
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 06 January 2023
Sec. Mucosal Immunity
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.1054119
This article is part of the Research TopicBridging the gap between immunology, virology, genetics, and epigenetics in bronchiolitis: The Multiomics pathway to asthma developmentView all 10 articles
 Zihan Dong1,2†
Zihan Dong1,2† Åsne Myklebust3,4†
Åsne Myklebust3,4† Ingvild Bjellmo Johnsen4
Ingvild Bjellmo Johnsen4 Tuomas Jartti5,6,7
Tuomas Jartti5,6,7 Henrik Døllner3,4
Henrik Døllner3,4 Kari Risnes3,4*‡
Kari Risnes3,4*‡ Andrew T. DeWan2,8*‡
Andrew T. DeWan2,8*‡Background: Genome-wide association studies of asthma have identified associations with variants in type-2 related genes. Also, specific interactions between genetic variants and viral bronchiolitis in the development of asthma has been suggested.
Objective: To conduct a gene-based analysis of genetic variants in type 2 cytokine related genes as risk factors for allergic asthma at school age, and further, to study their interaction with specific viral infections in early childhood.
Methods: A prospectively investigated cohort of children with previous bronchiolitis and controls came for follow-up at school age. The research visit, blinded to viral exposure, included detailed lung function tests, laboratory investigation, and questionnaires. Allergic asthma was defined as typical symptoms plus objective variable airway obstruction, in addition to laboratory verified atopy (elevated eosinophil count or sensitization to an allergen). Targeted and complete sequencing was performed for nine type 2 cytokine candidate genes: IL4, 5, 13, 25, 33 and 37, IL17RB, CRLF2 and TSLP.
Results: At follow-up, there were 109 children with genetic data, 91 with a history of bronchiolitis (46% respiratory syncytial virus, 24% human rhinovirus, 15% human metapneumovirus and 14% mixed viral etiology) and 18 without. The median age was 9.4 years (range 6-13) and 41 (38%) had laboratory verified atopy. Twenty-one children (19%) met the definition of allergic asthma. After adjusting for age, sex and five viral categories, IL33 achieved nominal significance (p = 0.017) for a positive association with allergic asthma development. In the gene-virus interaction analysis, the variant set in IL17RB demonstrated a nominally significant positive interaction with human metapneumovirus infection (p=0.05).
Conclusion: The results highlight the multifactorial nature of allergic asthma risk, with both viral infection and inherited genetic variants contributing to increasing risk. Results for IL33 and IL17RB were nominally significant and are potential candidate targets for designing therapeutics and early screening, but these results must be replicated in an independent study.
Asthma is the most common chronic disease among children, and is characterized by airway hyperresponsiveness, mucus hypersecretion, and an exaggerated inflammatory response usually dominated by type 2 cytokines (1). The possible role of inherited genetic risk variants related to type 2 cytokines and asthma development following viral bronchiolitis has not been considered in previous studies. There is also evidence that virus-induced bronchiolitis is associated with recurrent wheezing and asthma, particularly rhinovirus, that has been recognized as an important risk factor for asthma (2–4).
IL33 and thymic stromal lymphopoietin (TSLP) are cytokines functioning as alarmins that are released by the airway epithelium in response to viruses, allergens and other triggers and drive type 2 responses in asthma. IL33 and TSLP both have synergistic effects on type 2 cytokines production and eosinophil levels (5, 6). TSLP and IL33 may be important new targets individualized for asthma treatment, and antibody-based drugs for these pathways are being developed (7, 8). Tezepelumab is an anti-TSLP human monoclonal antibody that normalizes type 2 cytokine levels (9) and in clinical studies significantly reduced exacerbation rates vs placebo in patients with severe, uncontrolled asthma (10). Astegolimab is an anti-ST2 antibody that blocks IL33 signaling and has shown promise in reducing asthma exacerbations (11). Airway epithelial cells (AECs) from humans with asthma have increased TSLP mRNA levels (12), and over-expression of TSLP induces experimental asthma in mice (13). Rhinovirus (RV) infection in young children was found to be associated with elevated levels of TSLP (14). Further, respiratory syncytial virus (RSV) and metapneumovirus (MPV) infection induce production of TSLP in AECs in vitro and in vivo, respectively (15, 16).
IL25 also triggers expression of type 2 cytokines, activates type 2 innate lymphoid cells (ILC2 cells) and induces allergic asthma, all of which is dependent on its receptor, IL17RB (17–19). As with TSLP, overexpression of IL25 can induce asthma-like disease in mice (20, 21). Further, RV infected epithelial cells cultured from asthmatic patients have a higher capacity to produce IL25 (22). Using an established mouse model of allergic airway inflammation, infection with rhinovirus led to an increased production of IL25 as well as increased infiltration of eosinophils, neutrophils and basophils, secretion of mucus and production of type 2 cytokines. Importantly, these effects were neutralized if the animals were treated with an antibody directed against IL17RB (22).
While genome-wide association studies of asthma have identified associations with variants in genes that encode cytokines, cytokine receptors and related proteins [e.g. IL13, IL4, IL33, IL1RL1, IL1RL2, IL18R1, TSLP; see Demanais et al. (23)], an examination of genetic variants related to excessive type 2 cytokine production and their interactions with viral infections may shed insight into specific asthma mechanisms. Here we conduct a gene-based analysis of variants in type 2 cytokine related genes (i.e. IL4, 5, 13, 25, 33 and 37, IL17RB, CRLF2 and TSLP) as risk factors for allergic asthma at school age and study their interaction with specific viral infections in early childhood (Table 1). We hypothesized that variants in type 2 cytokine related genes are associated with the susceptibility to virus-induced early wheezing and development of allergic asthma at school age, and that such associations may show interactions with specific viral infections.

Table 1 Description of the selected candidate genes and their role in type 2 responses.
The study population was recruited from a prospective, regional, population-based surveillance cohort of children admitted for airway infections (The St Olav Hospital Airway Project Cohort) (29–31). Eligible participants for the present follow-up study had initially been hospitalized between 2006 and 2012 and had nasopharyngeal secretion (NPS) routinely tested with in-house polymerase chain reaction (PCR) for 17 viruses. The inclusion criteria for the follow-up study were lower respiratory tract infection before 2 years of life with referral to pediatric specialist care, or children from the original control group consisting of surgery patients and without lower viral infection exposure (see Main Effects section for details on how viral infections were classified). All journals were revised after the research visit, and two subjects were excluded because of bacterial pneumonia at exposure.
The study was approved by the Regional Committee on Medical Research Ethics (REK number 2016/540). Informed consent for the clinical follow-up at school age was collected and included consent for the genetic analyses.
The data from the hospitalization was prospectively collected (29–31). The follow up after 6 years of life was systematically set up for research and was not part of routine practice. Examination of the children and data collection took place at the Research Facility Ward at St Olav Hospital between March 2017 and June 2019 and was led by a pediatric asthma specialist together with a trained research nurse, both blinded for viral exposure. The research visit included lung function testing, blood sample draw after local anesthesia, a systematic medical history and clinical exam by the study pediatrician. A digital questionnaire for care givers was based on The International Study of Asthma and Allergies in Childhood Questionnaires (ISAAC) (32) related to history of asthma and atopy in childhood.
Lung function was measured according to established guidelines (33, 34) using a Vyntus Pneumo APS spirometer. The results were obtained both as absolute values and in % of predicted values according to EU standard data for age, height and sex. After initial baseline flow-volume spirometry attempts all children were considered for a methacholine provocation test (MPT) to measure eventual bronchial hyper-reactivity. Contraindications for the MPT was any airway infection, ongoing antibiotic treatment or asthma exacerbation during the last two weeks, FEV1 <70%, clinical signs of airway obstruction, technical inability to perform reproducible and repeated spirometries. The MPT test was performed with an inhalation-synchronized dosimetry nebulizer. The test procedure implicated doubling of doses of methacholine administered until a 20% fall in FEV1 or to a maximum cumulative dose of 1,447 mg methacholine. All children concluded the lung function reversibility testing with a spirometry after inhalation of salbutamol 0.4 mg (Ventoline) through a spacer (Optichamber Diamond by Philips Respironics).
The allergic asthma diagnosis required both a clinical diagnosis of asthma based on recent guidelines and one of the laboratory criteria (35). Clinical asthma was diagnosed if either 1) presence of one or more typical asthma symptoms in history plus variable expiratory airflow limitation at study visit or 2) recent asthma diagnosed by a pediatric specialist and ongoing treatment with inhaled corticosteroids at research visit. Asthma symptoms were defined as cough at night or prolonged cough > 14 days during viral infections, exercise induced either chest tightness, wheeze or shortness of breath, or recognition of these symptoms during methacholine testing when inducing airflow limitation. Variable airflow expiratory limitation, or reversibility, was defined as either a fall in FEV1 of ≥ 20% during the metacholine challenge, or for those who only performed spirometry an increase of FEV1 ≥ 12% compared to the baseline spirometry after salbutamol inhalation. The laboratory criteria were defined as either allergic sensitization (specific IgE > 0.35 kU/L to any of the tested 11 aeroallergens or 6 food allergens) or blood eosinophil count≥300 cells/µL.
Sequence data was generated for 111 of these subjects. Sequencing was done for nine candidate genes (i.e. IL4, 5, 13, 25, 33 and 37, IL17RB, CRLF2 and TSLP) to capture all exons, intron/exon boundaries and up- and down-stream regulatory regions. Library prep was conducted using AmpliSeq for Illumina custom DNA panel (Illumina Inc., CA), which contained 203 amplicons (in two pools) with average amplicon length of 244 bp and covered in total 28,546 base pairs (27,076 covered bases). In brief, 5 ng DNA was used as input in two target PCR amplification reactions (one of each amplicon pool) of 18 cycles (99°C for 15 sec and 60°C for 4 minutes). Secondly, FuPa reagent was used to digest primer dimers and partially digest amplicons. In the next step dual sample indexes were added. The amplicon products were then cleaned up by using Mag-Bind® TotalPure NGS (Omega Bio-tek, Inc., Georgia, GA, USA) to purify the specific amplicons away from free primers and primer dimer species. A second amplification step which amplifies the libraries to ensure sufficient quantity for sequencing on the Illumina platform were then performed, prior to a second clean up by using Mag-Bind® TotalPure NGS (Omega Bio-tek, Inc., Georgia, GA, USA). Finally, a validation of the libraries was performed using an Agilent High Sensitivity DNA Kit on a Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA).
Libraries were normalized and pooled to 8 pM and subjected to clustering on two MiSeq V3 flowcell. Finally, paired end read sequencing was performed for 2X150 cycles on a MiSeq instrument (Illumina, Inc. San Diego, CA, USA), according to the ‘manufacturer’s instructions. Base calling was done on the MiSeq instrument by RTA v1.18.54. FASTQ files were generated using bcl2fastq2 conversion software v2.17 (Illumina, Inc. San Diego, CA, USA).
Sequencing adapters and low-quality reads were removed using Atropos (v. 1.1.18, flags -q 15 and -m 15). The processed reads were aligned against the human genome (hg38) using BWA MEM (v. 0.7.15, flags -M and -v 2). The BAM files were sorted and read group (RG) information was added, followed by cleaning, soft-clipping, and marking of duplicate reads using AddOrReplaceReadGroups, CleanSam, and MarkDuplicates (OPTICAL_DUPLICATE_PIXEL_DISTANCE = 2500, CREATE_INDEX = TRUE) from Picard tools (v. 2.18.5). Base quality scores were recalibrated using GATK’s (v. 4.0.4.0) BaseRecalibrator and ApplyBQSR based on known sites of variation (dbSNP v. 151 and Mills and 1000G gold standard from GATK’s resource bundle). Variants were then called for each sample separately using GATK’s HaplotypeCaller, before merging all resulting gvcf files with GATK’s CombineGVCFs and calling variants on the joint file using GATK’s GenotypeGVCFs (flag -G StandardAnnotation). Called SNVs and indels were processed separately to filter high quality variants using ‘GATK’s VariantFiltration, keeping SNVs with QD < 2.0, FS > 60.0, MQ < 40.0, MQRankSum < -12.5, and ReadPosRankSum < -8.0, and indels with QD < 2.0, FS > 200.0, and ReadPosRankSum < -20.0.
Following variant calling and initial quality control, we examined the per subject call rate to determine if there were any subjects with an indication of low-quality data across all variants. All included subjects (N=109) had a call rate across the 350 variants > 85% and therefore all were included in the analysis.
Among all variants identified by sequencing, 350 were annotated to the nine genes by UCSC GTEx Gene V8 (within 20kb of the gene start and stop site). We excluded variants with call rate < 90% (N=41) and then with a Hardy–Weinberg equilibrium P-values < 10-4 (N=5) leaving 304 variants for analysis. The included variants were annotated by ANNOVAR (36) for functional annotation (Supplementary Table S1).
Our main analysis examined the association with allergic asthma case/controls status. Gene-based analysis integrates information across all included SNPs within one gene or genomic region to improve the statistical power of the association analysis. The gene-based analysis was conducted using SKAT (37). with all parameters being default values. When the sample size is less than 2000, SKAT automatically applies small sample size adjustment. Missing genotype values are imputed based on Hardy-Weinberg equilibrium for binary outcomes. Covariates include age and sex. Viral infections were classified into five categories: 1) sole RSV infection; 2) sole RV infection; 3) sole MPV infection or in combination with viruses other than RSV, RV or bocavirus (BoV); 4) mixed infection (co-infections including RSV, RV, MPV or BoV and any other virus); 5) no viral infection. Dummy coding was utilized to adjust for viral infections in the analysis. Study-wise statistical significance was determined by Bonferroni correction for the nine genes at alpha = 0.05 (p<0.05/9 = 0.0056) and nominal significance was considered at p <= 0.05.
SNP-level association tests were conducted using command “SKATBinary_Single” and p-values were computed. We ran logistic regression to calculate the direction of effect of each SNP.
Given the small sample size, we conducted several sensitivity analyses to test the robustness of our results to various parameters: 1) we applied the SKAT-O (38) method instead of SKAT since SKAT-O combines both the variance components and burden tests; 2) we analyzed only low-frequency variants with a minor MAF < 5% and 3) instead of dummy-coding the five indicator variables representing each viral category and adjusting for each, we merged this into one dichotomous covariate which indicates whether (viral categories 1-4) or not (viral category 5) an individual had been exposed to viral bronchiolitis.
We tested the interaction between the specific virus types, rhinovirus, RSV and MPV, and each of the nine gene-based variant sets using the R package “iSKAT” (39). We utilized the same categorization for rhinovirus, RSV, MPV described previously and adjustment for the other virus categories not contained in the interaction was done using dummy coding. In addition, age and sex were included as covariates in each model. All parameters were default values.
There were 135 participants who came to the follow-up visit, 109 of these children were available for genetic analysis. Table 2 shows their clinical characteristics. The age at follow-up ranged from 6 – 13 years and there were 41 females (38%). There were 42 from the RSV group, 22 from the RV group, 14 from the MPV group, 13 from the mixed infection group and 18 with no bronchiolitis exposure. Twenty-one children (19%) met the definition for allergic asthma, all of them corresponded to mild asthma. As expected children with RV bronchiolitis exposure were more likely to have allergic asthma (29%) than not (18%), the opposite was seen for the RSV group (19% with allergic asthma vs 43% without).
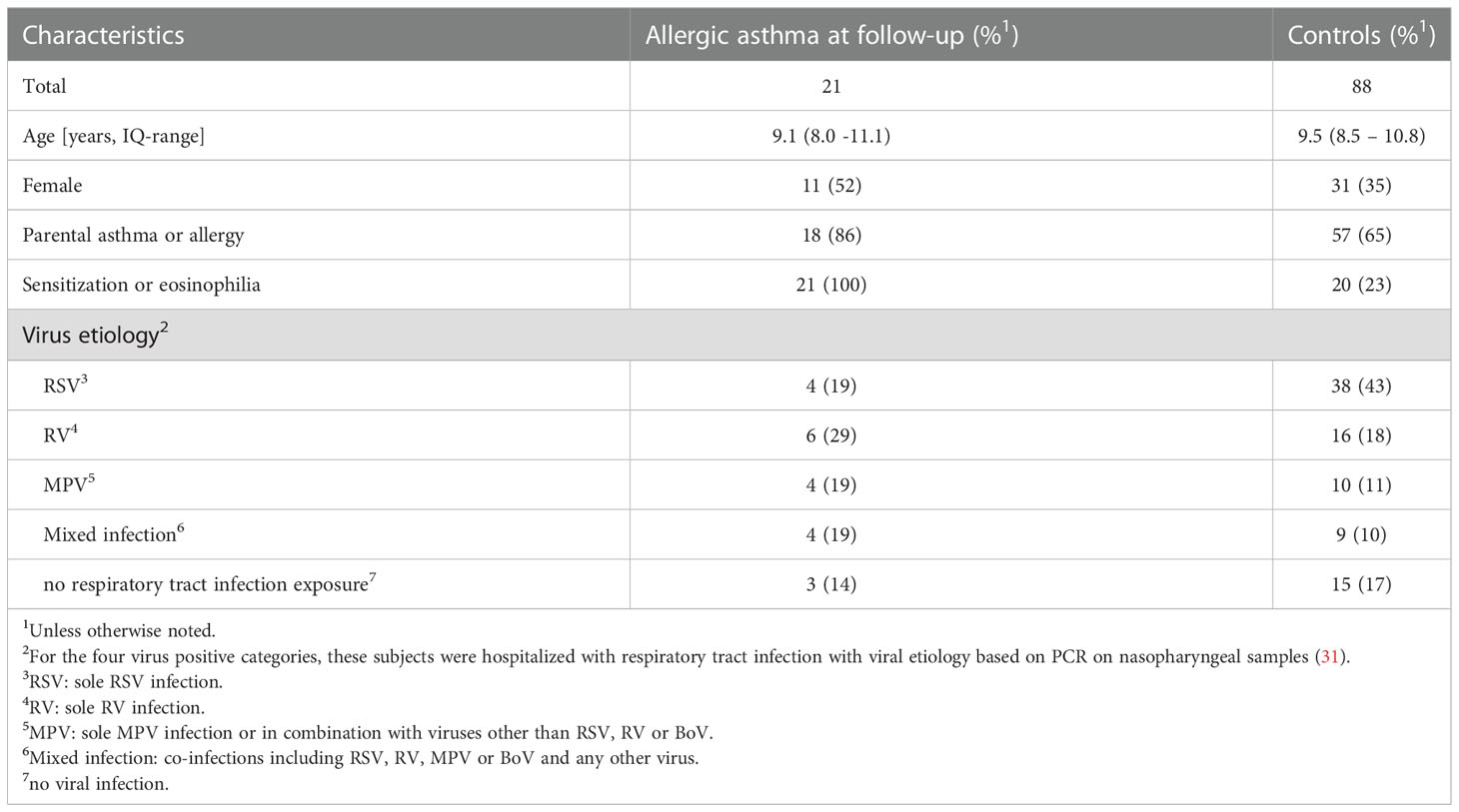
Table 2 Clinical characteristics study subjects.
Targeted sequencing of nine candidate genes was performed to identify all potential variants in each gene (N=350 variants with 304 that passed quality control [see Methods]). Using all coding variants annotated by ANNOVAR (36) to each of the nine genes, aggregate gene-based analyses were conducted for each of the variant sets. We examined the association between each of the nine gene-based aggregate of variants with allergic asthma case/controls status. After adjusting for age, sex and the five viral categories, IL33 achieved nominal significance (P = 0.017, Table 3). Among the samples, IL33 contained 37 coding variants, but none of the single variants achieved statistical significance after adjusting for all 304 variants but five achieved nominal significance (p<0.05), all of which showed an association between the minor allele and increased risk of allergic asthma (Table 4).
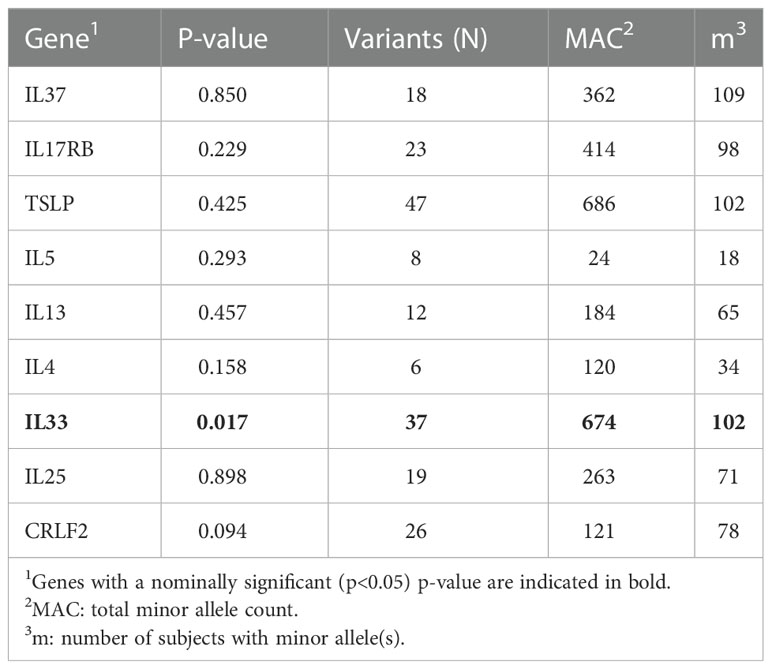
Table 3 Gene-based association results using SKAT.
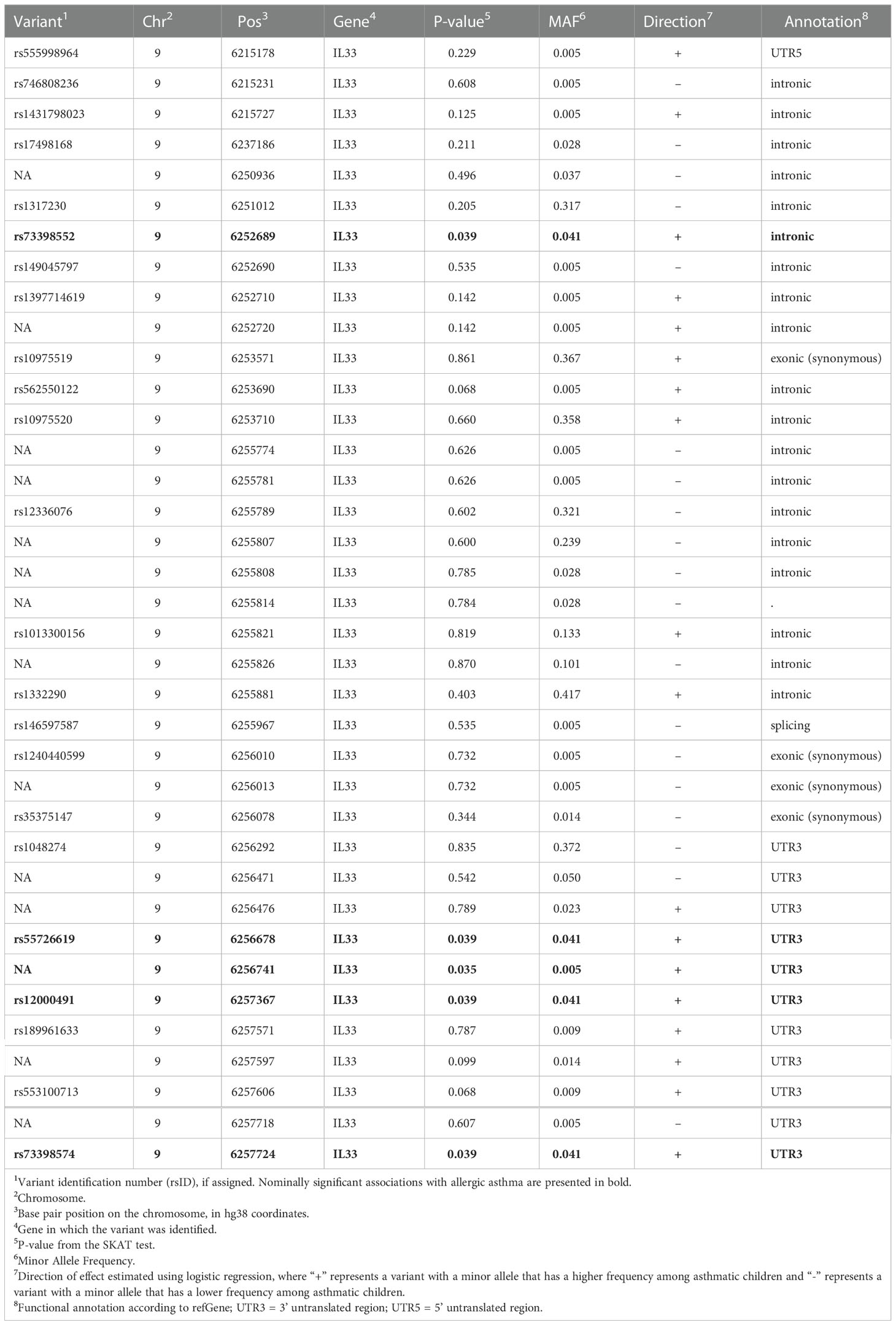
Table 4 Single variant results for IL33.
To test if the IL33 gene-based association was robust to different inclusion/exclusion criteria for variants, subjects and analysis method, we performed a series of sensitivity analyses (Table 5). All sensitivity analyses were nominally significant indicating that the results are robust to these inclusion/exclusion criteria. Further, restricting to low frequency variants (MAF < 5%) yielded the same p-value (p=0.017), indicating that low frequency variants are driving the signal in IL33 rather than common variants.
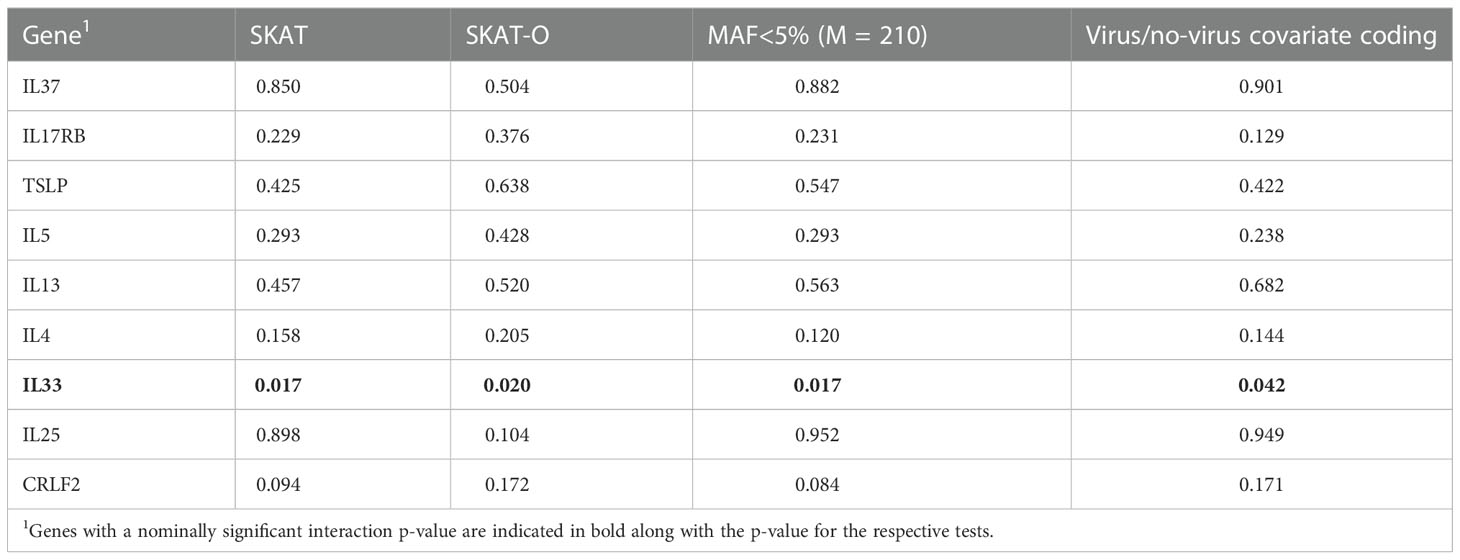
Table 5 Sensitivity analysis results (P-values).
We conducted a gene-virus analysis to identify potential interactions with specific viral types that may be interacting with the gene-based variant sets to alter the risk for asthma. The variant set in IL17RB demonstrated a nominally significant interaction with MPV infection (p=0.050; Table 6).
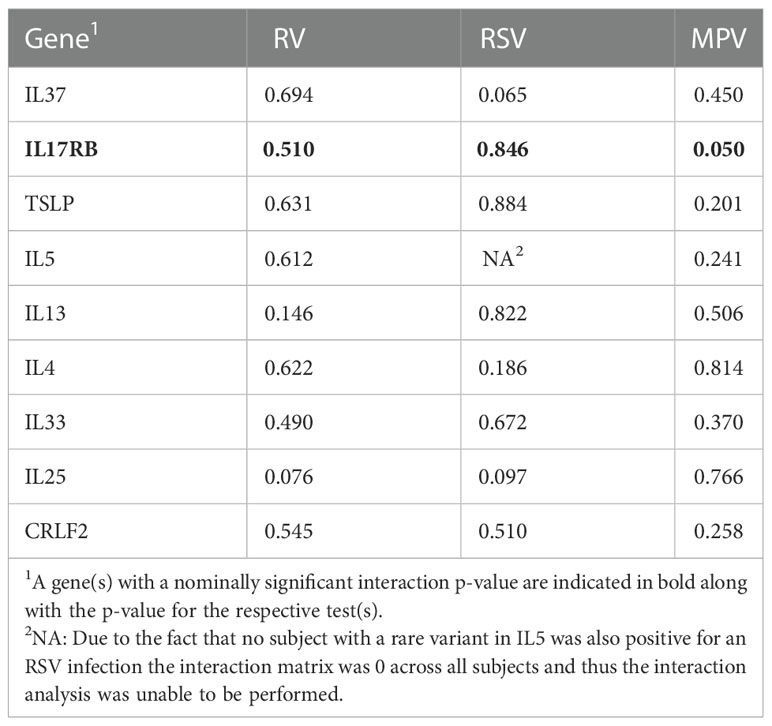
Table 6 Gene x virus interaction analysis results (P-values).
Using children from a well described bronchiolitis cohort and their controls that collected detailed phenotype and clinical data at school-age, we were able to detect a nominal association with asthma for the gene IL33. Further, we were able to detect a nominally significant interaction with MPV infection and variants in IL17RB despite a small sample size. These are promising findings that will need to be confirmed in a larger study sample and contribute to the growing literature of gene-virus interactions contributing to asthma development.
Type 2 cytokine hyperresponsiveness is a hallmark of both asthma and bronchiolitis following rhinovirus (40), RSV (41) and MPV (42, 43) infections. Therefore, an examination of variants within the genes that code for cytokines and their receptors is warranted to identify their potential role in interactions with viral infection and asthma risk. Previous studies identified an association between asthma susceptibility and a variant in IL17RB (44) as well as IL33 which was also associated across diverse ancestries (45, 46).
IL33 induces expression of a number of Th2 cytokines and thus increases eosinophilic inflammation. A rare loss of function variant in IL33 has been shown to be protective against asthma (8). While our analysis did not include any loss of function variants, this does further demonstrate the potential importance of low frequency and rare variants in the etiology of asthma. Variants included in our analysis of IL33 both increased and decreased risk for asthma development (Table 4), however, we do not yet know if the minor alleles increase or decrease expression of IL33 in relevant tissues following viral exposure.
IL17RB encodes the cytokine receptor interleukin-17 receptor B, a receptor specific for IL17B and IL25 (IL17E). IL17 knock-out mice appear to have a lower inflammatory response following MPV infection (42). Further, there is evidence of an intronic IL17RB variant (+5561G>A) in which the minor allele is protective against asthma and associated with lower IL17RB expression (44). Taken together, these observations suggest that genetic variations that increase IL17RB expression may increase the risk for asthma, particularly in those exposed to respiratory infections.
Unlike previous studies of gene-virus interactions for asthma, we chose a sequence-based approach to identify all variants in the coding regions of our selected candidate genes. Our primarily analyses aggregated variants across all those annotated to each gene rather than examining them individually. Our results for IL33 demonstrate that while several single variants within this gene were nominally associated with asthma, no single variant achieved the same level of statistical significance as they did in aggregate. By using a variance-components based method to analyze our aggregated variants, we increased our power to detect associations since the directions of effect of the variants in the IL33 gene were relatively evenly split between risk and protective. Further, our sensitivity results show that the gene-based association between IL33 and asthma is driven by low frequency variants (MAF <5%) which we had no power to detect associations with individually.
Unfortunately, we do not detect any association signals between allergic asthma and the remaining eight candidate genes, despite all of them being strong candidate genes for this phenotype. This could be due to out sequencing strategy of focusing primarily on coding and regulatory regions which may miss many of the tagging genetic variants that were identified in previous genetic studies. Further, we are likely underpowered to detect associations in many of these genes.
These results highlight the multifactorial nature of asthma risk, with both viral infection and inherited genetic variants contributing to increasing asthma risk. While the results presented for both IL33 and IL17RB are statistically significant and are potential targets for designing therapeutics and early screening, these results must be replicated in an independent study. Further work must also be done to determine if the genetic effects on asthma risk are dependent on specific treatment responses, a question requiring a much larger sample size.
The dataset presented in this article is not readily available because Norwegian law limits the sharing of sensitive data which includes genetic sequence data. Requests to access the dataset should be directed to Drs. Kari Risnes and Andrew DeWan.
The study was approved by the Regional Committee on Medical Research Ethics (REK number 2016/540). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
KR, IJ and AD conceived of the overall study design. IJ, AM, HD and KR led the clinical follow-up and data collection. AM performed the clinical data analysis and phenotype generation. ZD performed the genetic analyses. KR, IJ and AD supervised the work. ZD, AM, IJ, TJ, HD, KR and AD interpreted the results. ZD, AM, KR and AD drafted the initial manuscript. All authors contributed to and approved the final manuscript.
Liaison Committee for Education, Research, and Innovation in Central Norway (Samarbeidsorganet); The Faculty of Medicine and Health Sciences, NTNU and Children’s Clinic, St Olav University Hospital
The AmpliSeq custom DNA panel library prep and sequencing analysis were performed in close collaboration with the Genomics Core Facility (GCF), Norwegian University of Science and Technology (NTNU). Processing and alignment of sequencing reads, as well as calling and filtering of variants was performed by the Bioinformatics Core Facility (BioCore), NTNU. GCF and BioCore are funded by the Faculty of Medicine and Health Sciences at NTNU and Central Norway Regional Health Authority. The study visits took place at the Clinical Research Facility Ward at St. Olavs University Hospital.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.1054119/full#supplementary-material
AEC, airway epithelial cells; BoV, bocavirus; FEV1, forced expiratory volume in one second; ILC2, type 2 innate lymphoid cells; ISAAC, International Study of Asthma and Allergies in Childhood; MAF, minor allele frequency; MPT, metacholine provocation test; MPV, metapneumovirus; NPS, nasopharyngeal secretion; RSV, respiratory syncytial virus; RV, rhinovirus; SNP, single nucleotide polymorphism; TSLP, thymic stromal lymphopoietin.
1. Soumelis V, Reche PA, Kanzler H, Yuan W, Edward G, Homey B, et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat Immunol (2002) 3(7):673–80. doi: 10.1038/ni805
2. Lukkarinen M, Koistinen A, Turunen R, Lehtinen P, Vuorinen T, Jartti T. Rhinovirus-induced first wheezing episode predicts atopic but not nonatopic asthma at school age. J Allergy Clin Immunol (2017) 140(4):988–95. doi: 10.1016/j.jaci.2016.12.991
3. Jackson DJ, Gangnon RE, Evans MD, Roberg KA, Anderson EL, Pappas TE, et al. Wheezing rhinovirus illnesses in early life predict asthma development in high-risk children. Am J Respir Crit Care Med (2008) 178(7):667–72. doi: 10.1164/rccm.200802-309OC
4. Makrinioti H, Hasegawa K, Lakoumentas J, Xepapadaki P, Tsolia M, Castro-Rodriguez JA, et al. The role of respiratory syncytial virus- and rhinovirus-induced bronchiolitis in recurrent wheeze and asthma-a systematic review and meta-analysis. Pediatr Allergy Immunol (2022) 33(3):e13741. doi: 10.1111/pai.13741
5. Corren J, Pham T-H, Gil EG, Sałapa K, Ren P, Parnes JR, et al. Baseline type 2 biomarker levels and response to tezepelumab in severe asthma. Allergy (2021) 77(6):1786-96. doi: 10.1111/all.15197
6. Kearley J, Silver JS, Sanden C, Liu Z, Berlin AA, White N, et al. Cigarette smoke silences innate lymphoid cell function and facilitates an exacerbated type I interleukin-33-dependent response to infection. Immunity (2015) 42(3):566–79. doi: 10.1016/j.immuni.2015.02.011
7. Salvati L, Maggi L, Annunziato F, Cosmi L. Thymic stromal lymphopoietin and alarmins as possible therapeutical targets for asthma. Curr Opin Allergy Clin Immunol (2021) 21(6):590–6. doi: 10.1097/ACI.0000000000000793
8. Smith D, Helgason H, Sulem P, Bjornsdottir US, Lim AC, Sveinbjornsson G, et al. A rare IL33 loss-of-function mutation reduces blood eosinophil counts and protects from asthma. PloS Genet (2017) 13(3):e1006659. doi: 10.1371/journal.pgen.1006659
9. Pham T-H, Chen C, Colice G, Parnes JR, Griffiths JM, Cook B. Tezepelumab normalizes serum interleukin-5 and -13 levels in patients with severe, uncontrolled asthma. Ann Allergy Asthma Immunol (2021) 127(6):689–91. doi: 10.1016/j.anai.2021.08.008
10. Menzies-Gow A, Corren J, Bourdin A, Chupp G, Israel E, Wechsler ME, et al. Tezepelumab in adults and adolescents with severe, uncontrolled asthma. New Engl J Med (2021) 384(19):1800–9. doi: 10.1056/NEJMoa2034975
11. Kelsen SG, Agache IO, Soong W, Israel E, Chupp GL, Cheung DS, et al. Astegolimab (anti-ST2) efficacy and safety in adults with severe asthma: A randomized clinical trial. J Allergy Clin Immunol (2021) 148(3):790–8. doi: 10.1016/j.jaci.2021.03.044
12. Ying S, O'Connor B, Ratoff J, Meng Q, Mallett K, Cousins D, et al. Thymic stromal lymphopoietin expression is increased in asthmatic airways and correlates with expression of Th2-attracting chemokines and disease severity. J Immunol (2005) 174(12):8183–90. doi: 10.4049/jimmunol.174.12.8183
13. Zhou B, Comeau MR, De Smedt T, Liggitt HD, Dahl ME, Lewis DB, et al. Thymic stromal lymphopoietin as a key initiator of allergic airway inflammation in mice. Nat Immunol (2005) 6(10):1047–53. doi: 10.1038/ni1247
14. Perez GF, Pancham K, Huseni S, Preciado D, Freishtat RJ, Colberg-Poley AM, et al. Rhinovirus infection in young children is associated with elevated airway TSLP levels. Eur Respir J (2014) 44(4):1075–8. doi: 10.1183/09031936.00049214
15. Lee HC, Headley MB, Loo YM, Berlin A, Gale M Jr., Debley JS, et al. Thymic stromal lymphopoietin is induced by respiratory syncytial virus-infected airway epithelial cells and promotes a type 2 response to infection. J Allergy Clin Immunol (2012) 130(5):1187–96 e5. doi: 10.1016/j.jaci.2012.07.031
16. Lay MK, Cespedes PF, Palavecino CE, Leon MA, Diaz RA, Salazar FJ, et al. Human metapneumovirus infection activates the TSLP pathway that drives excessive pulmonary inflammation and viral replication in mice. Eur J Immunol (2015) 45(6):1680–95. doi: 10.1002/eji.201445021
17. Andreakos E, Papadopoulos NG. IL-25: The missing link between allergy, viral infection, and asthma? Sci Trans Med (2014) 6(256):256fs38. doi: 10.1126/scitranslmed.3010273
18. Klose CS, Artis D. Innate lymphoid cells as regulators of immunity, inflammation and tissue homeostasis. Nat Immunol (2016) 17(7):765–74. doi: 10.1038/ni.3489
19. Wilson SC, Caveney NA, Yen M, Pollmann C, Xiang X, Jude KM, et al. Organizing structural principles of the IL-17 ligand-receptor axis. Nature (2022), 609(7927):622-9. doi: 10.1038/s41586-022-05116-y
20. Tamachi T, Maezawa Y, Ikeda K, S-i K, Hatano M, Seto Y, et al. IL-25 enhances allergic airway inflammation by amplifying a TH2 cell–dependent pathway in mice. J Allergy Clin Immunol (2006) 118(3):606–14. doi: 10.1016/j.jaci.2006.04.051
21. Angkasekwinai P, Park H, Wang Y-H, Wang Y-H, Chang SH, Corry DB, et al. Interleukin 25 promotes the initiation of proallergic type 2 responses. J Exp Med (2007) 204(7):1509–17. doi: 10.1084/jem.20061675
22. Beale J, Jayaraman A, Jackson DJ, Macintyre JDR, Edwards MR, Walton RP, et al. Rhinovirus-induced IL-25 in asthma exacerbation drives type 2 immunity and allergic pulmonary inflammation. Sci Trans Med (2014) 6(256):256ra134. doi: 10.1126/scitranslmed.3009124
23. Demenais F, Margaritte-Jeannin P, Barnes KC, Cookson WOC, Altmüller J, Ang W, et al. Multiancestry association study identifies new asthma risk loci that colocalize with immune-cell enhancer marks. Nat Genet (2018) 50(1):42–53. doi: 10.1038/s41588-017-0014-7
24. Mjosberg J, Spits H. Human innate lymphoid cells. J Allergy Clin Immunol (2016) 138(5):1265–76. doi: 10.1016/j.jaci.2016.09.009
25. Momen T, Ahanchian H, Reisi M, Shamsdin SA, Shahsanai A, Keivanfar M. Comparison of interleukin-33 serum levels in asthmatic patients with a control group and relation with the severity of the disease. Int J Prev Med (2017) 8:65. doi: 10.4103/ijpvm.IJPVM_179_16
26. Zhu J, Dong J, Ji L, Jiang P, Leung TF, Liu D, et al. Anti-allergic inflammatory activity of interleukin-37 is mediated by novel signaling cascades in human eosinophils. Front Immunol (2018) 9:1445. doi: 10.3389/fimmu.2018.01445
27. Zhang L, Zhang J, Gao P. The potential of interleukin-37 as an effective therapeutic agent in asthma. Respir Res (2017) 18(1):192. doi: 10.1186/s12931-017-0675-x
28. Paplinska-Goryca M, Nejman-Gryz P, Proboszcz M, Kwiecien I, Hermanowicz-Salamon J, Grabczak EM, et al. Expression of TSLP and IL-33 receptors on sputum macrophages of asthma patients and healthy subjects. J Asthma (2020) 57(1):1–10. doi: 10.1080/02770903.2018.1543435
29. Schjelderup Nilsen H-J, Nordbø SA, Krokstad S, Døllner H, Christensen A. Human adenovirus in nasopharyngeal and blood samples from children with and without respiratory tract infections. J Clin Virol (2019) 111:19–23. doi: 10.1016/j.jcv.2018.12.005
30. Heimdal I, Moe N, Krokstad S, Christensen A, Skanke LH, Nordbø SA, et al. Human coronavirus in hospitalized children with respiratory tract infections: A 9-year population-based study from Norway. J Infect Dis (2019) 219(8):1198–206. doi: 10.1093/infdis/jiy646
31. Moe N, Stenseng IH, Krokstad S, Christensen A, Skanke LH, Risnes KR, et al. The burden of human metapneumovirus and respiratory syncytial virus infections in hospitalized Norwegian children. J Infect Dis (2017) 216(1):110–6. doi: 10.1093/infdis/jix262
32. Asher MI, Anderson HR, Krokstad S, Stewart AW, Crane J. Worldwide variations in the prevalence of asthma symptoms: The international study of asthma and allergies in childhood (ISAAC). Eur Respir J (1998) 12(2):315–35. doi: 10.1183/09031936.98.12020315
33. Crapo RO, Casaburi R, Coates AL, Enright L, Hankinson JL, Irvin CG, et al. Guidelines for methacholine and exercise challenge testing–1999. Am J Respir Crit Care Med (2000) 161(1):309–29. doi: 10.1164/ajrccm.161.1.ats11-99
34. Miller MR, Hankinson J, Brusasco V, Burgos F, Casaburi R, Coates A, et al. Standardisation of spirometry. Eur Respir J (2005) 26(2):319. doi: 10.1183/09031936.05.00034805
35. Global initiative for asthma. global strategy for asthma management adn prevention (2021). Available at: http://www.ginaasthma.org.
36. Wang K, Li M, Hakonarson H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res (2010) 38(16):e164. doi: 10.1093/nar/gkq603
37. Wu MC, Lee S, Cai T, Li Y, Boehnke M, Lin X. Rare-variant association testing for sequencing data with the sequence kernel association test. Am J Hum Genet (2011) 89(1):82–93. doi: 10.1016/j.ajhg.2011.05.029
38. Lee S, Emond MJ, Bamshad MJ, Barnes KC, Rieder MJ, Nickerson DA, et al. Optimal unified approach for rare-variant association testing with application to small-sample case-control whole-exome sequencing studies. Am J Hum Genet (2012) 91(2):224–37. doi: 10.1016/j.ajhg.2012.06.007
39. Lin X, Lee S, Christiani DC, Lin X. Test for interactions between a genetic marker set and environment in generalized linear models. Biostatistics (2013) 14(4):667–81. doi: 10.1093/biostatistics/kxt006
40. Rajput C, Han M, Ishikawa T, Lei J, Goldsmith AM, Jazaeri S, et al. Rhinovirus c infection induces type 2 innate lymphoid cell expansion and eosinophilic airway inflammation. Front Immunol (2021) 12:649520–. doi: 10.3389/fimmu.2021.649520
41. Norlander AE, Peebles RS Jr. Innate type 2 responses to respiratory syncytial virus infection. Viruses (2020) 12(5):521. doi: 10.3390/v12050521
42. Liu Y, Qin T, Zhao X, Dong S, Zhu J, Peng D, et al. Skewed balance of regulatory T cell and inflammatory T cell in IL-17 defect with human metapneumovirus infection. Cell Immunol (2018) 331:161–7. doi: 10.1016/j.cellimm.2018.06.007
43. Soto JA, Galvez NMS, Benavente FM, Pizarro-Ortega MS, Lay MK, Riedel C, et al. Human metapneumovirus: Mechanisms and molecular targets used by the virus to avoid the immune system. Front Immunol (2018) 9:2466. doi: 10.3389/fimmu.2018.02466
44. Jung J-S, Park BL, Cheong HS, Bae JS, Kim J-H, Chang HS, et al. Association of IL-17RB gene polymorphism with asthma. Chest (2009) 135(5):1173–80. doi: 10.1378/chest.08-1595
45. Moffatt MF, Gut IG, Demenais F, Strachan DP, Bouzigon E, Heath S, et al. A large-scale, consortium-based genomewide association study of asthma. N Engl J Med (2010) 363(13):1211–21. doi: 10.1056/NEJMoa0906312
Keywords: childhood asthma, viral respiratory infection, bronchiolitis, type 2 cytokines, genetic risk factors, gene-environment interaction (G × E)
Citation: Dong Z, Myklebust Å, Johnsen IB, Jartti T, Døllner H, Risnes K and DeWan AT (2023) Type 2 cytokine genes as allergic asthma risk factors after viral bronchiolitis in early childhood. Front. Immunol. 13:1054119. doi: 10.3389/fimmu.2022.1054119
Received: 26 September 2022; Accepted: 15 December 2022;
Published: 06 January 2023.
Edited by:
Raffaella Nenna, Sapienza University of Rome, ItalyReviewed by:
Patricia De NADAI, Université de Lille, FranceCopyright © 2023 Dong, Myklebust, Johnsen, Jartti, Døllner, Risnes and DeWan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrew T. DeWan, YW5kcmV3LmRld2FuQHlhbGUuZWR1; Kari Risnes, a2FyaS5yaXNuZXNAbnRudS5ubw==
†These authors have contributed equally to this work and share first authorship
‡These authors share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.