
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 22 April 2022
Sec. Viral Immunology
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.878630
This article is part of the Research Topic Perinatal HIV infection and Immunologic Pointers for the Path to a Cure View all 8 articles
 Laura Tarancon-Diez1,2†
Laura Tarancon-Diez1,2† Irene Consuegra1,2†
Irene Consuegra1,2† Elena Vazquez-Alejo1,2
Elena Vazquez-Alejo1,2 Ricardo Ramos-Ruiz3
Ricardo Ramos-Ruiz3 José Tomás Ramos4
José Tomás Ramos4 María Luisa Navarro5,6
María Luisa Navarro5,6 Mª Ángeles Muñoz-Fernández1,2*
Mª Ángeles Muñoz-Fernández1,2*Early antiretroviral treatment (ART) in vertically acquired HIV-1-infection is associated with a rapid viral suppression, small HIV-1 reservoir, reduced morbimortality and preserved immune functions. We investigated the miRNA profile from vertically acquired HIV-1-infected young adults based on ART initiation delay and its association with the immune system activation. Using a microRNA panel and multiparametric flow cytometry, miRNome profile obtained from peripheral blood mononuclear cells and its association with adaptive and innate immune components were studied on vertically HIV-1-infected young adults who started ART early (EARLY, 0-53 weeks after birth) and later (LATE, 120-300 weeks). miR-1248 and miR-155-5p, were significantly upregulated in EARLY group compared with LATE group, while miR-501-3p, miR-548d-5p, miR-18a-3p and miR-296-5p were significantly downregulated in EARLY treated group of patients. Strong correlations were obtained between miRNAs levels and soluble biochemical biomarkers and immunological parameters including CD4 T-cell count and maturation by CD69 expression on CD4 T-cells and activation by HLA-DR on CD16high NK cell subsets for miR-1248 and miR-155-5p. In this preliminary study, a distinct miRNA signature discriminates early treated HIV-1-infected young adults. The role of those miRNAs target genes in the modulation of HIV-1 replication and latency may reveal new host signaling pathways that could be manipulated in antiviral strategies. Correlations between miRNAs levels and inflammatory and immunological markers highlight those miRNAs as potential biomarkers for immune inflammation and activation in HIV-1-infected young adults who initiated a late ART.
Antiretroviral treatment (ART) implementation has greatly reduced mother to child HIV-1 transmission. Despite this, at the end of 2019, 1.7 million people living with HIV-1 were children aged 0 to 14 years old (1). WHO guidelines recommend that ART should be initiated within 1 year of age in all vertically acquired HIV-1 infected children as it is well known that the earlier the ART is initiated after infection, the greater the reduction in the reservoir and disease progression, not only in HIV-1 infected children but also in adults (2–4). Several observations suggested that a late timing of treatment initiation in HIV-1 infected children is associated with the induction of ineffective HIV-specific immune responses (5, 6), altered immune phenotype (7) and a distinct transcriptional signature and protein profile, showing upregulation of immune activation pathways (5, 8), indicated a proinflammatory state that may potentially trigger premature non-AIDS events including atherosclerotic and metabolic diseases. It is still unclear whether the effects of the infection on an immune system under development are reversible later in life, as first children infected since birth are now reaching adulthood. Immunological data on this population remain scarce, but our group along with others has described a premature immune aging profile associated with HIV-1 infection (7, 9).
MicroRNAs (miRNAs) are small noncoding RNAs implicated in the development and progression of multiple diseases and have recently been proposed as therapeutic targets in cancer, cardiovascular and metabolic diseases and infections due to their crucial roles in processes of cell maturation, proliferation, migration, differentiation and function in the immune system (10, 11). Focusing on HIV-1 infection, there is a strong effort centered on determining how HIV-1 can differentially modulate several miRNAs that potentially regulate host cellular pathways such as cell cycle, apoptosis, T-cell signalling and cytokine response (12, 13). Because of that, cellular miRNAs could possibly be involved in establishing HIV-1 latency. Recent strategies for clearing viral reservoir have resulted from studies on the mechanism of how miRNAs influence viral protein expression (14). Up to the moment, most findings related to miRNA in HIV-1-infection have been obtained from patients infected during adulthood focussing on the disease progression and there is no information on the miRNA profile related to an early ART initiation in vertically infected HIV-1-patients that reached adult status. Thus, the objective of the present preliminary study was to investigate the miRNA expression profile from vertically acquired HIV-1-infected young adults based on ART initiation delay as well as its association with inflammatory biomarkers and immune system activation and maturation state.
This cross-sectional study is based on young adults with vertically transmitted HIV-1 infection from the Paediatric Spanish AIDS Research Network Cohort (coRISpe). All participants were Caucasian. Eighteen patients, all of them on suppressive ART treatment, were included based on sample availability and the following inclusion criteria: 1) patients had evidence of virological suppression within the first year after ART initiation; 2) had subsequent maintenance of viral control (≤20 copies/mL) for at least five years before sampling; 3) were not coinfected by Hepatitis C Virus (HCV). Cryopreserved peripheral blood mononuclear cells (PBMCs) and associated clinical data were provided by the Spanish HIV Hospital General Universitario Gregorio Marañón BioBank (15) and by coRISpe (16) respectively. Patients were classified in two groups according to ART timing initiation in those who initiated ART early (EARLY, 0-53 weeks, n=7) and those who initiated ART late (LATE, 120-300 weeks, n=11). As reference group, samples of fresh whole blood from volunteers non-HIV-1-infected Healthy Donors (HD, n=6) paired by sex and age were collected in EDTA tubes and PBMCs were immediately isolated by Ficoll-Paque density gradient centrifugation and stored at -20°C until RNA extraction.
The study was approved by the ethic committee of Hospital General Universitario Gregorio Marañón (HGUGM) in Madrid (protocol miRNA-Ped-HIV_16, approved on March 21, 2017, acta06/2017). Written informed consent was obtained from all participants before inclusion in coRISpe and from all volunteers before inclusion in the study. Data was collected from October 2020 to March 2021.
Immunophenotyping of T lymphocytes and Natural Killer (NK) cells using multiparametric flow cytometry was performed following this hierarchy order according to sample availability. In future analysis, the number of patients will vary due to this limitation. For T lymphocyte immunophenotyping, two flow cytometer panels were designed. For the first panel PBMCs were thawed, washed and stained with LIVE/DEAD fixable Aqua Blue Dead Cell Stain (Life Technologies, CA, USA) for viability, and the surface antibodies CD56-CD19-CD14-BV510 (CD56 clone NCAM16.2; CD19 clone SJ25C1 and CD14 clone MOP9), CD3-PerCP-Cy5.5 (clone SK7) (BD Biosciences, San Diego, CA) and CD8-PB (clone RPA-T8; Biolegend, San Diego, CA) for lineage, CD45RA-ECD (clone 2H4), IL-7 receptor, CD127-PeCy7 (clone R34.34), HLA-DR-APC (clone GRB-1) (Beckman Coulter), CD38-FITC (clone HB7), CD69-APC-R700 (clone FN50) (BD Biosciences) for maturation, survival and activation.
The second T lymphocyte panel, in addition to LIVE/DEAD fixable Aqua Blue Dead Cell Stain for viability, CD56-CD19-CD14-BV510, (CD56 clone NCAM16.2; CD19 clone SJ25C1 and CD14 clone MOP9), CD3-APC-Cy7 (clone SK7) for lineage and CD45RA-ECD (clone 2H4; Beckman Coulter) and CD27-PerCP-Cy5.5 (clone M-T271, BD, Biosciences) for maturation, it also included CD4-APC-R700 (clone RPA-T4, BD Biosciences) for lineage, CD57-FITC (clone NC1; Beckman Coulter) for senescence and TIM-3-PE (clone 7D3), PD-1 (clone EH12.1) (BD Biosciences), LAG-3-Pe-Cy7 (clone 7H2C65) and TIGIT-AF647 (clone A15153G) (Biolegend), for exhaustion. Lymphocytes were defined as viable cells having low forward/side scatter and expressing CD3, and/or no CD8/CD4, but not CD19, CD14 and CD56. Isotypes controls were included for CD57, TIM-3, LAG-3, TIGIT and PD-1.
The NK immunophenotyping included: LIVE/DEAD fixable Aqua Blue Dead Cell Stain for viability, CD3-CD19-CD14-BV510 (CD3 clone SK7; CD19 clone SJ25C1 and CD14 clone MOP9), CD56-APC-Cy7 (clone NCAM16.2), CD16-PerCP-Cy5.5 (clone 3G8) (BD Biosciences) for NK subsets identification and CD57-FITC (clone NC1) for maturation, HLA-DR-APC (clone GRB-1) (Beckman Coulter) for activation, the constitutively expressed receptor, TIM-3-PE (clone 7D3) involved in NK cell cytokine secretion, the C-type lectin-like activating receptor NKG2D-PECF594 (clone 1D11), CD69-APC-R700 (clone FN50) for activation, the inhibiting receptor NKG2A-BV421 (clone 131411) (BD Biosciences) and the natural cytotoxicity family activating receptor NKp30-PE-Cy7 (clone P30-15; Biolegend). To identify NK cells, viable cells, negative for CD3, CD14 and CD19 were classified also in three subsets based on the expression of CD56 (CD56neg, CD56high and CD56dim) and CD16 that define the NK cytotoxic subset (CD16high) Isotype controls were included for TIM-3, NKG2D, NKG2A, NKp30 and CD69.
Acquisition was carried out in a Galios flow cytometer (Beckman Coulter). Before acquisition, cells were fixed with 4% paraformaldehyde. At least 1 million events were acquired for each condition. FlowJo software (v10.6.1) was used for data analysis.
Total RNA isolation was performed with mirVana miRNA isolation kit (Ambion, Huntingdon, UK), following the manufacturer’s instructions and RNA concentrations were estimated with Nanodrop ND-1000 (Thermo Scientific, Waltham, MA, USA) followed by RNA analysis of quality and integrity based on RIN values, determined using Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). TaqMan™ Advanced miRNA cDNA Synthesis Kit (Thermo Fisher) was used for reverse-transcription of isolated RNA. MiRNA gene expression profile was then assessed by using the 754 panel TaqMan™ OpenArray™ Human Advanced MicroRNA Panel and run in the Open Array block of a QuantStudio™ 12K Flex equipment (ThermoFisher).
The RQ module (version4.3) of the apps.thermofisher cloud (apps.thermofisher.com) was used to generate Crt values (herein referred as Cq) and to calculate deltaCq (∆Cq) data using Global Normalization. Cq values ≥ 32 were considered negative and microRNAs which did not show expression above this level in at least 12 out of the 24 samples were discarded. Therefore, 142 miRNAs were detected as positive in this study. MiRNA expression profile analysis from six healthy donors non-HIV-infected subjects, paired by sex and age with HIV-1 participants, was also included. Reactions and data analysis were carried out at Genomics Core Facility, Parque Científico de Madrid, Spain.
Shapiro-Wilk normality test was used for variables normality and they were non-normally distributed and then considered as non-parametric. Differences between categorical and continuous values were determined by the Chi-square test and Mann-Whitney U test, respectively. Correlations between variables were assessed using the Spearman rank test. p values <0.05 were considered statistically significant.
The statistical software used included SPSS 23.0 package (IBM, Madrid, Spain). Graphs were generated with Prism, version 9.0 (GraphPad Software, Inc.).
Clinical and demographic characteristics of patients are shown in Table 1. Comparing groups, statistical difference was only observed in age at sampling, being LATE group older than EARLY group (24 [22-25] and 21 [20-23] years respectively, p=0.035). No differences were found in the clinical and immunological parameters including CD4+ and CD8+ percentage and cell counts, CD4+/CD8+ ratio, nadir CD4+ cell count, time since HIV-1 diagnosis, time under ART and under virological control.
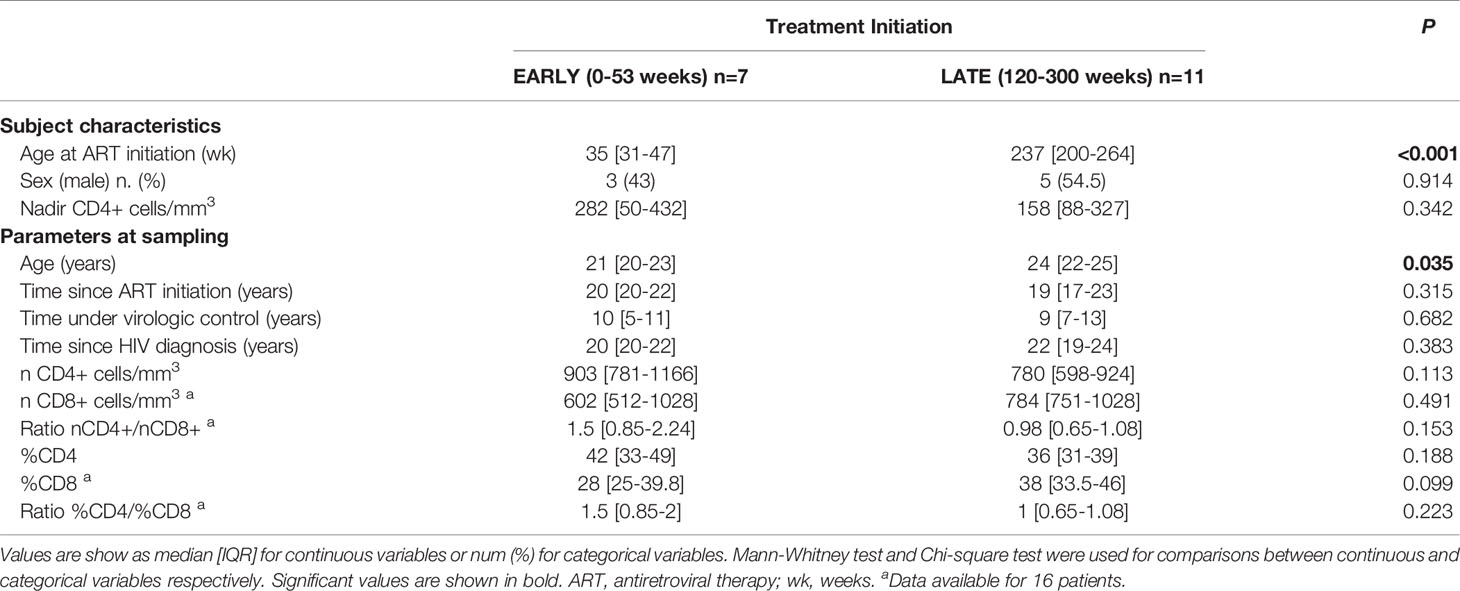
Table 1 Study cohort characteristics.
Biochemical and inflammatory biomarkers including plasma levels of glucose, GPT (ALT), GOT (AST), bilirubin, total cholesterol, triglycerides, HDL, LDL, LDL/HDL ratio and creatinine at sampling are showed in Table 2. EARLY group showed increased levels of triglycerides (p=0.044) and a trend to low frequency of HDL (p=0.097).
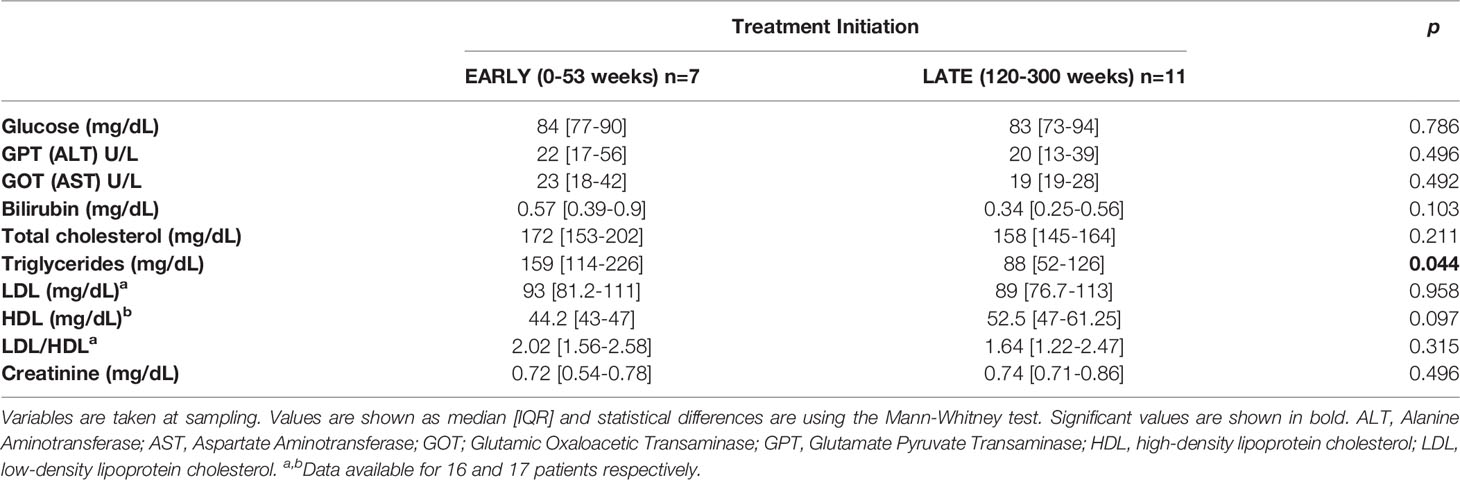
Table 2 Biochemical and inflammatory biomarkers.
The miRNA profiling analysis in PBMCs detected and quantified 142 miRNAs. Comparisons between HD (median age of 23 years, 50% male) and EARLY and LATE treated HIV-1-infected young adults showed 34 and 28 differentially expressed miRNAs respectively (see Supplementary Table 1).
Comparing both groups of HIV-infected young adults, six miRNAs were expressed differently: two were significantly upregulated in EARLY group compared with LATE group (miR-1248 and miR-155-5p) and four (miR-501-3p, miR-548d-5p, miR-18a-3p and miR-296-5p) were significantly downregulated in EARLY treated group of patients (Figure 1). Noteworthy, one of that subset of regulated miRNAs (miR-501-3p) was also found modulated in HD group compared with both groups of HIV-infected young adults. Focusing on EARLY versus LATE comparison, for target genes identification, the public database miRTarBase was accessed because it gathers functional studies of miRNA–target interactions, which are validated experimentally (http://mirtarbase.cuhk.edu.cn/) and genes with strong evidence of interactions when linked to targets related to HIV-1 infection based on information available in scientific literature. Regulated genes involved in HIV-1 infection by miR-18a-3p, miR-296-5p and miR-155-5p are shown in Table 3 while no evidence was found for miR-1248, miR-501-3p or miR-548d-5p.
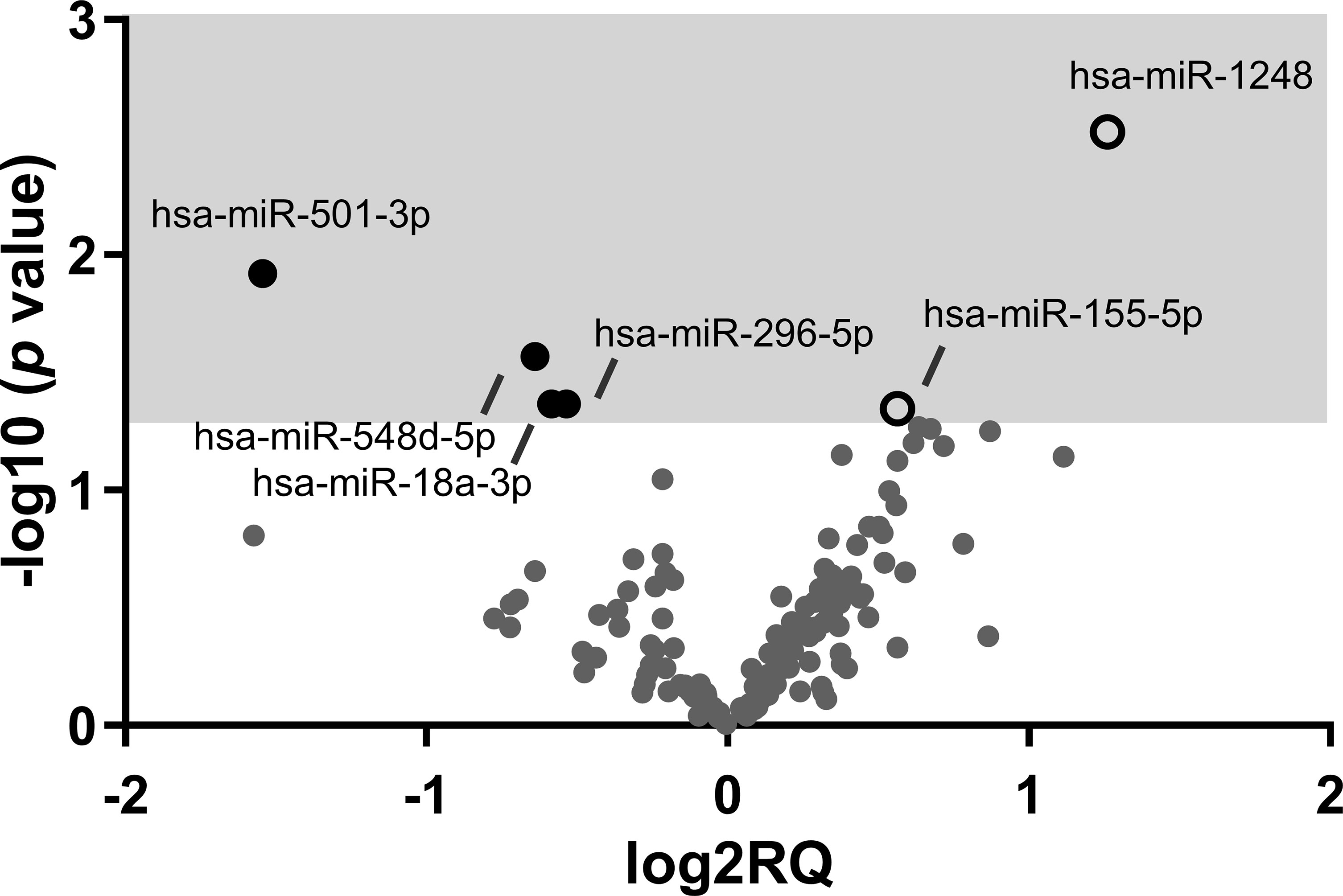
Figure 1 Volcano plot showing the results of the miRNA profile analysis comparing EARLY and LATE-treated groups of HIV-infected patients. Graph of log2 Relative Quantity ratio (log2RQ) against log10 p value significance (-log10 p value) from 142 quantified miRNAs. Open white spots represent upregulated genes and black spots represent downregulated genes on EARLY treated (n=7) compared to LATE (n=11) treated HIV-1-young adults.
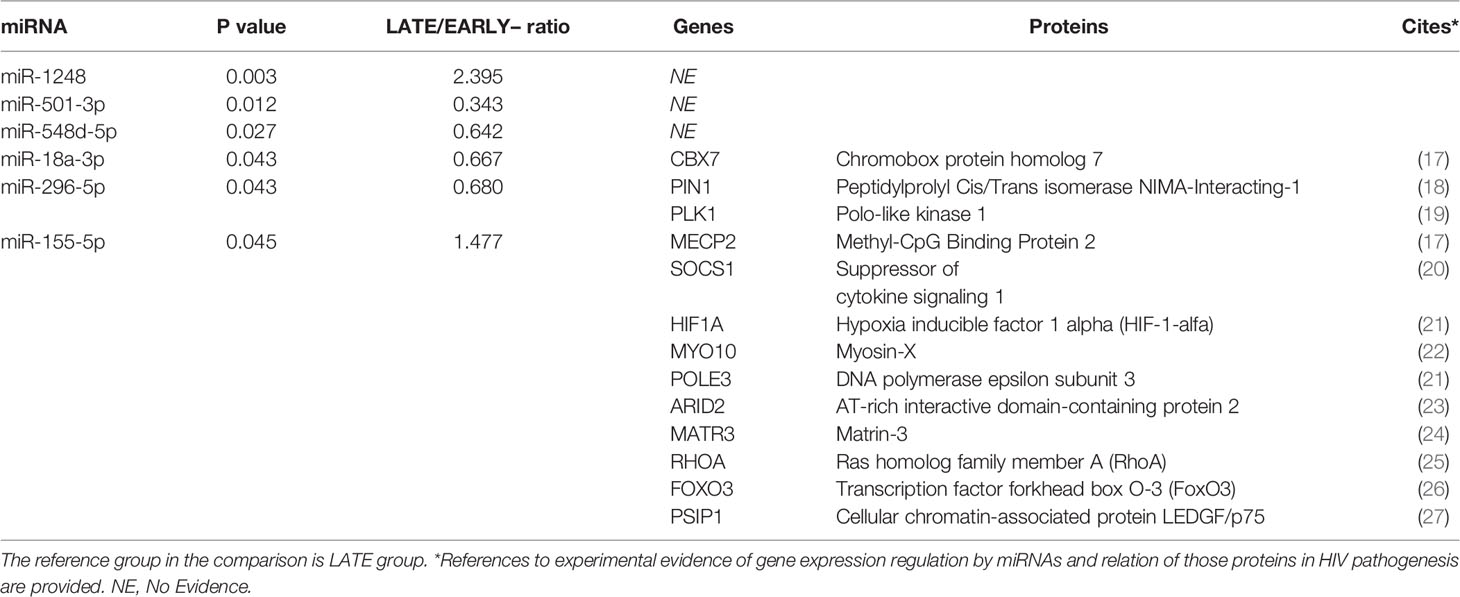
Table 3 Differentially expressed miRNAs between LATE and EARLY treated HIV-1-young adults and list of targeted genes involved in HIV immunopathogenesis.
To determine possible associations between miRNAs profile and clinical and immunological parameters on HIV-1 infected patients, the levels of six most significant miRNAs were correlated with immunological and biochemical parameters (Figure 2A). The relative levels of miR-1248, miR-501-3p and miR-155-5p were associated with CD4+ cell counts (p=0.016; ρ=0.55; p=0.058; ρ=-0.58 and p=0.028; ρ=0.51 respectively). Besides, miR-296-5p and miR-155-5p correlated directly with soluble GOT transaminase (p=0.028; ρ=0.51 and p=0.016; ρ=0.56 respectively) and concerning soluble lipoproteins, strong associations were found between HDL soluble levels with miR-1248, miR-501-3p and miR-155-5p relative levels (p=0.020; ρ=-0.56; p=0.021; ρ=0.56 and p=0.002; ρ=-0.70 respectively) and ratio LDL/HDL with miR-501-3p and miR-155-5p relative levels (p=0.030; ρ=-0.54 and p=0.042; ρ=0.54 respectively) (Figures 2B–E).
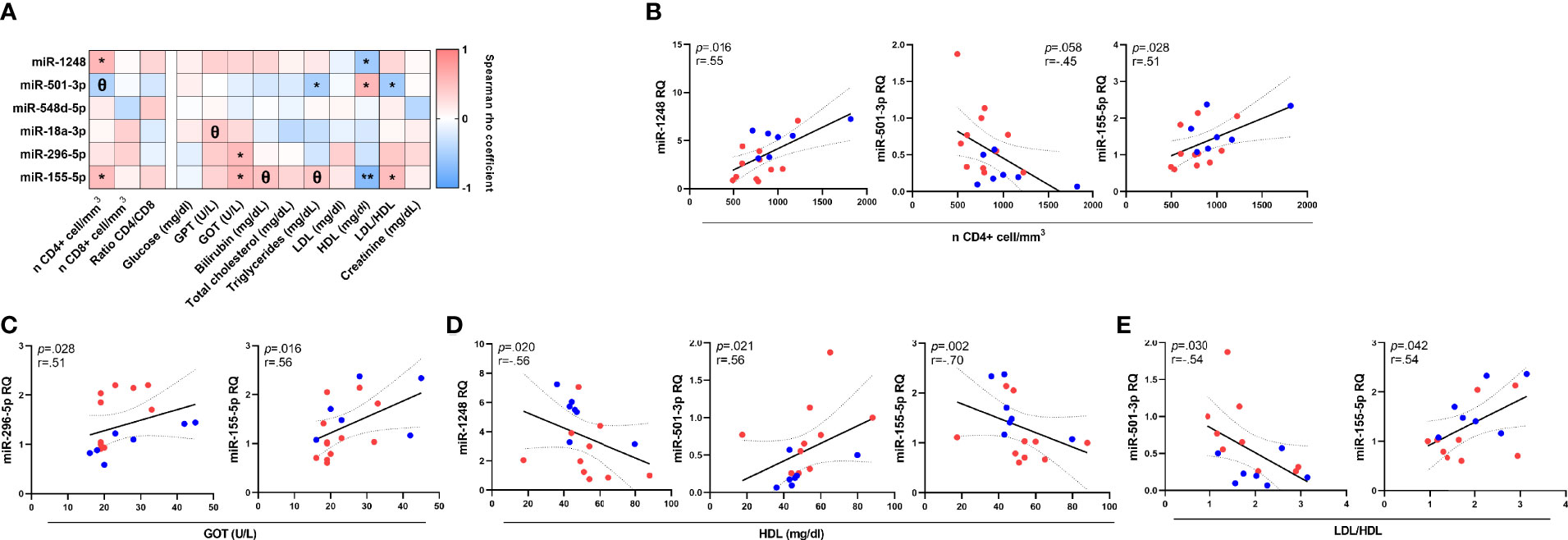
Figure 2 Associations between miRNA relative levels and immunological and biochemical parameters on HIV-1-infected young adults. Correlations of miRNAs relative quantity (RQ) with all soluble immunological and biochemical parameters (A). Significant and close to significant associations with T-CD4+ cell count (B), Glutamic Oxaloacetic Transaminase (Aspartate Aminotransferase) (GOT; AST) (C), high-density lipoprotein cholesterol (HDL) (data available for 16 patients) (D) and LDL/HDL ratio (LDL: low-density lipoprotein cholesterol) (data available for 16 patients) (E). Patients from EARLY treated group (n=7) are highlighted with blue dots and red dots represent LATE treated HIV-1-young adults (n=11). The Spearman rho correlation coefficient test was used. **p <0.01, *p <0.05, Θ0.05<p< 0.1.
CD4 and CD8 T-cell maturation subsets were defined as naïve (CD45RA+CD27+), Central Memory (CM; CD45RA-CD27+), Effector Memory (EM; CD45RA-CD27-) and Terminally Differentiated (TemRA; CD45RA+CD27-) (gate strategy in Figure 3A) and NK cells were classified also in subsets based on the expression of CD56 (CD56neg, CD56high and CD56dim) and CD16 that define the NK cytotoxic subset (CD16high) (gate strategy in Figure 3E).
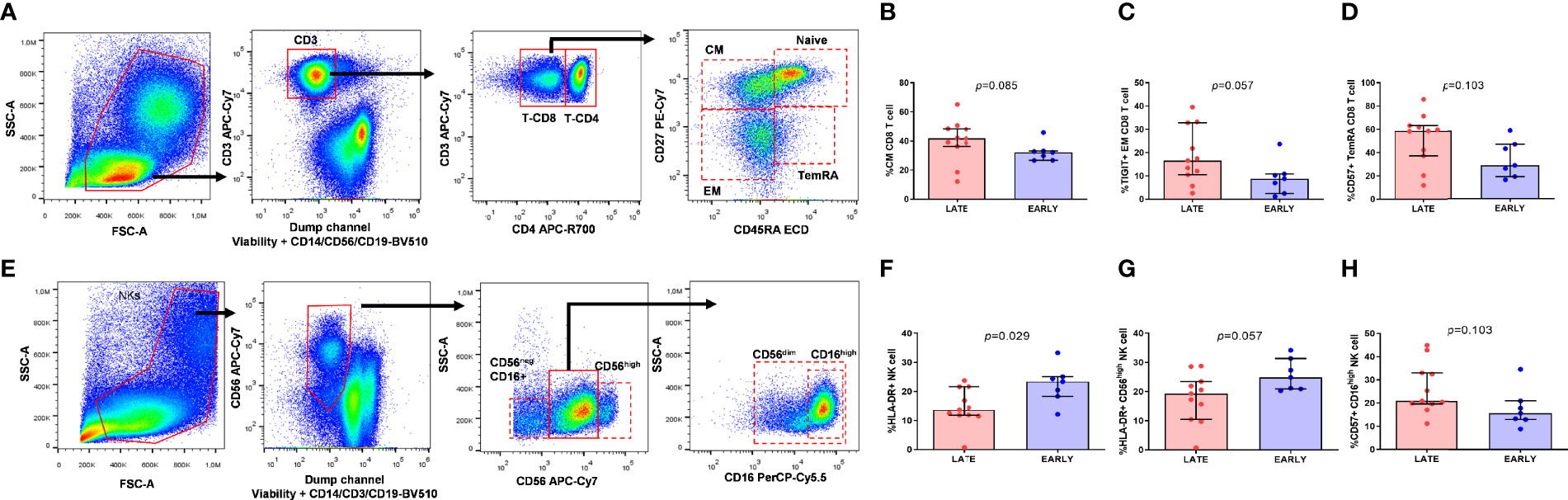
Figure 3 Memory CD8-T cell distribution and activation and exhaustion expression markers on CD8-T cell subsets and NK cell subsets from HIV-infected young adults. Gate strategy for memory subset distribution on T cell (A), lymphocytes were characterized by the patterns of forward and side scatter, followed by selection of CD3+. Central Memory (CM) (CD45RA-CD27+) CD8 T cell subset distribution (B), TIGIT expression on Effector Memory (EM) (CD45RA-CD27-) CD8 T cell subset (C) and CD57 expression on Terminally Differentiated (TemRA) (CD45RA+CD27-) CD8 T cell subset (D). Gate strategy for NK cell subsets (E) viable cells, negative for CD3, CD14 and CD19 were classified in three subsets according to the expression of CD56 and CD16. HLA-DR expression on total NK cells (F) and CD56high (G) NK cell subset and CD57 expression on CD16high NK cell subset (H) from EARLY treated group (n=7) and LATE treated HIV-1-young adults (n=11). Mann-Whitney U test was used.
Firstly, we wanted to determine if EARLY and LATE treated HIV-1 patients showed differences in subset distribution and activation, survival and exhaustion levels on T and NK cells. Only borderline statistical significance was observed, probably due to the scarce sample size, towards a higher frequency of CM CD8 T-cells and TIGIT and CD57 expression on EM and TemRA CD8 T-cells from LATE group, respectively (Figures 3B–D). Regarding NK cells, gate strategy is defined in Figure 3E, LATE group had significant lower activation levels defined by HLA-DR on total NK cells (p=0.029) and a borderline statistical significance on CD56high NK subset and higher CD57 levels on CD16high NK subset compared to EARLY group (Figures 3F–H).
We next analyzed the relationship between T and NK cell activation, survival and exhaustion levels and miRNAs expression, including those six that discriminate both groups of HIV-patients. Regarding T cells, CM CD4 T-cell subset distribution negatively correlated with miRNA-1248 relative levels (Figure 4A). Besides, expression levels of activation marker CD69 on naïve CD4 T-cells (defined as CD45RA+) was inversely correlated with miRNA-1248 relative levels, (ρ=-0.52; p=0.020) and similarly, CD69 expression levels on memory CD4 T-cells (defined as CD45RA-) also negatively correlated with miRNA-1248 and miRNA-155-5p relative levels (Figures 4B, C). No correlations were found for exhaustion markers on CD4 T-cells, neither for CD8 T-cell markers.
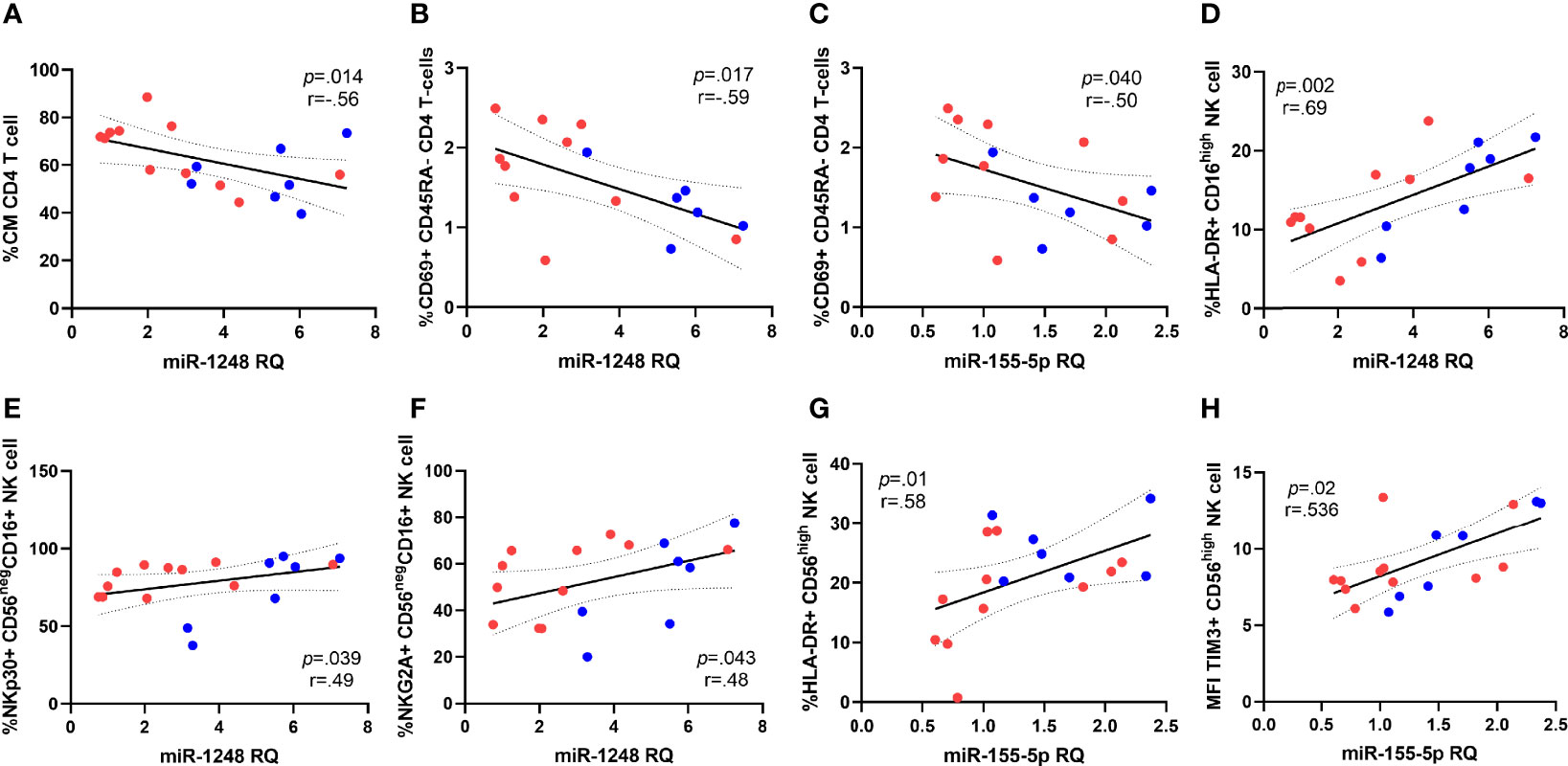
Figure 4 Correlations between miRNAs and subset distribution and activation markers expression on CD4 T cells and NK cells. Correlations of miRNA-1248 relative quantity (RQ) with CD4 Central Memory subset distribution (CM, defined as CD45RA-CD27+) (A) and CD69+ expression on CD45RA- CD4 T cells (B), data available for 16 patients. MiRNA-155-5p correlations with CD69+ expression on CD45RA- CD4 T cells (C), data available for 16 patients. Correlations of miRNA-1248 levels and expression of HLA-DR on CD16high NK subset (D) and NKp30 and NKG2A on CD56negCD16+ NK cell subset (E, F). Correlations of with miRNA-155-5p levels with HLA-DR expression on CD56high NK cells (G) and median intensity fluorescence (MFI) of TIM-3 expression on CD56high NK cell subset (H). Patients from EARLY treated group (n=7) are highlighted with blue dots and red dots represent LATE treated HIV-1-young adults (n=11). The Spearman rho correlation coefficient test was used.
Focusing on NK cell associations, miRNA-1248 levels strongly and directly correlated with the expression of the activation marker HLA-DR on total NK cells subsets (ρ=0.569; p=0.014) and that association remains significant on the cytolytic NK subset CD16high expressing HLA-DR and miRNA-1248 (Figure 4D). Besides, miRNA-1248 levels also correlated with the expression of NKp30 and NKG2A on CD56negCD16+ NK cells subset (Figures 4E, F). In the case of miRNA-155-5p, upregulated on EARLY treated group, its levels strongly and directly correlated with HLA-DR expression and MFI of TIM-3 expression on CD56high NK cell subset (Figures 4G, H). Most of those associations were lost after stratification by group probably a cause of the small number of studied cases (data not shown).
In this work, for the first time we describe a specific miRNA signature on peripheral blood mononuclear cells of vertically HIV-1-young adults who started an ART during the first year after birth compared to late treated HIV-1-young adults, with the purpose of performing a distinctive screening of miRNAs that potentially control the expression of multiple related genes in the context of HIV-1 control and future delayed progression due to their relevant role in HIV-1 cycle and in the establishment of reservoirs. Besides, the miRNome assay was completed with an in-depth phenotype characterization on innate and adaptive immune system components. The associations found between miRNAs and biochemical inflammatory biomarkers and activation and maturation markers expression highlight the potential role of those miRNAs as biomarkers for immune activation and can be considered good targets for therapies aimed at modulating immune response in HIV-1-young adults.
Our study confirms previous studies in which HIV-1 infection leads to altered miRNome compared to non-HIV-1 infected population. However, all evidences were described on HIV-1 patients infected at adulthood by sexual route among other ways of transmission and tried to find patters related to immune and viral progression (28). Up to the moment, none information had been carried out from vertically acquired HIV-1 infected group that still represents more than 150,000 new infections worldwide every year (1).
Focusing on most relevant identified miRNAs comparing EARLY and LATE treated HIV-1-young adults, miRNA-155-5p has been widely studied due to its role in the functionality of the immune system regulating the immune response and lymphocyte differentiation (29, 30). Besides, miRNA-155 has been highlighted as potential biomarker of long-term nonprogressors (LTNPs) who showed decreased levels compared to viremic naïve HIV-subjects suggesting an important role also in the control and pathogenesis of HIV-1 infection (31). Accordingly to that, but contrary to what we expected due to all well-known benefits associated to an EARLY antiretroviral therapy introduction described up to the moment (2, 4, 32, 33), our group of LATE treated patients would look like LTNPs. In LTNP patients, an increased adaptive immune activation has been described suggesting that spontaneous viral control may imply an immunological cost in those patients who maintain virologic control in the absence of treatment. Similarly, the activated immune phenotype we described herein on LATE treated vertically HIV-1 patients with higher viral reservoir, could reflect the exacerbated immune pressure needed against viral replication on this group. Interestingly, increased levels of miRNA-155 have been correlated with increased immune activation in HIV-scenario (34) accordingly to our results.
Regarding miRNA-18a, despite scarce evidences related to miR-18a-3p, low levels of miRNA family members like miRNA-18a and miR-18b-5p in LTNP compared to typical progressors indicated the potential role of that miRNA in viral replication and disease progression (35) by controlling genes of the inflammatory response as HIF-1α (36). The low levels of miRNA-18a-5p expression observed in EARLY treated vertically acquired HIV-patients studied in this work could explain the low viral reservoir and immune activation that has widely characterized this group of patients (37).
In addition to everything described so far related to miRNA-155-5p, that miRNA and miRNA-1248, which is associated with good survival prognoses in cancer (38), both upregulated on EARLY treated HIV-1 patients, strongly correlated with the activation marker HLA-DR, the natural cytotoxicity family activation receptor NKp30 and the inhibiting receptor NKG2A expressed on NK cells and more specifically on the main cytotoxic NK subset, CD16high and the activated and mature CD56negCD16+ NK cells, described as a subset with impaired effector functions in HIV-1 infection (39). Despite the limitation that miRNA profile was determined in the pool of PBMCs without isolated different cell subsets, our results are in agreement with recent insights that have highlighted the role of miRNAs in maintaining killing effect of NK cells in cancer scenario via activation markers modulation (40–42), in promoting antiviral immunity in the scenario of viral infection (43) and altogether suggests the possible role of both, miRNA-155-5p and miRNA-1248 in regulating inflammatory cytokine production in NK cells, potentially balancing disease progression during chronic HIV infection. Higher TIM-3 expression in NK cells has been considered a marker of better prognosis during HIV infection since its down regulation is indicative of IFN-γ deficient response (44) and interestingly, strong and direct correlations were also found between miRNA-155-5p levels, miRNA upregulated on EARLY treated HIV-1 young adults, and MFI expression of TIM-3 in CD56high NK cells.
MiRNAs have also arisen as critical regulators of lipidic metabolism controlling genes involved in HDL and LDL secretion, the most important risk factors of atherosclerosis, cholesterol biosynthesis and hepatic LDL receptor expression (45, 46). In our study EARLY treated HIV patients had decreased HDL levels traditionally associated with risk for developing cardiovascular diseases (CVD) and, while no differences were found in ratio LDL/HDL comparing groups, both biochemical parameters correlated with miRNA-501-3p and miRNA-155-5p relative levels. In the case of miRNA-501-3p, downregulated on EARLY treated HIV-1 young adults, that miRNA levels correlated directly with soluble HDL and inversely with ratio LDL/HDL and the opposite correlations were observed for miRNA-155-5p, upregulated on EARLY treated group.
Multiple miRNAs are being also evaluated as novel plasma biomarkers that out-perform or add value to the conventional liver injury biomarkers ALT, AST among others (47). Interestingly, miRNA-296-5p and miRNA-501-3p levels, both downregulated on EARLY group compared with LATE group, also correlated directly with AST levels. Recent evidences indicate that miRNA-296-5p regulates liver PIN1 and PLK1 genes that encode peptidylprolyl Cis/Trans isomerase NIMA-Interacting-1 and polo-like kinase 1 respectively, both involved in HIV-1 replication cycle stages (18, 19), suggesting that miRNA-296-5p may have a regulatory activity on virus replication in HIV pathogenesis and contributing to immune activation. These indicators could be potential biomarkers for disease progression and, in agreement with previous observations (48), good targets for therapies aimed at modulating the immune response in HIV infection. All explain herein, reinforce the idea that those distinct miRNAs could be considered regulators of cholesterol metabolism, hepatic lipoprotein synthesis, potential liver damage biomarkers and therapeutic targets for the treatment of CVD. However, genetic ablation of other miRNAs in the context of CVD development have led also to contradictory effects depending on animal model raising concerns about the use of therapies based on miRNAs (49).
There is unclear information on the risk of developing CVDs among other pathologies as our study is based on the time of treatment introduction and the cross-sectional design of our study as main limitation. In addition, the scarce number of participants and the low statistical power do not allow us to find differences in the immunological comparisons and results obtained here should be validated in a larger cohort of patients. Moreover, the in-depth pairing between groups in terms of CD4, CD8-T cell counts, time since ART initiation and since virological control can also explain the lack of immune phenotype differences on T- and NK cells. The panel used for miRNAs profile had limited number of them. However, samples of vertically infected patients reaching adulthood are scarce and mentioned limitations are also counterbalanced by the exhaustive follow-up of the participants during a median of 21 years since HIV diagnosis and during that period none of them has developed any inflammatory disease or non-AIDS-event. Further longitudinal observation would be necessary to determined clinical consequences that could be explained by the EARLY start of treatment.
In conclusion, our finding demonstrates that there is a miRNA signature that discriminates LATE treated from EARLY treated vertically acquired HIV-1-infected young adults. The role of those distinct miRNAs target genes in the modulation of HIV-1 replication and latency reveals new host targets that may be modulated in future antiviral strategies. Moreover, we found strong correlations between the most significant miRNAs and soluble inflammatory biomarkers and NK and T-cell activation and maturation suggesting that those miRNAs may be potential biomarkers for immune activation and progression disease in vertically acquired HIV-1-infected young adults who initiated a late ART and in general HIV-1-infected patients.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by Comité de Ética de Investigación con Medicamentos. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
MAM-F designed the research study. LT-D, IC, EV-A designed and performed the research studies. RR contributed essential tools. LT-D and EV-A analyzed the data. LT-D wrote the paper. JTR, MLN, and MAM-F critically discussed the paper. All authors have read and approved the final manuscript.
LT-D is supported by the Instituto de Salud Carlos III (ISCIII) under grant agreement “CD20/00025” through the Sara Borrell Program and by GeSIDA through the “Premio para Jóvenes Investigadores 2021”. EV-A was supported by Comunidad de Madrid, Programa de Empleo Joven “PEJ-2017 AI_BMD-7446”. This work has been (partially) supported by the RD16/0025/0019 projects as part of Acción Estratégica en Salud, Plan Nacional de Investigación Científica, Desarrollo e Innovación Tecnológica (2020–2022) and co-financed by Instituto de Salud Carlos III (Subdirección General de Evaluación) and Fondo Europeo de Desarrollo Regional (FEDER), RETIC PT17/0015/0042, Fondo de Investigación Sanitaria (FIS) 2020-2022 (grant number PI19/01638) and EPIICAL Project. Moreover, this work has been supported partially by a EUROPARTNER: Strengthening and spreading international partnership activities of the Faculty of Biology and Environmental Protection for interdisciplinary research and innovation of the University of Lodz Programme: NAWA International Academic Partnership Programme. This article/publication is based upon work from COST Action CA 17140 “Cancer Nanomedicine from the Bench to the Bedside” supported by COST (European Cooperation in Science and Technology) 2018-2022.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Iveta Waczulikova for her expert technical assistance and Laura Diaz of the Flow Cytometry Unit for her technical assistance as flow cytometry technician.We particularly acknowledge Santiago Jiménez de Ory for datasupport and valuable help.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.878630/full#supplementary-material
Supplementary Table 1 | Differentially expressed miRNAs comparing HD and EARLY and LATE treated HIV-1 infected young adults.
2. Barlow-Mosha L, Musiime V, Davies M-A, Prendergast AJ, Musoke P, Siberry G, et al. Universal Antiretroviral Therapy for HIV-Infected Children: A Review of the Benefits and Risks to Consider During Implementation. J Int AIDS Society (2017) 20(1):21552. doi: 10.7448/IAS.20.1.21552
3. Domínguez-Rodríguez S, Tagarro A, Palma P, Foster C, Puthanakit T, Jupimai T, et al. Reduced Time to Suppression Among Neonates With HIV Initiating Antiretroviral Therapy Within 7 Days After Birth. JAIDS J Acquired Immune Deficiency Syndromes (2019) 82(5):483–90. doi: 10.1097/QAI.0000000000002188
4. Tagarro A, Chan M, Zangari P, Ferns B, Foster C, De Rossi A, et al. Early and Highly Suppressive Antiretroviral Therapy Are Main Factors Associated With Low Viral Reservoir in European Perinatally HIV-Infected Children. JAIDS J Acquired Immune Deficiency Syndromes (2018) 79(2):269–76. doi: 10.1097/QAI.0000000000001789
5. Rinaldi S, Pallikkuth S, Cameron M, de Armas LR, Cotugno N, Dinh V, et al. Impact of Early Antiretroviral Therapy Initiation on HIV-Specific CD4 and CD8 T Cell Function in Perinatally Infected Children. JI (2020) 204(3):540–9. doi: 10.4049/jimmunol.1900856
6. Ananworanich J, Puthanakit T, Suntarattiwong P, Chokephaibulkit K, Kerr SJ, Fromentin R, et al. Reduced Markers of HIV Persistence and Restricted HIV-Specific Immune Responses After Early Antiretroviral Therapy in Children. AIDS (2014) 28(7):1015–20. doi: 10.1097/QAD.0000000000000178
7. Carrasco I, Tarancon-Diez L, Vázquez-Alejo E, Jiménez de Ory S, Sainz T, Apilanez M, et al. Innate and Adaptive Abnormalities in Youth With Vertically Acquired HIV Through a Multicentre Cohort in Spain. J Int AIDS Soc (2021) 24(10):e25804. doi: 10.1002/jia2.25804
8. Tarancón-Diez L, Rull A, Herrero P, Vazquez-Alejo E, Peraire J, Guillén S, et al. Early Antiretroviral Therapy Initiation Effect on Metabolic Profile in Vertically HIV-1-Infected Children. J Antimicrob Chemother (2021) 76(11):2993–3001. doi: 10.1093/jac/dkab277
9. Dalzini A, Petrara MR, Ballin G, Zanchetta M, Giaquinto C, De Rossi A. Biological Aging and Immune Senescence in Children With Perinatally Acquired HIV. J Immunol Res (2020) 2020:1–15. doi: 10.1155/2020/8041616
10. Colpaert RMW, Calore M. Epigenetics and microRNAs in Cardiovascular Diseases. Genomics (2021) 113(2):540–51. doi: 10.1016/j.ygeno.2020.12.042
11. Wang X, He Y, Mackowiak B, Gao B. MicroRNAs as Regulators, Biomarkers and Therapeutic Targets in Liver Diseases. Gut (2021) 70(4):784–95. doi: 10.1136/gutjnl-2020-322526
12. Wang P, Qu X, Zhou X, Shen Y, Ji H, Fu Z, et al. Two Cellular microRNAs, miR-196b and miR-1290, Contribute to HIV-1 Latency. Virology (2015) 486:228–38. doi: 10.1016/j.virol.2015.09.016
13. Reynoso R, Laufer N, Hackl M, Skalicky S, Monteforte R, Turk G, et al. MicroRNAs Differentially Present in the Plasma of HIV Elite Controllers Reduce HIV Infection In Vitro. Sci Rep (2015) 4(1):5915. doi: 10.1038/srep05915
14. Heinson AI, Woo J, Mukim A, White CH, Moesker B, Bosque A, et al. Micro RNA Targets in HIV Latency: Insights Into Novel Layers of Latency Control. AIDS Res Hum Retroviruses (2021) 37(2):109–21. doi: 10.1089/aid.2020.0150
15. García-Merino I, de las Cuevas N, Jiménez JL, García A, Gallego J, Gómez C, et al. Pediatric HIV BioBank: A New Role of the Spanish HIV BioBank in Pediatric HIV Research. AIDS Res Hum Retroviruses (2010) 26(2):241–4. doi: 10.1089/aid.2009.0122
16. Aguilera-Alonso D, Sainz T, Jimenez de Ory S, Bernardino I, Díez C, Torres B, et al. Clinical, Immunological, and Virological Outcomes Among Youths With Perinatal HIV After Transition to Adult Units in Spain From 1997 to 2016. JAIDS J Acquired Immune Deficiency Syndromes (2021) 86(2):240–7. doi: 10.1097/QAI.0000000000002539
17. Liu Y, Niu Y, Li L, Timani KA, He VL, Sanburns C, et al. Tat Expression Led to Increased Histone 3 Tri-Methylation at Lysine 27 and Contributed to HIV Latency in Astrocytes Through Regulation of MeCP2 and Ezh2 Expression. J Neurovirol (2019) 25(4):508–19. doi: 10.1007/s13365-019-00751-0
18. Manganaro L, Lusic M, Gutierrez MI, Cereseto A, Del Sal G, Giacca M. Concerted Action of Cellular JNK and Pin1 Restricts HIV-1 Genome Integration to Activated CD4+ T Lymphocytes. Nat Med (2010) 16(3):329–33. doi: 10.1038/nm.2102
19. Zhang S-M, Song M, Yang T-Y, Fan R, Liu X-D, Zhou P-K. HIV-1 Tat Impairs Cell Cycle Control by Targeting the Tip60, Plk1 and Cyclin B1 Ternary Complex. Cell Cycle (2012) 11(6):1217–34. doi: 10.4161/cc.11.6.19664
20. Ryo A, Tsurutani N, Ohba K, Kimura R, Komano J, Nishi M, et al. SOCS1 is an Inducible Host Factor During HIV-1 Infection and Regulates the Intracellular Trafficking and Stability of HIV-1 Gag. Proc Natl Acad Sci (2008) 105(1):294–9. doi: 10.1073/pnas.0704831105
21. Deshmane SL, Amini S, Sen S, Khalili K, Sawaya BE. Regulation of the HIV-1 Promoter by HIF-1α and Vpr Proteins. Virol J (2011) 8(1):477 doi:10.1186/1743-422X-8-477
22. Uhl J, Gujarathi S, Waheed AA, Gordon A, Freed EO, Gousset K. Myosin-X is Essential to the Intercellular Spread of HIV-1 Nef Through Tunneling Nanotubes. J Cell Commun Signal (2019) 13(2):209–24. doi: 10.1007/s12079-018-0493-z
23. Li H, Chi X, Li R, Ouyang J, Chen Y. HIV-1-Infected Cell-Derived Exosomes Promote the Growth and Progression of Cervical Cancer. Int J Biol Sci (2019) 15(11):2438–47. doi: 10.7150/ijbs.38146
24. Kula A, Guerra J, Knezevich A, Kleva D, Myers MP, Marcello A. Characterization of the HIV-1 RNA Associated Proteome Identifies Matrin 3 as a Nuclear Cofactor of Rev Function. Retrovirology (2011) 8(1):60. doi: 10.1186/1742-4690-8-60
25. Wang L, Zhang H, Solski PA, Hart MJ, Der CJ, Su L. Modulation of HIV-1 Replication by a Novel RhoA Effector Activity. J Immunol (2000) 164(10):5369–74. doi: 10.4049/jimmunol.164.10.5369
26. Dong H, Ye X, Zhong L, Xu J, Qiu J, Wang J, et al. Role of FOXO3 Activated by HIV-1 Tat in HIV-Associated Neurocognitive Disorder Neuronal Apoptosis. Front Neurosci (2019) 13:44. doi: 10.3389/fnins.2019.00044
27. Fadel HJ, Morrison JH, Saenz DT, Fuchs JR, Kvaratskhelia M, Ekker SC, et al. TALEN Knockout of the PSIP1 Gene in Human Cells: Analyses of HIV-1 Replication and Allosteric Integrase Inhibitor Mechanism. J Virol (2014) 88(17):9704–17. doi: 10.1128/JVI.01397-14
28. Houzet L, Yeung ML, de Lame V, Desai D, Smith SM, Jeang K-T. MicroRNA Profile Changes in Human Immunodeficiency Virus Type 1 (HIV-1) Seropositive Individuals. Retrovirology (2008) 5(1):118. doi: 10.1186/1742-4690-5-118
29. Seddiki N, Brezar V, Ruffin N, Lévy Y, Swaminathan S. Role of miR-155 in the Regulation of Lymphocyte Immune Function and Disease. Immunology (2013) 142(1):32–8 doi: 10.1111/imm.12227
30. Thai T-H, Calado DP, Casola S, Ansel KM, Xiao C, Xue Y, et al. Regulation of the Germinal Center Response by MicroRNA-155. Science (2007) 316(5824):604–8. doi: 10.1126/science.1141229
31. Bignami F, Pilotti E, Bertoncelli L, Ronzi P, Gulli M, Marmiroli N, et al. Stable Changes in CD4+ T Lymphocyte miRNA Expression After Exposure to HIV-1. Blood (2012) 119(26):6259–67. doi: 10.1182/blood-2011-09-379503
32. Violari A, Cotton MF, Gibb DM, Babiker AG, Steyn J, Madhi SA, et al. Early Antiretroviral Therapy and Mortality Among HIV-Infected Infants. N Engl J Med (2008) 359(21):2233–44. doi: 10.1056/NEJMoa0800971
33. Foster C, Domínguez-Rodríguez S, Tagarro A, Gkouleli T, Heaney J, Watters S, et al. The CARMA Study: Early Infant Antiretroviral Therapy—Timing Impacts on Total HIV-1 DNA Quantitation 12 Years Later. J Pediatr Infect Dis Soc (2021) 10(3):295–301. doi: 10.1093/jpids/piaa071
34. Jin C, Cheng L, Höxtermann S, Xie T, Lu X, Wu H, et al. MicroRNA-155 is a Biomarker of T-Cell Activation and Immune Dysfunction in HIV-1-Infected Patients. HIV Med (2017) 18(5):354–62. doi: 10.1111/hiv.12470
35. Ayala-Suárez R, Díez-Fuertes F, Calonge E, de la Torre Tarazona HE, Gracia-Ruíz de Alda M, Capa L, et al. Insight in Mirnome of Long-Term Non-Progressors and Elite Controllers Exposes Potential RNAi Role in Restraining HIV-1 Infection. JCM (2020) 9(8):2452 doi: 10.3390/jcm9082452
36. Duette G, Pereyra Gerber P, Rubione J, Perez PS, Landay AL, Crowe SM, et al. Induction of HIF-1α by HIV-1 Infection in Infection in CD4+ T Cells Promotes Viral Replication and Drives Extracellular Vesicle-Mediated Inflammation. Sarkar SN, Smithgall TE. mBio (2018) 9(5):e00757-18 doi: 10.1128/mBio.00757-18
37. Martínez-Bonet M, Puertas MC, Fortuny C, Ouchi D, Mellado MJ, Rojo P, et al. Establishment and Replenishment of the Viral Reservoir in Perinatally HIV-1-Infected Children Initiating Very Early Antiretroviral Therapy. Clin Infect Dis (2015) 61(7):1169–78. doi: 10.1093/cid/civ456
38. Zhang L, Chen J, Cheng T, Yang H, Pan C, Li H. Identification of Differentially Expressed Genes and miRNAs Associated With Esophageal Squamous Cell Carcinoma by Integrated Analysis of Microarray Data. BioMed Res Int (2020) 2020:1–16. doi: 10.1155/2020/2814548
39. Milush JM, López-Vergès S, York VA, Deeks SG, Martin JN, Hecht FM, et al. CD56negCD16+NK Cells are Activated Mature NK Cells With Impaired Effector Function During HIV-1 Infection. Retrovirology (2013) 10(1):158. doi: 10.1186/1742-4690-10-158
40. Chang Y, Cui M, Fu X, Zhang L, Li X, Li L, et al. MiRNA-155 Regulates Lymphangiogenesis in Natural Killer/T-Cell Lymphoma by Targeting BRG1. Cancer Biol Ther (2019) 20(1):31–41. doi: 10.1080/15384047.2018.1504721
41. Wei MF, Gu ZS, Zheng LL, Zhao MX, Wang XJ. Long non-Coding RNA GAS5 Promotes Natural Killer Cell Cytotoxicity Against Gastric Cancer by Regulating miR-18a. neo (2020) 67(05):1085–93. doi: 10.4149/neo_2020_191014N1034
42. Zhang J, Han X, Hu X, Jin F, Gao Z, Yin L, et al. IDO1 Impairs NK Cell Cytotoxicity by Decreasing NKG2D/NKG2DLs via Promoting miR-18a. Mol Immunol (2018) 103:144–55. doi: 10.1016/j.molimm.2018.09.011
43. Zawislak CL, Beaulieu AM, Loeb GB, Karo J, Canner D, Bezman NA, et al. Stage-Specific Regulation of Natural Killer Cell Homeostasis and Response Against Viral Infection by microRNA-155. Proc Natl Acad Sci (2013) 110(17):6967–72. doi: 10.1073/pnas.1304410110
44. Kared H, Martelli S, Tan SW, Simoni Y, Chong ML, Yap SH, et al. Adaptive NKG2C+CD57+ Natural Killer Cell and Tim-3 Expression During Viral Infections. Front Immunol (2018) 9:686. doi: 10.3389/fimmu.2018.00686
45. Rayner KJ, Suárez Y, Dávalos A, Parathath S, Fitzgerald ML, Tamehiro N, et al. MiR-33 Contributes to the Regulation of Cholesterol Homeostasis. Science (2010) 328(5985):1570–3. doi: 10.1126/science.1189862
46. Pedretti S, Brulhart-Meynet M-C, Montecucco F, Lecour S, James RW, Frias MA. HDL Protects Against Myocardial Ischemia Reperfusion Injury via miR-34b and miR-337 Expression Which Requires STAT3. Gallyas F. PloS One (2019) 14(6):e0218432. doi: 10.1371/journal.pone.0218432
47. Bailey WJ, Barnum JE, Erdos Z, LaFranco-Scheuch L, Lane P, Vlasakova K, et al. A Performance Evaluation of Liver and Skeletal Muscle-Specific miRNAs in Rat Plasma to Detect Drug-Induced Injury. Toxicol Sci (2019) 168(1):110–25. doi: 10.1093/toxsci/kfy282
48. Cárdenas-Bedoya J, Marquez-Pedroza J, Morán-Moguel MC, Escoto-Delgadillo M, Vázquez-Valls E, González-Enríquez GV, et al. MicroRNA-296-5p is Differentially Expressed in Individuals With and Without HIV-1 Infection. Genet Mol Biol (2020) 43(3):e20200017 doi: 10.1590/1678-4685-GMB-2020-0017
Keywords: miRNA profile, vertically acquired-HIV-1 infection, ART, youths, inflammatory profile
Citation: Tarancon-Diez L, Consuegra I, Vazquez-Alejo E, Ramos-Ruiz R, Ramos JT, Navarro ML and Muñoz-Fernández MA (2022) miRNA Profile Based on ART Delay in Vertically Infected HIV-1 Youths Is Associated With Inflammatory Biomarkers and Activation and Maturation Immune Levels. Front. Immunol. 13:878630. doi: 10.3389/fimmu.2022.878630
Received: 18 February 2022; Accepted: 23 March 2022;
Published: 22 April 2022.
Edited by:
Anita De Rossi, University of Padua, ItalyReviewed by:
Liz Simon, Louisiana State University, United StatesCopyright © 2022 Tarancon-Diez, Consuegra, Vazquez-Alejo, Ramos-Ruiz, Ramos, Navarro and Muñoz-Fernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mª Ángeles Muñoz-Fernández, bW11bm96LmhndWdtQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.