BCG Against Tuberculosis in Guinea Pigs. Sci Rep (2017) 7(1):17286. doi: 10.1038/s41598-017-17587-5
65. Smaill F, Jeyanathan M, Smieja M, Medina MF, Thanthrige-Don N, Zganiacz A, et al. A Human Type 5 Adenovirus-Based Tuberculosis Vaccine Induces Robust T Cell Responses in Humans Despite Preexisting Anti-Adenovirus Immunity. Sci Transl Med (2013) 5(205):205ra134. doi: 10.1126/scitranslmed.3006843
66. Jeyanathan M, Fritz DK, Afkhami S, Aguirre E, Howie KJ, Zganiacz A, et al. Aerosol Delivery, But Not Intramuscular Injection, of Adenovirus-Vectored Tuberculosis Vaccine Induces Respiratory-Mucosal Immunity in Humans. JCI Insight (2022) 7(3):e155655. doi: 10.1172/jci.insight.155655
67. Abel B, Tameris M, Mansoor N, Gelderbloem S, Hughes J, Abrahams D, et al. The Novel Tuberculosis Vaccine, AERAS-402, Induces Robust and Polyfunctional CD4+ and CD8+ T Cells in Adults. Am J Respir Crit Care Med (2010) 181(12):1407–17. doi: 10.1164/rccm.200910-1484OC
68. Kagina BM, Tameris MD, Geldenhuys H, Hatherill M, Abel B, Hussey GD, et al. The Novel Tuberculosis Vaccine, AERAS-402, is Safe in Healthy Infants Previously Vaccinated With BCG, and Induces Dose-Dependent CD4 and CD8T Cell Responses. Vaccine (2014) 32(45):5908–17. doi: 10.1016/j.vaccine.2014.09.001
69. Hoft DF, Blazevic A, Stanley J, Landry B, Sizemore D, Kpamegan E, et al. A Recombinant Adenovirus Expressing Immunodominant TB Antigens can Significantly Enhance BCG-Induced Human Immunity. Vaccine (2012) 30(12):2098–108. doi: 10.1016/j.vaccine.2012.01.048
70. van Zyl-Smit RN, Esmail A, Bateman ME, Dawson R, Goldin J, van Rikxoort E, et al. Safety and Immunogenicity of Adenovirus 35 Tuberculosis Vaccine Candidate in Adults With Active or Previous Tuberculosis. A Randomized Trial. Am J Respir Crit Care Med (2017) 195(9):1171–80. doi: 10.1164/rccm.201603-0654OC
71. Walsh DS, Owira V, Polhemus M, Otieno L, Andagalu B, Ogutu B, et al. Adenovirus Type 35-Vectored Tuberculosis Vaccine has an Acceptable Safety and Tolerability Profile in Healthy, BCG-Vaccinated, QuantiFERON((R))-TB Gold (+) Kenyan Adults Without Evidence of Tuberculosis. Vaccine (2016) 34(21):2430–6. doi: 10.1016/j.vaccine.2016.03.069
72. Churchyard GJ, Snowden MA, Hokey D, Dheenadhayalan V, McClain JB, Douoguih M, et al. The Safety and Immunogenicity of an Adenovirus Type 35-Vectored TB Vaccine in HIV-Infected, BCG-Vaccinated Adults With CD4(+) T Cell Counts >350 Cells/Mm(3). Vaccine (2015) 33(15):1890–6. doi: 10.1016/j.vaccine.2015.02.004
73. Sivakumaran D, Blatner G, Bakken R, Hokey D, Ritz C, Jenum S, et al. A 2-Dose AERAS-402 Regimen Boosts CD8(+) Polyfunctionality in HIV-Negative, BCG-Vaccinated Recipients. Front Immunol (2021) 12:673532. doi: 10.3389/fimmu.2021.673532
74. Wang J, Thorson L, Stokes RW, Santosuosso M, Huygen K, Zganiacz A, et al. Single Mucosal, But Not Parenteral, Immunization With Recombinant Adenoviral-Based Vaccine Provides Potent Protection From Pulmonary Tuberculosis. J Immunol (2004) 173(10):6357–65. doi: 10.4049/jimmunol.173.10.6357
75. Santosuosso M, Zhang X, McCormick S, Wang J, Hitt M, Xing Z. Mechanisms of Mucosal and Parenteral Tuberculosis Vaccinations: Adenoviral-Based Mucosal Immunization Preferentially Elicits Sustained Accumulation of Immune Protective CD4 and CD8 T Cells Within the Airway Lumen. J Immunol (2005) 174(12):7986–94. doi: 10.4049/jimmunol.174.12.7986
76. Santosuosso M, McCormick S, Zhang X, Zganiacz A, Xing Z. Intranasal Boosting With an Adenovirus-Vectored Vaccine Markedly Enhances Protection by Parenteral Mycobacterium Bovis BCG Immunization Against Pulmonary Tuberculosis. Infect Immun (2006) 74(8):4634–43. doi: 10.1128/IAI.00517-06
77. Santosuosso M, McCormick S, Roediger E, Zhang X, Zganiacz A, Lichty BD, et al. Mucosal Luminal Manipulation of T Cell Geography Switches on Protective Efficacy by Otherwise Ineffective Parenteral Genetic Immunization. J Immunol (2007) 178(4):2387–95. doi: 10.4049/jimmunol.178.4.2387
78. Xing Z, McFarland CT, Sallenave JM, Izzo A, Wang J, McMurray DN. Intranasal Mucosal Boosting With an Adenovirus-Vectored Vaccine Markedly Enhances the Protection of BCG-Primed Guinea Pigs Against Pulmonary Tuberculosis. PloS One (2009) 4(6):e5856. doi: 10.1371/journal.pone.0005856
79. Vordermeier HM, Villarreal-Ramos B, Cockle PJ, McAulay M, Rhodes SG, Thacker T, et al. Viral Booster Vaccines Improve Mycobacterium Bovis BCG-Induced Protection Against Bovine Tuberculosis. Infect Immun (2009) 77(8):3364–73. doi: 10.1128/IAI.00287-09
80. Perez de Val B, Villarreal-Ramos B, Nofrarias M, Lopez-Soria S, Romera N, Singh M, et al. Goats Primed With Mycobacterium Bovis BCG and Boosted With a Recombinant Adenovirus Expressing Ag85A Show Enhanced Protection Against Tuberculosis. Clin Vaccine Immunol (2012) 19(9):1339–47. doi: 10.1128/CVI.00275-12
81. Jeyanathan M, Shao Z, Yu X, Harkness R, Jiang R, Li J, et al. AdHu5Ag85A Respiratory Mucosal Boost Immunization Enhances Protection Against Pulmonary Tuberculosis in BCG-Primed Non-Human Primates. PloS One (2015) 10(8):e0135009. doi: 10.1371/journal.pone.0135009
82. Sekaly RP. The Failed HIV Merck Vaccine Study: A Step Back or a Launching Point for Future Vaccine Development? J Exp Med (2008) 205(1):7–12. doi: 10.1084/jem.20072681
83. Quinn KM, Da Costa A, Yamamoto A, Berry D, Lindsay RW, Darrah PA, et al. Comparative Analysis of the Magnitude, Quality, Phenotype, and Protective Capacity of Simian Immunodeficiency Virus Gag-Specific CD8+ T Cells Following Human-, Simian-, and Chimpanzee-Derived Recombinant Adenoviral Vector Immunization. J Immunol (2013) 190(6):2720–35. doi: 10.4049/jimmunol.1202861
84. Quinn KM, Zak DE, Costa A, Yamamoto A, Kastenmuller K, Hill BJ, et al. Antigen Expression Determines Adenoviral Vaccine Potency Independent of IFN and STING Signaling. J Clin Invest (2015) 125(3):1129–46. doi: 10.1172/JCI78280
85. Radosevic K, Wieland CW, Rodriguez A, Weverling GJ, Mintardjo R, Gillissen G, et al. Protective Immune Responses to a Recombinant Adenovirus Type 35 Tuberculosis Vaccine in Two Mouse Strains: CD4 and CD8 T-Cell Epitope Mapping and Role of Gamma Interferon. Infect Immun (2007) 75(8):4105–15. doi: 10.1128/IAI.00004-07
86. Darrah PA, Bolton DL, Lackner AA, Kaushal D, Aye PP, Mehra S, et al. Aerosol Vaccination With AERAS-402 Elicits Robust Cellular Immune Responses in the Lungs of Rhesus Macaques But Fails to Protect Against High-Dose Mycobacterium Tuberculosis Challenge. J Immunol (2014) 193(4):1799–811. doi: 10.4049/jimmunol.1400676
87. Magalhaes I, Sizemore DR, Ahmed RK, Mueller S, Wehlin L, Scanga C, et al. rBCG Induces Strong Antigen-Specific T Cell Responses in Rhesus Macaques in a Prime-Boost Setting With an Adenovirus 35 Tuberculosis Vaccine Vector. PloS One (2008) 3(11):e3790. doi: 10.1371/journal.pone.0003790
88. Hokey DA, Wachholder R, Darrah PA, Bolton DL, Barouch DH, Hill K, et al. A Nonhuman Primate Toxicology and Immunogenicity Study Evaluating Aerosol Delivery of AERAS-402/Ad35 Vaccine: Evidence for Transient T Cell Responses in Peripheral Blood and Robust Sustained Responses in the Lungs. Hum Vaccin Immunother (2014) 10(8):2199–210. doi: 10.4161/hv.29108
89. Stylianou E, Griffiths KL, Poyntz HC, Harrington-Kandt R, Dicks MD, Stockdale L, et al. Improvement of BCG Protective Efficacy With a Novel Chimpanzee Adenovirus and a Modified Vaccinia Ankara Virus Both Expressing Ag85A. Vaccine (2015) 33(48):6800–8. doi: 10.1016/j.vaccine.2015.10.017
90. Pinpathomrat N, Bull N, Pasricha J, Harrington-Kandt R, McShane H, Stylianou E. Using an Effective TB Vaccination Regimen to Identify Immune Responses Associated With Protection in the Murine Model. Vaccine (2021) 39(9):1452–62. doi: 10.1016/j.vaccine.2021.01.034
91. Stylianou E, Harrington-Kandt R, Beglov J, Bull N, Pinpathomrat N, Swarbrick GM, et al. Identification and Evaluation of Novel Protective Antigens for the Development of a Candidate Tuberculosis Subunit Vaccine. Infect Immun (2018) 86(7):e00014-18. doi: 10.1128/IAI.00014-18
92. Jeyanathan M, Thanthrige-Don N, Afkhami S, Lai R, Damjanovic D, Zganiacz A, et al. Novel Chimpanzee Adenovirus-Vectored Respiratory Mucosal Tuberculosis Vaccine: Overcoming Local Anti-Human Adenovirus Immunity for Potent TB Protection. Mucosal Immunol (2015) 8(6):1373–87. doi: 10.1038/mi.2015.29
93. Afkhami S, Lai R, D'Agostino MR, Vaseghi-Shanjani M, Zganiacz A, Yao Y, et al. Single-Dose Mucosal Immunotherapy With Chimpanzee Adenovirus-Based Vaccine Accelerates Tuberculosis Disease Control and Limits Its Rebound After Antibiotic Cessation. J Infect Dis (2019) 220(8):1355–66. doi: 10.1093/infdis/jiz306
94. Stukova MA, Sereinig S, Zabolotnyh NV, Ferko B, Kittel C, Romanova J, et al. Vaccine Potential of Influenza Vectors Expressing Mycobacterium Tuberculosis ESAT-6 Protein. Tuberculosis (Edinb) (2006) 86(3-4):236–46. doi: 10.1016/j.tube.2006.01.010
95. Sereinig S, Stukova M, Zabolotnyh N, Ferko B, Kittel C, Romanova J, et al. Influenza Virus NS Vectors Expressing the Mycobacterium Tuberculosis ESAT-6 Protein Induce CD4+ Th1 Immune Response and Protect Animals Against Tuberculosis Challenge. Clin Vaccine Immunol (2006) 13(8):898–904. doi: 10.1128/CVI.00056-06
96. WHO. WHO Global TB Report. (2017). https://apps.who.int/iris/bitstream/handle/10665/259366/9789241565516-eng.pdf?sequence=1
97. Florido M, Muflihah H, Lin LCW, Xia Y, Sierro F, Palendira M, et al. Pulmonary Immunization With a Recombinant Influenza A Virus Vaccine Induces Lung-Resident CD4(+) Memory T Cells That are Associated With Protection Against Tuberculosis. Mucosal Immunol (2018) 11(6):1743–52. doi: 10.1038/s41385-018-0065-9
98. Beverley PC, Ruzsics Z, Hey A, Hutchings C, Boos S, Bolinger B, et al. A Novel Murine Cytomegalovirus Vaccine Vector Protects Against Mycobacterium Tuberculosis. J Immunol (2014) 193(5):2306–16. doi: 10.4049/jimmunol.1302523
99. Hansen SG, Zak DE, Xu G, Ford JC, Marshall EE, Malouli D, et al. Prevention of Tuberculosis in Rhesus Macaques by a Cytomegalovirus-Based Vaccine. Nat Med (2018) 24(2):130–43. doi: 10.1038/nm.4473
100. Abad-Fernandez M, Goonetilleke N. Human Cytomegalovirus-Vectored Vaccines Against HIV. Curr Opin HIV AIDS (2019) 14(2):137–42. doi: 10.1097/COH.0000000000000524
101. Gugliesi F, Coscia A, Griffante G, Galitska G, Pasquero S, Albano C, et al. Where do We Stand After Decades of Studying Human Cytomegalovirus? Microorganisms (2020) 8(5):685. doi: 10.3390/microorganisms8050685
102. Stockdale L, Nash S, Farmer R, Raynes J, Mallikaarjun S, Newton R, et al. Cytomegalovirus Antibody Responses Associated With Increased Risk of Tuberculosis Disease in Ugandan Adults. J Infect Dis (2020) 221(7):1127–34. doi: 10.1093/infdis/jiz581
103. van der Heijden YF, Zhang B, Chougnet CA, Huaman MA. Cytomegalovirus Infection Is Associated With Increased Prevalence of Latent Tuberculosis Infection. Open Forum Infect Dis (2021) 8(11):ofab539. doi: 10.1093/ofid/ofab539
104. Hu Z, Gu L, Li CL, Shu T, Lowrie DB, Fan XY. The Profile of T Cell Responses in Bacille Calmette-Guerin-Primed Mice Boosted by a Novel Sendai Virus Vectored Anti-Tuberculosis Vaccine. Front Immunol (2018) 9:1796. doi: 10.3389/fimmu.2018.01796
105. Hu Z, Jiang W, Gu L, Qiao D, Shu T, Lowrie DB, et al. Heterologous Prime-Boost Vaccination Against Tuberculosis With Recombinant Sendai Virus and DNA Vaccines. J Mol Med (Berl) (2019) 97(12):1685–94. doi: 10.1007/s00109-019-01844-3
106. Hashimoto D, Nagata T, Uchijima M, Seto S, Suda T, Chida K, et al. Intratracheal Administration of Third-Generation Lentivirus Vector Encoding MPT51 From Mycobacterium Tuberculosis Induces Specific CD8+ T-Cell Responses in the Lung. Vaccine (2008) 26(40):5095–100. doi: 10.1016/j.vaccine.2008.03.101
107. Xu Y, Yang E, Wang J, Li R, Li G, Liu G, et al. Prime-Boost Bacillus Calmette-Guerin Vaccination With Lentivirus-Vectored and DNA-Based Vaccines Expressing Antigens Ag85B and Rv3425 Improves Protective Efficacy Against Mycobacterium Tuberculosis in Mice. Immunology (2014) 143(2):277–86. doi: 10.1111/imm.12308
108. Britton G, MacDonald DC, Brown JS, Collins MK, Goodman AL. Using a Prime and Pull Approach, Lentivector Vaccines Expressing Ag85A Induce Immunogenicity But Fail to Induce Protection Against Mycobacterium Bovis Bacillus Calmette-Guerin Challenge in Mice. Immunology (2015) 146(2):264–70. doi: 10.1111/imm.12498
109. Yang E, Wang F, Xu Y, Wang H, Hu Y, Shen H, et al. A Lentiviral Vector-Based Therapeutic Vaccine Encoding Ag85B-Rv3425 Potently Increases Resistance to Acute Tuberculosis Infection in Mice. Acta Biochim Biophys Sin (Shanghai) (2015) 47(8):588–96. doi: 10.1093/abbs/gmv059
110. Shakouri M, Moazzeni SM, Ghanei M, Arashkia A, Etemadzadeh MH, Azadmanesh K. A Novel Dendritic Cell-Targeted Lentiviral Vector, Encoding Ag85A-ESAT6 Fusion Gene of Mycobacterium Tuberculosis, Could Elicit Potent Cell-Mediated Immune Responses in Mice. Mol Immunol (2016) 75:101–11. doi: 10.1016/j.molimm.2016.04.014
111. Kalos M, Levine BL, Porter DL, Katz S, Grupp SA, Bagg A, et al. T Cells With Chimeric Antigen Receptors Have Potent Antitumor Effects and can Establish Memory in Patients With Advanced Leukemia. Sci Transl Med (2011) 3(95):95ra73. doi: 10.1126/scitranslmed.3002842
112. Gutierrez-Guerrero A, Cosset FL, Verhoeyen E. Lentiviral Vector Pseudotypes: Precious Tools to Improve Gene Modification of Hematopoietic Cells for Research and Gene Therapy. Viruses (2020) 12(9):1016. doi: 10.3390/v12091016
113. Roediger EK, Kugathasan K, Zhang X, Lichty BD, Xing Z. Heterologous Boosting of Recombinant Adenoviral Prime Immunization With a Novel Vesicular Stomatitis Virus-Vectored Tuberculosis Vaccine. Mol Ther (2008) 16(6):1161–9. doi: 10.1038/mt.2008.59
114. Jeyanathan M, Damjanovic D, Shaler CR, Lai R, Wortzman M, Yin C, et al. Differentially Imprinted Innate Immunity by Mucosal Boost Vaccination Determines Antituberculosis Immune Protective Outcomes, Independent of T-Cell Immunity. Mucosal Immunol (2013) 6(3):612–25. doi: 10.1038/mi.2012.103
115. Zhang M, Dong C, Xiong S. Vesicular Stomatitis Virus-Vectored Multi-Antigen Tuberculosis Vaccine Limits Bacterial Proliferation in Mice Following a Single Intranasal Dose. Front Cell Infect Microbiol (2017) 7:34. doi: 10.3389/fcimb.2017.00034
116. Zhang M, Dong C, Xiong S. Heterologous Boosting With Recombinant VSV-846 in BCG-Primed Mice Confers Improved Protection Against Mycobacterium Infection. Hum Vaccin Immunother (2017) 13(4):816–22. doi: 10.1080/21645515.2016.1261229
117. Watanabe K, Matsubara A, Kawano M, Mizuno S, Okamura T, Tsujimura Y, et al. Recombinant Ag85B Vaccine by Taking Advantage of Characteristics of Human Parainfluenza Type 2 Virus Vector Showed Mycobacteria-Specific Immune Responses by Intranasal Immunization. Vaccine (2014) 32(15):1727–35. doi: 10.1016/j.vaccine.2013.11.108
118. Nagatake T, Suzuki H, Hirata SI, Matsumoto N, Wada Y, Morimoto S, et al. Immunological Association of Inducible Bronchus-Associated Lymphoid Tissue Organogenesis in Ag85B-Rhpiv2 Vaccine-Induced Anti-Tuberculosis Mucosal Immune Responses in Mice. Int Immunol (2018) 30(10):471–81. doi: 10.1093/intimm/dxy046
119. Chen Z, Gupta T, Xu P, Phan S, Pickar A, Yau W, et al. Efficacy of Parainfluenza Virus 5 (PIV5)-Based Tuberculosis Vaccines in Mice. Vaccine (2015) 33(51):7217–24. doi: 10.1016/j.vaccine.2015.10.124
120. Vierboom MPM, Chenine AL, Darrah PA, Vervenne RAW, Boot C, Hofman SO, et al. Evaluation of Heterologous Prime-Boost Vaccination Strategies Using Chimpanzee Adenovirus and Modified Vaccinia Virus for TB Subunit Vaccination in Rhesus Macaques. NPJ Vaccines (2020) 5(1):39. doi: 10.1038/s41541-020-0189-2
121. Bettencourt P, Muller J, Nicastri A, Cantillon D, Madhavan M, Charles PD, et al. Identification of Antigens Presented by MHC for Vaccines Against Tuberculosis. NPJ Vaccines (2020) 5(1):2. doi: 10.1038/s41541-019-0148-y
122. Mosley JF 2nd, Smith LL, Brantley P, Locke D, Como M. Vaxchora: The First FDA-Approved Cholera Vaccination in the United States. P T (2017) 42(10):638–40.
123. Minassian AM, Satti I, Poulton ID, Meyer J, Hill AV, McShane H. A Human Challenge Model for Mycobacterium Tuberculosis Using Mycobacterium Bovis Bacille Calmette-Guerin. J Infect Dis (2012) 205(7):1035–42. doi: 10.1093/infdis/jis012
124. Harris SA, Meyer J, Satti I, Marsay L, Poulton ID, Tanner R, et al. Evaluation of a Human BCG Challenge Model to Assess Antimycobacterial Immunity Induced by BCG and a Candidate Tuberculosis Vaccine, MVA85A, Alone and in Combination. J Infect Dis (2014) 209(8):1259–68. doi: 10.1093/infdis/jit647
125. Kleinwaks G, Schmit V, Morrison J. Considering Human Challenge Trials for Tuberculosis Vaccine Development. Vaccine (2022) 40(2):173–4. doi: 10.1016/j.vaccine.2021.11.024
126. McShane H, Williams A. A Review of Preclinical Animal Models Utilised for TB Vaccine Evaluation in the Context of Recent Human Efficacy Data. Tuberculosis (Edinb) (2014) 94(2):105–10. doi: 10.1016/j.tube.2013.11.003
127. Dijkman K, Sombroek CC, Vervenne RAW, Hofman SO, Boot C, Remarque EJ, et al. Prevention of Tuberculosis Infection and Disease by Local BCG in Repeatedly Exposed Rhesus Macaques. Nat Med (2019) 25(2):255–62. doi: 10.1038/s41591-018-0319-9
128. Plumlee CR, Duffy FJ, Gern BH, Delahaye JL, Cohen SB, Stoltzfus CR, et al. Ultra-Low Dose Aerosol Infection of Mice With Mycobacterium Tuberculosis More Closely Models Human Tuberculosis. Cell Host Microbe (2021) 29(1):68–82.e5. doi: 10.1016/j.chom.2020.10.003
129. Park CO, Kupper TS. The Emerging Role of Resident Memory T Cells in Protective Immunity and Inflammatory Disease. Nat Med (2015) 21(7):688–97. doi: 10.1038/nm.3883
130. Parga-Vidal L, van Aalderen MC, Stark R, van Gisbergen K. Tissue-Resident Memory T Cells in the Urogenital Tract. Nat Rev Nephrol (2022) 18(4):209–23. doi: 10.1038/s41581-021-00525-0
131. Sollid LM. Gut Tissue-Resident Memory T Cells in Coeliac Disease. Scand J Immunol (2022) 95(1):e13120. doi: 10.1111/sji.13120
132. Lange J, Rivera-Ballesteros O, Buggert M. Human Mucosal Tissue-Resident Memory T Cells in Health and Disease. Mucosal Immunol (2021) 15(3):389–97. doi: 10.1038/s41385-021-00467-7
133. Ryan GE, Harris JE, Richmond JM. Resident Memory T Cells in Autoimmune Skin Diseases. Front Immunol (2021) 12:652191. doi: 10.3389/fimmu.2021.652191
134. Mueller SN, Mackay LK. Tissue-Resident Memory T Cells: Local Specialists in Immune Defence. Nat Rev Immunol (2016) 16(2):79–89. doi: 10.1038/nri.2015.3
135. Sakai S, Kauffman KD, Schenkel JM, McBerry CC, Mayer-Barber KD, Masopust D, et al. Cutting Edge: Control of Mycobacterium Tuberculosis Infection by a Subset of Lung Parenchyma-Homing CD4 T Cells. J Immunol (2014) 192(7):2965–9. doi: 10.4049/jimmunol.1400019
136. Torrado E, Fountain JJ, Liao M, Tighe M, Reiley WW, Lai RP, et al. Interleukin 27R Regulates CD4+ T Cell Phenotype and Impacts Protective Immunity During Mycobacterium Tuberculosis Infection. J Exp Med (2015) 212(9):1449–63. doi: 10.1084/jem.20141520
137. Sakai S, Kauffman KD, Sallin MA, Sharpe AH, Young HA, Ganusov VV, et al. CD4 T Cell-Derived IFN-Gamma Plays a Minimal Role in Control of Pulmonary Mycobacterium Tuberculosis Infection and Must Be Actively Repressed by PD-1 to Prevent Lethal Disease. PloS Pathog (2016) 12(5):e1005667. doi: 10.1371/journal.ppat.1005667
138. Hu Z, Zhao HM, Li CL, Liu XH, Barkan D, Lowrie DB, et al. The Role of KLRG1 in Human CD4+ T-Cell Immunity Against Tuberculosis. J Infect Dis (2018) 217(9):1491–503. doi: 10.1093/infdis/jiy046
139. Perdomo C, Zedler U, Kuhl AA, Lozza L, Saikali P, Sander LE, et al. Mucosal BCG Vaccination Induces Protective Lung-Resident Memory T Cell Populations Against Tuberculosis. mBio (2016) 7(6):e01686-16. doi: 10.1128/mBio.01686-16
140. Jeyanathan M, Afkhami S, Khera A, Mandur T, Damjanovic D, Yao Y, et al. CXCR3 Signaling Is Required for Restricted Homing of Parenteral Tuberculosis Vaccine-Induced T Cells to Both the Lung Parenchyma and Airway. J Immunol (2017) 199(7):2555–69. doi: 10.4049/jimmunol.1700382
141. Carpenter SM, Yang JD, Lee J, Barreira-Silva P, Behar SM. Vaccine-Elicited Memory CD4+ T Cell Expansion is Impaired in the Lungs During Tuberculosis. PloS Pathog (2017) 13(11):e1006704. doi: 10.1371/journal.ppat.1006704
142. Woodworth JS, Cohen SB, Moguche AO, Plumlee CR, Agger EM, Urdahl KB, et al. Subunit Vaccine H56/CAF01 Induces a Population of Circulating CD4 T Cells That Traffic Into the Mycobacterium Tuberculosis-Infected Lung. Mucosal Immunol (2017) 10(2):555–64. doi: 10.1038/mi.2016.70
143. Brinkmann V, Davis MD, Heise CE, Albert R, Cottens S, Hof R, et al. The Immune Modulator FTY720 Targets Sphingosine 1-Phosphate Receptors. J Biol Chem (2002) 277(24):21453–7. doi: 10.1074/jbc.C200176200
144. Connor LM, Harvie MC, Rich FJ, Quinn KM, Brinkmann V, Le Gros G, et al. A Key Role for Lung-Resident Memory Lymphocytes in Protective Immune Responses After BCG Vaccination. Eur J Immunol (2010) 40(9):2482–92. doi: 10.1002/eji.200940279
145. Netea MG, Quintin J, van der Meer JW. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe (2011) 9(5):355–61. doi: 10.1016/j.chom.2011.04.006
146. Netea MG, van der Meer JW. Trained Immunity: An Ancient Way of Remembering. Cell Host Microbe (2017) 21(3):297–300. doi: 10.1016/j.chom.2017.02.003
147. Netea MG, Dominguez-Andres J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, et al. Defining Trained Immunity and its Role in Health and Disease. Nat Rev Immunol (2020) 20(6):375–88. doi: 10.1038/s41577-020-0285-6
148. Hu Z, Lu SH, Lowrie DB, Fan XY. Trained Immunity: A Yin-Yang Balance. MedComm (2020) (2022) 3(1):e121. doi: 10.1002/mco2.121
149. Giamarellos-Bourboulis EJ, Tsilika M, Moorlag S, Antonakos N, Kotsaki A, Dominguez-Andres J, et al. Activate: Randomized Clinical Trial of BCG Vaccination Against Infection in the Elderly. Cell (2020) 183(2):315–23.e9. doi: 10.1016/j.cell.2020.08.051
150. Prentice S, Nassanga B, Webb EL, Akello F, Kiwudhu F, Akurut H, et al. BCG-Induced non-Specific Effects on Heterologous Infectious Disease in Ugandan Neonates: An Investigator-Blind Randomised Controlled Trial. Lancet Infect Dis (2021) 21(7):993–1003. doi: 10.1016/S1473-3099(20)30653-8
151. Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Ifrim DC, Saeed S, et al. Bacille Calmette-Guerin Induces NOD2-Dependent Nonspecific Protection From Reinfection via Epigenetic Reprogramming of Monocytes. Proc Natl Acad Sci U S A (2012) 109(43):17537–42. doi: 10.1073/pnas.1202870109
152. Verma D, Parasa VR, Raffetseder J, Martis M, Mehta RB, Netea M, et al. Anti-Mycobacterial Activity Correlates With Altered DNA Methylation Pattern in Immune Cells From BCG-Vaccinated Subjects. Sci Rep (2017) 7(1):12305. doi: 10.1038/s41598-017-12110-2
153. Vierboom MPM, Dijkman K, Sombroek CC, Hofman SO, Boot C, Vervenne RAW, et al. Stronger Induction of Trained Immunity by Mucosal BCG or MTBVAC Vaccination Compared to Standard Intradermal Vaccination. Cell Rep Med (2021) 2(1):100185. doi: 10.1016/j.xcrm.2020.100185
154. Moorlag S, Khan N, Novakovic B, Kaufmann E, Jansen T, van Crevel R, et al. Beta-Glucan Induces Protective Trained Immunity Against Mycobacterium Tuberculosis Infection: A Key Role for IL-1. Cell Rep (2020) 31(7):107634. doi: 10.1016/j.celrep.2020.107634
155. Khan N, Downey J, Sanz J, Kaufmann E, Blankenhaus B, Pacis A, et al. M. Tuberculosis Reprograms Hematopoietic Stem Cells to Limit Myelopoiesis and Impair Trained Immunity. Cell (2020) 183(3):752–70.e22. doi: 10.1016/j.cell.2020.09.062
156. Yao Y, Jeyanathan M, Haddadi S, Barra NG, Vaseghi-Shanjani M, Damjanovic D, et al. Induction of Autonomous Memory Alveolar Macrophages Requires T Cell Help and Is Critical to Trained Immunity. Cell (2018) 175(6):1634–50.e17. doi: 10.1016/j.cell.2018.09.042
157. D'Agostino MR, Lai R, Afkhami S, Khera A, Yao Y, Vaseghi-Shanjani M, et al. Airway Macrophages Mediate Mucosal Vaccine-Induced Trained Innate Immunity Against Mycobacterium Tuberculosis in Early Stages of Infection. J Immunol (2020) 205(10):2750–62. doi: 10.4049/jimmunol.2000532
158. Afkhami S, D'Agostino MR, Zhang A, Stacey HD, Marzok A, Kang A, et al. Respiratory Mucosal Delivery of Next-Generation COVID-19 Vaccine Provides Robust Protection Against Both Ancestral and Variant Strains of SARS-CoV-2. Cell (2022) 185(5):896–915.e19. doi: 10.1101/2021.07.16.452721
159. Goletti D, Lee MR, Wang JY, Walter N, Ottenhoff THM. Update on Tuberculosis Biomarkers: From Correlates of Risk, to Correlates of Active Disease and of Cure From Disease. Respirology (2018) 23(5):455–66. doi: 10.1111/resp.13272
160. Simmons JD, Stein CM, Seshadri C, Campo M, Alter G, Fortune S, et al. Immunological Mechanisms of Human Resistance to Persistent Mycobacterium Tuberculosis Infection. Nat Rev Immunol (2018) 18(9):575–89. doi: 10.1038/s41577-018-0025-3
161. Lu LL, Smith MT, Yu KKQ, Luedemann C, Suscovich TJ, Grace PS, et al. IFN-Gamma-Independent Immune Markers of Mycobacterium Tuberculosis Exposure. Nat Med (2019) 25(6):977–87. doi: 10.1038/s41591-019-0441-3
162. Chen ZY, Wang L, Gu L, Qu R, Lowrie DB, Hu Z, et al. Decreased Expression of CD69 on T Cells in Tuberculosis Infection Resisters. Front Microbiol (2020) 11:1901. doi: 10.3389/fmicb.2020.01901
163. Tait DR, Hatherill M, van der Meeren O, Ginsberg AM, Van Brakel E, Salaun B, et al. Final Analysis of a Trial of M72/AS01E Vaccine to Prevent Tuberculosis. N Engl J Med (2019) 381(25):2429–39. doi: 10.1056/NEJMoa1909953
164. COVID-19 Vaccine Tracker and Landscape (2022). Available at: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines.
Keywords: tuberculosis, vaccine, viral vector, mucosal immunity, trained immunity
Citation: Hu Z, Lu S-H, Lowrie DB and Fan X-Y (2022) Research Advances for Virus-vectored Tuberculosis Vaccines and Latest Findings on Tuberculosis Vaccine Development. Front. Immunol. 13:895020. doi: 10.3389/fimmu.2022.895020
Received: 12 March 2022; Accepted: 27 May 2022;
Published: 23 June 2022.
Edited by:
Hao Li, China Agricultural University, ChinaReviewed by:
Joanna Kirman, University of Otago, New ZealandRachel Tanner, University of Oxford, United Kingdom
Copyright © 2022 Hu, Lu, Lowrie and Fan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhidong Hu, aHV6aGlkb25nQGZ1ZGFuLmVkdS5jbg==; Xiao-Yong Fan, eHlmYW4wMDhAZnVkYW4uZWR1LmNu