
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 22 June 2022
Sec. Cancer Immunity and Immunotherapy
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.892750
This article is part of the Research Topic Biomarkers and Immunotherapy of Hepatic-Biliary-Pancreatic Cancers View all 53 articles
 Hongjun Yu1,2†
Hongjun Yu1,2† Chaoqun Wang1,2†Shanjia Ke1,2†Miaoyu Bai1,2†
Chaoqun Wang1,2†Shanjia Ke1,2†Miaoyu Bai1,2† Yanan Xu1,2Shounan Lu1,2Zhigang Feng1,2,3
Yanan Xu1,2Shounan Lu1,2Zhigang Feng1,2,3 Baolin Qian1,2Yue Xu4
Baolin Qian1,2Yue Xu4 Menghua Zhou1,2Zihao Li1,2Bing Yin1,2
Menghua Zhou1,2Zihao Li1,2Bing Yin1,2 Xinglong Li1,2
Xinglong Li1,2 Yongliang Hua2,5Yongzhi Zhou1Shangha Pan2Yao Fu6*
Yongliang Hua2,5Yongzhi Zhou1Shangha Pan2Yao Fu6* Yong Ma1,2*
Yong Ma1,2*Background: Complement factor H-related 4 (CFHR4) is a protein-coding gene that plays an essential role in multiple diseases. However, the prognostic value of CFHR4 in hepatocellular carcinoma (HCC) is unknown.
Methods: Using multiple databases, we investigated CFHR4 expression levels in HCC and multiple cancers. The relationship between CFHR4 expression levels and clinicopathological variables was further analyzed. Various potential biological functions and regulatory pathways of CFHR4 in HCC were identified by performing a Gene Ontology (GO) analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis and Gene Set Enrichment Analysis (GSEA). Single-sample gene set enrichment analysis (ssGSEA) was performed to confirm the correlation between CFHR4 expression and immune cell infiltration. The correlations between CFHR4 expression levels in HCC and N6-methyladenosine (m6A) modifications and the competing endogenous RNA (ceRNA) regulatory networks were confirmed in TCGA cohort.
Results: CFHR4 expression levels were significantly decreased in HCC tissues. Low CFHR4 expression in HCC tissues was significantly correlated with the patients’ sex, race, age, TNM stage, pathological stage, tumor status, residual tumor, histologic grade and alpha fetal protein (AFP) level. GO and KEGG analyses revealed that differentially expressed genes related to CFHR4 may be involved in the synaptic membrane, transmembrane transporter complex, gated channel activity, chemical carcinogenesis, retinol metabolism, calcium signaling pathway, PPAR signaling pathway, insulin and gastric acid secretion. GSEA revealed that the FCGR-activated reaction, PLK1 pathway, ATR pathway, MCM pathway, cascade reactions of PI3K and FGFR1, reactant-mediated MAPK activation and FOXM1 pathway were significantly enriched in HCC with low CFHR4 expression. Moreover, CFHR4 expression was inversely correlated the levels of infiltrating Th2 cells, NK CD56bright cells and Tfh cells. In contrast, we observed positive correlations with the levels of infiltrating DCs, neutrophils, Th17 cells and mast cells. CFHR4 expression showed a strong correlation with various immunomarker groups in HCC. In addition, high CFHR4 expression significantly prolonged the overall survival (OS), disease-specific survival (DSS) and progression-free interval (PFI). We observed a substantial correlation between the expression of CFHR4 and multiple N6-methyladenosine genes in HCC and constructed potential CFHR4-related ceRNA regulatory networks.
Conclusions: CFHR4 might be a potential therapeutic target for improving the HCC prognosis and is closely related to immune cell infiltration.
HCC is the sixth most common cancer worldwide. Over 900,000 new cases of HCC are confirmed each year, and approximately 800,000 people die of HCC annually, making it the third most common cause of cancer-related death. The morbidity and mortality rates of HCC are 2 to 3 times higher in men than in women in most areas (1). In China, the death rate of HCC is the highest among men over 60 years of age. The number of new cases of liver cancer diagnosed each year accounts for approximately 50% of all cases worldwide. The key determinants of liver cancer are chronic HBV infection, aflatoxin exposure or both (1, 2). The development of surgical procedures has improved the survival rate of patients with early-phase HCC, but many patients already have advanced HCC at the diagnosis, resulting in a poor overall survival rate. Therefore, the identification of new, relevant biomarkers is urgently needed to improve the early diagnosis, prognostic assessment and treatment of HCC (3–5).
Research shows that the complement system is a vitally important component of innate immunity and is extensively involved in innate immune recognition, adaptive cell stimulation and proinflammatory effector responses. The complement system exerts a regulatory effect on the tumor microenvironment, influencing the outcome of the immune response (6, 7). The factor H/CFHR family includes five complement F factor H-related proteins (CFHR1/2/3/4/5), factor H and complement factor H‐like protein (CFHL1) (8, 9). CFHRs are secreted plasma proteins synthesized mainly by hepatocytes. CFHR4 is a key component of the innate immune system, and its expression is restricted to the liver (10). To date, numerous studies have suggested a role for CFHR4 in immune system disorders, such as age-related macular degeneration (AMD) (10, 11), systemic lupus erythematosus (12) and atypical hemolytic uremic syndrome (AHUS) (13, 14). However, the association of CFHR4 with HCC has not yet been characterized.
The N6-methyladenosine (m6A) RNA and competing endogenous RNA (ceRNA) regulatory network is currently a new direction in cancer therapy, and the mechanisms have been extensively studied in HCC (15). Current studies mainly focus on methyltransferases, demethylases and binding proteins (16, 17). Although the mechanism of the m6A regulatory factor requires further study, the roles of the m6A regulatory factor in tumor proliferation, invasion and metastasis have been confirmed (18). In addition, ceRNA regulatory networks are also crucial for the emergence and development of multiple cancers, including ovarian cancer (19), esophageal cancer (20) and gastric cancer (21). However, no studies have examined the ceRNA regulatory network of CFHR4 in HCC or reported on its association with m6A regulators.
In the present study, we analyzed CFHR4 expression levels in HCC tumors and normal liver tissue from multiple datasets. An analysis of RNA sequencing (RNA-seq) data from TCGA revealed the clinical relevance and potential diagnostic and prognostic roles of CFHR4 in HCC. In addition, we further explored the biological significance of CFHR4 by performing enrichment analyses and a protein–protein interaction (PPI) network analysis and determining the correlation with immune cell infiltration. After analyzing the correlation of CFHR4 and m6A, we constructed ceRNA regulatory networks involving CFHR4 in HCC.
We first collected gene expression data and clinical data from 424 patients with HCC in TCGA (https://portal.gdc.cancer.gov). In addition, the RNA sequencing data (GSE14520) were downloaded from the Gene Expression Omnibus (GEO) database. HTSeq-FPKM of level 3 format was converted into transcripts per million (TPM). Screening was performed to exclude patients with incomplete information, and the TPM data from 374 patients were used in subsequent analyses (Supplementary Table 1). The evolution process used the “ggplot2” R package.
Normal human liver cells (WRL68) were purchased from AcceGen (Fairfield, USA), and HCC cell lines (BEL7402, SK-hep1, HCCLM3, HepG2 and Huh7) were purchased from the Chinese Academy of Science (Shanghai, China). WRL68 cells were cultured in RPMI-1640 medium, and other cell lines were cultured in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin. All cells were incubated in a 37°C incubator with 5% CO2.
We collected 30 pairs of HCC tissues and adjacent liver tissues at the First Affiliated Hospital of Harbin Medical University from 2006 to 2013 after obtaining informed consent from patients. The research project was conducted under the supervision of the Ethics Committee of the First Affiliated Hospital of Harbin Medical University.
Quantitative real-time PCR was performed on the samples as described previously (5). The following primers were used: CFHR4-F, 5’-TGCGGTTTAAGCTCCATGACA -3’; CFHR4-R, 5’-CCCATCTTCACCACACACTATG-3’; GAPDH-F, 5’ -TGACTTCAACAGCGACACCCA-3’ and GAPDH-R, 5’-CACCCTGTTGCTGTAGCCAAA-3’. GAPDH was used as a control to determine changes in mRNA levels using the 2-ΔΔCT method.
The differentially expressed genes (DEGs) between high CFHR4 expression and low CFHR4 expression samples from TCGA database were analyzed using the DEseq2 (1.26.0) R package (22) with Student’s t test. Differences were considered statistically significant for an adjusted p value < 0.05 and absolute log2-fold change > 1.5. Moreover, volcano plots and heatmaps were constructed to visualize the DEGs.
Pathway enrichment analyses were performed with the “clusterProfiler” R package (23, 24). The c2.cp.v7.2.symbols.gmt curated gene sets were retrieved from the Molecular Signatures Database (MSigDB). Each analytical technique was conducted repeatedly a thousand times. An FDR-corrected q value < 0.25 and adjusted p value< 0.05 were considered statistically significant.
We analyzed the levels of infiltration of 24 types of immune cells in HCC using the ssGSEA method with the GSVA package in R. We then quantified the enrichment score for each immune cell by performing gene expression profiling of each HCC sample based on the signature of immune cells (25, 26).
The univariate Cox regression analysis of the correlation between CFHR4 expression and the values multiple clinical prognostic parameters in patients with HCC was performed using R software with the “survival” package. Using the RMS package (version 6.2-0) and survival package (version 3.2-10), nomograms including important clinical features and calibration plots were constructed. The 45° line represents the best-predicted value, and calibration curves were graphically evaluated by mapping the nomogram-predicted probability against observed occurrences. The consistency index (C-index) was used to measure the discriminative capability of the nomogram and to compare the predictive accuracy of nomograms and individual prognostic indicators. This process was calculated using the bootstrap method and repeated 1000 times. In the present study, one-way analysis of variance (ANOVA) and two-tailed Student’s t test were used to analyze the data. A P value < 0.05 was considered statistically significant.
The TargetScan (http://www.targetscan.org), DIANA-microT (http://diana.imis.athena-innovation.gr/DianaTools/index) and RNAinter (http://www.rnainter.org) online sites were used together to predict and analyze the target miRNAs of CFHR4, compare the correlations between the expression of CFHR4 and target miRNAs and screen miRNAs that were more compatible with ceRNA networks. The target lncRNAs of the screened miRNAs were predicted and analyzed using miRNet2.0 (www.mirnet.ca/miRNet/home.xhtml) and starBase3.0 (www.starbase.sysu.edu.cn), and the correlation between the two was further analyzed to screen for additional eligible ceRNAs. A comprehensive analysis of negatively correlated miRNA–mRNA and miRNA-lncRNA expression levels was performed to establish an HCC-related lncRNA-miRNA–mRNA (CFHR4) ceRNA network.
The R package (version 3.6.3) was used for statistical analyses and plotting. CFHR4 expression in unpaired and paired samples was analyzed using the Wilcoxon rank sum test and Wilcoxon signed rank test, respectively, with the pROC (1.17.0.1) package for ROC analysis. In addition, the Kruskal–Wallis test and univariate Cox analysis were applied to investigate whether CFHR4 expression was associated with clinicopathological factors. Using the KM method and log-rank test, we compared the differences in 10-year OS, DSS and PFI between patients with high CFHR4 expression and those with low CFHR4 expression in TCGA. In all studies, a P value < 0.05 was defined as statistically significant.
By analyzing GTEx and TCGA datasets, we investigated the CFHR4 mRNA levels across cancer types using the Wilcoxon rank sum test, including adrenocortical carcinoma (ACC), bladder urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA), cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC), cholangiocarcinoma (CHOL), colon adenocarcinoma (COAD), esophageal carcinoma (ESCA), glioblastoma multiforme (GBM), head and neck squamous cell carcinoma (HNSC), kidney chromophobe (KICH), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), acute myeloid leukemia (LAML), brain lower grade glioma (LGG), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), mesothelioma (MESO), ovarian serous cystadenocarcinoma (OV), pancreatic adenocarcinoma (PAAD), pheochromocytoma and paraganglioma (PCPG), prostate adenocarcinoma (PRAD), rectum adenocarcinoma (READ), stomach adenocarcinoma (STAD), skin cutaneous melanoma (SKCM), testicular germ cell tumors (TGCT), thyroid carcinoma (THCA), uterine corpus endometrial carcinoma (UCEC) and uterine carcinosarcoma (UCS). We found that CFHR4 expression was significantly decreased in LIHC and CHOL compared with normal tissues (Figure 1A). We obtained similar results from the Timer and GEPIA databases (Supplementary Figures 1A, B). According to the expression of CFHR4 in 374 HCC tissues and 50 normal liver tissues, we confirmed that the CFHR4 expression level was also noticeably decreased in HCC tissues (P<0.001) (Figure 1B). Furthermore, CFHR4 was underexpressed in the GSE14520 HCC cohort (P<0.001) (Figure 1C). Similar results were obtained for adjacent HCC tissues among the 50 matched HCC tissues and adjacent HCC tissues (P<0.05) (Figure 1D). We extracted protein from human normal hepatic cells (WRL68) and HCC cells (BEL7402, SK-hep1, HCCLM3, HepG2 and Huh7) and confirmed the low expression of CFHR4 in HCC cells using Western blot (Figure 1E). Subsequently, 30 pairs of HCC samples were validated, and similar conclusions were reached (Figure 1F). CFHR4 mRNA expression levels were further validated using quantitative real-time PCR analyses (P<0.001) (Figures 1G, H). In addition, we constructed the receiver operating characteristic (ROC) curve. The area under the curve (AUC) for CFHR4 was 0.698, and it has a significant diagnostic value for HCC (Figure 1I).
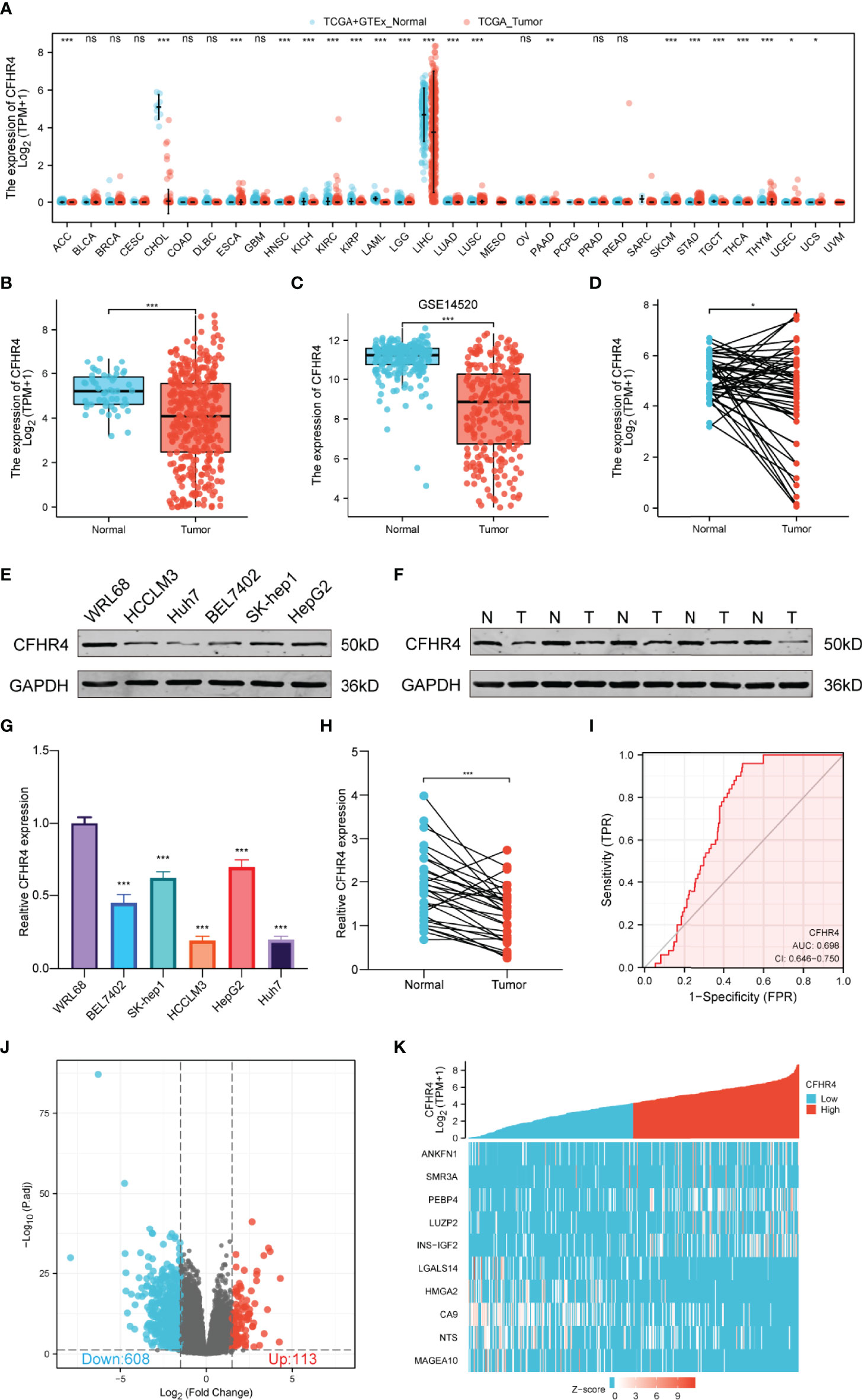
Figure 1 Differences in the expression of CFHR4 and CFHR4-associated DEGs. (A) CFHR4 expression levels in different cancer tissues compared to normal tissues (TCGA). (B–D) CFHR4 expression in HCC samples. (E) CFHR4 expression was detected in WRL68, BEL7402, SK-Hep1, HCCLM3, HepG2, and Huh7 cell lines using Western blotting. (F) CFHR4 protein expression in 30 paired adjacent noncancerous tissues and HCC tissues. (G) CFHR4 expression was detected in WRL68, BEL7402, SK-Hep1, HCCLM3, HepG2, and Huh7 cell lines using PCR. (H) CFHR4 mRNA expression in 30 paired adjacent noncancerous tissues and HCC tissues. (I) ROC curves were created to investigate the value of CFHR4 in identifying HCC tissues. (J, K) Volcano plots of the DEGs and heatmap showing the top 10 DEGs. *p < 0.05, **p < 0.01, ***p < 0.001, NS, no significance.
According to the CFHR4 expression level, we divided the data from patients with HCC into high and low CFHR4 expression groups for comparison. The DESeq2 package was used to infer CFHR4-associated genes and analyze the DEGs between the high and low expression groups. An adjusted p value < 0.05 and absolute log2-fold change > 1.5 were considered statistically significant. A total of 721 significant DEGs were identified. 113 DEGs were associated with the high CFHR4 expression group, and 608 DEGs were associated with the low CFHR4 expression group (Figure 1J and Supplementary Table 2). The top 10 DEGs were identified, further analyzed using HTSeq-Counts and sorted by relative expression (Figure 1K).
GO and KEGG enrichment analyses were performed using the “clusterProfiler” R package to further analyze the potential biological functions of CFHR4-related DEGs. The GO analysis indicated that CFHR4-related DEGs may be involved in gated channel activity, regulation of signal release, regulation of ion transmembrane transport, metal ion transmembrane transporter activity, synaptic membrane, transmembrane transporter complex and passive transmembrane transporter activity (Figures 2A, B; Supplementary Table 3). In the KEGG enrichment analysis, CFHR4-related DEGs were mainly involved in chemical carcinogenesis, retinol metabolism, the calcium signaling pathway, the PPAR signaling pathway, bile secretion, insulin secretion and gastric acid secretion (Figures 2C, D).
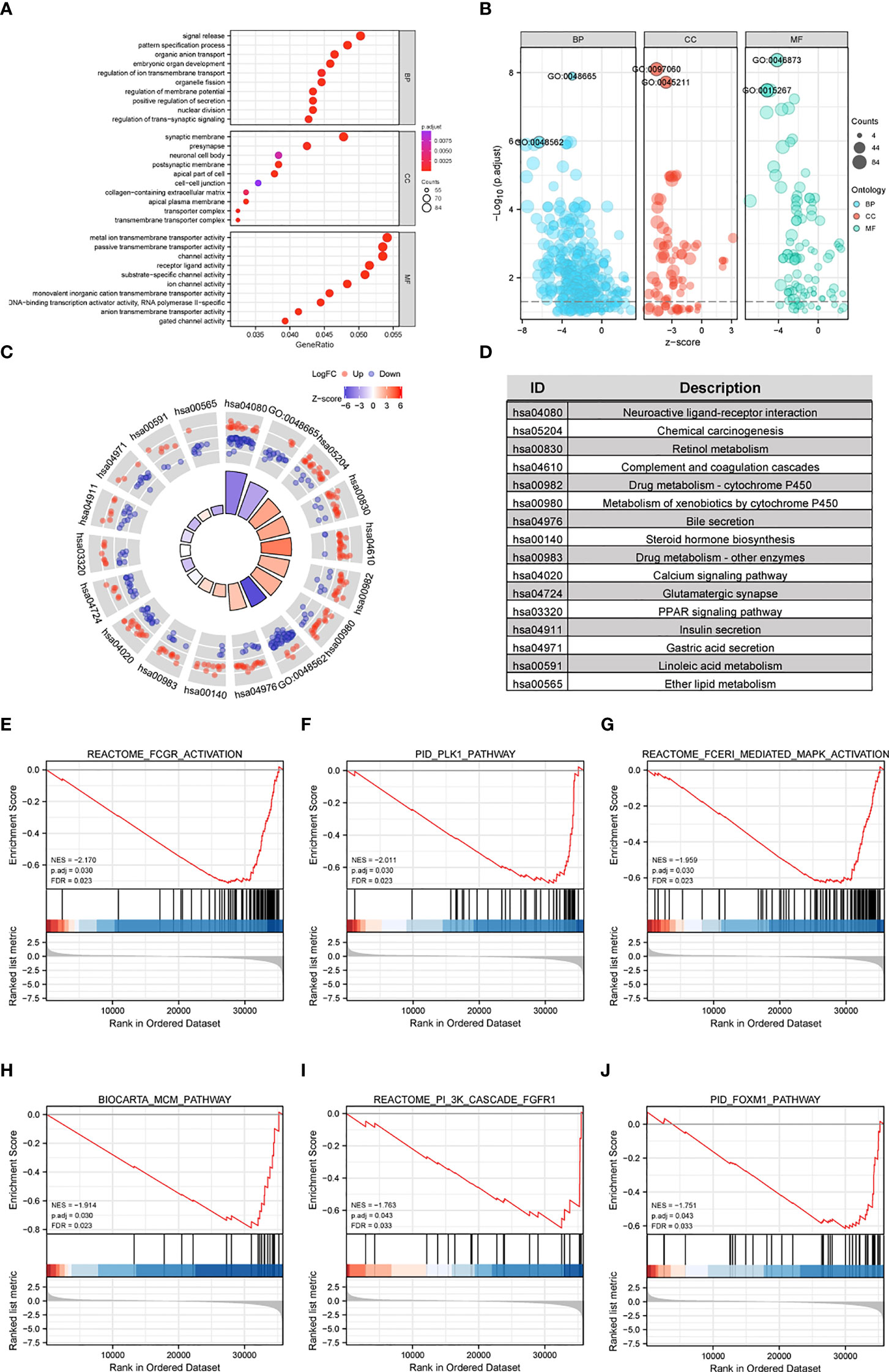
Figure 2 Functional enrichment analyses of CFHR4-related genes in HCC. (A, B) The enriched terms in GO categories in HCC. (C, D) KEGG pathway analysis based on CFHR4-associated DEGs. (E–J) GSEA enrichment plots, including FCGR, activated reaction, PLK1 pathway, reactant FCERI-mediated MAPK activation, ATR pathway, MCM pathway, cascade reactions of PI3K and FGFR1, reactant-mediated MAPK activation and FOXM1 pathway.
GSEA was conducted between the high and low CFHR4 expression groups to further reveal CFHR4-related signaling pathways in HCC. The following pathways were significantly enriched in patients with low CFHR4 expression: FCGR-activated reaction, PLK1 pathway, reactant FCERI-mediated MAPK activation, ATR pathway, MCM pathway, cascade reaction of PI3K and FGFR1, reactant-mediated MAPK activation and FOXM1 pathway (Figures 2E–J; Supplementary Table 4).
We explored the association between 721 DEGs in the HCC group using the STRING database by setting the interaction threshold to 0.70 and constructed a PPI network to further investigate the underlying mechanisms (Figure 3A; Supplementary Table 5). Subsequently, 301 proteins and 420 edges were screened, and five central gene clusters were identified using a total score ≥5000 (Figures 3B–F). In addition, the top 7 central genes were screened, including CENPA, CDC20, UBE2C, CEP55, BIRC5, FAM64A and TRIP13 (Figure 3G). By analyzing the GeneMANIA and STRING online datasets, potential CFHR4-interacting target genes were identified (Supplementary Figures 2A, B). CFHR4-related genes were selected by performing a crossover analysis, including C3, CRP, CFHR1, CFHR3 and CFHR5 (Supplementary Figure 2C). We subsequently analyzed the association between CFHR4 and the 5 intersecting genes (Supplementary Figures 2D–H).
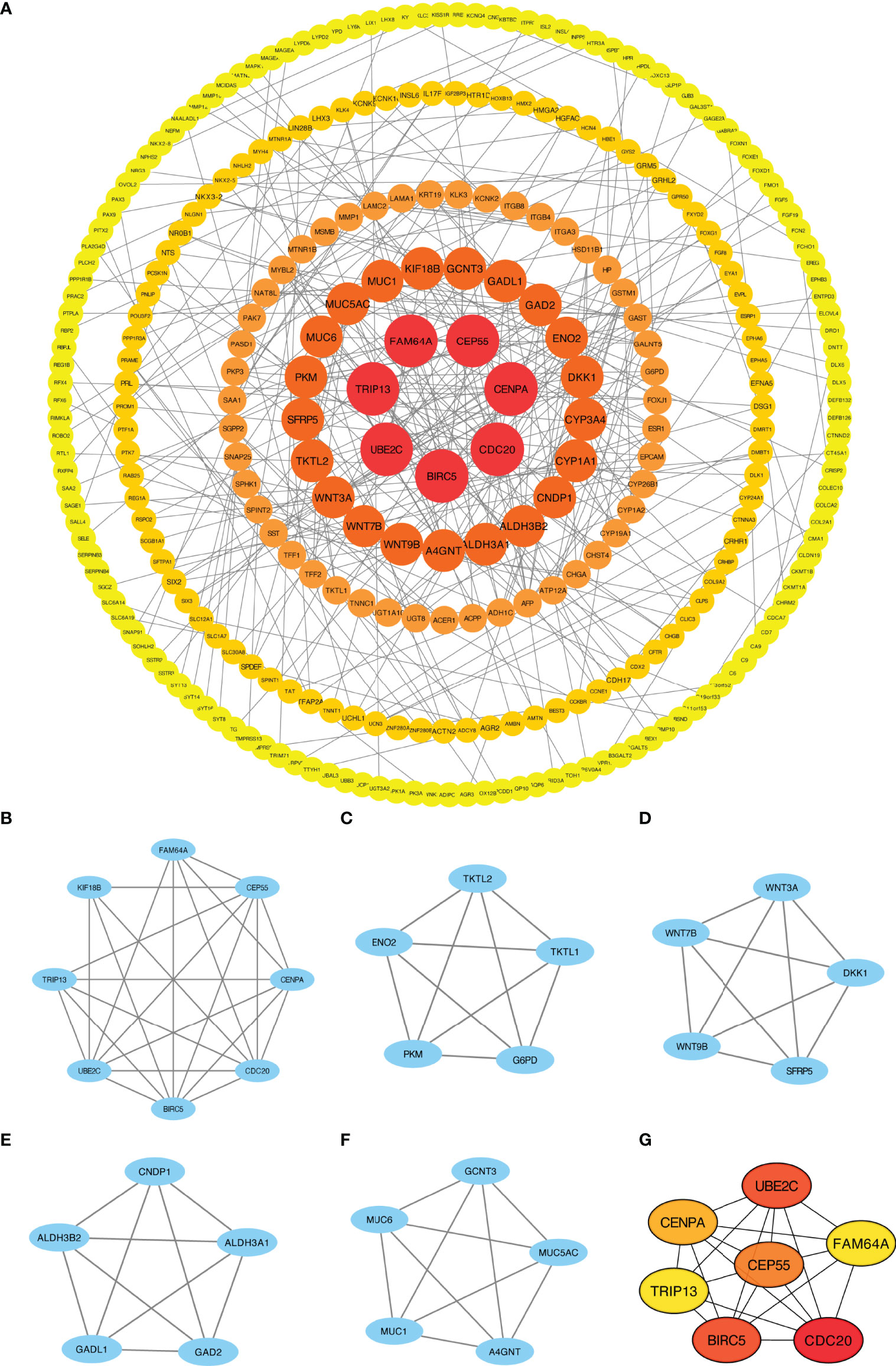
Figure 3 PPI network enrichment analysis. (A) The PPI network was built based on PPI pairs identified by the STRING dataset. (B–F) Hub gene clusters were selected from the PPI network (criteria of total scores ≥ 5,000). (G) Top 7 hub genes in the PPI network.
Based on the ssGSEA algorithm, we confirmed and quantified the correlations between CFHR4 expression and the immune cell infiltration levels (Figure 4A). The expression of CFHR4 was negatively correlated with aDCs, TFH cells, NK CD56bright cells and Th2 cells, and it has positive correlations with Th17 cells, DCs, neutrophils, mast cells, Tgd cells, Tcm cells, cytotoxic cells, Tregs, NK cells, pDCs, eosinophils, iDCs, B cells, T cells, CD8 T cells, Tems, NK CD56dim cells, T helper cells, macrophages and Th1 cells (Figures 4B–H). We further confirmed the correlation between CFHR4 expression with immunomarker of various immune cells in HCC. The results showed that CFHR4 expression was significantly correlated with the immunomarkers IRF5 and INOS of M1 macrophages in HCC (Table 1). It indicated that CFHR4 may induce macrophages to M1 polarization in HCC. This analysis of immune markers of different functions T cells showed that CFHR4 expression was highly correlated with the most immunomarkers (CD8B, CD3D, STAT1, IFN-γ, STAT5A, IL21, TGFβ, PD-1, CTLA4, LAG3 and TIM-3) of T cells in HCC (Table 1). It turns out that CFHR4 may perform an indispensable role in the T cells’ immune response to HCC. Especially for T cells exhaustion, consistent results with the GISTIC analysis were obtained. The somatic copy number alteration (SCNA) module demonstrated that the arm-level deletion of CFHR4 was markedly associated with immune cell infiltration levels in HCC (Figure 4I). In addition, the results also showed a correlation between CFHR4 expression and the immunomarkers of TAMs, neutrophils and dendritic cells (Table 1). Subsequently, according to the expression level of CFHR4, HCC samples were dichotomized into CFHR4-high and low expression groups, we aimed to reveal whether different expression groups of CFHR4 differ in the tumor immune microenvironment of HCC (Figure 4J). We found that cytotoxic cells, DCs, iDCs, mast cells, neutrophils, NK cells, pDCs, Tcm cells, Tem cells, Tgd cells, Th17 cells and Tregs were increased in the CFHR4 high expression group (P < 0.05), while the NK CD56bright cells, TFH cells and Th2 cells decreased (P < 0.05). These findings confirmed that reduced expression of CFHR4 in HCC was closely associated with immune cell infiltration.
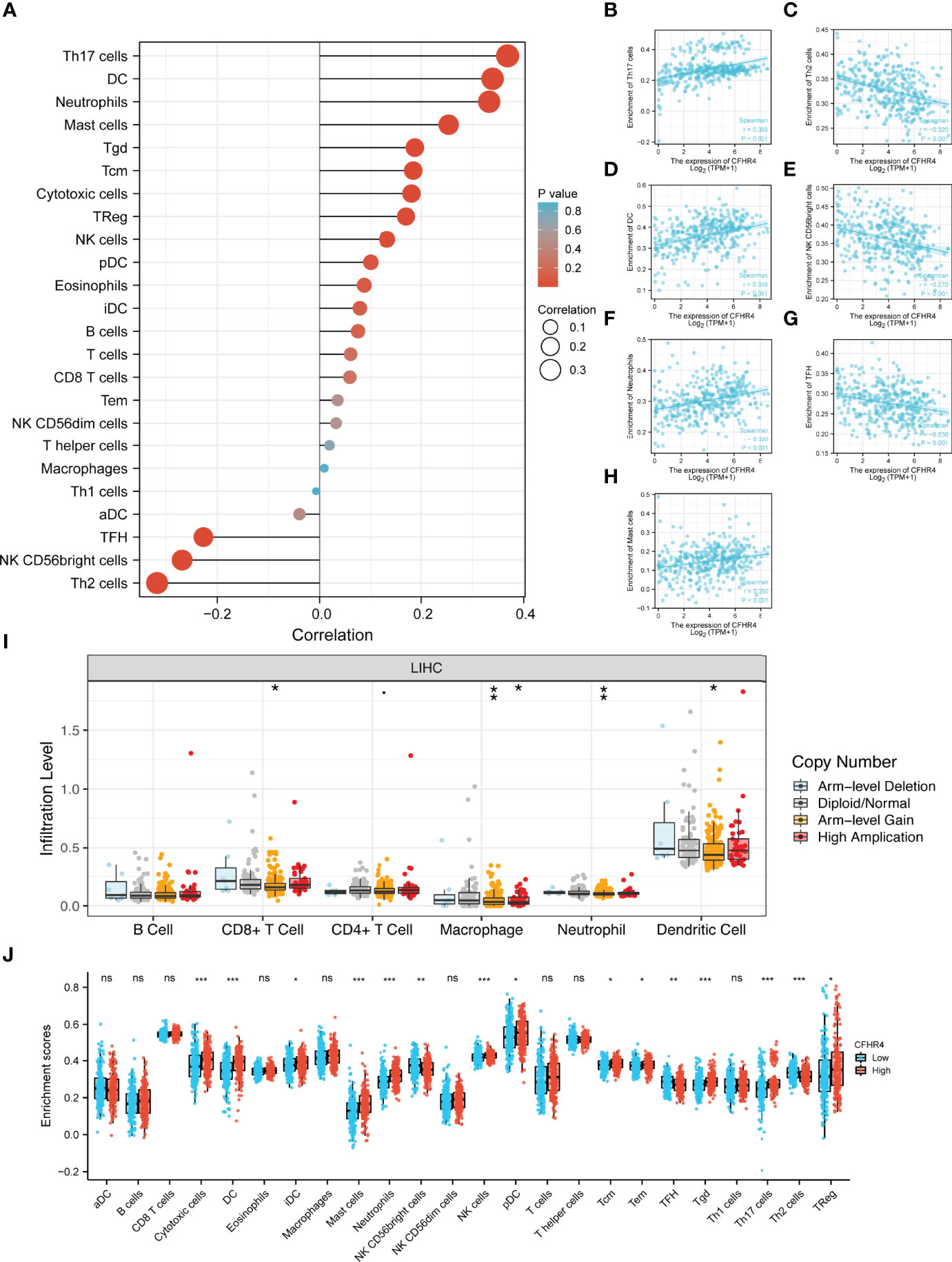
Figure 4 Integrative analysis of CFHR4 expression in the infiltrating immune microenvironment. (A) The forest plot depicts the relationship between the level of CFHR4 expression and the relative abundances of 24 immune cells. (B–H) Scatter plots showing the differentiation of Th17 cells, Th2 cells, DCs, NK CD56bright cells, neutrophils, TFH cells and mast cells infiltration levels between high and low groups of CFHR4 expression. (I) The SCNA showed that CFHR4 expression correlated with the level of immune cell infiltration. (J) Scatter plots showing the correlations between 24 immune cells and CFHR4 expression levels. *p < 0.05, **p < 0.01, ***p < 0.001, NS, no significance.
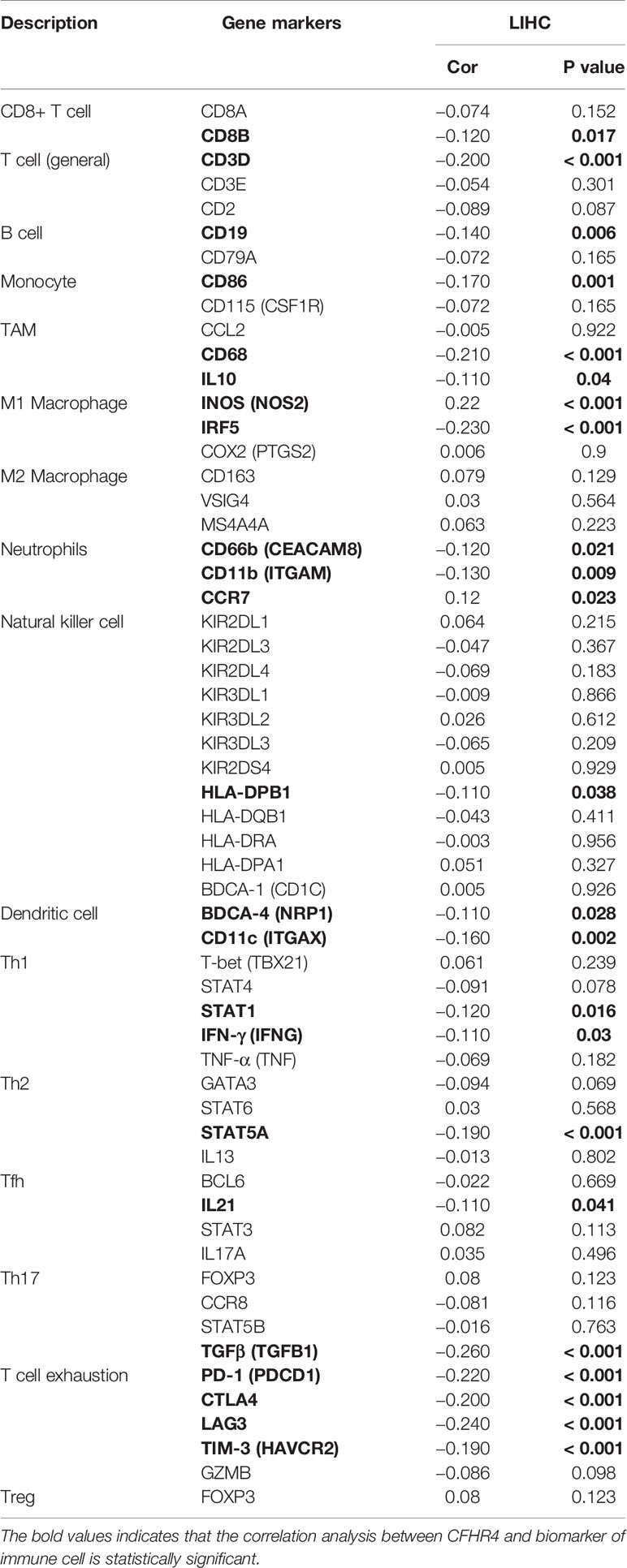
Table 1 Correlation analysis between CFHR4 expression and biomarkers of immune cells.
The clinical data from patients with HCC in TCGA database were obtained to investigate the clinical characteristics of patients with different CFHR4 expression levels. After removing patients with incomplete clinical data, 374 patients remained for further analysis; the average age was 61.5 years (49.25 to 70.00 years), and 67% were male. Table 2 provides a detailed description of the clinical data. We evaluated the differences in clinicopathological variables after stratifying patients based on CFHR4 expression using the Kruskal–Wallis test, and the level of CFHR4 was strongly correlated with age, sex, race, TNM stage, histologic grade, pathological stage, tumor status, residual tumor, vascular invasion and AFP level (Figures 5A–L). Notably, CFHR4 was expressed at higher levels in the older age group (age>60 years) than in the younger age group (age ≤ 60 years) (P<0.05). Significant differences in CFHR4 expression levels were also noted in different races (P<0.001). Moreover, a higher histological grade, TNM grade, pathological stage and tumor status were also significantly associated with low CFHR4 expression. Subsequently, we further confirmed the lower CFHR4 expression level in the group with a high AFP level (>400 ng/mL) (P<0.001). Based on these results, patients with HCC presenting lower CFHR4 expression seemed to have a more advanced tumor stage.
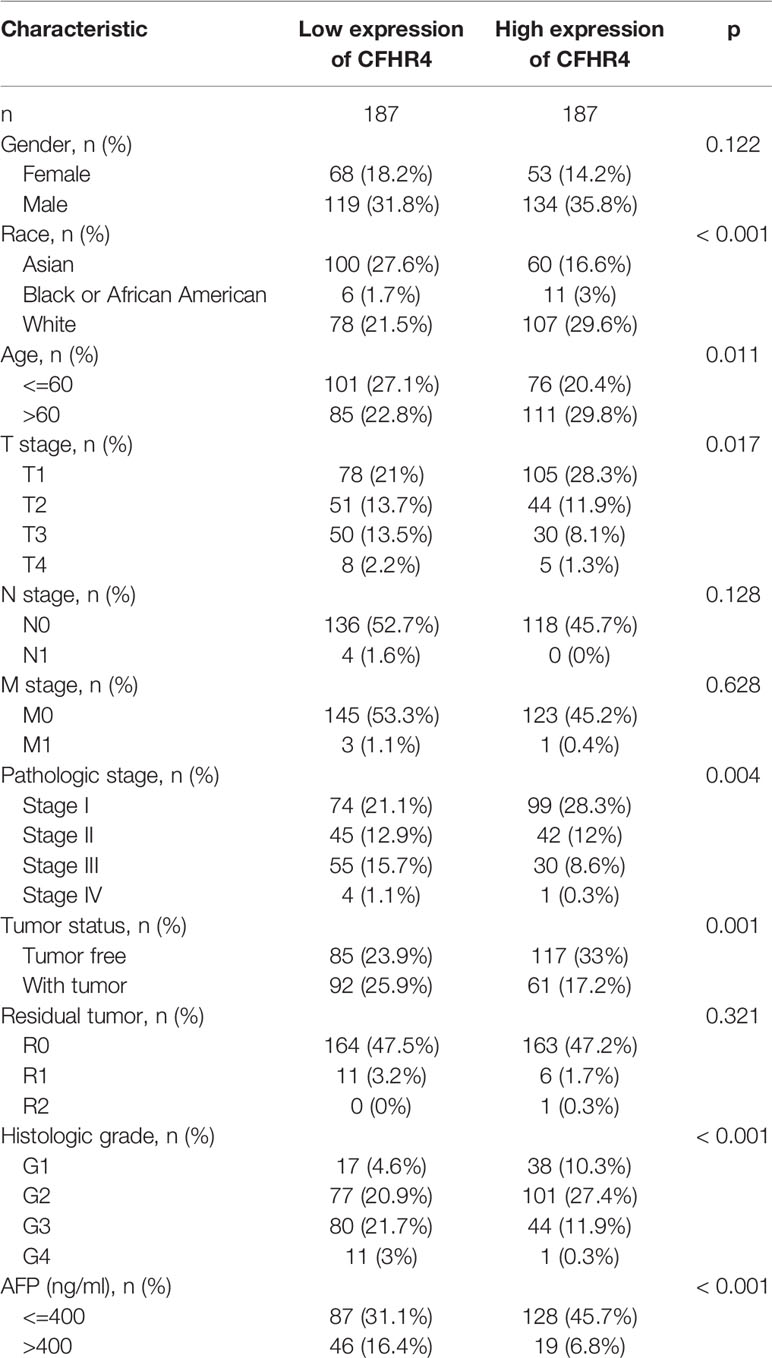
Table 2 The correlations between clinicopathological variables and CFHR4 expression.
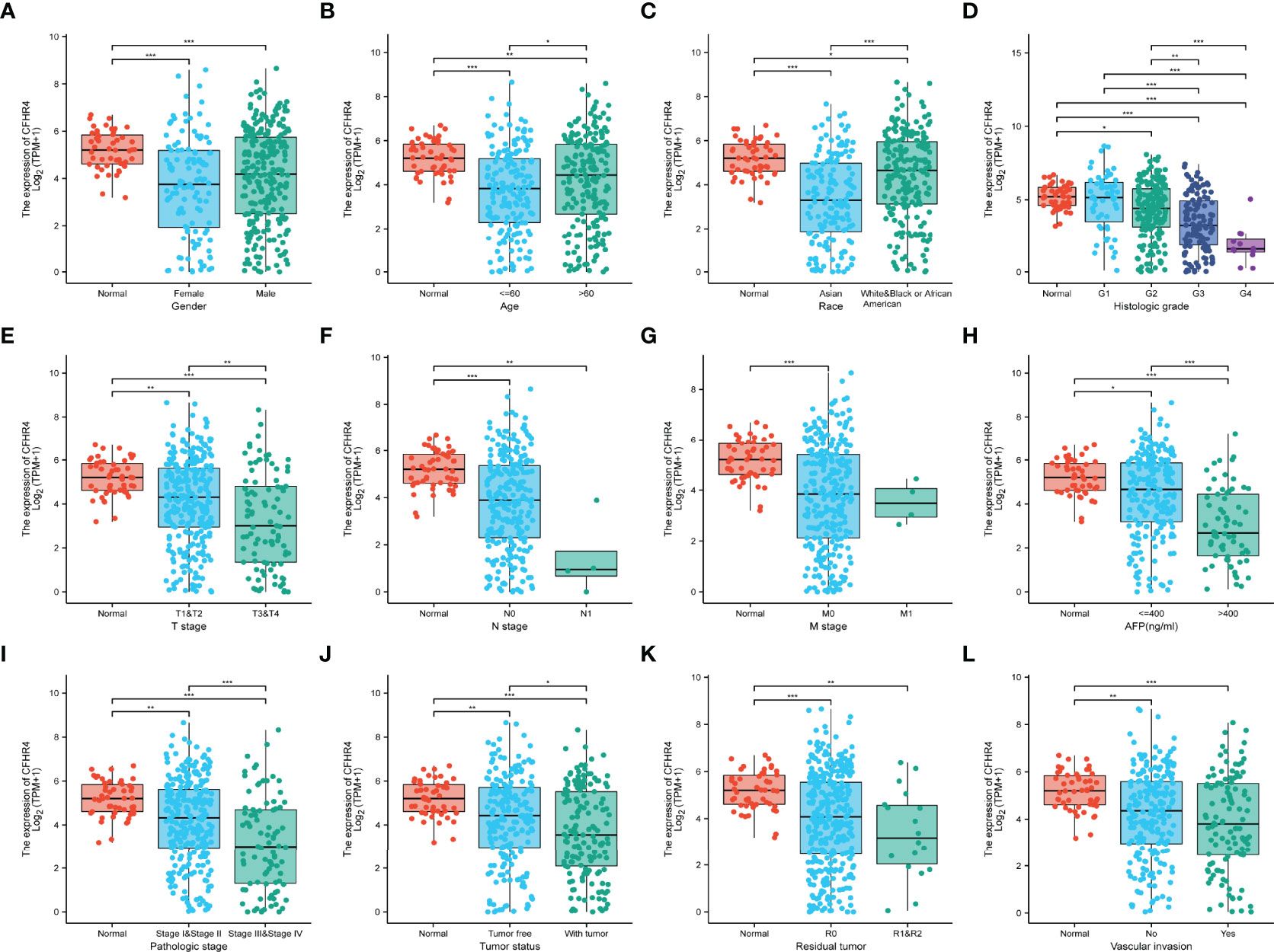
Figure 5 Correlation of CFHR4 expression with clinicopathological characteristics. (A) Sex. (B) Age. (C) Race. (D) Histologic grade. (E) T stage. (F) N stage. (G) M stage. (H) AFP level. (I) Pathological stage. (J) Tumor status. (K) Residual tumor. (L) Vascular invasion. *p < 0.05, **p < 0.01, ***p < 0.001.
Afterward, we performed a series of studies to determine the association of CFHR4 expression levels with the prognosis of patients with HCC. The Kaplan–Meier Plotter analysis revealed an association between low CFHR4 expression and a poor prognosis (Figures 6A–C). Moreover, we performed subgroup analyses of OS, DSS and PFI. Patients with high CFHR4 expression had a correspondingly better prognosis for OS, DSS and PFI in the Asian group (Figures 6D–F). However, OS, DSS and PFI in the white and black or African–American subgroups were not significantly different (Supplementary Figures 3A–C). In addition, patients with HCC presenting high CFHR4 expression who were aged ≤ 60 years experienced longer OS and DSS but had a worse prognosis in terms of PFI (Figure 6G–I). However, no significant differences were observed in the younger age subgroups for OS, DSS and PFI (age ≤ 60 years) (Supplementary Figures 3D–F). We further confirmed that the T3 and T4 subgroups and the stage III and stage IV subgroups experienced poorer OS (Supplementary Figures 3G, H).
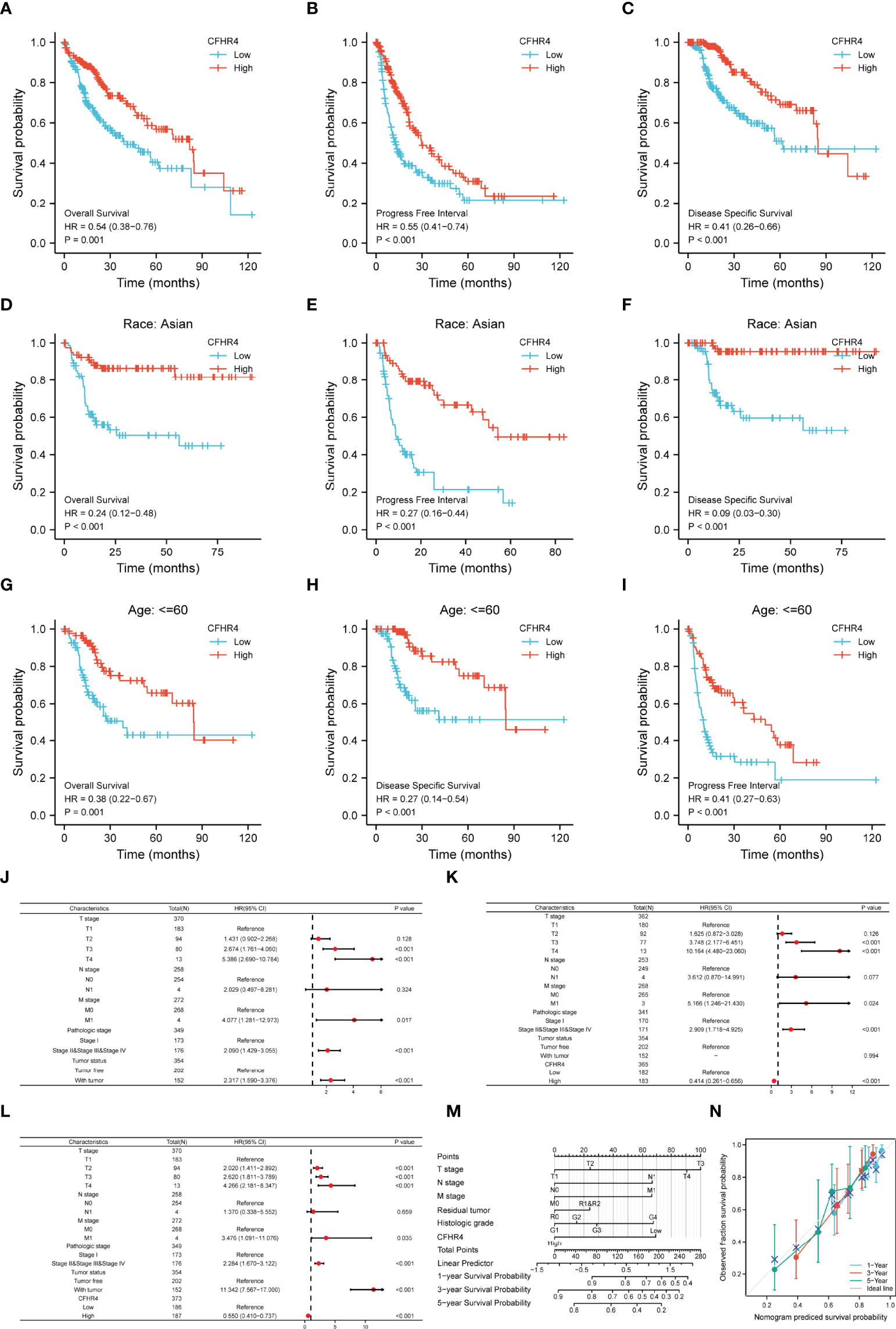
Figure 6 The prognostic value of CFHR4 in HCC. (A–C) Survival curves showing a comparison of OS, DSS and PFI between patients with HCC presenting high and low CFHR4 expression. (D–F) OS, DSS and PFI survival curves for Asian patients with HCC presenting high and low CFHR4 expression. (G–I) OS, DSS and PFI survival curves for patients with HCC aged ≤60 years presenting with high and low CFHR4 expression. (J–L) Univariate survival analysis of OS, PFI, and DSS in patients from different subgroups stratified according to TNM stage, pathological grade, tumor status, and CFHR4 expression levels. (M) For patients with HCC, a nomogram was constructed to estimate the probability of 1-, 3-, and 5-year OS. (N) Nomogram calibration plots for determining the probability of OS at 1, 3, and 5 years.
A univariate Cox regression analysis was performed with TNM stage, pathological grade, tumor status and CFHR4 expression levels to further identify factors associated with different prognoses (Supplementary Table 6). The forest plot illustrated that low expression of CFHR4 was a risk factor for the OS (Figure 6J; Supplementary Table 7), DSS (Figure 6K; Supplementary Table 6) and PFI (Figure 6L; Supplementary Table 8) of patients with HCC. According to the results of the univariate Cox regression analysis, CFHR4 expression and other independent clinicopathological factors were used to construct the point scale of the nomogram. Each variable was scored with reference to the scale of the nomogram, and the total scores were dispatched to the outcome line and predicted the prognosis of patients with at 1, 3 and 5 years. The C-index of the nomogram was 0.706 (95% confidence interval: 0.671-0.741). This result suggested that the prognostic nomogram of CFHR4 had good discriminatory power (Figure 6M). The deviation correction line in the calibration analysis approached the ideal curve, indicating that the predicted values were consistent with the observed values (Figure 6N). Consistent results were obtained with the univariate Cox regression analysis.
As reported in previous studies, m6A RNA methylation exerts an important effect on the development of HCC (27–29). The correlations between CFHR4 expression and the expression of 23 m6A-related genes were analyzed in TCGA (Figure 7A). The correlation analysis showed significant negative correlations between the expression of CFHR4 (P < 0.05) and 15 m6A-related genes in HCC (Figures 7B–P). Furthermore, groups were established based on the median CFHR4 expression, and 211 patients were assigned to the high expression group and 210 patients were assigned to in the low expression group. We determined the relationship between the CFHR4 expression level and m6A modification level in HCC by analyzing the differential expression of 23 m6A-related genes in different expression groups (Figure 7Q). The expression of YTHDC1, IGF2BP1, IGF2BP2, IGF2BP3, YTHDF1, YTHDF2, HNRNPA2B1, LRPPRC, HNRNPC, RBMX, METTL16, METTL3, RBM15, RBM15B, VIRMA, WTAP and ALKBH5 was reduced in the high CFHR4 expression group (P < 0.05). In summary, a strong correlation was observed between m6A RNA methylation in HCC and the CFHR4 expression level.
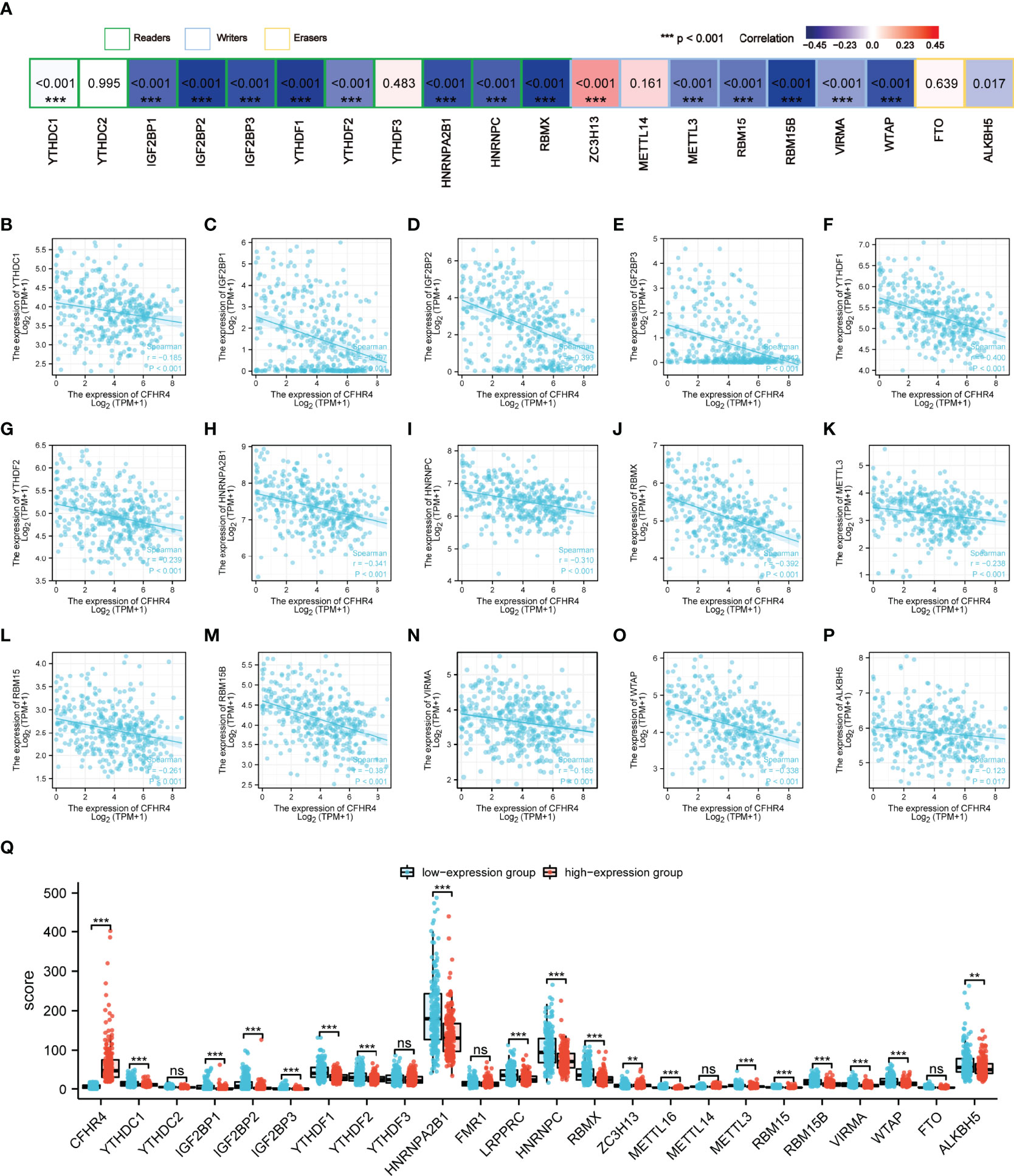
Figure 7 Analysis of the association between the CFHR4 expression level and the expression of m6A-related genes in HCC. (A) Correlation of CFHR4 expression levels with m6A gene expression in HCC. (B–P) Scatter plot showing the relationship between CFHR4 and m6A genes. (Q) Correlation of m6A genes in the CFHR4 high and low expression groups of HCC tumor samples. **p < 0.01, ***p < 0.001, NS, no significance.
Accumulating evidence highlights the regulatory role of lncRNA–miRNA–mRNA ceRNA networks in cancers. Therefore, we analyzed and constructed a ceRNA regulatory network for CFHR4 in HCC. Through TargetScan, DIANA-microT and RNAinter database predictions, the following 11 miRNAs were jointly predicted: hsa-miR-32-3p, hsa-miR-142-5p, hsa-miR-146a-5p, hsa-miR-302c-5p, hsa-miR-361-5p, hsa-miR-4775, hsa-miR-4786-5p, hsa-miR-4795-3p, hsa-miR-5590-3p, hsa-miR-580-3p and hsa-miR-590-3p (Figure 8A). Based on the regulatory relationship in the ceRNA network, a negative correlation was observed between mRNAs and miRNAs. Four miRNAs negatively correlated with CFHR4 expression were identified and screened by performing a correlation analysis. The scatter plots showed the correlation between CFHR4 expression and the target miRNAs, and the TargetScan database was used to predict the potential binding sites in CFHR4 for target miRNAs (Figures 8B–E). Subsequently, the lncRNAs that may interact with the target miRNAs (hsa-miR-146a-5p, hsa-miR-361-5p and hsa-miR-580-3p) were further predicted using the miRNet and starBase databases (Figures 8F–H). This interaction is due to the negative correlation between the expression of lncRNAs and miRNAs. Consequently, using the starBase database, we further screened and confirmed the lncRNAs in HCC that were negatively correlated with the three target miRNAs. Based on these results, the following 10 ceRNA regulatory networks that play a role in HCC were constructed: TMEM161B-AS1-hsa-miR-146a-5p-CFHR4, CCDC183-AS1-hsa-miR-146a-5p-CFHR4, NEAT1-hsa-miR-146a-5p-CFHR4, MALAT1-hsa-miR-146a-5p-CFHR4, XIST-hsa-miR-146a-5p-CFHR4, DNAAF4-CCPG1-hsa-miR-361-5p-CFHR4, NEAT1-hsa-miR-580-3p-CFHR4, LINC00641-hsa-miR-580-3p-CFHR4, DNAAF4-CCPG1-hsa-miR-580-3p-CFHR4 and DSCAM-AS1-hsa-miR-580-3p-CFHR4 (Figure 8I).
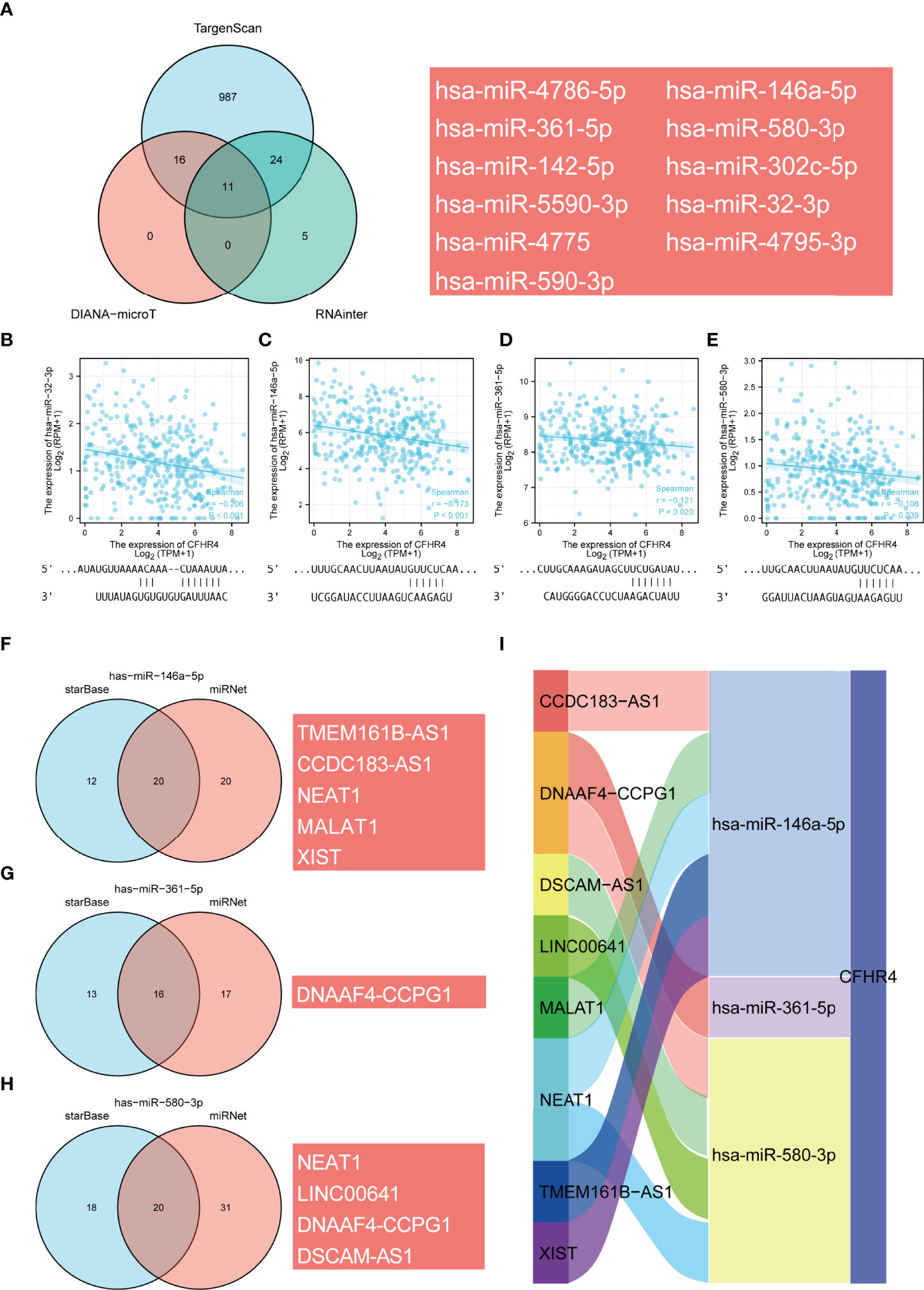
Figure 8 Prediction of the ceRNA network in HCC. (A) Venn diagram showing the results for CFHR4 targets predicted using the TargetScan, DIANA-microT and RNAinter databases. (B–E) Scatter plots were generated to show miRNAs-mRNAs with significant correlations. TargetScan prediction of the potential binding sites in CFHR4 for the target miRNAs. (F–H) The lncRNAs that bind to target miRNAs were predicted using the miRNet and starBase online databases and displayed in a Venn diagram, including hsa−miR−146−5p, hsa−miR−361−5p and hsa−miR−580−3p. (I) Sankey diagram showing the CFHR4-related ceRNA regulatory network.
The CFHR family consists of five highly related proteins. Each CFHR gene has a completely duplicated structural domain in the plasma proteins, and they share high sequence identity (8, 9). Members of the CFHR family of proteins play key roles in the progression of multiple diseases through multiple mechanisms. For example, CFHR1 exacerbates atherosclerotic cardiovascular disease by altering the expression levels of C-reactive protein apolipoprotein and serum amyloid protein A (30). All CFHR genes are genetic risk factors for AMD (31). The CFHR family of genes is also important in AHUS and C3 glomerulopathy (11, 13, 32). In addition, some members of the CFHR family of proteins have been proven to exert a marked effect on the progression of a variety of cancers (33–35). However, few studies on CFHR4 have been conducted, and no studies have determined its role in cancer.
In the present study, we measured the expression level and prognostic value of CFHR4. We confirmed that CFHR4 mRNA expression was markedly downregulated in HCC and CHOL tissues, and these results were validated in multiple databases. The ROC curve analysis suggested that CFHR4 may be a promising diagnostic biomarker for differentiating HCC from normal tissue.
We confirmed the reduced expression of CFHR4 in HCC cell lines and HCC samples by performing in vitro experiments. We analyzed the DEGs related to CFHR4 to further assess the role of CFHR4 in HCC. By conducting GO and KEGG analyses, we found that differences in CFHR4 expression were significantly correlated with regulating signal release, regulation of ion transmembrane transport, gated channel activity, metal ion transmembrane transporter activity, calcium signaling pathway and the PPAR signaling pathway. Using GSEA, we also revealed that low CFHR4 expression was significantly associated with FCGR-activated reactions, the PLK1 pathway, reactant FCERI-mediated MAPK activation, the ATR pathway, the MCM pathway, the cascade reactions of PI3K and FGFR1, reactant-mediated MAPK activation and the FOXM1 pathway in patients. PLK1 (36), MAPK (37), ATR (38), MCM (39), PI3K and FGFR1 (40) have been shown to play increasingly crucial regulatory roles in HCC, and these studies and our results indicated that CFHR4 may inhibit the development and progression of HCC by regulating these signaling pathways. However, the association of CFHR4 with these signaling pathways was first discovered here, and the regulatory mechanisms require further exploration. Furthermore, based on the DEGs, we constructed the PPI networks using the Cytoscape tool. Five central gene clusters (a total score ≥ 5000) and the top 7 central genes were screened, including CENPA, CDC20, UBE2C, CEP55, BIRC5, FAM64A and TRIP13. The CFHR4-interacting genes were generated using STRING and GeneMANIA online databases, and we observed five intersecting genes, including C3, CRP, CFHR1, CFHR3 and CFHR5. Existing studies have confirmed that CFHR4 regulates complement activation and opsonization on biological surfaces by interacting with native CRP (Hebecker et al., 2010). CFHR4 interacts with C3b (C3 activation fragment) (Hellwage et al., 1999, Hebecker and Jozsi, 2012). These conclusions promote the credibility of the predictions from the STRING database and will provide critical insights into the design of follow-up studies and experimental validation.
Among the results, tumor infiltrating immune cells (TIICs) were recently shown to play a pivotal regulatory role in tumor progression (41). The substantial accumulation of TIICs in HCC affects the prognosis of HCC (42). By revealing the relationship between CFHR4 expression and the level of immune cell infiltration in HCC, CFHR4 expression was clearly associated with the infiltration of Th17 cells, DCs, neutrophils and Th2 cells. Th17 cells are a major effector subset of CD4+ T cells that play a vital role in host protection and autoinflammatory disorders (43, 44). The differentiation of Th17 cells into Th1 and Th2 cell subsets participates in regulating the response to intracellular pathogens and extracellular organisms (45). Th1/17 cells produce IFN-γ to drive antitumor immune responses (46). Multiple studies reported that increased infiltration of Th17 cells inhibits the progression of breast cancer (47). Moreover, DCs are specialized antigen-presenting cells that play important roles in the initiation and regulation of innate and adaptive immune responses (48). The antitumor effect of DCs has been confirmed (49). Neutrophils have also been proven to exert bidirectional regulatory effects on the tumor immune microenvironment (50). Our studies indicated that high CFHR4 expression activated Th17 cells, DCs and neutrophils to promote antitumor immune responses. In addition, antigen-presenting cells might promote the polarization of CD4+ T cells toward Th1 and Th2 cell subsets. Th1 cells are mainly involved in cellular immunity and tumor clearance, and Th2 cells are involved in the stimulation of antibody production (51). Th2 cells have also been confirmed as an independent risk factor for cancer growth and progression (52, 53). The number of NK CD56bright cells is significantly increased in various cancers (54–56). Multiple studies reported that Tfh cells are a specialized subset of CD4+ T cells that support the germinal centers, which secrete high-affinity antibodies and provide help for memory B cells (57, 58). Additionally, Tfh cells were confirmed to be involved in human autoimmune responses and cancers (59, 60). Based on this information, CFHR4 modulates immune responses mediated by Th2 cells, NK CD56bright cells and Tfh cells in HCC. We also found that the CFHR4 CNV was significantly correlated with the levels of infiltrating CD8+ T cells, macrophages, neutrophils, and dendritic cells. In addition, CFHR4 expression is strongly correlated with various immunomarker groups in HCC. We confirmed significant correlations between CFHR4 expression and CD8+ T cells (CD8B), monocytes (CD86), TAMs (CD68 and IL10), M1 macrophages (NOS2 and IRF5), neutrophils (CD66b, CD11b, and CCR7), natural killer cells (HLA-DPB1), dendritic cells (NRP1 and ITGAX), Th1 cells (STAT1 and IFN-γ), Th2 cells (STAT5A), Tfh cells (IL21), Th17 cells (TGFβ) and exhausted T cells (PD-1, CTLA4, LAG3, and TIM-3). Our identified a potentially indispensable role for CFHR4 in regulating immune cell infiltration in HCC. We explored the relationship between CFHR4 expression with OS, PFI, DSS and clinical characteristics (TNM stage, residual tumor, and histological grade) by performing univariate Cox regression analysis. Calibration plots showed good agreement between predicted values of CFHR4-related column line plots and forecasted and observed values for 1-, 3- and 5-year OS probabilities. These results were consistent with those of the univariate Cox regression analysis.
The m6A methylation exerts a substantial effect on tumor cell proliferation, invasion and migration (61). Currently, m6A RNA and ceRNA regulatory networks are widely studied to determine HCC mechanisms (15). We further analyzed the relationship between CFHR4 expression and m6A modifications and determined that CFHR4 expression had inseparable relationships with IGF2BP2, IGF2BP3, YTHDF1, HNRNPA2B1, LRPPRC, HNRNPC, RBMX, RBM15B and WTAP expression. We also observed significant correlations between high CFHR4 expression and YTHDC1, IGF2BP1, IGF2BP2, IGF2BP3, YTHDF1, YTHDF2, HNRNPA2B1, LRPPRC, HNRNPC, RBMX, METTL16, METTL3, RBM15, RBM15B, VIRMA, WTAP and ALKBH5 expression. Multiple studies have now reported that IGF2BP1 (62), IGF2BP2 (28), IGF2BP3 (63), YTHDF1 (64), YTHDF2 (29), BRMX (65), RBM15 (66), METTL3 (67) and WTAP (27) are significantly upregulated in HCC, and their overexpression promotes HCC progression and is associated with a poor prognosis for patients with HCC. These discussions further supported our results. Thus, these findings suggested that the CFHR4 gene may be modified by m6A to increase the stability of its mRNA, which further inhibits the proliferation, invasion and migration of HCC. Subsequently, we constructed ceRNA regulatory networks based on the prediction. Because the ceRNA regulatory networks of CFHR4 were derived from a bioinformatics analysis, more experiments are needed to validate this network in future studies.
Although we increased our awareness of the regulatory mechanism of CFHR4 in HCC, the study had several limitations. Initially, the expression levels of CFHR4 and the important regulatory mechanisms and pathways related to CFHR4 in HCC should be further validated and evaluated by analyzing clinical samples from more centers. Secondly, However, the potential diagnostic value of the circulating CFHR4 content in HCC patients is not clear, and the clinical significance of circulating tumor markers remains to be further explored. In addition, the relationship between CFHR4 and interacting genes and m6A genes in HCC should be further explored and validated. In future studies, we will further elucidate the potential regulatory mechanisms of CFHR4 in HCC by performing more experiments.
In summary, this study represents the first in-depth analysis of CFHR4 in HCC. Our study suggested that CFHR4 was abnormally downregulated in HCC and that its reduced expression was correlated with a poorer prognosis. We confirmed the correlation between CFHR4 expression and the m6A modification, indicating that CFHR4 may be modified by m6A to improve mRNA stability. The construction of ceRNA networks suggested that CFHR4 may be involved in multiple molecular regulatory mechanisms of HCC. More importantly, CFHR4 expression was associated with multiple immune cells and may affect HCC tumor immunity by inducing M1 macrophage polarization and altering the infiltration of exhausted T cells. These findings provide additional insights into the mechanism by which CFHR4 may represent an important independent prognostic marker for HCC. The potential molecular mechanisms and regulatory networks of CFRH4 provide a basis for follow-up studies. The study also provides important insights into the treatment of HCC based on genomics.
The article/Supplementary Material contains the original contributions presented in the study. Any additional questions can be forwarded to the corresponding authors.
The Ethics Committee of the First Affiliated Hospital of Harbin Medical University provided ethical review and approval. The patients provided their written informed consent to participate in this study.
HY, CW, SK, and MB made equal contributions to this article. HY, CW, and SK planned the research trials and engaged in article writing. YNX, SL, ZF, BQ, YX, and YF attended in information generation and analysis. MZ, ZL, BY, XL, YH, YZ, and SP also provided assistance with analysis. All authors made great efforts to the article and agreed to the version submitted.
This work was supported by the Research Fund of the National Natural Scientific Foundation of China (81100305, 81470876 and 81270527), Natural Science Foundation of Heilongjiang Province of China (QC2013C094, LC2018037), Chen Xiaoping Foundation for the Development of Science and Technology of Hubei Province (CXPJJH11900001-2019349), Outstanding Youth Training Fund from Academician Yu Weihan of Harbin Medical University (2014), and the First Affiliated Hospital of Harbin Medical University (2019L01, HYD2020JQ0007).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.892750/full#supplementary-material
Supplementary Figure 1 | CFHR4 expression levels in different cancer tissues compared to normal tissues. (A) CFHR4 expression levels in different cancer tissues compared to normal tissues in the Timer database. (B) CFHR4 expression levels in different cancer tissues compared to normal tissues in the GEPIA database. *p < 0.05, **p < 0.01, ***p < 0.001, NS, no significance.
Supplementary Figure 2 | PPI network and potential CFHR4-interacting target genes. (A) PPI networks were built using the STRING database. (B) PPI networks were built using the GeneMANIA database. (C) The intersecting genes identified by the STRING and GeneMANIA online databases are displayed in a Venn diagram. (D) Scatter plot showing the correlation between CFHR1 and CFHR4 expression. (E) Scatter plot showing the correlation between CFHR3 and CFHR4 expression. (F) Scatter plot showing the correlation between CFHR5 and CFHR4 expression. (G) Scatter plot showing the correlation between C3 and CFHR4 expression. (H) Scatter plot showing the correlation between CRP and CFHR4 expression.
Supplementary Figure 3 | The prognostic value of CFHR4 in HCC. (A–C) OS, DSS and PFI survival curves for white, black or African–American patients with HCC presenting high and low CFHR4 expression. (D-F) OS, DSS and PFI survival curves for patients with HCC aged > 60 years presenting with high and low CFHR4 expression. (G) OS curves for patients with stage T3 and T4 HCC presenting with high and low CFHR4 expression. (H) OS curves for patients with pathological stage III and IV HCC presenting with high and low CFHR4 expression.
Supplementary Table 1 | Characteristics of patients with HCC in TCGA.
Supplementary Table 2 | CFHR4-related DEGs.
Supplementary Table 3 | GO enrichment analysis of CFHR4.
Supplementary Table 4 | GSEA enrichment analysis of CFHR4.
Supplementary Table 5 | PPI network of CFHR4.
Supplementary Table 6 | OS of patients with HCC based on prognostic covariates.
Supplementary Table 7 | DSS patients with HCC based on prognostic covariates.
Supplementary Table 8 | PFI of patients with HCC based on prognostic covariates.
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin (2021) 71(3):209–49. doi: 10.3322/caac.21660
2. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, et al. Cancer Statistics in China, 2015. CA Cancer J Clin (2016) 66(2):115–32. doi: 10.3322/caac.21338
3. Forner A, Reig M, Bruix J. Hepatocellular Carcinoma. Lancet (2018) 391(10127):1301–14. doi: 10.1016/S0140-6736(18)30010-2
4. Singal AG, Lampertico P, Nahon P. Epidemiology and Surveillance for Hepatocellular Carcinoma: New Trends. J Hepatol (2020) 72(2):250–61. doi: 10.1016/j.jhep.2019.08.025
5. Wang C, Dong L, Li X, Li Y, Zhang B, Wu H, et al. The PGC1alpha/NRF1-MPC1 Axis Suppresses Tumor Progression and Enhances the Sensitivity to Sorafenib/Doxorubicin Treatment in Hepatocellular Carcinoma. Free Radic Biol Med (2021) 163:141–52. doi: 10.1016/j.freeradbiomed.2020.11.035
6. Reis ES, Mastellos DC, Ricklin D, Mantovani A, Lambris JD. Complement in Cancer: Untangling an Intricate Relationship. Nat Rev Immunol (2018) 18(1):5–18. doi: 10.1038/nri.2017.97
7. Yarmoska SK, Alawieh AM, Tomlinson S, Hoang KB. Modulation of the Complement System by Neoplastic Disease of the Central Nervous System. Front Immunol (2021) 12:689435. doi: 10.3389/fimmu.2021.689435
8. Poppelaars F, Goicoechea de Jorge E, Jongerius I, Baeumner AJ, Steiner MS, Jozsi M, et al. A Family Affair: Addressing the Challenges of Factor H and the Related Proteins. Front Immunol (2021) 12:660194. doi: 10.3389/fimmu.2021.660194
9. Skerka C, Chen Q, Fremeaux-Bacchi V, Roumenina LT. Complement Factor H Related Proteins (CFHRs). Mol Immunol (2013) 56(3):170–80. doi: 10.1016/j.molimm.2013.06.001
10. Lores-Motta L, Paun CC, Corominas J, Pauper M, Geerlings MJ, Altay L, et al. Genome-Wide Association Study Reveals Variants in CFH and CFHR4 Associated With Systemic Complement Activation: Implications in Age-Related Macular Degeneration. Ophthalmology (2018) 125(7):1064–74. doi: 10.1016/j.ophtha.2017.12.023
11. Cipriani V, Lores-Motta L, He F, Fathalla D, Tilakaratna V, McHarg S, et al. Increased Circulating Levels of Factor H-Related Protein 4 Are Strongly Associated With Age-Related Macular Degeneration. Nat Commun (2020) 11(1):778. doi: 10.1038/s41467-020-14499-3
12. Zhao J, Wu H, Khosravi M, Cui H, Qian X, Kelly JA, et al. Association of Genetic Variants in Complement Factor H and Factor H-Related Genes With Systemic Lupus Erythematosus Susceptibility. PLoS Genet (2011) 7(5):e1002079. doi: 10.1371/journal.pgen.1002079
13. Zipfel PF, Wiech T, Stea ED, Skerka C. CFHR Gene Variations Provide Insights in the Pathogenesis of the Kidney Diseases Atypical Hemolytic Uremic Syndrome and C3 Glomerulopathy. J Am Soc Nephrol (2020) 31(2):241–56. doi: 10.1681/ASN.2019050515
14. Moore I, Strain L, Pappworth I, Kavanagh D, Barlow PN, Herbert AP, et al. Association of Factor H Autoantibodies With Deletions of CFHR1, CFHR3, CFHR4, and With Mutations in CFH, CFI, CD46, and C3 in Patients With Atypical Hemolytic Uremic Syndrome. Blood (2010) 115(2):379–87. doi: 10.1182/blood-2009-05-221549
15. Wang P, Wang X, Zheng L, Zhuang C. Gene Signatures and Prognostic Values of M6a Regulators in Hepatocellular Carcinoma. Front Genet (2020) 11:540186. doi: 10.3389/fgene.2020.540186
16. Yang Y, Hsu PJ, Chen YS, Yang YG. Dynamic Transcriptomic M(6)A Decoration: Writers, Erasers, Readers and Functions in RNA Metabolism. Cell Res (2018) 28(6):616–24. doi: 10.1038/s41422-018-0040-8
17. Zaccara S, Ries RJ, Jaffrey SR. Reading, Writing and Erasing mRNA Methylation. Nat Rev Mol Cell Biol (2019) 20(10):608–24. doi: 10.1038/s41580-019-0168-5
18. Pan Y, Xiao K, Li Y, Li Y, Liu Q. RNA N6-Methyladenosine Regulator-Mediated Methylation Modifications Pattern and Immune Infiltration Features in Glioblastoma. Front Oncol (2021) 11:632934. doi: 10.3389/fonc.2021.632934
19. Zhang F, Luo BH, Wu QH, Li QL, Yang KD. LncRNA HCG18 Upregulates TRAF4/TRAF5 to Facilitate Proliferation, Migration and EMT of Epithelial Ovarian Cancer by Targeting miR-29a/B. Mol Med (2022) 28(1):2. doi: 10.1186/s10020-021-00415-y
20. Xue ST, Zheng B, Cao SQ, Ding JC, Hu GS, Liu W, et al. Long non-Coding RNA LINC00680 Functions as a ceRNA to Promote Esophageal Squamous Cell Carcinoma Progression Through the miR-423-5p/PAK6 Axis. Mol Cancer (2022) 21(1):69. doi: 10.1186/s12943-022-01539-3
21. Li D, Xu M, Wang Z, Huang P, Huang C, Chen Z, et al. The EMT-Induced lncRNA NR2F1-AS1 Positively Modulates NR2F1 Expression and Drives Gastric Cancer via miR-29a-3p/VAMP7 Axis. Cell Death Dis (2022) 13(1):84. doi: 10.1038/s41419-022-04540-2
22. Love MI, Huber W, Anders S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data With Deseq2. Genome Biol (2014) 15(12):550. doi: 10.1186/s13059-014-0550-8
23. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc Natl Acad Sci USA (2005) 102(43):15545–50. doi: 10.1073/pnas.0506580102
24. Yu G, Wang LG, Han Y, He QY. Clusterprofiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS (2012) 16(5):284–7. doi: 10.1089/omi.2011.0118
25. Hanzelmann S, Castelo R, Guinney J. GSVA: Gene Set Variation Analysis for Microarray and RNA-Seq Data. BMC Bioinf (2013) 14:7. doi: 10.1186/1471-2105-14-7
26. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, et al. Spatiotemporal Dynamics of Intratumoral Immune Cells Reveal the Immune Landscape in Human Cancer. Immunity (2013) 39(4):782–95. doi: 10.1016/j.immuni.2013.10.003
27. Chen Y, Peng C, Chen J, Chen D, Yang B, He B, et al. WTAP Facilitates Progression of Hepatocellular Carcinoma via M6a-HuR-Dependent Epigenetic Silencing of ETS1. Mol Cancer (2019) 18(1):127. doi: 10.1186/s12943-019-1053-8
28. Pu J, Wang J, Qin Z, Wang A, Zhang Y, Wu X, et al. IGF2BP2 Promotes Liver Cancer Growth Through an M6a-FEN1-Dependent Mechanism. Front Oncol (2020) 10:578816. doi: 10.3389/fonc.2020.578816
29. Zhang C, Huang S, Zhuang H, Ruan S, Zhou Z, Huang K, et al. YTHDF2 Promotes the Liver Cancer Stem Cell Phenotype and Cancer Metastasis by Regulating OCT4 Expression via M6a RNA Methylation. Oncogene (2020) 39(23):4507–18. doi: 10.1038/s41388-020-1303-7
30. Irmscher S, Zipfel SLH, Halder LD, Ivanov L, Gonzalez-Delgado A, Waldeyer C, et al. Factor H-Related Protein 1 (FHR-1) Is Associated With Atherosclerotic Cardiovascular Disease. Sci Rep (2021) 11(1):22511. doi: 10.1038/s41598-021-02011-w
31. Cipriani V, Tierney A, Griffiths JR, Zuber V, Sergouniotis PI, Yates JRW, et al. Beyond Factor H: The Impact of Genetic-Risk Variants for Age-Related Macular Degeneration on Circulating Factor-H-Like 1 and Factor-H-Related Protein Concentrations. Am J Hum Genet (2021) 108(8):1385–400. doi: 10.1016/j.ajhg.2021.05.015
32. Zuber J, Frimat M, Caillard S, Kamar N, Gatault P, Petitprez F, et al. Use of Highly Individualized Complement Blockade Has Revolutionized Clinical Outcomes After Kidney Transplantation and Renal Epidemiology of Atypical Hemolytic Uremic Syndrome. J Am Soc Nephrol (2019) 30(12):2449–63. doi: 10.1681/ASN.2019040331
33. Riihila PM, Nissinen LM, Ala-Aho R, Kallajoki M, Grenman R, Meri S, et al. Complement Factor H: A Biomarker for Progression of Cutaneous Squamous Cell Carcinoma. J Invest Dermatol (2014) 134(2):498–506. doi: 10.1038/jid.2013.346
34. Fan WL, Yang LY, Hsieh JC, Lin TC, Lu MJ, Liao CT. Prognostic Genetic Biomarkers Based on Oncogenic Signaling Pathways for Outcome Prediction in Patients With Oral Cavity Squamous Cell Carcinoma. Cancers (Basel) (2021) 13(11):2709. doi: 10.3390/cancers13112709
35. Zhu H, Li Q, Zhao Y, Peng H, Guo L, Zhu J, et al. Vaccinia-Related Kinase 2 Drives Pancreatic Cancer Progression by Protecting Plk1 From Chfr-Mediated Degradation. Oncogene (2021) 40(28):4663–74. doi: 10.1038/s41388-021-01893-4
36. Mok WC, Wasser S, Tan T, Lim SG. Polo-Like Kinase 1, a New Therapeutic Target in Hepatocellular Carcinoma. World J Gastroenterol (2012) 18(27):3527–36. doi: 10.3748/wjg.v18.i27.3527
37. Dimri M, Satyanarayana A. Molecular Signaling Pathways and Therapeutic Targets in Hepatocellular Carcinoma. Cancers (Basel) (2020) 12(2):491. doi: 10.3390/cancers12020491
38. Sheng H, Huang Y, Xiao Y, Zhu Z, Shen M, Zhou P, et al. ATR Inhibitor AZD6738 Enhances the Antitumor Activity of Radiotherapy and Immune Checkpoint Inhibitors by Potentiating the Tumor Immune Microenvironment in Hepatocellular Carcinoma. J Immunother Cancer (2020) 8(1):e000340. doi: 10.1136/jitc-2019-000340
39. Lei Y, Wang S, Liu J, Yan W, Han P, Tian D. Identification of MCM Family as Potential Therapeutic and Prognostic Targets for Hepatocellular Carcinoma Based on Bioinformatics and Experiments. Life Sci (2021) 272:119227. doi: 10.1016/j.lfs.2021.119227
40. Liu Z, Mo H, Liu R, Niu Y, Chen T, Xu Q, et al. Matrix Stiffness Modulates Hepatic Stellate Cell Activation Into Tumor-Promoting Myofibroblasts via E2F3-Dependent Signaling and Regulates Malignant Progression. Cell Death Dis (2021) 12(12):1134. doi: 10.1038/s41419-021-04418-9
41. Domingues P, Gonzalez-Tablas M, Otero A, Pascual D, Miranda D, Ruiz L, et al. Tumor Infiltrating Immune Cells in Gliomas and Meningiomas. Brain Behav Immun (2016) 53:1–15. doi: 10.1016/j.bbi.2015.07.019
42. Sun H, Liu L, Huang Q, Liu H, Huang M, Wang J, et al. Accumulation of Tumor-Infiltrating CD49a(+) NK Cells Correlates With Poor Prognosis for Human Hepatocellular Carcinoma. Cancer Immunol Res (2019) 7(9):1535–46. doi: 10.1158/2326-6066.CIR-18-0757
43. Stockinger B, Veldhoen M. Differentiation and Function of Th17 T Cells. Curr Opin Immunol (2007) 19(3):281–6. doi: 10.1016/j.coi.2007.04.005
44. Lee YK, Turner H, Maynard CL, Oliver JR, Chen D, Elson CO, et al. Late Developmental Plasticity in the T Helper 17 Lineage. Immunity (2009) 30(1):92–107. doi: 10.1016/j.immuni.2008.11.005
45. Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. Reciprocal Developmental Pathways for the Generation of Pathogenic Effector TH17 and Regulatory T Cells. Nature (2006) 441(7090):235–8. doi: 10.1038/nature04753
46. Chatterjee S, Daenthanasanmak A, Chakraborty P, Wyatt MW, Dhar P, Selvam SP, et al. CD38-NAD(+)Axis Regulates Immunotherapeutic Anti-Tumor T Cell Response. Cell Metab (2018) 27(1):85–100.e8. doi: 10.1016/j.cmet.2017.10.006
47. Karpisheh V, Ahmadi M, Abbaszadeh-Goudarzi K, Mohammadpour Saray M, Barshidi A, Mohammadi H, et al. The Role of Th17 Cells in the Pathogenesis and Treatment of Breast Cancer. Cancer Cell Int (2022) 22(1):108. doi: 10.1186/s12935-022-02528-8
48. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, Sancho D. Dendritic Cells in Cancer Immunology and Immunotherapy. Nat Rev Immunol (2020) 20(1):7–24. doi: 10.1038/s41577-019-0210-z
49. Martinek J, Wu TC, Cadena D, Banchereau J, Palucka K. Interplay Between Dendritic Cells and Cancer Cells. Int Rev Cell Mol Biol (2019) 348:179–215. doi: 10.1016/bs.ircmb.2019.07.008
50. Shaul ME, Fridlender ZG. Neutrophils as Active Regulators of the Immune System in the Tumor Microenvironment. J Leukoc Biol (2017) 102(2):343–9. doi: 10.1189/jlb.5MR1216-508R
51. Roybal KT, Williams JZ, Morsut L, Rupp LJ, Kolinko I, Choe JH, et al. Engineering T Cells With Customized Therapeutic Response Programs Using Synthetic Notch Receptors. Cell (2016) 167(2):419–32.e16. doi: 10.1016/j.cell.2016.09.011
52. Orecchioni M, Bedognetti D, Newman L, Fuoco C, Spada F, Hendrickx W, et al. Single-Cell Mass Cytometry and Transcriptome Profiling Reveal the Impact of Graphene on Human Immune Cells. Nat Commun (2017) 8(1):1109. doi: 10.1038/s41467-017-01015-3
53. Mantovani A, Romero P, Palucka AK, Marincola FM. Tumour Immunity: Effector Response to Tumour and Role of the Microenvironment. Lancet (2008) 371(9614):771–83. doi: 10.1016/S0140-6736(08)60241-X
54. Tonetti CR, de Souza-Araujo CN, Yoshida A, da Silva RF, Alves PCM, Mazzola TN, et al. Ovarian Cancer-Associated Ascites Have High Proportions of Cytokine-Responsive CD56bright NK Cells. Cells (2021) 10(7):1702. doi: 10.3390/cells10071702
55. Nie Y, Liu D, Yang W, Li Y, Zhang L, Cheng X, et al. Increased Expression of TIGIT and KLRG1 Correlates With Impaired CD56(bright) NK Cell Immunity in HPV16-Related Cervical Intraepithelial Neoplasia. Virol J (2022) 19(1):68. doi: 10.1186/s12985-022-01776-4
56. Duan J, Lv G, Zhu N, Chen X, Shao Y, Liu Y, et al. Multidimensional Profiling Depicts Infiltrating Immune Cell Heterogeneity in the Tumor Microenvironment of Stage IA Non-Small Cell Lung Cancer. Thorac Cancer (2022) 13(7):947–55. doi: 10.1111/1759-7714.14329
57. Chen Z, Wang N, Yao Y, Yu D. Context-Dependent Regulation of Follicular Helper T Cell Survival. Trends Immunol (2022) 43(4):309–21. doi: 10.1016/j.it.2022.02.002
58. Walker LSK. The Link Between Circulating Follicular Helper T Cells and Autoimmunity. Nat Rev Immunol (2022) 11:1–9. doi: 10.1038/s41577-022-00693-5
59. Galdiero MR, Varricchi G, Marone G. The Immune Network in Thyroid Cancer. Oncoimmunology (2016) 5(6):e1168556. doi: 10.1080/2162402X.2016.1168556
60. Bian J, Lin J, Long J, Yang X, Yang X, Lu X, et al. T Lymphocytes in Hepatocellular Carcinoma Immune Microenvironment: Insights Into Human Immunology and Immunotherapy. Am J Cancer Res (2020) 10(12):4585–606.
61. Chen H, Gao S, Liu W, Wong CC, Wu J, Wu J, et al. RNA N(6)-Methyladenosine Methyltransferase METTL3 Facilitates Colorectal Cancer by Activating the M(6)A-GLUT1-Mtorc1 Axis and Is a Therapeutic Target. Gastroenterology (2021) 160(4):1284–300.e16. doi: 10.1053/j.gastro.2020.11.013
62. Zhang J, Hu K, Yang YQ, Wang Y, Zheng YF, Jin Y, et al. LIN28B-AS1-IGF2BP1 Binding Promotes Hepatocellular Carcinoma Cell Progression. Cell Death Dis (2020) 11(9):741. doi: 10.1038/s41419-020-02967-z
63. Jiang W, Cheng X, Wang T, Song X, Zheng Y, Wang L. LINC00467 Promotes Cell Proliferation and Metastasis by Binding With IGF2BP3 to Enhance the mRNA Stability of TRAF5 in Hepatocellular Carcinoma. J Gene Med (2020) 22(3):e3134. doi: 10.1002/jgm.3134
64. Su T, Huang M, Liao J, Lin S, Yu P, Yang J, et al. Insufficient Radiofrequency Ablation Promotes Hepatocellular Carcinoma Metastasis Through N6-Methyladenosine mRNA Methylation-Dependent Mechanism. Hepatology (2021) 74(3):1339–56. doi: 10.1002/hep.31766
65. Song Y, He S, Ma X, Zhang M, Zhuang J, Wang G, et al. RBMX Contributes to Hepatocellular Carcinoma Progression and Sorafenib Resistance by Specifically Binding and Stabilizing BLACAT1. Am J Cancer Res (2020) 10(11):3644–65.
66. Cai X, Chen Y, Man D, Yang B, Feng X, Zhang D, et al. RBM15 Promotes Hepatocellular Carcinoma Progression by Regulating N6-Methyladenosine Modification of YES1 mRNA in an IGF2BP1-Dependent Manner. Cell Death Discov (2021) 7(1):315. doi: 10.1038/s41420-021-00703-w
Keywords: CFHR4, prognosis, biomarker, immune Infiltrate, hepatocellular carcinoma
Citation: Yu H, Wang C, Ke S, Bai M, Xu Y, Lu S, Feng Z, Qian B, Xu Y, Zhou M, Li Z, Yin B, Li X, Hua Y, Zhou Y, Pan S, Fu Y and Ma Y (2022) Identification of CFHR4 as a Potential Prognosis Biomarker Associated With lmmune Infiltrates in Hepatocellular Carcinoma. Front. Immunol. 13:892750. doi: 10.3389/fimmu.2022.892750
Received: 09 March 2022; Accepted: 16 May 2022;
Published: 22 June 2022.
Edited by:
Xuesong Gu, Beth Israel Deaconess Medical Center and Harvard Medical School, United StatesReviewed by:
Simon John Clark, University of Tübingen, GermanyCopyright © 2022 Yu, Wang, Ke, Bai, Xu, Lu, Feng, Qian, Xu, Zhou, Li, Yin, Li, Hua, Zhou, Pan, Fu and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yao Fu, ZHJmdXlhb0AxNjMuY29t; Yong Ma, bWF5b25nQGVtcy5ocmJtdS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.