 Peter D. Burbelo1*
Peter D. Burbelo1* Riccardo Castagnoli2,3
Riccardo Castagnoli2,3 Chisato Shimizu4
Chisato Shimizu4 Ottavia M. Delmonte2
Ottavia M. Delmonte2 Kerry Dobbs2Valentina Discepolo5
Kerry Dobbs2Valentina Discepolo5 Andrea Lo Vecchio5
Andrea Lo Vecchio5 Alfredo Guarino5
Alfredo Guarino5 Francesco Licciardi6
Francesco Licciardi6 Ugo Ramenghi6Emma Rey-Jurado7
Ugo Ramenghi6Emma Rey-Jurado7 Cecilia Vial7
Cecilia Vial7 Gian Luigi Marseglia3
Gian Luigi Marseglia3 Amelia Licari3
Amelia Licari3 Daniela Montagna3Camillo Rossi8Gina A. Montealegre Sanchez9Karyl Barron10
Daniela Montagna3Camillo Rossi8Gina A. Montealegre Sanchez9Karyl Barron10 Blake M. Warner1
Blake M. Warner1 John A. Chiorini1
John A. Chiorini1 Yazmin Espinosa11Loreani Noguera7Lesia Dropulic12Meng Truong2
Yazmin Espinosa11Loreani Noguera7Lesia Dropulic12Meng Truong2 Dana Gerstbacher13
Dana Gerstbacher13 Sayonara Mató14John Kanegaye4Adriana H. Tremoulet4 Pediatric Emergency Medicine Kawasaki GroupEli M. Eisenstein15
Sayonara Mató14John Kanegaye4Adriana H. Tremoulet4 Pediatric Emergency Medicine Kawasaki GroupEli M. Eisenstein15 Helen C. Su2
Helen C. Su2 Luisa Imberti16
Luisa Imberti16 Maria Cecilia Poli7,11Jane C. Burns4†
Maria Cecilia Poli7,11Jane C. Burns4† Luigi D. Notarangelo2†
Luigi D. Notarangelo2† Jeffrey I. Cohen12†
Jeffrey I. Cohen12†- 1National Institute of Dental and Craniofacial Research, NIH, Bethesda, MD, United States
- 2Laboratory of Clinical Immunology and Microbiology, National Institute of Allergy and Infectious Diseases, NIH, Bethesda, MD, United States
- 3Department of Pediatrics, Fondazione IRCCS Policlinico San Matteo, University of Pavia, Pavia, Italy
- 4Department of Pediatrics, Rady Children’s Hospital, University of California San Diego, San Diego, CA, United States
- 5Department of Translational Medical Science, Section of Pediatrics, University of Naples Federico II, Naples, Italy
- 6Division of Pediatric Immunology and Rheumatology, Department of Public Health and Pediatric Sciences, “Regina Margherita” Children’s Hospital, University of Turin, Turin, Italy
- 7Instituto de Ciencias e Innovación en Medicina (ICIM), Clínica Alemana Universidad del Desarrollo, Santiago, Chile
- 8Direzione Sanitaria, ASST Spedali Civili, Brescia, Italy
- 9Intramural Clinical Management and Operations Branch (ICMOB), Division of Clinical Research NIAID, NIH, Bethesda, MD, United States
- 10Division of Intramural Research, National Institute of Allergy and Infectious Disease, NIH, Bethesda, MD, United States
- 11Hospital Roberto del Río, Santiago, Chile
- 12Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, NIH, Bethesda, MD, United States
- 13Pediatric Rheumatology, Stanford Children’s Hospital, Stanford, CA, United States
- 14Pediatric Infectious Diseases, Randall Children’s Hospital at Legacy Emanuel, Portland, OR, United States
- 15Department of Pediatrics, Hadassah Medical Center, Faculty of Medicine, Hebrew University, Jerusalem, Israel
- 16CREA Laboratory, Diagnostic Department, ASST Spedali Civili di Brescia, Brescia, Italy
The antibody profile against autoantigens previously associated with autoimmune diseases and other human proteins in patients with COVID-19 or multisystem inflammatory syndrome in children (MIS-C) remains poorly defined. Here we show that 30% of adults with COVID-19 had autoantibodies against the lung antigen KCNRG, and 34% had antibodies to the SLE-associated Smith-D3 protein. Children with COVID-19 rarely had autoantibodies; one of 59 children had GAD65 autoantibodies associated with acute onset of insulin-dependent diabetes. While autoantibodies associated with SLE/Sjögren’s syndrome (Ro52, Ro60, and La) and/or autoimmune gastritis (gastric ATPase) were detected in 74% (40/54) of MIS-C patients, further analysis of these patients and of children with Kawasaki disease (KD), showed that the administration of intravenous immunoglobulin (IVIG) was largely responsible for detection of these autoantibodies in both groups of patients. Monitoring in vivo decay of the autoantibodies in MIS-C children showed that the IVIG-derived Ro52, Ro60, and La autoantibodies declined to undetectable levels by 45-60 days, but gastric ATPase autoantibodies declined more slowly requiring >100 days until undetectable. Further testing of IgG and/or IgA antibodies against a subset of potential targets identified by published autoantigen array studies of MIS-C failed to detect autoantibodies against most (16/18) of these proteins in patients with MIS-C who had not received IVIG. However, Troponin C2 and KLHL12 autoantibodies were detected in 2 of 20 and 1 of 20 patients with MIS-C, respectively. Overall, these results suggest that IVIG therapy may be a confounding factor in autoantibody measurements in MIS-C and that antibodies against antigens associated with autoimmune diseases or other human proteins are uncommon in MIS-C.
Introduction
Coronavirus disease 2019 (COVID-19) caused by the SARS-CoV-2 virus, is associated with a high rate of respiratory-related mortality (1). Besides pulmonary disease, SARS-CoV-2 infection can result in acute injury to the kidney, liver, and heart, disseminated intravascular coagulation, rhabdomyolysis, and chronic fatigue (2). The variability in clinical symptoms with COVID-19 likely involves sites of virus and viral antigen localization, vascular complications associated with the infection, the level of inflammation, and host factors including genetics, age, gender, and coexistence of other comorbid conditions (3). In addition, the host immune response has a significant impact on disease severity and outcome (4, 5). One major immune pathway involved in limiting SARS-CoV-2 infection is the type I interferon (IFN) pathway. Evidence for its role is based on genetic studies showing that individuals with inborn errors of type I IFN are at higher risk of severe COVID-19 (6). Furthermore, neutralizing autoantibodies to IFN-α and/or -ω occur in approximately 10% of patients with severe COVID-19 pneumonia, especially men, and blunt effective clearance of the virus, likely contributing to death (7).
Compared to adults, most children and young adolescents with acute SARS-CoV-2 infection remain asymptomatic or have mild symptoms (8, 9). However, a rare, severe condition, multisystem inflammatory syndrome in children (MIS-C), presents with high-grade fever, rash, multi-organ dysfunction, and elevated markers of inflammation, which typically occur 4-6 weeks after SARS-CoV-2 infection (10). MIS-C clinical manifestations resemble in part those of Kawasaki Disease (KD), a vasculitis typically affecting children. Children with MIS-C more frequently show myocardial involvement and gastrointestinal symptoms. The mechanisms involved in the pathogenesis of MIS-C are uncertain. Common treatments for MIS-C and KD include high dose intravenous immunoglobulin (IVIG), corticosteroids, and biologic agents (11). In particular, IVIG, containing pooled IgG purified from >10,000 donors (12), is the standard first-line treatment for KD and is often administered to MIS-C patients.
Increasing efforts are being directed at understanding the differences in symptoms and complications seen in COVID-19 patients with the goal of improving treatment. One strategy is to identify biomarkers associated with, and predictive of, severe COVID-19 outcomes and involves analysis of soluble molecules in blood such as cytokine, chemokines, and other inflammatory markers (13–15). From these and additional studies, a large body of evidence implicates the importance of the immune system in both controlling infection and promoting tissue damage. Several recent studies have identified autoantibodies in cross-sectional serum/plasma samples of adults and children with COVID-19 and children with MIS-C (16–21). Here, we assessed the presence of autoantibodies against a panel of autoantigens associated with known autoimmune diseases in adults and children with COVID-19. Our analysis showed a moderate prevalence of autoantibodies targeting the lung antigen KCNRG in adults, but not in children with COVID-19. We also demonstrate a high prevalence of autoantibodies associated with systemic lupus erythematosus (SLE)/Sjögren’s syndrome and autoimmune gastritis in the serum/plasma of patients with MIS-C, which we found to be associated with previous administration of high-dose IVIG and were absent from MIS-C patients not receiving IVIG. Based on recent autoantigen array studies identifying numerous potential antibodies against diverse proteins in MIS-C (16–18), we tested MIS-C patients who had not received IVG for autoantibodies to 16 additional proteins using an immunoprecipitation assay that detects both linear and conformational epitopes associated with the native form of the these proteins. Both IgG and IgA antibodies to most (16/18) of these proteins were not detected in the patients. However, one patient with MIS-C had low levels of autoantibodies against Troponin-C2 and another patient had autoantibodies against both Troponin-C2 and the KLHL12 autoantigen associated with Sjögren’s syndrome/primary biliary cirrhosis. Overall, our findings differ from prior studies that reported extensive production of autoantibodies against multiple proteins in MIS-C.
Material and Methods
Patient Population
Deidentified serum/plasma samples were obtained from cases with MIS-C based on CDC criteria https://emergency.cdc.gov/han/2020/han00432.asp, other COVID-19 patients or control subjects from multiple sites under an NIH IRB exemption after local IRB approval: Brescia, Italy Comitato Etico Provinciale (NP 4000 and NP 4408); Pavia, Italy (Comitato Etico Pavia Prot. 20200037677); Turin, Italy (Comitato Etico Interaziendale A.O.U. Città della Salute e della Scienza di Torino, Protocol: 00282/2020); Naples, Italy (Ethics Committee of the University of Naples Federico II, Protocol: 158/20); Chile (Comité Ético Científico Facultad de Medicina Clínica Alemana Universidad del Desarrollo, Santiago, Chile, Protocol: 2020-41); and Israel (Hadassah Medical Organization Institutional Review Board (IRB) for studies involving human subjects, Protocol: HMO-235-20). Additional MIS-C patients were enrolled from other sources including Stanford, California and Portland, Oregon, after written informed consent to NIH IRB-approved research protocols (NIH Protocols l1-1-0109, 18-I-0041 and 18-I-0128). Sera from children with KD or acute febrile illnesses were obtained at the University of California, San Diego under IRB approved protocols (UCSD #140220). All patients and parents gave written informed consent and assent as appropriate.
Serum/plasma samples from adults with COVID-19 (n=80) in Brescia, Italy were obtained from a higher number of male than female (48:32) patients with a median age of 55 years [interquartile range (IQR): 45-66], and a median time between blood draw and onset of symptoms of 25 days (IQR: 13-52). These adults had varying degrees of severity of disease (critical, severe, moderate, and mild) based on a previously described clinical rating system (22). Serum/plasma samples from SARS-CoV-2 uninfected, healthy children (n=16) were obtained from Pavia, Italy (Table 1). Serum/plasma samples from children approximately 8 days after onset of acute febrile infections unrelated to KD or SARS-CoV-2 (n=16), from children during the acute phase of KD (median time of 9 days after disease onset, n=39), and from children during the subacute phase of KD (median time of 40 days after disease onset, n=37) were obtained from the University of California, San Diego. Samples from children with COVID-19 and MIS-C were from multiple locations in the US, Israel, Italy, and Chile. Among the children with COVID-19 without MIS-C, 59 were in the acute phase of COVID-19 and 8 had recovered from COVID-19 that had occurred approximately two months earlier. All patients with MIS-C (n=54) were SARS-CoV-2 PCR and/or antibody positive (Table 1). Forty-four percent of the patients with MIS-C had 2 or more longitudinal samples that were obtained from the time of initial hospitalization to several weeks later.
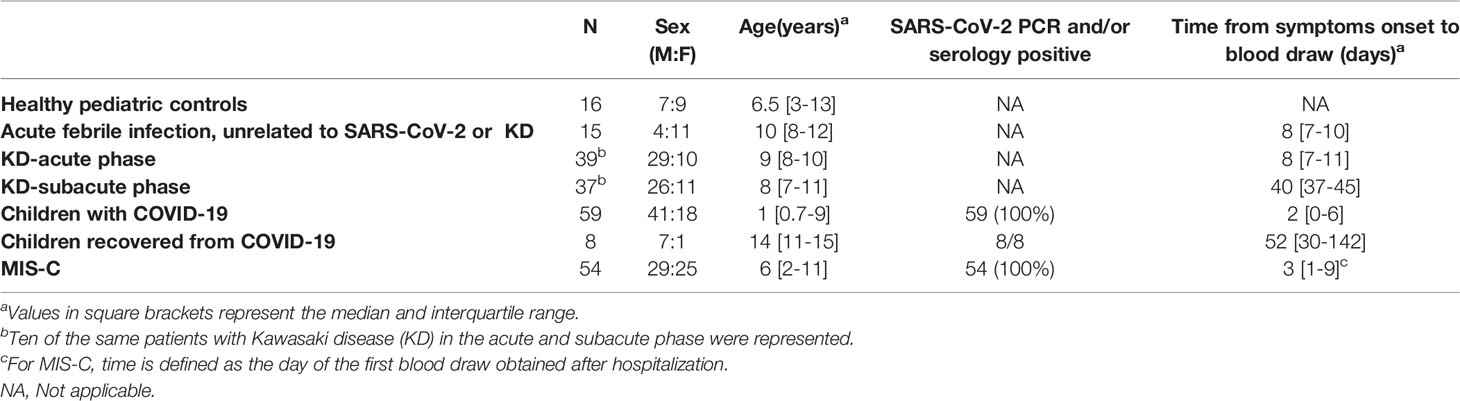
Table 1 Subject characteristics of the pediatric cohort (n = 228).
LIPS Antibody and Autoantibody Testing
Luciferase Immunoprecipitation Systems (LIPS) assays use a 96-well format to detect antibodies to both conformational and linear epitopes in a fluid phase system (23). A key aspect of the LIPS technology is the use of custom luciferase-antigen fusion proteins, containing the antigen of interest, which provide high sensitivity and specificity and a wide dynamic range of detection. Panels of autoantigen targets used for LIPS testing have been previously described and validated in many autoimmune diseases (23), including Sjogren’s syndrome, SLE (24), systemic sclerosis, myositis, anti-cytokine autoimmune diseases, corpus autoimmune gastritis (25), ANCA-associated vasculitis, autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) (26), type 1 diabetes (27), autoimmune encephalitis, and membranous nephropathy. A selected panel of autoantigens representing both ubiquitous and tissue-specific antigens were first tested in adults and included: Ro52, Ro60, La, RNP-A, Sm-D3, JO-1, gastric ATPase (ATPB4 subunit), LGI1, U1-70K, CENP-A, RNAP-K, BPIFB1, defensin 5A, KCNRG, glutamic acid decarboxylase (GAD65), gastric inhibitory factor (GIF), and M-type phospholipase A2 receptor (PLA2R). Based on the results of the adult COVID-19 study and the limited availability of serum/plasma for the COVID-19 children with and without MIS-C, a smaller number of autoantigens were selected for study in these children. Due to a single case showing GAD65 autoantibodies in a patient with symptoms of acute onset of type 1 diabetes, follow-up LIPS studies were performed to determine immunoreactivity to IA2 and ZNT-8, two autoantigens associated with diabetes mellitus. While healthy adult blood donor samples were used for to determine negative cut-off values for LIPS assays in adult COVID-19 patients, the control children without COVID-19 were used to determine the cut-off values in children with KD, COVID-19 and MIS-C. The value derived from the mean plus three standard deviations of the controls were used to set the cut-off for determining seropositivity (23). For the gastric ATPase, one elevated outlier from the control group of children was excluded in determining the cut-off value.
Besides examining autoantibodies by LIPS to well-established proteins associated with autoimmunity, we also tested the immunoreactivity of a subset of proteins previously identified by three autoantigen array studies (16–18). Due to the limited amount sera availability from MIS-C cases, we tested for IgG against 16 proteins and IgA autoantibodies against 6 proteins.
Sera from the initial cohort of MIS-C patients (who had not received IVIG) that were tested using LIPS assays were from subjects in Turin, Italy and additional MIS-C patients (not given IVIG) were from other locations including Chile and Israel. The proteins that had been previously identified as IgG targets using autoantigen arrays that we tested for IgG autoantibodies using LIPS included CSNK1A and LGAL3 (17), ECE1, IFNGR2, IL-1A, and Jo-1 (18) and ANX11, ATP4A, ECE1, FAM84A, KLHL12, NAPB, PDLIM5, TNNC2, and UBE3A (16). In addition, six previously reported targets of IgA autoantibodies (16, 18) included CALM1, Hepatitis B virus (HBV) X protein, JO-1 and IL-1A and FAM84A and TNNC2 which we tested by LIPS in our MIS-C patients. For detecting IgA autoantibodies, goat anti-human IgA-agarose conjugated beads (Sigma) were substituted for protein A/G beads as previously described (27). Of note, autoantibodies to four of the previously reported proteins detected in protein array studies in MIS-C patients including ATPA4, IL-1A, Jo-1, and KLHL12, have been detected by LIPS assays in known autoimmune diseases (23, 25) and in other published and unpublished studies.
LIPS testing was used to evaluate the autoantibody levels in three different batches of IVIG. For these experiments, the stock 10% IVIG solution (Gamunex®, Grifols Therapeutics Inc.) was diluted 1:10 in PBS to achieve approximately the same concentration of IgG present in human sera (20 μg/μL). In addition to testing the IVIG preparation for autoantibodies against Ro52, Ro60, La and the gastric ATPase, several additional autoantigens were tested including Jo-1 and IL-1A.
Statistical Analysis
GraphPad Prism software (San Diego, CA) was used for analyzing the antibody levels in this study. Median antibody levels, expressed as mean log10 light units (LU) and interquartile range (IQR), were calculated and presented as antilog values. In some cases, the non-parametric Mann-Whitney U statistical test was used for comparison of antibody levels in the control and COVID-19 and MIS-C patient groups. The Fishers exact test was used to evaluate the statistical significance of the prevalence of autoantibodies in patient groups and controls. JMP® 14.0.0 was used to illustrate the kinetics of antibody levels over time using a smoothing spline (cubic spline) with a λ of 0.77 and a shaded area representing the bootstrap confidence region for each fit.
Results
Characteristics of Adults and Children With COVID-19 and Children With MIS-C
Plasma from adults and children with acute COVID-19 and children with MIS-C were obtained from multiple geographical sites including the US, Italy, Israel, and Chile. The median age of the MIS-C children (n=54) was 6 years (IQR; 2 to 11 years) which was older than in children with COVID-19 (median age 1 year, IQR; 0.7 to 9 years) (Table 1). Furthermore, there was an approximately equal number of males and females in MIS-C patients, while the children with COVID-19 had a male predominance (69%).
Sixty-one percent of the patients with MIS-C were Hispanic/Latino, 32% were Caucasian, and 7% were black (Table 2). The most common symptom in the MIS-C cohort was fever (100%) followed by gastrointestinal symptoms of abdominal pain, vomiting, and diarrhea (91%). Of the total cohort of MIS-C patients, 87% were treated with corticosteroids and 70% were given IVIG.

Table 2 Demographics and clinical characteristics of the MIS-C cohort (N = 54).
Autoantibodies in Adults With COVID-19
A cohort of adults with COVID-19 and with different levels of clinical severity was analyzed for autoantibodies against known antigens associated with multiple autoimmune diseases. A small number of COVID-19 patients had antibodies against Ro52, Ro60, and La comprising the SSA and SSB rheumatological autoantigens (Figures 1A–C). Only one subject had autoantibodies to all three targets and no correlation of these autoantibodies was seen with disease severity. Testing of two SLE-associated antigens showed a lack of autoantibodies against RNP-A (Figure 1D); however, there was a high frequency of Sm-D3 autoantibodies in 34% (27/80) of adult COVID-19 patients (Figure 1E). Approximately 60% of the severe patients had Sm-D3 autoantibodies compared to 4%, 43%, and 27% in the subjects with critical, moderate, and mild disease, respectively (Figure 1E). The high prevalence of autoantibodies was statistically significant by the Fisher exact test in severe (P=0.005) and moderate disease (P=0.03) compared to the control group. Low levels of autoantibodies to KCNRG, a lung antigen, were found in 30% (24/80) of adult COVID-19 patients and were significantly enriched in the patients with severe or moderate disease and generally absent from patients with critical or mild disease (Fisher Exact test; P<0.003) (Figure 1F). Autoantibodies against GAD65, an autoantigen associated with type 1 diabetes and autoimmune encephalitis, was also detected in 8% of the severe and 4% of the critical patients (Figure 1G). Finally, high levels of autoantibodies against gastric ATPase, which are associated with autoimmune gastritis, were detected in approximately 9% of the COVID-19 patients, but the presence of these autoantibodies did not correlate with clinical severity (Figure 1H). Testing of antibodies to several other autoantigens including CENP-A, PLA2R, and LGI1 that are found in patients with systemic sclerosis, membranous nephropathy, and limbic encephalitis, respectively, revealed little or no autoantibodies in any of COVID-19 adults (data not shown). Taken together, these results suggest that adults with COVID-19 had a diverse repertoire of serum autoantibodies to known autoantigens and at least two of them, the lung KCNRG and stomach gastric ATPase, might reflect infection-related damage to these tissues in selected patients.

Figure 1 Autoantibodies against known autoantigens in adults with COVID-19 not receiving IVIG. Autoantibody levels against eight well known autoantigens (panels A–H) were determined in adults with COVID-19 having varying levels of disease severity: critical, severe, moderate, and mild. Each symbol represents a sample from an individual control subject (SARS-CoV-2 uninfected health care worker (NIH HCW), a patient with COVID-19 (critical, severe, or moderate, or mild). Autoantibody levels are plotted in light units on a log10-scale and the solid horizontal line represents the mean value in each group. The dashed lines represent the cut-off level for determining seropositive autoantibodies as described in the Methods. Only statistically significant P values from the Fisher exact test examining autoantibody prevalence differences between the uninfected HCW and the different severity groups of COVID-19 adults are shown.
Autoantibody Profiles of Children With COVID-19 or MIS-C
Levels of autoantibodies were also measured in cross-sectional and longitudinal serum/plasma samples of children with COVID-19 or with MIS-C, with many (70%) of the latter receiving IVIG. As controls, plasma samples collected from healthy children without SARS-CoV-2 infection were used as controls and to determine cut-off values for these autoantibodies. Approximately 50% of MIS-C children had Ro52, Ro60, and/or La autoantibodies (Figures 2A–C), which were nearly absent in children with COVID-19 (1/59) and in children (0/8) who had recovered from COVID-19. One child with acute COVID-19 that was seropositive for Ro52, Ro60, and La autoantibodies had a complex of neurological findings not fitting a clear diagnosis and received IVIG for autoimmune thrombocytopenia six days before the serum sample was collected. In MIS-C, the high prevalence of autoantibodies against Ro52, Ro60, La and the gastric ATPase were often found together in the same individuals. The relative antibody levels were comparable to those seen in some patients with SLE or Sjögren’s syndrome and were on average over 10 times higher than the cut-off value. The prevalence of autoantibodies against Ro52, Ro60, and La antigens among the individual MIS-C patients was 57% (31/54), 67% (36/54), and 59% (32/54), respectively, and these autoantibodies were often present at more than one time point. None of the recovered COVID-19 children, and only one child at two different time points with COVID-19 had all three autoantibodies against Ro52, Ro60, La, and gastric ATPase autoantibodies. Autoantibodies against gastric ATPase were also present in 74% (40/54) of the children with MIS-C (Figure 2D). The median level of these gastric ATPase autoantibodies in MIS-C children was 122,705 LU (IQR 11,810-253,100) which was approximately 10-fold higher than in children with COVID-19 (8790 LU; IQR 7,567-10,670) and 12-fold higher than in uninfected children (9,825 LU; IQR 9190-11,314). Examination of whether autoantibody positivity against Ro52, Ro60, La, and gastric ATPase were associated with clinical features of the MIS-C patients revealed that patients with autoantibodies had more severe disease based on a statistically higher frequency of ICU admission than those without autoantibodies (P=0.0003) and all were treated with IVIG.

Figure 2 Many children with MIS-C treated with IVIG show a high prevalence of autoantibodies against Ro52, Ro60, La, and gastric ATPase. (A–D) Autoantibody levels against Ro52, Ro60, La, and gastric ATPase were determined in 16 children not infected with SARS-CoV-2 (Controls), 59 children with COVID-19 (COVID-19-C), 116 serum/plasma samples from 54 children with MIS-C, and 8 children who recovered from acute COVID-19 (COVID-19-C late). Each symbol represents a sample from an individual patient or different time points from an individual patient (A–D) Autoantibody levels against four autoantigens, KCNRG, PLA2R, GAD65, and GIF (E–H) were tested in the same children as in panels (A–D), except for MIS-C children where a single sample from the time of peak Ro52, Ro60, La, and/or gastric autoantibodies was used. Autoantibody levels are plotted on the Y-axis in light units on a log10 scale and the solid horizontal line represents the mean value in each group The dashed lines represent the cutoff levels for determining positive autoantibody levels for each target antigen as described in the Methods. The sample from the child with acute COVID-19 with high levels of GAD65 autoantibodies is denoted by the open black circle. Only statistically significant P values from the Fisher exact test examining autoantibody prevalence differences between the controls and the groups of COVID-19 and MIS-C children are shown.
Samples from children with COVID-19 or MIS-C were also examined to determine if there was immunoreactivity against additional autoantigens including KCNRG, PLA2R, GAD65, and gastric intrinsic factor (GIF). No autoantibodies were detected above the cut-off values against the lung (KCNRG) and kidney (PLA2R) antigens (Figures 2E, F). High levels of autoantibodies against GAD65, but not to Ro52, Ro60 and/or gastric ATPase were detected in a 10-year-old girl with acute SARS-CoV-2 infection who had not received IVIG (Figure 2G). Review of the medical records indicated that this girl with COVID-19 was hospitalized with severe ketoacidosis and required intravenous insulin. Additional autoantibody testing against other type 1 diabetes-associated autoantigens including IA2, and Znt-8, was negative. These results suggest the possibility that acute COVID-19, at least in one child, may have contributed to an immune response triggering type 1 diabetes. Autoantibodies against GAD65 were also detected in approximately 26% of children with MIS-C, but their levels were only slightly above the cut-off threshold (Figure 2G). Importantly, all the GAD65 seropositive MIS-C children had received IVIG prior to blood sampling. Testing for antibodies against GIF, which is produced by parietal cells and whose loss is associated with B12 deficiency, showed only sporadic low levels of GIF autoantibodies in children with COVID-19 and MIS-C (Figure 2H).
No autoantibodies against IA2 and defensin-5A (a self-antigen that is expressed in the ileum and is targeted by autoantibodies in patients with autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy patients) were detected in children with COVID-19 or MIS-C (data not shown).
Ro52, Ro60, La, and Gastric ATPase Autoantibodies in MIS-C and KD Are Due to IVIG Administration
Based on the high prevalence of Ro52, Ro60, La, and gastric ATPase autoantibodies in MIS-C, additional testing was performed to determine if these autoantibodies could help to distinguish MIS-C from KD and whether their presence could be affected by previous administration of high-dose IVIG. To address this, we analyzed serum/plasma from control children (n=8), children with acute febrile infections other than COVID-19 (n=15), children with MIS-C before they received IVIG (n=10), children with KD prior to receipt of IVIG (n=39), and children with KD after receiving IVIG (n=37). In addition, three batches of IVIG were tested for autoantibodies. Only one child with acute febrile illness not related to COVID-19, who was later diagnosed with SLE, showed Ro52, Ro60, and La autoantibodies, but not gastric ATPase autoantibodies (Figure 3). Interestingly, no MIS-C children who had not received IVIG had autoantibodies to any of the four antigens (Figure 3). Moreover, children with acute KD, who had not yet received IVIG, showed no autoantibodies against Ro52, but two showed seropositivity for gastric autoantibodies and a third patient had both Ro60 and La autoantibodies. In contrast, subacute KD patients from later time points, all of whom received IVIG in the previous 6 weeks, showed a very high rate of autoantibodies against Ro52, Ro60, La, and gastric ATPase with a frequency of 46%, 68%, 35% and 86%, respectively (Figure 3). Five subacute KD patients showed no detectable autoantibodies. Testing of three different IVIG preparations diluted 1:10, which approximates the IgG antibody concentration in human serum showed remarkably high levels of autoantibodies against the four autoantigens. The median levels of Ro52, Ro60, La, and gastric ATPase autoantibodies approximated the highest levels seen in the KD patients with geometric mean levels of 125,000, 359,300, 120,000 and 117,800 LU, respectively (Figure 3). In addition, testing of two commonly found autoantibody targets, IL-1α and Jo-1, that have been observed in both healthy persons and patients with myositis, respectively, revealed low level autoantibodies in IVIG just above the cut-off value derived from control children (data not shown). Together these results suggest that the Ro52, Ro60, La, and gastric ATPase autoantibodies observed in MIS-C and KD patients were due to prior infusion of high-dose IVIG and that other autoantibodies found in IVIG preparations may contribute to anomalous autoantibody seropositivity,

Figure 3 Autoantibodies in the serum/plasma of patients with MIS-C and Kawasaki disease (KD) are due to administration of IVIG. Autoantibody levels are plotted in light units on a log10 scale. The dashed lines represent the cut-off level for determining positive antibody titers as described in the Methods. Results from 8 sera from children not infected with SARS-CoV-2 (Controls), 15 sera from children with acute febrile infection not due to COVID-19 or KD (child acute infection), 10 sera from patients with MIS-C who did not receive IVIG (MISC-no IVIG), 39 sera from children with acute KD (KD-acute), 37 sera children with subacute KD (KD-subacute), and three preparations of IVIG are shown. Only statistically significant P values from the Fisher exact test examining autoantibody prevalence differences between the control group and the groups of children are shown.
Longitudinal Analysis of SARS-CoV-2 Autoantibodies in Children With MIS-C and KD
To formally assess if the Ro52, Ro60, La, and gastric ATPase autoantibodies detected in the children with MIS-C and KD were due to IVIG, serial serum/plasma samples were tested from three MIS-C and three KD patients that were collected both before and after receiving IVIG. Analysis of the sera of MIS-C and KD patients before IVIG administration revealed that none of the patients had Ro52, Ro60, La, and gastric ATPase autoantibodies (Figure 4). However, serum samples taken within 24 hours after IVIG administration in these patients showed that they all contained high levels of autoantibodies. Furthermore, analysis of longitudinal samples from these six patients demonstrated high levels of these autoantibodies that persisted over the next 12 days (Figure 4). Taken together, these results confirm that the Ro52, Ro60, La, and gastric ATPase autoantibodies observed in these patients are directly due to administration of IVIG.

Figure 4 Longitudinal autoantibody profiles before and after administration of IVIG in MIS-C and KD patients. Antibody levels were determined in serial samples from representative MIS-C (MIS-C, Pt1-3) and KD patients (KD, Pt1-3), in which time zero represents the day of hospital admission. The autoantibody profiles for Ro52 (blue), Ro60 (red), La (green), and gastric ATPase (purple) are shown by the colored lines, in which the cut-off value for determining positivity is shown by the dotted lines. The arrow shows the day when IVIG was administered.
To accurately explore the kinetics of decay of autoantibodies, we assessed levels of Ro52, Ro60, La, and gastric ATPase autoantibodies in 21 MIS-C patients for which samples were available at two or more time points after IVIG administration (Figure 5). Based on the known time of IVIG administration, the Ro52, Ro60, and La autoantibodies showed a relatively rapid decline back to the seronegative cut-off value by 35-60 days (Figures 5A–C). However, the gastric ATPase autoantibody decline trajectory was generally more delayed, in that most of the samples did not approach the seronegative cut-off value in the available serial samples. Modeling of gastric ATPase autoantibodies suggested a slower decay trajectory lasting >100 days (Figure 5D).

Figure 5 Ro52, Ro60, La, and gastric ATPase autoantibody decay in patients with MIS-C receiving IVIG. Decay plots (A–D) were generated for Ro52, Ro60, La, and gastric ATPase (ATPB4) autoantibody data from the MIS-C (n = 21) patients having two or more different time points. Day 0 is the time the patients received IVIG. The solid-colored line for each autoantigen indicates the decay obtained using JMP® 14.0.0 and a smoothing spline (cubic spline) with a λ of 0.77 with the shaded area representing the bootstrap confidence region for each fit. The dotted lines indicate the cut-off value for determining seronegativity for each autoantigen.
LIPS Assay Analysis for Autoantibodies Previously Reported in MIS-C
Based on the large number of autoantibodies identified previously by protein array studies in MIS-C (16–18), LIPS assays were used to examine IgG autoantibodies against some of these proteins in 8 SARS-CoV-2 uninfected children and 10 children with MIS-C who had not received IVIG. The potential targets of IgG autoantibodies included CSNK1A and LGAL3S (17), CALM1, ECE1, IFNGR2, IL1A, and JO-1 (18), and ANX11, ATP4A, ECE1, FAM84A, KLHL12, NAPB, PDLIM5, TNNC2, and UBE3A (16). Based on LIPS testing, elevated antibody titers were not detected to most (14/16) of these proteins in the 10 children with MIS-C including antibodies against CSNK1A, LGAL3S, ANX11, ATP4A, FAM84A, JO-1, NAPB, PDLIM5, IFNGR2, and ECE1 (Figures 6A–H and 7A, B). No IgG autoantibodies were detected against UBE3A and the two IgA targets, CALM1 and HBV X protein (data not shown). However, two patients with MIS-C had autoantibodies against Troponin-C2 (TNNC2), a protein enriched in skeletal muscle (Figure 7C); neither of these patients had myocarditis. One of the two MIS-C patients had high titers of autoantibodies against Troponin C2 and against KLHL12, a known autoantigen (28, 29) in Sjögren’s syndrome/primary biliary cirrhosis (Figure 7D). Based on the identification of antibodies against TNNC2 and KLHL2, another independent set of 10 control children and 10 MIS-C patients who did not receive IVIG were tested. However, no additional patients had antibodies against these two proteins. These results suggest that in patients with MIS-C the prevalence of antibodies against TNNC2 and KLHL12 is very low (~5%). Furthermore, none of the patients with MIS-C had IgA antibodies against CALM1, JO-1, IL-1A, HBV X protein, FAM84A and TNNC2 (Figures 7E–H and data not shown).

Figure 6 Lack of IgG autoantibodies to previously reported autoantigens in MIS-C. SARS-CoV2 uninfected children (controls) and MIS-C children who did not receive IVIG (MISC-no IVIG) were screened for IgG autoantibodies against potential target proteins (panels A–H) discovered previously by autoantigen array studies. Autoantibody levels are plotted in light units on a log10 scale. The dashed lines represent the cut-off level for determining positive antibody titers as described in the Methods. No statistically significant seropositive autoantibodies in MIS-C were detected in these previous array-identified proteins.

Figure 7 Paucity of IgG and IgA autoantibodies to previously reported autoantigens in MIS-C. SARS-CoV2 uninfected children (controls) and MIS-C children who did not receive IVIG (MISC-no IVIG) were screened for IgG (A–D) and IgA (E–H) autoantibodies against potential target proteins discovered previously by autoantigen array studies. Autoantibody levels are plotted in light units on a log10 scale. The dashed lines represent the cut-off level for determining positive antibody titers as described in the Methods. Only two antigens, TNNC2 and KLHL12 showed occasional IgG seropositivity in MIS-C and no seropositives IgA responses were detected.
Discussion
Here we evaluated the prevalence of autoantibodies in adults and children with COVID-19 and children with MIS-C. Autoantibodies commonly found in autoimmune disease were chosen as target antigens because of their known association with different types of tissue damage. Analysis of adult patients with COVID-19 showed a moderate frequency of autoantibody positivity against several autoantigens including the lung protein KCNRG (30%) and the SLE antigen Sm-D3 (34%). Several other autoantigens, including Ro52, Ro60, La, and GAD65 were detected less frequently (< 8%) in the adults with COVID-19. While the levels of KCNRG autoantibodies seen in adults with COVID-19 occurred at a higher frequency than that seen in adults with ARDS (23%) or sepsis (25%) (30), they were lower than those reported in other diseases such as APECED complicated by pneumonitis (29%) (26). Detection of autoantibodies to KCNRG is consistent with the lung injury and dysfunction that occur in many hospitalized COVID-19 patients. We found that some adults with COVID-19 had elevated antibodies to Sm-D3 as to well as Ro52, Ro60, and La. While RNP-A and Sm-D3 autoantibodies usually cluster together in patients with SLE (24, 31), only Sm-D3 autoantibodies were seen in adult COVID-19 patients. Moreover, both KCNRG and Sm-D3 autoantibodies were enriched in COVID-19 adult patients with severe and moderate disease, but not in critically ill COVID-19 patients. While the presence of specific autoantibodies may be driven by the release of the proteins that these antibodies target from damaged or dying cells in combination with immune activation and plasmablast expansion (32), possible explanations for the paradoxical lack of autoantibodies in critically ill patients is that these patients have a immunodeficiency state that can be similarly seen in late stages of sepsis or they might have received immunosuppressive medications that could have blunted their antibody response. A recent study by Su and Yuan, (33) identified many of the same autoantigens in adults reported here, including Ro52 and Ro60 (SSA), La (SSB) and RNP autoantigens, as biomarkers of post-acute sequelae of COVID (PASC/long-COVID), which were independent of COVID-19 infection severity (33). Based on these findings, future studies are needed to assess whether the autoantibodies detected here in adults are long lasting and whether they correlate with long-COVID.
In contrast to adults, most children with COVID-19 demonstrated little or no autoantibodies against KCNRG, Sm-D3, and the other autoantigens we tested. The relative differences in the frequency of autoantibodies between children and adults is likely impacted by age-related differences in immunological function and increased comorbidity seen in adults (8). One 10-year-old girl with COVID-19 was found to have very high levels of GAD65 autoantibodies, a well-known marker of type 1 diabetes. This child presented with ketoacidosis and required insulin upon hospital admission. Although viruses have long been implicated in pancreatic beta cell destruction in type 1 diabetes (34, 35), recent studies show that SARS-CoV-2 can infect, replicate, and destroy beta cells that produce insulin in pancreatic islets (36, 37). SARS-CoV-2 infection has been reported to induce both autoantibody-positive (38) and autoantibody-negative diabetes (37, 39). SARS-CoV-2 infection has also been implicated in triggering the acute onset of several other autoimmune diseases, including encephalitis (40, 41), Guillain-Barre syndrome (42), and myasthenia gravis (43). Based on these and other studies, the finding of SARS-CoV-2 infection associated type 1 diabetes onset further highlights that the virus may be a risk factor associated with the future onset of diabetes or other autoimmune diseases in children.
Children with MIS-C have inflammation involving multiple organ systems, which is usually not observed in children with COVID-19. KD typically present in childhood and shares certain clinical features with MIS-C including fever, skin, mucous inflammation, and increased inflammatory markers (44). One feature of MIS-C that is usually not seen in KD is the high frequency of gastrointestinal symptoms including abdominal pain, vomiting, and diarrhea (45, 46). Several groups have reported the presence of a broad range of tissue-specific and non-organ-specific autoantibodies in MIS-C (16–18). Our initial autoantibody analysis suggested the possibility that there might be autoantibodies in MIS-C cases that were associated with various rheumatological diseases and autoimmune gastritis. However, further evaluation revealed that the autoantibodies against Ro52, Ro60, La, and gastric ATPase in the MIS-C patients’ sera were due to IVIG administration. Approximately 57% of the patients with MIS-C had all four autoantibodies, and the presence of these autoantibodies in MIS-C directly tracked with the administration of IVIG administration.
Multiple lines of evidence established that the autoantibodies against Ro52, Ro60, La, and gastric ATPase seen in MIS-C and KD children was due to IVIG administration. First, serum/plasma samples from both MIS-C and KD patients who did not receive IVIG did not have autoantibodies against Ro52, Ro60, La, and gastric ATPase. In contrast, known KD and MIS-C children who received IVIG, showed a very high frequency (86-100%) of autoantibodies to these autoantigens. The few seronegative cases were likely confounded by different batches of IVIG and collection of serum samples weeks after IVIG administration. Second, our studies with multiple serial samples from both MIS-C and KD before and after IVIG revealed that the autoantibodies were only found after IVIG administration. Third, testing of three different lots of IVIG revealed high levels of autoantibodies against Ro52, Ro60, La and gastric ATPase, although the relative levels of these autoantibodies varied from the different lots of IVIG. These findings of existing autoantibodies in IVIG are also consistent with other published findings reporting autoantibodies in IVIG or in patients who received IVIG (47–51). One explanation for the seropositive autoantibodies in IVIG against these Ro52, Ro60, La and gastric autoantibodies relates to the large IVIG donor pool who harbor high levels of these autoantibodies because they likely include samples from asymptomatic or pre-symptomatic individuals with autoimmune conditions. Our results and work of others (50) also suggests that commercial IVIG preparations may differ in autoantibody concentrations due to differences in donor pools and manufacturing. Taken together these results show that IVIG given to MIS-C patients contains a variety of autoantibodies and without testing before IVIG treatment or allowing sufficient time after the dose of IVIG to allow decay of these antibodies before repeat testing, can be mistakenly interpreted as associated with the manifestations of MIS-C. From our analysis of MIS-C IVIG recipients, Ro52, Ro60, and La autoantibodies showed a similar decay profile, in which all three autoantibodies were present in the blood for several weeks before becoming undetectable by 35-60 days. In contrast, the decay kinetics of autoantibodies directed against the gastric ATPase was much longer (>100 days). One potential explanation for this decay discrepancy is that Ro52, Ro60, and La are intracellular proteins, and the shorter autoantibody decay may reflect limited persistence in specific tissue due to inaccessibility of the antibodies to the intracellular compartment where these antigens are located. In contrast, gastric ATPase is a surface membrane protein that is antibody accessible, which might allow gastric ATPase autoantibodies to bind gastric tissue and thereby slowly be released resulting in persistent presence in the blood. Based on the availability of serial samples from IVIG recipients and additional studies of the decay of both anti-pathogen and autoantibodies present in preparations, further insights may be gained into the therapeutic action of IVIG.
Three published studies have used antigen arrays for autoantibody discovery in MIS-C (16–18). It is worth noting that few overlapping target proteins were identified among the three publications and little validation of the identified target autoantigens by orthogonal techniques was provided to eliminate potential false positives that are known to occur with autoantigen arrays (52). Most of the target autoantigens identified in these array studies were ubiquitous, intracellular proteins and did represent established autoantigens associated with autoimmune disease. Consiglio et al. analyzed serum samples taken exclusively from MIS-C subjects before they received IVIG, and identified several autoantibodies including high levels against three members of the intracellular casein kinase family (CSNK1A, CSNK2A1, and CSNK1E), which were some of the most informative biomarkers (17). However, no validation immunoassays were used to independently confirm the relevance of these targets and the number of negative controls was very limited. Using the sensitive and specific LIPS technology, we were unable to detect any autoantibodies against CSNK1A and LGAL3, another reported target in any of our MIS-C patients not receiving IVIG. These findings raise questions about the usefulness and validity of these autoantibodies as markers of MIS-C.
Two additional antigen array publications also discovered autoantibodies using a small number of MIS-C patients. Gruber et al. identified seropositive IgG autoantibodies against La and Jo-1 associated with known autoimmune diseases in their MIS-C cohort (18). Our autoantibody testing of La showed a complete absence (i.e., 0/21 MIS-C patients) of the antibody in our MIS-C patients who had not received IVIG including some with severe disease. Moreover, we found no evidence of IgG autoantibodies against several other previously reported MIS-C targets including JO-1, ECE1, IL-1A and IFNGR2 receptor, and no IgA autoantibodies against IL-1A, JO-1 and HBV X protein. While an ELISA was used to independently validate one array target, CD244 a potential extracellular protein, additional follow-up assays related to the functional significance of these autoantibodies were not performed and additional experiments are needed to independently validate the presence of these autoantibodies in other MIS-C patients. We also examined IgG and IgA autoantibodies against proteins identified by the third autoantigen array study (16) using LIPS. No patients with MIS-C who did not receive IVIG had IgG autoantibodies against most of the previously reported proteins including ANX11, ATP4A, ECE1, FAM84A, NAPB, TROVE2 (Ro60), PDLIM5, and UBE3A, and none had IgA autoantibodies against FAM84A and TNNC2 with one exception. Ten percent of the MIS-C patients had antibodies to TNNC2. In addition, 5% of patients had antibodies against KLHL12, a known autoantigen in Sjögren’s syndrome and primary biliary cirrhosis. One patient had antibodies to both KLHL12 and TNNC2 which might have been due to an underlying autoimmune condition, or a predisposition to autoantibodies that was triggered by COVID-19.
In conclusion, our study found little evidence that children with MIS-C or COVID-19 produce autoantibodies associated with known autoimmune diseases. Moreover, from testing 18 candidate targets previously reported by protein microarrays from patients with MIS-C (16–18), we were unable to confirm elevated levels of these antibodies in MIS-C in our LIPS assays. We found antibodies to two of these proteins in a minority of our patients. While we did not test all the autoantigens reported by the array studies, many of the targets we tested were postulated to be the most informative. Our inability to replicate the reportedly broad autoantibody immunoreactivity seen in MIS-C with our assays casts in doubts the role of these autoantibodies as biomarkers in MIS-C. Furthermore, the lack of studies reporting autoantibodies in MIS-C against several common antigens (e.g., Ro60/Trove2 [part of SSA], La [SSB], JO-1, gastric ATPase and for HBV infection), which are widely available as clinical immunoassays, provides circumstantial evidence against the usefulness of these biomarkers in MIS-C and KD. Future studies focusing on identifying autoantibodies directed against extracellular protein targets may be particularly informative due the possibility that they might have a pathogenic role. Along these lines, Ramaswamy et al. using flow cytometry found evidence for MIS-C autoantibody binding to activated endothelial cells relevant for severe disease pathology, yet no specific autoantigen targets were identified (20). In light of recent studies showing increased somatic hypermutation in plasmablasts of MIS-C (20, 53) additional studies are needed in MIS-C to determine if specific, yet to be identified, autoantibodies play a role in immune complex formation or autoantibody-driven damage to host cells.
Pediatric Emergency Medicine Kawasaki Group
Naomi Abe, Amy Bryl, J. Joelle Donofrio-Odmann, Atim Ekpenyong, Michael Gardiner, David J. Gutglass, Margaret B. Nguyen, Stacey Ulrich.
Department of Pediatrics, Rady Children’s Hospital, University of California San Diego, San Diego, CA, USA.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The studies involving human participants were reviewed and approved by under an NIH IRB exemption after local IRB approval: Brescia, Italy Comitato Etico Provinciale (NP 4000 and NP 4408); Pavia, Italy (Comitato Etico Pavia Prot. 20200037677); Turin, Italy (Comitato Etico Interaziendale A.O.U. Città della Salute e della Scienza di Torino, Protocol: 00282/2020); Naples, Italy (Ethics Committee of the University of Naples Federico II, Protocol: 158/20); Chile (Comité Ético Científico Facultad de Medicina Clínica Alemana Universidad del Desarrollo, Santiago, Chile, Protocol: 2020-41); and Israel (Hadassah Medical Organization Institutional Review Board (IRB) for studies involving human subjects, Protocol: HMO-235-20). Additional MIS-C patients were enrolled from other sources including Stanford, California and Portland, Oregon, after written informed consent to NIH IRB-approved research protocols (NIH Protocols l1-1-0109, 18-I-0041 and 18-I-0128). Sera from children with KD or acute febrile illnesses were obtained at the University of California, San Diego under IRB approved protocols (UCSD #140220). All patients and parents gave written informed consent and assent as appropriate. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
PB, JB, LDN, and JIC conceived and designed the study. CS, OD, VD, ALV, AG, FL, UR, ER-J, CV, GLM, AL, DM, CR, YE, LN, LD, MT, DG, SM, JK, AT, EE, LI, MP, and JB consented patients and provided biological specimens. HS, RC, and KD coordinated the collection and processing of the MIS-C samples. PB performed antibody profiling experiments and drafted the manuscript. RC, GMS, and KB reviewed clinical metadata. RC and BW analyzed results for tables and figure. PB, JC, JB, LDN, and JIC critically reviewed and revised the draft of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported in part by the Intramural Research Programs of NIDCR and NIAID of the National Institutes of Health and Regione Lombardia (project “Immune response in patients with COVID-19 and co-morbidities”), FONDECYT 11181222 (MP), ANID-COVID19 0999 (MV) and NIH 3R01HL140898-03S1 and NICHD 1R61HD105590 (JB and AT). We thank the NIAID Office of Cyber Infrastructure and Computational Biology, Bioinformatics and Computational Biosciences Branch (Contract HHSN316201300006W/HHSN27200002 to MSC, Inc) and Operations Engineering Branch for developing the HGRepo system to enable streamlined access to the data and the NCI Advanced Biomedical Computational Science (ABCS) for data transformation support. This research was also supported in part by the NIDCR Combined Technical Research Core (ZIC DE000729-09).
Author Disclaimer
The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, et al. A Novel Coronavirus From Patients With Pneumonia in China, 2019. N Engl J Med (2020) 382(8):727–33. doi: 10.1056/NEJMoa2001017
2. Richardson S, Hirsch JS, Narasimhan M, Crawford JM, McGinn T, Davidson KW, et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA (2020) 323(20):2052–9. doi: 10.1001/jama.2020.6775
3. Godri Pollitt KJ, Peccia J, Ko AI, Kaminski N, Dela Cruz CS, Nebert DW, et al. COVID-19 Vulnerability: The Potential Impact of Genetic Susceptibility and Airborne Transmission. Hum Genomics (2020) 14(1):17. doi: 10.1186/s40246-020-00267-3
4. Tay MZ, Poh CM, Renia L, MacAry PA, Ng LFP. The Trinity of COVID-19: Immunity, Inflammation and Intervention. Nat Rev Immunol (2020) 20(6):363–74. doi: 10.1038/s41577-020-0311-8
5. Casanova JL, Su HC, Effort CHG. A Global Effort to Define the Human Genetics of Protective Immunity to SARS-CoV-2 Infection. Cell (2020) 181(6):1194–9. doi: 10.1016/j.cell.2020.05.016
6. Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, Chen J, et al. Inborn Errors of Type I IFN Immunity in Patients With Life-Threatening COVID-19. Science (2020) 370(6515):eabd4570. doi: 10.1126/science.abd4570
7. Bastard P, Gervais A, Le Voyer T, Rosain J, Philippot Q, Manry J, et al. Autoantibodies Neutralizing Type I IFNs are Present in ~4% of Uninfected Individuals Over 70 Years Old and Account for ~20% of COVID-19 Deaths. Sci Immunol (2021) 6(62):eabl4340. doi: 10.1126/sciimmunol.abl4340
8. Wong LSY, Loo EXL, Kang AYH, Lau HX, Tambyah PA, Tham EH. Age-Related Differences in Immunological Responses to SARS-CoV-2. J Allergy Clin Immunol Pract (2020) 8(10):3251–8. doi: 10.1016/j.jaip.2020.08.026
9. Castagnoli R, Votto M, Licari A, Brambilla I, Bruno R, Perlini S, et al. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection in Children and Adolescents: A Systematic Review. JAMA Pediatr (2020) 174(9):882–9. doi: 10.1001/jamapediatrics.2020.1467
10. Sharma C, Ganigara M, Galeotti C, Burns J, Berganza FM, Hayes DA, et al. Multisystem Inflammatory Syndrome in Children and Kawasaki Disease: A Critical Comparison. Nat Rev Rheumatol (2021) 17(12):731–48. doi: 10.1038/s41584-021-00709-9
11. McMurray JC, May JW, Cunningham MW, Jones OY. Multisystem Inflammatory Syndrome in Children (MIS-C), a Post-Viral Myocarditis and Systemic Vasculitis-A Critical Review of Its Pathogenesis and Treatment. Front Pediatr (2020) 8:626182. doi: 10.3389/fped.2020.626182
12. Nimmerjahn F, Ravetch JV. Anti-Inflammatory Actions of Intravenous Immunoglobulin. Annu Rev Immunol (2008) 26:513–33. doi: 10.1146/annurev.immunol.26.021607.090232
13. Galani IE, Rovina N, Lampropoulou V, Triantafyllia V, Manioudaki M, Pavlos E, et al. Untuned Antiviral Immunity in COVID-19 Revealed by Temporal Type I/III Interferon Patterns and Flu Comparison. Nat Immunol (2021) 22(1):32–40. doi: 10.1038/s41590-020-00840-x
14. Notz Q, Schmalzing M, Wedekink F, Schlesinger T, Gernert M, Herrmann J, et al. Pro- and Anti-Inflammatory Responses in Severe COVID-19-Induced Acute Respiratory Distress Syndrome-An Observational Pilot Study. Front Immunol (2020) 11:581338. doi: 10.3389/fimmu.2020.581338
15. Zeng Z, Yu H, Chen H, Qi W, Chen L, Chen G, et al. Longitudinal Changes of Inflammatory Parameters and Their Correlation With Disease Severity and Outcomes in Patients With COVID-19 From Wuhan, China. Crit Care (2020) 24(1):525. doi: 10.1186/s13054-020-03255-0
16. Porritt RA, Binek A, Paschold L, Rivas MN, McArdle A, Yonker LM, et al. The Autoimmune Signature of Hyperinflammatory Multisystem Inflammatory Syndrome in Children. J Clin Invest (2021) 131(20):e151520. doi: 10.1172/JCI151520
17. Consiglio CR, Cotugno N, Sardh F, Pou C, Amodio D, Rodriguez L, et al. The Immunology of Multisystem Inflammatory Syndrome in Children With COVID-19. Cell (2020) 183(4):968–81.e7. doi: 10.1016/j.cell.2020.09.016
18. Gruber CN, Patel RS, Trachtman R, Lepow L, Amanat F, Krammer F, et al. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (MIS-C). Cell (2020) 183(4):982–95.e14. doi: 10.1016/j.cell.2020.09.034
19. Zuo Y, Estes SK, Ali RA, Gandhi AA, Yalavarthi S, Shi H, et al. Prothrombotic Autoantibodies in Serum From Patients Hospitalized With COVID-19. Sci Transl Med (2020) 12(570):eabl4340. doi: 10.1126/scitranslmed.abd3876
20. Ramaswamy A, Brodsky NN, Sumida TS, Comi M, Asashima H, Hoehn KB, et al. Immune Dysregulation and Autoreactivity Correlate With Disease Severity in SARS-CoV-2-Associated Multisystem Inflammatory Syndrome in Children. Immunity (2021) 54(5):1083–95.e7. doi: 10.1016/j.immuni.2021.04.003
21. Wang EY, Mao T, Klein J, Dai Y, Huck JD, Jaycox JR, et al. Diverse Functional Autoantibodies in Patients With COVID-19. Nature (2021) 595(7866):283–8. doi: 10.1038/s41586-021-03631-y
22. Diagnosis and Treatment Protocol for Novel Coronavirus Pneumonia (Trial Version 7). Chin Med J (Engl) (2020) 133(9):1087–95. doi: 10.1097/CM9.0000000000000819
23. Burbelo PD, Lebovitz EE, Notkins AL. Luciferase Immunoprecipitation Systems for Measuring Antibodies in Autoimmune and Infectious Diseases. Transl Res (2015) 165(2):325–35. doi: 10.1016/j.trsl.2014.08.006
24. Ching KH, Burbelo PD, Tipton C, Wei C, Petri M, Sanz I, et al. Two Major Autoantibody Clusters in Systemic Lupus Erythematosus. PloS One (2012) 7(2):e32001. doi: 10.1371/journal.pone.0032001
25. Lahner E, Marzinotto I, Lampasona V, Dottori L, Bazzigaluppi E, Brigatti C, et al. Autoantibodies Toward ATP4A and ATP4B Subunits of Gastric Proton Pump H+,K+-ATPase Are Reliable Serological Pre-Endoscopic Markers of Corpus Atrophic Gastritis. Clin Transl Gastroenterol (2020) 11(10):e00240. doi: 10.14309/ctg.0000000000000240
26. Ferre EMN, Break TJ, Burbelo PD, Allgauer M, Kleiner DE, Jin D, et al. Lymphocyte-Driven Regional Immunopathology in Pneumonitis Caused by Impaired Central Immune Tolerance. Sci Transl Med (2019) 11(495). doi: 10.1126/scitranslmed.aav5597
27. Burbelo PD, Lebovitz EE, Bren KE, Bayat A, Paviol S, Wenzlau JM, et al. Extrapancreatic Autoantibody Profiles in Type I Diabetes. PloS One (2012) 7(9):e45216. doi: 10.1371/journal.pone.0045216
28. Norman GL, Yang CY, Ostendorff HP, Shums Z, Lim MJ, Wang J, et al. Anti-Kelch-Like 12 and Anti-Hexokinase 1: Novel Autoantibodies in Primary Biliary Cirrhosis. Liver Int (2015) 35(2):642–51. doi: 10.1111/liv.12690
29. Uchida K, Akita Y, Matsuo K, Fujiwara S, Nakagawa A, Kazaoka Y, et al. Identification of Specific Autoantigens in Sjogren’s Syndrome by SEREX. Immunology (2005) 116(1):53–63. doi: 10.1111/j.1365-2567.2005.02197.x
30. Burbelo PD, Seam N, Groot S, Ching KH, Han BL, Meduri GU, et al. Rapid Induction of Autoantibodies During ARDS and Septic Shock. J Transl Med (2010) 8:97. doi: 10.1186/1479-5876-8-97
31. To CH, Petri M. Is Antibody Clustering Predictive of Clinical Subsets and Damage in Systemic Lupus Erythematosus? Arthritis Rheum (2005) 52(12):4003–10. doi: 10.1002/art.21414
32. Mathew D, Giles JR, Baxter AE, Oldridge DA, Greenplate AR, Wu JE, et al. Deep Immune Profiling of COVID-19 Patients Reveals Distinct Immunotypes With Therapeutic Implications. Science (2020) 369(6508). doi: 10.1126/science.abc8511
33. Su Y, Yuan D, Chen DG, Ng RH, Wang K, Choi J, et al. Multiple Early Factors Anticipate Post-Acute COVID-19 Sequelae. Cell (2022) 185(5):881–95.e20. doi: 10.1016/j.cell.2022.01.014
34. Coppieters KT, Boettler T, von Herrath M. Virus Infections in Type 1 Diabetes. Cold Spring Harb Perspect Med (2012) 2(1):a007682. doi: 10.1101/cshperspect.a007682
35. Spagnuolo I, Patti A, Sebastiani G, Nigi L, Dotta F. The Case for Virus-Induced Type 1 Diabetes. Curr Opin Endocrinol Diabetes Obes (2013) 20(4):292–8. doi: 10.1097/MED.0b013e328362a7d7
36. Muller JA, Gross R, Conzelmann C, Kruger J, Merle U, Steinhart J, et al. SARS-CoV-2 Infects and Replicates in Cells of the Human Endocrine and Exocrine Pancreas. Nat Metab (2021) 3(2):149–65. doi: 10.1038/s42255-021-00347-1
37. Liu F, Long X, Zhang B, Zhang W, Chen X, Zhang Z. ACE2 Expression in Pancreas May Cause Pancreatic Damage After SARS-CoV-2 Infection. Clin Gastroenterol Hepatol (2020) 18(9):2128–30.e2. doi: 10.1016/j.cgh.2020.04.040
38. Marchand L, Pecquet M, Luyton C. Type 1 Diabetes Onset Triggered by COVID-19. Acta Diabetol (2020) 57(10):1265–6. doi: 10.1007/s00592-020-01570-0
39. Hollstein T, Schulte DM, Schulz J, Gluck A, Ziegler AG, Bonifacio E, et al. Autoantibody-Negative Insulin-Dependent Diabetes Mellitus After SARS-CoV-2 Infection: A Case Report. Nat Metab (2020) 2(10):1021–4. doi: 10.1038/s42255-020-00281-8
40. Pilotto A, Masciocchi S, Volonghi I, Crabbio M, Magni E, De Giuli V, et al. Clinical Presentation and Outcomes of Severe Acute Respiratory Syndrome Coronavirus 2-Related Encephalitis: The ENCOVID Multicenter Study. J Infect Dis (2021) 223(1):28–37. doi: 10.1093/infdis/jiaa609
41. Ellul MA, Benjamin L, Singh B, Lant S, Michael BD, Easton A, et al. Neurological Associations of COVID-19. Lancet Neurol (2020) 19(9):767–83. doi: 10.1016/S1474-4422(20)30221-0
42. Fragiel M, Miro O, Llorens P, Jimenez S, Pinera P, Burillo G, et al. Incidence, Clinical, Risk Factors and Outcomes of Guillain-Barre in Covid-19. Ann Neurol (2021) 89(3):598–603. doi: 10.1002/ana.25987
43. Assini A, Gandoglia I, Damato V, Rikani K, Evoli A, Del Sette M. Myasthenia Gravis Associated With Anti-MuSK Antibodies Developed After SARS-CoV-2 Infection. Eur J Neurol (2021) 28(10):3537–9. doi: 10.1111/ene.14721
44. Kabeerdoss J, Pilania RK, Karkhele R, Kumar TS, Danda D, Singh S. Severe COVID-19, Multisystem Inflammatory Syndrome in Children, and Kawasaki Disease: Immunological Mechanisms, Clinical Manifestations and Management. Rheumatol Int (2021) 41(1):19–32. doi: 10.1007/s00296-020-04749-4
45. Dufort EM, Koumans EH, Chow EJ, Rosenthal EM, Muse A, Rowlands J, et al. Multisystem Inflammatory Syndrome in Children in New York State. N Engl J Med (2020) 383(4):347–58. doi: 10.1056/NEJMoa2021756
46. Feldstein LR, Rose EB, Horwitz SM, Collins JP, Newhams MM, Son MBF, et al. Multisystem Inflammatory Syndrome in U.S. Children and Adolescents. N Engl J Med (2020) 383(4):334–46. doi: 10.1056/NEJMoa2021680
47. van der Molen RG, Hamann D, Jacobs JF, van der Meer A, de Jong J, Kramer C, et al. Anti-SSA Antibodies Are Present in Immunoglobulin Preparations. Transfusion (2015) 55(4):832–7. doi: 10.1111/trf.12922
48. Gruter T, Ott A, Meyer W, Jarius S, Kinner M, Motte J, et al. Effects of IVIg Treatment on Autoantibody Testing in Neurological Patients: Marked Reduction in Sensitivity But Reliable Specificity. J Neurol (2020) 267(3):715–20. doi: 10.1007/s00415-019-09614-4
49. Smith TD, Cunningham-Rundles C. Detection of Anti-Glutamic Acid Decarboxylase Antibodies in Immunoglobulin Products. J Allergy Clin Immunol Pract (2018) 6(1):260–1. doi: 10.1016/j.jaip.2017.04.042
50. Khan S, Sims S, Raine D, Sewell WA. Both Autoantibodies and Pathogen-Specific Antibodies are Present in Immunoglobulin Preparations and Reflect Characteristics of the Donor Population. J Am Acad Dermatol (2008) 59(6):1089–90. doi: 10.1016/j.jaad.2008.07.048
51. Khan S, Davies L, Cowley D, Wild G, Sewell WA. Anti-Acetylcholine Receptor Antibody Reactivity of IgG in Commercial Immunoglobulin Preparations. Clin Neurol Neurosurg (2010) 112(9):835–6. doi: 10.1016/j.clineuro.2010.06.014
52. Ayoglu B, Schwenk JM, Nilsson P. Antigen Arrays for Profiling Autoantibody Repertoires. Bioanalysis (2016) 8(10):1105–26. doi: 10.4155/bio.16.31
Keywords: autoantibodies, MIS-C multisystem inflammatory syndrome in children, IVIG, autoimmunity, COVID-19
Citation: Burbelo PD, Castagnoli R, Shimizu C, Delmonte OM, Dobbs K, Discepolo V, Lo Vecchio A, Guarino A, Licciardi F, Ramenghi U, Rey-Jurado E, Vial C, Marseglia GL, Licari A, Montagna D, Rossi C, Montealegre Sanchez GA, Barron K, Warner BM, Chiorini JA, Espinosa Y, Noguera L, Dropulic L, Truong M, Gerstbacher D, Mató S, Kanegaye J, Tremoulet AH, Pediatric Emergency Medicine Kawasaki Group, Eisenstein EM, Su HC, Imberti L, Poli MC, Burns JC, Notarangelo LD and Cohen JI (2022) Autoantibodies Against Proteins Previously Associated With Autoimmunity in Adult and Pediatric Patients With COVID-19 and Children With MIS-C. Front. Immunol. 13:841126. doi: 10.3389/fimmu.2022.841126
Received: 21 December 2021; Accepted: 16 February 2022;
Published: 11 March 2022.
Edited by:
Luca Quartuccio, University of Udine, ItalyReviewed by:
Donato Amodio, Bambino Gesù Children’s Hospital (IRCCS), ItalyJagadeesh Bayry, Indian Institute of Technology Palakkad, India
Copyright © 2022 Burbelo, Castagnoli, Shimizu, Delmonte, Dobbs, Discepolo, Lo Vecchio, Guarino, Licciardi, Ramenghi, Rey-Jurado, Vial, Marseglia, Licari, Montagna, Rossi, Montealegre Sanchez, Barron, Warner, Chiorini, Espinosa, Noguera, Dropulic, Truong, Gerstbacher, Mató, Kanegaye, Tremoulet, Pediatric Emergency Medicine Kawasaki Group, Eisenstein, Su, Imberti, Poli, Burns, Notarangelo and Cohen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter D. Burbelo, YnVyYmVsb3BAbmlkY3IubmloLmdvdg==
†These authors have contributed equally to this work