
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 22 April 2022
Sec. Autoimmune and Autoinflammatory Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.809590
This article is part of the Research Topic Adhesion Molecules and Autoimmune Diseases View all 11 articles
 Carla Guenther1,2,3*
Carla Guenther1,2,3*Leukocyte trafficking is an essential process of immunity, occurring as leukocytes travel within the bloodstream and as leukocyte migration within tissues. While it is now established that leukocytes can utilize the mesenchymal migration mode or amoeboid migration mode, differences in the migratory behavior of leukocyte subclasses and how these are realized on a molecular level in each subclass is not fully understood. To outline these differences, first migration modes and their dependence on parameters of the extracellular environments will be explained, as well as the intracellular molecular machinery that powers migration in general. Extracellular parameters are detected by adhesion receptors such as integrins. β2-integrins are surface receptors exclusively expressed on leukocytes and are essential for leukocytes exiting the bloodstream, as well as in mesenchymal migration modes, however, integrins are dispensable for the amoeboid migration mode. Additionally, the balance of different RhoGTPases – which are downstream of surface receptor signaling, including integrins – mediate formation of membrane structures as well as actin dynamics. Individual leukocyte subpopulations have been shown to express distinct RhoGTPase profiles along with their differences in migration behavior, which will be outlined. Emerging aspects of leukocyte migration include signal transduction from integrins via actin to the nucleus that regulates DNA status, gene expression profiles and ultimately leukocyte migratory phenotypes, as well as altered leukocyte migration in tumors, which will be touched upon.
Leukocyte trafficking is a multifaceted process that not only encompasses the cell adhesion cascade, transmigration and migration through tissues, but also gains further depth by the fact that leukocytes can use the mesenchymal migration mode as well as a unique amoeboid migration mode that sets them apart from other cell types. Additionally, our understanding of leukocyte migration is rapidly evolving to the extent that not differentiating between these aspects – be it transmigration, crawling on 2D or migration through 3D environments – and instead refering to them in equal fashion as ‘leukocyte migration’ hinders us from appreciating the true complexity of these processes. The importance of this is heightened by the fact that leukocyte subpopulations show distinct migratory behavior across all these aspects. Additionally, while it is now appreciated that environmental parameters dictate which migration mode is possible for all leukocytes and these parameters are integrated into the cell by adhesion receptors such as β2-integrins, it is becoming clear that how the appropriate migration modes are executed is down to a distinct combination of β2-integrin downstream components which are, in turn, acting uniquely and are expressed in different leukocyte populations. It is clear that our collective understanding of leukocyte migration is just scratching the surface and this is due to several factors. One hindering factor is that not all aspects of leukocyte migration are researched to the same degree. Transwell migration assays represent the vast majority of assays used to study leukocyte migration; this means we have much more data on transmigration than on migration in tissues. Additionally, the 2D or 3D migration assays which are carried out often rely on single-protein coatings or matrices, respectively, and this fails to represent the complexity of living endothelium and tissues. Furthermore, the structural organization of proteins in bundles or fibers is neglected, just as is their stiffness and elasticity. Finally, leukocyte subpopulations such as neutrophils, macrophages and T-cells have been studied most, while eosinophils and basophils have been almost entirely ignored. This review seeks to outline some of the field’s complexity and why the aforementioned aspects of the environment are important, in the context of β2-integrins, which are essential for the leukocyte adhesion cascade.
The majority of leukocyte trafficking occurs in the bloodstream, in which leukocytes – typically T- and B-cells, neutrophils, monocytes, eosinophils and basophils – are passively carried by the bloodstream close to sites of infections. In response to infections, the endothelium lining the blood vessels closest to the site of infection expresses chemokines and adhesion receptor ligands which initiate and mediate the cell adhesion cascade. This cascade is the process through which leukocytes leave the bloodstream and extravasate into the tissue. In this multistep process, leukocytes first tether on the endothelium, followed by fast and slow rolling along the vessel wall until the leukocyte is arrested on the endothelium (1, 2). Leukocytes then typically begin to spread and crawl across the endothelium (3). The distinctive steps in the adhesion cascade are: step 1 – tethering and fast rolling, step 2 – slow rolling and arrest, step 3 – spreading and crawling, step 4 – transmigration. The adhesion cascade step of crawling across the endothelium has been termed locomotion (3). This process is depicted in Figure 1 and has been reviewed multiple times in detail [for example (4)]. Selectins are the first adhesion receptors mediating tethering and fast rolling during which their activity typically overlaps with β2-integrins (4). β2-integrins are especially crucial for the fast and slow rolling, cell arrest, and spreading and crawling steps (4). They have been identified as mechanoreceptors (5) and therefore likely play a role in probing the endothelium for points of exit as well. Further detail in regards to involvement of each adhesion receptor at each step and their typical ligands are depicted and described in Figure 1.
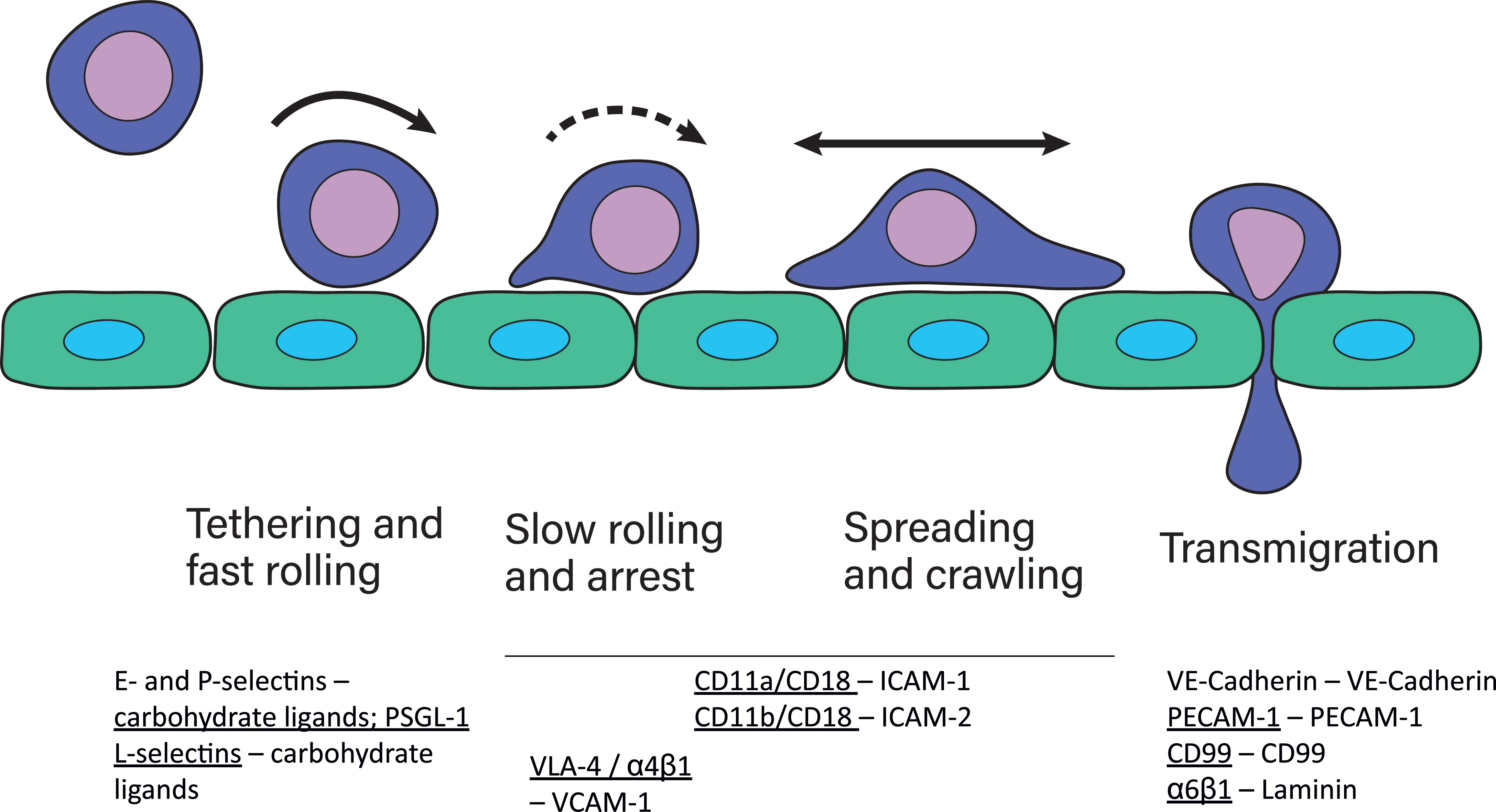
Figure 1 The cell adhesion cascade. During adhesion to and extravasation through the endothelium, leukocytes undergo several steps, namely fast rolling and tethering, slow rolling and arrest, spreading and crawling and lastly transmigration. These steps are mediated by several different adhesion receptors expressed by leukocytes, indicated by underscore, and the endothelium (2). P-selectins and E-selectins expressed by the endothelium bind to their carbohydrate ligands on leukocytes during fast rolling. Leukocytes express L-selectins, which bind to ligands on the endothelium during fast rolling. The most well known selectin ligand is P-selectin glycoprotein ligand-1 (PSGL-1). Selectin-ligand binding results in integrin activation in leukocytes, which then mediates slow rolling, arrest, spreading and crawling. During transmigration, several homophilic adhesion receptors, including VE-cadherins important for tight junctions between endothelial cells, and CD99 and PECAM-1 are expressed by endothelial cells and leukocytes. Additionally, leukocytes express α6β1 integrins that bind to laminin.
β2-integrins are a group of essential adhesion receptors, consisting of CD11a/CD18 (LFA-1, αLβ2), CD11b/CD18 (MAC1, αMβ2), CD11c/CD18 (αXβ2), and CD11d/CD18 (αDβ2) (6). Leukocytes express several different types of integrins (7) (Table 1 and Figure 2), and they express β2- and β7-integrins as the only cells [β2-integrins are also expressed on platelets (28)]. Furthermore, in most circulating leukocytes β2-integrins are the integrin class expressed in the highest amounts (13). β2-integrins are differently expressed on different leukocytes and they differ in terms of ligand binding. CD11a and CD11d bind limited ligands (CD11a/ICAMs, CD11d/ICAM-3, VCAM-1), while CD11c and CD11b form diverse receptor–ligand interactions with more than 40 identified ligands. β2-integrins have been shown to be essential for pro- as well as anti-inflammatory processes (19, 29–31).
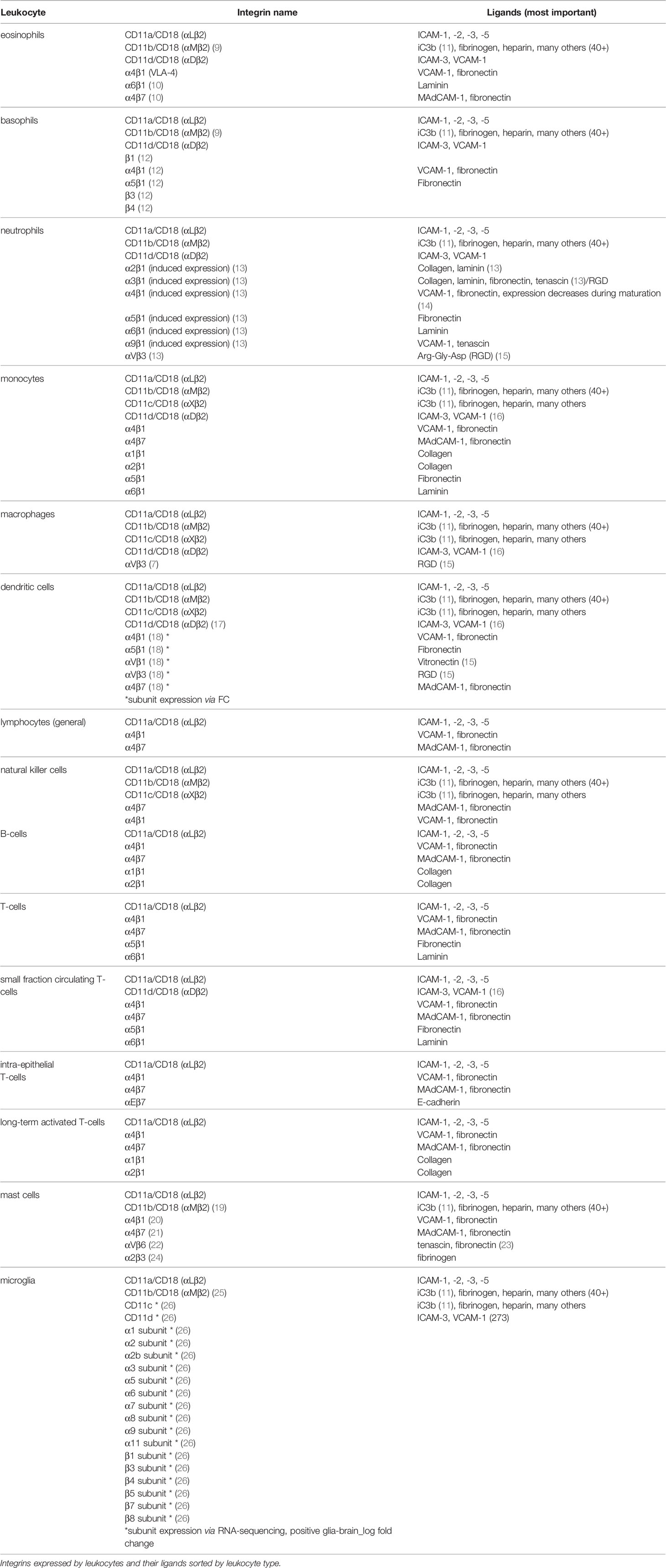
Table 1 Leukocyte Integrins And Their Ligands, Expanded On From (8).
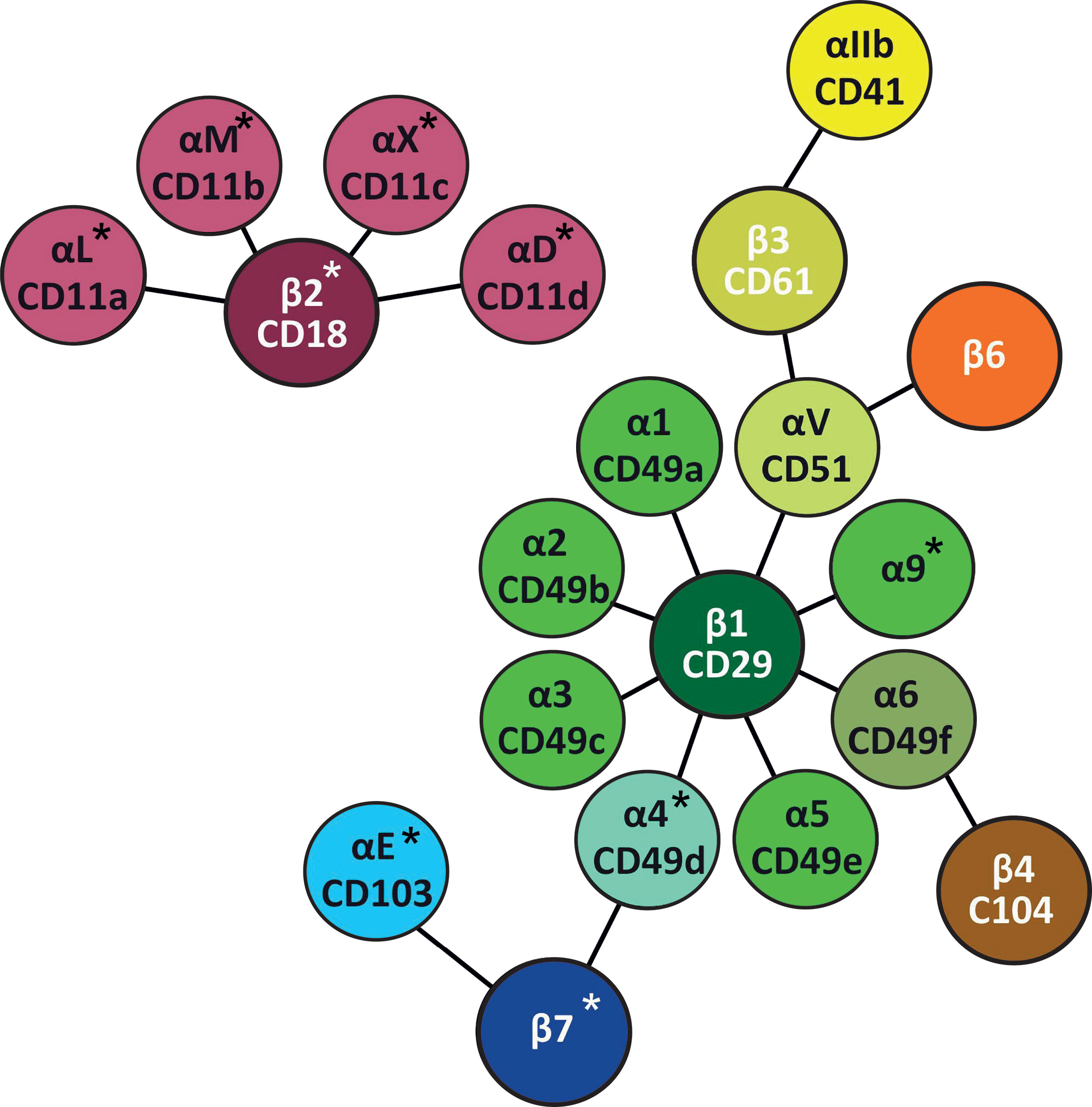
Figure 2 Leukocyte integrins. Leukocyte integrins arranged around their β chains. α and β chains marked with * are specific for leukocytes (27), while platelets also express β2-integrins (28).
Integrins can signal from the inside to the outside, which is especially important during the cell adhesion cascade. A central aspect of such signaling is typically conformational changes of integrins, which in turn regulate the individual bond strength between individual integrin heterodimers and ligands (30, 32). Integrins can only bind ligands if they are in an active – meaning open, extended (high affinity) conformation (33) (depicted in Figure 3). A ‘bent’, inactive integrin conformation (33) predominates when leukocytes travel through the bloodstream to prevent cell aggregation (19). For example, a 9:1 ratio between inactive and active conformations was established for a mutant form of CD11b in the absence of ligands (34, 35). When leukocytes bump into endothelial cells producing chemokines and expressing selectin ligands, chemokine-receptor and selectin engagement initiates the binding of talin and kindlin to integrin cytoplasmic tails (36, 37). This leads to integrin switching from the bent into the extended conformation, akin to a switch blade, and is an example of integrin inside-out signaling. This conformational switch reveals ligand binding sites in the extracellular domain in parts that were facing the cell membrane in the bent conformation (38). At the same time, the conformational switch leads to integrin cytoplasmic tail separation which enables the binding of further interaction partners like tensin and vinculin (39).

Figure 3 Integrin conformation. In leukocytes traveling through the bloodstream, in absence of ligands, 90% of integrins are in the bent inactive conformation, depicted on the left. Inside-out signaling as a result of chemokine receptor/ligand and selectin/ligand binding during fast rolling results in talin and kindlin-3 binding to β chains which results in unbending and changing to the active conformation (right). The unbending results in ligand binding sites being revealed.
During crawling and spreading, leukocytes probe the endothelium for suitable sites of exit, through which they transmigrate. Such sites of exit can be cell–cell junctions, but they also travel through endothelial cell bodies (1, 4, 40, 41). Monocytes have been shown to typically choose gap-junctions to transmigrate. If crawling/locomotion is disabled, for example due to CD11b/CD18 knockdown, neutrophils transmigrate transcellularly, meaning through endothelial cells (3). During transmigration, leukocytes change their integrin expression profiles, for example α6β1-integrin is upregulated, which mediates transmigration (4). Transmigration is the migration process modeled best in transwell/Boyden chamber assays (42). This assay consists of two champers, one filled with normal media and one with media including chemokines. These chambers are separated by a porous membrane, which creates a chemokine gradient (43). However, these assays fail to present leukocyte–endothelial cell interactions as well as gap junctions. Based on the number of papers tagged with ‘transwell leukocyte migration’ vs ‘2D leukocyte migration’ or ‘3D leukocyte migration’ in Pubmed to date, transwell assays are the most used assay to investigate leukocyte migration, however, data generated with this model is often referred to as ‘migration’ or ‘chemotaxis’ instead of ‘transmigration’.
After transmigrating, leukocytes proceed to actively migrate through tissues towards the site of infection, guided by chemokines. This means all migrating leukocytes navigate numerous tissues that present different microenvironments and necessitate different migration modes. This is especially true for leukocytes that migrate long distances. Leukocytes, that have transmigrated from the bloodstream, however typically migrate relatively short distances. For example, the typical maximum distance of (tissue) cells to their nearest blood vessel is 100 µm and rarely exceeds 200 µm (44), and leukocytes likely don’t migrate into other tissues or microenvironments. Leukocytes that actively migrate long distances across several microenvironments are typically antigen–presenting cells, such as dendritic cells, neutrophils and macrophages (45, 46), which migrate through the tissues towards and transmigrate into the lymph system. The lymph system is closely related to and connected to the blood system, however, lymph flow rates are much lower. For example, in mice the blood flow rate in microvessels (45.8 µm diameter) in the retina can be up to 1.55 µl/min (47), while in the lymph system a flow rate of 0.3 µl/h has been measured (48). The slow flow rates necessitate the active migration of leukocytes to the nearest draining lymph node to present antigens. Besides antigen presenting cells, high numbers of T-cells and low numbers of monocytes, all types of granulocytes, and B-cells have also been found in afferent lymph vessels (49–53), however, it is unclear how much active migration is involved in the presence of these leukocytes. Additionally, mast cells migrate from the skin to lymph nodes after UV exposure (54).
The most obvious and most acknowledged difference in the microenvironments that leukocytes migrate through is dimensionality. Crawling on the endothelium of blood- or lymph vessels happens in 2D environments (4, 55) and migration through tissues happens in 3D environments (56). While direct in vivo microscopy has shed much light on migratory processes (57, 58), the molecular mechanisms are often investigated using in vitro models. 2D migration is often modeled in assays using Zigmond or Dunn chambers, which involves two or three tissue plastic chambers, one or two of which are filled with media containing chemokines and one without chemokines to establish a chemokine gradient (59). Since 2D migration in vivo typically happens during crawling on the endothelium (4, 55), leukocyte–endothelial cell interactions are neglected in these models as are shear forces from the bloodstream.
3D migration is typically investigated using collagen or matrigel matrices in which cells are embedded (60). These matrices can be sandwiched between chambers of media with different initial chemokine concentrations, establishing a chemokine gradient. One example of a commercially-available setup is ‘µ-Slides Chemotaxis’ by ibidi (59). Additionally gel matrices can be set atop transwells to first model 3D migration and then transmigration (61), however, direct observation of 3D migration would be difficult. The downside of collagen and matrigel matrices is that they fail to replicate the complexity of tissues, be it in structure or in the variety of components. Cancer spheroids are used to model leukocyte invasion into tumors (62). These are often limited to cancer cells and fail to account for the stroma that support cancer progression. Ex vivo models are also used, in which mice ears are utilized that allow direct observation by microscopy of fluorescently labeled leukocytes (63). Mice ears offer few specific tissue environments for leukocytes to migrate in and would not be comparable to other tissues and organs.
On 2D surfaces such as the endothelium of blood and lymph vessels (55), leukocytes use the mesenchymal migration mode, which is heavily dependent on integrins. Monocyte crawling on the endothelium of blood vessels is particularly dependent on CD11b/CD18 and ligand ICAM-1 (3, 64).
The molecular mechanism of the mesenchymal migration mode is the following: Integrins establish adhesion sites at the edge of the cell in the direction the cell is migrating. At this leading edge, lamellipodia are formed, that contain adhesion receptors, which bind ligands and transduce pulling forces (65). The cell then pulls its body towards this adhesion site. Meanwhile, at the cell rear the adhesion sites are broken and recycled to the front. The force required for the cell’s movement is generated by the actin filament treadmill. This treadmill consists of actin polymerization at the leading cell edge and actin depolymerization at the cell rear (66–68). Actin polymerization ultimately generates power by pushing against elastic proteins that mediate the force by unbending at the cell front (69). The actin network as a whole presses against the cell’s leading edge and the membrane that forms the lamellipodia containing the adhesion receptors (65).
Leukocytes migrate differently in 3D. In particular, dendritic cells (during homeostasis) (18, 53) and neutrophils (70) have been shown to use an amoeboid migration mode independent of integrins. So instead of the actin treadmill, this mode is facilitated by actin polymerization and squeezing and pushing (18). In a groundbreaking study, Lämmermann et al. knocked down all integrins in dendritic cells and proved that this migration mode is independent of integrin-mediated adhesion (18). Instead of lamellipodia, such migrating cells form blebs that do not strongly adhere or pull on the substrate (18, 71–73).
Mechanistically, this migration mode is powered by myosin II filaments sliding along actin filaments and thereby generating contractile forces (65). These contractile forces are present in the cell’s rear and push the cytoplasmic fluid (74), as well as move the cell body (71) forward. At a molecular level, actin and myosin II form the actomyosin complex. In this complex, myosin is bound and travels along actin filaments in an ATP-dependent manner. Myosin travels by binding to consecutive actin binding sites further along the actin filament and in this manner generates force (75, 76). Consecutive binding and detachment in turn is dependent on conformational changes that induce rotation of myosin’s motor protein structures that lead to the ‘power stroke’. The power stroke signifies myosin changing from one actin binding site to another (75). Both migration modes are depicted in Figure 4.
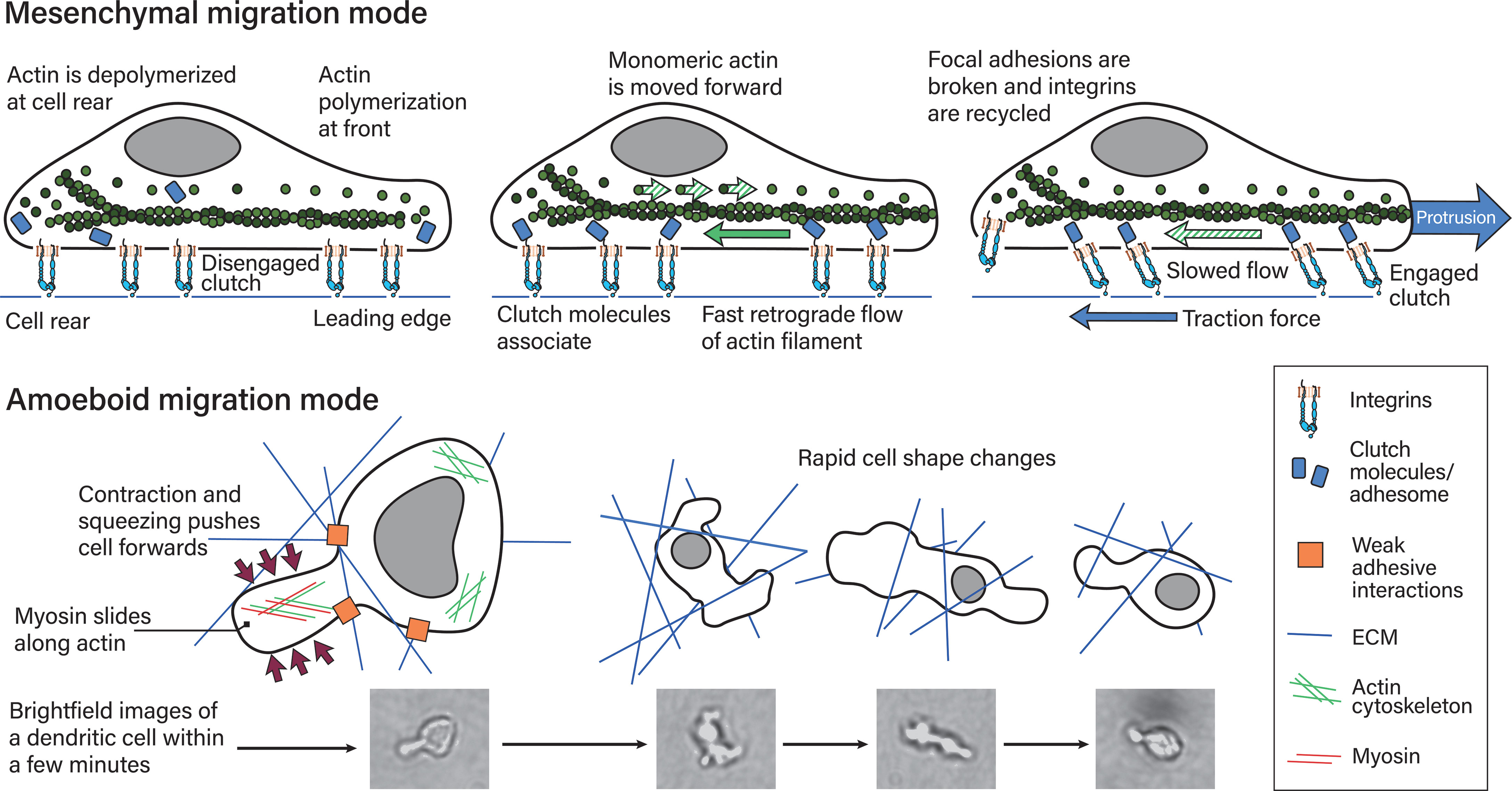
Figure 4 Mesenchymal and amoeboid migration modes. The mesenchymal migration mode, depicted on top, is mediated by strong focal adhesions including integrin–ligand bonds to the substrate. Actin polymerization at the cell’s leading edge and actin retrograde flow powers this migration mode, which requires the counterforce of the adhesion sites. In more detail (77): (left) Actin is polymerized at the leading edge while it is depolymerized at the rear. In this depiction adhesome/clutch proteins have not bound integrin cytoplasmic tails and are instead distributed in the cytoplasm. This means the integrin-actin clutch is disengaged. (middle) Actin polymerization at the front necessitates actin monomers being moved to the front. The growing actin polymer at the front pushes against the elastic fibers at the cell membrane and generates an actin retrograde flow relative to actin’s position in the cell. If the integrin-actin clutch is not engaged this flow is fast. Integrin–ligand bonds are reinforced by adhesome protein binding, which results in focal adhesions, along with the metaphorical clutch, being formed. The clutch is still disengaged if the clutch does not bind actin. (right) The adhesome proteins within the clutch mediate binding to polymerized actin moving towards the rear. This results in slowing of the retrograde actin flow, traction force generation and ultimately protrusion. At the cell rear, focal adhesions are broken and proteins, including integrins recycled. During amoeboid migration (bottom), leukocytes exert weak adhesive forces on the extracellular matrix. This mode is powered by myosin II sliding along actin filaments at the cell’s rear, which translates into contraction and squeezing forces that push the cytoplasm and cell body forwards. Leukocytes find their paths through complex 3D environments and navigating these involves rapid cell shape changes within minutes and shorter, as depicted on the right. The reference pictures were taken during a timelapse 3D dendritic cell migration experiment through a collagen matrix and are of a single dendritic cell, with most cell shapes not being depicted.
The regulation of the two different cell migration modes occurs by a combination of both intracellular mechanisms and sensed extracellular parameters. Intracellularly, cell migration modes are regulated by cell shape and adhesiveness. These processes in turn are regulated by the balance of Ras homologous A (RhoA), Ras-related C3 botulinum toxin substrate (Rac) proteins and cell division cycle 42 (Cdc42), which are small GTPases central to actin remodeling dynamics and are characterized further below.
Extracellular parameters are sensed by integrins. In order to function properly, integrins are typically located at the edges of filopodia and lamellipodia, which are organized membrane structures that T-cells have been shown to use to navigate the extracellular matrix (ECM) (78), a mechanism most likely shared by all leukocytes (depicted in Figure 5). Filopodia are highly dynamic, finger-like protrusions containing parallel bundles of filamentous actin (F-actin) (79). The lamellipodia is a flat membrane protrusion at the leading cell edge, mainly consisting of F-actin, and the persistence of the lamellipodia is important for directionality during cell migration (80). Lamellipodia formation is associated with the mesenchymal migration mode (81). The area directly behind the lamellipodia is termed the lamella and is also relatively flat but contains myosin II (82–84). The lamellipodia has been proposed to be dispensable for cell migration, while the lamella is essential in epithelial cell migration (84). The cell rear of migrating cells is called the uropod. Migrating leukocytes form pseudopodial protrusions (membrane structures that facilitate material uptake) at the leading edge during amoeboid migration (85). Macrophages, dendritic cells and neutrophils form podosomes, which are actin-rich membrane protrusions used to degrade the ECM (86). Additionally, leukocytes form membrane blebs during amoeboid migration modes through which they push into the ECM and generate anchoring stresses, as well as shear stress at bleb protrusions (81). Amoeboid migrating neutrophils in a zebrafish model have been found to form knobs, as the beginning of long trailing extensions that undergo dynamic retractions at the cell rear (87). However, it is not clear what kind of proteins are present in these trailing extensions and what function they serve.
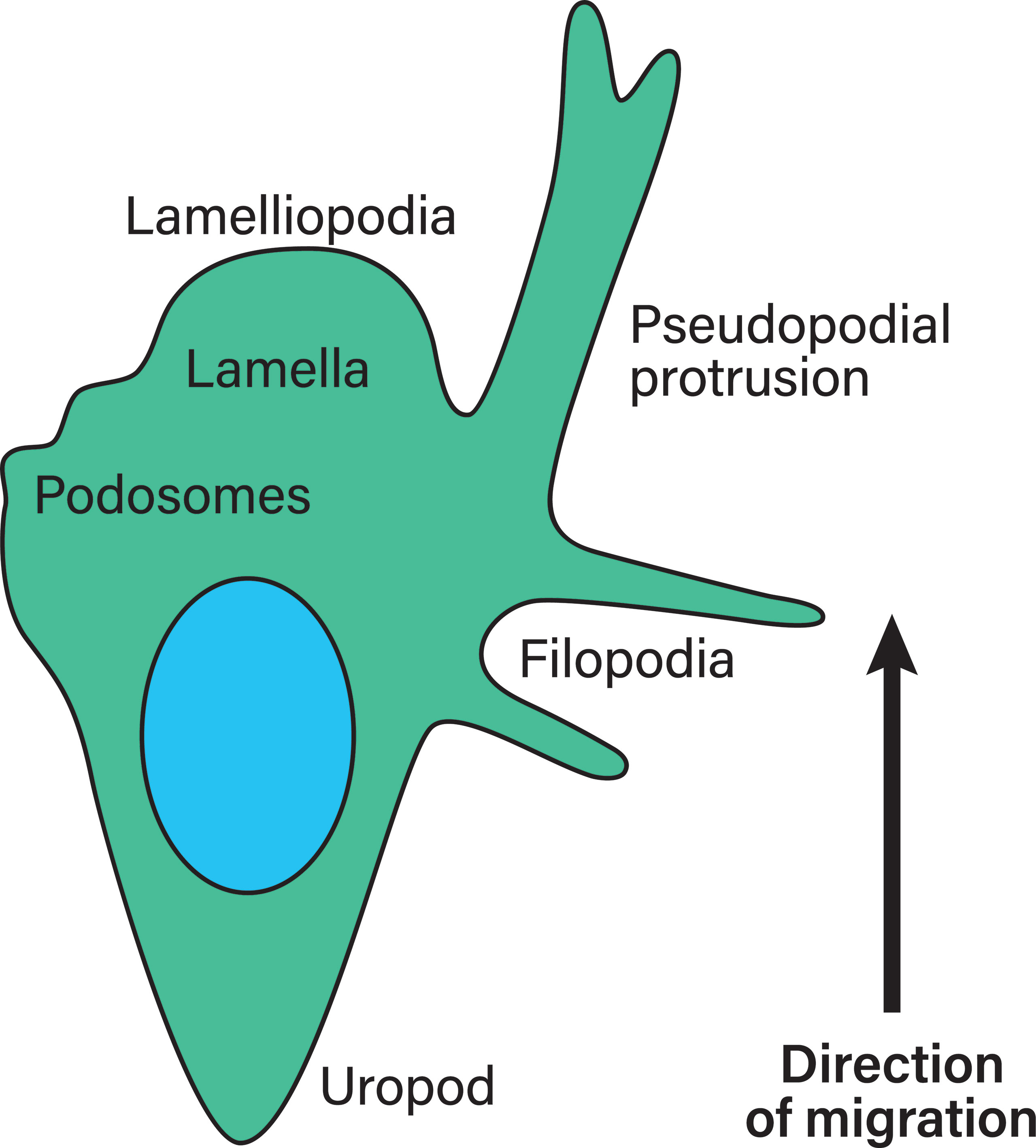
Figure 5 Membrane structures in migrating leukocytes. Actively migrating leukocytes on 2D surfaces form distinct membrane structures. Depicted is the lamellipodia, the actin-rich leading cell edge, the lamella behind it (containing myosin II), as well as the uropod or the cell rear of migrating leukocytes, which are membrane structures that need to be formed during polarization to achieve directional migration. Pseudopodial protrusions are thick, arm-like membrane protrusions in the direction of migration that mediate material uptake. Podosomes are small membrane protrusions which are associated with ECM degradation using proteases. Finally, filopodia are thin finger-like membrane protrusions used to probe the surrounding environment. During amoeboid migration in 3D, leukocytes form small membrane blebs to enact low adhesive forces on the ECM.
Extracellular matrix parameters that regulate migratory modes include ligand availability (for adhesion receptors), density, stiffness, elasticity and pore size. Of these parameters, pore size is a critical factor. Pores that are many times larger than cells render matrices into essentially 2D surfaces and thereby prevent leukocytes from utilizing the 3D migration mode. Pores that are many times smaller than cells block cell migration (71, 88). The nucleus is a major obstacle in leukocyte migration because it is the stiffest and largest organelle (89, 90), and if a cell is unable to move its nucleus through a pore, it has to find a way around (91–93). In order to efficiently navigate ECM environments, leukocytes use their nuclei as mechanical force sensors to scan their environment for adequate pore sizes (94). Leukocytes can further use their nuclei to push ECM components such as collagen apart, and test ECM qualities like stiffness and elasticity, which sets them apart from other migrating cells, that typically drag their nuclei behind (71, 88, 95, 96). Stiffness has been shown to guide cell migration towards stiffer matrices and this process is called durotaxis (71, 88, 95). Elasticity is closely linked to stiffness. Adhesion receptor availability dictates which migration mode can be used. Integrin ligand availability is critical for the mesenchymal migration mode, which is powered by pushing filaments, causing the membrane to protrude (97). This can only happen if there is actin retrograde flow and this in turn is dependent on a counter force such as integrin engagement (77, 97). Dependent on integrin or adhesion receptor ligand availability, dendritic cells switch from adhesion-dependent to independent modes (97). This is probably some of the most direct evidence regarding how integrins regulate migration modes as engagement of the integrin–actin clutch translates to protrusion, whereas disengagement of this clutch leads to fast actin retrograde flow and slippage (amoeboid migration mode) (77, 97). Basically, ECM parameters dictate which migration modes are possible for leukocytes, while RhoA, Rac and Cdc42 mediate migration modes intracellularly.
Integrins function as regulatory and executive bridges between these two components since they bind ECM components and are upstream of RhoA and Rac. The function as executive bridges is based on the characteristic integrin signaling in which they ‘integrate’ extracellular information into the inside of the cell. This type of signaling is called ‘outside-in’ signaling. The ability of integrins to mediate ‘outside-in’ signaling is due to their structure as well as binding of a multitude of cytoplasmic interaction partners. Integrins are transmembrane proteins and are linked to the intracellular cytoskeleton via interaction partners such as talin and kindlin (98, 99). Extracellular information is further transmitted via the actin cytoskeleton directly to the nucleus (100, 101) or is translated into chemical information in a multitude of signaling pathways downstream of integrins. Many such pathways contain focal adhesion kinase (FAK), Src kinase family members, adaptor molecules bearing immunoreceptor tyrosine-based activation motifs (ITAMs, e.g. Fc receptor γ-chain IIa), Syk and Rho GTPases that regulate actin polymerization (98, 102–104). For example, β1-integrins (105, 106) and β2-integrins (17) regulate actin dynamics via RhoA. Whether β2-integrin signaling stimulates or inhibits Rac seems to be cell type–dependent. In neutrophils, β2-integrin–mediated adhesion has been shown to downregulate Rac and stimulate Cdc42 (107). In human T-cells, CD11a/CD18 has been shown to activate Rac-1 and Cdc42, which mediates cell adhesion (108).
Inside-out and outside-in signaling can happen seamlessly during adhesion processes. An example is, again, adhesion in the bloodstream. In order to withstand the shear forces leukocytes are subjected to, adhesion sites are strengthened under force. Single adhesion points consisting of individual or a few integrin-ligand bonds can be reinforced by recruiting other integrins to form multiprotein clusters (focal adhesion points). This reinforcement is a result of outside-in signaling followed by inside-out signaling (109).
As indicated throughout, both ways of integrin signaling are mediated by cytoplasmic interaction partners. Cytoplasmic interaction partners and proteins that are recruited to integrin adhesion sites are collectively termed the integrin adhesome. Hundreds to thousands (110) of integrin adhesome proteins have been identified (111, 112); 60 integrin adhesome proteins are shared between integrin classes such as α-actinin, talin, tensin and filamin (113). Talin, kindlin(-2) and filamin crosslink integrins to the actin cytoskeleton (114, 115) and recruit other proteins to mediate downstream signaling via Src kinases (Lck, Lyn, Fyn) and Syk (116).
In the following sections β2-integrin downstream signaling components will be characterized to outline rough pathways before the components’ involvement in migration of individual leukocyte subpopulations is explored.
Talin is an important integrin cofactor in general, and its binding to integrin cytoplasmic tails is involved in activation of β3-integrins during inside-out signaling (117). Talin is mechanosensitive and unfolds under applied forces of 5–25 pN (118, 119). In neurons, talin signaling initiates RhoA/ROCK activation via Src (120). Talin binds DLC1 (121), which activates RhoA, RhoB, RhoC and Cdc42 and is a negative regulator of RhoA in non-small cell lung carcinoma cells (122).
Kindlin-3 is important for integrin inside-out signaling mediating the active integrin conformation and is essential for proper function of multiple integrin classes (β1, β2 and β3 at least) (123–125). In fact, this signaling is so important that mutations affecting the kindlin-3 gene result in leukocyte adhesion deficiency III (LAD III; mutations mainly affecting CD18 lead to LAD I). LAD III is prevented by the expression of only 5% of the typical kindlin-3 level during homeostasis, however, higher amounts are needed for leukocytes to function properly (126). Kindlin-3 is mainly expressed in the haematopoietic system, so it is important for leukocytes and platelets (123, 127). Kindlin-3 is essential for CD11a/CD18 inside-out signaling (125).
Filamin A is an integrin cytoplasmic cofactor interacting with β-1, β-2, β-3 and β-7 integrins (128–134), mechanosensitive (135) and an actin crosslinking protein (136). Filamin A-actin crosslinking establishes leading-edge actin networks which distribute forces over the entire cytoskeleton (137). This force distribution is especially important during adhesion to and crawling across the endothelium since it allows cells to withstand shear forces (138).
Stretching Filamin A/actin crosslinks under forces of 2–5 pN (135) leads to Filamin A unfolding and dissociates the FilaminA-binding GTPase-activating protein (FilGAP) from filamin. Instead, Filamin A binding to integrins is increased (139). FilGAP inactivates Rac1 in a Rho-associated kinase (ROCK)-dependent manner, and it inactivates Cdc42 to a lesser extent. The forced expression of Filamin A together with FilGAP decreases active Rac1 amounts, and since FilGAP is localized at lamellae, it inhibits lamellae formation and integrin-mediated cell spreading, which could be important for amoeboid migration (140). Filamin A itself is an important regulator of leukocyte adhesion, migration and homing.
Actin is indispensable for both migration modes. In accordance with the importance of the actin network, RhoGTPases – integral to actin remodeling – have been shown to be involved in many migrating leukocytes. RhoGTPases include 20 members and are excellently reviewed in (141). Of these 20 members, RhoA, Rac proteins and Cdc42 will be characterized below and their downstream effectors are depicted in Figure 6.
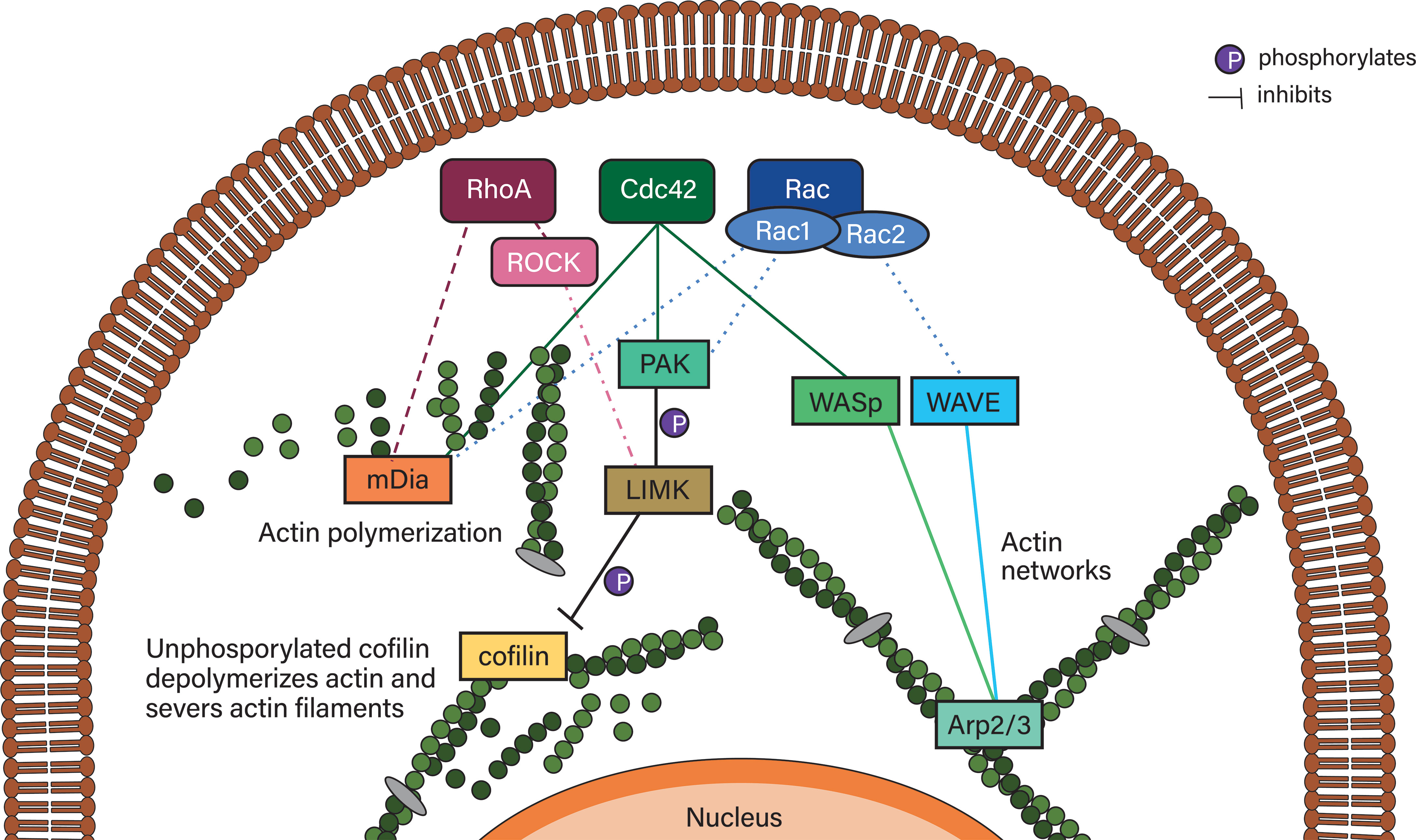
Figure 6 Simplified RhoGTPase signaling to their downstream effectors. RhoA signals to mDia2 and, via ROCK, can inhibit actin depolymerization via LIMK/cofilin. Cdc42 can activate Rac proteins, which activates PAK proteins that phosphorylate LIMK family proteins, which phosphorylate and thereby deactivate cofilin. Deactivated phosphorylated cofilin is reactivated by dephosphorylation mediated by slingshot proteins such as SSHL1 (not shown). Cdc42 also initiates actin polymerization via mDia2 and actin network formation (typically in lamellipodia/lamella) via WASp, which activates Arp2/3. Arp2/3 is also activated by the WAVE complex of which hem1 is an associated protein. WAVE is activated by Rac proteins such as Rac1 and Rac2. Rac proteins also activate mDia2 as well as the PAK/LIMK/cofilin axis.
RhoA has been linked to cell migration in general. It has been shown to be essential for, for example, colon cancer (142), stellate cell (143) and mature osteoclast migration (144). In leukocytes, RhoA is important for actin contractility; this enables retraction of the cell rear during monocyte transmigration (145) and pushes cell migration in several cell types (68, 71, 73, 146), which is characteristic of amoeboid migration and its associated round cell shapes and weak adhesion (18, 71). However, RhoA plays a role in both active migration modes. RhoA induces ROCK, which regulates actin via LIMK and cofilin (141). This has been shown to be essential for leukocyte migration, especially in 2D conditions in which RhoA/ROCK is essential for establishing one leading membrane lamella and thereby directionality (147). RhoA also enhances the activity of mammalian Diaphanous-related formins (mDia)2, which is important for actin polymerization (148, 149).
RhoA also regulates leukocyte arrest on the endothelium, by mediating CD11a/CD18 conformational change into the active conformation as part of an inside-out signaling pathway (4, 150, 151) downstream of RAP1 (in thymocytes) (152). It is important to note that (β2)-integrins are not the only adhesion receptors upstream of RhoA. For example, P2Y1, a platelet purinergic receptor, has been shown to be upstream of RhoA activation, and both were essential for platelet-dependent leukocyte chemotaxis in vitro and leukocyte recruitment in thrombocytopenic mice (153).
Rac, or Rac proteins (Rac1, Rac2, Rac3 and RhoG), is another group of GTPases that can activate RhoA in fibroblasts (154, 155). Rac proteins are critical for generating actin rich, stimulated lamellipodia formation and membrane ruffling, and are important for establishing adhesion sites to the extracellular matrix together with Cdc42 in macrophages (156). While Rac1 is ubiquitously (141) and RhoG widely expressed (157), Rac2 is mostly only expressed in cells of the hematopoietic lineage (158, 159), and Rac3 is mostly found in the brain (141, 160, 161). Rac, along with increased adhesion, is associated with increased spreading and the slower mesenchymal migration mode (68, 71, 73, 146). Rac in leukocytes is critically dependent on DOCK2, a haematopoietic cell-specific CDM family protein downstream of chemokine receptors (162). T-lymphocytes and B-lymphocytes deficient in DOCK2 exhibit homing and migratory defects in vivo, however, these migratory defects where not present in monocytes (162). Rac signals via several different downstream proteins to initiate lamellipodia formation. Actin polymerization is initiated by Rac via the WAVE complex, which activates actin-related protein (Arp) 2/3 that activates actin polymerization in a branched network (163), or Rac initiates mDia2 which mediates actin polymerization directly. Similarly to Cdc42, Rac initiates actin turnover via active P-21–activated kinases (PAK), LIM kinase (LIMK) and Cofilin (141).
Cdc42 in turn can activate Rac1 (154, 155, 164). Not only does Cdc42 regulate the actin cytoskeleton, it also has an important function in regulating cell polarity, which is well conserved in many eukaryotic organisms (141). Cdc42 controls actin polymerization via WASp and IRSp53, which activate ARP2/3 and mDia2 – which initiates actin polymerization directly – and PAK, which inhibits actin turnover via LIMK and Cofilin (141).
RhoGTPases are clearly involved in regulating adhesion and migration of many different leukocytes. However, expression levels differ across leukocyte subpopulations and even within monocytes. A study by van Helden et al. (165) investigated mRNA levels of RhoGTPases in neutrophils, differentiating dendritic cells and macrophages. They found that neutrophils expressed high levels of Cdc42, RhoQ, Rac1, Rac2, RhoA and RhoC and low and specific expression of RhoG, RhoB, RhoF and RhoV. While this GTPase expression profile was similar between different cell types, differences emerged during differentiation. During dendritic cell differentiation, Rac1 was up- and Rac2 downregulated and the dendritic cell populations expressed very low levels of RhoV, which was also present at low levels in macrophages and at higher levels in CD34+ progenitors. Macrophage GTPase expression profiles looked similar to those of dendritic cells, however, M-CSF–differentiated macrophages also expressed low levels of RhoBTB1. Apart from these minuscule differences, neutrophils, dendritic cells and macrophages differed regarding whether RhoA, Rac1, Rac2 or CdC42 was most expressed. The individual percentages were also dependent on which growth factor had been used to differentiate the cells. For example, GM-CSF–differentiated macrophages expressed high Rac2 levels while M-CSF–differentiated macrophages expressed high levels of RhoA.
Furthermore, RhoH is a GTPase only expressed in the haematopoietic linage (166). While myeloid cells and lymphocytes have both been shown to express RhoH, lymphocytes express high levels of this GTPase (167, 168). Thymocyte development and T-cell receptor signaling is critically dependent on RhoH (169, 170). B-cell receptor signaling is intact in RhoH-deficient cells, however B-cell chronic lymphocytic leukemia is dependent on RhoH (171). RhoH has been described as an inhibitor of other RhoGTPase activity, because it negatively regulates RhoA-induced NfKB and p38 signaling in Jurkat cells (172), however, its active contribution to T-cell receptor signaling does not involve Rac inhibition (170). Knockdown of RhoH in T-cells leads to the activation of CD11a/CD18, but does not affect the heterodimer’s ability to bind ligands and mediate adhesion (170). It is important to note that RhoH is regulated on a transcriptional level (172).
These results might explain some of the discrepancies between cell types and even studies. RhoGTPase expression profiles are clearly closely linked to individual cell identity, based on which stimuli the cell had experienced. Additionally, leukocyte migration mechanisms should be more closely investigated based on cell types and even sub-populations.
Integrins are mechanosensors and they transmit mechanical forces directly, via the actin cytoskeleton, to the nucleus (173) and the nuclear lamina, consisting of lamin. Lamin A/C has been shown to be force-sensitive and scales in response to higher extracellular stiffness in order to maintain the cell’s tensional integrity (174, 175). Lamin A/C is responsible for nuclear stiffness (176) and high lamin A/C levels hinder 3D leukocyte migration through porous environments (92). If the β2-integrin/kindlin-3 binding is abolished by mutating the β2-cytoplasmic tail, lamin A/C levels are downregulated. Furthermore, this leads to an increase in H3K4me3 levels, a methylation mark associated with active transcription, changes in open chromatin formation and activation of an IKAROS network, which includes RelA. Compared to WT dendritic cells, dendritic cells that had an abolished β2-integrin/kindlin-3 interaction have been shown to migrate faster in a 3D collagen matrix, express higher levels of maturation surface receptors and induce superior tumor rejection responses in a B16.Ova and B16.F10 melanoma model. Directly targeting H3K4 methylation and disrupting the adhesion in WT dendritic cells was shown to be enough to replicate this phenotype (177). Confinement of Jurkat cells in a 3D collagen I matrix resulted in H3K4me3 upregulation via WD repeat domain 5 (178). Furthermore, in Jurkat cells or primary CD4+ T-cells VCAM1 adhesion mediated by α1β1-integrin leads to triple methylation of another lysine, H3K9me2/3. This involves interaction of methyltransferase G9a with lamin B1 and mediates nuclear stiffness and viscoelasticity and ultimately regulates T-cell migration in 2D and 3D as G9a depletion blocks T-cell migration (179). Finally, histone deacetylation has been shown to be essential for CD4+ T-cell extravasation as histone deacetylase 1 deficiency resulted in reduced transwell transmigration, aberrant migration on ICAM-1 surfaces in vitro and decreased migration into the intestinal epithelium and lamina propria in vivo. Additionally, during the cell adhesion cascade fewer histone deacetylase 1 deficient T-cells were found to probe the surface for places to transmigrate. Abolished histone deacetylation further resulted in downregulation of CD11a, CD18 and β7 chain as well as selectins on a transcriptional level, however, deficient T-cells spread more on ICAM-1 and formed more F-actin, while their random cell migration speed was increased by 10% (180). It is clear, that as with RhoGTPases, the mechanism of how epigenetic marks regulate leukocyte migration is complex and context dependent and much effort is required to unravel this mechanism fully.
Leukocyte migration and specific migratory elements such as chemotaxis, transmigration, membrane architecture and adhesion are regulated differently by varying RhoGTPases and their downstream effectors across leukocyte cell types. In accordance with this, migratory behavior also differs between leukocyte cell types, however, some leukocytes have been much more extensively studied, while leukocytes such as basophils and eosinophils have not been studied much regarding migration and trafficking.
Blood-derived monocytes enter tissues from the bloodstream via the cell adhesion cascade, which involves utilizing the 2D mesenchymal migration mode. Additionally, they are able to use the amoeboid migration mode in fibrous, porous collagen I matrices, however, they are unable to infiltrate dense matrigel matrices or to form podosomes (181). Successful invasion of such matrices is dependent on proteolytic degradation (see section on macrophages). Monocyte migration seems to be intracellularly governed by RhoA and ROCK, while Filamin A merely restricts monocyte transmigration in vitro (182). RhoA and ROCK control β2-integrin localization (145) and leading edge formation during transmigration (147). Furthermore, RhoA/ROCK promote monocyte adhesion, migration and transendothelial transmigration (183). RhoA, as well as Cdc42, regulate filopodia formation (184, 185), and Cdc42 also regulates monocyte migration across endothelial cells (185).
Dendritic cell migration is dependent on their maturity, with immature dendritic cells being relatively immobile, while sampling antigens and intrinsically activated or extrinsically stimulated maturation leads to CCR7 upregulation, which initiates haptotaxis (directed migration along an immobilized chemokine gradient) (186). Dendritic cells use different migration modes when migrating to lymph nodes. To exit constrained porous environments, such as in ear tissues and the skin, dendritic cells use the amoeboid migration mode (18, 187, 188). Like macrophages, dendritic cells can use a protease-dependent, ROCK-independent migration mode in dense 3D environments like matrigel. In such dense 3D environments, podosome formation, rather than maturation status, has been shown to be essential for 3D migration. Dendritic cells migrate through heterogenous environments such as tumor spheroids using both mesenchymal and amoeboid migration modes (188). Once dendritic cells are inside the lymph vessels, they form lamellipodia to crawl along the endothelium (55). CCL-21/CCR7 activation initiates chemotaxis/haptotaxis (189, 190) and arrests dendritic cells on the endothelium. In addition to the chemokine cue, dendritic cells also sense the lymph flow which directs them to proceed downstream (55). Dendritic cells can enter lymph nodes by transmigrating through the subcapsular sinus on the afferent side and then prepare the entry of T-cells into these sites. During intranodal migration, dendritic cells have been shown to exhibit extensive polarization, which established leading edges and a long uropod, which was necessary to translocate to the paracortex. Dendritic cell polarization is dependent on CCR7 (191). Motile dendritic cells are also found in the gut, the lamina propria, Peyer’s patches and solitary intestinal lymphoid tissues from where dendritic cells migrate to gut-draining lymph nodes. During gut infections accompanied by rupture of afferent lymph vessels, dendritic cell migration is prevented (186). Dendritic cell migration from the lung tissue to draining lymph nodes has been shown to be dependent on CCR7, but CCR8 was also implicated (186, 192). Intracellularly, β2-integrin/kindlin-3 signaling restricts the mature, migratory phenotype of dendritic cells, and abolishing the β2-integrin/kindlin-3 interaction leads to increased expression of maturity markers, increased migration speed in 3D towards CCL19, increased IL-12a production and induction of superior anti-tumor responses in B16.OVA and B16.F10 melanoma models in vivo (177, 193). Furthermore, in dendritic cells the β2-integrin/kindlin-3 signaling is upstream of RhoA in a ROCK-independent pathway that regulates adhesion but not 3D migration (17).
Macrophages use mesenchymal migration modes, as well as amoeboid migration modes in 3D environments, and this depends on the specific environment/matrix architecture. For example, the amoeboid migration mode is utilized in porous fibrillar collagen I, and mesenchymal migration mode in dense matrigel (194). During mesenchymal migration modes, macrophages form podosomes that can lyse collagen, however, this migration mode is not dependent on matrix metalloproteinase (MMPs) and might be mediated by other proteolytic systems (194). If collagen I is in the form of a gel, macrophages also utilize mesenchymal migration modes and this could be due to the increased stiffness and viscosity of this form of collagen I (194) or the matrix structure, such as the availability of pores. Macrophages use a migration mode dependent on ROCK and MMPs to infiltrate tumor cell spheroids encapsulated in matrigel (195). These spheroids would present a heterogeneous matrix architecture similar to another collagen I matrix polymerized at 3.6 mg/ml. The 3.6 mg/ml collagen I matrix has been found to contain fibrillar collagen structures, be porous and have intermediate stiffness (G’ is around 30 Pa, though this is quite soft) and viscosity (G’’ is around 8–9 Pa) (195). Macrophages used a combination of amoeboid migration mode and MMP-dependent mesenchymal migration mode in this collagen I matrix, which resembled spheroid infiltration. In a follow-up study, Filamin A was shown to be essential for a macrophage mesenchymal migration mode utilizing proteolytic podosomes. Filamin A is, however, dispensable for amoeboid migration modes (196), and restricts macrophage transmigration in vitro (197). Macrophage migration mode seems to be additionally dependent on macrophage status and β2-integrin expression levels (198). CD11d/CD18 is upregulated on M1 proinflammatory macrophages, while CD11b/CD18 is highly expressed on resident macrophages. High expression levels of these integrins inhibit migration and promote adhesion, while intermediate integrin expression promotes amoeboid and mesenchymal migration modes. Knocking out these heterodimers has also been shown to result in reduced migration (198). Finally, individual levels of RhoGTPases seem to be critical for macrophage migration: RhoA mediates actin cable formation, macrophage rounding up and contractility (156) and has to be precisely concentrated in order to facilitate macrophage migration (199). Rac1 and Rac2 are important GTPases in macrophages (200), with Rac1 being the most abundant Rac isoform (201, 202) and being essential for lamellipodia formation and macrophage migration (199). Rac1 is also important for macrophage invasion through matrigel (203), which would have likely utilized the mesenchymal migration mode (194). Rac2 mediates transmigration; additionally, its role in macrophage migration is somewhat substratum-dependent, since it mediates migration across laminin and fibronectin (203).
Microglia are a type of leukocyte found in the brain. They express CD11b (a β2-integrin alpha chain) (204, 205) and are phenotypically similar to macrophages. While a study with β2-integrin knockout mice has shown that β2-integrins are not necessary for microglia migration (206), similar to dendritic cells, this experiment reduced β2-integrins to just 2–16 % of CD18 expression (206), therefore not precluding their involvement to a small degree. Microglia in vitro were shown to migrate with similar speeds (ca. 60 µm/min) to leukocytes. During migration, microglia were shown to use thin transparent lamellipodia as ‘sails’ to glide forward (207). In murine brain slices, microglia migrated at an average of 5.02 ± 0.06 μm/min, with spurts of 10 μm/min speeds and migrated individually and not highly directionally (208). In another study using murine brain slices, CD11a was knocked down in a microglia cell line and this inhibited microglia migration. Although CD11a-deficient microglia were able to migrate up to 50 µm into the slices, WT microglia were able to migrate down to 80 µm (209).
Neutrophils are capable of using the mesenchymal migration mode, as well as the amoeboid migration mode (18). In matrices such as fibrillar, porous collagen I matrices, neutrophils use the amoeboid migration mode, however, they are unable to invade dense matrigel matrices or to form podosomes (181). It is important to note that neutrophils express adhesion receptors dynamically (210); for example, α4β1-integrin expression is downregulated during neutrophil maturation (14). Neutrophils have been observed to swarm into tissues and form clusters as a response to sterile inflammation or infections with bacteria, fungi and parasites. Neutrophil swarming behavior can be categorized into two patterns, transient swarms and persistent swarms. Transient swarms are formed by neutrophils migrating towards a chemoattractant, forming a cluster for a few minutes and migrating out again to join nearby clusters. Persistent swarms are formed by sustained neutrophil migration into clusters that can last for hours. There seems to be tissue-specific parameters dictating which kind of swarming behavior occurs, although in some tissues swarming behaviors can also co-occur. Transient swarms occur in lungs, lymph nodes and intestine, while persistent swarms occur in skin, liver, spleen and cornea. Further parameters that have been shown to influence which swarming behavior occurs are the extent of initial and secondary tissue damage, presence of pathogens and number of recruited neutrophils (211). CD11a/CD18 and CD11b/CD18 were both shown to be essential for the final stages of accumulation in collagen-free zones (212).
The role of β2-integrin cofactors in neutrophil trafficking is dependent on the dimensionality of the environment: in 2D talin mediates adhesion, spreading, polarization and migration, while in 3D it plays a role in chemotactic velocity (213, 214). Furthermore, talin is important for activating integrins to mediate neutrophil entry into sites of infections, to arrest in venules and to emigrate out of venules (214). Kindlin-3 mediates directionality in neutrophils crawling on 2D surfaces (213). Filamin A seems to be an important regulator of neutrophil migration. Not only does it enable fast migration speed in 2D and 3D (215), it also mediates RhoA activation and localization in uropods. Furthermore, Filamin A regulates myosin II–mediated uropod contractility via RhoA, and Filamin A mediates spreading on β1- and β2-integrin ligands (216, 217).
Different RhoGTPases regulate different aspects of neutrophil migration: Rac1 is important for directed neutrophil migration (218), while Rac2 mediates chemokinesis (random movement), spreading as well as proper F-actin generation, which regulates polarization and directed movement (219, 220). Conversely, Cdc42 restricts neutrophil migration speed, but is important for proper directed migration and steering in vitro (221, 222). Cdc42 regulates directed neutrophil migration by controlling polarity via its effector Wiskott-Aldrich Syndrome Protein (WASp), whose activation and localization in uropods is controlled by Cdc42. WASp in turn controls CD11b localization in uropods, and the integrin heterodimer then recruits and stabilizes tubulin (223).
Eosinophil transmigration has been mainly shown to be mediated by CD11a/CD18 and CD11b/CD18 when the cells were stimulated by IL-1. IL-4–mediated transmigration, however, seems to be dependent on α4β1 (VLA-4). Inhibiting CD11b/CD18 and α4β1 has been shown to impair eosinophil accumulation in the airways in in vivo models (10). Similarly, inhibiting CD18 with antibodies inhibited basophil transendothelial transmigration in transwell assays (224).
Mast cell progenitors traffic from the bone marrow into tissues where they mature, and their homing to different (healthy and inflamed mucosal) tissues is dependent on different integrins (225, 226). Homing to the intestine is dependent on integrin α4β7 (the α4 chain, which can also pair with β1, but especially the β7 chain in this case) (21, 225), while homing to the lungs is dependent on α4β7 as well as α4β1 (20, 225). Mast cell homing to the peritoneum is dependent on CD11b/CD18 and αIIbβ3 (23, 227). Migrating mast cells form pericentral actin clusters that prevent cell flattening and localize secretory granules at the cell center, and this actin organization is mDia(1) dependent. mDia1 knockdown was shown to impair mast cell transmigration in a transwell model, however, mDia1 knockout mice have normal mast cell distribution in tissues (228). Mast cells migrate in response to UV exposure from the skin to draining lymph nodes, and this is important for immune suppression. The extent of such migrating mast cells is dependent on the degree of UV exposure, and mast cells primarily migrate to B-cell areas in lymph nodes guided by CXCL12 secreted by B-cells (receptor CXCR4) (54). Additionally, mast cells accumulate at sites of angiogenesis (i.e. blood vessel formation), and are guided by angiogenic factors, such as VEGF, PDGF-AB and bFGF (229). Interestingly, mast cells can secrete heparin, a polysaccharide, and use it to induce endothelial cell migration in vitro (230).
Similarly to T-cells and B-cells, NK cells exit the bone marrow to enter secondary lymphoid organs in a β2-integrin–dependent mode (231). In lymph nodes, NK cells are activated by dendritic cells during infections, however, in vivo they have been shown to barely form stable dendritic cell interactions, instead forming short-lived contacts with dendritic cells. During this process, NK cells are highly motile in order to establish multiple interactions with dendritic cells and migrate with a less straight trajectory than T-cells (232). Conversely, in superficial lymph node regions, NK cells have been shown to exhibit low motility and long interactions with dendritic cells (233). On stroma, human NK cell subsets have different migratory profiles, with CD56 bright NK cells migrating in short, multidirectional tracks, CD56 dim NK cells migrating in linear and rapid fashion and CD56 neg NK cells (CD56low CD3− CD16+ CD57+ KIR+) migrating to a minimal extent. NK cell migration is dependent on CD56 expression, and more mature NK cells exhibit the most motility (234). Furthermore, NK cell migration on 2D surfaces in vitro can consist of a series of short, constrained motions mixed with highly directed motions or migration in a straight line for the entirety of migration, the latter being observed in few NK cells (235). NK cells are proposed to be capable of transmigration and invasion in response to chemokine and cytokine stimulation and without being activated by dendritic cells. NK cell invasion into cancer spheroids was further proposed to be β2-integrin–dependent and, unlike T-cells, NK cells do not recirculate (236).
T-cells in the afferent lymph system are transported by lymph flow into the medullary sinus system. These T-cells then cross the endothelium and enter lymph nodes by directly transmigrating into medullary cords (191). Intranodal migration of T-cells has been shown to be dependent on CCR7, with the chemokine receptor stimulating migration in T-cell areas but not in subcapsular regions in vivo (237). T-cells exit the lymph nodes and travel to sites of infections, which involves adhesion to the endothelium under shear conditions. As mentioned, this adhesion is dependent on the CD18/Kindlin-3 interaction (238). Arrested T-cells that crawl on endothelial cells and ICAM-1–coated surfaces form lamellipodia, which contain low amounts of CD11a/CD18, and uropods, which contain high amounts of CD11a/CD18. Additionally, T-cells that crawl on ICAM-1, incorporated in lipid bilayers, form a specific zone in the middle of their bodies. This zone contains high affinity CD11a/CD18 and mediates adhesion to and migration on ICAM-1, and its stability is dependent on talin (239). In tissues as well as fibrillar, porous collagen I matrixes, T-cells – like monocytes and neutrophils – can use the amoeboid migration mode (181). However T-cells are unable to form podosomes or to invade dense matrigel matrices (181). Intracellularly, T-cell homing and trafficking is critically dependent on Kindlin-3, which mediates CD11a/CD18 inside-out signaling (125, 238). The involvement of further β2-integrin signaling components for T-cell trafficking is context dependent. T-cell transmigration is dependent on RhoA (240), while the tight binding of Filamin A to integrin tails inhibits transmigration (134). Rac1 signaling mediates T-cell adhesion (241) and can, together with Cdc42, compensate for loss of β2-integrin/14-3-3 signaling to mediate T-cell spreading (108).
B-cells are capable of using the amoeboid migration mode (242, 243), however, their 2D migration seems somewhat atypical. During migration on CXCL13-coated surfaces, the leading membrane edge has been shown to undergo dilation and shrinking events, and the balance of these events was predictive of turning versus directional persistence. During this migration, an imbalance of dilation versus shrinking events was associated with changes in direction (244). This phenomenon might involve β2-integrins, since CXCL13/CXCR5 mediates CD11a/CD18 adhesion in B-cells (243). In large B-cell lymphoma, the amoeboid migration mode is mediated by an IL-10-JAK-STAT-3-RhoH pathway that was shown to increase RhoA activity and suppress tubulin acetylation (245).
Leukocyte migration also has a huge implication in cancer therapy, as therapeutic antibodies targeting immune checkpoints have revolutionized cancer treatment in recent years. However, for 70-80% of patients their tumors do not respond to this treatment (246). This poor response has been linked to the presence or absence of T-cells in the tumor environments (247). T-cell tumor invasion is hindered by the prolonged interaction of T-cells with tumor-associated macrophages (TAMs), and depletion of TAMs can restore T-cell migration and invasion (248). In order to mount an anti-tumor immune response, antigen-presenting cells have to present antigens to and fully activate T-cells first. However, this mechanism is warped in order to support tumor growth by dendritic cell–like cells in the tumor periphery that engage CD8+ T-cells in long interactions without fully activating the T-cells (249, 250). This results in ‘immune excluded’ tumors with leukocytes stuck in the periphery. Macrophages and dendritic cells have, in turn, been found to be regulated by myeloid-derived suppressor cells (MDSCs) in glioma (251). MDSCs arise as monocytes from the bone marrow and then effectuate homing to secondary lymphatic organs before being recruited into the tumor, where they are retained. This process is thought to be a result of crosstalk between the bone marrow and the tumor, involving growth factors and chemokines produced in the tumor microenvironment. MDSC homing culminates in their retention in the tumor microenvironment, however, less is known about this process then about T-cell retention (252). Our understanding of leukocyte migration is limited in general and is even more elusive in regards to how it is impaired on a molecular level specifically in tumors.
It is now established in the literature that leukocytes use a distinct migration mode – the amoeboid migration mode – which sets them apart from other migrating cells. However, there are distinct differences between migration profiles of leukocyte populations, and especially the individual molecular mechanisms that mediate these, which were broadly outlined in this review. Future research will hopefully flesh out these differences and should address exactly how the molecular balance between RhoGTPases is regulated. The focus of this review was on β2-integrins – some of the most studied adhesion receptors. However, there are many other adhesion and surface receptors that signal via RhoGTPases and have been shown to play a role in leukocyte migration, such as NOTCH (253–255) and cadherins (256–258). Incorporating other ECM ligands for these receptors could reveal more RhoGTPase profiles and even new migration modes or unique leukocyte migration behavior. Furthermore, individual surface receptors might be key regulators or could strengthen and perpetuate signaling. The ultimate goal would be to establish exactly how collective surface receptor signaling mediates distinct RhoGTPase profiles and how this translates to exact migratory behavior. This should also include unraveling how leukocyte migration is regulated by histone methylation and acetylation downstream of integrin and adhesion receptor signaling. Thus far, deacetylation, H3K4me3, as well as H3K9me2/3, have been implicated in dendritic cell and T-cell migration, respectively. It would be important to see if other methylation marks are associated with the migration of distinct leukocyte subpopulations. Moreover, histone methylation and acetylation have received the most attention thus far but are only a small portion of the many types of histone modifications: it is possible that the others also play a role in regulating leukocyte migration.
Progress in cancer therapy has focused on combination treatments. Combining immune checkpoint inhibitors as well as combination therapy using radiotherapy first and then immune checkpoint inhibitors can improve objective response rates as well as progression free survival (259, 260). Ultimately similar improvements could be possible by mapping out the intracellular signaling that governs leukocyte migration. This could empower us to reestablish migratory behavior in effector T-cells stuck in the periphery of immune-excluded tumors, which could further increase patient survival rates after immune checkpoint inhibitor combination treatment, as well as control leukocyte migration in other diseases.
The author confirms being the sole contributor of this work and has approved it for publication.
IFReC Kishimoto Foundation for supporting the study with a fellowship.
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
My biggest gratitude for improving the manuscript goes to James Alexander Geraets and Mercedes Said. I would also like to very much thank my editor Alyce M Whipp. I thank the University of Helsinki Library for covering the article publishing costs and I thank Susanna Fagerholm for helpful discussion.
1. McEver RP, Zu C. Rolling Cell Adhesion. Annu Rev Cell Dev Biol (2010) 26:363–96. doi: 10.1146/annurev.cellbio.042308.113238
2. Muller WA. Getting Leukocytes to the Site of Inflammation. Vet Pathol (2013) 50(1):7–22. doi: 10.1177/0300985812469883
3. Schenkel AR, Mamdouh Z, Muller WA. Locomotion of Monocytes on Endothelium is a Critical Step During Extravasation. Nat Immunol (2004) 5:393–400. doi: 10.1038/ni1051
4. Klaus Ley CL, Cybulsky MI, Nourshargh S. Getting to the Site of Inflammation: The Leukocyte Adhesion Cascade Updated. Nat Rev Immunol (2007) 7(9):678–89. doi: 10.1038/nri2156
5. Baker EL, Zaman MH. The Biomechanical Integrin. J Biomech (2010) 43(1):38–44. doi: 10.1016/j.jbiomech.2009.09.007
6. Evans R, Patzak I, Svensson L, De Filippo K, Jones K, McDowall A, et al. Integrins in Immunity. J Cell Sci (2009) 122:215–25. doi: 10.1242/jcs.019117
7. Harris ES, McIntyre TM, Prescott SM, Zimmerman GA. The Leukocyte Integrins. J Cell Biol (2000) 275(31):23409–12. doi: 10.1074/jbc.R000004200
8. Luo B-H, Carman CV, Springer TA. Structural Basis of Integrin Regulation and Signaling. Annu Rev Immunol (2007) 25:619–47. doi: 10.1146/annurev.immunol.25.022106.141618
9. Lima CSP, Franco-Penteado C, Canalli AA, Conran N, Lorand-Metze I, Costa F, et al. Expressions of the VLA-4, LFA-1 and Mac-1 Integrins in Eosinophil Migration in a Case of Chronic Eosinophilic Leukaemia. Leukemia Res (2007) 31(5):695–7. doi: 10.1016/j.leukres.2006.07.025
10. Wardlaw AJ, Walsh GM, Symon FA. Mechanisms of Eosinophil and Basophil Migration. Allergy (1994) 49:797–807. doi: 10.1111/j.1398-9995.1994.tb00778.x
11. Moser M, Bauer M, Schmid S, Ruppert R, Schmidt S, Sixt M, et al. Kindlin-3 is Required for Beta2 Integrin-Mediated Leukocyte Adhesion to Endothelial Cells. Nat Med (2009) 15(3):300–5. doi: 10.1038/nm.1921
12. Füreder W, Agis H, Sperr WR, Lechner K, Valent P. The Surface Membrane Antigen Phenotype of Human Blood Basophils. Allergy (1994) 49(10):861–5. doi: 10.1111/j.1398-9995.1994.tb00788.x
13. Lindbom L, Werr J. Integrin-Dependent Neutrophil Migration in Extravascular Tissue. Immunology (2002) 14:115–21. doi: 10.1006/smim.2001.0348
14. Petty JM, Lenox CC, Weiss DJ, Poynter ME, Suratt BT. Crosstalk Between CXCR4/Stromal Derived Factor-1 and VLA-4/VCAM-1 Pathways Regulates Neutrophil Retention in the Bone Marrow. J Immunol (2009) 182:604–12. doi: 10.4049/jimmunol.182.1.604
15. Plow EF, Haas TA, Zhang L, Loftusi J, Smith JW. Ligand Binding to Integrins. J Biol Chem (2000) 275(29):21785–8. doi: 10.1074/jbc.R000003200
16. Van der Vieren M, Crowe DT, Hoekstra D, Vazeux R, Hoffman PA, Grayson MH, et al. The Leukocyte Integrin αdβ2 Binds VCAM-1: Evidence for a Binding Interface Between I Domain and VCAM-1. J Immunol (1999) 163(4):1984–90.
17. Guenther C, Faisal I, Uotila LM, Asens ML, Harjunpää H, Savinko T, et al. A β2-Integrin/MRTF-A/SRF Pathway Regulates Dendritic Cell Gene Expression, Adhesion, and Traction Force Generation. Front Immunol (2019) 10. doi: 10.3389/fimmu.2019.01138
18. Lämmermann T, Bader BL, Monkley SJ, Worbs T, Wedlich-Söldner R, Hirsch K, et al. Rapid Leukocyte Migration by Integrin-Independent Flowing and Squeezing. Nature (2008) 453(7191):51–5. doi: 10.1038/nature06887
19. Arnaout MA. Biology and Structure of Leukocyte β Integrins and Their Role in Inflammation. F1000 Res (2016) 5(2433):13. doi: 10.12688/f1000research.9415.1
20. Abonia JP, Hallgren J, Jones T, Shi T, Xu Y, Koni P, et al. Alpha-4 Integrins and VCAM-1, But Not MAdCAM-1, are Essential for Recruitment of Mast Cell Progenitors to the Inflamed Lung. Blood (2006) 108:1588–94. doi: 10.1182/blood-2005-12-012781
21. Abonia JP, Austen KF, Rollins BJ, Joshi SK, Flavell RA, Kuziel WA, et al. Constitutive Homing of Mast Cell Progenitors to the Intestine Depends on Autologous Expression of the Chemokine Receptor CXCR2. Blood (2005) 105:4308–13. doi: 10.1182/blood-2004-09-3578
22. Knight PA, Wright SH, Brown JK, Huang X, Sheppard D, Mille HRP. Enteric Expression of the Integrin alphaV Beta6 Is Essential for Nematode-Induced Mucosal Mast Cell Hyperplasia and Expression of the Granule Chymase, Mouse Mast Cell Protease-1. Am J Pathol (2002) 161(3):771–9. doi: 10.1016/S0002-9440(10)64236-8
23. Berlanga O, Emambokus N, Frampton J. GPIIb (CD41) Integrin is Expressed on Mast Cells and Influences Their Adhesion Properties. Exp Hematol (2005) 33:403–12. doi: 10.1016/j.exphem.2005.01.011
24. Weinacker A, Ferrando R, Elliott M, Hogg J, Balmes J, Sheppard D. Distribution of Integrins Alpha V Beta 6 and Alpha 9 Beta 1 and Their Known Ligands, Fibronectin and Tenascin, in Human Airways. Am J Respir Cell Mol Biol (1995) 12(5):547–57. doi: 10.1165/ajrcmb.12.5.7537970
25. Li Q, Barres BA. Microglia and Macrophages in Brain Homeostasis and Disease. Nat Rev Immunol (2018) 18:225–41. doi: 10.1038/nri.2017.125
26. Galatro TF, Holtman IR, Lerario AM, Vainchtein ID, Brouwer N, Sola PR, et al. Transcriptomic Analysis of Purified Human Cortical Microglia Reveals Age-Associated Changes. Nat Neurosci (2017) 20:1162–71. doi: 10.1038/nn.4597
27. Goswami S. Importance of Integrin Receptors in the Field of Pharmaceutical & Medical Science. Adv Biol Chem (2013) 3:224–52. doi: 10.4236/abc.2013.32028
28. Philippeaux M, Vesin C, Tacchini-Cottier F, Piguet PF. Activated Human Platelets Express Beta2 Integrin. Eur J Haematol (1996) 56(3):130–7. doi: 10.1111/j.1600-0609.1996.tb01331.x
29. Fagerholm SC, Guenther C, Asens ML, Savinko T, Uotila LM. Beta2-Integrins and Interacting Proteins in Leukocyte Trafficking, Immune Suppression, and Immunodeficiency Disease. Front Immunol (2019) 10(254). doi: 10.3389/fimmu.2019.00254
30. Mitroulis I, Alexaki VI, Kourtzelis I, Ziogas A, Hajishengallis G, Chavakis T. Leukocyte Integrins: Role in Leukocyte Recruitment and as Therapeutic Targets in Inflammatory Disease. Pharmacol Ther (2015), 123–35. doi: 10.1016/j.pharmthera.2014.11.008
31. Schittenhelm L, Hilkens CM, Morrison VL. β2 Integrins As Regulators of Dendritic Cell, Monocyte, and Macrophage Function. Front Immunol (2017) 8(1866). doi: 10.3389/fimmu.2017.01866
32. Carman CV, Springer TA. Integrin Avidity Regulation: Are Changes in Affinity and Conformation Underemphasized? Curr Opin Cell Biol (2003) 15:547–56. doi: 10.1016/j.ceb.2003.08.003
33. Askari JA, Buckley PA, Mould AP, Humphries MJ. Linking Integrin Conformation to Function. J Cell Sci (2009) 122(2):165–70. doi: 10.1242/jcs.018556
34. Li R, Rieu P, Griffith DL, Scott D, Arnaout MA. Two Functional States of the CD11b A-Domain: Correlations With Key Features of Two Mn2+-Complexed Crystal Structures. J Cell Biol (1998) 218(9):1523–34. doi: 10.1083/jcb.143.6.1523
35. McCleverty CJ, Liddington RC. Engineered Allosteric Mutants of the Integrin Alphambeta2 I Domain: Structural and Functional Studies. Biochem J (2003) 372(1):121–7. doi: 10.1042/bj20021273
36. Zarbock A, Abram CL, Hundt M, Altman A, Lowell CA, Ley K. PSGL-1 Engagement by E-Selectin Signals Through Src Kinase Fgr and ITAM Adapters DAP12 and Fcrγ to Induce Slow Leukocyte Rolling. J Exp Med (2008) 205(10):2339–47. doi: 10.1084/jem.20072660
37. Tadayuki Yago NZ, Zhao L, Abrams CS, McEver RP. Selectins and Chemokines Use Shared and Distinct Signals to Activate β2 Integrins in Neutrophils. Blood Adv (2018) 2(7):731–44. doi: 10.1182/bloodadvances.2017015602
38. Takagi J, Petre BM, Walz T, Springer TA. Global Conformational Rearrangements in Integrin Extracellular Domains in Outside-In and Inside-Out Signaling. Cell (2002) 110(5):599–611. doi: 10.1016/S0092-8674(02)00935-2
39. Wegener KL, Partridge AW, Han J, Pickford AR, Liddington RC, Ginsberg MH, et al. Structural Basis of Integrin Activation by Talin. Cell (2007) 128(1):171–82. doi: 10.1016/j.cell.2006.10.048
40. Choi EY, Santoso S, Chavakis T. Mechanisms of Neutrophil Transendothelial Migration. Front Biosci (Landmark edition) (2009) 14:1596–605. doi: 10.2741/3327
41. Nourshargh S, Hordijk PL, Sixt M. Breaching Multiple Barriers: Leukocyte Motility Through Venular Walls and the Interstitium. Nat Rev Moclec Cell Biol (2010) 11:366–78. doi: 10.1038/nrm2889
42. Partsch G, Schwarzer C. An Indirect Bioluminescence Method for the Quantitative Measurement of Polymorphonuclear Cell Chemotaxis. J Biolumin Chemilumin (1991) 6(3):159–67. doi: 10.1002/bio.1170060305
43. Kramer N, Walzl A, Unger C, Rosner M, Krupitza G, Hengstschläger M, et al. In Vitro Cell Migration and Invasion Assays. Mutat Res/Rev Mutat Res (2013) 752(1):10–24. doi: 10.1016/j.mrrev.2012.08.001
44. Place TL, Domann FE, Case AJ. Limitations of Oxygen Delivery to Cells in Culture: An Underappreciated Problem in Basic and Translational Research. Free Radical Biol Med (2017) 113:311–22. doi: 10.1016/j.freeradbiomed.2017.10.003
45. Trombetta ES, Mellman I. Cell Biology of Antigen Processing In Vitro and In Vivo. Annu Rev Immunol (2005) 23:975–1028. doi: 10.1146/annurev.immunol.22.012703.104538
46. Culshaw S, Millington OR, Brewer JM, McInnes IB. Murine Neutrophils Present Class II Restricted Antigen. Immunol Lett (2008) 118:49–54. doi: 10.1016/j.imlet.2008.02.008
47. Joseph A, Guevara-Torres A, Schallek J. Imaging Single-Cell Blood Flow in the Smallest to Largest Vessels in the Living Retina. eLIFE (2019) 8:1–36:e45077. doi: 10.7554/eLife.45077
48. Blatter C, Meijer EFJ, Nam AS, Jones D, Bouma BE, Padera TP, et al. In Vivo Label-Free Measurement of Lymph Flow Velocity and Volumetric Flow Rates Using Doppler Optical Coherence Tomography. Sci Rep (2016) 6:10. doi: 10.1038/srep29035
49. Hall JG, Morris B. The Output of Cells IN Lymph From THE Popliteal Node Of Sheep. Exp Physiol Cognate Med Sci (1962) 47(4):360–9. doi: 10.1113/expphysiol.1962.sp001620
50. Smith JB, McIntosh GH, Morris B. The Traffic of Cells Through Tissues: A Study of Peripheral Lymph in Sheep. J Anat (1970) 107(1):87–100. doi: 10.1002/path.1711000104
51. Sokolowski J, Jakobsen E, Johannessen JV. Cells in Peripheral Leg Lymph of Normal Men, Lymphology. Lymphology (1978) 11(4):202–7.
52. Olszewski WL, Grzelak I, Ziolkowska A, Engeset A. Immune Cell Traffic From Blood Through the Normal Human Skin to Lymphatics. Clinics Dermatol (1995) 13(5):473–83. doi: 10.1016/0738-081X(95)00087-V
53. Schineis P, Runge P, Halin C. Cellular Traffic Through Afferent Lymphatic Vessels. Vasc Pharmacol (2019) 112:31–41. doi: 10.1016/j.vph.2018.08.001
54. Byrne SN, Limo´n-Flores AY, Ullrich SE. Mast Cell Migration From the Skin to the Draining Lymph Nodes Upon Ultraviolet Irradiation Represents a Key Step in the Induction of Immune Suppression. J Immunol (2008) 180:4648–55. doi: 10.4049/jimmunol.180.7.4648
55. Tal O, Lim HY, Gurevich I, Milo I, Shipony Z, Ng LG, et al. DC Mobilization From the Skin Requires Docking to Immobilized CCL21 on Lymphatic Endothelium and Intralymphatic Crawling. J Exp Med (2011) 208(10):2141–53. doi: 10.1084/jem.20102392
56. Friedl P, Bröcker E-B. Reconstructing Leukocyte Migration in 3D Extracellular Matrix by Time-Lapse Videomicroscopy and Computer-Assisted Tracking. Cell Migration Inflammation Immunity Methods Mol Biol (2004) 239:77–90. doi: 10.1385/1-59259-435-2:77
57. Mempel TR, Scimone ML, Mora JR, von Andrian UH. In Vivo Imaging of Leukocyte Trafficking in Blood Vessels and Tissues. Curr Opin Immunol (2004) 16(4):406–17. doi: 10.1016/j.coi.2004.05.018
58. Torcellan T, Stolp J, Chtanova T. In Vivo Imaging Sheds Light on Immune Cell Migration and Function in Cancer. Front Immunol (2017) 8:309. doi: 10.3389/fimmu.2017.00309
59. Rumianek AN, Greaves DR. How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas About Macrophage Migration? Biology (2020) 9(439):1–13. doi: 10.3390/biology9120439
60. Friedl P, Zänker KS, Bröcker E-B. Cell Migration Strategies in 3-D Extracellular Matrix: Differences in Morphology, Cell Matrix Interactions, and Integrin Function. Microscopy Res Technique (1998) 43(5):369–78. doi: 10.1002/(SICI)1097-0029(19981201)43:5<369::AID-JEMT3>3.0.CO;2-6
61. Repesh LA. A New In Vitro Assay for Quantitating Tumor Cell Invasion. Invasion Metastasis (1989) 9(3):192–208.
62. Giannattasio A, Weil S, Kloess S, Ansari N, Stelzer EHK, Cerwenka A, et al. Cytotoxicity and Infiltration of Human NK Cells in In Vivo-Like Tumor Spheroids. BMC Cancer (2015) 15(351):13. doi: 10.1186/s12885-015-1321-y
63. Weber M, Sixt M. Live Cell Imaging of Chemotactic Dendritic Cell Migration in Explanted Mouse Ear Preparations. Chemokines Methods Mol Biol (Methods Protocols) (2013) 1013:1013. doi: 10.1007/978-1-62703-426-5_14
64. Phillipson M, Heit B, Colarusso P, Liu L, Ballantyne CM, Kubes P. Intraluminal Crawling of Neutrophils to Emigration Sites: A Molecularly Distinct Process From Adhesion in the Recruitment Cascade. J Exp Med (2006) 203(12):2569–75. doi: 10.1084/jem.20060925
65. Blanchoin L, Boujemaa-Paterski R, Sykes C, Plastino J. Actin Dynamics, Architecture, and Mechanics in Cell Motility. Physiol Rev (2014) 94(1):235–63. doi: 10.1152/physrev.00018.2013
66. Gardel ML, Schneider IC, Aratyn-Schaus Y, Waterman CM. Mechanical Integration of Actin and Adhesion Dynamics in Cell Migration. Annu Rev Cell Dev Biol (2010) 26:315–33. doi: 10.1146/annurev.cellbio.011209.122036
67. Huse M. Mechanical Forces in the Immune System. Nat Rev Immunol (2017) 17(11):679–90. doi: 10.1038/nri.2017.74
68. Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, et al. Cell Migration: Integrating Signals From Front to Back. Science (2003) 302(5651):1704–9. doi: 10.1126/science.1092053
69. Pollard TD, Borisy GG. Cellular Motility Driven by Assembly and Disassembly of Actin Filaments. Cell (2003) 112(4):453–65. doi: 10.1016/S0092-8674(03)00120-X
70. Smith LA, Aranda-Espinoza H, Haun JB, Dembo M, Hammer DA. Neutrophil Traction Stresses are Concentrated in the Uropod During Migration. Biophys J (2007) 92(7):L58–60. doi: 10.1529/biophysj.106.102822
71. Friedl P, Wolf K. Plasticity of Cell Migration: A Multiscale Tuning Model. J Cell Biol (2009) 188(1):11–9. doi: 10.1083/jcb.200909003
72. Fackler OT, Grosse R. Cell Motility Through Plasma Membrane Blebbing. J Cell Biol (2008) 181(6):879–84. doi: 10.1083/jcb.200802081
73. Sanz-Moreno, Marshall CJ. Rho-GTPase Signaling Drives Melanoma Cell Plasticity. Cell Cycle (2009) 8(10):1484–7. doi: 10.4161/cc.8.10.8490
74. Janson LW, Taylor DL. In Vitro Models of Tail Contraction and Cytoplasmic Streaming in Amoeboid Cells. J Cell Biol (1993) 123(2):345–56. doi: 10.1083/jcb.123.2.345
75. Houdusse A, Sweeney HL. How Myosin Generates Force on Actin Filaments. Trends Biochem Sci (2016) 41(12):989–97. doi: 10.1016/j.tibs.2016.09.006
76. Kato Y, Miyakawa T, Tanokura M. Overview of the Mechanism of Cytoskeletal Motors Based on Structure. Biophys Rev (2018) 10:571–81. doi: 10.1007/s12551-017-0368-1
77. Case LB, Waterman CM. Integration of Actin Dynamics and Cell Adhesion by a Three-Dimensional, Mechanosensitive Molecular Clutch. Nat Cell Biol (2015) 17:955–63. doi: 10.1038/ncb3191
78. Song KH, Kwon KW, Choi J-C, Jung J, Park Y, Suhb K-Y, et al. T Cells Sense Biophysical Cues Using Lamellipodia and Filopodia to Optimize Intraluminal Path Finding. Integr Biol (2014) 6:450–9. doi: 10.1039/c4ib00021h
79. Gupton SL, Gertler FB. Filopodia: The Fingers That Do the Walking. Sci Signaling (2007) 2007(400):re5. doi: 10.1126/stke.4002007re5
80. Krause M, Gautreau A. Steering Cell Migration: Lamellipodium Dynamics and the Regulation of Directional Persistence. Nat Rev Mol Cell Biol (2014) 15:577–90. doi: 10.1038/nrm3861
81. Yip AK, Chiam K-H, Matsudaira P. Traction Stress Analysis and Modeling Reveal That Amoeboid Migration in Confined Spaces is Accompanied by Expansive Forces and Requires the Structural Integrity of the Membrane–Cortex Interactions. Integr Biol (2015) 7(1196):16. doi: 10.1039/C4IB00245H
82. Vallotton P, Small JV. Shifting Views on the Leading Role of the Lamellipodium in Cell Migration: Speckle Tracking Revisited. J Cell Sci (2009) 122(12):1955–8. doi: 10.1242/jcs.042036
83. Heath JP, Holifield BF. Cell Locomotion: New Research Tests Old Ideas on Membrane and Cytoskeletal Flow. Cell Motil Cytoskeleton (1991) 18:245–57. doi: 10.1002/cm.970180402
84. Gupton SL, Anderson KL, Kole TP, Fischer RS, Ponti A, Hitchcock-DeGregori SE, et al. Cell Migration Without a Lamellipodium: Translation of Actin Dynamics Into Cell Movement Mediated by Tropomyosin. J Cell Biol (2005) 168:619–31. doi: 10.1083/jcb.200406063
85. Hiromi M, Tsubota K, Hoyano T, Adachi T, Liu H. Three-Dimensional Modulation of Cortical Plasticity During Pseudopodial Protrusion of Mouse Leukocytes. Biochem Biophys Res Commun (2013) 438:594–9. doi: 10.1016/j.bbrc.2013.08.010
86. Murphy DA, Courtneidge SA. The ‘Ins’ and ‘Outs’ of Podosomes and Invadopodia: Characteristics, Formation and Function. Nat Rev Moclec Cell Biol (2011) 12:413–26. doi: 10.1038/nrm3141
87. Manley HR, Potter DL, Heddleston JM, Chew T-L, Keightley MC, Lieschke GJ. Frontline Science: Dynamic Cellular and Subcellular Features of Migrating Leukocytes Revealed by In Vivo Lattice Lightsheet Microscopy. J Leukocyte Biol (2020) 108:455–68. doi: 10.1002/JLB.3HI0120-589R
88. Doyle AD, Petrie RJ, Kutys ML, Yamada KM. Dimensions in Cell Migration. Curr Opin Cell Biol (2013) 25:642–9. doi: 10.1016/j.ceb.2013.06.004
89. Caille N, Thoumine O, Tardy Y, Meister J-J. Contribution of the Nucleus to the Mechanical Properties of Endothelial Cells. J Biomech (2002) 35(2):177–87. doi: 10.1016/S0021-9290(01)00201-9
90. Lammerding J. Mechanics of the Nucleus. Compr Physiol (2011) 1(2):783–807. doi: 10.1002/cphy.c100038
91. Kameritsch P, Renkawitz J. Principles of Leukocyte Migration Strategies. Trends Cell Biol (2020) 30(10):818–32. doi: 10.1016/j.tcb.2020.06.007
92. Harada T, Swift J, Irianto J, Shin J-W, Spinler KR, Athirasala A, et al. Nuclear Lamin Stiffness is a Barrier to 3D Migration, But Softness can Limit Survival. J Cell Biol (2014) 204(5):669–82. doi: 10.1083/jcb.201308029
93. Wolf K, te Lindert M, Krause M, Alexander S, te Riet J, Willis AL, et al. Physical Limits of Cell Migration: Control by ECM Space and Nuclear Deformation and Tuning by Proteolysis and Traction Force. J Cell Biol (2013) 201(7):1069–84. doi: 10.1083/jcb.201210152
94. Renkawitz J, Kopf A, Stopp J, de Vries I, Driscoll MK, Merrin J, et al. Nuclear Positioning Facilitates Amoeboid Migration Along the Path of Least Resistance. Nature (2019) 568:546–50. doi: 10.1038/s41586-019-1087-5
95. Lo C-M, Wang H-B, Dembo M, Wang Y-l. Cell Movement Is Guided by the Rigidity of the Substrate. Biophys J (2000) 79:144–52. doi: 10.1016/S0006-3495(00)76279-5
96. Calero-Cuenca FJ, Janota CS, Gomes ER. Dealing With the Nucleus During Cell Migration. Curr Opin Cell Biol (2018) 50:35–41. doi: 10.1016/j.ceb.2018.01.014
97. Renkawitz J, Schumann K, Weber M, Lämmermann T, Pflicke H, Piel M, et al. Adaptive Force Transmission in Amoeboid Cell Migration. Nat Cell Biol (2009) 11(12):1438–43. doi: 10.1038/ncb1992
98. Harburger DS, Calderwood DA. Integrin Signalling at a Glance. J Cell Sci (2009) 122:159–63. doi: 10.1242/jcs.018093
99. Legate KR, Wickström SA, Fässler R. Genetic and Cell Biological Analysis of Integrin Outside-in Signaling. Genes Dev (2009) 23(4):397–418. doi: 10.1101/gad.1758709
100. Maniotis AJ, Chen CS, Ingber DE. Demonstration of Mechanical Connections Between Integrins, Cytoskeletal Filaments, and Nucleoplasm That Stabilize Nuclear Structure. PNAS (1997) 94:849–54. doi: 10.1073/pnas.94.3.849
101. Madrazo E, Conde AC, Redondo-Muñoz J. Inside the Cell: Integrins as New Governors of Nuclear Alterations? Cancers (2017) 9(82):1–17. doi: 10.3390/cancers9070082
102. Hogg N, Patzak I, Willenbrock F. The Insider’s Guide to Leukocyte Integrin Signalling and Function. Nat Rev Immunol (2011) 11:416–26. doi: 10.1038/nri2986
103. Huveneers S, Danen EHJ. Adhesion Signaling – Crosstalk Between Integrins, Src and Rho. Oncogene (2009) 122:1059–69. doi: 10.1242/jcs.039446
104. Zhi H, Rauova L, Hayes V, Gao C, Boylan B, Newman DK, et al. Cooperative Integrin/ITAM Signaling in Platelets Enhances Thrombus Formation In Vitro and In Vivo. Blood (2013) 121(10):1858–67. doi: 10.1182/blood-2012-07-443325
105. Kalappurakkal JM, Anilkumar AA, Patra C, van Zanten TS, Sheetz MP, Mayor S. Integrin Mechano-Chemical Signaling Generates Plasma Membrane Nanodomains That Promote Cell Spreading. Cell (2019) 177(7):1738–56. doi: 10.1016/j.cell.2019.04.037
106. Chronopoulos A, Thorpe SD, Cortes E, Lachowski D, Rice AJ, Mykuliak VV, et al. Syndecan-4 Tunes Cell Mechanics by Activating the Kindlin-Integrin-RhoA Pathway. Nat Mater (2020) 19:669–78. doi: 10.1038/s41563-019-0567-1
107. Dib K, Melander F, Axelsson L, Dagher M-C, Aspenstrom P, Andersson T. Down-Regulation of Rac Activity During Beta2 Integrin-Mediated Adhesion of Human Neutrophils. J Biol Chem (2003) 278(26):24181–8. doi: 10.1074/jbc.M302300200
108. Nurmi SM, Autero M, Raunio AK, Gahmberg CG, Fagerholm SC. Phosphorylation of the LFA-1 Integrin β2-Chain on Thr-758 Leads to Adhesion, Rac-1/Cdc42 Activation, and Stimulation of CD69 Expression in Human T Cells. J Biol Chem (2006) 282:968–75. doi: 10.1074/jbc.M608524200
109. Schiller HB, Fässler R. Mechanosensitivity and Compositional Dynamics of Cell-Matrix Adhesions. EMBO Rep (2013) 14(6):509–19. doi: 10.1038/embor.2013.49
110. Winograd-Katz SE, Fässler R, Geiger B, Legate KR. The Integrin Adhesome: From Genes and Proteins to Human Disease. Nat Rev Mol Cell Biol (2014) 15:273–88. doi: 10.1038/nrm3769
111. Bachmann M, Kukkurainen S, Hytönen VP, Wehrle-Haller XB. Cell Adhesion by Integrins. Physiol Rev (2019) 99:1655–99. doi: 10.1152/physrev.00036.2018
112. Kuo J-C, Han X, Hsiao C-T, Yates JR III., Waterman CM. Analysis of the Myosin-II-Responsive Focal Adhesion Proteome Reveals a Role for β−Pix in Negative Regulation of Focal Adhesion Maturation. Nat Cell Biol (2011) 13(4):383–93. doi: 10.1038/ncb2216
113. Horton ER, Byron A, Askari JA, Ng DHJ, Millon-Frémillon A, Robertson J, et al. Definition of a Consensus Integrin Adhesome and its Dynamics During Adhesion Complex Assembly and Disassembly. Nat Cell Biol (2015) 17:1577–87. doi: 10.1038/ncb3257
114. Calderwood DA, Ginsberg MH. Talin Forges the Links Between Integrins and Actin. Nat Cell Biol (2003) 5(8):694–6. doi: 10.1038/ncb0803-694
115. Bledzka K, Bialkowska K, Sossey-Alaoui K, Vaynberg J, Pluskota E, Qin J, et al. Kindlin-2 Directly Binds Actin and Regulates Integrin Outside-in Signaling. J Cell Biol (2016) 213:97–108. doi: 10.1083/jcb.201501006
116. Zhang M, March ME, Lane WS, Long EO. A Signaling Network Induced by β2 Integrin Controls the Polarization of Lytic Granulesin Cytotoxic Cells. Sci Signaling (2014) 7(346):96. doi: 10.1126/scisignal.2005629
117. Calderwood DA, Zent R, Grant R, Rees DJG, Hynes RO, Ginsberg MH. The Talin Head Domain Binds to Integrin β Subunit Cytoplasmic Tails and Regulates Integrin Activation. J Biol Chem (1999) 274:28071–4. doi: 10.1074/jbc.274.40.28071
118. Rio AD, Perez-Jimenez R, Liu R, Roca-Cusachs P, Fernandez JM, Sheetz MP. Stretching Single Talin Rod Molecules Activates Vinculin Binding. Science (2009) 323(5914):638–41. doi: 10.1126/science.1162912
119. Yao M, Goult BT, Chen H, Cong P, Sheetz MP, Yan J. Mechanical Activation of Vinculin Binding to Talin Locks Talin in an Unfolded Conformation. Sci Rep (2014) 4(4610):7. doi: 10.1038/srep04610
120. Sakai Y, Tsunekawa M, Ohta K, Shimizu T, Pastuhov SI, Hanafusa H, et al. The Integrin Signaling Network Promotes Axon Regeneration via the Src–Ephexin-RhoA GTPase Signaling Axis. J Neurosci (2021) 41(22):4754–67. doi: 10.1523/JNEUROSCI.2456-20.2021
121. Haining AWM, Rahikainen R, Cortes E, Lachowski D, Rice A, von Essen M, et al. Mechanotransduction in Talin Through the Interaction of the R8 Domain With DLC1. PloS Biol (2018) 16(7):20. doi: 10.1371/journal.pbio.2005599
122. Healy KD, Hodgson L, Kim T-Y, Shutes A, Maddileti S, Juliano RL, et al. DLC-1 Suppresses Non-Small Cell Lung Cancer Growth and Invasion by RhoGAP-Dependent and Independent Mechanisms. Mol Carcinogenesis (2008) 47:326–37. doi: 10.1002/mc.20389
123. Moser M, Nieswandt B, Ussar S, Pozgajova M, Fässler R, et al. Kindlin-3 is Essential for Integrin Activation and Platelet Aggregation. Nat Med (2008) 14:325–30. doi: 10.1038/nm1722
124. Lu L, Lin C, Yan Z, Wang S, Zhang Y, Wang S, et al. Kindlin-3 Is Essential for the Resting Alpha4beta1 Integrin-Mediated Firm Cell Adhesion Under Shear Flow Conditions. J Biol Chem (2016) 291(19):10363–71. doi: 10.1074/jbc.M116.717694
125. Manevich-Mendelson E, Feigelson SW, Pasvolsky R, Aker M, Grabovsky V, Shulman Z, et al. Loss of Kindlin-3 in LAD-III Eliminates LFA-1 But Not VLA-4 Adhesiveness Developed Under Shear Flow Conditions. Blood (2009) 114:2344–53. doi: 10.1182/blood-2009-04-218636
126. Klapproth S, Moretti FA, Zeiler M, Ruppert R, Breithaupt U, Mueller S, et al. Minimal Amounts of Kindlin-3 Suffice for Basal Platelet and Leukocyte Functions in Mice. Blood (2015) 126(24):2592–600. doi: 10.1182/blood-2015-04-639310
127. Ussar S, Wang H-V, Linder S, Fässler R, Moser M. The Kindlins: Subcellular Localization and Expression Duringmurine Development. Exp Cell Res (2006) 312(16):3142–51. doi: 10.1016/j.yexcr.2006.06.030
128. Chatterjee D, Zhiping LL, Tan S-M, Bhattacharjya S. NMR Structure, Dynamics and Interactions of the Integrin β2 Cytoplasmic Tail With Filamin Domain Igflna21. Sci Rep (2018) 8(5490):14. doi: 10.1038/s41598-018-23866-6
129. Kiema T, Lad Y, Jiang P, Oxley CL, Baldassarre M, Wegener KL, et al. The Molecular Basis of Filamin Binding to Integrins and Competition With Talin. Mol Cell (2006) 21(3):337–47. doi: 10.1016/j.molcel.2006.01.011
130. Takala H, Nurminen E, Nurmi SM, Aatonen M, Strandin T, Takatalo M, et al. β2 Integrin Phosphorylation on Thr758 Acts as a Molecular Switch to Regulate 14-3-3 and Filamin Binding. Blood (2008) 112(5):1853–62. doi: 10.1182/blood-2007-12-127795
131. Valmu L, Fagerholm S, Suila H, Gahmberg CG. The Cytoskeletal Association of CD11/CD18 Leukocyte Integrins in Phorbol Ester-Activated Cells Correlates With CD18 Phosphorylation. Eur J Immunol (1999) 29:2107–18. doi: 10.1002/(SICI)1521-4141(199907)29:07<2107::AID-IMMU2107>3.0.CO;2-T
132. Loo DT, Kanner SB, Aruffo A. Filamin Binds to the Cytoplasmic Domain of the Beta1-Integrin. Identification of Amino Acids Responsible for This Interaction. J Biol Chem (1998) 273:23304–12. doi: 10.1074/jbc.273.36.23304
133. Rosa J-P, Raslova H, Bryckaert M. Filamin A: Key Actor in Platelet Biology. blood (2019) 134(16):1279–88. doi: 10.1182/blood.2019000014
134. Calderwood DA, Huttenlocher A, Kiosses WB, Rose DM, Woodside DG, Schwartz MA, et al. Increased Filamin Binding to β-Integrin Cytoplasmic Domains Inhibits Cell Migration. Nat Cell Biol (2001) 3:1060–8. doi: 10.1038/ncb1201-1060
135. Rognoni L, Stigler J, Pelz B, Ylänne J, Rief M. Dynamic Force Sensing of Filamin Revealed in Single-Molecule Experiments. PNAS (2012) 109(48):19679–84. doi: 10.1073/pnas.1211274109
136. Nakamura F, Stossel TP, Hartwig JH. The Filamins Organizers of Cell Structure and Function. Cell Adhesion Migration (2011) 5(2):160–9. doi: 10.4161/cam.5.2.14401
137. Kumar A, Shutova MS, Tanaka K, Iwamoto DV, Calderwood DA, Svitkina TM, et al. Filamin A Mediates Isotropic Distribution of Applied Force Across the Actin Network. J Cell Biol (2019) 218(8):2481–91. doi: 10.1083/jcb.201901086
138. Glogauer M, Pam A, Chou D, Janmey PA, Downey GP, McCulloch CAG. The Role of Actin-Binding Protein 280 in Integrin-Dependent Mechanoprotection. J Biol Chem (1998) 273:1689–98. doi: 10.1074/jbc.273.3.1689
139. Ehrlicher AJ, Nakamura F, Hartwig JH, Weitz DA, Stossel TP. Mechanical Strain in Actin Networks Regulates FilGAP and Integrin Binding to Filamin A. Nature (2011) 478:260–3. doi: 10.1038/nature10430
140. Ohta Y, Hartwig JH, Stossel TP. FilGAP, a Rho- and ROCK-Regulated GAP for Rac Binds Filamin A to Control Actin Remodelling. Nat Cell Biol (2006) 8:803–14. doi: 10.1038/ncb1437
141. Heasman SJ, Ridley AJ. Mammalian Rho GTPases: New Insights Into Their Functions From In Vivo Studies. Nat Rev Mol Cell Biol (2008) 9:690–701. doi: 10.1038/nrm2476
142. Algaber A, AlHaidari A, Madhi R, Rahman M, Syk I, Thorlacius H. MicroRNA−340−5p Inhibits Colon Cancer Cell Migration via Targeting of RhoA. Sci Rep (2020) 10(16934):1–7. doi: 10.1038/s41598-020-73792-9
143. Moon M-Y, Kim H-J, Kim M-J, Uhm S, Park J-W, Suk K-T. Rap1 Regulates Hepatic Stellate Cell Migration Through the Modulation of RhoA Activity in Response to TGF−β1. Int J Mol Med (2019) 44:491–502. doi: 10.3892/ijmm.2019.4215
144. Kim J-M, Kim MY, Lee K, Jeong D. Distinctive and Selective Route of PI3K/PKCa-PKCd/RhoA-Rac1 Signaling in Osteoclastic Cell Migration. Mol Cell Endocrinol (2016) p:261–7. doi: 10.1016/j.mce.2016.08.042
145. Worthylake RA, Lemoine S, Watson JM, Burridge K. RhoA is Required for Monocyte Tail Retraction During Transendothelial Migration. J Cell Biol (2001) 154(1):147–60. doi: 10.1083/jcb.200103048
146. Sahai E, Marshall CJ. Differing Modes of Tumour Cell Invasion Have Distinct Requirements for Rho/ROCK Signalling and Extracellular Proteolysis. Nat Cell Biol (2003) 5:711–9. doi: 10.1038/ncb1019
147. Worthylake RA, Burridge K. RhoA and ROCK Promote Migration by Limiting Membrane Protrusions. J Biol Chem (2003) p:13578–84. doi: 10.1074/jbc.M211584200
148. Watanabe S, Okawa K, Miki T, Sakamoto S, Morinaga T, Segawa K, et al. Rho and Anillin-Dependent Control of Mdia2 Localization and Function in Cytokinesis. Mol Biol Cell (2010) 21:3193–204. doi: 10.1091/mbc.e10-04-0324
149. Staus DP, Taylor JM, Mack CP. Enhancement of Mdia2 Activity by Rho-Kinase-Dependent Phosphorylation of the Diaphanous Autoregulatory Domain. Biochem J (2011) 439:57–65. doi: 10.1042/BJ20101700
150. Giagulli C, Scarpini E, Ottoboni L, Narumiya S, Butcher EC, Constantin G, et al. RhoA and Zeta PKC Control Distinct Modalities of LFA-1 Activation by Chemokines: Critical Role of LFA-1 Affinity Triggering in Lymphocyte In Vivo Homing. Immunity (2004) 20(1):25–35. doi: 10.1016/S1074-7613(03)00350-9
151. Kinashi T. Intracellular Signalling Controlling Integrin Activation in Lymphocytes. Nat Rev Immunol (2005) 5:546–59. doi: 10.1038/nri1646
152. Vielkind S, Gallagher-Gambarelli M, Gomez M, Hinton HJ, Cantrell DA. Integrin Regulation by RhoA in Thymocytes. J Immunol (2005) 175:350–7. doi: 10.4049/jimmunol.175.1.350
153. Amison RT, Momi S, Morris A, Manni G, Keir S, Gresele P, et al. RhoA Signaling Through Platelet P2Y1 Receptor Controls Leukocyte Recruitment in Allergic Mice. J Allergy Clin Immunol (2015) 135(2):528–38. doi: 10.1016/j.jaci.2014.09.032
154. Ridley AJ, Paterson HF, Johnston CL, Diekmann D, Hall A. The Small Gtp-Binding Protein Rac Regulates Growth Factor-Induced Membrane Ruffling. Cell (1992) 70:401–10. doi: 10.1016/0092-8674(92)90164-8
155. Nobes CD, Hall A. Rho, Rac, and Cdc42 GTPases Regulate the Assembly of Multimolecular Focal Complexes Associated With Actin Stress Fibers, Lamellipodia, and Filopodia. Cell (1995) 81:53–62. doi: 10.1016/0092-8674(95)90370-4
156. Allen WE, Jones GE, Pollard JW. Rho, Rac and Cdc42 Regulate Actin Organization and Cell Adhesion in Macrophages. J Cell Sci (1997) 110:707–20. doi: 10.1242/jcs.110.6.707
157. Vincent S, Jeanteur P, Fort P. Growth-Regulated Expression of Rhog, a New Member of the Ras Homolog Gene Family. Mol Cell Biol (1992) 12(7):3138–48. doi: 10.1128/MCB.12.7.3138
158. Didsbury J, Weber RF, Bokoch GM, Evans T, Snyderma R. Rac, a Novel Ras-Related Family of Proteins That Are Botulinum Toxin Substrates. J Biol Chem (1989) 264(28):16378–82. doi: 10.1016/S0021-9258(19)84716-6
159. Shirsat NV, Pignolo RJ, Kreider BL, Rovera G. A Member of the Ras Gene Superfamily is Expressed Specifically in T, B and Myeloid Hemopoietic Cells. Oncogene (1990) 5(5):769–72.
160. Corbetta S, Gualdoni S, Albertinazzi C, Paris S, Croci L, Consalez GG, et al. Generation and Characterization of Rac3 Knockout Mice. Mol Cell Biol (2005) 25(13):5763–76. doi: 10.1128/MCB.25.13.5763-5776.2005
161. Haataja L, Groffen J, Heisterkamp N. Characterization of RAC3, a Novel Member of the Rho Family. J Biol Chem (1997) 272(33):20384–8. doi: 10.1074/jbc.272.33.20384
162. Fukui Y, Hashimoto O, Sanui T, Oono T, Koga H, Abe M, et al. Haematopoietic Cell-Specific CDM Family Protein DOCK2 is Essential for Lymphocyte Migration. Nature (2001) 412:826–31. doi: 10.1038/35090591
163. Pollard TD. Regulation of Actin Filament Assembly by Arp2/3 Complex and Formins. Annu Rev Biophys Biomolec Structures (2007) 36:451–77. doi: 10.1146/annurev.biophys.35.040405.101936
164. Atat OE, Fakih A, El-Sibai M. RHOG Activates RAC1 Through CDC42 Leading to Tube Formation in Vascular Endothelial Cells. cells (2019) 8(2):17. doi: 10.3390/cells8020171
165. Helden S. Rho GTPase Expression in Human Myeloid Cells. PLOoS One (2012) 7(8):14. doi: 10.1371/journal.pone.0042563
166. Dallery E, Galiègue-Zouitina S, Collyn-d'Hooghe M, Quief S, Denis C, Hildebrand MP, et al. TTF, a Gene Encoding a Novel Small G Protein, Fuses to the Lymphoma-Associated LAZ3 Gene by T(3;4) Chromosomal Translocation. Oncogene (1995) 10(11):2171–8.
167. Lahousse SB, Smorowski A-L, Denis C, Daniéle L, Kerckaert J-P, Galiégue-Zouitina S. Structural Features of Hematopoiesis-Specific RhoH/ARHH Gene: High Diversity of 5V-UTR in Different Hematopoietic Lineages Suggests a Complex Post-Transcriptional Regulation. Gene (2004) 343:55–68. doi: 10.1016/j.gene.2004.08.022
168. Gu Y, Jasti AC, Jansen M, Siefring JE. RhoH, a Hematopoietic-Specific Rho GTPase, Regulates Proliferation, Survival, Migration, and Engraftment of Hematopoietic Progenitor Cells. blood (2005) 105(4):1467–75. doi: 10.1182/blood-2004-04-1604
169. Gu Y, Chae H-D, Siefring JE, Jasti AC, Hildeman DA, Williams DA. RhoH GTPase Recruits and Activates Zap70 Required for T Cell Receptor Signaling and Thymocyte Development. Nat Immunol (2006) 7(11):1182–90. doi: 10.1038/ni1396
170. Dorn T, Kuhn U, Bungartz G, Stiller S, Bauer M, Ellwart J, et al. RhoH is Important for Positive Thymocyte Selection and T-Cell Receptor Signaling. Blood (2007) 109(6):2346–55. doi: 10.1182/blood-2006-04-019034
171. Sanchez-Aguilera A, Rattmann I, Drew DZ, Müller LUW, Summey V, Lucas DM, et al. Involvement of RhoH GTPase in the Development of B-Cell Chronic Lymphocytic Leukemia. Leukemia (2010) 24(1):97–104. doi: 10.1038/leu.2009.217
172. Li X, Bu X, Lu B, Avraham H, Flavell RA, Lim B. The Hematopoiesis-Specific GTP-Binding Protein RhoH Is GTPase Deficient and Modulates Activities of Other Rho GTPases by an Inhibitory Function. Mol Cell Biol (2002) 22(4):1158–71. doi: 10.1128/MCB.22.4.1158-1171.2002
173. Poh Y-C, Shevtsov SP, Chowdhury F, Wu DC, Na S, Dundr M, et al. Dynamic Force-Induced Direct Dissociation of Protein Complexes in a Nuclear Body in Living Cells. Nat Commun (2012) 3(866):10. doi: 10.1038/ncomms1873
174. Swift J, Ivanovska IL, Buxboim A, Harada T, Dingal PCDP, Pinter J, et al. Nuclear Lamin-A Scales With Tissue Siffness and Enhances Matrix-Directed Differentiation. Science (2013) 341. doi: 10.1126/science.1240104
175. Stephens AD, Banigan EJ, Adam SA, Goldman RD, Marko JF. Chromatin and Lamin A Determine Two Different Mechanical Response Regimes of the Cell Nucleus. Mol Biol Cell (2017) 28(14):1984–96. doi: 10.1091/mbc.e16-09-0653
176. Lammerding J, Fong LG, Ji JY, Reue K, Stewart CL, Young SG, et al. Lamins A and C But Not Lamin B1 Regulate Nuclear Mechanics. J Biol Chem (2006) 281:25768–80. doi: 10.1074/jbc.M513511200
177. Guenther C, Faisal I, Fusciello M, Sokolova M, Harjunpää H, Ilander M, et al. β2-Integrin Adhesion Regulates Dendritic Cell Epigenetic and Transcriptional Landscapes to Restrict Dendritic Cell Maturation and Tumor Rejection. Cancer Immunol Res (2021) 9:1354–69. doi: 10.1158/2326-6066.CIR-21-0094
178. Wang P, Dreger M, Madrazo E, Williams CJ, Samaniego R, Hodson NW, et al. WDR5 Modulates Cell Motility and Morphology and Controls Nuclear Changes Induced by a 3D Environment. PNAS (2018) 115(34):8581–6. doi: 10.1073/pnas.1719405115
179. Zhang X, Cook PC, Zindy E, Williams CJ, Jowitt TA, Streuli CH, et al. Integrin Alpha4beta1 Controls G9a Activity That Regulates Epigenetic Changes and Nuclear Properties Required for Lymphocyte Migration. Nucleic Acids Res (2015) 44(7):3031–44. doi: 10.1093/nar/gkv1348
180. Hamminger P, Marchetti L, Preglej T, Platzer R, Zhu C, Kamnev A, et al. Histone Deacetylase 1 Controls CD4+ T Cell Trafficking in Autoinflammatory Diseases. J Autoimmun (2021) 119:13. doi: 10.1016/j.jaut.2021.102610
181. Cougoule C, Van Goethem E, Le Cabec V, Lafouresse F, Dupré L, Mehraj V, et al. Blood Leukocytes and Macrophages of Various Phenotypes Have Distinct Abilities to Form Podosomes and to Migrate in 3D Environments. Eur J Cell Biol (2012) 91:938–49. doi: 10.1016/j.ejcb.2012.07.002
182. Leung R, Wang Y, Cuddy K, Sun C, Magalhaes J, Grynpas M, et al. Filamin A Regulates Monocyte Migration Through Rho Small GTPases During Osteoclastogenesis. J Bone Mineral Res (2010) 25(5):1077–91. doi: 10.1359/jbmr.091114
183. Honing H, van den Berg TK, van der Pol SMA, Dijkstra CD, van der Kammen RA, Collard JG, et al. RhoA Activation Promotes Transendothelial Migration of Monocytes via ROCK. J Leukocyte Biol (2004) 75:6. doi: 10.1189/jlb.0203054
184. Aepfelbacher M, Essler M, Huber E, Czech A, Weber PC. Rho Is a Negative Regulator of Human Monocyte Spreading. J Immunol (1996) 157:5070–5.
185. Weber KSC, Klickstein LB, Weber PC, Weber C. Chemokine-Induced Monocyte Transmigration Requires Cdc42-Mediated Cytoskeletal Changes. Eur J Immunol (1998) 28:2245–51. doi: 10.1002/(SICI)1521-4141(199807)28:07<2245::AID-IMMU2245>3.0.CO;2-V
186. Worbs T, Hammerschmidt SI, Förster R. Dendritic Cell Migration in Health and Disease. Nat Rev Immunol (2017) 17:30–48. doi: 10.1038/nri.2016.116
187. Nitschké M, Aebischer D, Abadier M, Haener S, Lucic M, Vigl B, et al. Differential Requirement for ROCK in Dendritic Cell Migration Within Lymphatic Capillaries in Steady-State and Inflammation. Blood (2012) 120(11):2249–58. doi: 10.1182/blood-2012-03-417923
188. Cougoule C, Lastrucci C, Guiet R, Mascarau R, Meunier E, Lugo-Villarino G, et al. Podosomes, But Not the Maturation Status, Determine the Protease-Dependent 3d Migration in Human Dendritic Cells. Front Immunol (2018) 9:846. doi: 10.3389/fimmu.2018.00846
189. Dieu M-C, Vanbervliet B, Vicari A, Bridon J-M, Oldham E, Aït-Yahia S, et al. Selective Recruitment of Immature and Mature Dendritic Cells by Distinct Chemokines Expressed in Different Anatomic Sites. J Exp Med (1998) 188(2):373–86. doi: 10.1084/jem.188.2.373
190. Saeki H, Moore AM, Brown MJ, Hwang ST. Cutting Edge: Secondary Lymphoid Tissue Chemokine (SLC) and CC Chemokine Receptor 7 (CCR7) Participate in the Emigration Pathway of Mature Dendritic Cells From the Skin to Regional Lymph Nodes. J Immunol (1999) 162:2472–5.
191. Braun A, Worbs T, Moschovakis GL, Halle S, Hoffmann K, Bölter J, et al. Afferent Lymph–Derived T Cells and DCs Use Different Chemokine Receptor CCR7-Dependent Routes for Entry Into the Lymph Node and Intranodal Migration. Nat Immunol (2011) 12:879–87. doi: 10.1038/ni.2085
192. Jakubzick C, Tacke F, Llodra J, van Rooijen N, Randolph GJ. Modulation of Dendritic Cell Trafficking to and From the Airways. J Immunol (2006) 176:3578–84. doi: 10.4049/jimmunol.176.6.3578
193. Morrison VL, James MJ, Grzes K, Cook P, Glass DG, Savinko T, et al. Loss of Beta2-Integrin-Mediated Cytoskeletal Linkage Reprograms Dendritic Cells to a Mature Migratory Phenotype. Nat Commun (2014) 5:13. doi: 10.1038/ncomms6359
194. van Goethem E, Poincloux R, Gauffre F, Maridonneau-Parini I, Le Cabec V. Matrix Architecture Dictates Three-Dimensional Migration Modes of Human Macrophages: Differential Involvement of Proteases and Podosome-Like Structures. J Immunol (2010) 184:1049–61. doi: 10.4049/jimmunol.0902223
195. Guiet R, Van Goethem E, Cougoule C, Balor S, Valette A, Al Saati T, et al. The Process of Macrophage Migration Promotes Matrix Metalloproteinase-Independent Invasion by Tumor Cells. J Immunol (2011) 187(7):3806–14. doi: 10.4049/jimmunol.1101245
196. Guiet R, Vérollet C, Lamsoul I, Cougoule C, Poincloux R, Labrousse A, et al. Macrophage Mesenchymal Migration Requires Podosome Stabilization by Filamin A. J Biol Chem (2012) 287:13051–62. doi: 10.1074/jbc.M111.307124
197. Bandaru S, Ala C, Salimi R, Akula MK, Ekstrand M, Devarakonda S, et al. Targeting Filamin A Reduces Macrophage Activity and Atherosclerosis. Circulation (2019) 140:67–79. doi: 10.1161/CIRCULATIONAHA.119.039697
198. Cui K, Ardell CL, Podolnikova NP, Yakubenko VP. Distinct Migratory Properties of M1, M2, and Resident Macrophages Are Regulated by αdβ2 and αmβ2 Integrin-Mediated Adhesion. Front Immunol (2018) 9. doi: 10.3389/fimmu.2018.02650
199. Allen WE, Zicha D, Ridley AJ, Jones GE. A Role for Cdc42 in Macrophage Chemotaxis. J Cell Biol (1998) 141(5):1147–57. doi: 10.1083/jcb.141.5.1147
200. Pradip D, Peng X, Durden DL. Rac2 Specificity in Macrophage Integrin Signaling. J Biol Chem (2003) 278(43):41661–9. doi: 10.1074/jbc.M306491200
201. Wells CM, Walmsley M, Ooi S, Tybulewicz V, Ridley AJ. Rac1-Deficient Macrophages Exhibit Defects in Cell Spreading and Membrane Ruffling But Not Migration. J Cell Sci (2003) 117(7):1259–68. doi: 10.1242/jcs.00997
202. Yamauchi A, Kim C, Li S, Marchal CC, Towe J, Atkinson SJ, et al. Rac2-Deficient Murine Macrophages Have Selective Defects in Superoxide Production and Phagocytosis of Opsonized Particles. J Immunol (2004) 173:5971–9. doi: 10.4049/jimmunol.173.10.5971
203. Wheeler AP, Wells CM, Smith SD, Vega FM, Henderson RB, Tybulewicz VL, et al. Rac1 and Rac2 Regulate Macrophage Morphology But are Not Essential for Migration. J Cell Sci (2006) 119:2749–57. doi: 10.1242/jcs.03024
204. Harry GJ. Microglia During Development and Aging. Pharmacol Ther (2013) 139(3):313–26. doi: 10.1016/j.pharmthera.2013.04.013
205. Marshall GP, Demir M, Steindler DA, Laywell ED. Subventricular Zone Microglia Possess a Unique Capacity for Massive In Vitro Expansion. GLIA (2008) 56(16):1799–808. doi: 10.1002/glia.20730
206. Kurpius D, Wilson N, Fuller L, Hoffman A, Dailey ME. Early Activation, Motility, and Homing of Neonatal Microglia to Injured Neurons Does Not Require Protein Synthesis. Glia (2006) 54:58–70. doi: 10.1002/glia.20355
207. Haapaniemi H, Tomita M, Tanahashi N, Takeda H, Yokoyama M, Fukuuchi Y. Non-Amoeboid Locomotion of Cultured Microglia Obtained From Newborn Rat Brain. Neurosci Lett (1995) 193:121–4. doi: 10.1016/0304-3940(95)11683-N
208. Carbonell WS, Murase S-I, Horwitz AF, Mandell JW. Migration of Perilesional Microglia After Focal Brain Injury and Modulation by CC Chemokine Receptor 5: An In Situ Time-Lapse Confocal Imaging Study. J Neurosci (2005) 25(30):7040–7. doi: 10.1523/JNEUROSCI.5171-04.2005
209. Neumann J, Gunzer M, Gutzeit HO, Ullrich O, Reymann KG, Dinkel K. Microglia Provide Neuroprotection After Ischemia. FASEB J (2006) 20:714–6. doi: 10.1096/fj.05-4882fje
210. Hyun Y-M, Hong C-W. Deep Insight Into Neutrophil Trafficking in Various Organs. J Leukocyte Biol (2017) 102:617–29. doi: 10.1189/jlb.1RU1216-521R
211. Kienle K, Lämmermann T. Neutrophil Swarming: An Essential Process of the Neutrophil Tissue Response. Immunol Rev (2016) 273:76–93. doi: 10.1111/imr.12458
212. Lämmermann T, Afonso PV, Angermann BR, Wang JM, Kastenmüller W, Parent CA, et al. Neutrophil Swarms Require LTB4 and Integrins at Sites of Cell Death In Vivo. Nature (2013) 498(7454):371–5. doi: 10.1038/nature12175
213. Yamahashi Y, Cavnar PJ, Hind LE, Berthier E, Bennin DA, Beebe D, et al. Integrin Associated Proteins Differentially Regulate Neutrophil Polarity and Directed Migration in 2D and 3D. Biomed Microdevices (2015) 17(100):9. doi: 10.1007/s10544-015-9998-x
214. Yago T, Petrich BG, Zhang N, Liu Z, Shao B, Ginsberg MH, et al. Blocking Neutrophil Integrin Activation Prevents Ischemia–Reperfusion Injury. J Exp Med (2015) 212(8):1267–81. doi: 10.1084/jem.20142358
215. Roth H, Samereier M, Begandt D, Pick R, Salvermoser M, Brechtefeld D, et al. Filamin A Promotes Efficient Migration and Phagocytosis of Neutrophil-Like HL-60 Cells. Eur J Cell Biol (2017) 96(6):553–66. doi: 10.1016/j.ejcb.2017.05.004
216. Sun C, Forster C, Nakamura F, Glogauer M. Filamin-A Regulates Neutrophil Uropod Retraction Through RhoA During Chemotaxis. PloS One (2013) 8(10):8. doi: 10.1371/journal.pone.0079009
217. Uotila LM, Guenther C, Savinko T, Lehti TA, Fagerholm SC. Filamin A Regulates Neutrophil Adhesion, Production of Reactive Oxygen Species, and Neutrophil Extracellular Trap Release. J Immunol (2017) 199(210):3644–53. doi: 10.4049/jimmunol.1700087
218. Sun CX, Downey GP, Zhu F, Koh ALY, Thang H, Glogauer M. Rac1 is the Small GTPase Responsible for Regulating the Neutrophil Chemotaxis Compass. Blood (2004) 104(12):3758–65. doi: 10.1182/blood-2004-03-0781
219. Roberts AW, Kim C, Zhen L, Lowe JB, Kapur R, Petryniak B, et al. Deficiency of the Hematopoietic Cell-Specific Rho Family GTPase Rac2 Is Characterized by Abnormalities in Neutrophil Function and Host Defense. Immunity (1999) 10:183–96. doi: 10.1016/s1074-7613(00)80019-9
220. Boutboul D, Kuehn HS, Van de Wyngaert Z, Niemela JE, Callebaut I, Stoddard J, et al. Dominant-Negative IKZF1 Mutations Cause a T, B, and Myeloid Cell Combined Immunodeficiency. J Clin Invest (2018) 128(7):3071–87. doi: 10.1172/JCI98164
221. Szczur K, Xu H, Atkinson S, Zheng Y, Filippi M-D. Rho GTPase CDC42 Regulates Directionality and Random Movement via Distinct MAPK Pathways in Neutrophils. Blood (2006) 108(13):4205–13. doi: 10.1182/blood-2006-03-013789
222. Yang HW, Collins SR, Meyer T. Locally Excitable Cdc42 Signals Steer Cells During Chemotaxis. Nat Cell Biol (2015) 18(2):191–201. doi: 10.1038/ncb3292
223. Kumar S, Xu J, Perkins C, Guo F, Snapper S, Finkelman FD, et al. Cdc42 Regulates Neutrophil Migration via Crosstalk Between WASp, CD11b, and Microtubules. Blood (2012) 120(17):3563–74. doi: 10.1182/blood-2012-04-426981
224. Iikura M, Ebisawa M, Yamaguchi M, Tachimoto H, Ohta K, Yamamoto K, et al. Transendothelial Migration of Human Basophils. J Immunol (2004) 173:5189–95. doi: 10.4049/jimmunol.173.8.5189
225. Dahlin JS, Hallgren J. Mast Cell Progenitors: Origin, Development and Migration to Tissues. Mol Immunol (2015) 63:9–17. doi: 10.1016/j.molimm.2014.01.018
226. Okayama Y, Kawakami T. Development, Migration, and Survival of Mast Cells. Immunol Res (2006) 34(2):97–115. doi: 10.1385/IR:34:2:97
227. Rosenkranz AR, Coxon A, Maurer M, Gurish MF, Austen KF, Friend DS, et al. Cutting Edge: Impaired Mast Cell Development and Innate Immunity in Mac-1 (CD11b/CD18, CR3)-Deficient Mice. J Immunol (1998) 161:6463–7.
228. Klein O, Krier-Burris RA, Lazki-Hagenbach P, Gorzalczany Y, Mei Y, Ji P, et al. Mammalian Diaphanous-Related Formin 1 (Mdia1) Coordinates Mast Cell Migration and Secretion Through its Actin-Nucleating Activity. J Allergy Clin Immunol (2019) 144(4):1074–90. doi: 10.1016/j.jaci.2019.06.028
229. Gruber BL, Marchese MJ, Kew R. Angiogenic Factors Stimulate Mast-Cell Migration. Blood (1995) 86(7):2488–93. doi: 10.1182/blood.V86.7.2488.2488
230. Azizkhan RG, Azizkhan JC, Zetter BR, Folkman J. Mast Cell Heparin Stimulates Migration Of Capillary Endothelial Cells In Vitro. J Exp Med (1980) 152:931–44. doi: 10.1084/jem.152.4.931
231. Lee BJ, Mace EM. From Stem Cell to Immune Effector: How Adhesion, Migration, and Polarity Shape T-Cell and Natural Killer Cell Lymphocyte Development In Vitro and In Vivo. Mol Biol Cell (2020) 31(10):981–91. doi: 10.1091/mbc.E19-08-0424
232. Beuneu H, Deguine J, Breart B, Mandelboim O, Di Santo JP, Bousso P. Dynamic Behavior of NK Cells During Activation in Lymph Nodes. Blood (2009) 114(15):3227–34. doi: 10.1182/blood-2009-06-228759
233. Bajénoff M, Breart B, Huang AYC, Qi H, Cazareth J, Braud VM, et al. Natural Killer Cell Behavior in Lymph Nodes Revealed by Static and Real-Time Imaging. J Exp Med (2006) 203(3):619–31. doi: 10.1084/jem.20051474
234. Mace EM, Gunesch JT, Dixon A, Orange JS. Human NK Cell Development Requires CD56-Mediated Motility and Formation of the Developmental Synapse. Nat Commun (2016) 7(12171):13. doi: 10.1038/ncomms12171
235. Lee BJ, Mace EM. Acquisition of Cell Migration Defines NK Cell Differentiation From Hematopoietic Stem Cell Precursors. Mol Biol Cell (2017) 28:3573–81. doi: 10.1091/mbc.e17-08-0508
236. Timonen T. Natural Killer Cells: Endothelial Interactions, Migration, and Target Cell Recognition. J Leukocyte Biol (1997) 62(6):693–701. doi: 10.1002/jlb.62.6.693
237. Worbs T, Mempel TR, Bölter J, von Andrian UH, Förster R. CCR7 Ligands Stimulate the Intranodal Motility of T Lymphocytes In Vivo. J Exp Med (2007) 204(3):489–95. doi: 10.1084/jem.20061706
238. Morrison VL, MacPherson M, Savinko T, Lek HS, Prescott A, Fagerholm SC. The β2 Integrin–Kindlin-3 Interaction is Essential for T-Cell Homing But Dispensable for T-Cell Activation In Vivo. Blood (2013) 122(8):1428–36. doi: 10.1182/blood-2013-02-484998
239. Smith A, Carrasco YR, Stanley P, Kieffer N, Batista FD, Hogg N. A Talin-Dependent LFA-1 Focal Zone is Formed by Rapidly Migrating T Lymphocytes. J Cell Biol (2005) 170(1):141–51. doi: 10.1083/jcb.200412032
240. Luo J, Li D, Wei D, Wang X, Wang L, Zeug X. RhoA and RhoC are Involved in Stromal Cell-Derived Factor-1-Induced Cell Migration by Regulating F-Actin Redistribution and Assembly. Mol Cell Biochem (2017) 436:13–21. doi: 10.1007/s11010-017-3072-3
241. Garcı´a-Bernal D, Wright N, Sotillo-Mallo E, Nombela-Arrieta C, Stein JV, Bustelo XR, et al. Vav1 and Rac Control Chemokine-Promoted T Lymphocyte Adhesion Mediated by the Integrin Alpha4 Beta1. Mol Biol Cell (2005) 16:3223–5. doi: 10.1091/mbc.e04-12-1049
242. Banon-Rodriguez I, de Guinoa JS, Bernardini A, Ragazzini C, Fernandez E, Carrasco YR, et al. WIP Regulates Persistence of Cell Migration and Ruffle Formation in Both Mesenchymal and Amoeboid Modes of Motility. PloS One (2013) 8(8):14. doi: 10.1371/journal.pone.0070364
243. Guinoa J, Barrio L, Mellado M, Carrasco YR. CXCL13/CXCR5 Signaling Enhances BCR-Triggered B-Cell Activation by Shaping Cell Dynamics. Blood (2011) 118(6):1560–9. doi: 10.1182/blood-2011-01-332106
244. Liu X, Carman CV, Springer TA. Quantitative Analysis of B-Lymphocyte Migration Directed by CXCL13. Integr Biol (2016) 8:894–903. doi: 10.1039/c6ib00128a
245. Pan Y-R, Chen C-C, Chan Y-T, Wang H-J, Chien F-T, Chen Y-L, et al. STAT3-Coordinated Migration Facilitates the Dissemination of Diffuse Large B-Cell Lymphomas. Nat Commun (2018) 9(3696):16. doi: 10.1038/s41467-018-06134-z
246. Sharma P, Siddiqui BA, Anandhan S, Yadav SS, Subudhi SK, Gao J, et al. The Next Decade of Immune Checkpoint Therapy. Cancer Discov (2021) 11(4):838–57. doi: 10.1158/2159-8290.CD-20-1680
247. Bonaventura P, Shekarian T, Alcazer V, Valladeau-Guilemond J, Valsesia-Wittmann S, Amigorena S, et al. Cold Tumors: A Therapeutic Challenge for Immunotherapy. Front Immunol (2019) 10. doi: 10.3389/fimmu.2019.00168
248. Peranzoni E, Lemoine J, Vimeux L, Feuillet V, Barrin S, Kantari-Mimoun C, et al. Macrophages Impede CD8 T Cells From Reaching Tumor Cells and Limit the Efficacy of Anti–PD-1 Treatment. PNAS (2018) 115(17):E4041–50. doi: 10.1073/pnas.1720948115
249. Boissonnas A, Licata F, Poupel L, Jacquelin S, Fetler L, Krumeich S, et al. CD8+ Tumor-Infiltrating T Cells Are Trapped in the Tumor-Dendritic Cell Network. Neoplasia (2013) 15(1):85–94. doi: 10.1593/neo.121572
250. Engelhardt JJ, Boldajipour B, Beemiller P, Pandurangi P, Sorensen C, Werb Z, et al. Marginating Dendritic Cells of the Tumor Microenvironment Cross-Present Tumor Antigens and Stably Engage Tumor-Specific T Cells. Cell (2012) 21(3):402–17. doi: 10.1016/j.ccr.2012.01.008
251. Mi Y, Guo N, Luan J, Cheng J, Hu Z, Jiang P, et al. The Emerging Role of Myeloid-Derived Suppressor Cells in the Glioma Immune Suppressive Microenvironment. Front Immunol (2020) 11. doi: 10.3389/fimmu.2020.00737
252. Karin N. The Development and Homing of Myeloid-Derived Suppressor Cells: From a Two-Stage Model to a Multistep Narrative. Front Immunol (2020) 11. doi: 10.3389/fimmu.2020.557586
253. Liu L, Zhang L, Zhao S, Zhao X-Y, Min P-X, Ma Y-D, et al. Non-Canonical Notch Signaling Regulates Actin Remodeling in Cell Migration by Activating PI3K/AKT/Cdc42 Pathway. Front Pharmacol (2019) 10. doi: 10.3389/fphar.2019.00370
254. Redmond L, Ghosh A. The Role of Notch and Rho GTPase Signaling in the Control of Dendritic Development. Curr Opin Neurobiol (2001) 11:111–7. doi: 10.1016/S0959-4388(00)00181-1
255. Liu LY, Wang H, Xenakis JJ, Spencer LA. Notch Signaling Mediates Granulocyte-Macrophage Colony-Stimulating Factor Priming-Induced Transendothelial Migration of Human Eosinophils. Allergy (2015) 70:805–12. doi: 10.1111/all.12624
256. Noren NK, Niessen CM, Gumbiner BM, Burridge K. Cadherin Engagement Regulates Rho Family GTPases. J Biol Chem (2001) 276(36):33305–8. doi: 10.1074/jbc.C100306200
257. Nelson CM, Pirone DM, Tan JL, Chen CS. Vascular Endothelial-Cadherin Regulates Cytoskeletal Tension, Cell Spreading, and Focal Adhesions by Stimulating RhoA. Mol Biol Cell (2004) 15:2943–53. doi: 10.1091/mbc.e03-10-0745
258. Siddiqui KRR, Laffont S, Powrie F. E-Cadherin Marks a Subset of Inflammatory Dendritic Cells That Promote T Cell-Mediated Colitis. Immunity (2010) 32(4):557–67. doi: 10.1016/j.immuni.2010.03.017
259. Vafaei S, Zekiy AO, Khanamir RA, Zaman BA, Ghayourvahdat A, Azimizonuzi H, et al. Combination Therapy With Immune Checkpoint Inhibitors (ICIs); a New Frontier. Cancer Cell Int (2022) 22(2):27. doi: 10.1186/s12935-021-02407-8
260. Yamaguchi O, Kaira K, Hashimoto K, Mouri A, Miura Y, Shiono A, et al. Radiotherapy is an Independent Prognostic Marker of Favorable Prognosis in non-Small Cell Lung Cancer Patients After Treatment With the Immune Checkpoint Inhibitor, Nivolumab. Thorac Cancer (2019) 10(4):992–1000. doi: 10.1111/1759-7714.13044
Keywords: leukocyte migration, β2-integrins, RhoGTPases, epigenetics, actin tread milling, amoeboid migration, LFA-1
Citation: Guenther C (2022) β2-Integrins – Regulatory and Executive Bridges in the Signaling Network Controlling Leukocyte Trafficking and Migration. Front. Immunol. 13:809590. doi: 10.3389/fimmu.2022.809590
Received: 05 November 2021; Accepted: 11 March 2022;
Published: 22 April 2022.
Edited by:
Hao Sun, University of California, San Diego, United StatesReviewed by:
Ralph Böttcher, Max Planck Institute of Biochemistry, GermanyCopyright © 2022 Guenther. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carla Guenther, Y2FybGEuZ3VlbnRoZXJAYXJjb3IuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.