 Pamela A. McCombe
Pamela A. McCombe Judith M. Greer
Judith M. Greer
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CONCEPTUAL ANALYSIS article
Front. Immunol., 28 November 2022
Sec. Autoimmune and Autoinflammatory Disorders : Autoimmune Disorders
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.1059833
This article is part of the Research TopicSexual Dimorphism in Autoimmune and Immune-dysregulated DiseasesView all 7 articles
Experimental autoimmune encephalomyelitis (EAE) can be induced in many animal strains by inoculation with central nervous system antigens and adjuvant or by the passive transfer of lymphocytes reactive with these antigens and is widely used as an animal model for multiple sclerosis (MS). There are reports that female sex and pregnancy affect EAE. Here we review the effects of biological sex and the effects of pregnancy on the clinical features (including disease susceptibility) and pathophysiology of EAE. We also review reports of the possible mechanisms underlying these differences. These include sex-related differences in the immune system and in the central nervous system, the effects of hormones and the sex chromosomes and molecules unique to pregnancy. We also review sex differences in the response to factors that can modify the course of EAE. Our conclusion is that the effects of biological sex in EAE vary amongst animal models and should not be widely extrapolated. In EAE, it is therefore essential that studies looking at the effects of biological sex or pregnancy give full information about the model that is used (i.e. animal strain, sex, the inducing antigen, timing of EAE induction in relation to pregnancy, etc.). In addition, it would be preferable if more than one EAE model were used, to show if any observed effects are generalizable. This is clearly a field that requires further work. However, understanding of the mechanisms of sex differences could lead to greater understanding of EAE, and suggest possible therapies for MS.
The biological sex and the features of sexual dimorphism in mammals are determined by the complement of X and Y chromosomes (1), which evolved many millions of years ago with the development of sexual reproduction (2). Sexual dimorphism occurs not only in differences in external appearances, but also in internal organs and biological functions. There are thought to be sex differences in all cells of the body (3). In mice, it has been reported that sex has an impact on ~57% of quantitative traits (i.e. continuous measurements such as body weight) and on 10% of qualitative traits (i.e. categorical measurements such as normal head shape) (4). Aside from sex differences due to effects of genes on the sex chromosomes, many of the physiological effects of sex differences are due to sex hormones (5, 6).
Many autoimmune diseases show sexual dimorphism in incidence, prevalence, and features of disease (7). One such autoimmune disease is multiple sclerosis (MS), an inflammatory and demyelinating disorder of the central nervous system (CNS). More than 80% of people initially develop a relapsing-remitting form of MS (RR-MS), whereas the remainder develop a form of disease that is progressive from the outset (primary progressive (PP)-MS). RR-MS is approximately three times more prevalent in females than males, whereas the ratio of females to males with PP-MS is closer to 1:1 (8, 9). There is also a marked effect of pregnancy on the course of MS (10, 11). There are a multitude of potential mechanisms underlying these sex differences in humans, many of which currently remain unexplored.
It has also been reported that biological sex and pregnancy can modulate the outcomes of experimental autoimmune encephalomyelitis (EAE), an animal model widely used in research on MS. In this review, we focus on how biological sex and pregnancy influence the incidence, severity, and clinical course of EAE. We conclude that there is much variability in these effects between different EAE models, suggesting that many intrinsic and extrinsic modifiers make this a much more nuanced topic than sometimes portrayed. Nevertheless, understanding how these effects occur in the animal model could point to strategies that might modify the course of MS.
EAE developed from investigations into the cause of paralysis following anti-rabies vaccination, which used vaccines produced from desiccated spinal cords of rabies-infected rabbits. The first description of EAE in the English language literature appeared in 1933 by Rivers, Sprunt and Berry (12). The original forms of EAE were induced by multiple inoculations with emulsions of CNS tissue. Modern forms of EAE came about with the introduction of adjuvants (13). EAE can be actively induced in a range of animal species by injection of CNS antigens emulsified in adjuvant or by adoptive transfer of T cells reactive to such antigens (14–17). EAE is now most commonly induced with peptides of myelin oligodendrocyte glycoprotein (MOG), myelin basic protein (MBP) or myelin proteolipid protein (PLP), although a wide range of CNS antigens, including homogenates of brain and spinal cord, can be used (18).
Soon after its first description, some of the pathological changes in the CNS in EAE were noted to be similar to those of MS (13). However, EAE differs from MS in needing to be induced, rather than occurring spontaneously (with the exception of a small number of transgenic mouse models in which the majority of the T cells and/or B cells in the mice have receptors specific for myelin antigens (19, 20)). There are also marked differences in clinical and some of the pathological features of disease, compared to MS (21), although, similar to MS, the pathological features of EAE always include some degree of neuroinflammation and usually some demyelination and some degree of axonal injury and loss (22–25).
Nowadays, EAE is most frequently induced in mice and rats, although it can also be induced in non-human primates (26–31), guineas pigs (32–34), rabbits (35, 36), hamsters (37), dogs (38) sheep (39, 40) or opossums (41). Within species, animal strains differ in their susceptibility to EAE and the clinical and pathological features of disease (28, 31, 34, 36, 39). It should also be noted that many factors in addition to the animal strain and encephalitogen can affect the development of EAE, including the source of the animals and the sterility of the environment in which they are housed (which affects the gut microbiome), the age of the animals, the adjuvant(s) in which the encephalitogen is emulsified and the way in which emulsions are prepared and injected, and whether or not pertussis toxin is also given to the animals (commonly used in mice, but rarely in other species). Several detailed standard protocols for induction of actively- or passively-induced EAE have been published (42–45), but there are many variations on a theme in the literature.
Interpreting whether or not the sex of an animal affects the incidence of disease or the severity of EAE can be complicated, as these parameters also vary with the species, strain and age of animal and the antigens and adjuvants that are used to induce disease (46–53). In addition, in many EAE studies, animals of only one sex have been tested (most often females, which are often favored as they tend to be less aggressive to each other when housed in groups and also because of the higher prevalence of MS in females), or else the sex of the animals is sometimes not reported in papers.
In older studies using outbred animals (e.g. non-human primates and guinea pigs), where both sexes have been tested, males and females generally appear to be equally susceptible to development of EAE (54–57). Only some rabbit strains are able to develop EAE, but, in those susceptible strains, the incidence of disease is reported to be similar in males and females (36).
In inbred rat and mouse strains, however, there are many differences reported between males and females (Table 1 – details of variables such as strain, age, antigen etc are given when this information is available). For example, adult (8-10 week old) Lewis rats of both sexes can develop EAE, but in males disease is generally monophasic, whereas females of the same age often develop relapses of disease (58–60); however, juvenile (6-week-old) male Lewis rats develop more severe relapsing EAE than juvenile females. In contrast, in adult DA rats, both males and females develop relapsing EAE with similar clinical severity; however, by histology, males show more severe inflammation, whereas females have more astrocyte activation (47). To round out the spectrum of responses, in the PVG strain of rats, males are much more susceptible than females to development of EAE and disease severity is greater in the males (63).

Table 1 Effects of biological sex on incidence and severity of actively-induced EAE in rodents.
Similarly, in mice, there are strain-specific relationships in the effects of sex on EAE, but these can be quite complex (Table 1). For example, in the SJL mouse strain with EAE induced with a variety of antigens (CNS tissue homogenates, PLP or MBP), females have generally been found to be significantly more susceptible to EAE than males (>80% vs <30%, respectively), have an earlier time of onset of disease, and have higher clinical scores compared to males (49, 52, 67). However, this female predominance in SJL mice is not seen when EAE is induced with MOG92-106 peptide (49). In addition, the observed sex differences can be age dependent: 6-8 week adult SJL males are resistant to induction of PLP-induced EAE, however, the risk for development of EAE in males increases by 4% for each increasing week of age; in contrast, disease severity does not change in females with ageing (51, 68, 69).
It does not appear that the major histocompatibility complex (called the H-2 complex in mice) allele carried by the mice has a major role in the effects of biological sex on development of EAE, as other strains of mice carrying the H-2s allele (same as SJL) either show no significant differences in disease incidence, clinical scores, or disease course between males and females (e.g. A.SW strain), or else both sexes are resistant to development of EAE (e.g. B10.S strain). The only other mouse strains in which females are reported to be significantly more susceptible to actively-induced EAE compared to males are (B10.S x SJL)F1 mice (H-2s) and the NZW/LAC strain (H-2d), although, in the latter strain, males that show clinical signs have more severe disease than females (49). In mouse strains such as C57BL/6 (H-2b), NOD (H-2g7), and PL/J and B10.PL (both H-2u), the incidence of EAE in male and female mice does not differ significantly between the sexes (48–50, 70), but male PL/J and B10.PL mice show increased EAE-related mortality compared to females (49). Interestingly, a recent report suggests that a female bias in disease severity becomes more apparent in C57BL/6 mice when lower amounts of pertussis toxin are used during the induction phase of EAE, suggesting that a sex difference in the response to pertussis toxin could confound other underlying sexual dimorphism in this strain (53).
Several “spontaneous” EAE models have also been developed, in which there is transgenic expression of immunoglobulin heavy (IgH) chains and/or T cell receptors (TCR) specific for peptides of MOG on either C57BL/6, NOD, or SJL backgrounds (20, 71–73). Some of these models exhibit sexual dimorphism (Table 2). Double transgenic (i.e. IgH X TCR) mice on a C57BL/6 background show no sex differences (72), however all the other models show a higher incidence of spontaneous disease in females. In spontaneous models on an SJL background, female mice are more likely to develop a relapsing-remitting type of disease, whereas males mostly develop progressive disease from the outset (20, 73).

Table 2 Effects of biological sex on incidence and severity of spontaneous EAE mouse models.
Adoptive transfer studies have been done to try to determine if the sex of the donor of the encephalitogenic T cells affects the susceptibility to development of EAE, or if is the sex of the recipient that determines the outcome. These studies have yielded conflicting results (Table 3). A confounder in the interpretation of these studies is that each has used a different mouse strain and/or antigen. The different studies variously suggest that :i) the sex of the recipient determines the differences (52: ii) that disease severity is related to the sex of the donor cells, with female cells giving greater disease, and suggesting that the sex differences play a greater role during the induction phase of the disease (74); or iii) that cells from male donors induce more severe disease, irrespective of whether they are transferred into male or female recipients (75). In vitro conditions under which the cells to be transferred are prepared may also affect the outcome, as differences in findings from adoptive transfer studies do not always reflect the sex bias seen in actively induced EAE (66).

Table 3 Effects of biological sex in adoptive transfer EAE.
Overall, we can conclude that some mouse strains show a female bias in disease, but this may either not occur in other strains, or be masked by sex-related effects of the antigens or adjuvants used. In most studies, females appear more likely to develop a relapsing-remitting disease course, whereas males are more likely to develop a chronic progressive disease course with greater inflammation than seen in females, which is somewhat reminiscent of what occurs in humans with MS.
A summary of some of the cells and molecular pathways involved in sex differences in the pathophysiology of EAE are given in Tables 4 and 5 and are discussed in more detail below.
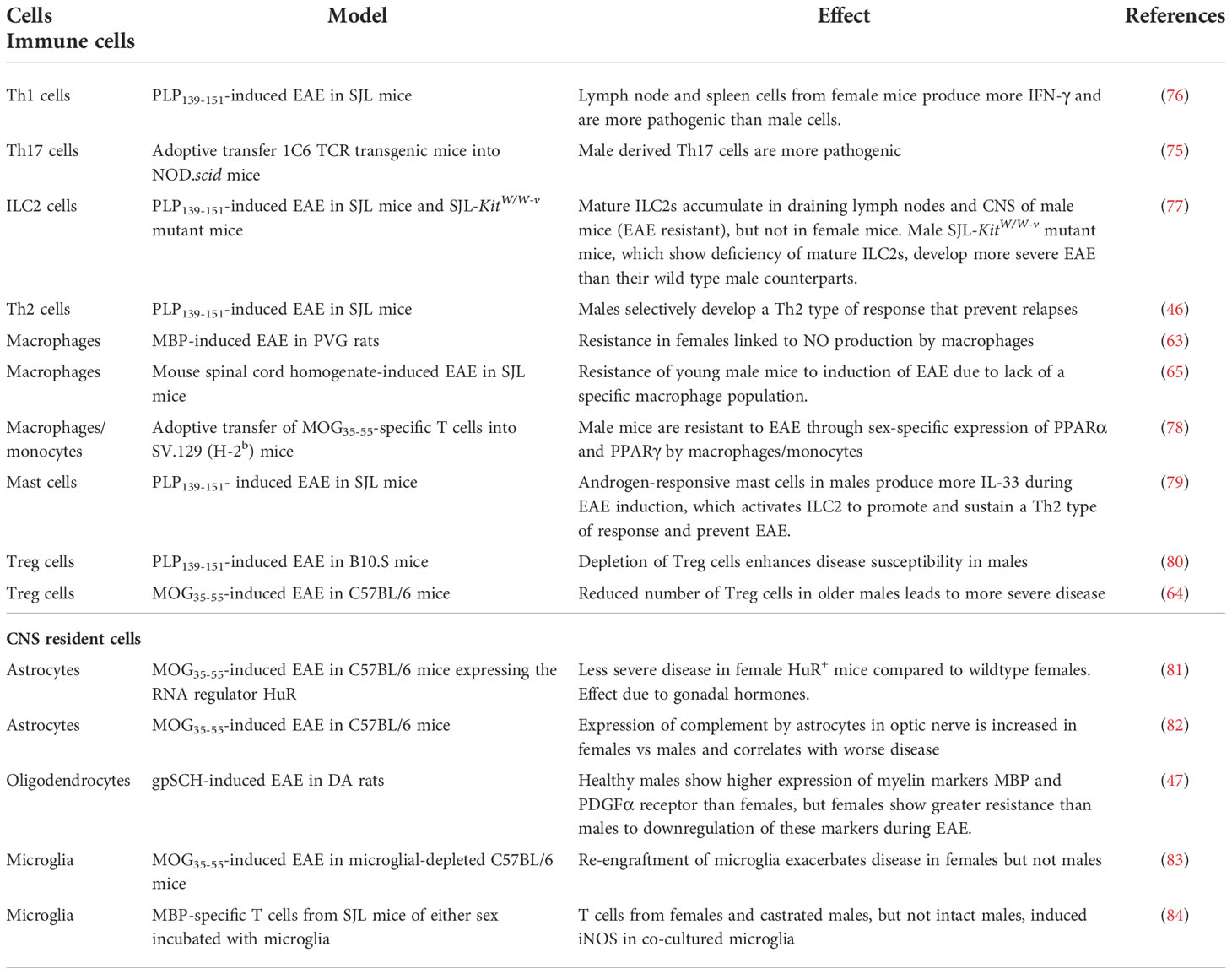
Table 4 Cells that are implicated in sex differences in EAE.
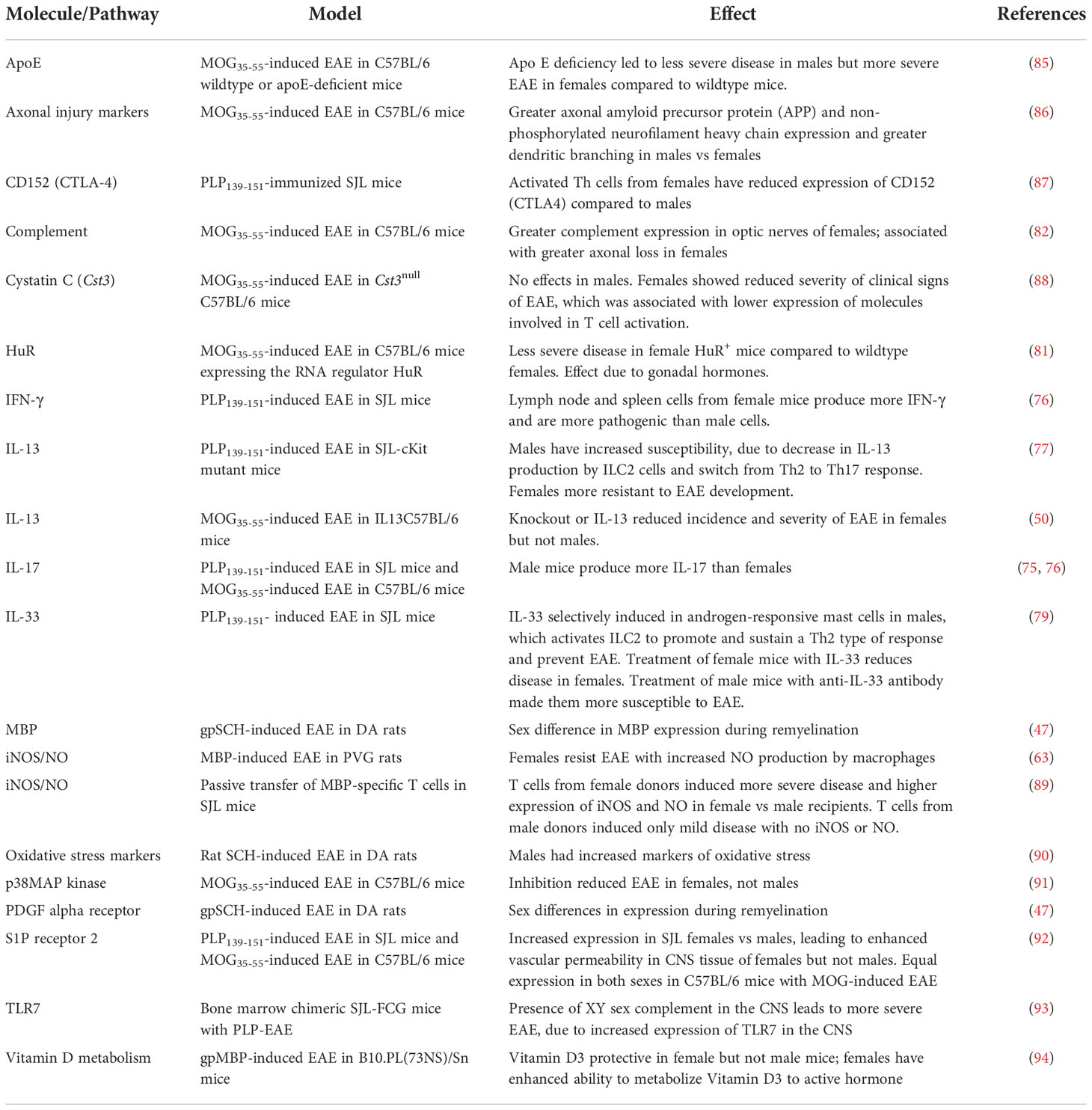
Table 5 Molecules and pathways implicated in sex differences in EAE.
When considering the effects of biological sex on the outcome of EAE, it is important to remember that the sex of an animal impacts the makeup of the immune cells and organs under physiologically normal conditions, as well as when they are pathologically challenged. This is true of humans, as well as experimental animals, where (for example) CD4+ cells from female healthy volunteers show a much higher levels of production of IFNγ following T cell activation with anti-CD3/anti-CD28 than do cells from males, whereas IL-17 production is higher in cells from males (76). In humans, elevated Th17 responses have been linked to male sex in some autoimmune diseases (95).
In mice, biological sex affects the numbers and activity of Th1, Th17 and Th2 T cells, regulatory T cells (Treg), and antibody levels (96, 97). However, these differences are not necessarily seen to an equal degree in all immune organs in an individual (e.g. there can be variation between the sex-related differences seen in the spleen and those seen in other secondary lymphoid organs) and they can occur in a mouse strain-specific manner (96).
Effects of biological sex on adaptive immune responses in EAE are summarized in Table 4. Neuroantigen-specific CD4+ Th1 and Th17 cells are the main cells responsible for the induction of EAE. Although many papers state that Th17 cells are critical for disease development, there are quite a number showing that, at least in both the models of MOG-induced EAE in C57BL/6 mice and PLP-induced EAE in SJL mice, Th1 cells, Th17 cells, and cells producing both IFNγ and IL-17 appear to be equally able to induce EAE (76, 98–101). However, there do appear to be differences in the site of induction of antigen-specific T cells in male and female mice. For example, in PLP-induced EAE in SJL mice, lymph nodes cells from female mice produce copious amounts of both IFN-γ and IL-17 in response to a secondary challenge with the autoantigen, whereas lymph node cells from male mice produce neither of these. In contrast, spleen cells from the same mice produce predominantly IFN-γ if they are from females, but IL-17 if they are from males (76). These observations have been interpreted as providing an explanation for the ability (or not) of male mice to develop EAE, based on how much IL-17 they can produce.
However, the interpretation of these results may be more complicated. A recent study found (using adoptive transfer of MOG-specific transgenic Th17 cells from males and females into sex-matched NOD.scid recipients) that male Th17 cells are more encephalitogenic than female Th17 cells, but only if they rapidly switch (once they are transferred into the recipient) from producing IL-17 to producing large amounts of IFN-γ- cells from males that produce some IFN-γ, but also retain IL-17 production, as is seen in females, are less encephalitogenic (75). Relative levels of IFN-γ and IL-17 therefore appear to play different roles in the pathogenesis of EAE in males vs females, but much work remains to determine the exact relationship between these measures and the susceptibility of different strains of mice to EAE and how (or whether) this relates to humans with MS, in whom similar mixtures of myelin antigen-specific Th1, Th17 and mixed phenotype cells have also been reported (102).
There are several molecules that are of potential interest in these effects, including PPARγ, one of the central regulatory molecules in determining T cell fate, which selectively inhibits Th17 cell differentiation only in male cells and which modulates Th1, Th2 and Th17 differentiation in female T cells, based on the levels of estrogen (103), and Jarid1c, an X-linked immune regulator that represses severity of Th17-mediated EAE in males (75).
There also appear to be sex-based differences in the action of regulatory types of T cells in EAE. In PLP-induced EAE in SJL mice, male mice develop IL-10 producing cells that prevent relapses of disease (46) and this appears to be due to direct effects of testosterone on CD4+ T cells (104), whereas females have increased T cell proliferation and reduced expression of the inhibitory molecule CD152 (CTLA-4) compared to males (87). In B10.S mice, which are normally resistant to PLP-induced EAE, male mice show greater susceptibility to disease after depletion of Treg cells, but depletion of Treg cells has no effect on susceptibility of female mice (80). In MOG-induced EAE in C57BL/6 mice, where there are no sex differences in incidence of disease, there appear to be underlying sex differences in pathophysiology, as male mice have both increased inflammatory processes and increased regulatory processes, which cancel out, so that disease is similar to that in females (97). Another study using the same model found that young male mice develop less severe EAE than middle aged male mice, due to reduced numbers of CD4+CD25+ Treg cells in the middle-aged vs the younger males (64).
As with cells of the adaptive immune response, there are differences between numbers and location of dendritic cells (DC), macrophages, natural killer (NK) cells and other innate cells in male and female mice (96, 97). In actively-induced EAE studies, an important consideration when looking at innate immune responses is the effects of the adjuvants (typically complete Freund’s adjuvant and pertussis toxin) used in the induction of disease, as a recent study (97) found that there were sex differences in the induction of key innate inflammatory factors by these adjuvants alone.
Several sex-related differences in macrophages have been reported in EAE (Table 4). Resistance of male SJL mice to induction of EAE is reportedly dependent on a subpopulation of macrophages (Mac-1+, Mac-2-, Mac-3+) that play a regulatory role in the induction of encephalitogenic T cells (65). Additionally, several rodent models have associated higher levels of inducible nitric oxide synthase (iNOS) and nitric oxide (NO) production by macrophages with sexual dimorphism in EAE; however, in PVG rats, this is linked to resistance to EAE in females (63), whereas in SJL mice it is linked to increased severity of EAE in females (89). Other molecules produced or expressed by macrophages or monocytes that have been implicated in the sexual dimorphism seen in EAE include PPARα and PPARγ (78), and several Th2 cytokines, including IL-33. In addition, apolipoprotein E (apoE), which is produced by macrophages and DCs, has been reported to enhance anti-inflammatory macrophage phenotypes and downregulate Th1 and Th17 responses (105, 106). It is therefore not surprising to find that apoE deficiency in C57BL/6 mice with MOG-induced EAE leads to more severe EAE in females; however, it is surprising to find less severe EAE in apoE-deficient males (85). The mechanisms underlying these varying effects of apoE deficiency in males and females remain to be elucidated. Finally, inhibition of p38 MAPK activity in C57BL/6 mice with MOG-induced EAE ameliorates EAE in female but not male mice, and this is due specifically to affects in myeloid cells of the females, which have a greater dependence on p38 signaling for expression of proinflammatory genes than do male myeloid cells (91).
Other IL-13- and IL-33-expressing cells, including mast cells and innate lymphoid cell type 2 (ILC2) cells, are also of relevance for sex-related differences in EAE. In PLP-induced EAE in SJL mice, androgen receptor-expressing mast cells from males produce higher levels of IL-33 in response to testosterone and other inflammatory activators (79). IL-33 then activates ILC2 cells to promote and sustain a Th2 type of response (via production of IL-13) and prevent EAE. Treatment of male mice with anti-IL-33 antibody makes them more susceptible to EAE, whereas treatment of females with IL-33 leads to a reduction in EAE severity (79). Conversely, decreased accumulation of IL-13-secreting ILC2 cells in the brains of male, but not female, SJL–c-Kit mutant mice leads to increased susceptibility to development of EAE in the male mice, but resistance to the development of EAE in females (77), via a switch from a Th2- to a Th17-dominated T cell response that is regulated by the presence or absence of mature ILC2 cells in the draining lymph nodes and the CNS. In C57BL/6 mice, knockout of IL-13 reduces the incidence and severity of MOG-induced EAE in females but not in males (50).
Within the CNS, microglia and astrocytes are the main cell types thought to be involved in innate responses. They differ regionally and numerically between male and female mice (107–111) and there is sexual dimorphism in their responses to CNS assault in adult mice (107, 112). For microglia, these can include differences in inflammatory sensitivity and reactivity, their ability to affect cellular repair, and in their response to oxidative stress (113, 114). Studies in which microglia have been depleted from brains of mice using the CSF1R inhibitor, PLX5622, result in adverse sex-specific behavioral effects, with females developing long-term hyperactivity and anxiolytic-like behaviour (115). Astrocytes display sex differences in uptake of glutamate and their responses to fatty acids, and are known to express the estrogen receptor alpha (ER-α) (116–118). There are also sex differences in mitochondrial bioenergetics of astrocytes, but not microglia (119).
In EAE studies, some sex-specific effects on or by these CNS cell types have also been reported (Table 4). In microglia, for example, iNOS can be induced by MBP-primed T cells from female and castrated male SJL mice, but not from intact males, and this is dependent on activation of a CCAAT-enhancer-binding protein (C/EBPβ) in the microglia (84). In MOG-induced EAE in microglial-depleted C57BL/6 mice, re-engraftment of microglia exacerbates disease in females but not males (83). Astrocytes are also involved in the pathology of EAE (120) and sex differences in the astrocyte response during EAE have been described. For example, there is expression of complement by astrocytes in optic nerves of C57BL/6 mice with MOG-induced EAE, and this is greater in females than males and correlates with worse retinal ganglion cell and axonal loss in females (82). Similarly, transgenic expression in astrocytes of C57BL/6 mice of the RNA regulator HuR, which plays a major role in regulating production of cytokine and growth factors, results in attenuation of MOG-induced EAE in females, but not males (81). Furthermore, this effect can be reversed by ovariectomy.
One molecule that is widely expressed in both peripheral and CNS innate immune cells, and which has previously been implicated in both MS and EAE, is cystatin C. However, cystatin C appears to have different functions in immune and CNS cells, and there has been much debate on whether it plays a protective or pathogenic role in MS and EAE. In MOG-induced EAE in C57BL/6 mice, deficiency of Cst3, the gene encoding cystatin C, leads to improvement of EAE in females, but has no effect in males. Furthermore, the sex-dependent effects of cystatin C are negated by ovariectomy of females, and are revealed by treatment of males with female hormones (88).
Immunometabolism, the metabolic reprogramming of anaerobic glycolysis, oxidative phosphorylation and metabolite synthesis that occurs when immune cells are activated, is an important regulator of immune responses and immune homeostasis. Immunometabolic dysfunction has been linked to an inflammatory phenotype of immune cells and has been implicated in development of autoimmune diseases (121), including EAE (122). There is an interaction of immunometabolism with sex, especially through the actions of reproductive hormones (123). Male DA rats with EAE have more severe disease and greater oxidative stress than females (90), consistent with studies that show sex differences in levels of pro-oxidant and anti-oxidant enzymes (124, 125). It has also been found that high sodium (126), high fat (127), or high carbohydrate (128) diets can all exacerbate EAE. In SJL mice fed a high sodium diet from prior to induction of PLP-induced EAE, there is an increase in disease severity in female mice, whereas male mice are not affected (129). In contrast, compared to mice fed a normal diet, both male and female C57BL/6 mice with MOG-induced EAE and fed a high sodium diet show some increase in disease severity, particularly during the peak of the acute disease (129). In vitro studies have suggested that the effects of high levels of salt appear to be due to stabilization of the Th17 cell phenotype, via the activation of several pathways, including p38 MAPK, nuclear factor of activated T cells 5 (NFAT5), and the salt-sensing serum glucocorticoid kinase 1 (SGK1) (126, 130). However, a high salt diet has also been reported to elevate serum sodium, leading to increased blood-brain barrier permeability and more severe brain pathology (129). Additionally, it cannot be ruled out of these studies that different effects of high salt diets in mice of different sexes may relate to changes to the gut microbiome. The gut microbiome appears to play an important role in both EAE and MS (131). Sex hormones influence the composition of the gut microbiota (132) and its interaction with the brain (“the gut-brain axis”) (133). In MOG-induced EAE in C57BL/6 mice, protection due to estrogen is associated with changes in the gut microbiota and the gut lymphoid tissue (134). In the same model, oral administration of 2.6% alcohol for 3 weeks prior to EAE induction is protective in both sexes, but the protective effect is greater in males, and also appears to be mediated though changes in the gut microbiota (135).
Several therapeutic interventions also show sex-specific effects. Supplementation with Vitamin D3 protects against EAE in female but not in male mice of several strains (94, 136), and this effect appears to be due to sex differences in vitamin D3 metabolism, with females having an enhanced ability to metabolize vitamin D3 to the active hormone 1,25-(OH)2D3 (94). Ovariectomy abrogates this protective effect (94). Other therapeutic approaches show more of a male-specific effect, e.g. oral tolerance can be induced in male, but not female, B10.PL mice fed with an altered peptide ligand of MBP (137).
In several autoimmune diseases it is recognized that the severity of disease can be influenced by the resistance of the target tissue to damage (138, 139). In EAE and MS, target organ resistance could involve variability in the processes that lead to damage to the myelin-producing oligodendrocytes and neurons, and in the processes of repair. Since myelin is the target of immune attack in EAE and MS, several studies have investigated the effects of biological sex on oligodendrocytes. These studies have shown that there is sexual dimorphism in oligodendrocytes’ capacity for myelination and their responses to stress (140, 141). There are reports of sex differences in expression of mRNA for MBP and PDGF alpha receptor (markers of myelination) in DA rats with EAE (47), and markers of axonal injury and dendritic aborisation are greater in male than female C57BL/6 mice with MOG-induced EAE (86). However, it is not clear at present if these findings merely reflect the strength of the immune response in the animals of different sexes in these models or if they truly reflect sex differences in target organ resistance in EAE. Adoptive transfer studies of male and female cells (not only of peripheral immune cells into naïve recipients, but also of microglia into microglia-depleted recipients) could help to answer this question. This is a topic that will likely be difficult to decipher, but one that may prove very useful in ultimately answering some important questions related to the differential prognosis of males and females with MS.
Other molecules that are expressed in the CNS can also influence sex differences in other pathophysiological mechanisms in EAE. For example, sphingosine-1-phosphate receptor 2 (S1PR2) is a molecule involved in regulation of the integrity of the blood-brain barrier (BBB). Naïve female SJL mice have significantly elevated levels of S1PR2 compared to SJL males and to C57BL/6 mice of both sexes (92). Induction of PLP-induced EAE in the SJL mice further increases expression of S1PR2 and causes increased vascular permeability in CNS tissue from females, but not males. Furthermore, S1PR2-deficient mice showed decreased EAE severity compared with wild-type controls (92). Interestingly, however, adoptive transfer of CD4+ cells from TCR1640 transgenic mice (MOG92-106-specific cells on SJL background) into naïve male or female SJL recipients show no sex differences in BBB integrity (73).
It may even be that the sex of the experimenter can affect outcomes. Whilst this has not been systematically investigated in EAE, as far as we are aware, there are several reports that rodents of both sexes are more stressed when the person doing the experiments on them is male, and that this can lead to differences in behavioral assays of pain and stress (142, 143). This could potentially add another layer of complexity to investigation of the effects of biological sex on the pathophysiology of EAE.
Overall, biological sex can affect many of the elements of immune function and CNS biology that are required for development of EAE. Sex differences could occur during the induction of disease, where there could be sex differences in response to adjuvant and in antigen presentation or in the effector stage of disease when there could be sex differences in the pathological processes in the CNS. To clarify this, appropriate controls (naïve and adjuvant only) of both sexes need to be included in all experiments looking at mechanisms underlying sex differences in EAE. Much more work remains to understand the basic biological differences between the sexes, both in mice and humans.
The sex chromosomes, X and Y, are the only chromosomes in the body that are represented differently in females and males, and they vary greatly (144). Thus, effects of sex could be due differences in the sex chromosomes and/or sex-related differences in gene expression.
The Y chromosome contains only 50 to 60 unique protein-coding genes that encode primarily for molecules required for gonadal formation and fertility, including the Sry gene, which is necessary for the development of male gonads (145) and the Azf (azoospermia factor) locus that has a role in organs outside the testis, including the brain (146), and in inflammation (147, 148). Variable amounts of gene replication occur on the Y chromosome from different species, so that the Y chromosomes from humans and mice are quite different (149, 150). Even within mouse strains differences can be observed: e.g. some strains have a Y chromosome gene known as Yaa (y accelerated autoimmunity) which has resulted from a translocation of the gene encoding Toll-like receptor 7 (Tlr7) from the X chromosome onto the Y chromosome. Yaa can modulate a mouse’s potential to develop autoimmunity via epistatic interactions with other autoimmunity-related genes (151) (152).
In contrast to the Y chromosome, the X chromosome contains over 900 genes, and is highly conserved across species. Genes on the human X chromosome include genes for reproduction and a disproportionately large number that code for brain function, such as the BEX family (brain expressed X-linked genes), NF2 (neurofibromin 2), GDNF (glial cell derived neurotrophic factor) and PLP1 (myelin proteolipid protein, the most abundant protein in CNS myelin, an autoantigen for EAE, and a target of immune attack in MS (153, 154)). The X chromosome also contains genes for the testis that are subject to selection pressure because they confer advantage in males (155). There are also many genes of relevance to immune function and susceptibility for autoimmunity located on the X chromosome, including those encoding IL2-Rγ, IL13-Rα, CXCR3, CD40 ligand, TLR7 and TLR8, TIMP1 and Bruton’s tyrosine kinase (BTK).
In males, the X chromosome is always of maternal origin. In females, one X chromosome is randomly inactivated in each cell of an early-stage embryo, so that the embryo is a mosaic of cells with either an active paternal X chromosome or an active maternal X chromosome. All descendants of that cell will have the same active X chromosome, and most females have an approximately even spread of paternally- and maternally-derived active X chromosomes (156). However, this inactivation is not always complete and can differ in different tissues and cell types from a single individual (157), potentially leading to X dosage effects, with higher expression of X-linked genes in females compared to males. DNA methylation plays an important role in X chromosome inactivation and recent studies have reported that maternally derived X chromosomes have less DNA methylation than paternally-derived X chromosomes, and that therefore there is greater expression of X-linked genes in male cells (158). These factors could have implication for sex differences in autoimmunity.
The development of a mouse model known as the “4 core genotypes” (FCG) model, in which the sex chromosome complement is unrelated to the animal’s gonadal sex (159), has allowed studies of the effects of the sex chromosome complement on susceptibility to EAE. Early studies compared gonadally-intact FCG mice on a SJL background, (SJL-FCG mice) with FCG mice in which the gonads were removed. These studies show that the presence of XY (male) complement of sex chromosomes leads to greater immune responses to MBP than XX chromosomes, whereas the presence of male sex gonads (and therefore sex hormones) inhibits immune responses to MBP (160). In that same model, when gonads are removed, mice carrying the XX genotype are more susceptible to PLP-induced EAE than are mice carrying a XY genotype, thus demonstrating an autoimmune-enhancing effect of the XX genotype independent from hormonal influences (161). Whether this outcome could be due to dosage effects of the target antigen, PLP, which is encoded on the X chromosome, was not investigated. Bone marrow chimeric SJL-FCG mice, in which the mice have either a XX or XY sex complement in the CNS and an immune system with a common sex chromosomal type, demonstrate that the presence of an XY sex complement in the CNS associates with greater clinical and neuropathological severity of EAE, apparently due to higher expression of TLR7 in the CNS of XY vs XX mice (93). FCG mice on a C57BL/6 background and immunized with MOG show parent of origin effects of DNA methylation of a set of five X chromosome genes in autoantigen-stimulated T cells and B cells, with higher expression in XY than XX genotypes. SJL-FCG mice immunized with PLP also show similar effects, but the X-linked genes affected differ from those seen in the C57BL/6-FCG mice, suggesting that parent of origin effects are not strain specific (158).
The Y chromosome also has an effect on immune cell gene transcription (162) and copy number variation in Y chromosome genes plays a role in paternal transmission of autoimmune disease (163). There is an effect of Y chromosome genes on the incidence and severity of EAE, since studies using progeny of C57BL/6 Y-chromosome substitution strains show that the Y chromosome influences the severity of EAE in both male and female mice (164).
Given the differences between the ability of male SJL and C57BL/6 mice to develop EAE, it is of also of interest to note that the Y chromosome in SJL mice has been inherited from the Mus musculus domesticus subspecies, whereas the Y chromosome in C57BL/6 and most other commonly used inbred laboratory mouse strains comes from Mus musculus musculus (165). Y chromosome substitution (Y consomic) SJL male mice carrying a Mus musculus musculus Y chromosome are more susceptible to EAE than are age-matched SJL males, suggesting that a Y chromosome-linked polymorphism controls the sexual dimorphism to development of EAE seen in SJL mice (68).
It is of interest to note that, as well as genes on the sex chromosomes, there are other sex-specific genetic differences. A study of crosses between EAE-susceptible SJL mice and EAE-resistant B10.S mice identified sex-specific loci that governed susceptibility of SJL mice to relapsing EAE (eae12) and monophasic EAE (eae7 and eae13) (69). In a large study of 26 chromosome substitution (consomic) strains, in which mice inherited chromosomes from the EAE-resistant wild-derived PWD mouse strain on a C57BL/6 background, 19 genetic loci on 18 chromosomes were identified that influenced susceptibility to MOG-induced EAE: of these, six were male-specific and four were female specific (166).
Males and females also differ in the levels of circulating sex hormones, which change over the lifetime. In females this is especially true during pregnancy, when there are increased levels of many different hormones and also increased levels of mediators related to pregnancy (see section 6 for more on pregnancy-related effects). The effects of physiological levels of the major sex hormones could explain some of the observed effects of biological sex in EAE, as shown through comparisons of intact and gonadectomized mice (167, 168). The effects of sex hormones on EAE could be due to immune modulation and/or neuroprotection, as summarized in Table 6. In addition, it appears that there could be bi-directional effects of sex hormones and the pro-inflammatory molecules produced during EAE (178), as SJL mice with PLP-induced EAE have a significantly shorter estrous cycle (normally around 4-6 days (179)) than mice just immunized with adjuvant. The effects of the major sex hormones are summarized below.

Table 6 Summary of immune and neuroprotective effects of sex and pregnancy hormones.
Estrogens comprise estrone, estradiol (E2) and estriol (E3). Females have higher levels of estrogens than males, and in females the levels fluctuate with the menstrual cycle and decline after menopause. In pregnancy there are high levels of E2 and estrone in early pregnancy and high levels of E3 in the later stages of pregnancy.
Low doses of estrogen are reported to enhance pro-inflammatory responses, whereas high doses appear to inhibit such responses (180). In humans, higher doses of estrogens can inhibit the T cell-stimulatory capacity of DCs (181), modulate the effects of lipopolysaccharide (LPS) on monocytes (182), and inhibit monocyte adhesion to endothelium (183). Estrogens also influence production of cytokines (including IL-1, IL-6, TNF-α, M-CSF, GM-CSF, and TGF-β) and chemokines and affect T cell trafficking, probably via modulation of chemokine receptor expression (184). Furthermore, estrogen can convert CD25− cells to CD25+ Treg cells (185) (in humans, pregnancy is accompanied by increased levels of CD25+ Treg cells (186)). Thus, enhancement of Th2 responses and induction of Treg cells by high doses of estrogens could ameliorate Th1/Th17-mediated autoimmune diseases and could be one of the mechanisms underlying the beneficial effects of pregnancy on MS. Estrogens, particularly E2, also alter nerve conduction, and there is in vitro evidence that estrogen can modulate potassium channels, with effects of nerve conduction (187), and be neuroprotective (168).
Ovariectomy of females has been reported to have variable effects on the outcome of EAE. Ovariectomy of female B10.RIII mice with MBP-induced EAE or of C57BL/6 mice with MOG-induced EAE causes worse disease than is seen in intact female mice, showing that even physiological non-pregnant levels of female sex hormones confer some protection (188) (189). In contrast, in SJL and (B10.S x SJL)F1 mice with PLP-induced EAE, ovariectomy of females does not have a major impact on disease development (76, 190), or even has a slightly protective effect (67). Some of these studies show a role for ERα, one of the three estrogen receptors, which is widely expressed in the immune system and in the nervous system (191, 192), in these effects.
As listed in Table 7, there are many studies showing that administration of exogenous estrogen, particularly E2 and E3, suppresses EAE in a variety of models, most likely via ERα (encoded by esr1 in mice) signaling (206). A variety of mechanisms have been reported to underlie the beneficial effects of therapeutic administration of estrogen in different EAE models. These include: i) reduced secretion of matrix metalloproteinase 9 (MMP9) (169), a molecule produced primarily by macrophages and astrocytes in EAE and involved in entry of inflammatory cells into the CNS; ii) enhanced numbers of regulatory immune cells (204, 205), particularly regulatory B cells and microglia (203); iii) induction of a novel population of VLA-4+ cells, without lymphocyte markers, that suppress EAE (207); iv) prevention of the changes to the gut microbiota that normally occur with EAE (134); v) reduced astrocyte production of the pro-inflammatory chemokine CCL2 (201); and vi) reduced DC antigen presentation and activation (196). Estrogen is also synergistic with vitamin D3 in protection from EAE in females, although not in males (200).
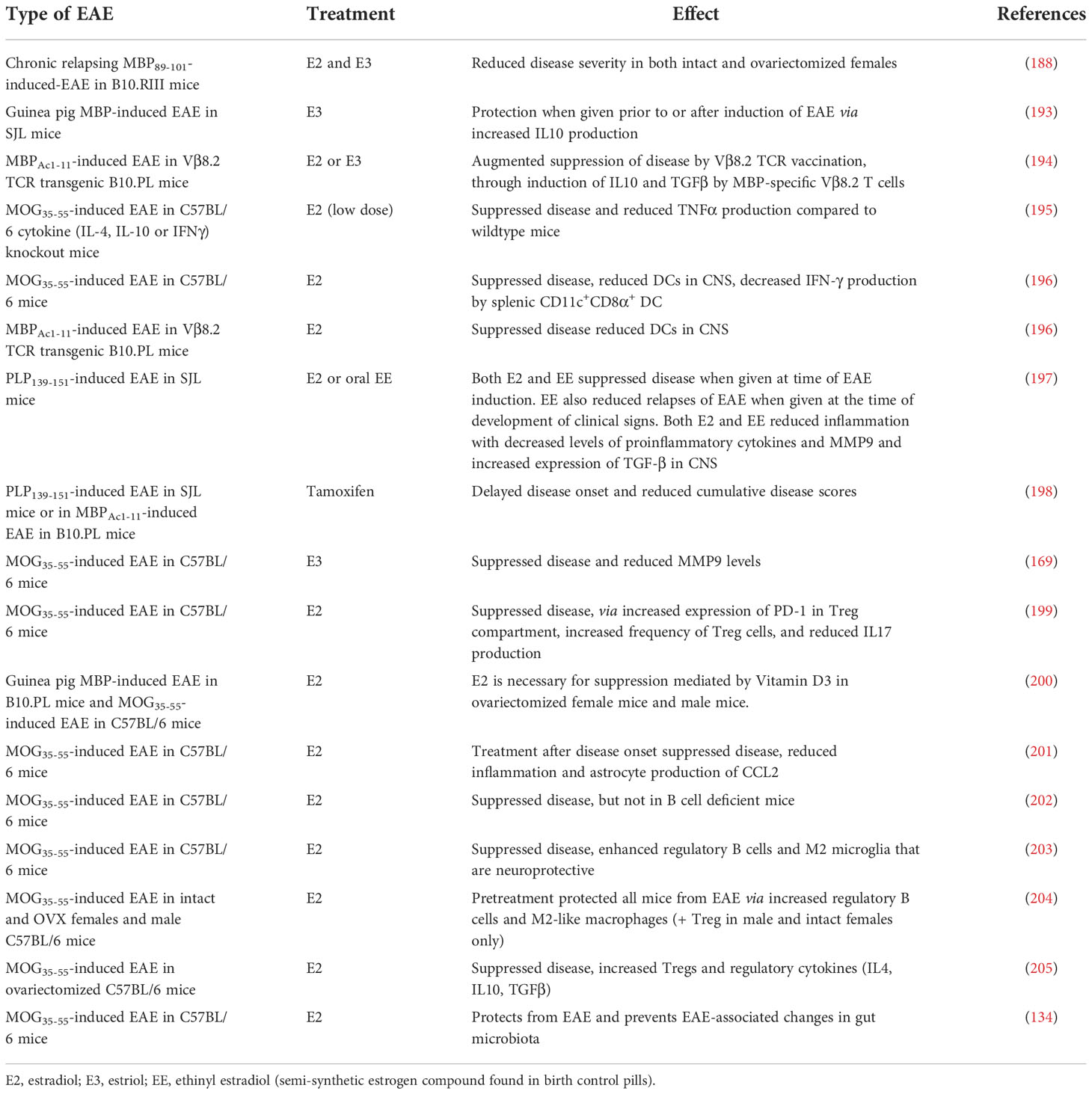
Table 7 Estrogen and EAE.
Although there is a significant amount of evidence that exogenous estrogen can inhibit EAE, clinical application of estrogens in human patients has been slow, due to undesirable side effects of treatment mediated via the intracellular ERα. There have therefore been studies of drugs that target other elements of the estrogen pathway, including an agonist, G-1, that selectively activates the putative membrane estrogen receptor, GPR30, without engagement of ERα. G-1 protects C57BL/6 mice against development of MOG-induced EAE via enhancement of CD4+Foxp3+ regulatory T cells (208). Other studies have used selective estrogen receptor modulators such as tamoxifen and raloxifene, which behave as estrogen agonists in some tissues but are either inert or behave like estrogen antagonists in other tissues. These drugs inhibit PLP-induced EAE in SJL mice via reduction of antigen presentation by DC and induction of a Th2 bias in myelin-specific T cells (198). They also ameliorate MOG-induced EAE in female C57BL/6 mice by inhibiting NF-κB signaling and expression of CCL20 in reactive astrocytes (209), similar to the effects of E2 (201).
Progesterone is present in higher levels in females than males. It is also present in high levels in pregnancy and could contribute to the effects of pregnancy on EAE. Progesterone has immunomodulatory effects that are generally anti-inflammatory, including inhibition of glucocorticoid-mediated thymocyte apoptosis, and reduction of nitric oxide production and expression of toll-like receptors (TLR) by macrophages (210). Progesterone promotes Th2 differentiation in vitro and also has neuroprotective effects (211).
In MOG-induced EAE in C57BL/6 mice, progesterone therapy (either prophylactically or therapeutically) leads to a reduction in the severity of disease, increased IL10 production, reduced axonal damage, and increases production of neurosteroids, which may contribute to neuroprotection (212–215). In chronic EAE induced in DA rats, progesterone reduces severity of disease and is neuroprotective (216). Similarly, in Wistar rats with EAE, progesterone therapy slows the rate of deterioration and enhances remyelination (217).
Androgens control the development and maintenance of masculine characteristics; however, they have other functions, including maintenance of bone mass in both women and men. Androgens are also the precursors of all estrogens. In males, androgens, primarily testosterone, are present in high levels, but these levels decline with increasing age (218). In females, low levels of androgens (particularly the adrenal-derived androgens DHEA and DHEA-S) are present throughout life (219). Testosterone is immunosuppressive and leads to thymic atrophy, while castration of males results in thymic hypertrophy; these effects occur via interaction of sex hormones with receptors on thymic stromal cells (220). Testosterone impairs thymocyte proliferation in response to Concanavalin A (175). In men treated with medical castration, there is a reduction in levels of Treg cells, reduced mitogen-induced activation of CD8+ T cells, and an increased percentage of NK cells (176), suggesting that testosterone can help preserve immune homeostatis to help prevent autoimmunity.
There is also evidence for effects, some positive and some negative, of testosterone on cells of the nervous system. In vitro studies suggest that testosterone can exacerbate cell death in human neurons under oxidative stress conditions, which could be detrimental in MS and EAE, particularly in males (221). However, testosterone is also reported play a role in regeneration of myelin (177); such a mechanism could play a protective role in MS and EAE.
Castration has no effect on disease in C57BL/6 male mice, which are normally susceptible to EAE (222). However, castration of male SJL and (B10.S x SJL)F1 mice, which are normally resistant to development of EAE, increases the incidence and severity of EAE, suggesting that endogenous androgens are protective in males (76, 190). This is also suggested from experiments using male DA rats with SCH-induced EAE, in which levels of testosterone drop during the onset and peak of EAE (223). Androgens have been found to alter the cytokine profile and reduce the pathogenicity of myelin-reactive T cells (224).
Androgen derivatives suppress PLP-induced EAE in female SJL mice and MBP-induced EAE in male B10.PL mice (225). In female SJL mice with MBP-induced EAE, treatment with dihydrotestosterone ameliorates disease and increases IL-10 secretion by T lymphocytes (226). Castration of males removes the sex differences in the induction of oral tolerance to MBP-induced EAE in B10.PL mice (137).
Prolactin, a peptide hormone produced mainly by the anterior pituitary gland, has immunomodulatory effects (227), likely due to the presence of the prolactin receptor on T lymphocytes and other cells of the immune system (228). Prolactin promotes the differentiation of thymocytes (229). In general, prolactin appears to enhance autoimmune diseases, and serum levels of prolactin are increased in some autoimmune diseases (230).
Treatment with bromocriptine, which reduces prolactin secretion, ameliorates EAE in female Lewis rats when given up to one week after induction of EAE (231). Since prolactin levels are elevated in the post-partum period, it is possible that the prolactin could induce the relapses and/or worsening of EAE that is often seen during this period.
There are effects of both the sex chromosomes and the sex hormones on sexual dimorphism in EAE, but these are not always clear-cut and vary from one EAE model to another. The Y chromosome appears to exert an influence on EAE in both female and male mice through paternal transmission of autoimmune susceptibility. The X chromosome also carries many genes relevant to development of EAE, and X dosage effects may occur. The female sex hormones show fairly subtle effects at physiological levels, but are highly immunosuppressive at higher levels in most models tested. In contrast, male sex hormones appear to have stronger effects at physiological levels.
Pregnancy involves physiological changes in the mother, including elevation of cardiac output, increased basal metabolic rate, increased lipid levels, weight gain and alterations to the immune system (232–237). These changes in maternal physiology during pregnancy and lactation are largely mediated through and reflect the net effects of pregnancy hormones (238). In pregnancy there are changes in the levels of estrone, E2, E3, progesterone, prolactin, early pregnancy factor (EPF) (239), alpha-fetoprotein (240) and leptin (241), and elevated levels of growth factors such as insulin-like growth factor (IGF) (242). After delivery, there is a rapid decline in the levels of pregnancy hormones and maternal physiology rapidly returns to normal, although during lactation the levels of prolactin and oxytocin are elevated (243).
Immune changes in pregnancy tend to suppress the maternal immune system; this is thought to be important in preventing rejection of the fetus, which contains foreign (paternal) antigens (244–246). During pregnancy, there are increased Th2 responses and reduced Th1- and Th17- responses (237, 247) and increased levels of circulating Treg cells (186, 248, 249). There are also increased Treg cell numbers in the placenta and draining lymph nodes (250–252). Other evidence of increased Treg activity in pregnancy comes from findings of increased foxp3 expression and increased functional suppression in pregnant and estrogen treated rats (253). The increased level of Treg cells in pregnancy is thought to be due to the effects on the immune system of E2 (185). The changes in pregnancy also include a shift to Th2 immune responses and a shift to greater humoral than cell mediated immunity (254). B cells also show changes in pregnancy, with increases in numbers of regulatory B cells (255).
The effects of pregnancy on EAE are summarized in Table 8. In general, pregnancy reduces the incidence and severity of disease in all models at some time point; however, the specific details of when the effects were most notable differ, depending on the timing of EAE induction vs the beginning of pregnancy.
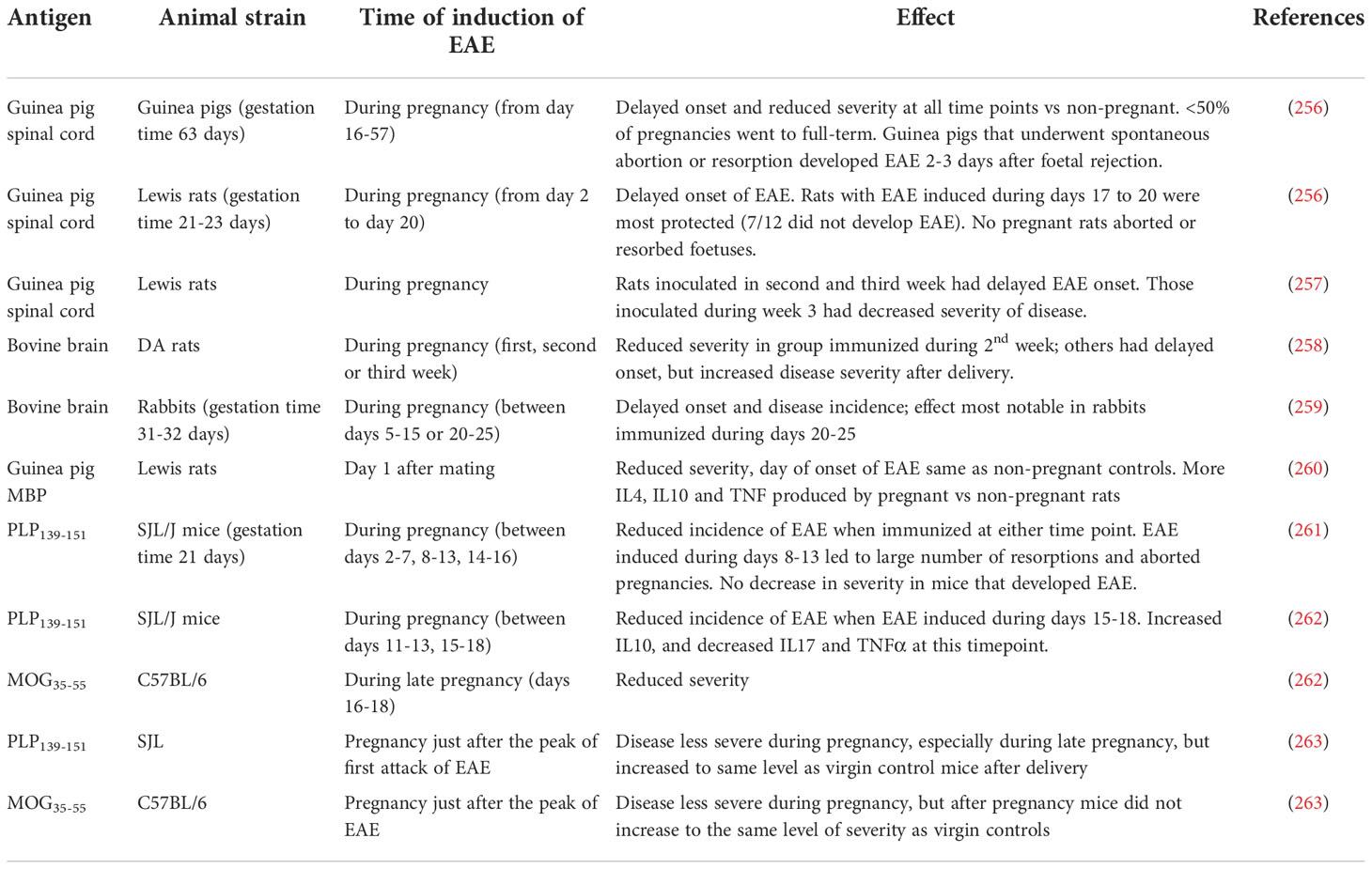
Table 8 Effects of pregnancy in EAE.
In guinea pigs, which have a much longer gestation period (63 days) than mice and rats (~21 days), induction of EAE at any timepoint during pregnancy delays the onset of disease and reduces the severity of EAE compared to induction of disease in non-pregnant animals (256). In rabbits (31-32 days gestation), pregnancy suppresses EAE, with the effects most noticeable when EAE is induced during mid to late pregnancy (day 20-25) (259). Similarly, Lewis rats with EAE induced during the third weeks of pregnancy show the greatest level of protection against the development of disease; rats inoculated in the first week or second week of pregnancy are less protected (256, 257). Interestingly, however, when those rats are re-challenged again 3 weeks after weaning of their litters rats that were protected in pregnancy are more susceptible to the second encephalitogenic challenge, suggesting either that the protective effects during pregnancy are not antigen specific, or that they are not very long-lasting (257). In DA rats, EAE is less severe when disease is induced during the second week of pregnancy, although the rats develop more severe relapses post-partum (258).
In various mouse models, active induction of EAE during late pregnancy leads to a reduction in the incidence of EAE and/or a decrease in clinical severity, compared to virgin controls (261, 262). In contrast, adoptive transfer of activated MBP-specific T cells into pregnant mice during mid-pregnancy, but not in late pregnancy, resulted in a delayed onset of EAE and reduced clinical severity in comparison to virgin controls (264).
The effects of becoming pregnant after the onset of EAE have also been investigated. In both SJL mice with PLP-induced EAE and in C57BL/6 mice with MOG-induced EAE, onset of pregnancy leads to clinical improvement of EAE, but the pathological lesions persist (263). In mice followed until after delivery, EAE scores in the SJL mice return to the same level as in virgin controls, whereas C57BL/6 mice continue to have milder disease than the virgin controls (263).
There have been attempts to transfer the protective effects of pregnancy. Offspring from rats that had EAE during pregnancy show transient protection from EAE, and rats suckled by mothers that had EAE in pregnancy also acquire protection (265).
Several potential mechanisms have been put forward to explain the beneficial effects of pregnancy on EAE. There are alterations of cytokine production (increased expression of mRNA for IL-4 and IL-10) in the spinal cords of pregnant compared to non-pregnant Lewis rats with EAE induced one day after mating, and serum from pregnant rats suppresses lymphocyte proliferation (260, 261). A suppressive serum factor found in late pregnancy has also been implicated in the amelioration of EAE in SJL mice when EAE is induced during the second or third week of pregnancy. Mice with EAE induced during the first week of pregnancy are not protected, suggesting that the serum factor acts on the induction and early effector phases of EAE (261).
Exosomes from both pregnant and non-pregnant mice suppress EAE. However, exosomes are more abundant in pregnancy, and it is suggested that they contribute to protection during pregnancy by suppressing T cell activation and promoting maturation of oligodendrocyte precursor cells and their migration into CNS lesions (266).
Other studies have shown that pregnancy does not appear to affect the induction of myelin-specific T cells, and that there is actually an increase in numbers of such cells in the spleen compared to non-pregnant mice; however, these T cells produce less TNF and IL-17 than those from non-pregnant mice, and there is a higher frequency of IL-10-secreting cells within the CD11b+, CD11c+, CD19+, and CD4+/CD25+ Treg populations (262). Treg cells bearing TCR clonotypes related to the encephalitogenic TCR are increased in pregnant mice in which EAE has been induced on day 7 of pregnancy, whereas such cells are not seen in virgin controls (267). These results suggest that when an antigen is introduced during pregnancy, an immunoregulatory environment towards that antigen predominates.
Although pregnancy is a state when there are circulating growth factors, and although estrogen and progesterone are neuroprotective, there is little information about the effects of pregnancy on neuroprotection and remyelination in EAE.
Several other factors that could contibute to the suppression of EAE by pregnancy have been identified, but only a limited number of studies has been done on each of them. They include:
EPF was first identified as a factor appearing in the serum shortly after conception (268). EPF is identical to chaperonin 10 (269, 270). It is immunomodulatory and has some growth factor-like properties, including enhancing survival of oligodendrocyte precursors in culture (271, 272). EPF suppresses EAE in Lewis rats and in SJL/J mice (273–276).
Alpha-fetoprotein, a glycoprotein produced by the fetus during pregnancy, has immunomodulatory properties (277). In guinea pigs, alpha-fetoprotein administered daily from the time of induction of EAE is able to prevent the development of clinical signs of EAE, and treatment with alpha-fetoprotein commencing after onset of disease also results in a decrease in disease severity (278). In rabbits, treatment with alpha-fetoprotein (daily injections after the onset of clinical signs) also suppresses disease (279). In female C57BL/6 mice with MOG-induced EAE and treated with human alpha-fetoprotein there is increased expression of apoptosis genes and reduced severity of disease (280).
IFN-τ is a type I interferon that is produced in ruminants during pregnancy, and binds to the same receptors as other type I interferons (281). Oral administration of IFN-τ suppresses MBP-induced EAE in NZW mice (282) and both oral and intra-peritoneal administration of IFN-τ after the onset of disease suppresses relapses of EAE induced by MBP in SJL/J mice (283).
Insulin-like growth factor (IGF) has immunomodulatory and immunosuppressive effects (284) and is present in increased levels in serum during pregnancy (242). IGF treatment reduces the severity of EAE in Lewis rats (285).
PIF is a peptide found in the maternal circulation (286). It has a role in immune modulation and neuroprotection (287), and a synthetic version of PIF can reduce signs of PLP-induced EAE in SJL mice, via induction of changes to the phosphorylation state of many molecules in the brain (288).
Overall, pregnancy (particularly the latter part) is a highly immunosuppressive state in almost every EAE model, and studies of pregnancy across different models show more consistent results than do other studies of the effects of sexual dimorphism in EAE. The effects of pregnancy appear to be driven largely, but not solely by estrogen levels, but there are also numerous other pregnancy-related molecules that have been identified that likely contribute to the overall effects of pregnancy. Further study of these molecules is warranted.
Many studies provide evidence that biological sex and pregnancy influence the clinical course of EAE. EAE is a complex disease that has an induction phase involving immune cells in the periphery, an effector phase when the immune cells move into the CNS and when the ability of the nervous system to both take part in the inflammatory response and also to resist damage plays a role, and a recovery phase, when restorative and neuroprotective processes play a role. As indicated in our review, sex- or pregnancy-determined effects can affect each stage of EAE, and their overall effects reflect the net effect of sex differences in many aspects of the pathophysiology of EAE. Additionally, there are other intrinsic and extrinsic factors that can modify the course of EAE and that can also be affected by biological sex. Importantly, the effects of sex vary from one animal model to another and are complex and cannot be widely extrapolated. In EAE, it is therefore essential that studies looking at the effects of biological sex or pregnancy provide full information about the model that is used (i.e. animal strain, sex, the inducing antigen, timing of EAE induction in relation to pregnancy etc.) and, preferably, that more than one EAE model is used to show if any observed effects are generalizable.
There is some evidence of the underlying mechanisms by which sex and pregnancy alter the pathogenesis of EAE. Tables 4 and 5 list the many cell types and cellular processes that have been implicated thus far. These studies show that sex differences in EAE can be found at all stages of disease (induction stage, effector stage, recovery stage), in many cell types and in many molecules and pathways. However, due to the aforementioned complexity and variability from one model to another, studies that comprehensively identify the full gamut of mechanisms underlying the effects of sex or pregnancy on EAE are lacking and some of the effects are likely to be specific to different animal strains. However, despite the variability amongst studies, there are clearly effects of sex and pregnancy in EAE, likely due to the effects of sex hormones and sex chromosomes, and these effects could all be relevant to MS. Going forward, we suggest that even though there is variability among animal strains species and, molecules and pathways that show sex differences are likely to be important in disease. To study these more thoroughly will require investigators to be mindful of the possibility of sex differences and to consider adding additional control groups to experiments. Because the effects of sex hormones vary with age, it would also be useful to study young, adult and aged animals. This is clearly a field that requires further work in order to better our understanding of the mechanisms of sex differences, which are fundamental to biology, and to suggest possible targets or therapies for MS.
PM and JG conceived the idea, conducted literature searches, and drafted the manuscript. Both authors approve the manuscript for publication.
Funded by Grant 1164036 from the National Health and Medical Research Council, Australia.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
BBB, blood–brain barrier; bSCH, bovine spinal cord homogenate; DC, dendritic cells; E2, estradiol; E3, estriol; EAE, experimental autoimmune encephalomyelitis; EPF, early pregnancy factor; ERα, estrogen receptor alpha; FCG, four core genotype; GM-CSF, granulocyte/macrophage colony stimulating factor; gpSCH, guinea pig spinal cord homogenate; IFNγ, interferon gamma; IFNτ, interferon tau; IGF, insulin-like growth factor; IL, interleukin; ILC2, innate lymphoid cell type 2; LPS, lipopolysaccharide; M-CSF, macrophage colony stimulating factor; MBP, myelin basic protein; MOG, myelin/oligodendrocyte glycoprotein; MS, multiple sclerosis; NK, natural killer; PPAR-γ, peroxisome proliferator-activated receptor gamma; PIF, preimplantation factor; PLP, myelin proteolipid protein; S1PR2, sphingosine-1-phosphate receptor 2; TCR, T cell receptor; TGF-β, transforming growth factor beta; TNF, tumor necrosis factor; Treg, regulatory T cell; VLA-4, very late antigen 4 (integrin α4β1).
1. Mank JE. Sex chromosomes and the evolution of sexual dimorphism: lessons from the genome. Am Nat (2009) 173(2):141–50. doi: 10.1086/595754
2. Livernois AM, Graves JA, Waters PD. The origin and evolution of vertebrate sex chromosomes and dosage compensation. Heredity (Edinb). (2012) 108(1):50–8. doi: 10.1038/hdy.2011.106
3. Parsch J, Ellegren H. The evolutionary causes and consequences of sex-biased gene expression. Nat Rev Genet (2013) 14(2):83–7. doi: 10.1038/nrg3376
4. Karp NA, Mason J, Beaudet AL, Benjamini Y, Bower L, Braun RE, et al. Prevalence of sexual dimorphism in mammalian phenotypic traits. Nat Commun (2017) 8:15475. doi: 10.1038/ncomms15475
5. LoMauro A, Aliverti A. Sex and gender in respiratory physiology. Eur Respir Review. (2021) 30(162):210038. doi: 10.1183/16000617.0038-2021
6. McEwen BS, Milner TA. Understanding the broad influence of sex hormones and sex differences in the brain. J Neurosci Res (2017) 95(1-2):24–39. doi: 10.1002/jnr.23809
7. McCombe PA, Greer JM, Mackay IR. Sexual dimorphism in autoimmune disease. Curr Mol Med (2009) 9(9):1058–79. doi: 10.2174/156652409789839116
8. Greer JM, McCombe PA. Role of gender in multiple sclerosis: clinical effects and potential molecular mechanisms. J Neuroimmunol (2011) 234(1-2):7–18. doi: 10.1016/j.jneuroim.2011.03.003
9. Miller DH, Fazekas F, Montalban X, Reingold SC, Trojano M. Pregnancy, sex and hormonal factors in multiple sclerosis. Mult Scler. (2014) 20(5):527–36. doi: 10.1177/1352458513519840
10. McCombe PA. The short and long-term effects of pregnancy on multiple sclerosis and experimental autoimmune encephalomyelitis. J Clin Med (2018) 7(12):494. doi: 10.3390/jcm7120494
11. McCombe PA, Greer JM. Female reproductive issues in multiple sclerosis. Mult Scler. (2013) 19(4):392–402. doi: 10.1177/1352458512452331
12. Rivers TM, Sprunt DH, Berry GP. Observations on attempts to produce acute disseminated encephalomyelitis in monkeys. J Exp Med (1933) 58(1):39–53. doi: 10.1084/jem.58.1.39
13. Kabat EA, Wolf A, Bezer AE. The rapid production of acute disseminated encephalomyelitis in rhesus monkeys by injection of heterologous and homologous brain tissue with adjuvants. J Exp Med (1947) 85(1):117–30. doi: 10.1084/jem.85.1.117
14. Baxter AG. The origin and application of experimental autoimmune encephalomyelitis. Nat Rev Immunol (2007) 7(11):904–12. doi: 10.1038/nri2190
15. Burrows DJ, McGown A, Jain SA, De Felice M, Ramesh TM, Sharrack B, et al. Animal models of multiple sclerosis: From rodents to zebrafish. Mult Scler. (2019) 25(3):306–24. doi: 10.1177/1352458518805246
16. Glatigny S, Bettelli E. Experimental autoimmune encephalomyelitis (EAE) as animal models of multiple sclerosis (MS). Cold Spring Harb Perspect Med (2018) 8(11):a028977. doi: 10.1101/cshperspect.a028977
17. van der Star BJ, Vogel DY, Kipp M, Puentes F, Baker D, Amor S. In vitro and in vivo models of multiple sclerosis. CNS Neurol Disord Drug Targets. (2012) 11(5):570–88. doi: 10.2174/187152712801661284
18. Gold R, Linington C, Lassmann H. Understanding pathogenesis and therapy of multiple sclerosis via animal models: 70 years of merits and culprits in experimental autoimmune encephalomyelitis research. Brain (2006) 129(Pt 8):1953–71. doi: 10.1093/brain/awl075
19. Krishnamoorthy G, Lassmann H, Wekerle H, Holz A. Spontaneous opticospinal encephalomyelitis in a double-transgenic mouse model of autoimmune T cell/B cell cooperation. J Clin Invest. (2006) 116(9):2385–92. doi: 10.1172/JCI28330
20. Pollinger B, Krishnamoorthy G, Berer K, Lassmann H, Bosl MR, Dunn R, et al. Spontaneous relapsing-remitting EAE in the SJL/J mouse: MOG-reactive transgenic T cells recruit endogenous MOG-specific b cells. J Exp Med (2009) 206(6):1303–16. doi: 10.1084/jem.20090299
21. Lassmann H, Bradl M. Multiple sclerosis: experimental models and reality. Acta Neuropathol. (2017) 133(2):223–44. doi: 10.1007/s00401-016-1631-4
22. Constantinescu CS, Farooqi N, O'Brien K, Gran B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br J Pharmacol (2011) 164(4):1079–106. doi: 10.1111/j.1476-5381.2011.01302.x
23. Linington C, Engelhardt B, Kapocs G, Lassman H. Induction of persistently demyelinated lesions in the rat following the repeated adoptive transfer of encephalitogenic T cells and demyelinating antibody. J Neuroimmunol. (1992) 40(2-3):219–24. doi: 10.1016/0165-5728(92)90136-9
24. McCombe PA, de Jersey J, Pender MP. Inflammatory cells, microglia and MHC class II antigen-positive cells in the spinal cord of Lewis rats with acute and chronic relapsing experimental autoimmune encephalomyelitis. J Neuroimmunol. (1994) 51(2):153–67. doi: 10.1016/0165-5728(94)90077-9
25. Muller DM, Pender MP, Greer JM. A neuropathological analysis of experimental autoimmune encephalomyelitis with predominant brain stem and cerebellar involvement and differences between active and passive induction. Acta Neuropathol. (2000) 100(2):174–82. doi: 10.1007/s004019900163
26. Dunham J, van de Vis R, Bauer J, Wubben J, van Driel N, Laman JD, et al. Severe oxidative stress in an acute inflammatory demyelinating model in the rhesus monkey. PloS One (2017) 12(11):e0188013. doi: 10.1371/journal.pone.0188013
27. Jagessar SA, Dijkman K, Dunham J, t Hart BA, Kap YS. Experimental autoimmune encephalomyelitis in marmosets. Methods Mol Biol (2016) 1304:171–86. doi: 10.1007/7651_2014_113
28. Lerner EM, Stone SH, Myers RE. Autoimmune encephalomyelitis and hemorrhagic retinal disease in neonatal, infant, juvenile, and adult monkeys. J Neuroimmunol. (1985) 7(5-6):299–313.
29. t'Hart BA. Experimental autoimmune encephalomyelitis in the common marmoset: a translationally relevant model for the cause and course of multiple sclerosis. Primate Biol (2019) 6(1):17–58. doi: 10.5194/pb-6-17-2019
30. Uccelli A, Giunti D, Mancardi G, Caroli F, Fiorone M, Seri M, et al. Characterization of the response to myelin basic protein in a non human primate model for multiple sclerosis. Eur J Immunol (2001) 31(2):474–9. doi: 10.1002/1521-4141(200102)31:2<474::AID-IMMU474>3.0.CO;2-9
31. van Lambalgen R, Jonker M. Experimental allergic encephalomyelitis in rhesus monkeys: I. immunological parameters in EAE resistant and susceptible rhesus monkeys. Clin Exp Immunol (1987) 68(1):100–7.
32. Meyer TJ, Azuma I, Ribi EE. Biologically active components from mycobacterial cell walls. III. production of experimental allergic encephalomyelitis in guinea-pigs. Immunology. (1975) 28(2):219–29. doi: 10.1016/0008-8749(75)90181-1
33. Shaw CM, Alvord EC Jr., Fahlberg WJ, Kies MW. Adjuvant-antigen relationships in the production of experimental "allergic" encephalomyelitis in the guinea pig. J Exp Med (1962) 115:169–79. doi: 10.1084/jem.115.1.169
34. Stone SH, Lerner EM, Goode JH. Adoptive autoimmune encephalomyelitis in inbred guinea pigs: immunological and histological aspects. Science. (1968) 159(3818):995–7. doi: 10.1126/science.159.3818.995
35. Paterson PY, Steele FM. Cerebrospinal fluid beta-glucuronidase activity in rabbits with experimental allergic encephalomyelitis. Clin Exp Immunol (1970) 7(5):759–68.
36. Villarroya H, Dalix AM, Paraut M, Oriol R. Differential susceptibility to experimental allergic encephalomyelitis (EAE) in genetically defined a+ and a- rabbits. Autoimmunity (1990) 6(1-2):47–60. doi: 10.3109/08916939008993369
37. Massanari RM, Paterson PY, Lipton HL. Potentiation of experimental allergic encephalomyelitis in hamsters with persistent encephalitis due to measles virus. J Infect Dis (1979) 139(3):297–303. doi: 10.1093/infdis/139.3.297
38. Thomas L, Paterson PY, Smithwick B. Acute disseminated encephalomyelitis following immunization with homologous brain extracts; studies on the role of a circulating antibody in the production of the condition in dogs. J Exp Med (1950) 92(2):133–52. doi: 10.1084/jem.92.2.133
39. Ainsworth SK, Brostoff S, Sharbaugh RJ, Fitts CT. Brief note. induction of experimental allergic encephalomyelitis in sheep with purified bovine basic protein and adjuvant. Pathology. (1979) 11(3):457–60. doi: 10.3109/00313027909059022
40. Willenborg DO. Transfer of lesions of allergic encephalomyelitis in sheep with cell-free lymph from animals sensitized with homologous spinal cord plus adjuvant. Scand J Immunol (1982) 16(5):437–41. doi: 10.1111/j.1365-3083.1982.tb00744.x
41. Taylor DL, Burrell R. The immunologic responses of the north American opossum (Didelphys virginiana). J Immunol (1968) 101(6):1207–16. doi: 10.4049/jimmunol.101.6.1207
42. Stromnes IM, Goverman JM. Passive induction of experimental allergic encephalomyelitis. Nat Protoc (2006) 1(4):1952–60. doi: 10.1038/nprot.2006.284
43. Stromnes IM, Goverman JM. Active induction of experimental allergic encephalomyelitis. Nat Protoc (2006) 1(4):1810–9. doi: 10.1038/nprot.2006.285
44. Miller SD, Karpus WJ, Davidson TS. Experimental autoimmune encephalomyelitis in the mouse. Curr Protoc Immunol (2007) 15:1.1-1.18. doi: 10.1002/0471142735.im1501s77
45. Bittner S, Afzali AM, Wiendl H, Meuth SG. Myelin oligodendrocyte glycoprotein (MOG35-55) induced experimental autoimmune encephalomyelitis (EAE) in C57BL/6 mice. J Vis Exp (2014) 86:e51275. doi: 10.3791/51275
46. Bebo BF Jr., Vandenbark AA, Offner H. Male SJL mice do not relapse after induction of EAE with PLP 139-151. J Neurosci Res (1996) 45(6):680–9. doi: 10.1002/(SICI)1097-4547(19960915)45:6<680::AID-JNR4>3.0.CO;2-4
47. Massella A, D'Intino G, Fernandez M, Sivilia S, Lorenzini L, Giatti S, et al. Gender effect on neurodegeneration and myelin markers in an animal model for multiple sclerosis. BMC Neurosci (2012) 13:12. doi: 10.1186/1471-2202-13-12
48. Okuda Y, Okuda M, Bernard CC. Gender does not influence the susceptibility of C57BL/6 mice to develop chronic experimental autoimmune encephalomyelitis induced by myelin oligodendrocyte glycoprotein. Immunol Lett (2002) 81(1):25–9. doi: 10.1016/S0165-2478(01)00339-X
49. Papenfuss TL, Rogers CJ, Gienapp I, Yurrita M, McClain M, Damico N, et al. Sex differences in experimental autoimmune encephalomyelitis in multiple murine strains. J Neuroimmunol. (2004) 150(1-2):59–69. doi: 10.1016/j.jneuroim.2004.01.018
50. Sinha S, Kaler LJ, Proctor TM, Teuscher C, Vandenbark AA, Offner H. IL-13-mediated gender difference in susceptibility to autoimmune encephalomyelitis. J Immunol (2008) 180(4):2679–85. doi: 10.4049/jimmunol.180.4.2679
51. Teuscher C, Bunn JY, Fillmore PD, Butterfield RJ, Zachary JF, Blankenhorn EP. Gender, age, and season at immunization uniquely influence the genetic control of susceptibility to histopathological lesions and clinical signs of experimental allergic encephalomyelitis: implications for the genetics of multiple sclerosis. Am J Pathol (2004) 165(5):1593–602. doi: 10.1016/S0002-9440(10)63416-5
52. Voskuhl RR, Pitchekian Halabi H, MacKenzie Graham A, McFarland HF, Raine CS. Gender differences in autoimmune demyelination in the mouse: implications for multiple sclerosis. Ann Neurol (1996) 39(6):724–33. doi: 10.1002/ana.410390608
53. Alvarez Sanchez N, Dunn SE. Immune cell contributors to the female sex bias in multiple sclerosis and experimental autoimmune encephalomyelitis. Curr Top Behav Neurosci (2022). doi: 10.1007/7854_2022_324
54. Haanstra KG, Jagessar SA, Bauchet AL, Doussau M, Fovet CM, Heijmans N, et al. Induction of experimental autoimmune encephalomyelitis with recombinant human myelin oligodendrocyte glycoprotein in incomplete freund's adjuvant in three non-human primate species. J Neuroimmune Pharmacol (2013) 8(5):1251–64. doi: 10.1007/s11481-013-9487-z
55. Kerlero de Rosbo N, Brok HP, Bauer J, Kaye JF, t Hart BA, Ben Nun A. Rhesus monkeys are highly susceptible to experimental autoimmune encephalomyelitis induced by myelin oligodendrocyte glycoprotein: characterisation of immunodominant T- and b-cell epitopes. J Neuroimmunol. (2000) 110(1-2):83–96. doi: 10.1016/S0165-5728(00)00306-4
56. Kopeloff LM, Kopeloff N. Neurologic manifestations in laboratory animals produced by organ (adjuvant) emulsions. J Immunol (1947) 57(3):229–37.
57. Wisniewski HM, Keith AB. Chronic relapsing experimental allergic encephalomyelitis: an experimental model of multiple sclerosis. Ann Neurol (1977) 1(2):144–8. doi: 10.1002/ana.410010207
58. Keith AB. Sex difference in Lewis rats in the incidence of recurrent experimental allergic encephalomyelitis. Nature. (1978) 272(5656):824–5. doi: 10.1038/272824a0
59. Pender MP, Sears TA. Involvement of the dorsal root ganglion in acute experimental allergic encephalomyelitis in the Lewis rat. a histological and electrophysiological study. J Neurol Sci (1986) 72(2-3):231–42. doi: 10.1016/0022-510X(86)90011-0
60. Chalk JB, McCombe PA, Smith R, Pender MP. Clinical and histological findings in proteolipid protein-induced experimental autoimmune encephalomyelitis (EAE) in the Lewis rat. distribution of demyelination differs from that in EAE induced by other antigens. J Neurol Sci (1994) 123(1-2):154–61. doi: 10.1016/0022-510x(94)90218-6
61. Panitch H, Ciccone C. Induction of recurrent experimental allergic encephalomyelitis with myelin basic protein. Ann Neurol (1981) 9(5):433–8. doi: 10.1002/ana.410090504
62. Flytzani S, Guerreiro Cacais AO, N'Diaye M, Lindner M, Linington C, Meinl E, et al. MOG-induced experimental autoimmune encephalomyelitis in the rat species triggers anti-neurofascin antibody response that is genetically regulated. J Neuroinflammation. (2015) 12:194. doi: 10.1186/s12974-015-0417-2
63. Staykova MA, Cowden W, Willenborg DO. Macrophages and nitric oxide as the possible cellular and molecular basis for strain and gender differences in susceptibility to autoimmune central nervous system inflammation. Immunol Cell Biol (2002) 80(2):188–97. doi: 10.1046/j.1440-1711.2002.01072.x
64. Matejuk A, Hopke C, Vandenbark AA, Hurn PD, Offner H. Middle-age male mice have increased severity of experimental autoimmune encephalomyelitis and are unresponsive to testosterone therapy. J Immunol (2005) 174(4):2387–95. doi: 10.4049/jimmunol.174.4.2387
65. Cua DJ, Hinton DR, Kirkman L, Stohlman SA. Macrophages regulate induction of delayed-type hypersensitivity and experimental allergic encephalomyelitis in SJL mice. Eur J Immunol (1995) 25(8):2318–24. doi: 10.1002/eji.1830250830
66. Massilamany C, Thulasingam S, Steffen D, Reddy J. Gender differences in CNS autoimmunity induced by mimicry epitope for PLP 139-151 in SJL mice. J Neuroimmunol. (2011) 230(1-2):95–104. doi: 10.1016/j.jneuroim.2010.09.011
67. Fillmore PD, Blankenhorn EP, Zachary JF, Teuscher C. Adult gonadal hormones selectively regulate sexually dimorphic quantitative traits observed in experimental allergic encephalomyelitis. Am J Pathol (2004) 164(1):167–75. doi: 10.1016/S0002-9440(10)63107-0
68. Spach KM, Blake M, Bunn JY, McElvany B, Noubade R, Blankenhorn EP, et al. Cutting edge: the y chromosome controls the age-dependent experimental allergic encephalomyelitis sexual dimorphism in SJL/J mice. J Immunol (2009) 182(4):1789–93. doi: 10.4049/jimmunol.0803200
69. Butterfield RJ, Blankenhorn EP, Roper RJ, Zachary JF, Doerge RW, Sudweeks J, et al. Genetic analysis of disease subtypes and sexual dimorphisms in mouse experimental allergic encephalomyelitis (EAE): relapsing/remitting and monophasic remitting/nonrelapsing EAE are immunogenetically distinct. J Immunol (1999) 162(5):3096–102.
70. Columba Cabezas S, Iaffaldano G, Chiarotti F, Alleva E, Cirulli F. Early handling increases susceptibility to experimental autoimmune encephalomyelitis (EAE) in C57BL/6 male mice. J Neuroimmunol. (2009) 212(1-2):10–6. doi: 10.1016/j.jneuroim.2009.05.007
71. Anderson AC, Chandwaskar R, Lee DH, Sullivan JM, Solomon A, Rodriguez Manzanet R, et al. A transgenic model of central nervous system autoimmunity mediated by CD4+ and CD8+ T and b cells. J Immunol (2012) 188(5):2084–92. doi: 10.4049/jimmunol.1102186
72. Bettelli E, Baeten D, Jager A, Sobel RA, Kuchroo VK. Myelin oligodendrocyte glycoprotein-specific T and b cells cooperate to induce a devic-like disease in mice. J Clin Invest. (2006) 116(9):2393–402. doi: 10.1172/JCI28334
73. Dhaeze T, Lachance C, Tremblay L, Grasmuck C, Bourbonnière L, Larouche S, et al. Sex-dependent factors encoded in the immune compartment dictate relapsing or progressive phenotype in demyelinating disease. JCI Insight (2019) 4(6):e124885. doi: 10.1172/jci.insight.124885
74. Bebo BF Jr., Schuster JC, Vandenbark AA, Offner H. Gender differences in experimental autoimmune encephalomyelitis develop during the induction of the immune response to encephalitogenic peptides. J Neurosci Res (1998) 52(4):420–6. doi: 10.1002/(SICI)1097-4547(19980515)52:4<420::AID-JNR5>3.0.CO;2-B
75. Doss PMIA, Umair M, Baillargeon J, Fazazi R, Fudge N, Akbar I, et al. Male Sex chromosomal complement exacerbates the pathogenicity of Th17 cells in a chronic model of central nervous system autoimmunity. Cell Rep (2021) 34(10):108833. doi: 10.1016/j.celrep.2021.108833
76. Zhang MA, Rego D, Moshkova M, Kebir H, Chruscinski A, Nguyen H, et al. Peroxisome proliferator-activated receptor (PPAR)alpha and -gamma regulate IFNgamma and IL-17A production by human T cells in a sex-specific way. Proc Natl Acad Sci U S A. (2012) 109(24):9505–10. doi: 10.1073/pnas.1118458109
77. Russi AE, Walker Caulfield ME, Ebel ME, Brown MA. Cutting edge: c-kit signaling differentially regulates type 2 innate lymphoid cell accumulation and susceptibility to central nervous system demyelination in Male and female SJL mice. J Immunol (2015) 194(12):5609. doi: 10.4049/jimmunol.1500068
78. Dunn SE, Ousman SS, Sobel RA, Zuniga L, Baranzini SE, Youssef S, et al. Peroxisome proliferator-activated receptor (PPAR)alpha expression in T cells mediates gender differences in development of T cell-mediated autoimmunity. J Exp Med (2007) 204(2):321–30. doi: 10.1084/jem.20061839
79. Russi AE, Ebel ME, Yang Y, Brown MA. Male-Specific IL-33 expression regulates sex-dimorphic EAE susceptibility. Proc Natl Acad Sci (2018) 115(7):e1520. doi: 10.1073/pnas.1710401115
80. Reddy J, Waldner H, Zhang X, Illes Z, Wucherpfennig KW, Sobel RA, et al. Cutting edge: CD4+CD25+ regulatory T cells contribute to gender differences in susceptibility to experimental autoimmune encephalomyelitis. J Immunol (2005) 175(9):5591–5. doi: 10.4049/jimmunol.175.9.5591
81. Wheeler C, Nabors LB, Barnum S, Yang X, Hu X, Schoeb TR, et al. Sex hormone-dependent attenuation of EAE in a transgenic mouse with astrocytic expression of the RNA regulator HuR. J Neuroimmunol. (2012) 246(1-2):34–7. doi: 10.1016/j.jneuroim.2012.02.014
82. Tassoni A, Farkhondeh V, Itoh Y, Itoh N, Sofroniew MV, Voskuhl RR. The astrocyte transcriptome in EAE optic neuritis shows complement activation and reveals a sex difference in astrocytic C3 expression. Sci Rep (2019) 9(1):10010. doi: 10.1038/s41598-019-46232-6
83. Han J, Zhu K, Zhou K, Hakim R, Sankavaram SR, Blomgren K, et al. Sex-specific effects of microglia-like cell engraftment during experimental autoimmune encephalomyelitis. Int J Mol Sci (2020) 21(18):6824. doi: 10.3390/ijms21186824
84. Dasgupta S, Jana M, Liu X, Pahan K. Myelin basic protein-primed T cells of female but not male mice induce nitric-oxide synthase and proinflammatory cytokines in microglia: implications for gender bias in multiple sclerosis. J Biol Chem (2005) 280(38):32609–17. doi: 10.1074/jbc.M500299200
85. Schrewe L, Lill CM, Liu T, Salmen A, Gerdes LA, Guillot Noel L, et al. Investigation of sex-specific effects of apolipoprotein e on severity of EAE and MS. J Neuroinflammation. (2015) 12:234. doi: 10.1186/s12974-015-0429-y
86. Catuneanu A, Paylor JW, Winship I, Colbourne F, Kerr BJ. Sex differences in central nervous system plasticity and pain in experimental autoimmune encephalomyelitis. Pain. (2019) 160(5):1037–49. doi: 10.1097/j.pain.0000000000001483
87. Luna RM, Kormendy D, Brunner Weinzierl MC. Female-biased incidence of experimental autoimmune encephalomyelitis reflects sexually dimorphic expression of surface CTLA-4 (CD152) on T lymphocytes. Gend Med (2010) 7(4):296–308. doi: 10.1016/j.genm.2010.08.005
88. Hoghooghi V, Palmer AL, Frederick A, Jiang Y, Merkens JE, Balakrishnan A, et al. Cystatin c plays a sex-dependent detrimental role in experimental autoimmune encephalomyelitis. Cell Rep (2020) 33(1):108236. doi: 10.1016/j.celrep.2020.108236
89. Ding M, Wong JL, Rogers NE, Ignarro LJ, Voskuhl RR. Gender differences of inducible nitric oxide production in SJL/J mice with experimental autoimmune encephalomyelitis. J Neuroimmunol. (1997) 77(1):99–106. doi: 10.1016/S0165-5728(97)00065-9
90. Dimitrijevic M, Kotur Stevuljevic J, Stojic Vukanic Z, Vujnovic I, Pilipovic I, Nacka Aleksic M, et al. Sex difference in oxidative stress parameters in spinal cord of rats with experimental autoimmune encephalomyelitis: Relation to neurological deficit. Neurochem Res (2017) 42(2):481–92. doi: 10.1007/s11064-016-2094-7
91. Krementsov DN, Noubade R, Dragon JA, Otsu K, Rincon M, Teuscher C. Sex-specific control of central nervous system autoimmunity by p38 mitogen-activated protein kinase signaling in myeloid cells. Ann Neurol (2014) 75(1):50–66. doi: 10.1002/ana.24020
92. Cruz Orengo L, Daniels BP, Dorsey D, Basak SA, Grajales Reyes JG, McCandless EE, et al. Enhanced sphingosine-1-phosphate receptor 2 expression underlies female CNS autoimmunity susceptibility. J Clin Invest. (2014) 124(6):2571–84. doi: 10.1172/JCI73408
93. Du S, Itoh N, Askarinam S, Hill H, Arnold AP, Voskuhl RR. XY sex chromosome complement, compared with XX, in the CNS confers greater neurodegeneration during experimental autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. (2014) 111(7):2806–11. doi: 10.1073/pnas.1307091111
94. Spach KM, Hayes CE. Vitamin D3 confers protection from autoimmune encephalomyelitis only in female mice. J Immunol (2005) 175(6):4119–26. doi: 10.4049/jimmunol.175.6.4119
95. Gracey E, Yao Y, Green B, Qaiyum Z, Baglaenko Y, Lin A, et al. Sexual dimorphism in the Th17 signature of ankylosing spondylitis. Arthritis Rheumatol (2016) 68(3):679–89. doi: 10.1002/art.39464
96. Elderman M, van Beek A, Brandsma E, de Haan B, Savelkoul H, de Vos P, et al. Sex impacts Th1 cells, tregs, and DCs in both intestinal and systemic immunity in a mouse strain and location-dependent manner. Biol Sex Differ (2016) 7:21. doi: 10.1186/s13293-016-0075-9
97. Wiedrick J, Meza Romero R, Gerstner G, Seifert H, Chaudhary P, Headrick A, et al. Sex differences in EAE reveal common and distinct cellular and molecular components. Cell Immunol (2021) 359:104242. doi: 10.1016/j.cellimm.2020.104242
98. Cloake NC, Beaino W, Trifilieff E, Greer JM. Thiopalmitoylation of altered peptide ligands enhances their protective effects in an animal model of multiple sclerosis. J Immunol (2014) 192(5):2244–51. doi: 10.4049/jimmunol.1301871
99. Carbajal KS, Mironova Y, Ulrich Lewis JT, Kulkarni D, Grifka Walk HM, Huber AK, et al. Th Cell diversity in experimental autoimmune encephalomyelitis and multiple sclerosis. J Immunol (2015) 195(6):2552–9. doi: 10.4049/jimmunol.1501097
100. Hirota K, Duarte JH, Veldhoen M, Hornsby E, Li Y, Cua DJ, et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat Immunol (2011) 12(3):255–63. doi: 10.1038/ni.1993
101. Suryani S, Sutton I. An interferon-gamma-producing Th1 subset is the major source of IL-17 in experimental autoimmune encephalitis. J Neuroimmunol. (2007) 183(1-2):96–103. doi: 10.1016/j.jneuroim.2006.11.023
102. Segal BM. The diversity of encephalitogenic CD4+ T cells in multiple sclerosis and its animal models. J Clin Med (2019) 8(1):120. doi: 10.3390/jcm8010120
103. Park HJ, Park HS, Lee JU, Bothwell AL, Choi JM. Sex-based selectivity of PPARgamma regulation in Th1, Th2, and Th17 differentiation. Int J Mol Sci (2016) 17(8):1347. doi: 10.3390/ijms17081347
104. Liva SM, Voskuhl RR. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. J Immunol (2001) 167(4):2060–7. doi: 10.4049/jimmunol.167.4.2060
105. Baitsch D, Bock HH, Engel T, Telgmann R, Muller Tidow C, Varga G, et al. Apolipoprotein e induces antiinflammatory phenotype in macrophages. Arterioscler Thromb Vasc Biol (2011) 31(5):1160–8. doi: 10.1161/ATVBAHA.111.222745
106. Wei J, Zheng M, Liang P, Wei Y, Yin X, Tang Y, et al. Apolipoprotein e and its mimetic peptide suppress Th1 and Th17 responses in experimental autoimmune encephalomyelitis. Neurobiol Dis (2013) 56:59–65. doi: 10.1016/j.nbd.2013.04.009
107. Crespo Castrillo A, Arevalo MA. Microglial and astrocytic function in physiological and pathological conditions: Estrogenic modulation. Int J Mol Sci (2020) 21(9):3219. doi: 10.3390/ijms21093219
108. McCarthy MM, Todd BJ, Amateau SK. Estradiol modulation of astrocytes and the establishment of sex differences in the brain. Ann N Y Acad Sci (2003) 1007:283–97. doi: 10.1196/annals.1286.027
109. Santos-Galindo M, Acaz Fonseca E, Bellini MJ, Garcia Segura LM. Sex differences in the inflammatory response of primary astrocytes to lipopolysaccharide. Biol Sex Differ (2011) 2:7. doi: 10.1186/2042-6410-2-7
110. Yanguas Casas N, Crespo Castrillo A, Arevalo MA, Garcia Segura LM. Aging and sex: Impact on microglia phagocytosis. Aging Cell (2020) 19(8):e13182. doi: 10.1111/acel.13182
111. Rurak GM, Simard S, Freitas-Andrade M, Lacoste B, Charih F, Van Geel A, et al. Sex differences in developmental patterns of neocortical astroglia: A mouse translatome database. Cell Rep (2022) 38(5):110310. doi: 10.1016/j.celrep.2022.110310
112. Morrison HW, Filosa JA. Sex differences in astrocyte and microglia responses immediately following middle cerebral artery occlusion in adult mice. Neuroscience. (2016) 339:85–99. doi: 10.1016/j.neuroscience.2016.09.047
113. Hanamsagar R, Alter MD, Block CS, Sullivan H, Bolton JL, Bilbo SD. Generation of a microglial developmental index in mice and in humans reveals a sex difference in maturation and immune reactivity. Glia. (2017) 65(9):1504–20. doi: 10.1002/glia.23176
114. Daly CM, Saxena J, Singh J, Bullard MR, Bondy EO, Saxena A, et al. Sex differences in response to a high fat, high sucrose diet in both the gut microbiome and hypothalamic astrocytes and microglia. Nutr Neurosci (2022) 25(2):321–35. doi: 10.1080/1028415X.2020.1752996
115. Rosin JM, Vora SR, Kurrasch DM. Depletion of embryonic microglia using the CSF1R inhibitor PLX5622 has adverse sex-specific effects on mice, including accelerated weight gain, hyperactivity and anxiolytic-like behaviour. Brain Behav Immun (2018) 73:682–97. doi: 10.1016/j.bbi.2018.07.023
116. Morizawa Y, Sato K, Takaki J, Kawasaki A, Shibata K, Suzuki T, et al. Cell-autonomous enhancement of glutamate-uptake by female astrocytes. Cell Mol Neurobiology. (2012) 32(6):953–6. doi: 10.1007/s10571-012-9829-z
117. Spence RD, Hamby ME, Umeda E, Itoh N, Du S, Wisdom AJ, et al. Neuroprotection mediated through estrogen receptor-alpha in astrocytes. Proc Natl Acad Sci U S A. (2011) 108(21):8867–72. doi: 10.1073/pnas.1103833108
118. Chowen JA, Argente Arizón P, Freire-Regatillo A, Argente J. Sex differences in the neuroendocrine control of metabolism and the implication of astrocytes. Front Neuroendocrinology. (2018) 48:3–12. doi: 10.1016/j.yfrne.2017.05.003
119. Jaber SM, Bordt EA, Bhatt NM, Lewis DM, Gerecht S, Fiskum G, et al. Sex differences in the mitochondrial bioenergetics of astrocytes but not microglia at a physiologically relevant brain oxygen tension. Neurochem Int (2018) 117:82–90. doi: 10.1016/j.neuint.2017.09.003
120. Brambilla R. The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. (2019) 137(5):757–83. doi: 10.1007/s00401-019-01980-7
121. Stathopoulou C, Nikoleri D, Bertsias G. Immunometabolism: an overview and therapeutic prospects in autoimmune diseases. Immunotherapy. (2019) 11(9):813–29. doi: 10.2217/imt-2019-0002
122. Graham KL, Werner BJ, Moyer KM, Patton AK, Krois CR, Yoo HS, et al. DGAT1 inhibits retinol-dependent regulatory T cell formation and mediates autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. (2019) 116(8):3126–35. doi: 10.1073/pnas.1817669116
123. Eaton SA, Sethi JK. Immunometabolic links between estrogen, adipose tissue and female reproductive metabolism. Biol (Basel) (2019) 8(1):8. doi: 10.3390/biology8010008
124. Azevedo RB, Lacava ZG, Miyasaka CK, Chaves SB, Curi R. Regulation of antioxidant enzyme activities in male and female rat macrophages by sex steroids. Braz J Med Biol Res (2001) 34(5):683–7. doi: 10.1590/S0100-879X2001000500018
125. Dantas AP, Franco Mdo C, Silva-Antonialli MM, Tostes RC, Fortes ZB, Nigro D, et al. Gender differences in superoxide generation in microvessels of hypertensive rats: role of NAD(P)H-oxidase. Cardiovasc Res (2004) 61(1):22–9. doi: 10.1016/j.cardiores.2003.10.010
126. Kleinewietfeld M, Manzel A, Titze J, Kvakan H, Yosef N, Linker RA, et al. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature. (2013) 496(7446):518–22. doi: 10.1038/nature11868
127. Ji Z, Wu S, Xu Y, Qi J, Su X, Shen L. Obesity promotes EAE through IL-6 and CCL-2-Mediated T cells infiltration. Front Immunol (2019) 10:1881. doi: 10.3389/fimmu.2019.01881
128. Zhang D, Jin W, Wu R, Li J, Park SA, Tu E, et al. High glucose intake exacerbates autoimmunity through reactive-Oxygen-Species-Mediated TGF-β cytokine activation. Immunity. (2019) 51(4):671–81.e5. doi: 10.1016/j.immuni.2019.08.001
129. Krementsov DN, Case LK, Hickey WF, Teuscher C. Exacerbation of autoimmune neuroinflammation by dietary sodium is genetically controlled and sex specific. FASEB J (2015) 29(8):3446–57. doi: 10.1096/fj.15-272542
130. Wu C, Yosef N, Thalhamer T, Zhu C, Xiao S, Kishi Y, et al. Induction of pathogenic TH17 cells by inducible salt-sensing kinase SGK1. Nature. (2013) 496(7446):513–7. doi: 10.1038/nature11984
131. Chu F, Shi M, Lang Y, Shen D, Jin T, Zhu J, et al. Gut microbiota in multiple sclerosis and experimental autoimmune encephalomyelitis: Current applications and future perspectives. Mediators Inflamm (2018) 2018:8168717. doi: 10.1155/2018/8168717
132. Gomez A, Luckey D, Taneja V. The gut microbiome in autoimmunity: Sex matters. Clin Immunol (2015) 159(2):154–62. doi: 10.1016/j.clim.2015.04.016
133. Jaggar M, Rea K, Spichak S, Dinan TG, Cryan JF. You've got male: Sex and the microbiota-gut-brain axis across the lifespan. Front Neuroendocrinol. (2020) 56:100815. doi: 10.1016/j.yfrne.2019.100815
134. Benedek G, Zhang J, Nguyen H, Kent G, Seifert HA, Davin S, et al. Estrogen protection against EAE modulates the microbiota and mucosal-associated regulatory cells. J Neuroimmunol. (2017) 310:51–9. doi: 10.1016/j.jneuroim.2017.06.007
135. Caslin B, Maguire C, Karmakar A, Mohler K, Wylie D, Melamed E. Alcohol shifts gut microbial networks and ameliorates a murine model of neuroinflammation in a sex-specific pattern. Proc Natl Acad Sci U S A. (2019) 116(51):25808–15. doi: 10.1073/pnas.1912359116
136. Krementsov DN, Asarian L, Fang Q, McGill MM, Teuscher C. Sex-specific gene-by-Vitamin d interactions regulate susceptibility to central nervous system autoimmunity. Front Immunol (2018) 9:1622. doi: 10.3389/fimmu.2018.01622
137. Bebo BF Jr., Adlard K, Schuster JC, Unsicker L, Vandenbark AA, Offner H. Gender differences in protection from EAE induced by oral tolerance with a peptide analogue of MBP-Ac1-11. J Neurosci Res (1999) 55(4):432–40. doi: 10.1002/(SICI)1097-4547(19990215)55:4<432::AID-JNR4>3.0.CO;2-2
138. Hill NJ, Hultcrantz M, Sarvetnick N, Flodstrom Tullberg M. The target tissue in autoimmunity–an influential niche. Eur J Immunol (2007) 37(3):589–97. doi: 10.1002/eji.200636368
139. Liston A. Autoimmunity: beyond the immune system. Immunol Cell Biol (2008) 86(4):295–6. doi: 10.1038/icb.2008.10
140. Swamydas M, Bessert D, Skoff R. Sexual dimorphism of oligodendrocytes is mediated by differential regulation of signaling pathways. J Neurosci Res (2009) 87(15):3306–19. doi: 10.1002/jnr.21943
141. Yasuda K, Maki T, Kinoshita H, Kaji S, Toyokawa M, Nishigori R, et al. Sex-specific differences in transcriptomic profiles and cellular characteristics of oligodendrocyte precursor cells. Stem Cell Res (2020) 46:101866. doi: 10.1016/j.scr.2020.101866
142. Georgiou P, Zanos P, Mou TCM, An X, Gerhard DM, Dryanovski DI, et al. Experimenters’ sex modulates mouse behaviors and neural responses to ketamine via corticotropin releasing factor. Nat Neurosci (2022) 25(9):1191–200. doi: 10.1038/s41593-022-01146-x
143. Sorge RE, Martin LJ, Isbester KA, Sotocinal SG, Rosen S, Tuttle AH, et al. Olfactory exposure to males, including men, causes stress and related analgesia in rodents. Nat Methods (2014) 11(6):629–32. doi: 10.1038/nmeth.2935
144. Graves JAM. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu Rev Genet (2008) 42(1):565–86. doi: 10.1146/annurev.genet.42.110807.091714
145. Capel B. WOMEN IN REPRODUCTIVE SCIENCE: To be or not to be a testis. Reproduction (2019) 158(6):F101–F11. doi: 10.1530/REP-19-0151
146. Colaco S, Modi D. Consequences of y chromosome microdeletions beyond male infertility. J Assist Reprod Genet (2019) 36(7):1329–37. doi: 10.1007/s10815-019-01492-z
147. Khan SI, Andrews KL, Jennings GL, Sampson AK, Chin Dusting JPF. Y chromosome, hypertension and cardiovascular disease: Is inflammation the answer? Int J Mol Sci (2019) 20(12):2892. doi: 10.3390/ijms20122892
148. Maan AA, Eales J, Akbarov A, Rowland J, Xu X, Jobling MA, et al. The y chromosome: a blueprint for men's health? Eur J Hum Genet (2017) 25(11):1181–8. doi: 10.1038/ejhg.2017.128
149. Hughes JF, Skaletsky H, Pyntikova T, Graves TA, van Daalen SK, Minx PJ, et al. Chimpanzee and human y chromosomes are remarkably divergent in structure and gene content. Nature. (2010) 463(7280):536–9. doi: 10.1038/nature08700
150. Soh YQ, Alföldi J, Pyntikova T, Brown LG, Graves T, Minx PJ, et al. Sequencing the mouse y chromosome reveals convergent gene acquisition and amplification on both sex chromosomes. Cell. (2014) 159(4):800–13. doi: 10.1016/j.cell.2014.09.052
151. Jansson L, Holmdahl R. The y chromosome-linked "autoimmune accelerating" yaa gene suppresses collagen-induced arthritis. Eur J Immunol (1994) 24(5):1213–7. doi: 10.1002/eji.1830240531
152. Subramanian S, Tus K, Li QZ, Wang A, Tian XH, Zhou J, et al. A Tlr7 translocation accelerates systemic autoimmunity in murine lupus. Proc Natl Acad Sci U S A. (2006) 103(26):9970–5. doi: 10.1073/pnas.0603912103
153. Greer JM, Csurhes PA, Muller DM, Pender MP. Correlation of blood T cell and antibody reactivity to myelin proteins with HLA type and lesion localization in multiple sclerosis. J Immunol (2008) 180(9):6402–10. doi: 10.4049/jimmunol.180.9.6402
154. Greer JM, Trifilieff E, Pender MP. Correlation between anti-myelin proteolipid protein (PLP) antibodies and disease severity in multiple sclerosis patients with PLP response-permissive HLA types. Front Immunol (2020) 11:1891. doi: 10.3389/fimmu.2020.01891
155. Graves JA. Review: Sex chromosome evolution and the expression of sex-specific genes in the placenta. Placenta. (2010) 31 Suppl:S27–32. doi: 10.1016/j.placenta.2009.12.029
156. Lyon MF. Gene action in the X-chromosome of the mouse (Mus musculus l.). Nature (1961) 190:372–3. doi: 10.1038/190372a0
157. Fang H, Disteche CM, Berletch JB. X inactivation and escape: Epigenetic and structural features. Front Cell Dev Biol (2019) 7(219). doi: 10.3389/fcell.2019.00219
158. Golden LC, Itoh Y, Itoh N, Iyengar S, Coit P, Salama Y, et al. Parent-of-origin differences in DNA methylation of X chromosome genes in T lymphocytes. Proc Natl Acad Sci USA (2019) 16(52):26779–87. doi: 10.1073/pnas.1910072116
159. Arnold AP, Chen X. What does the "four core genotypes" mouse model tell us about sex differences in the brain and other tissues? Front Neuroendocrinol (2009) 30(1):1–9. doi: 10.1016/j.yfrne.2008.11.001
160. Palaszynski KM, Smith DL, Kamrava S, Burgoyne PS, Arnold AP, Voskuhl RR. A yin-yang effect between sex chromosome complement and sex hormones on the immune response. Endocrinology. (2005) 146(8):3280–5. doi: 10.1210/en.2005-0284
161. Smith Bouvier DL, Divekar AA, Sasidhar M, Du S, Tiwari Woodruff SK, King JK, et al. A role for sex chromosome complement in the female bias in autoimmune disease. J Exp Med (2008) 205(5):1099–108. doi: 10.1084/jem.20070850
162. Case LK, Wall EH, Dragon JA, Saligrama N, Krementsov DN, Moussawi M, et al. The y chromosome as a regulatory element shaping immune cell transcriptomes and susceptibility to autoimmune disease. Genome Res (2013) 23(9):1474–85. doi: 10.1101/gr.156703.113
163. Case LK, Wall EH, Osmanski EE, Dragon JA, Saligrama N, Zachary JF, et al. Copy number variation in y chromosome multicopy genes is linked to a paternal parent-of-origin effect on CNS autoimmune disease in female offspring. Genome Biol (2015) 16:28. doi: 10.1186/s13059-015-0591-7
164. Teuscher C, Noubade R, Spach K, McElvany B, Bunn JY, Fillmore PD, et al. Evidence that the y chromosome influences autoimmune disease in male and female mice. Proc Natl Acad Sci U S A. (2006) 103(21):8024–9. doi: 10.1073/pnas.0600536103
165. Bishop CE, Boursot P, Baron B, Bonhomme F, Hatat D. Most classical mus musculus domesticus laboratory mouse strains carry a mus musculus musculus y chromosome. Nature. (1985) 315(6014):70–2. doi: 10.1038/315070a0
166. Bearoff F, Case LK, Krementsov DN, Wall EH, Saligrama N, Blankenhorn EP, et al. Identification of genetic determinants of the sexual dimorphism in CNS autoimmunity. PloS One (2015) 10(2):e0117993. doi: 10.1371/journal.pone.0117993
167. Voskuhl RR, Palaszynski K. Sex hormones in experimental autoimmune encephalomyelitis: implications for multiple sclerosis. Neuroscientist. (2001) 7(3):258–70. doi: 10.1177/107385840100700310
168. Spence RD, Voskuhl RR. Neuroprotective effects of estrogens and androgens in CNS inflammation and neurodegeneration. Front Neuroendocrinol. (2012) 33(1):105–15. doi: 10.1016/j.yfrne.2011.12.001
169. Gold SM, Sasidhar MV, Morales LB, Du S, Sicotte NL, Tiwari Woodruff SK, et al. Estrogen treatment decreases matrix metalloproteinase (MMP)-9 in autoimmune demyelinating disease through estrogen receptor alpha (ERalpha). Lab Invest. (2009) 89(10):1076–83. doi: 10.1038/labinvest.2009.79
170. Khalaj AJ, Hasselmann J, Augello C, Moore S, Tiwari Woodruff SK. Nudging oligodendrocyte intrinsic signaling to remyelinate and repair: Estrogen receptor ligand effects. J Steroid Biochem Mol Biol (2016) 160:43–52. doi: 10.1016/j.jsbmb.2016.01.006
171. Shah NM, Imami N, Johnson MR. Progesterone modulation of pregnancy-related immune responses. Front Immunol (2018) 9:1293. doi: 10.3389/fimmu.2018.01293
172. Zhu X, Frechou M, Schumacher M, Guennoun R. Cerebroprotection by progesterone following ischemic stroke: Multiple effects and role of the neural progesterone receptors. J Steroid Biochem Mol Biol (2019) 185:90–102. doi: 10.1016/j.jsbmb.2018.07.014
173. Borba VV, Zandman Goddard G, Shoenfeld Y. Prolactin and autoimmunity. Front Immunol (2018) 9:73. doi: 10.3389/fimmu.2018.00073
174. Cabrera Reyes EA, Limon Morales O, Rivero Segura NA, Camacho Arroyo I, Cerbon M. Prolactin function and putative expression in the brain. Endocrine. (2017) 57(2):199–213. doi: 10.1007/s12020-017-1346-x
175. Yao G, Shang XJ. A comparison of modulation of proliferation of thymocyte by testosterone, dehydroisoandrosterone and androstenedione in vitro. Arch Androl. (2005) 51(4):257–65. doi: 10.1080/014850190924115
176. Page ST, Plymate SR, Bremner WJ, Matsumoto AM, Hess DL, Lin DW, et al. Effect of medical castration on CD4+ CD25+ T cells, CD8+ T cell IFN-gamma expression, and NK cells: a physiological role for testosterone and/or its metabolites. Am J Physiol Endocrinol Metab (2006) 290(5):E856–63. doi: 10.1152/ajpendo.00484.2005
177. Bielecki B, Mattern C, Ghoumari AM, Javaid S, Smietanka K, Abi Ghanem C, et al. Unexpected central role of the androgen receptor in the spontaneous regeneration of myelin. Proc Natl Acad Sci U S A. (2016) 113(51):14829–34. doi: 10.1073/pnas.1614826113
178. Jaini R, Altuntas CZ, Loya MG, Tuohy VK. Disruption of estrous cycle homeostasis in mice with experimental autoimmune encephalomyelitis. J Neuroimmunology. (2015) 279:71–4. doi: 10.1016/j.jneuroim.2015.01.002
179. Ajayi AF, Akhigbe RE. Staging of the estrous cycle and induction of estrus in experimental rodents: an update. Fertility Res Practice. (2020) 6(1):5. doi: 10.1186/s40738-020-00074-3
180. Lang TJ. Estrogen as an immunomodulator. Clin Immunol (2004) 113(3):224–30. doi: 10.1016/j.clim.2004.05.011
181. Segerer SE, Muller N, van den Brandt J, Kapp M, Dietl J, Reichardt HM, et al. Impact of female sex hormones on the maturation and function of human dendritic cells. Am J Reprod Immunol (2009) 62(3):165–73. doi: 10.1111/j.1600-0897.2009.00726.x
182. Pioli PA, Jensen AL, Weaver LK, Amiel E, Shen Z, Shen L, et al. Estradiol attenuates lipopolysaccharide-induced CXC chemokine ligand 8 production by human peripheral blood monocytes. J Immunol (2007) 179(9):6284–90. doi: 10.4049/jimmunol.179.9.6284
183. Nilsson BO. Modulation of the inflammatory response by estrogens with focus on the endothelium and its interactions with leukocytes. Inflammation Res (2007) 56(7):269–73. doi: 10.1007/s00011-007-6198-z
184. Mo R, Chen J, Grolleau Julius A, Murphy HS, Richardson BC, Yung RL. Estrogen regulates CCR gene expression and function in T lymphocytes. J Immunol (2005) 174(10):6023–9. doi: 10.4049/jimmunol.174.10.6023
185. Tai P, Wang J, Jin H, Song X, Yan J, Kang Y, et al. Induction of regulatory T cells by physiological level estrogen. J Cell Physiol (2008) 214(2):456–64. doi: 10.1002/jcp.21221
186. Somerset DA, Zheng Y, Kilby MD, Sansom DM, Drayson MT. Normal human pregnancy is associated with an elevation in the immune suppressive CD25+ CD4+ regulatory T-cell subset. Immunology. (2004) 112(1):38–43. doi: 10.1111/j.1365-2567.2004.01869.x
187. Roepke TA, Qiu J, Smith AW, Ronnekleiv OK, Kelly MJ. Fasting and 17beta-estradiol differentially modulate the m-current in neuropeptide y neurons. J Neurosci (2011) 31(33):11825–35. doi: 10.1523/JNEUROSCI.1395-11.2011
188. Jansson L, Olsson T, Holmdahl R. Estrogen induces a potent suppression of experimental autoimmune encephalomyelities and collagen-induced arthritis in mice. J Neuroimmunology. (1994) 53(2):203–7. doi: 10.1016/0165-5728(94)90030-2
189. Lelu K, Delpy L, Robert V, Foulon E, Laffont S, Pelletier L, et al. Endogenous estrogens, through estrogen receptor alpha, constrain autoimmune inflammation in female mice by limiting CD4+ T-cell homing into the CNS. Eur J Immunol (2010) 40(12):3489–98. doi: 10.1002/eji.201040678
190. Bebo BF Jr., Zelinka Vincent E, Adamus G, Amundson D, Vandenbark AA, Offner H. Gonadal hormones influence the immune response to PLP 139-151 and the clinical course of relapsing experimental autoimmune encephalomyelitis. J Neuroimmunol. (1998) 84(2):122–30. doi: 10.1016/S0165-5728(97)00214-2
191. Khan D, Ansar Ahmed S. The immune system is a natural target for estrogen action: Opposing effects of estrogen in two prototypical autoimmune diseases. Front Immunol (2016) 6. doi: 10.3389/fimmu.2015.00635
192. Toran Allerand CD. Minireview: A plethora of estrogen receptors in the brain: Where will it end? Endocrinology (2004) 145(3):1069–74. doi: 10.1210/en.2003-1462
193. Kim S, Liva SM, Dalal MA, Verity MA, Voskuhl RR. Estriol ameliorates autoimmune demyelinating disease. Neurology. (1999) 52(6):1230. doi: 10.1212/wnl.52.6.1230
194. Offner H, Adlard K, Zamora A, Vandenbark AA. Estrogen potentiates treatment with T-cell receptor protein of female mice with experimental encephalomyelitis. J Clin Invest. (2000) 105(10):1465–72. doi: 10.1172/JCI9213
195. Ito A, Bebo BF Jr., Matejuk A, Zamora A, Silverman M, Fyfe-Johnson A, et al. Estrogen treatment down-regulates TNF-alpha production and reduces the severity of experimental autoimmune encephalomyelitis in cytokine knockout mice. J Immunol (2001) 167(1):542–52. doi: 10.4049/jimmunol.167.1.542
196. Liu HY, Buenafe AC, Matejuk A, Ito A, Zamora A, Dwyer J, et al. Estrogen inhibition of EAE involves effects on dendritic cell function. J Neurosci Res (2002) 70(2):238–48. doi: 10.1002/jnr.10409
197. Subramanian S, Matejuk A, Zamora A, Vandenbark AA, Offner H. Oral feeding with ethinyl estradiol suppresses and treats experimental autoimmune encephalomyelitis in SJL mice and inhibits the recruitment of inflammatory cells into the central nervous system. J Immunol (2003) 170(3):1548–55. doi: 10.4049/jimmunol.170.3.1548
198. Bebo BF Jr., Dehghani B, Foster S, Kurniawan A, Lopez FJ, Sherman LS. Treatment with selective estrogen receptor modulators regulates myelin specific T-cells and suppresses experimental autoimmune encephalomyelitis. Glia. (2009) 57(7):777–90. doi: 10.1002/glia.20805
199. Wang C, Dehghani B, Li Y, Kaler LJ, Vandenbark AA, Offner H. Oestrogen modulates experimental autoimmune encephalomyelitis and interleukin-17 production via programmed death 1. Immunology. (2009) 126(3):329–35. doi: 10.1111/j.1365-2567.2008.03051.x
200. Nashold FE, Spach KM, Spanier JA, Hayes CE. Estrogen controls vitamin D3-mediated resistance to experimental autoimmune encephalomyelitis by controlling vitamin D3 metabolism and receptor expression. J Immunol (2009) 183(6):3672–81. doi: 10.4049/jimmunol.0901351
201. Giraud SN, Caron CM, Pham Dinh D, Kitabgi P, Nicot AB. Estradiol inhibits ongoing autoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in reactive astrocytes. Proc Natl Acad Sci U S A. (2010) 107(18):8416–21. doi: 10.1073/pnas.0910627107
202. Bodhankar S, Wang C, Vandenbark AA, Offner H. Estrogen-induced protection against experimental autoimmune encephalomyelitis is abrogated in the absence of b cells. Eur J Immunol (2011) 41(4):1165–75. doi: 10.1002/eji.201040992
203. Benedek G, Zhang J, Bodhankar S, Nguyen H, Kent G, Jordan K, et al. Estrogen induces multiple regulatory b cell subtypes and promotes M2 microglia and neuroprotection during experimental autoimmune encephalomyelitis. J Neuroimmunol. (2016) 293:45–53. doi: 10.1016/j.jneuroim.2016.02.009
204. Seifert HA, Benedek G, Nguyen H, Kent G, Vandenbark AA, Offner H. Estrogen protects both sexes against EAE by promoting common regulatory cell subtypes independent of endogenous estrogen. Metab Brain Dis (2017) 32(5):1747–54. doi: 10.1007/s11011-017-0063-8
205. Haghmorad D, Salehipour Z, Nosratabadi R, Rastin M, Kokhaei P, Mahmoudi MB, et al. Medium-dose estrogen ameliorates experimental autoimmune encephalomyelitis in ovariectomized mice. J Immunotoxicol. (2016) 13(6):885–96. doi: 10.1080/1547691X.2016.1223768
206. Polanczyk M, Zamora A, Subramanian S, Matejuk A, Hess DL, Blankenhorn EP, et al. The protective effect of 17beta-estradiol on experimental autoimmune encephalomyelitis is mediated through estrogen receptor-alpha. Am J Pathol (2003) 163(4):1599–605. doi: 10.1016/S0002-9440(10)63516-X
207. Matejuk A, Bakke AC, Hopke C, Dwyer J, Vandenbark AA, Offner H. Estrogen treatment induces a novel population of regulatory cells, which suppresses experimental autoimmune encephalomyelitis. J Neurosci Res (2004) 77(1):119–26. doi: 10.1002/jnr.20145
208. Wang C, Dehghani B, Li Y, Kaler LJ, Proctor T, Vandenbark AA, et al. Membrane estrogen receptor regulates experimental autoimmune encephalomyelitis through up-regulation of programmed death 1. J Immunol (2009) 182(5):3294–303. doi: 10.4049/jimmunol.0803205
209. Li R, Xu W, Chen Y, Qiu W, Shu Y, Wu A, et al. Raloxifene suppresses experimental autoimmune encephalomyelitis and NF-kappaB-dependent CCL20 expression in reactive astrocytes. PloS One (2014) 9(4):e94320. doi: 10.1371/journal.pone.0094320
210. Benagiano M, Bianchi P, D'Elios MM, Brosens I, Benagiano G. Autoimmune diseases: Role of steroid hormones. Best Pract Res Clin Obstet Gynaecol. (2019) 60:24–34. doi: 10.1016/j.bpobgyn.2019.03.001
211. Singh M, Su C. Progesterone and neuroprotection. Horm Behav (2013) 63(2):284–90. doi: 10.1016/j.yhbeh.2012.06.003
212. De Nicola AF, Garay LI, Meyer M, Guennoun R, Sitruk Ware R, Schumacher M, et al. Neurosteroidogenesis and progesterone anti-inflammatory/neuroprotective effects. J Neuroendocrinol (2018) 30:e12502. doi: 10.1111/jne.12502
213. Yates MA, Li Y, Chlebeck P, Proctor T, Vandenbark AA, Offner H. Progesterone treatment reduces disease severity and increases IL-10 in experimental autoimmune encephalomyelitis. J Neuroimmunol. (2010) 220(1-2):136–9. doi: 10.1016/j.jneuroim.2010.01.013
214. Garay L, Gonzalez Deniselle MC, Meyer M, Costa JJ, Lima A, Roig P, et al. Protective effects of progesterone administration on axonal pathology in mice with experimental autoimmune encephalomyelitis. Brain Res (2009) 1283:177–85. doi: 10.1016/j.brainres.2009.04.057
215. Garay L, Gonzalez Deniselle MC, Lima A, Roig P, De Nicola AF. Effects of progesterone in the spinal cord of a mouse model of multiple sclerosis. J Steroid Biochem Mol Biol (2007) 107(3-5):228–37. doi: 10.1016/j.jsbmb.2007.03.040
216. Giatti S, Caruso D, Boraso M, Abbiati F, Ballarini E, Calabrese D, et al. Neuroprotective effects of progesterone in chronic experimental autoimmune encephalomyelitis. J Neuroendocrinol. (2012) 24(6):851–61. doi: 10.1111/j.1365-2826.2012.02284.x
217. Yu HJ, Fei J, Chen XS, Cai QY, Liu HL, Liu GD, et al. Progesterone attenuates neurological behavioral deficits of experimental autoimmune encephalomyelitis through remyelination with nucleus-sublocalized Olig1 protein. Neurosci Lett (2010) 476(1):42–5. doi: 10.1016/j.neulet.2010.03.079
218. Mitchell R, Hollis S, Rothwell C, Robertson WR. Age related changes in the pituitary-testicular axis in normal men; lower serum testosterone results from decreased bioactive LH drive. Clin Endocrinol (Oxf). (1995) 42(5):501–7. doi: 10.1111/j.1365-2265.1995.tb02669.x
219. Davison SL, Bell R, Donath S, Montalto JG, Davis SR. Androgen levels in adult females: changes with age, menopause, and oophorectomy. J Clin Endocrinol Metab (2005) 90(7):3847–53. doi: 10.1210/jc.2005-0212
220. Hince M, Sakkal S, Vlahos K, Dudakov J, Boyd R, Chidgey A. The role of sex steroids and gonadectomy in the control of thymic involution. Cell Immunol (2008) 252(1-2):122–38. doi: 10.1016/j.cellimm.2007.10.007
221. Massa MG, David C, Jorg S, Berg J, Gisevius B, Hirschberg S, et al. Testosterone differentially affects T cells and neurons in murine and human models of neuroinflammation and neurodegeneration. Am J Pathol (2017) 187(7):1613–22. doi: 10.1016/j.ajpath.2017.03.006
222. Palaszynski KM, Loo KK, Ashouri JF, Liu HB, Voskuhl RR. Androgens are protective in experimental autoimmune encephalomyelitis: implications for multiple sclerosis. J Neuroimmunol. (2004) 146(1-2):144–52. doi: 10.1016/j.jneuroim.2003.11.004
223. Milosevic A, Bjelobaba I, Bozic ID, Lavrnja I, Savic D, Tesovic K, et al. Testicular steroidogenesis is suppressed during experimental autoimmune encephalomyelitis in rats. Sci Rep (2021) 11(1):8996. doi: 10.1038/s41598-021-88305-5
224. Bebo BF Jr., Schuster JC, Vandenbark AA, Offner H. Androgens alter the cytokine profile and reduce encephalitogenicity of myelin-reactive T cells. J Immunol (1999) 162(1):35–40.
225. Offner H, Zamora A, Drought H, Matejuk A, Auci DL, Morgan EE, et al. A synthetic androstene derivative and a natural androstene metabolite inhibit relapsing-remitting EAE. J Neuroimmunol. (2002) 130(1-2):128–39. doi: 10.1016/S0165-5728(02)00214-X
226. Dalal M, Kim S, Voskuhl RR. Testosterone therapy ameliorates experimental autoimmune encephalomyelitis and induces a T helper 2 bias in the autoantigen-specific T lymphocyte response. J Immunol (1997) 159(1):3–6.
227. Chuang E, Molitch ME. Prolactin and autoimmune diseases in humans. Acta Biomed (2007) 78 Suppl 1:255–61.
228. Russell DH. New aspects of prolactin and immunity: a lymphocyte-derived prolactin-like product and nuclear protein kinase c activation. Trends Pharmacol Sci (1989) 10(1):40–4. doi: 10.1016/0165-6147(89)90106-5
229. Carreno PC, Sacedon R, Jimenez E, Vicente A, Zapata AG. Prolactin affects both survival and differentiation of T-cell progenitors. J Neuroimmunol (2005) 160(1-2):135–45. doi: 10.1016/j.jneuroim.2004.11.008
230. Orbach H, Shoenfeld Y. Hyperprolactinemia and autoimmune diseases. Autoimmun Rev (2007) 6(8):537–42. doi: 10.1016/j.autrev.2006.10.005
231. Riskind PN, Massacesi L, Doolittle TH, Hauser SL. The role of prolactin in autoimmune demyelination: suppression of experimental allergic encephalomyelitis by bromocriptine. Ann Neurol (1991) 29(5):542–7. doi: 10.1002/ana.410290514
232. Forsum E, Lof M. Energy metabolism during human pregnancy. Annu Rev Nutr (2007) 27:277–92. doi: 10.1146/annurev.nutr.27.061406.093543
233. Granger JP. Maternal and fetal adaptations during pregnancy: lessons in regulatory and integrative physiology. Am J Physiol Regul Integr Comp Physiol (2002) 283(6):R1289–92. doi: 10.1152/ajpregu.00562.2002
234. Lain KY, Catalano PM. Metabolic changes in pregnancy. Clin Obstet Gynecol. (2007) 50(4):938–48. doi: 10.1097/GRF.0b013e31815a5494
235. Tan EK, Tan EL. Alterations in physiology and anatomy during pregnancy. Best Pract Res Clin Obstet Gynaecol. (2013) 27(6):791–802. doi: 10.1016/j.bpobgyn.2013.08.001
236. Piccinni MP, Lombardelli L, Logiodice F, Kullolli O, Parronchi P, Romagnani S. How pregnancy can affect autoimmune diseases progression? Clin Mol Allergy (2016) 14:11. doi: 10.1186/s12948-016-0048-x
237. Saito S, Nakashima A, Shima T, Ito M. Th1/Th2/Th17 and regulatory T-cell paradigm in pregnancy. Am J Reprod Immunol (2010) 63(6):601–10. doi: 10.1111/j.1600-0897.2010.00852.x
238. Ysrraelit MC, Correale J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology. (2019) 156(1):9–22. doi: 10.1111/imm.13004
239. Morton H, Rolfe B, Clunie GJ. An early pregnancy factor detected in human serum by the rosette inhibition test. Lancet (1977) 1(8008):394–7. doi: 10.1016/S0140-6736(77)92605-8
240. Brunton PJ, Russell JA. Endocrine induced changes in brain function during pregnancy. Brain Res (2010) 1364:198–215. doi: 10.1016/j.brainres.2010.09.062
241. Molvarec A, Szarka A, Walentin S, Beko G, Karadi I, Prohaszka Z, et al. Serum leptin levels in relation to circulating cytokines, chemokines, adhesion molecules and angiogenic factors in normal pregnancy and preeclampsia. Reprod Biol Endocrinol (2011) 9:124. doi: 10.1186/1477-7827-9-124
242. Lauszus FF, Klebe JG, Bek T, Flyvbjerg A. Increased serum IGF-I during pregnancy is associated with progression of diabetic retinopathy. Diabetes. (2003) 52(3):852–6. doi: 10.2337/diabetes.52.3.852
243. Uvnas-Moberg K, Widstrom AM, Werner S, Matthiesen AS, Winberg J. Oxytocin and prolactin levels in breast-feeding women. correlation with milk yield and duration of breast-feeding. Acta Obstet Gynecol Scand (1990) 69(4):301–6. doi: 10.3109/00016349009036151
244. Munoz-Suano A, Hamilton AB, Betz AG. Gimme shelter: the immune system during pregnancy. Immunol Rev (2011) 241(1):20–38. doi: 10.1111/j.1600-065X.2011.01002.x
245. Veenstra van Nieuwenhoven AL, Heineman MJ, Faas MM. The immunology of successful pregnancy. Hum Reprod Update. (2003) 9(4):347–57. doi: 10.1093/humupd/dmg026
246. Yip L, McCluskey J, Sinclair R. Immunological aspects of pregnancy. Clin Dermatol (2006) 24(2):84–7. doi: 10.1016/j.clindermatol.2005.10.022
247. Doria A, Iaccarino L, Arienti S, Ghirardello A, Zampieri S, Rampudda ME, et al. Th2 immune deviation induced by pregnancy: the two faces of autoimmune rheumatic diseases. Reprod Toxicol (2006) 22(2):234–41. doi: 10.1016/j.reprotox.2006.04.001
248. Sanchez Ramon S, Navarro AJ, Aristimuno C, Rodriguez Mahou M, Bellon JM, Fernandez Cruz E, et al. Pregnancy-induced expansion of regulatory T-lymphocytes may mediate protection to multiple sclerosis activity. Immunol Lett (2005) 96(2):195–201. doi: 10.1016/j.imlet.2004.09.004
249. Ruocco MG, Chaouat G, Florez L, Bensussan A, Klatzmann D. Regulatory T-cells in pregnancy: historical perspective, state of the art, and burning questions. Front Immunol (2014) 5:389. doi: 10.3389/fimmu.2014.00389
250. Tilburgs T, Scherjon SA, van der Mast BJ, Haasnoot GW, Versteeg VDV MM, Roelen DL, et al. Fetal-maternal HLA-c mismatch is associated with decidual T cell activation and induction of functional T regulatory cells. J Reprod Immunol (2009) 82(2):148–57. doi: 10.1016/j.jri.2009.05.003
251. Tsuda S, Zhang X, Hamana H, Shima T, Ushijima A, Tsuda K, et al. Clonally expanded decidual effector regulatory T cells increase in late gestation of normal pregnancy, but not in preeclampsia, in humans. Front Immunol (2018) 9:1934. doi: 10.3389/fimmu.2018.01934
252. Aluvihare VR, Kallikourdis M, Betz AG. Regulatory T cells mediate maternal tolerance to the fetus. Nat Immunol (2004) 5(3):266–71. doi: 10.1038/ni1037
253. Polanczyk MJ, Hopke C, Huan J, Vandenbark AA, Offner H. Enhanced FoxP3 expression and treg cell function in pregnant and estrogen-treated mice. J Neuroimmunol (2005) 170(1-2):85–92. doi: 10.1016/j.jneuroim.2005.08.023
254. Jamieson DJ, Theiler RN, Rasmussen SA. Emerging infections and pregnancy. Emerg Infect Dis (2006) 12(11):1638–43. doi: 10.3201/eid1211.060152
255. Fettke F, Schumacher A, Canellada A, Toledo N, Bekeredjian Ding I, Bondt A, et al. Maternal and fetal mechanisms of b cell regulation during pregnancy: Human chorionic gonadotropin stimulates b cells to produce IL-10 while alpha-fetoprotein drives them into apoptosis. Front Immunol (2016) 7:495. doi: 10.3389/fimmu.2016.00495
256. Keith AB. Effect of pregnancy on experimental allergic encephalomyelitis in guinea pigs and rats. J Neurol Sci (1978) 38(3):317–26. doi: 10.1016/0022-510X(78)90138-7
257. Mertin LA, Rumjanek VM. Pregnancy and the susceptibility of Lewis rats to experimental allergic encephalomyelitis. J Neurol Sci (1985) 68(1):15–24. doi: 10.1016/0022-510X(85)90046-2
258. Barac Latas V, Muhvic D, Radosevic Stabic B. The influence of pregnancy on development and course of chronic relapsing experimental autoimmune encephalomyelitis in rats: implications for multiple sclerosis. Coll Antropol. (2010) 34 Suppl 1:267–71.
259. Evron S, Brenner T, Abramsky O. Suppressive effect of pregnancy on the development of experimental allergic encephalomyelitis in rabbits. Am J Reprod Immunol (1984) 5(3):109–13. doi: 10.1111/j.1600-0897.1984.tb00298.x
260. Harness J, McCombe PA. The effects of pregnancy on myelin basic protein-induced experimental autoimmune encephalomyelitis in Lewis rats: suppression of clinical disease, modulation of cytokine expression in the spinal cord inflammatory infiltrate and suppression of lymphocyte proliferation by pregnancy sera. Am J Reprod Immunol (2001) 46(6):405–12. doi: 10.1034/j.1600-0897.2001.d01-32.x
261. Langer Gould A, Garren H, Slansky A, Ruiz PJ, Steinman L. Late pregnancy suppresses relapses in experimental autoimmune encephalomyelitis: evidence for a suppressive pregnancy-related serum factor. J Immunol (2002) 169(2):1084–91. doi: 10.4049/jimmunol.169.2.1084
262. McClain MA, Gatson NN, Powell ND, Papenfuss TL, Gienapp IE, Song F, et al. Pregnancy suppresses experimental autoimmune encephalomyelitis through immunoregulatory cytokine production. J Immunol (2007) 179(12):8146–52. doi: 10.4049/jimmunol.179.12.8146
263. Gatson NN, Williams JL, Powell ND, McClain MA, Hennon TR, Robbins PD, et al. Induction of pregnancy during established EAE halts progression of CNS autoimmune injury via pregnancy-specific serum factors. J Neuroimmunol. (2011) 230(1-2):105–13. doi: 10.1016/j.jneuroim.2010.09.010
264. Song F, Gienapp I, Shawler T, Williams J, Smith K, Kithcart A, et al. Pregnancy modulates adoptively transferred experimental autoimmune encephalomyelitis (EAE). FASEB J (2008) 22(S1):853. 16-.16. doi: 10.1096/fasebj.22.1_supplement.853.16
265. Brenner T, Ovadia H, Evron S, Mizrachi R, Abramsky O. Experimental allergic encephalomyelitis: passive transfer of resistance during lactation. J Neuroimmunol. (1986) 12(4):317–27. doi: 10.1016/0165-5728(86)90038-X
266. Williams JL, Gatson NN, Smith KM, Almad A, McTigue DM, Whitacre CC. Serum exosomes in pregnancy-associated immune modulation and neuroprotection during CNS autoimmunity. Clin Immunol (2013) 149(2):236–43. doi: 10.1016/j.clim.2013.04.005
267. Engler JB, Heckmann NF, Jager J, Gold SM, Friese MA. Pregnancy enables expansion of disease-specific regulatory T cells in an animal model of multiple sclerosis. J Immunol (2019) 203(7):1743–52. doi: 10.4049/jimmunol.1900611
268. Morton H, Hegh V, Clunie GJA. Immunosuppression detected in pregnant mice by rosette inhibition test. Nature. (1974) 249(5456):459–60. doi: 10.1038/249459a0
269. Cavanagh AC, Morton H. The purification of early-pregnancy factor to homogeneity from human platelets and identification as chaperonin 10. Eur J Biochem (1994) 222(2):551–60. doi: 10.1111/j.1432-1033.1994.tb18897.x
270. Cavanagh AC. Identification of early pregnancy factor as chaperonin 10: implications for understanding its role. Rev Reprod (1996) 1(1):28–32. doi: 10.1530/ror.0.0010028
271. Morton H, Cavanagh AC, Athanasas Platsis S, Quinn KA, Rolfe BE. Early pregnancy factor has immunosuppressive and growth factor properties. Reprod Fertil Dev (1992) 4(4):411–22. doi: 10.1071/RD9920411
272. McCombe PA. Recombinant EPF/chaperonin 10 promotes the survival of O4-positive pro-oligodendrocytes prepared from neonatal rat brain. Cell Stress Chaperones. (2008) 13(4):467–74. doi: 10.1007/s12192-008-0045-1
273. Athanasas Platsis S, Zhang B, Hillyard NC, Cavanagh AC, Csurhes PA, Morton H, et al. Early pregnancy factor suppresses the infiltration of lymphocytes and macrophages in the spinal cord of rats during experimental autoimmune encephalomyelitis but has no effect on apoptosis. J Neurol Sci (2003) 214(1-2):27–36. doi: 10.1016/S0022-510X(03)00170-9
274. Harness J, Cavanagh A, Morton H, McCombe P. A protective effect of early pregnancy factor on experimental autoimmune encephalomyelitis induced in Lewis rats by inoculation with myelin basic protein. J Neurol Sci (2003) 216(1):33–41. doi: 10.1016/S0022-510X(03)00212-0
275. Zhang B, Harness J, Somodevilla Torres MJ, Hillyard NC, Mould AW, Alewood D, et al. Early pregnancy factor suppresses experimental autoimmune encephalomyelitis induced in Lewis rats with myelin basic protein and in SJL/J mice with myelin proteolipid protein peptide 139-151. J Neurol Sci (2000) 182(1):5–15. doi: 10.1016/S0022-510X(00)00432-9
276. Zhang B, Walsh MD, Nguyen KB, Hillyard NC, Cavanagh AC, McCombe PA, et al. Early pregnancy factor treatment suppresses the inflammatory response and adhesion molecule expression in the spinal cord of SJL/J mice with experimental autoimmune encephalomyelitis and the delayed-type hypersensitivity reaction to trinitrochlorobenzene in normal BALB/c mice. J Neurol Sci (2003) 212(1-2):37–46. doi: 10.1016/S0022-510X(03)00103-5
277. Zamorina SA, Litvinova LS, Yurova KA, Khaziakhmatova OG, Timganova VP, Bochkova MS, et al. Role of alpha-fetoprotein in regulation of proliferation and functional activity of naive T cells and immune memory T cells. Bull Exp Biol Med (2019) 167(4):470–4. doi: 10.1007/s10517-019-04552-7
278. Abramsky O, Brenner T, Mizrachi R, Soffer D. Alpha-fetoprotein suppresses experimental allergic encephalomyelitis. J Neuroimmunol. (1982) 2(1):1–7. doi: 10.1016/0165-5728(82)90070-4
279. Brenner T, Evron S, Soffer D, Abramsky O. Treatment of experimental allergic encephalomyelitis in rabbits with alpha-fetoprotein. Isr J Med Sci (1985) 21(12):945–9.
280. Irony Tur Sinai M, Grigoriadis N, Tsiantoulas D, Touloumi O, Abramsky O, Brenner T. Immunomodulation of EAE by alpha-fetoprotein involves elevation of immune cell apoptosis markers and the transcription factor FoxP3. J Neurol Sci (2009) 279(1-2):80–7. doi: 10.1016/j.jns.2008.12.014
281. Bazer FW, Johnson HM. Type I conceptus interferons: maternal recognition of pregnancy signals and potential therapeutic agents. Am J Reprod Immunol (1991) 26(1):19–22. doi: 10.1111/j.1600-0897.1991.tb00696.x
282. Soos JM, Mujtaba MG, Subramaniam PS, Streit WJ, Johnson HM. Oral feeding of interferon tau can prevent the acute and chronic relapsing forms of experimental allergic encephalomyelitis. J Neuroimmunol. (1997) 75(1-2):43–50. doi: 10.1016/S0165-5728(97)00003-9
283. Mujtaba MG, Streit WJ, Johnson HM. IFN-tau suppresses both the autoreactive humoral and cellular immune responses and induces stable remission in mice with chronic experimental allergic encephalomyelitis. Cell Immunol (1998) 186(2):94–102. doi: 10.1006/cimm.1998.1300
284. Bilbao D, Luciani L, Johannesson B, Piszczek A, Rosenthal N. Insulin-like growth factor-1 stimulates regulatory T cells and suppresses autoimmune disease. EMBO Mol Med (2014) 6(11):1423–35. doi: 10.15252/emmm.201303376
285. Liu X, Yao DL, Webster H. Insulin-like growth factor I treatment reduces clinical deficits and lesion severity in acute demyelinating experimental autoimmune encephalomyelitis. Mult Scler. (1995) 1(1):2–9. doi: 10.1177/135245859500100102
286. Barnea ER, Almogi-Hazan O, Or R, Mueller M, Ria F, Weiss L, et al. Immune regulatory and neuroprotective properties of preimplantation factor: From newborn to adult. Pharmacol Ther (2015) 156:10–25. doi: 10.1016/j.pharmthera.2015.10.008
287. Mueller M, Schoeberlein A, Zhou J, Joerger Messerli M, Oppliger B, Reinhart U, et al. PreImplantation factor bolsters neuroprotection via modulating protein kinase a and protein kinase c signaling. Cell Death Differ (2015) 22(12):2078–86. doi: 10.1038/cdd.2015.55
Keywords: experimental autoimmune encephalomyelitis, biological sex, pregnancy, sex chromosomes, sex hormones, central nervous system
Citation: McCombe PA and Greer JM (2022) Effects of biological sex and pregnancy in experimental autoimmune encephalomyelitis: It’s complicated. Front. Immunol. 13:1059833. doi: 10.3389/fimmu.2022.1059833
Received: 02 October 2022; Accepted: 03 November 2022;
Published: 28 November 2022.
Edited by:
Trine N. Jorgensen, Case Western Reserve University, United StatesReviewed by:
Melissa Anne Cunningham, Medical University of South Carolina, United StatesCopyright © 2022 McCombe and Greer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Judith M. Greer, ai5ncmVlckB1cS5lZHUuYXU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.