 Yang Du
Yang Du Xiaoman Hu1,2,3†
Xiaoman Hu1,2,3† Jiong Chen
Jiong Chen- 1State Key Laboratory for Managing Biotic and Chemical Threats to the Quality and Safety of Agro-products, Ningbo University, Ningbo, China
- 2Laboratory of Biochemistry and Molecular Biology, School of Marine Sciences, Ningbo University, Ningbo, China
- 3Key Laboratory of Applied Marine Biotechnology of Ministry of Education, Ningbo University, Ningbo, China
- 4Key Laboratory of Marine Biotechnology of Fujian Province, Institute of Oceanology, Fujian Agriculture and Forestry University, Fuzhou, China
Diseases are a significant impediment to aquaculture’s sustainable and healthy growth. The aquaculture industry is suffering significant financial losses as a result of the worsening water quality and increasing frequency of aquatic disease outbreaks caused by the expansion of aquaculture. Drug control, immunoprophylaxis, ecologically integrated control, etc. are the principal control strategies for fish infections. For a long time, the prevention and control of aquatic diseases have mainly relied on the use of various antibiotics and chemical drugs. However, long-term use of chemical inputs not only increases pathogenic bacteria resistance but also damages the fish and aquaculture environments, resulting in drug residues in aquatic products, severely impeding the development of the aquaculture industry. The development and use of aquatic vaccines are the safest and most effective ways to prevent aquatic animal diseases and preserve the health and sustainability of aquaculture. To give references for the development and implementation of aquatic vaccines, this study reviews the development history, types, inoculation techniques, mechanisms of action, development prospects, and challenges encountered with aquatic vaccines.
1 Introduction
Global aquatic products, including aquatic plants, fish, crustaceans (shrimps and crabs, etc.), molluscs (scallops, abalone, oysters, mussels, etc.) and other species (bullfrogs, jellyfish, etc.), are the third largest source of food protein for human consumption after cereals and milk, accounting for 16.4% of the total animal protein supply (1, 2). Among them, fish are the dominant species of farmed and harvested aquatic products worldwide, accounting for more than 40% of the aquaculture industry. The growing demand for fish and other aquatic foods has changed the face of fisheries and aquaculture, especially accelerating the development of aquaculture and increasing the proportion of aquaculture production to approximately 7.7% by 2020, the total fisheries and aquaculture production rises to a record high of 214 million tons, including 178 million tons of aquatic, and the proportion is expected to grow by a further 15% by 2030 (3). However, the growth of aquaculture has often come at the expense of the environment, resulting in the potential for major disease problems and food safety issues gradually increasing. Especially in recent years, viral diseases of aquatic animals such as grass carp hemorrhagic disease, carp spring virus disease, grouper iridovirus disease, infectious pancreatic necrosis, infectious spleen and kidney necrosis, infectious hematopoietic organ necrosis and viral hemorrhagic septicemia and other high incidences of disease, almost involving the main species of aquaculture, causing serious economic losses to aquaculture. It is estimated that 10% of all farmed aquatic animals die due to infectious diseases, costing more than $10 billion per year globally (4). The overall trend of aquaculture diseases shows a high incidence, diversified types, long duration of onset, a wide range of infections, and increasing year by year etc, so it is apparent that disease control in aquaculture is a matter of urgency (5, 6). Due to the lack of basic theoretical research on pharmacodynamics, pharmacogenetics, and toxicology and their impact on the aquaculture ecosystem, as well as serious problems such as resistance of aquatic pathogens, it is no longer possible to solve the current disease problems by continuing to rely on the use of various chemical drugs and antibiotics. The use of microecological preparation, immunostimulants, vaccines etc. for integrated disease control is more advocated in the new ecological farming model. Therefore, vaccination, as a green ecological control means, has become a mainstream technology for global aquatic animal disease control and has a good prospect in the development of the aquaculture industry in the world (7).
2 Aquatic vaccines overview
Vaccination is the most effective way to prevent and control diseases caused by viruses and bacteria. Research work on aquatic vaccines started in the 1940s. Duff et al. (1942) (8) performed the first successful oral immunization with inactivated Bacterium salmonicida on trout. Since the 1980s, significant progress has been made in the development of aquatic vaccines. The number of commercial vaccines available for use in fish against the major infectious bacterial and viral diseases has increased from 2 in the 1980s to over 50 currently (Table 1) (13, 17, 26). According to incomplete statistics, as of 2020, more than 140 aquatic vaccines have been approved worldwide (27). These include whole inactivated, peptide subunit, recombinant protein, nucleic acid, and live attenuated vaccines. However, some commercial vaccines are not always protective, and aquatic diseases continue to be severe (28, 29). Current disease vaccination is routinely used in some economically important fish species such as Atlantic salmon, Rainbow trout, Nile tilapia, Amberjack, and Striped catfish (14, 30), whereas corresponding vaccines are lacking, ineffective, or expensive in many other economic fish species. Currently, parasite vaccines face significant challenges, with only one commercial parasite vaccine available against ectoparasite sea lice. In addition, only a few trials have reported potential vaccine candidates against endoparasites (31). Furthermore, most commercial vaccines are administered via intraperitoneal injection (32), which may not be the best route of immunization for large-scale population vaccination and is frequently influenced by factors such as animal species, immune system status, production cycle, environment, nutrition and cost-effectiveness (33). In some countries and regions, the high cost of vaccination has led to the abandonment of vaccine defence due to widespread drug use. In light of the rapidly thriving global aquaculture industry, it is especially crucial to solve these concerns.
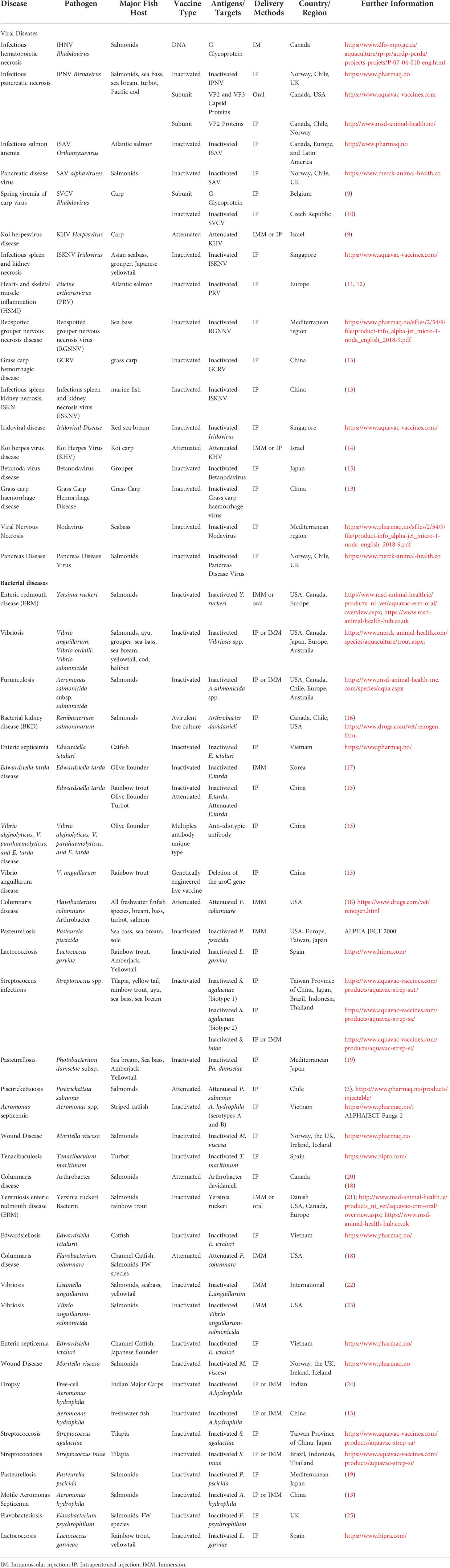
Table 1 Global commercial aquatic vaccines.
In recent years, with the continuous reduction of technological costs (e.g., genome sequencing, high-throughput screening of antigens, expanded cell culture, etc.), the development of novel antigen expression and delivery systems, and the massive accumulation of basic knowledge on fish mucosal immunity, aquatic vaccines are bound to see rapid development in the coming years (34). Developing highly effective mucosal vaccines and corresponding adjuvants is an important direction for future development, and optimizing vaccine delivery will undoubtedly facilitate the development of new vaccines. Aquaculture in low-income nations will be able to adopt routine immunization in the future, which will drastically reduce the use of drugs and decrease disease outbreaks and transmission on a worldwide scale. Since vaccine development processes are lengthy, it is impossible to have a vaccine on hand when a disease outbreak first starts. As a result, it is of great significance to analyze the common factors that exist among various known pathogens, find the trend rules, and conduct more clinical trials to discover vaccines with cross-protection based on existing targeted successful vaccines. In general, efficient vaccine development and utilization will contribute to the global aquaculture industry’s healthy and sustainable development.
3 Classification and preparation technology of aquatic vaccines
Aquatic vaccines are an effective preventative measure against many diseases and have recently gained popularity. According to the antipathogen, aquatic vaccines can be classified as bacterial, viral, and parasitic. They can also be classified according to composition, including monovalent, multivalent, or mixed (multivalent) vaccines, or classify by the preparation method, such as live, inactivated, or genetically engineered vaccines (35). It is worth noting that each type of vaccine has advantages and disadvantages and that different vaccines need to be developed and used for different pathogens and animals (as shown in Table 2). Aquatic vaccine production technology has developed quickly since the introduction of the Aeromonas salmonicida inactivated vaccine in 1942, and vaccine types have also diversified. Currently, most commercial aquatic vaccines are still inactivated, and the primary methodology for vaccine manufacture is still the inactivation of virulent wild-type pathogens by physical and chemical methods. However, inactivated vaccines have drawbacks such as short immunity duration, large inoculation dosage, adverse responses, and strict inactivation requirements. The utilization of live attenuated vaccines in aquaculture provides several benefits, including simple vaccination, low manufacturing costs, rapid delivery, low immunization dosage, and prolonged immunity. Positive outcomes have been attained with several significant economic fish. For instance, the use of antibiotics in Norwegian and UK aquaculture has significantly decreased as a result of the Arthrobacter Vaccine’s favourable immunological response in salmonids (12). Deficiencies are mostly caused by live attenuated vaccines’ poor safety in natural settings, which can result in virus transformation and loss of ecological environment control. It also has a limited shelf life, low thermal stability, high transport and storage requirements, and the danger of resuming mutations. Subunit vaccine keeps only the effective immunogen components and discards the irrelevant or dangerous pathogen components that trigger protective immunity. Subunit vaccines eliminate irrelevant or harmful pathogen components that induce protective immunity while retaining only the effective immunogenic components, resulting in significantly improved and stable immune effects, increased reliability, and fewer adverse reactions in the body. However, they frequently need optional adjuvants and are incapable of producing mucosal and cellular protection (36). But the plant-produced subunit vaccines are easier to manufacture, transport and store, and the immunization route is simpler and safer. They can be added to feeds as additives to induce mucosal and systemic immune responses that are more effective than traditional immunization routes and have great potential for application in aquaculture (37). Heterologous vaccines are those that are created with heterologous pathogens that cross-react with pathogens. It is safe and has a long immunity period, but it is difficult to obtain. With the advancement of immunology and molecular biology technology in recent years, there are more technical means for the development of aquatic vaccines, such as recombinant vaccine preparation technology, gene deletion attenuated vaccine preparation technology, genetic engineering live vector preparation technology, nucleic acid vaccine preparation technology, and plant vaccine preparation technology, etc. Vaccines developed using molecular biology methods provide a variety of benefits: They have immune properties that are chemically well-defined and stable; they have known chemical structures that can be engineered and modified to stimulate particular immune responses; they are free of infectious components and have no residual toxicity or risk of toxicity reversion; they can be synthesized directly or expressed through recombinant expression, which makes it easier to manufacture and enables the development of multivalent vaccines (38–40). Using genetic engineering technology to prepare vaccines is especially significant and advantageous for pathogens that cannot or are difficult to culture, pathogens with potential carcinogenic or immunopathological effects, or pathogens with poor effects or high adverse reactions to traditional vaccines. However, genetically engineered vaccines currently face varieties of challenges, including expensive development and production costs, protracted development cycles, and a single protective property. As a result, improved design strategies for the advanced, novel, and effective genetically engineered multivalent vaccines will be an important development direction for the preparation of vaccines for aquaculture.

Table 2 Types and characteristics of aquatic vaccines.
3.1 Live-attenuated vaccine
In veterinary medicine, there are three types of live vaccines: strong, weak, and heterologous vaccines (41). Live aquatic vaccines are now more commonly used with pathogenic strains that have been weakened or mutated, such as the F25 (9) antipyretic strain of viral hemorrhagic septicemia virus (VHSV), infectious spleen and kidney necrosis virus (ISKNV), attenuated channel catfish virus (CCV) vaccine, attenuated Furunculosis vaccine, attenuated haematopoietic necrosis virus (IHNV) vaccine (42–47). The outstanding benefit of live vaccines is that the pathogen replicates in the host to produce antigenic stimuli, and the quantity, type, and location of these antigens are similar to those of natural infections. As a consequence, immunogenicity is typically strong, even without the need for a booster vaccination and the addition of adjuvants, and immunity lasts longer, displaying better immune protection (48). The live vaccine can be immunized by injection, but it is also very effective by immersion, and oral or nasal vaccination (49–52). This exceptional benefit is, however, accompanied by potential hazards, including inconveniences with storage and transportation, a short shelf life, the potential for infection to be triggered in those with low immunity, and the potential for mutations to reestablish virulence. Additionally, live attenuated/weakly virulent vaccines are excellent starting strains for vector vaccines due to their ability to retain high host invasiveness and restricted host multiplication.
There are several techniques for producing live attenuated vaccines, including chemical/physical mutagenesis, genetic engineering, attenuated culture, and antibiotic-induced attenuation. Early live attenuated vaccines were continuously transmitted to attenuated strains for screening, but the process was time-consuming and the virulence returned. Antibiotic-induced attenuation is the term for the in vitro screening of weak strains that are not antibiotic-dependent using media that contain antibiotics (53). Antibiotic-induced attenuated mutant strains, on the other hand, are haphazard, lack a clear genetic background, and are virulently unstable. It is difficult to ensure their safety when used in vaccine preparation and therefore hasn’t been frequently applied in following attenuated investigations. Chemical/physical mutagenesis attenuation refers to the use of chemical mutagens or altered physical conditions to induce non-situated mutations in the causative agent thereby obtaining a weakly virulent strain. The mutations produced by this induction method are random and the corresponding vaccine strain is selected due to the lack of virulence in the host animal (54, 55). Currently, with the development of genome sequencing and genetic engineering technologies, a clear understanding of the genetic background and pathogenic mechanisms of various pathogens has been achieved, and safe and efficient live attenuated vaccine strains can be constructed by knocking out virulence and metabolism-related genes of virulent strains (56). As a new means of genetic engineering, homologous recombination technology can modify DNA without any restriction endonuclease and DNA ligase. Attenuated strains constructed by knocking out one or more virulence genes using genetic engineering techniques have the advantages of a clear genetic background, weaker virulence and less susceptibility to reassortment. Some of the common virulence-related genes include housekeeping genes involved in encoding structural components of bacteria, outer membrane protein genes (57, 58), aromatic amino acid genes involved in essential metabolite synthesis (59), virulence genes involved in bacterial host resistance mechanisms, population sensing genes (60–64), iron uptake system genes (65, 66), virulence island (SPI)-related genes (67–71), secretion system-related genes, etc (72–75). Delete multiple functional genes, such as flagellin and enzymes, present more desirable immune protection (76–80), and even produce a good cross-protection (81–83).
In recent years, important progress has been made in the research of live aquatic vaccines (Table 3). The attenuated vaccine against Edwardsiella tarda and the genetically engineered weakened vaccine against Vibrio eels used in turbot and flounder culture have resulted in important protection of the fish (106–108). The Nile tilapia (Oreochromis niloticus) (109–111), silver pompano (Pampus argenteus) (112), giant Queensland grouper (Epinephelus lanceolatus) (113), Australian jewel perch (Scortumco) (114), bighead carp (Aristichthys nobilis) (115), and many other fish species can be infected by the important pathogen Streptococcus lactis, which threatens the aquaculture industry and causes high mortality. Liu et al. (2019) (100) developed an attenuated S. lactis TFJ0901 (named TFJ-ery) from a naturally low virulence S. lactis strain through erythromycin resistance screening, and they found TFJ-ery to be an effective attenuated vaccine candidate to protect tilapia from S. lactis infection. Zhang et al. (2020a) (102) developed an effective live attenuated vaccine against S. lactis that was also able to induce humoral and cellular immune responses in tilapia, significantly increasing the level of specific antibodies. A live attenuated vaccine was created by Liu et al. (2018) (116) to protect against the Vibrio disease in Takifugu rubripes, and it was discovered that the vaccination had no negative impacts on fish growth. T. rubripes also produced an efficient immune response after receiving the live attenuated vaccination. High-density farming of loach and deteriorating farming conditions have accelerated the outbreak of Aeromonas hydrophila and are a potential threat to food quality. Zhang et al. (2020) (117) developed a live attenuated vaccine to treat loach infected with A. hydrophila TH0426 and found that the vaccine induced an increase in enzyme activity in the serum and skin mucus of loach and upregulated immune-related genes, including IL-1β, TNF-α, IL-10 and pIgR, which effectively protected the loach from infection by A. hydrophila TH0426. Zhou et al. (2020) (96) developed a live attenuated vaccine against Vibrio alginolyticus infection that increased the expression of IgM, IL-1β, IL-6 and TNF-α and induced a protective immune response in zebrafish.

Table 3 Attenuated vaccine.
In general, live attenuated vaccines have great potential in the aquaculture industry because of their weakened toxicity, which cannot cause diseases, and their ability to be administered via natural infection routes. These vaccines provide extended and consistent antigen presentation, stimulate humoral and cell-mediated immune responses, induce host mucosal immunity, and offer the organism broad immune protection. In light of this, live attenuated vaccines have enormous promise in the aquaculture industry, and the development of novel, diverse live attenuated vaccines using genetic engineering technologies is a key area of focus for the future development of fishery vaccines.
3.2 Inactivated vaccine
Inactivated vaccines are vaccines that use physical or chemical methods to inactivate highly virulent pathogenic microorganisms, but they still maintain their immunogenicity and produce specific resistance in aquatic animals after vaccination. Physical inactivation methods include ultraviolet light (118), high-temperature heating (119–121), ultrasound (122) and γ-ray (123). Chemical inactivation is the process of rendering pathogenic bacteria dormant by destroying their nucleic acids or proteins with chemical chemicals. Physical inactivation is complex, costly, limiting and unstable, while in contrast, chemical inactivation is simple, low cost and reliable, and is the most commonly used method of inactivation (124, 125). In 1942, Duff first applied an inactivated vaccine against A. salmonicida on trout to give them immune protection, which became a precedent for fish vaccine application. An inactivated cell culture vaccine for grass carp disease was developed in China in 1986 (126), which was the first successful step in the development of aquatic vaccines. Inactivated vaccines also have limitations in that their immunogenicity is inevitably affected by the way they are prepared, which destroys the integrity of some pathogens, thus affecting the protective effect of the vaccine (127). Meanwhile, inactivated vaccines are not biologically active after entering the host and cannot colonize and reproduce, thus requiring higher doses and shorter duration of immunity, and need to be improved by adding appropriate adjuvants and making multivalent vaccines or combination vaccines. However, inactivated vaccines have the advantages of a short development period, stable storage, low cost and no virulence problems, and are currently the most reported and used vaccines in aquaculture (128).
Bacterial diseases can cause significant biological harm and thus economic losses. Although it can be controlled with antibiotics, these drugs eventually pose a threat to human health due to the development and transfer of resistance mechanisms among bacterial species (129, 130). Therefore, inactivated bacterial vaccines have been the subject to combat the spread of bacterial diseases among aquatic animals, as shown in Table 4, many inactivated aquatic bacterial vaccines have been reported. Liu et al. (2015) (132) used 0.5% formalin to inactivate eel vibrio at 30°C for 48 h. They found that the survival of immunized zebrafish and turbot was effectively improved with relative survival rates of (89.0 ± 4.5) % and (80.0 ± 6.9) %, respectively. Nguyen et al. (2017) (152) inactivated Vibrio harveyi with 0.3% formalin at 25°C for 48 h. Immunization of grouper for 6 weeks resulted in 100% relative survival and a significant increase in interleukins. Inactivation of Haemonchus contortus with formalin at an optimal vaccination rate of 1.0×108 CFU/fish significantly promoted antibody production in turbot, and enhanced lysozyme activity, total serum protein and antimicrobial properties in the serum of immunized fish (153). Formalin-inactivated F. harveyi ZJ0603 vaccine (FKC) combined with β-glucan improved immune protection in pearl grouper, and the expression levels of IgM, TNF-α, MHC-Iα, IL-1β and IL-16 in the spleen and the antibody potency, lysozyme and superoxide dismutase activities in the blood of immunized fish were significantly increased (154). Aeromonas veronii is an extremely important infectious pathogen in carp culture. Inactivated A. veronii TH0426 significantly improves immune protection against crucian carp, while the addition of adjuvants can promote vaccine efficacy (139).
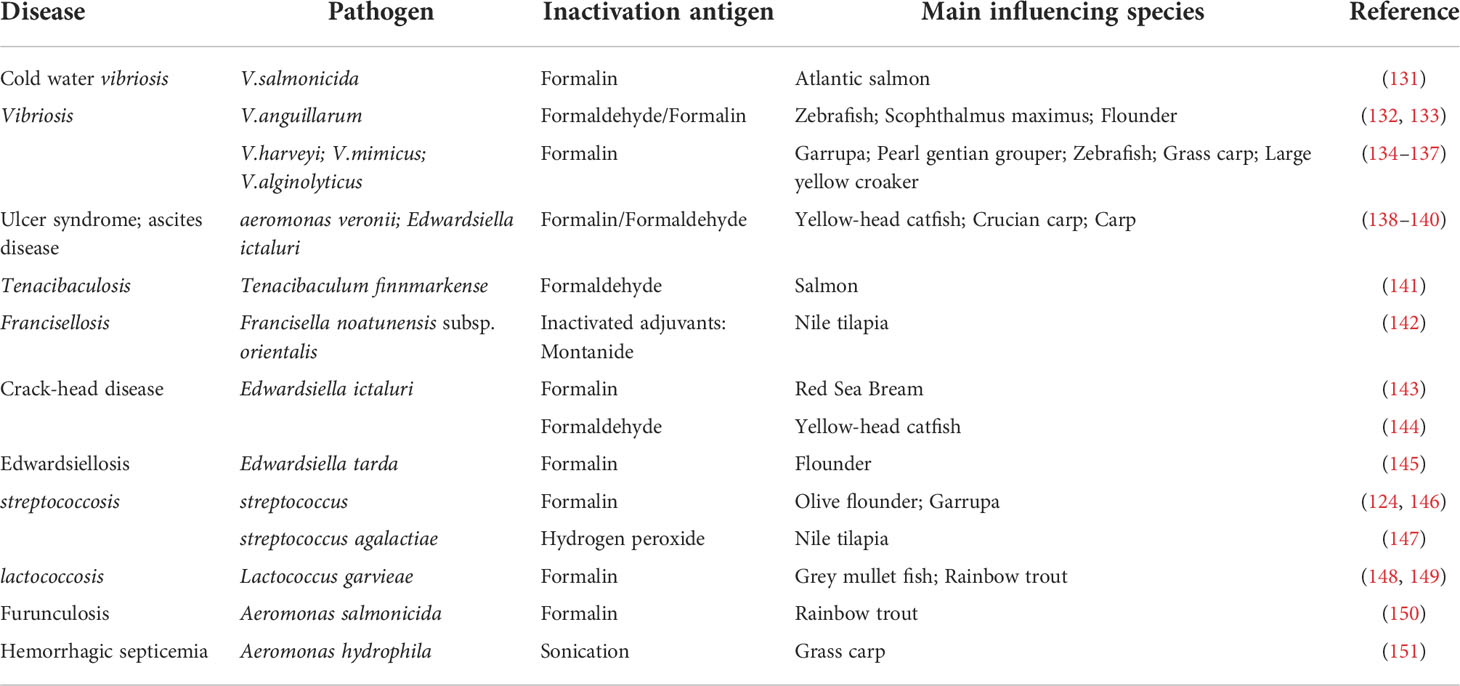
Table 4 Inactivated Bacterial vaccine.
Inactivated viral vaccines are safe and won’t cause any infections because the virus has lost its activity, many inactivated aquatic viral vaccines have been reported in studies (Table 5). However, the immune response is lessened or the duration of immunity is reduced since the virus is only a fragment and not an intact virus (173). It has been reported that the infectious hematopoietic necrosis virus (IHNV) vaccine inactivated by 1.5 mmol/L Beinylating agents can significantly improve the survival rate of rainbow trout (155). Neural necrosis virus vaccine inactivated by 4 mmol/L BEI can not only significantly promote the expression of various immune genes in grouper, but also cause body fluids and cellular immunity (162). Inactivated cyprinid herpesvirus type 2 (CyHV-2) vaccine with 0.1% BPL (chemical formula C3H4O2) at 4°C significantly induced non-specific and specific antiviral immune responses in carp, resulting in relative immune protection of 71.4% (164). Grass carp had a relative immune protection rate of more than 80% after receiving the grass carp reovirus (GCRV) vaccine inactivated with 0.1% BPL (165). The addition of adjuvants is effective in improving the effectiveness of inactivated viral vaccines, for example, because squalene and aluminium hydroxide adjuvants prolong and improve the efficiency of formalin-inactivated VHSV in flounder, which can be controlled by a single dose of this vaccine administered before winter (174). Montanide IMS 1312 adjuvant significantly improves the immune efficiency of formaldehyde and β-propiolactone inactivated tilapia lake virus-inactivated vaccine and increases protection against tilapia (175).
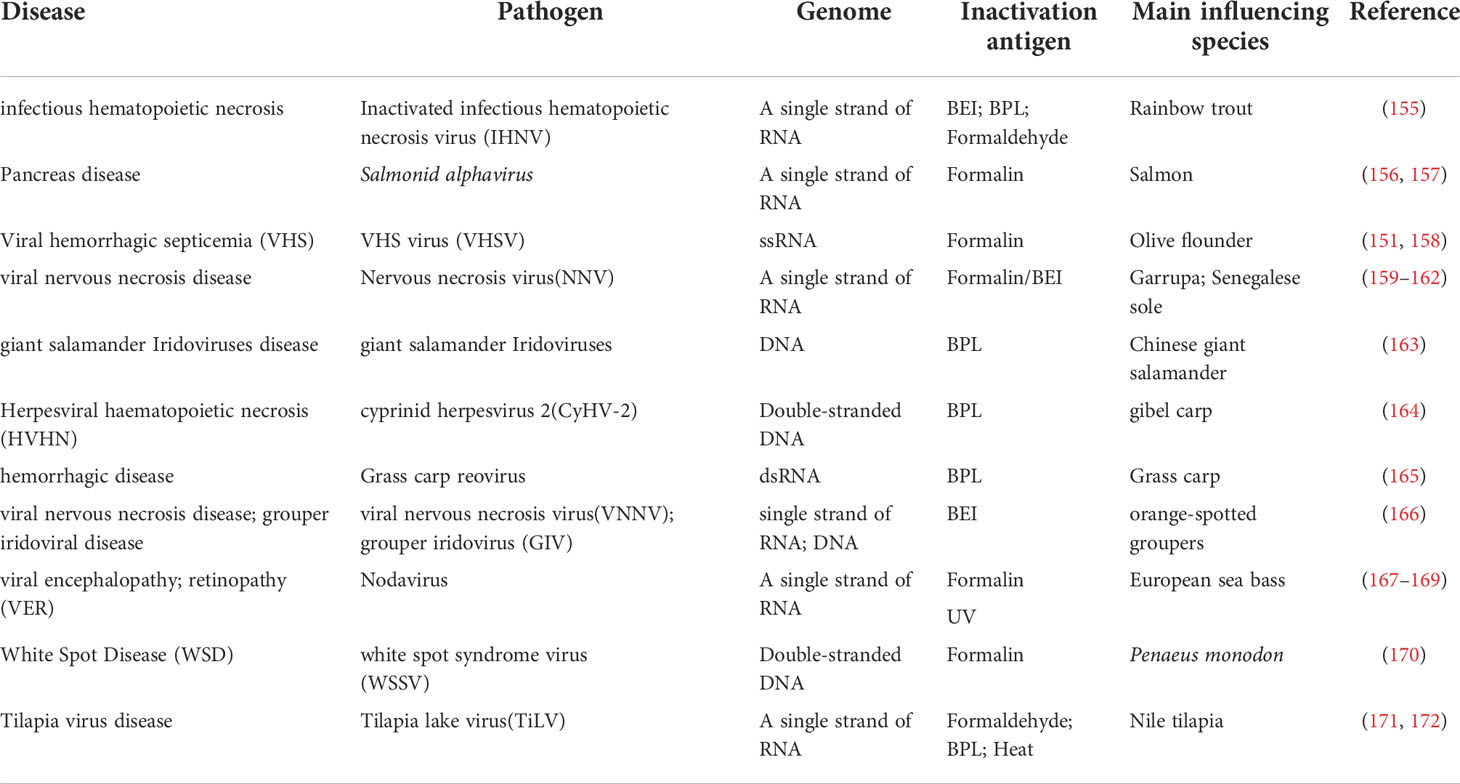
Table 5 Inactivated Viral vaccine.
3.3 Genetically engineered vaccine
Genetic engineering vaccines primarily use genetic engineering technology to isolate immune antigen genes from bacteria or viruses and transfer them into animal cells or recipient organisms, allowing the organism to produce a large number of protective antigens and improve disease resistance. The main types include recombinant subunit vaccines, nucleic acid vaccines, recombinant live vector vaccines, gene deletion/mutation vaccines and plant vaccines. Aquatic vaccines have been widely used in aquaculture since the 1980s, but the majority of commercial vaccines are inactivated bacterial vaccines, with only a few being genetically engineered recombinant vaccines.
3.3.1 Subunit vaccine
A subunit vaccine is a vaccine prepared by encoding a pathogen antigen into a recombinant expression vector and expressing a gene product (recombinant peptide or recombinant protein) using a prokaryotic or eukaryotic expression system. Subunit vaccines utilize only the antigenic component for vaccination and pose no risk of causing disease in the host or non-target species because subunit vaccines cannot replicate in the host. Furthermore, subunit vaccines can be manufactured in a highly characterized state, they can target immune responses to specific microbial determinants, they can incorporate non-natural components, and they can be freeze-dried, allowing for non-refrigerated transport and storage. Heterologously expressed recombinant subunit vaccines address both difficulties with antigen supply and vaccination safety, are easily accessible when testing is required and can be produced and used consistently. Some of the most important characteristics of vaccine candidate molecules are that they are highly conserved among different strains of the same species and that they are expressed on the pathogen’s surface so that antigen-presenting cells can easily recognize them due to their immunogenicity. The high immunogenicity of outer membrane proteins (OMP) is considered a promising candidate for the design of antimicrobial drugs and vaccines in various pathogenic strains. Numerous other studies have reported that outer membrane proteins (OMPs) are generally very immunogenic due to their exposed epitopes on the cell surface (176) and have the potential of a vaccine candidate for fish against V. anguillarum (177, 178), Vibrio mimicus (179), V. harveyi (180), Vibrio ichthyoenteri (181), P. olivaceus (182), E. tarda (183) and A. hydrophila (184) infection. Flagellin is also well-studied and effective subunit vaccine candidate (185). Viral capsid proteins are also regarded as significant possible antigens for the development of subunit vaccines (186, 187). It is concerning that subunit vaccines frequently contain a single antigen, resulting in low immunization efficiency, necessitating multiple immunizations and the addition of adjuvants to improve the immunization effect or the development of multiple vaccines (188–192). Cytokines can act as signalling molecules involved in the immune system and as low molecular weight glycoproteins or peptides regulating the host defence network (193). Compared to the most commonly used adjuvants in fish culture (e.g., aluminium plate adjuvants and oil adjuvants), cytokines have advantages as immune adjuvants in initiating the expression of co-stimulatory molecules and polarization of antigen-presenting cells (194). A large number of cytokines have been identified in various fish species, for example, IL-1β, IL-8, G-CS, TNF-α, IL-6, IL-2 and other cytokines have been found to enhance the immune effect of subunits in flounder (195, 196).
Subunit antigens have been produced in bacteria, yeast, transgenic plants, insects, mammalian cell cultures, and cell-free platforms. Among them, bacteria can easily and abundantly express antigens, but cannot post-translationally modify the expression products; therefore, eukaryotic antigens are usually not expressed in bacteria. And the expressed exogenous proteins have damaging effects on the host bacteria, and there may be a risk of endotoxin contamination in the expression end products. Yeast is widely used in food, beverage and feed production, and its genetic and biochemical background is clear. Yeast not only expresses proteins in a secreted form but also post-translationally modifies them. Yeast is easy to operate, has a fast growth rate, and has now mastered a lot of experience in large-scale fermentation production, with great potential in aquatic vaccines. The advantage of the mammalian expression system is that it can post-translationally modify the expression products close to the natural conformation, which is important for inducing immune responses to vaccine antigens. However, the cellular production of vaccines is too costly and potentially carries the risk of contamination with foreign or oncogenic components (2, 197). The plant-based vaccine also referred to as a transgenic plant vaccine, involves the introduction of antigen genes into plant cells and their expression, or the replication of antigen genes in plants using pathogenic vectors to produce high yields of the target protein. Transgenic plants are currently being used to produce vaccines against viruses, bacteria, and parasites, and clinical studies have shown that plant-produced recombinant drug proteins are safe and effective (198, 199). Compared to microbial or mammalian cell expression, plants have a higher biosynthetic capacity of which plant seeds, tubers and fruits are excellent locations for protein aggregation and conservation. Stably expressed plants integrating antigenic genes can be produced in large quantities by asexual or sexual reproduction. They can also undergo post-translational modifications like glycosylation and complex folding and assembly to improve vaccine immunogenicity (200). Furthermore, there are no safety problems with plant-made subunit vaccines compared to live vaccines, and no harmful substances like bacterial endotoxins or highly glycosylated proteins from yeast have been discovered in plant production systems (201). Plant-produced recombinant subunit vaccines can also deliver multiple antigenic proteins simultaneously (202). Thus, plant-based platforms offer appealing benefits for recombinant subunit vaccines. Plant-produced vaccines are appropriate for oral delivery, as the vaccine antigen is absorbed by M cells in the epithelium after passing through the stomach and into the intestine, eliciting mucosal and systemic immune responses (203). Aquatic vaccines made from edible plants hold great promise for oral vaccines in aquaculture, as such transgenic plants can be directly crushed and added to feeds as additives to trigger a particular immune response. The best plants for edible vaccines include vegetables and fruits that have been shown to generate substantial amounts of exogenous proteins, such as lettuce, tomatoes, potatoes, rice, and cabbage (204, 205). According to a study, feeding striped bream transgenic rice healing tissue with recombinantly expressed Iridovirus coat protein (MCP) dramatically increased the fish’s disease resistance (206).
Microalgae are natural baits and nutritional additives for aquatic animals, especially for the small fry, and are not only rich in proteins, lipids and essential nutrients and potential immunogenicity, but they also have a shorter culture cycle than other macrophytes to complete complex protein folding to form active recombinant exogenous protein, making them an ideal system for aquatic vaccines production and oral delivery (207–209). Chlamydomonas reinhardtii, Dunaliella salina, and cyanobacteria have been reported to express antigenic genes for the prevention and control of infectious diseases. For example, oral administration of D. salina and C. reinhardtii integrated with VP28 improved the survival of shrimp exposed to WSSV (210, 211). Feeds prepared with C. reinhardtii expressing Antiviral double-stranded (ds) RNA-YHV increased the survival of shrimp by 22% after a yellow head virus (YHV) attack (212). Through successful expression in Nicotiana benthamiana, Atlantic cod neuro necrosis virus (ACNNV) VLPs be effective in protecting sea bass from viral attack (213). Diatom recombinantly expressed heat shock protein (GroEL) and outer membrane protein (IglC) of Francisella orientalis (Fo), both of which enhance the immune response of Nile tilapia and improve immune protection in fish (214). To date, however, many microalgae have not become an effective and established genetic manipulation system, and no plant-produced aquatic vaccines have been commercialized (215). Therefore, further research on the production of aquatic vaccines using plant biotechnology is essential. To improve the activity of vaccines and heighten the level of induced animal immune responses, research on plant-produced vaccines is currently concentrated on increasing the concentration and purity of antigens in transgenic plants as well as addressing shortcomings in the ambiguous efficiency of modifications, such as glycosylation, methylation, and polymerization (216). In addition, animal species, animal diet, disease type, vaccine characteristics, production cycle, production scale, biosafety issues, future commercial prospects, and management difficulties need to be considered before selecting a plant expression system for fish vaccine production (5).
Overall, subunit vaccines offer a wide range of uses and are a good option for some hard-to-culture microorganisms. They can be used to protect both homologous pathogens and multiple infections by combining the antigenic genes of different pathogens. The effectiveness of subunit vaccinations to generate mucosal and cellular immunity in vertebrate hosts has significantly improved even though they are less immunogenic than inactivated vaccines due to ongoing advancements in antigen delivery and vaccine adjuvants. Subunit vaccines are currently plagued by the difficulty of expressing recombinant viral and protozoan membrane antigens in their natural structural state, as well as the production of misfolded or misprocessed proteins by microbial systems (particularly E. coli and yeast) that lack the conformational epitopes required to produce protective antibodies in the host. Furthermore, the fact that subunit vaccines are primarily administered by injection and the need for adjuvants raises the cost of their use.
3.3.2 Nucleic acid vaccine
Nucleic acid vaccine, which includes DNA and RNA vaccines, refers to the introduction of plasmids containing a gene encoding an antigen protein into the host to express the antigen protein through the host cell and induce the host cell to produce an immune response to the antigen protein to achieve immune effects. In 1996, Anderson et al. (1996) (217) prepared a DNA vaccine pCMV4-G containing the IHNV G gene and immunized trout by intramuscular injection, which improved trout resistance, launching research into the use of nucleic acid vaccines for fish. After traditional vaccines and genetically engineered subunit vaccines, nucleic acid vaccines have emerged as a hot spot for aquatic vaccine research and development. Although nucleic acid vaccines provide enhanced immune protection, are simple to prepare, save time and effort saving, are safer in an application, and have a long duration of an immune response, they are prone to immune tolerance and have a low probability of plasmid DNA integration into the host genome and causing autoimmune diseases and insertional mutations. Nucleic acid vaccines are frequently very effective in preventing viral infections, such as fish bouncing viruses (rhabdovirus), because they use the same cellular mechanisms that viruses use when they enter the host cell (218, 219). Huo et al. (2020) (220) developed recombinant plasmids pcORF25 and pcCCL35.2 to serve as a vaccine and molecular adjuvant against CyHV-2, respectively. They discovered that pcORF25/pcCCL35.2 effectively increased mRNA expression of critical immune genes (IL-1, IL-2, IFN-γ2, and viperin) and significantly inhibited CyHV-2 replication in the head kidney and spleen tissues. DNA vaccines are also effective in preventing bacterial infection in fish, such as Vibrio anguillarum (221–223), E. tarda (224), V. harveyi (225) and so on. Several DNA vaccines have been approved for marketing, and the first licensed DNA vaccine ever was a DNA vaccination for IHNV that was approved in Colombia, UK, in 2003 (226), and was approved for marketing in Canada in 2005 (218). In 2018, a DNA vaccine against the Salmon pancreatic disease virus (SPDV) was approved for marketing in Norway and the EU (227). A commercially available vaccine based on DNA plasmid has been authorized to be used in Norwegian aquaculture against Pancreas disease (PD) caused by salmonid alphavirus subtype 3 (SAV3) in Atlantic salmon SAV3 in salmon since 2018 (228, 229). Studies on DNA vaccines against parasitic fish diseases have also been reported, for example, immunization of obliquely banded grouper with DNA vaccines prepared by stimulation of Cryptococcus repressor antigens improved the resistance of the fish (230). Intramuscular injection of DNA vaccine against the parasite Ichthyophthirius multifiliis (Ich) induced significant upregulation of immune genes in the head kidney of catfish, including IgM, CD4, MHCI, TCR-α, IFN-γ, complement component 3 (C3) and Toll-like receptor (TLR), and the fish produced anti-Ich antibodies, obtaining a higher immunoprotective effect (231). A DNA vaccine against Ichthyophthirius multifiliis was also effective in immunizing channel catfish (232).
Compared to conventional vaccines, DNA vaccines are more stable, safer, and largely free of toxicity reversal. However, DNA vaccines are not mature enough and, at present, it is not possible to determine whether the host genome will integrate with exogenous DNA to disrupt the genetic homeostasis in the host. What’s more, DNA vaccines have potential side effects, including immune tolerance of the organism to the expressed antigen, chromosomal integration, and injection site inflammation (219). RNA vaccines have received significant attention in recent years, and they have many advantages. For example, RNA is not infectious, it can be degraded by normal cellular processes, and there is no potential risk of infection or insertional mutagenesis. Therefore, RNA vaccines have a promising future in aquatic vaccines (233).
3.3.3 Recombinant live vector vaccine
A live vector vaccine is a genetically modified vaccine that uses a non-pathogenic virus or bacteria as a carrier to express protective immune-related antigens and deliver the antigens to the lymphoid tissue of the intestine, generating intestinal mucosal immune responses and system immunity. It is characterized by the combination of the high immunogenicity of live attenuated vaccines and the precision of subunit vaccines. A significant benefit of the live vector vaccines is their ability to effectively induce cellular and humoral immunization and even mucosal immunization by inducing target antigen expression in vivo, resulting in endogenous antigen processing and MHC class I restriction antigen presentation. Second, the production cost of live vector vaccine is low and the risk of virulence is low. The antigen genes of different pathogens can be inserted into the vector and expressed simultaneously, thus achieving the goal of preventing multiple diseases with one needle, providing the possibility for the development of multiple or multiple vaccines (234). At present, bacterial live-carrier vaccines have been studied in fish live-carrier vaccines, such as E. coli, E. tarda, Salmonella, Listeria, Lactobacillus, Bacillus subtilis, etc. have been reported. It is worth noting that with the in-depth research on the function of gut microbes in recent years, the role of probiotics has been paid more and more attention. Probiotics are safe and non-toxic, can promote the intestinal absorption of nutrients better, enhance the body’s immunity, and also have a good adjuvant effect as a live vaccine carrier. Studies have shown that recombinant live vector vaccines constructed with Lactococcus lactis and B. subtilis can elicit a protective immune response in the host (235–238). Oral administration of Lactobacillus casei CC16 strain expressing the flaB and OmpAI antigens of A. veronii significantly induced carp-specific antibody responses and carp-specific antibody responses, up-regulated IL-10, IL-β, IFN-γ, TNF-α and other immune genes expression, and improved the fish immune protection (239, 240). The vectors of live viral vector vaccines are generally attenuated strains, and exogenous antigens are inserted into non-essential regions of the vector genome to form a new recombinant virus, with the exogenous genes being expressed concurrently with vector replication. Baculovirus (BmNPV), a pathogen of silkworm (Bombyx mori), can be used as a potential vaccine vector for clinical application (241). Baculovirus (Baculovirus) and IHNV can both be used as vaccine carriers to induce effective immune protection of the host (242, 243). It has been reported that the recombinant BacCarassius-D4ORFs-BmNPV vector vaccine can effectively protect crucian carp from CyHV-2 infection, with relative survival rates of 59.3% and 80.01% by oral or injection vaccination, respectively (244).
Although live vector vaccines have many advantages in terms of immunization mode, strain culture, and gene delivery, there are still many drawbacks, such as the fact that some bacteria with reduced virulence can still be harmful to the body’s health and that long-term immunity causes the body to become tolerant to producing antigen, which hurts the immune system. Therefore, further optimization of vector selection and immunization strategy is needed to improve the protective effect of live vector vaccines.
4 Aquatic vaccination methods
The development of protective vaccines and their proper use is essential for successful vaccination. Vaccination should be administered before pathogen exposure to allow sufficient time for immunity to develop. The route of immunization used is also selective, depending on the type of vaccine, cost, type, size, and several fish to be immunized, as well as the purpose of immunization. In aquaculture, three vaccination methods are commonly used: injection, immersion, and oral immunization (245). As shown in Table 6, each immunization method has advantages and disadvantages, and in practice, we must select the best vaccination method based on the characteristics of the vaccine, the type of animal, etc.
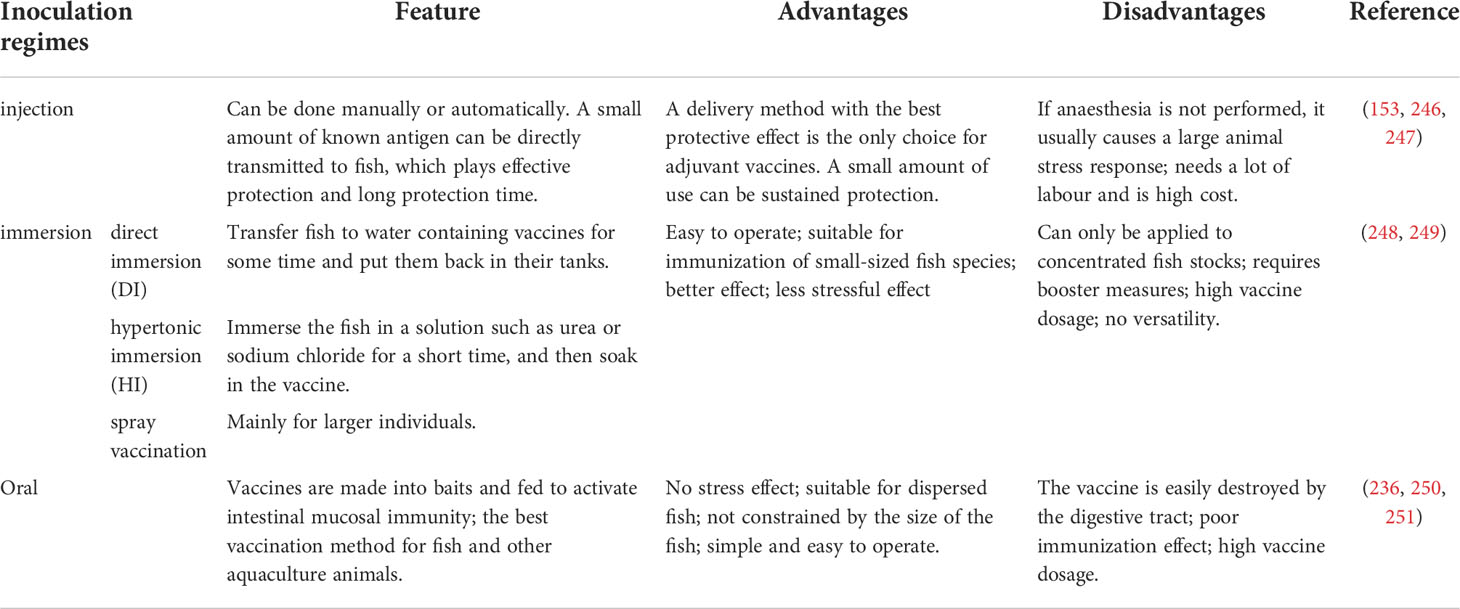
Table 6 Comparison of vaccination methods.
4.1 Injection immunization
The most effective method of vaccination is injectable immunization. It ensures that each immunized fish receives an equal amount of vaccine to stimulate the body’s immune response, allowing the immunized fish to achieve long-term and stable immune protection. This vaccination method requires fewer vaccines and is especially appropriate for inactivated, subunit, and nucleic acid vaccines, as well as mixed, monovalent, and multivalent vaccines. However, there are numerous issues with the injection immunization process, such as the difficulty of handling and, in particular, the inconvenience of immunizing small fish under 20g, but fish smaller than this are generally the most susceptible to disease. It can also harm the fish mechanically, causing ulceration at the injection site and adhesions between internal organs and the peritoneal wall. Second, the handling, anaesthesia, and injection processes not only stress the fish but also increase labour costs, particularly when immunizing large numbers of fish. Machine injection, as opposed to manual injection, can reduce stress on fish, reduce labour intensity, shorten injection time, and overcome some disadvantages of the injection immunization method itself (252). Many patented inventions of vaccination apparatus are now commercially available, which not only greatly facilitates large-scale vaccination operations by farmers, but also improves the efficiency of vaccine utilization and facilitates the vaccine development process. Injectable immunizations typically include intraperitoneal and intramuscular injections. Inactivated vaccines are frequently administered intraperitoneally, subunit vaccines usually require adjuvants, and some nucleic acid vaccines are only suitable for intramuscular immunization due to their high uptake by muscle tissue, which can induce a systemic immune response. Corbeil et al. (2000) (253) evaluated the vaccination effect via various vaccination routes including intramuscular injection, scarification of the skin, intraperitoneal injection, intrabuccal administration, gene gun, or immersion with pIHNVw-G, and found only the intramuscular injection and using gene gun induced protective immunity in Rainbow trout fry. The pSAV/HE DNA vaccine elicited a strong immune response at the injection site after intramuscular injection and provided high protection in Salmon, whereas neither the pSAV/HE alone nor the mixed inactivated low-dose virus vaccine by intraperitoneal injection elicited an immune response in Caenorhabditis elegans or improved the effectiveness of the inactivated vaccine (254). An SWCNTs-DNA vaccine encoding matrix protein provides a good protective effect against spring viremia of carp virus by intramuscular vaccination (246). Ramírez-Paredes et al. (2019) (247) developed an autologous whole-cell inactivated vaccine to effectively protect red Nile tilapia from Francisellosis by intraperitoneal injection. Xu et al. (2019) (153) developed an effective inactivated vaccine against Vibrio harzianus in turbot. They found that the vaccine not only induced humoral immunity but also enhanced the innate immune response to provide long-term effective protection by intraperitoneal injection with different concentrations of the vaccine in turbot. The commercial use of the Yersinia ruckeri vaccine without adjuvant is possible, but efficiency is insufficient, wherein the use of aluminium hydroxide is harmless and does not cause complications (255).
4.2 Immersion immunization
Immersion immunization is a unique way of vaccination for aquatic vaccines. Antigens can be uptaken by mucosal tissues such as gills, skin, lateral line and gastrointestinal tract, which can quickly and effectively activate the mucosal immune system of fish, and then spread to the peripheral blood, kidney and spleen and other systemic immune tissues through blood circulation to generate a systemic immune response (256). Its advantages are that it causes less mechanical damage and stress stimulation to fish bodies, is easy for herd immunity, has less labour intensity, and less time-consuming immunity. It is suitable for fry and smaller fish. The application of immersion vaccine has been reported in many fish, such as Anguilla japonica (257), Atlantic cod (258), grouper (259), Crucian carp (260), and rainbow trout (261). Immersion immunity mainly includes Direct immersion (DI), Hypertonic immersion (HI), Flush and Spray, etc (262). DI involves transferring fish to water containing the vaccine for some time and then returning them to the breeding pond. This method is simple and suitable for sterilization vaccines and attenuation vaccines and causes less stress to fish bodies. Therefore, it can improve the uptake of antigens by prolonging the immersion time, thus improving the effect of immersion immunity and reducing the cost of vaccines (263, 264). Many studies show that the immune protection of direct immersion immunization is low, and needs to be strengthened to achieve the ideal immune effect (265). The hyperosmotic method is to soak the fish in 3% ~ 5% salt solution for 5 minutes, then move the fish herd to the vaccine solution for immersing, to force the antigen to infiltrate into the fish by the change of osmotic pressure (248, 266). Ultrasonic treatment can increase the permeability of biological tissues such as skin and muscles, and the number of antigens entering a fish’s body can be significantly increased by ultrasound in spray (267, 268). Although hypertonic and spray methods can improve the efficacy of immersion vaccines, their application is limited by stress on fish bodies. The uptake of antigen by immersion is limited, so the immune effect of immersion on fish is lower than that of injection immunization. Moreover, the immersion environment has a great influence on the vaccine effect, and it is difficult to control the amount and time of immersion. Immersion usually requires a large amount of vaccine, and in practice, vaccine cost must be a primary consideration. In recent years, many studies have focused on how to improve the effect of immersion immunity using different methods. Examples are booster vaccination, administration of immunostimulants/adjuvants, use of delivery vehicles, preconditioning by physical methods, use of novel live attenuated vaccines and DNA vaccines, etc (269–271). Mucoadhesive polymers are widely used in drug delivery and can also be developed and utilized as vaccine carriers in immersion immunization (272–274).
4.3 Oral immunization
Oral administration is considered to be the most practicable form of immunisation for aquatic vaccines and is suitable for a wide range of vaccine types including inactivated, live attenuated and nucleic acid vaccines. Compared to injectable vaccination, oral administration is not restricted by the size and age of the animal, can reduce stress and mechanical damage to the animal, can provide some savings in labour costs and meets animal welfare requirements (275). At present, aquaculture is most intensive and factory-based, and oral immunization is more suitable for large-scale farm animals than other immunization methods. However, the development and application of oral vaccines for fish are not yet widespread. The main constraint is that the vaccine is susceptible to destruction by digestive tract enzymes, cannot induce intestinal mucosal immunity and systemic immune responses, and may induce immune tolerance. In addition, the number of orally immunised fish and the dose of vaccine cannot be effectively guaranteed, resulting in a lower immune protection effect than that of injection immunization at present. To improve the immune effect of oral vaccines, it is necessary to avoid naked antigens as much as possible. At present, the common materials used to encapsulate vaccines include alginate, chitosan, liposomes and other polymeric microspheres and biofilms (275–278). It has been shown that DNA vaccine can improve fish defence against disease, but the immune effect of orally administered naked DNA vaccines is not ideal, instead, the immunological effect of the vaccine can be significantly improved after encapsulating with sodium alginate (279–281). In addition, the recombinant subunit vaccine can also be administered orally after being encapsulated. Caruffo et al. (2016) (282) used brewer’s yeast to simultaneously express the surface protein of the Hemagglutinin-esterase virus and the F protein of the salmon anaemia virus (ISAV) and encapsulated it in a cationic polysaccharide matrix capsule. The salmon obtained a survival rate of 66.7% after feeding. Biofilm (BF) are Extracellular polymeric substances (EPSs) attached to the surface of microorganisms and serve as a protective barrier for microorganisms to enhance their tolerance to environmental stresses, such as desiccation, osmotic pressure, UV radiation, disinfection and antibiotic therapy (283). The biofilm can protect the antigen components of the vaccine from the destruction of the digestive fluid as it reaches the gut lymphoid tissue, allowing the oral vaccine to perform its proper immune function (284, 285). Due to the high cost of the encapsulated material and its large amount of use, it is unrealistic in practical application. And the mechanism of immune activation in the animal intestinal mucosa is not clear, some materials may cause non-specific reactions and reduce vaccine effectiveness (286, 287). Therefore, in recent years, the research of another biological carrier vaccine has become the focus of oral vaccine research. Genetic engineering allows the expression of exogenous antigen components through non-pathogenic microorganisms. Reported biological carriers include insects (halogen larvae, beet noctule, silkworm pupae, etc.), probiotics (lactobacillus, bacillus subtilis, yeast, etc.), plants (Chlamydomonas, fishy algae and potatoes, etc.) and viruses (288). Compared to traditional vaccines, biocarrier vaccines are more effective at presenting antigens and antigen selectivity (205, 289). Different vector vaccines can be prepared according to different antigen properties, or insert multiple exogenous antigens into the same vector vaccine to achieve the effect of one vaccine against several diseases. It is noteworthy, however, that biological vector vaccines often receive interference from vector-derived antibodies during booster immunization, and that the components of vectors such as viruses can also pose a potential threat to the host. Therefore, the selection of a suitable biocarrier for the antigen and the improvement of the safety and applicability of biocarrier vaccines have become prerequisites for the mass production of biocarrier vaccines.
Currently, the immune response to oral vaccines is still different from that of vaccines administered via injection, and in some cases, it is just half as strong. Further in-depth research is required on several topics, including the immune mechanism of oral vaccines, in particular the immune effect on the intestinal mucosa, the handling of antigens during vaccine production, the effective dose, the type of vaccine coating material, carrier, and adjuvant, the form of the vaccine, as well as the duration of immunization, the frequency of immunization, and the physiological developmental stage of the experimental fish.
5 Mechanism of aquatic vaccines
Teleost fish possess both primary and secondary lymphoid tissues. Primary lymphoid tissues include the thymus, where T-cell development occurs, and the head kidney, which performs hematopoietic functions similar to mammalian bone marrow. Secondary lymphoid tissues include the spleen and mucosa-associated lymphoid tissue (MALT). There are four kinds of MALTs, nasopharynx-associated lymphoid tissue (NALT), gill-associated lymphoid tissue (GIALT), skin-associated lymphoid tissue (SALT), and gut-associated lymphoid tissue (GALT), which contain specific adaptive immune systems (290, 291). In general, teleost MALT does not have organogenic lymph nodes or lymphatic aggregates but consists of a diffuse network of myeloid and lymphoid cells (292). However, there are some T lymphocyte aggregates within the GALT, called interbranchial lymphoid tissue (ILT), despite the lack of a complete regional growth centre for B and T cells (293). At present, the mechanisms of antigen uptake and antigen presentation in the teleost MALT are unclear. In all organisms, the mucosal surface is the main site of entry for pathogens, while the mucosal surface is covered with an immune-enhanced mucus protective layer, which acts as the first line of defence against pathogens. Fish mucus contains a variety of immune-related factors such as lysozyme, defensins, immunoglobulins, etc (294). Teleost fish also possess an adaptive immune system that relies on somatic recombination of germline-encoded VDJ fragments to produce a large number of antigen receptors expressed on T and B lymphocyte membranes (295). Studies have shown that teleost MALT has a large number of antigen-presenting cells in mucosal sites, including dendritic cells (DCs), macrophages, IgT/Z+ B cells and granulocytes, all of which have been shown to have antigen uptake functions (296–298).
5.1 Mechanism of injection immunization
The immune organs of fish are simpler, mainly including the thymus and spleen, as well as the anterior kidney, which functions like bone marrow and lymph nodes, and the scattered lymph-like tissues containing lymphocytes, monocytes, macrophages, granulocytes, platelets, mast cells, and non-specific cytotoxic cells (299). Fish have mammalian-like T and B lymphocytes. Different subpopulations of fish T cells have not been well characterized, but homologs of CD8 and CD4 have been reported, while B lymphocytes were also found to have phagocytic activity (300, 301). Fish MHC I, TCR and TCR co-receptor CD8 are homologous to mammals, suggesting similarities in the antigen presentation process, i.e., antigens presented by MHC are recognized by TCR and activate immune responses. In addition to TCR-specific recognition of MHC I/II, natural antigens are also recognized directly by B lymphocyte receptors. Currently, it is generally accepted that the specific immune response in fish is the basis of Injectable immunization. When the antigen invades the fish, it is presented by antigen-presenting cells (APCs) to T cells, and then presented to B cells by T cells, leading to B cell activation, proliferation, and differentiation into plasma cells and memory cells, which eventually produce a large number of antibodies, resulting in a specific immune response. Antibodies are the main functional factors of the fish immune system, and in recent years four classes of Ig have been identified in fish, namely IgM (302), IgD (303), IgZ/T (304, 305) and IgM-IgZ (306). Injection immunization mainly causes the immune system of fish to produce IgM. After intraperitoneal immunization, mucosal IgM antibodies and pIgR responses are triggered, and the pIgR produced by local plasma cells in the lamina propria (LP) mediate the IgM-antigen complex through the intestinal epithelium into the intestinal mucus (307). Antigens being injected intramuscularly can evoke a local adaptive immune response, however, the mechanisms regarding the dynamics and transport of antigens after injection have not been elucidated. There are differences in the immune response of various organs caused by injecting immunity, mainly inducing a response in the systemic immune organs such as the spleen and kidney, and a weaker immune response in the mucosa tissues such as gill, skin and intestinal (308). Interestingly, we found that the difference in antigen uptake after intraperitoneal and intramuscular injection followed the same pattern: the highest antigen uptake was in the spleen and head kidney, followed by blood, liver, and gills, and the least in the hindgut, muscle, and skin. However, the peak antigen uptake in the spleen, head and kidney, liver, blood, and hindgut was significantly higher in the intraperitoneal injection than in the intramuscular injection. The peak antigen uptake in skin and muscle was significantly higher in the intramuscular injection (309). Moreover, the expression of antigen presentation-related genes (CD4-1, MHC IIα, CD8α, MHC Iα) in the spleen, head kidney, liver and hindgut of flounder, and the proportion of sIgM+ cells in the spleen and peripheral blood were higher in the intraperitoneal injection group than in the intramuscular immunization group (310, 311). Both cellular immunity (including cytotoxicity) and humoral immunity are activated by nucleic acid vaccination. Cellular immunity is primarily initiated by antigen-presenting cells (APCs) such as macrophages and dendritic cells (DCs) (312, 313). DNA motifs and mRNA transcripts express immunogenic proteins in fish that simulate infection by intracellular pathogens and allow the presentation of exogenous antigenic peptides on the APC surface via major histocompatibility complex (MHC) class I. Subsequently, APC at the site of administration may trigger immune cells (314, 315). Antigen-presenting cells can also absorb soluble antigens released from transfected myoblasts (or other cells) or in the presence of antigenic apoptotic vesicles, delivered to MHC class II molecules on the cell surface. Antigenic peptides in MHC class I and MHC class II are recognized by the T cell receptor (TCR), which activates CD8+ T cell (cytotoxic T cells) and CD4+ T cell (helper T cells) responses, while the adaptive humoral immune response is manifested by B lymphocyte activation and antibody production. The unmethylated CpG motif is identified by the vertebrate immune system as a “foreign” and “dangerous” signal. For the vectors of nucleic acid vaccine derived from bacterial and viral DNA, which carries unmethylation CpG motif can be identified by the body as a pathogen-related molecular pattern, activating the macrophage, and B cells to induce the humoral immune also can directly activate monocytes, macrophages and dendritic cells to secrete Th1 cytokines to induce a cellular immune response (316, 317). In addition, CpG motifs can be recognized by Toll-like receptor 9 (TLR9) to activate leukocytes by the uptake of DNA into lysosomes containing specific receptors (318, 319). In addition, humoral factors such as complement, cytolysin, interferon, lectin, etc. that mediate fish’s non-specific immunity are important ways for fish bodies to resist the external environment. While macrophages were found to play the phagocytic function of non-specific immunity as well as phagocytosis, processing and presentation of antigens, indicating that specific and non-specific immunity in fish complements each other in the process of injection immunization. Studies have found that phagocytic cells similar to mammalian M cells are distributed in the hindgut of fish. After intraperitoneal injection, antigens are recognized, processed and presented by these cells, thus causing immune responses in fish.
5.2 Mechanism of immersion immunization
The gills and skin mucosa not only have non-specific immune functions but also have specific immune response functions (187, 320). The mucosal tissues and their secreted mucus contain abundant non-specific immune factors such as hydrolase, enzyme/transfer factor, chitin, C-reactive protein, lectin and specific antibody proteins, which together constitute the first line of defence against the invasion of pathogenic microorganisms in fish (321, 322). It was found that there were a certain number of T and B lymphocytes, mucous cells, cystic cells and antibody-secreting cells in the skin of fish. Similarly, various immune cells such as lymphocytes, macrophages, goblet cells and neutrophils were found in gill tissues. The level of antibodies and the number of antibody-secreting cells on the surface of gills, skin and intestinal mucosal tissues were significantly increased after vaccine immersion, which indicates immersion immunization can cause an obvious immune response (323, 324). Numerous studies have now shown that mucosal immunity can produce a local immune response independent of the systemic immune response. Immersion immunity mainly induces a mucosal immune response in fish, while systemic immune response is weaker and later than mucosal immunity. During immersion immunization, antigens are first taken up by mucosal tissues such as gills, skin and intestine, which activate lymphocytes in mucosal tissues to produce antibodies. Through blood circulation, antigens are transported to tissues such as the spleen and kidney and activate systemic immune responses (256). Antigen uptake was positively correlated with immersion concentration in a certain range, and antigen particles on the gills were mainly taken up by phagocytes, while antigen recognition and presentation in the skin were presumably done by macrophages (325). Three immunoglobulin isotypes (IgM, IgT and IgD) were detected in the gills, skin and nose of fish, with IgT being the predominant functional Ig isotype in these mucosal tissues and the strongest response of IgT in immersion immunization (326, 327). Some researchers believe that gills have an independent immune response, because MHC II -positive cells were detected in a variety of fish gill tissues, and found that the proportion of ASC on the gills was significantly higher than that in peripheral blood of immersion immunized fish, and the number of antibody-secreting cells at the different time showed a kinetic response pattern (328–330).
5.3 Mechanism of oral immunization
Current studies have shown that oral immunization can produce both local mucosal immunity and systemic immune responses, but the intestinal mucosal immune response is predominant (331). The possible mechanism is that the macromolecular antigens were swallowed by the hindgut epithelial cells, and the small molecule soluble antigens were infiltrate the blood through the intestinal mucosal cell gap. While large molecule particle antigens are the first uptake by macrophages into the cells after processing, then transported to the relevant lymphatic tissues or cells, then transported to the circulatory system, and finally transported to the lymph nodes via blood or lymphatic fluid to produce the relevant immune response (332). A large number of lymphocytes, mainly including macrophages, granulocytes and plasma cells, are distributed in the entire intestine of fish, especially in the hindgut (333, 334). It is now generally accepted that the hindgut is the main site of antigen uptake and processing in fish and that antigens entering the hindgut are taken up by macrophages to induce an immune response in the fish (334, 335). Companjen et al. (2005) (336) found lymphoid tissue in the gut of Dicentrarchus labrax, and the number of lymphocytes showed an obvious numerical gradient from the digestive tract to the anus. This phenomenon suggests that the hindgut produces a stronger immune response than any other part of the intestine, and it is presumed that oral vaccines work mainly in the hindgut. The intestinal mucosa layer of fish can be divided into two layers, the epithelial layer and the lamina propria, in which granulocytes, macrophages and other leukocytes and Ig+ cells mainly exist in the lamina propria of the intestinal mucosa layer, while a large number of Ig- cells also exist in the epithelial layer of the mid and hind intestine (337, 338). In addition, T cells are distributed in the epithelial layer of the intestinal mucosa, while B cells are distributed in the lamina propria of the intestinal mucosa to participate in the mucosal immune response. Mucosal immunoglobulins (e.g., IgT or IgZ) are relatively new findings in fish immunology. In rainbow trout intestine, IgT+ and IgM+ lymphocytes account for approximately 54% and 46% of the total B-cell population, the IgT/IgM ratio is higher in intestinal mucus than in serum, and the percentage of bacteria phagocytosed by IgT (48%) is significantly higher than that of IgM (24%), these studies suggest that IgT is the major immunoglobulin in intestinal mucus (339). In orally vaccinated rainbow trout intestine, IgM+ and IgT+ cells were found to be distributed throughout the intestine, with IgM+ cells located mainly in the lamina propria (LP) and IgT+ cells localized mainly as intraepithelial lymphocytes (IEL), and a significant increase in the number of IgM+ and IgT+ IEL was observed in the pyloric cecum region. Not only that, the authors found that B cells responsive to vaccination were detected in adipose tissue in the digestive tract, suggesting that these cells surrounded by adipocytes also play a role in mucosal defence (340). Oral tolerance is a low response to fed antigens and a result of inhibition of cellular and/or humoral immune responses, a phenomenon also prevalent in fish (341, 342). Many factors, including excessive antigen dose, inhibition of Treg production, repeated administration, low temperature, antigen type, and genetic effects, are prone to induce tolerance in fish (343, 344). Decreased antibody response after repeated exposure to antigens may be associated with the induction of upregulation of FoxP3, TGF-β and IL-10 (345).
6 Aquatic vaccines adjuvants
In recent years, immunization control has become an important initiative for aquatic animal disease control, and there has been rapid development of aquatic vaccines and more diversified vaccine types. Traditional inactivated vaccines and emerging recombinant vaccines have problems with high antigen dosage, short duration of immunity, and low immune efficacy, and cannot achieve the desired immunization effect when used alone. An adjuvant is a non-specific immune enhancer that is used before or simultaneously with an antigen to enhance the body’s immune response to the antigen or to alter the type of immune response. The use of adjuvants can reduce the immunization dose, lower the cost of vaccines, reduce vaccine irritation, enhance the body’s immune response to antigens, prolong the protection time, etc. The ideal adjuvant enhances the effectiveness of the vaccine, is free of side effects, and is easily available and universally applied. The development of new aquatic vaccines requires new adjuvants to improve the effectiveness of vaccine use. An appropriate adjuvant enables an effective vaccine to produce efficient immune protection by different vaccination methods, such as the Fno vaccine, which can also improve the protection of Nile tilapia by oral administration with an oral adjuvant (346). New adjuvants should be studied for vaccine type, mode of action and carrier, immune efficiency, the durability of immune effect, and toxicity of the adjuvant itself, to match vaccines better and improve immune protection. Available evidence suggests that adjuvants may generate an immune response in a variety of ways, such as the sustained release of antigen at the injection site and recruitment of immune-related immune cells, upregulation of cytokines and chemokines, promotion of antigen uptake and proliferation of antigen-presenting cells (APCs), and activation of inflammatory responses (347). Moreover, the majority of adjuvants are currently on the market for injectable vaccines, while immersion and oral vaccines are less available, which limits the development of immersion and oral vaccines to a certain extent. Compared with injectable vaccines, immersion and oral vaccines have the advantages of convenience, low stress and low cost, so the application prospect of immersion and oral vaccine adjuvants is promising. Based on their mode of action, adjuvants can be categorized broadly into antigen-presenting and immune-enhancing types.
6.1 Antigen-presenting adjuvants
Antigen-recurrent adjuvants include aluminium salts, oil emulsions and granules, which are usually presented to the immune system in the form of endocytosis, pinocytosis, and membrane fusion to promote an immune response to antigens. Among them, aluminium adjuvant has a powerful delivery effect and is widely used in many types of vaccines. Aluminium adjuvants can enhance the expression and duration of major histocompatibility complexes (MHCs) on the surface of dendritic cells, promote antigen delivery to intracellular antigens, and thus produce adaptive immune responses (348, 349). Aluminium adjuvants cause less tissue damage and few adverse immune reactions and hypersensitivity reactions and are considered safe adjuvants. Currently, most of the vaccines used in production are aluminium-adsorbed vaccines (350).
Oil adjuvants are the most commonly used adjuvants in aquatic vaccines, and almost all salmon are vaccinated against oil adjuvants. Of these, the complete Freund’s adjuvant (CFA) is composed of heat-inactivated mycobacteria and mineral oils containing surfactants. The adjuvants act as carriers and reservoirs for the presentation of antigens in animals after injection, allowing slow release of antigens and continuous stimulation of strong Th1 and Th17 reactions in animals through MyD88 pathways to produce higher immunity (351). CFA is the most widely used adjuvant (352). CFA causes fish in long-term antigen environmental stimulation, although it improves antibody levels, it also has side effects associated with inflammatory response, and usually causes damage in injection sites such as the pyloric blind sac, spleen, muscles, oesophagus, etc., affecting the growth of fish (353). Incomplete Freund’s adjuvant (IFA) does not contain mycobacteria, can improve cell phagocytosis, promote leukocyte infiltration and cytokine production, and produce long-lasting immune protection, IFA also has serious toxicity (354). The MONATINE series of mineral oil-based adjuvants developed by SEPPIC is easy to emulsify, low viscosity, stable and safe to absorb, and widely used in fisheries vaccines (355).
Microparticle is a new type of delivery vector widely used in vaccine adjuvants, including virions, nanoparticles, polymers, etc. Antigens such as polypeptides, proteins and DNA are transported inside particles to lymphatic organs through covalent binding or physical burial and are released continuously through void diffusion and particle degradation. At present, covalent coupling of antigen and micro grain has become a new technique, which can reduce antigen dose, protect antigen integrity, and improve antigen stability and immune efficiency (356–358). Studies have shown that the micro granule adjuvant effect is closely related to micro granule size, and the smaller the particle, the higher the immune efficiency. For example, microparticles often induce humoral immunity and nanoparticles mainly produce cellular immunity (359–364).
Polylacticacid (PLA) is a biodegradable histocompatibility polymer. PLAs can quickly reach the cytoplasm by phagocytic action or clathrin-mediated endocytosis causing an early immune response (365). As an adjuvant, polylactic-co-glycolicacid (PLGA) can cause cellular and humoral immunity and improve antigen delivery. It also has the advantages of good histocompatibility, biodegradability and non-toxicity. In an application of an outer membrane protein (OMP) based vaccine against A. hydrophila, the PLGA adjuvant showed a better vaccine immune effect compared with the IFA adjuvant (366).
6.2 Immune-enhancing adjuvants
Immuno-enhanced adjuvants indirectly cause a congenital immune response, or directly activate a congenital immune response, mainly by activating the pathogen recognition receptor (PRRS) (367). PRRS is mainly divided into membrane receptors (including Toll-like receptor, TLR, C-type lectin, etc.), intracellular receptors (including NOD-like receptor, RIG-I-like receptor) and secretory receptors, which are expressed in a variety of immune cells, to activate congenital immune responses by identifying PAMPs, or by stimulating the secretion of inflammatory cytokines and activating antigen-transitive cells, thus establishing an adaptive immune response. A total of 17 different types of TLR genes are found in fish, far more than in mammals, and immune enhancers are expected to play an important role in aquatic vaccines.
Saponins are natural steroids or terpenoids, widespread in animals and plants, low-grade marine animals and bacteria. Saponins can stimulate the secretion of specific antibodies, enhance the phagocytosis of macrophages and respond to exogenous antigens by cytotoxic t-lymphocytes (CTL). QS-21 is a saponin extracted from a soap tree that enhances the secretion of Th1 cytokines (IL-2 and IFNγ) and IgG2-a. In aquatic, saponins can enhance the non-specific immunity of shrimp and stimulate fish growth, but high-dose saponins can easily cause severe tissue damage in fish when using Intraperitoneal injection (368, 369). In the study of immersion immunity, it is shown that saponins can significantly improve the delivery of vaccine antigens in the skin and other tissues, trigger the inflammatory response, and improve the effect of non-specific immune levels in fish (370). Usually, the longer the immersion immunization time, the better the immunization effect and the adjuvant can accelerate the absorption of antigens, shorten the immunization time and reduce the stress on the fish body (371). Therefore, the application of saponins as an immersion vaccine adjuvant is of great value, but it is necessary to solve its instability in the water phase.
Cytokines are a class of small molecular proteins secreted by immune cells and non-specific immune cells, which play an important role in pro-inflammatory response and immune system regulation. The structural function of fish cytokines is similar to that of mammals, including IL, IFN, tumour necrosis factor and chemokine, etc., with good adjuvant effects. Studies have shown that a large number of B cells and cytotoxic T cells accumulate at the muscle site where IFN is injected. Different IFNs have different biological activities, while IFN, as an autoantigen, can maintain long-term adjuvant activity without attack by the immune system (372, 373). Cytokine IL (e.g., IL-8, IL-6) has been shown to upregulate genes associated with inflammatory response, humoral immunity and cellular immunity, improve immunoprotection and maintain long-term vaccine protection (370).
Others such as β-glucan, mannose (374, 375), chitosan, (376, 377)monophosphoryl lipid A (MPLA), flagelloprotein, CpG ODNs (CpGoligodeoxy nucleotides), poly-nosinicopolycytidylicacid, Poly (I-C) (68), insect hemocyanin (378), propolis (379), complement, etc. can also be used as adjuvants. Among them, β-glucan is the best adjuvant currently used in fish oral vaccines (380, 381). β-glucan or anisodamine as adjuvant β-glucan can enhance the immersion immune efficacy of inactivated CyHV-2 vaccine in gibel carp (382). More work is needed to present propolis as a proper candidate for the development of a natural adjuvant in aquatic vaccines (383). Clinically, MPL has been used in combination with liposomes or emulsions, demonstrating a good adjuvant effect (384). The inactivation vaccine of E. tarda in mixed flagelloprotein can significantly improve vaccination protection on flounder (385), which proves that flagelloprotein is an important candidate for the fish vaccine.
7 Challenges and prospects for aquatic vaccines
At present, the number and variety of aquatic vaccines with production approvals worldwide are relatively limited, and they cannot fully cover the major diseases of the main breeding species, which is insufficient to support the development of a comprehensive integrated disease immunization and control system. Inadequate supply of efficient vaccine varieties, as well as a lengthy development process, reduce the overall effect of disease immunization and control in the aquaculture production process and reduce farmers’ confidence in the industrial application of vaccines, which harms vaccine promotion and popularization, as well as investment in the fish vaccine industry. Given the current state of fish vaccine development, more research in the field of fish vaccine engineering is required, primarily to address the following technical challenges and accelerate the development of high-efficiency, multiplexed aquatic vaccines.
7.1 Research on types of fish vaccine and vaccination routes
Most vaccines are currently administered via injection, which is undeniably a very effective way to vaccinate expensive nucleic acid vaccines and subunit vaccines and is practical for the majority of vaccine types. However, given the small size of aquatic animals and the large number of vaccinations, oral and immersion vaccines appear to have more potential. While immersion vaccination necessitates vaccines that are inexpensive and simple to prepare in large quantities, such as whole bacteria-inactivated vaccines, attenuated vaccines, and so on. It is especially suitable for mass immunization of juvenile fish, which have tender skin and easily absorb antigens, whereas slightly larger fish must be vaccinated with the help of adjuvants, penetrants, carriers, and so on to achieve the desired effect, and research in this area is still limited. Oral immunization is the simplest and quickest method of immunization, with a low workload and little stress on the fish, and it is not limited by fish size, species, location, or time, making it a more ideal method of vaccine administration. However, current research indicates that an oral vaccine is less effective than injectable immunization because the integrity of the antigen is destroyed in the digestive tract, resulting in a weaker response from the intestinal mucosal system; second, the mucosal immune system of aquatic animals is not perfect. Nonetheless, antigens absorbed by mucosal tissues are transmitted to systemic immune tissues, resulting in a systemic immune response. As a result, future research on the mechanism of mucosal immunity and the relationship between mucosal and systemic immunity will be required. Furthermore, more detailed research is required on antigen treatment in vaccine production, effective doses, vaccine coating materials, carriers and adjuvants, vaccine forms, as well as the duration of immunization, the number of immunizations, and the physiological developmental stages of experimental fish.
7.2 Improve the evaluation system of fish vaccine
Survival of animals after the attack was the most primitive method for evaluating the effectiveness of vaccines. With the advancement of flow cytometry, monoclonal antibody preparation technology, and multi-omics, vaccine efficacy can now be evaluated comprehensively by detecting changes in gene and protein levels, as well as antibody and cellular responses. In general, vaccine evaluation systems have been rapidly developed, but more in-depth studies, particularly at the cellular response level, are required. The investigation of vaccine response at different levels in the animal organism not only evaluates vaccine effectiveness but also promotes in-depth research on immune mechanisms in aquatic animals. More and more immune cell types are being discovered and the role these cells play in the vaccine immunization process should be appreciated. The lack of antibody probes for fish cells has resulted in many more cells being limited in their detection during vaccine immunization. Despite the current high cost of single-cell sequencing, it is undeniably important to assess vaccine efficacy at the single-cell level, as this will aid in the development of efficient vaccines.
7. 3 Strengthen basic research related to fish pathogens and diseases
With a global shortage of fishery resources, an increasing number of aquatic animals are shifting from wild to factory farming, resulting in an increase in diseases that is becoming increasingly complex. The complex farming environment has resulted in disease outbreaks, with the majority of infections being conditional pathogenic and multi-pathogenic. Serotyping of fish epidemic pathogens is the foundation for effective vaccine research, and the lack of efficacy of many currently promoted vaccines is directly related to significant mutation of the relevant epidemic pathogens. However, the rapid development of genomic sequencing technology in recent years has made molecular typing and systematic identification of pathogens much easier, which is very helpful for ongoing research on pathogen serology and epidemiological patterns. Furthermore, there is a need to expand research into the mechanisms of pathogenic conditions that cause disease and disease outbreaks. Environmental factors such as temperature, dissolved oxygen, salinity, and nutrient salts may be causative factors of fish disease outbreaks in the “triad” system of pathogen, environment, and host, and an in-depth understanding of the signal transduction mechanism of pathogenic bacteria to related factors can lay a solid theoretical foundation for vaccine development and integrated immune control of diseases.
7. 4 Promoting the molecular design of efficient aquatic vaccines using multi-omics technologies
The screening of critical antigens is required for the efficient design of novel live attenuated, nucleic acid, and genetically engineered vaccines. Furthermore, with the rapid development of various omics technologies (genome, functional genome, proteome, and metabolome) and genome editing technologies, the genetic background and genomic information of various pathogens, as well as the dynamic process of pathogen infection, are better defined, allowing for the systematic screening of genetic targets for highly effective vaccines, particularly attenuated targets and safety targets of live attenuated vaccines. Furthermore, in recent years, structural biology has resolved an increasing number of critical protein structures of pathogenic bacteria, allowing for more precise screening and design of antigenic targets, which will greatly facilitate the molecular design of efficient vaccines, the discovery of antigenic epitopes, and the development of subunit vaccines, multiplex vaccines, and so on. There is no doubt that reverse genetics and reverse vaccinology are important directions for future development, and nucleic acid vaccines such as DNA vaccines, live vector vaccines, and mRNA vaccines have emerged as hot spots for viral infectious disease prevention. Synthetic biology should be used in the future to overcome key technical bottlenecks in the design and delivery of nucleic acid vaccines. Simultaneously, the mechanism of action and application potential of nucleic acid vaccines in the immune protection of fish in theropods should be investigated, as should the synergistic effects of different vaccination/delivery routes on systemic and local mucosal immunity.
7. 5 Construction of a technological platform for fish vaccine engineering research
Fish vaccine development is a complex systemic project that necessitates additional in-depth research on the immune system of fish, particularly on immune response generation, immune memory, immune tolerance after fish vaccination, and the homeostatic balance and conversion mechanisms of natural, adaptive, and trained immunity. Meanwhile, systematic research on vaccination protocols, disease early warning, animal models, vaccine adjuvants, immune boosters/modulators, therapeutic vaccines, manufacturing, and registration protocols should be strengthened. To promote the efficient development and application of fish multiplex vaccines, a complete and systematic engineering technology platform for aquatic vaccines can be formed. Vaccination can provide complete disease prevention and control coverage, particularly for key fish culture species and major diseases.
In general, there is a lack of a large amount of experimental data and information to support the immune mechanism of vaccine timing, immunization mode, number of immunizations, and time of booster immunization due to imperfect research on the basic immunology of aquatic animals, combined with the large range and variety of species spanned by aquatic animals. The aforementioned factors have severely hampered the development and application of aquatic vaccines. Therefore, it is critical to strengthen basic theoretical research on aquatic vaccines. The structure, function, and biological properties of pathogens, the technical basis and methodology of fish vaccine design and production, and the relationship between the immune effect of fish vaccine and the environment and organism will all be important foundations for fish vaccine development.
8 Conclusion
In recent decades, aquatic vaccines have been developed with some success and their application in aquaculture production has become increasingly popular. The promotion of aquatic vaccines can reduce the use of drugs, reduce diseases, protect the environment and improve the quality and safety of aquatic products. Although some aquatic vaccines against bacterial and viral diseases are currently available on the market, due to the numerous serotypes and easy variation of most pathogens, the strains isolated from different regions and fish species vary significantly, resulting in uneven vaccine effects. Parasitic and fungal diseases are causing increasing damage to aquaculture, but there is a serious lack of commercial vaccine. Genetic engineering vaccines consisting of protective antigen genes and a newer generation of nucleic acid vaccines have shown better immunity, but many studies are still in the experimental stage and fewer vaccines are commercially available. Combining vaccines with immunostimulants such as probiotics and herbal additives to improve the animal’s immune effect is an important measure. In addition, important progress has been made in recent years to address disease problems by breeding resistant strains of aquatic animals. On this basis, exploring the functional genes and proteins of resistant strains through gene and proteomics, resolving the differences in animal immune systems, and revealing the mechanisms of pathogenic infestation and pathogen-cell interaction will provide important theoretical guidance for vaccine design. Overall, it is a long process for the aquatic vaccine from research to commercialization, in which the lack of technical content, cumbersome approval procedures, and insufficient promotion efforts are seriously hindering the development of the aquatic vaccine industry. At present, many aquatic vaccine developments are in the clinical trial stage, and it is believed that there will be many commercial vaccines available in the next few years, which will greatly promote the development of the aquaculture industry.
Author contributions
YD, LM and JC designed the article framework and content. YD and XMH wrote the review content. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (41906107), the Key Laboratory of Marine Biotechnology of Fujian Province (2021MB04), and the Key Scientific and Technological Grant of Zhejiang for Breeding New Agricultural Varieties (2021C02069-1).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Huy DTN, Nam VQ, Hanh HT, Minh PN, Huong LTT. A review and further analysis on seafood processing and the development of the fish pangasius from the food industry perspective. Food Sci Technol (2022) 42:e7642. doi: 10.1590/fst.76421
2. Mondal H, Thomas J. A review on the recent advances and application of vaccines against fish pathogens in aquaculture. Aquac Int (2022) 30:1971–2000. doi: 10.1007/s10499-022-00884-w
3. FAO. Food and agriculture organization of the united nations (2022). Available at: http://www.fao.org.
4. Sneeringer S, Bowman M, Clancy M. Economic research report 290026. United States: Department of Agriculture, Economic Research Service (2019).
5. Su H, van Eerde A, Steen HS, Heldal I, Haugslien S, Ørpetveit I, et al. Establishment of a piscine myocarditis virus (PMCV) challenge model and testing of a plant-produced subunit vaccine candidate against cardiomyopathy syndrome (CMS) in Atlantic salmon salmo salar. Aquaculture (2021) 541:736806. doi: 10.1016/j.aquaculture.2021.736806
6. Chukwu-Osazuwa J, Cao T, Vasquez I, Gnanagobal H, Hossain A, Machimbirike VI, et al. Comparative reverse vaccinology of piscirickettsia salmonis, aeromonas salmonicida, yersinia ruckeri, vibrio anguillarum and moritella viscosa, frequent pathogens of Atlantic salmon and lumpfish aquaculture. Vaccines (2022) 10:473. doi: 10.3390/vaccines10030473
7. Su H, Yakovlev IA, Van Eerde A, Su J, Clarke JL. Plant-produced vaccines: future applications in aquaculture. Front Plant Sci (2021) 12:718775. doi: 10.3389/fpls.2021.718775
8. Duff DCB. The oral immunization of trout against bacterium salmonicida. J Immunol (1942) 44:87–94.
9. Dhar AK, Manna SK, Thomas Allnutt FC. Viral vaccines for farmed finfish. Virusdisease (2014) 25:1–17. doi: 10.1007/s13337-013-0186-4
10. Dixon P, Stone D. Spring viraemia of carp. Fish Viruses Bact Pathobiol Prot (2017) 79:90. doi: 10.1079/9781780647784.0079
11. Wessel Ø, Haugland Ø, Rode M, Fredriksen BN, Dahle MK, Rimstad E. Inactivated piscine orthoreovirus vaccine protects against heart and skeletal muscle inflammation in Atlantic salmon. J Fish Dis (2018) 41:1411–9. doi: 10.1111/jfd.12835
12. Miccoli A, Manni M, Picchietti S, Scapigliati G. State-of-the-art vaccine research for aquaculture use: The case of three economically relevant fish species. Vaccines (2021) 9:140. doi: 10.3390/vaccines9020140
13. Wang Q, Ji W, Xu Z. Current use and development of fish vaccines in China. Fish Shellfish Immunol (2020) 96:223–34. doi: 10.1016/j.fsi.2019.12.010
14. Adams A. Progress, challenges and opportunities in fish vaccine development. Fish Shellfish Immunol (2019) 90:210–4. doi: 10.1016/j.fsi.2019.04.066
15. Costa JZ, Thompson KD. Understanding the interaction between betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol (2016) 53:35–49. doi: 10.1016/j.fsi.2016.03.033
16. Salonius K, Siderakis C, MacKinnon AM, Griffiths SG. As a live vaccine against. Dev Biol Basel (2005) 121:189–97. Available at: https://www.researchgate.net/publication/7781797.
17. Hwang JY, Kwon MG, Seo JS, Hwang SD, Jeong JM, Lee JH, et al. Current use and management of commercial fish vaccines in Korea. Fish Shellfish Immunol (2020) 102:20–7. doi: 10.1016/j.fsi.2020.04.004
18. Shoemaker CA, Klesius PH, Drennan JD, Evans JJ. Efficacy of a modified live flavobacterium columnare vaccine in fish. Fish Shellfish Immunol (2011) 30:304–8. doi: 10.1016/j.fsi.2010.11.001
19. Sommerset I, Krossøy B, Biering E, Frost P. Vaccines for fish in aquaculture. Expert Rev Vaccines (2005) 4:89–101. doi: 10.1586/14760584.4.1.89
20. Salonius K, Siderakis C, MacKinnon AM, Griffiths SG. Use of arthrobacter davidanieli as a live vaccine against renibacterium salmoninarum and piscirickettsia salmonis in salmonids. Developments Biologicals. (2005) 121:189–98.
21. Yamaguchi T, Quillet E, Boudinot P, Fischer U. What could be the mechanisms of immunological memory in fish? Fish Shellfish Immunol (2019) 85:3–8. doi: 10.1016/j.fsi.2018.01.035
22. Angelidis P, Karagiannis D, Crump EM. Efficacy of a listonella anguillarum (syn. vibrio anguillarum) vaccine for juvenile sea bass dicentrarchus labrax. Dis Aquat Organ (2006) 71:19–24. doi: 10.3354/dao071019
23. Johnson KA, Flynn JK, Amend DF. Duration of immunity in salmonids vaccinated by direct immersion with yersinia ruckeri and vibrio anguillarum bacterins. J Fish Dis (1982) 5:207–13. doi: 10.1111/j.1365-2761.1982.tb00475.x
24. Nayak SK. Current prospects and challenges in fish vaccine development in India with special reference to aeromonas hydrophila vaccine. Fish Shellfish Immunol (2020) 100:283–99. doi: 10.1016/j.fsi.2020.01.064
25. Hoare R, Jung SJ, Ngo TPH, Bartie K, Bailey J, Thompson KD, et al. Efficacy and safety of a non-mineral oil adjuvanted injectable vaccine for the protection of Atlantic salmon (Salmo salar l.) against flavobacterium psychrophilum. Fish Shellfish Immunol (2019) 85:44–51. doi: 10.1016/j.fsi.2017.10.005
26. Kayansamruaj P, Areechon N, Unajak S. Development of fish vaccine in southeast Asia: A challenge for the sustainability of SE Asia aquaculture. Fish Shellfish Immunol (2020) 103:73–87. doi: 10.1016/j.fsi.2020.04.031
27. Jeong KH, Kim HJ, Kim HJ. Current status and future directions of fish vaccines employing virus-like particles. Fish Shellfish Immunol (2020) 100:49–57. doi: 10.1016/j.fsi.2020.02.060
28. Figueroa C, Torrealba D, Morales-Lange B, Mercado L, Dixon B, Conejeros P, et al. Commercial vaccines do not confer protection against two genogroups of piscirickettsia salmonis, LF-89 and EM-90, in Atlantic salmon. Biology-Basal (2022) 11:993. doi: 10.3390/biology11070993
29. Malecki JK, Roy LA, Arias CR, Nhat Truong T, Hanson TR, Lange MD, et al. Bioeconomic analysis of flavobacterium columnare vaccine pond trials with channel catfish. N Am J Aquac (2021) 83:207–17. doi: 10.1002/naaq.10191
30. Assefa A, Abunna F. Maintenance of fish health in aquaculture: review of epidemiological approaches for prevention and control of infectious disease of fish. Vet Med Int (2018) 2018:1-10. doi: 10.1155/2018/5432497
31. Shivam S, El-Matbouli M, Kumar G. Development of fish parasite vaccines in the OMICs era: Progress and opportunities. Vaccines (2021) 9:179. doi: 10.3390/vaccines9020179
32. Adams A, Subasinghe R. Use of fish vaccines in aquaculture (including methods of administration). Food Agric Organ United Nations Rome Italy (2019).
33. Bogwald J, Dalmo RA. Protection of teleost fish against infectious diseases through oral administration of vaccines: Update 2021. Int J Mol Sci (2021) 22:10932. doi: 10.3390/ijms222010932
34. Jose Priya TA, Kappalli S. Modern biotechnological strategies for vaccine development in aquaculture - prospects and challenges. Vaccine (2022) 40:5873–81. doi: 10.1016/j.vaccine.2022.08.075
35. Dadar M, Dhama K, Vakharia VN, Hoseinifar SH, Karthik K, Tiwari R, et al. Advances in aquaculture vaccines against fish pathogens: global status and current trends. Rev Fish Sci Aquac (2017) 25:184–217. doi: 10.1080/23308249.2016.1261277
36. Dong ZR, Mu QJ, Kong WG, Qin DC, Zhou Y, Wang XY, et al. Gut mucosal immune responses and protective efficacy of oral yeast cyprinid herpesvirus 2 (CyHV-2) vaccine in carassius auratus gibelio. Front Immunol (2022) 13:932722. doi: 10.3389/fimmu.2022.932722
37. Tamer C, Albayrak H, Gumusova S. Comparison of immune response between escherichia coli-derived recombinant subunit vaccine and formol-inactivated whole particle vaccine against viral haemorrhagic septicaemia virus (VHSV) in rainbow trout. Aquac Res (2021) 52:2706–14. doi: 10.1111/are.15121
38. Soltani M, Lymbery A, Song SK, Hosseini Shekarabi P. Adjuvant effects of medicinal herbs and probiotics for fish vaccines. Rev Aquac (2019) 11:1325–41. doi: 10.1111/raq.12295
39. Lv P, Song Y, Liu C, Yu L, Shang Y, Tang H, et al. Application of bacillus subtilis as a live vaccine vector: A review. J Vet Med Sci (2020) 82:1693–9. doi: 10.1292/jvms.20-0363
40. Zhu L, Yuan G, Wang X, Zhao T, Hou L, Li C, et al. The construction of a duckweed expression and delivery system for grass carp reovirus VP35. Aquaculture (2022) 553:738059. doi: 10.1016/j.aquaculture.2022.738059
41. Vetter V, Denizer G, Friedland LR, Krishnan J, Shapiro M. Understanding modern-day vaccines: what you need to know. Ann Med (2018) 50:110–20. doi: 10.1080/07853890.2017.1407035
42. Ström-Bestor M, Mustamäki N, Heinikainen S, Hirvelä-Koski V, Verner-Jeffreys D, Wiklund T. Introduction of yersinia ruckeri biotype 2 into Finnish fish farms. Aquaculture (2010) 308:1–5. doi: 10.1016/j.aquaculture.2010.08.009
43. Kim MS, Park JS, Kim KH. Generation of G gene-deleted viral hemorrhagic septicemia virus (VHSV) and evaluation of its vaccine potential in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol (2015) 45:666–71. doi: 10.1016/j.fsi.2015.05.031
44. Vargas D, Vallejos-Vidal E, Reyes-Cerpa S, Oyarzun-Arrau A, Acuna-Castillo C, Imarai M, et al. The analysis of live-attenuated piscirickettsia salmonis vaccine reveals the short-term upregulation of innate and adaptive immune genes in Atlantic salmon (Salmo salar): An In situ open-Sea cages study. Microorganisms (2021) 9:703. doi: 10.3390/microorganisms9040703
45. Zeng R, Pan W, Lin Y, He J, Luo Z, Li Z, et al. Development of a gene-deleted live attenuated candidate vaccine against fish virus (ISKNV) with low pathogenicity and high protection. iScience (2021) 24:102750. doi: 10.1016/j.isci.2021.102750
46. Saito H, Okamura T, Shibata T, Kato G, Sano M. Development of a live attenuated vaccine candidate against herpesviral hematopoietic necrosis of goldfish. Aquaculture (2022) 552:737974. doi: 10.1016/j.aquaculture.2022.737974
47. Aksnes I, Braaen S, Markussen T, Akesson CP, Villoing S, Rimstad E. Genetically modified attenuated salmonid alphavirus: A potential strategy for immunization of Atlantic salmon. J Fish Dis (2021) 44:923–37. doi: 10.1111/jfd.13352
48. Liu G, Zhu J, Chen K, Gao T, Yao H, Liu Y, et al. Development of streptococcus agalactiae vaccines for tilapia. Dis Aquat Organ (2016) 122:163–70. doi: 10.3354/dao03084
49. Liu YL, He TT, Jiang XL, Sun SS, Wang LK, Nie P, et al. Development of a hyper-adhesive and attenuated edwardsiella ictaluri strain as a novel immersion vaccine candidate in yellow catfish (Pelteobagrus fulvidraco). Microb Pathog (2022) 167:105577. doi: 10.1016/j.micpath.2022.105577
50. Klafack S, Schroeder L, Jin Y, Lenk M, Lee PY, Fuchs W, et al. Development of an attenuated vaccine against koi herpesvirus disease (KHVD) suitable for oral administration and immersion. NPJ Vaccines (2022) 7:106. doi: 10.1038/s41541-022-00525-6
51. Adamek M, Matras M, Rebl A, Stachnik M, Falco A, Bauer J, et al. Don’t let it get under your skin! vaccination protects the skin barrier of common carp from disruption caused by cyprinid herpesvirus 3. Front Immunol (2022) 13:787021. doi: 10.3389/fimmu.2022.787021
52. Ma J, Casadei E, Bruce TJ, Sepahi A, Cain KD, Salinas I. Long-term efficacy of nasal vaccination against enteric red mouth (ERM) disease and infectious hematopoietic necrosis (IHN) in juvenile rainbow trout (Oncorhynchus mykiss). Vaccine (2022) 40:229–38. doi: 10.1016/j.vaccine.2021.11.077
53. Curtiss IIIR, Wanda S-Y, Gunn BM, Zhang X, Tinge SA, Ananthnarayan V, et al. Salmonella enterica serovar typhimurium strains with regulated delayed attenuation in vivo. Infect Immun (2009) 77:1071–82. doi: 10.1128/IAI.00693-08
54. Shoemaker CA, Klesius PH, Evans JJ. Immunization of eyed channel catfish, ictalurus punctatus, eggs with monovalent flavobacterium columnare vaccine and bivalent f. columnare and edwardsiella ictaluri vaccine. Vaccine (2007) 25:1126–31. doi: 10.1016/j.vaccine.2006.09.055
55. Yang J, Yang X, Su Y, Peng X, Li H. Activation of the TCA cycle to provide immune protection in zebrafish immunized by high magnesium-prepared vibrio alginolyticus vaccine. Front Immunol (2021) 12:739591. doi: 10.3389/fimmu.2021.739591
56. Staib C, Drexler I, Ohlmann M, Wintersperger S, Erfle V, Sutter G. Transient host range selection for genetic engineering of modified vaccinia virus Ankara. Biotechniques (2000) 28:1137–48. doi: 10.2144/00286st04
57. Lery L, Frangeul L, Tomas A, Passet V, Almeida AS, Bialek-Davenet S, et al. Comparative analysis of klebsiella pneumoniae genomes identifies a phospholipase d family protein as a novel virulence factor. BMC Biol (2014) 12:1–15. doi: 10.1186/1741-7007-12-41
58. Erfanmanesh A, Mohajerfar T, Nikaein D, Mokhtari A, Beikzadeh B. Comparative protection of two antigens (whole-cell and outer membrane vesicle) of yersinia ruckeri in rainbow trout (Oncorhynchus mykiss). Iran J Fish Sci (2022) 21:187–201. doi: 10.22092/ijfs.2022.125884
59. Sasse A, Hamer SN, Amich J, Binder J, Krappmann S. Mutant characterization and in vivo conditional repression identify aromatic amino acid biosynthesis to be essential for aspergillus fumigatus virulence. Virulence (2016) 7:56–62. doi: 10.1080/21505594.2015.1109766
60. Ye J, Ma Y, Liu Q, Zhao DL, Wang QY, Zhang YX. Regulation of vibrio alginolyticus virulence by the LuxS quorum-sensing system. J Fish Dis (2008) 31:161–9. doi: 10.1111/j.1365-2761.2007.00882.x
61. Liu H, Wang Q, Liu Q, Cao X, Shi C, Zhang Y. Roles of hfq in the stress adaptation and virulence in fish pathogen vibrio alginolyticus and its potential application as a target for live attenuated vaccine. Appl Microbiol Biotechnol (2011) 91:353–64. doi: 10.1007/s00253-011-3286-3
62. Liu H, Gu D, Cao X, Liu Q, Wang Q, Zhang Y. Characterization of a new quorum sensing regulator luxT and its roles in the extracellular protease production, motility, and virulence in fish pathogen vibrio alginolyticus. Arch Microbiol (2012) 194:439–52. doi: 10.1007/s00203-011-0774-x
63. Gao X, Wang X, Mao Q, Xu R, Zhou X, Ma Y, et al. VqsA, a novel LysR-type transcriptional regulator, coordinates quorum sensing (QS) and is controlled by QS to regulate virulence in the pathogen vibrio alginolyticus. Appl Environ Microbiol (2018) 84:e00444–18. doi: 10.1128/AEM.00444-18
64. Ye H, Xu Z, Tao Z, Li W, Li Y, Yang A, et al. Efficacy and safety of pseudomonas plecoglossicida mutant ΔtssD-1 as a live attenuated vaccine for the large yellow croaker (Larimichthys crocea). Aquaculture (2021) 531:735976. doi: 10.1016/j.aquaculture.2020.735976
65. Alvarez B, Alvarez J, Menendez A, Guijarro JA. A mutant in one of two exbD loci of a TonB system in flavobacterium psychrophilum shows attenuated virulence and confers protection against cold water disease. Microbiology (2008) 154:1144–51. doi: 10.1099/mic.0.2007/010900-0
66. Bruce TJ, Ma J, Sudheesh PS, Cain KD. Quantification and comparison of gene expression associated with iron regulation and metabolism in a virulent and attenuated strain of flavobacterium psychrophilum. J Fish Dis (2021) 44:949–60. doi: 10.1111/jfd.13354
67. Figueira R, Watson KG, Holden DW, Helaine S. Identification of salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of s. enterica serovar typhimurium: implications for rational vaccine design. MBio (2013) 4:e00065–13. doi: 10.1128/mBio.00065-13
68. Jennings E, Thurston TLM, Holden DW. Salmonella SPI-2 type III secretion system effectors: molecular mechanisms and physiological consequences. Cell Host Microbe (2017) 22:217–31. doi: 10.1016/j.chom.2017.07.009
69. Knuff-Janzen K, Tupin A, Yurist-Doutsch S, Rowland JL, Finlay BB. Multiple salmonella-pathogenicity island 2 effectors are required to facilitate bacterial establishment of its intracellular niche and virulence. PloS One (2020) 15:e0235020. doi: 10.1371/journal.pone.0235020
70. Sayed M, Ozdemir O, Essa M, Olivier A, Karsi A, Lawrence ML, et al. Virulence and live vaccine potential of edwardsiella piscicida phoP and phoQ mutants in catfish against edwardsiellosis. J Fish Dis (2021) 44:1463–74. doi: 10.1111/jfd.13453
71. Sebastiao F de A, Hansen JD, Soto E. Evaluation of francisella orientalis delta pdpA as a live attenuated vaccine against piscine francisellosis in Nile tilapia. J Aquat Anim Health (2022) 34:134–9. doi: 10.1002/aah.10166
72. Yang W, Wang L, Zhang L, Qu J, Wang Q, Zhang Y. An invasive and low virulent edwardsiella tarda esrB mutant promising as live attenuated vaccine in aquaculture. Appl Microbiol Biotechnol (2015) 99:1765–77. doi: 10.1007/s00253-014-6214-5
73. Liu Y, Zhao L, Yang M, Yin K, Zhou X, Leung KY, et al. Transcriptomic dissection of the horizontally acquired response regulator EsrB reveals its global regulatory roles in the physiological adaptation and activation of T3SS and the cognate effector repertoire in edwardsiella piscicida during infection toward turbot. Virulence (2017) 8:1355–77. doi: 10.1080/21505594.2017.1323157
74. Jin J, Li Y, Huang M, Li S, Mao Z. Preliminary studies on the different roles of T6SSs in pathogenicity of pseudomonas plecoglossicida NB2011. J Fish Dis (2021) 44:1669–79. doi: 10.1111/jfd.13479
75. Wang J, Zeng F, Feng H, Chen L, Zhang W, Xiao X, et al. A T3SS regulator mutant HY9901 delta araC of vibrio alginolyticus decreased the expression of HopPmaJ and provided protection to danio rerio as a live-attenuated vaccine. Front Mar Sci (2022) 9:938742. doi: 10.3389/fmars.2022.938742
76. Choe Y, Lee D, Seong M, Bin YJ, JH Y, JY Y, et al. Characterization of edwardsiella piscicida CK108 flagellin genes and evaluation of their potential as vaccine targets in the zebrafish model. J Fish Dis (2022) 45:249–59. doi: 10.1111/jfd.13550
77. Li J, Ma S, Li Z, Yu W, Zhou P, Ye X, et al. Construction and characterization of an aeromonas hydrophila multi-gene deletion strain and evaluation of its potential as a live-attenuated vaccine in grass carp. Vaccines (2021) 9:451. doi: 10.3390/vaccines9050451
78. Tan H, Da F, Lin G, Wan X, Cai S, Cai J, et al. Construction of a phosphodiesterase mutant and evaluation of its potential as an effective live attenuated vaccine in pearl gentian grouper (♀ epinephelus fuscoguttatus × ♂ epinephelus lanceolatu). Fish Shellfish Immunol (2022) 124:543–51. doi: 10.1016/j.fsi.2022.04.016
79. Hao J, Wang S, Wei Z, Zhang Q, Wu Z, Lin Y, et al. Construction of streptococcus agalactiae sialic acid mutant and evaluation of its potential as a live attenuated vaccine in Nile tilapia (Oreochromis niloticus). J Appl Microbiol (2022) 133:2403–16. doi: 10.1111/jam.15706
80. Li Y, Chi Y, Li S, Jia T, Mao Z. Characterization of 4 deletion mutants of pseudomonas plecoglossicida and their potential for live attenuated vaccines in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol (2022) 127:264–70. doi: 10.1016/j.fsi.2022.06.034
81. Lopez-Porras A, Griffin MJ, Ware C, Richardson BM, Greenway TE, Rosser TG, et al. Cross-protective efficacy of a live-attenuated edwardsiella ictaluri vaccine against heterologous edwardsiella piscicida isolates in channel and channel x blue catfish hybrids. J Fish Dis (2022) 45:1001–10. doi: 10.1111/jfd.13623
82. Malik MS, Teige LH, Braaen S, Olsen AB, Nordberg M, Amundsen MM, et al. Piscine orthoreovirus (PRV)-3, but not PRV-2, cross-protects against PRV-1 and heart and skeletal muscle inflammation in Atlantic salmon. Vaccines (2021) 9:230. doi: 10.3390/vaccines9030230
83. Pang H, Chang Y, Zheng H, Tan H, Zhou S, Zeng F, et al. A live attenuated strain of HY9901AvscB provides protection against vibrio alginolyticus in pearl gentian grouper (♀ epinephelus fuscoguttatus × ♂ epinephelus lanceolatu). Aquaculture (2022) 546:737353. doi: 10.1016/j.aquaculture.2021.737353
84. Thune RL, Fernandez DH, Battista JR. An aroa mutant of edwardsiella ictaluri is safe and efficacious as a live, attenuated vaccine. J Aquat Anim Health (1999) 11:358–72. doi: 10.1577/1548-8667(1999)011<0358:AAMOEI>2.0.CO;2
85. Abdelhamed H, Lu J, Shaheen A, Abbass A, Lawrence ML, Karsi A. Construction and evaluation of an edwardsiella ictaluri fhuC mutant. Vet Microbiol (2013) 162:858–65. doi: 10.1016/j.vetmic.2012.11.006
86. Pridgeon JW, Klesius PH. Development of a novobiocin-resistant edwardsiella ictaluri as a novel vaccine in channel catfish (Ictalurus punctatus). Vaccine (2011) 29:5631–7. doi: 10.1016/j.vaccine.2011.06.016
87. Ronen A, Perelberg A, Abramowitz J, Hutoran M, Tinman S, Bejerano I, et al. Efficient vaccine against the virus causing a lethal disease in cultured cyprinus carpio. Vaccine (2003) 21:4677–84. doi: 10.1016/S0264-410X(03)00523-1
88. Perelberg A, Ronen A, Hutoran M, Smith Y, Kotler M. Protection of cultured cyprinus carpio against a lethal viral disease by an attenuated virus vaccine. Vaccine (2005) 23:3396–403. doi: 10.1016/j.vaccine.2005.01.096
89. Eide KE, Miller-Morgan T, Heidel JR, Kent ML, Bildfell RJ, LaPatra S, et al. Investigation of koi herpesvirus latency in koi. J Virol (2011) 85:4954–62. doi: 10.1128/jvi.01384-10
90. Vivas J, Riaño J, Carracedo B, Razquin BE, López-Fierro P, Naharro G, et al. The auxotrophic aroA mutant of aeromonas hydrophila as a live attenuated vaccine against a. salmonicida infections in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol (2004) 16:193–206. doi: 10.1016/S1050-4648(03)00078-0
91. Vivas J, Carracedo B, Riaño J, Razquin BE, López-Fierro P, Acosta F, et al. Behavior of an aeromonas hydrophila aroA live vaccine in water microcosms. Appl Environ Microbiol (2004) 70:2702–8. doi: 10.1128/AEM.70.5.2702-2708.2004
92. Yamasaki M, Araki K, Maruyoshi K, Matsumoto M, Nakayasu C, Moritomo T, et al. Comparative analysis of adaptive immune response after vaccine trials using live attenuated and formalin-killed cells of edwardsiella tarda in ginbuna crucian carp (Carassius auratus langsdorfii). Fish Shellfish Immunol (2015) 45:437–42. doi: 10.1016/j.fsi.2015.04.038
93. Kim MS, Kim KH. Genetically engineered viral hemorrhagic septicemia virus (VHSV) vaccines. Fish Shellfish Immunol (2019) 95:11–5. doi: 10.1016/j.fsi.2019.10.031
94. Kim H, Baek KW, Ko EJ, Luan NT, Lim Y, Roh HJ, et al. Genome based quantification of VHSV in multiple organs of infected olive flounder (Paralichthys olivaceus) using real-time PCR. Genes Genomics (2020) 42:773–7. doi: 10.1007/s13258-020-00951-7
95. Liu CH, Wu K, Chu TW, Wu TM. Dietary supplementation of probiotic, bacillus subtilis E20, enhances the growth performance and disease resistance against vibrio alginolyticus in parrot fish (Oplegnathus fasciatus). Aquaculture Int (2018) 26:63–74. doi: 10.1007/s10499-017-0189-z
96. Zhou S, Tu X, Pang H, Hoare R, Monaghan SJ, Luo J, et al. A T3SS regulator mutant of vibrio alginolyticus affects antibiotic susceptibilities and provides significant protection to danio rerio as a live attenuated vaccine. Front Cell Infect Microbiol (2020) 10:183. doi: 10.3389/fcimb.2020.00183
97. Chen Y, Wu F, Pang H, Tang J, Cai S, Jian J. Superoxide dismutase b (sodB), an important virulence factor of vibrio alginolyticus, contributes to antioxidative stress and its potential application for live attenuated vaccine. Fish Shellfish Immunol (2019) 89:354–60. doi: 10.1016/j.fsi.2019.03.061
98. Chen Y, Cai S, Jian J. Protection against vibrio alginolyticus in pearl gentian grouper (♀ epinephelus fuscoguttatus×♂ epinephelus lanceolatu) immunized with an acfA-deletion live attenuated vaccine. Fish Shellfish Immunol (2019) 86:875–81. doi: 10.1016/j.fsi.2018.12.030
99. Chen Y, Wu F, Wang Z, Tang J, Cai S, Jian J. Construction and evaluation of vibrio alginolyticus ΔclpP mutant, as a safe live attenuated vibriosis vaccine. Fish Shellfish Immunol (2020) 98:917–22. doi: 10.1016/j.fsi.2019.11.054
100. Liu L, Lu DQ, Xu J, Luo HL, Li AX. Development of attenuated erythromycin-resistant streptococcus agalactiae vaccine for tilapia (Oreochromis niloticus) culture. J Fish Dis (2019) 42:693–701. doi: 10.1111/jfd.12977
101. Mo XB, Wang J, Guo S, Li AX. Potential of naturally attenuated streptococcus agalactiae as a live vaccine in Nile tilapia (Oreochromis niloticus). Aquaculture (2020) 518:734774. doi: 10.1016/j.aquaculture.2019.734774
102. Zhang D, Gao Y, Li Q, Ke X, Liu Z, Lu M, et al. An effective live attenuated vaccine against streptococcus agalactiae infection in farmed Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol (2020) 98:853–9. doi: 10.1016/j.fsi.2019.11.044
103. Qi W, Xu W, Zhao L, Xu X, Luo Z, Huang L, et al. Protection against pseudomonas plecoglossicida in epinephelus coioides immunized with a cspA1-knock-down live attenuated vaccine. Fish Shellfish Immunol (2019) 89:498–504. doi: 10.1016/j.fsi.2019.04.029
104. Huang L, Zhang Y, He R, Zuo Z, Luo Z, Xu W, et al. Phenotypic characterization, virulence, and immunogenicity of pseudomonas plecoglossicida rpoE knock-down strain. Fish Shellfish Immunol (2019) 87:772–7. doi: 10.1016/j.fsi.2019.02.028
105. Zhang H, Chen M, Xu Y, Xu GY, Chen JR, Wang YM, et al. An effective live attenuated vaccine against aeromonas veronii infection in the loach (Misgurnus anguillicaudatus). Fish Shellfish Immunol (2020) 104:269–78. doi: 10.1016/j.fsi.2020.05.027
106. Zhou L, Wang X, Liu Q, Wang Q, Zhao Y, Zhang Y. A novel multivalent vaccine based on secretary antigen-delivery induces protective immunity against vibrio anguillarum and aeromonas hydrophila. J Biotechnol (2010) 146:25–30. doi: 10.1016/j.jbiotec.2009.12.010
107. Li XP, Zhang J. A live attenuated edwardsiella tarda vaccine induces immunological expression pattern in Japanese flounder (Paralichthys olivaceus) in the early phase of immunization. Comp Biochem Physiol C-Toxicology Pharmacol (2021) 239:108872. doi: 10.1016/j.cbpc.2020.108872
108. Kordon AO, Abdelhamed H, Karsi A, Pinchuk LM. Adaptive immune responses in channel catfish exposed to edwardsiella ictaluri live attenuated vaccine and wild type strains through the specific gene expression profiles. Dev Comp Immunol (2021) 116:103950. doi: 10.1016/j.dci.2020.103950
109. Zhang D, Liu Z, Ren Y, Wang Y, Pan H, Liang D, et al. Epidemiological characteristics of streptococcus agalactiae in tilapia in China from 2006 to 2020. Aquaculture (2022) 549:737724. doi: 10.1016/j.aquaculture.2021.737724
110. Heckman IT, Shahin K, Henderson EE, Griffin MJ, Soto E. Development and efficacy of streptococcus iniae live-attenuated vaccines in Nile tilapia, oreochromis niloticus. Fish Shellfish Immunol (2022) 121:152–62. doi: 10.1016/j.fsi.2021.12.043
111. Li B, Chen J, Huang P, Weng T, Wen Y, Yang H, et al. Induction of attenuated nocardia seriolae and their use as live vaccine trials against fish nocardiosis. Fish Shellfish Immunol (2022) 131:10–20. doi: 10.1016/j.fsi.2022.09.053
112. Azad IS, Al-Marzouk A, James CM, Almatar S, Al-Gharabally H, Qasem JA. Outbreak of natural streptococcosis in hatchery produced silver pomfret (Pampus argenteus euphrasen) larvae in Kuwait. Aquaculture (2012) 330:15–20. doi: 10.1016/j.aquaculture.2011.12.010
113. Delamare-Deboutteville J, Bowater R, Condon K, Reynolds A, Fisk A, Aviles F, et al. Infection and pathology in Queensland grouper, epinephelus lanceolatus,(Bloch), caused by exposure to streptococcus agalactiae via different routes. J Fish Dis (2015) 38:1021–35. doi: 10.1111/jfd.12273
114. Liu L, Li YW, He RZ, Xiao XX, Zhang X, Su YL, et al. Outbreak of streptococcus agalactiae infection in barcoogrunter, scortumbarcoo (M c c ulloch W aite), in an intensive fish farm in China. J Fish Dis (2014) 37:1067–72. doi: 10.1111/jfd.12187
115. Zhang D, Ke X, Liu L, Lu M, Shi C, Liu Z. Streptococcus agalactiae from tilapia (Oreochromis sp.) transmitted to a new host, bighead carp (Aristichthys nobilis), in China. Aquac Int (2018) 26:885–97. doi: 10.1007/s10499-018-0254-2
116. Liu X, Jiao C, Ma Y, Wang Q, Zhang Y. A live attenuated vibrio anguillarum vaccine induces efficient immunoprotection in tiger puffer (Takifugu rubripes). Vaccine (2018) 36:1460–6. doi: 10.1016/j.vaccine.2018.01.067
117. peng Zhang H, yao Chen M, xuan Xu Y, yi Xu G, rui Chen J, ming Wang Y, et al. An effective live attenuated vaccine against aeromonas veronii infection in the loach (Misgurnus anguillicaudatus). Fish Shellfish Immunol (2020) 104:269–78. doi: 10.1016/j.fsi.2020.05.027
118. Zhang Y, Zhou L, Zhang Y. Investigation of UV-TiO2 photocatalysis and its mechanism in bacillus subtilis spore inactivation. J Environ Sci (China) (2014) 26:1943–8. doi: 10.1016/j.jes.2014.07.007
119. Hussain I, Price GW, Farid AH. Inactivation of aleutian mink disease virus through high temperature exposure in vitro and under field-based composting conditions. Vet Microbiol (2014) 173:50–8. doi: 10.1016/j.vetmic.2014.07.014
120. Duan K, Tang X, Zhao J, Ren G, Shao Y, Lu T, et al. An inactivated vaccine against infectious pancreatic necrosis virus in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol (2022) 127:48–55. doi: 10.1016/j.fsi.2022.06.008
121. Afsharipour E, Zorriehzahra MJ, Takami GA, Kakoolaki S, Motallebi AA, Sharifpour I, et al. An investigation on protective effects of the new killed vaccine against nervous necrosis virus (NNV) using histopathology and immunohistochemistry approach on the brain and eye tissues of acipenser stellatus pallas 1771. Fish Shellfish Immunol (2021) 116:91–7. doi: 10.1016/j.fsi.2021.06.018
122. Li J, Suo Y, Liao X, Ahn J, Liu D, Chen S, et al. Analysis of staphylococcus aureus cell viability, sublethal injury and death induced by synergistic combination of ultrasound and mild heat. Ultrason Sonochem (2017) 39:101–10. doi: 10.1016/j.ultsonch.2017.04.019
123. Boudarkov VA, Sereda AD, Carpov ON, Ponomarev VN. Using gamma rays to inactivate African swine fever virus. Russ Agric Sci (2016) 42:375–7. doi: 10.3103/s1068367416050025
124. Jeong YU, Subramanian D, Jang YH, Kim DH, Park SH, Park K, et al. Protective efficiency of an inactivated vaccine against streptococcus iniae in olive flounder, paralichthys olivaceus. Arch Polish Fish (2016) 24:23–32. doi: 10.1515/aopf-2016-0003
125. Zhang J, Hu Y, Sun Q, Li X, Sun L. An inactivated bivalent vaccine effectively protects turbot (Scophthalmus maximus) against vibrio anguillarum and vibrio harveyi infection. Aquaculture (2021) 544:737158. doi: 10.1016/j.aquaculture.2021.737158
126. Morohoshi T, Yokoyama Y, Ouchi M, Kato N, Ikeda T. Motility and the expression of the flagellin protein FliC are negatively regulated by quorum sensing in edwardsiella tarda. J Biosci Bioeng (2009) 108:314–8. doi: 10.1016/j.jbiosc.2009.04.006
127. Sanders B, Koldijk M, Schuitemaker H. Inactivated viral vaccines[M]//Vaccine analysis: strategies, principles, and control. Berlin, Heidelberg: Springer (2015) p. 45–80.
128. Pridgeon JW, Klesius PH. Major bacterial diseases in aquaculture and their vaccine development. CAB Rev Perspect Agric Vet Sci Nutr Nat Resour (2012) 7:1–16. doi: 10.1079/PAVSNNR20127048
129. Ben Hamed S, Teresa Tapia-Paniagua S, Angel Morinigo M, Tavares Ranzani-Paiva MJ. Advances in vaccines developed for bacterial fish diseases, performance and limits. Aquac Res (2021) 52:2377–90. doi: 10.1111/are.15114
130. Hossain A, Habibullah-Al-Mamun M, Nagano I, Masunaga S, Kitazawa D, Matsuda H. Antibiotics, antibiotic-resistant bacteria, and resistance genes in aquaculture: risks, current concern, and future thinking. Environ Sci pollut Res (2022) 29:1–22. doi: 10.1007/s11356-021-17825-4
131. Schrøder MB, Espelid S, Jørgensen T. Two serotypes of vibrio salmonicida isolated from diseased cod (Gadus morhua l.); virulence, immunological studies and vaccination experiments. Fish Shellfish Immunol (1992) 2:211–21. doi: 10.1016/S1050-4648(05)80060-9
132. Liu X, Zhang H, Gao Y, Zhang Y, Wu H, Zhang Y. Efficacy of chitosan oligosaccharide as aquatic adjuvant administrated with a formalin-inactivated vibrio anguillarum vaccine. Fish Shellfish Immunol (2015) 47:855–60. doi: 10.1016/j.fsi.2015.10.012
133. Sheng X, Chai B, Wang Z, Tang X, Xing J, Zhan W. Polymeric immunoglobulin receptor and mucosal IgM responses elicited by immersion and injection vaccination with inactivated vibrio anguillarum in flounder (Paralichthys olivaceus). Aquaculture (2019) 505:1–11. doi: 10.1016/j.aquaculture.2019.02.045
134. Nguyen HT, Nguyen TTT, Tsai MA, Ya-Zhen E, Wang PC, Chen SC, et al. A formalin-inactivated vaccine provides good protection against vibrio harveyi infection in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol (2017) 65:118–26. doi: 10.1016/j.fsi.2017.04.008
135. Wei G, Cai S, Wu Y, Ma S, Huang Y. Immune effect of vibrio harveyi formalin-killed cells vaccine combined with chitosan oligosaccharide and astragalus polysaccharides in ♀ epinephelus fuscoguttatus × ♂ epinephelus lanceolatus. Fish Shellfish Immunol (2020) 98:186–92. doi: 10.1016/j.fsi.2020.01.015
136. Cao J, Huang AL, Zhu XC, Li L, Li JN. Construction of vibrio mimicus ghosts as a novel inactivated vaccine candidate and its protective efficacy against ascites disease in grass carps (Ctenopharyngodon idella). Aquaculture (2018) 485:147–53. doi: 10.1016/j.aquaculture.2017.11.002
137. Cao J, Zhang J, Ma L, Li L, Zhang W, Li J. Identification of fish source vibrio alginolyticus and evaluation of its bacterial ghosts vaccine immune effects. Microbiologyopen (2018) 7:e00576. doi: 10.1002/mbo3.576
138. Zeng J, Ouyang AT, Wang H, Liu W, Xue M, Zhou Y, et al. A bivalent vaccine comprised of inactivated aeromonas veronii and edwardsiella ictaluri stimulates protective immune responses in yellow-head catfish, pelteobagrus fulvidraco. Aquaculture Res (2021) 52:5673–81. doi: 10.1111/are.15441
139. Song H, Zhang S, Yang B, Liu Y, Kang Y, Li Y, et al. Effects of four different adjuvants separately combined with aeromonas veronii inactivated vaccine on haematoimmunological state, enzymatic activity, inflammatory response and disease resistance in crucian carp. Fish Shellfish Immunol (2022) 120:658–73. doi: 10.1016/j.fsi.2021.09.003
140. Song MF, Kang YH, Zhang DX, Chen L, Bi JF, Zhang HP, et al. Immunogenicity of extracellular products from an inactivated vaccine against aeromonas veronii TH0426 in koi, cyprinus carpio. Fish Shellfish Immunol (2018) 81:176–81. doi: 10.1016/j.fsi.2018.07.004
141. Småge SB, Frisch K, Vold V, Duesund H, Brevik ØJ, Olsen RH, et al. Induction of tenacibaculosis in Atlantic salmon smolts using tenacibaculum finnmarkense and the evaluation of a whole cell inactivated vaccine. Aquaculture (2018) 495:858–64. doi: 10.1016/j.aquaculture.2018.06.063
142. Shahin K, Shinn AP, Metselaar M, Ramirez-Paredes JG, Monaghan SJ, Thompson KD, et al. Efficacy of an inactivated whole-cell injection vaccine for nile tilapia, oreochromis niloticus (L), against multiple isolates of francisella noatunensis subsp. orientalis from diverse geographical regions. Fish Shellfish Immunol (2019) 89:217–27. doi: 10.1016/j.fsi.2019.03.071
143. Takano T, Matsuyama T, Sakai T, Nakayasu C. Protective efficacy of a formalin-killed vaccine against atypical edwardsiella tarda infection in red sea bream pagrus major. Fish Pathol (2011) 46:120–2. doi: 10.3147/jsfp.46.120
144. Zhu W, Zhang Y, Zhang J, Yuan G, Liu X, Ai T, et al. Astragalus polysaccharides, chitosan and poly (I: C) obviously enhance inactivated edwardsiella ictaluri vaccine potency in yellow catfish pelteobagrus fulvidraco. Fish Shellfish Immunol (2019) 87:379–85. doi: 10.1016/j.fsi.2019.01.033
145. Tang X, Gong J, Zeng C, Sheng X, Xing J, Zhan W. Dynamic distribution of formalin-inactivated edwardsiella tarda in flounder (Paralichthys olivaceus) post intraperitoneal vaccination. Fish Shellfish Immunol (2019) 89:393–402. doi: 10.1016/j.fsi.2019.04.022
146. Huang HY, Chen YC, Wang PC, Tsai MA, Yeh SC, Liang HJ, et al. Efficacy of a formalin-inactivated vaccine against streptococcus iniae infection in the farmed grouper epinephelus coioides by intraperitoneal immunization. Vaccine (2014) 32:7014–20. doi: 10.1016/j.vaccine.2014.08.039
147. Ramos-Espinoza FC, Cueva-Quiroz VA, Yunis-Aguinaga J, Alvarez-Rubio NC, de Mello NP, de Moraes JRE. Efficacy of two adjuvants administrated with a novel hydrogen peroxide-inactivated vaccine against streptococcus agalactiae in Nile tilapia fingerlings. Fish Shellfish Immunol (2020) 105:350–8. doi: 10.1016/j.fsi.2020.07.051
148. Rao S, Byadgi O, Pulpipat T, Wang PC, Chen SC. Efficacy of a formalin-inactivated lactococcus garvieae vaccine in farmed grey mullet (Mugil cephalus). J Fish Dis (2020) 43:1579–89. doi: 10.1111/jfd.13260
149. Zahrei-Abdevand L, Soltani M, Sh S. Adjuvant effect of licorice (Glycyrrhiza glabra) extract on the efficacy of lactococcosis vaccine in rainbow trout (Oncorhynchus mykiss). Iran J Fish Sci (2021) 20:646–62. doi: 10.22092/ijfs.2021.124006
150. Liu S, Wang D, Cao Y, Lu T, Liu H, Li S. Effects of propolis on the immune enhancement of the formalin-inactivated aeromonas salmonicida vaccine. Aquac Res (2020) 51:4759–70. doi: 10.1111/are.14822
151. Zhang Z, Liu G, Ma R, Qi X, Wang G, Zhu B, et al. The immunoprotective effect of whole-cell lysed inactivated vaccine with SWCNT as a carrier against aeromonas hydrophila infection in grass carp. Fish Shellfish Immunol (2020) 97:336–43. doi: 10.1016/j.fsi.2019.12.069
152. Nguyen HT, Nguyen TTT, Tsai MA, Ya-Zhen E, Wang PC, Chen SC. A formalin-inactivated vaccine provides good protection against vibrio harveyi infection in orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol (2017) 65:118–26. doi: 10.1016/j.fsi.2017.04.008
153. Xu W, Jiao C, Bao P, Liu Q, Wang P, Zhang R, et al. Efficacy of montanide ISA 763 a VG as aquatic adjuvant administrated with an inactivated vibrio harveyi vaccine in turbot (Scophthalmus maximus l.). Fish Shellfish Immunol (2019) 84:56–61. doi: 10.1016/j.fsi.2018.09.024
154. Wei G, Tan H, Ma S, Sun G, Zhang Y, Wu Y, et al. Protective effects of β-glucan as adjuvant combined inactivated vibrio harveyi vaccine in pearl gentian grouper. Fish Shellfish Immunol (2020) 106:1025–30. doi: 10.1016/j.fsi.2020.09.027
155. Anderson E, Clouthier S, Shewmaker W, Weighall A, LaPatra S. Inactivated infectious haematopoietic necrosis virus (IHNV) vaccines. J Fish Dis (2008) 31:729–45. doi: 10.1111/j.1365-2761.2008.00960.x
156. Karlsen M, Tingbø T, Solbakk IT, Evensen Ø, Furevik A, Aas-Eng A. Efficacy and safety of an inactivated vaccine against salmonid alphavirus (family togaviridae). Vaccine (2012) 30:5688–94. doi: 10.1016/j.vaccine.2012.05.069
157. Xu C, Mutoloki S, Evensen Ø. Superior protection conferred by inactivated whole virus vaccine over subunit and DNA vaccines against salmonid alphavirus infection in Atlantic salmon (Salmo salar l.). Vaccine (2012) 30:3918–28. doi: 10.1016/j.vaccine.2012.03.081
158. Kim HJ, Park JS, Choi MC, Kwon SR. Comparison of the efficacy of Poly(I: C) immunization with live vaccine and formalin-killed vaccine against viral hemorrhagic septicemia virus (VHSV) in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol (2016) 48:206–11. doi: 10.1016/j.fsi.2015.11.035
159. Kai YH, Chi SC. Efficacies of inactivated vaccines against betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine (2008) 26:1450–7. doi: 10.1016/j.vaccine.2007.12.043
160. Kai YH, Su HM, Tai KT, Chi SC. Vaccination of grouper broodfish (Epinephelus tukula) reduces the risk of vertical transmission by nervous necrosis virus. Vaccine (2010) 28:996–1001. doi: 10.1016/j.vaccine.2009.10.132
161. Valero Y, Olveira JG, López-Vázquez C, Dopazo CP, Bandín I. Bei inactivated vaccine induces innate and adaptive responses and elicits partial protection upon reassortant betanodavirus infection in senegalese sole. Vaccines (2021) 9:458. doi: 10.3390/vaccines9050458
162. Kai YH, Wu YC, Chi SC. Immune gene expressions in grouper larvae (Epinephelus coioides) induced by bath and oral vaccinations with inactivated betanodavirus. Fish Shellfish Immunol (2014) 40:563–9. doi: 10.1016/j.fsi.2014.08.005
163. Liu W, Xu J, Ma J, LaPatra SE, Meng Y, Fan Y, et al. Immunological responses and protection in Chinese giant salamander andrias davidianus immunized with inactivated iridovirus. Vet Microbiol (2014) 174:382–90. doi: 10.1016/j.vetmic.2014.10.028
164. Zhang L, Ma J, Fan Y, Zhou Y, Xu J, Liu W, et al. Immune response and protection in gibel carp, carassius gibelio, after vaccination with β-propiolactone inactivated cyprinid herpesvirus 2. Fish Shellfish Immunol (2016) 49:344–50. doi: 10.1016/j.fsi.2016.01.003
165. Zeng W, Wang Q, Wang Y, Zhao C, Li Y, Shi C, et al. Immunogenicity of a cell culture-derived inactivated vaccine against a common virulent isolate of grass carp reovirus. Fish Shellfish Immunol (2016) 54:473–80. doi: 10.1016/j.fsi.2016.04.133
166. Huang SM, Cheng JH, Tu C, Chen TI, Lin CT, Chang SK. A bivalent inactivated vaccine of viral nervous necrosis virus and grouper iridovirus applied to grouper broodfish (Epinephelus coioides) reduces the risk of vertical transmission. Taiwan Vet J (2017) 43:171–6. doi: 10.1142/s1682648517500032
167. Nuñez-Ortiz N, Pascoli F, Picchietti S, Buonocore F, Bernini C, Toson M, et al. A formalin-inactivated immunogen against viral encephalopathy and retinopathy (VER) disease in European sea bass (Dicentrarchus labrax): Immunological and protection effects. Vet Res (2016) 47:1–11. doi: 10.1186/s13567-016-0376-3
168. Valero Y, Mokrani D, Chaves-Pozo E, Arizcun M, Oumouna M, Meseguer J, et al. Vaccination with UV-inactivated nodavirus partly protects European sea bass against infection, while inducing few changes in immunity. Dev Comp Immunol (2018) 86:171–9. doi: 10.1016/j.dci.2018.05.013
169. Buonocore F, Nuñez-Ortiz N, Picchietti S, Randelli E, Stocchi V, Guerra L, et al. Vaccination and immune responses of European sea bass (Dicentrarchus labrax l.) against betanodavirus. Fish Shellfish Immunol (2019) 85:78–84. doi: 10.1016/j.fsi.2017.11.039
170. Amar EC, Faisan JP, Gapasin RSJ. Field efficacy evaluation of a formalin-inactivated white spot syndrome virus (WSSV) vaccine for the preventive management of WSSV infection in shrimp grow-out ponds. Aquaculture (2021) 531:735907. doi: 10.1016/j.aquaculture.2020.735907
171. Mai TT, Kayansamruaj P, Taengphu S, Senapin S, Costa JZ, del-Pozo J, et al. Efficacy of heat-killed and formalin-killed vaccines against tilapia tilapinevirus in juvenile Nile tilapia (Oreochromis niloticus). J Fish Dis (2021) 44:2097–109. doi: 10.1111/jfd.13523
172. Aich N, Paul A, Choudhury TG, Saha H. Tilapia lake virus (TiLV) disease: Current status of understanding. Aquac Fish (2022) 7:7–17. doi: 10.1016/j.aaf.2021.04.007
173. Mugimba KK, Byarugaba DK, Mutoloki S, Evensen O, Munang’andu HM. Challenges and solutions to viral diseases of finfish in marine aquaculture. Pathogens (2021) 10:673. doi: 10.3390/pathogens10060673
174. Vinay TN, Kim YJ, Jung MH, Kim WS, Kim DH, Jung SJ. Inactivated vaccine against viral hemorrhagic septicemia (VHS) emulsified with squalene and aluminum hydroxide adjuvant provides long term protection in olive flounder (Paralichthys olivaceus). Vaccine (2013) 31:4603–10. doi: 10.1016/j.vaccine.2013.07.036
175. Zeng W, Wang Y, Hu H, Wang Q, Bergmann SM, Wang Y, et al. Cell culture-derived tilapia lake virus-inactivated vaccine containing montanide adjuvant provides high protection against viral challenge for tilapia. Vaccines (2021) 9:4603–10. doi: 10.3390/vaccines9020086
176. Khushiramani RM, Maiti B, Shekar M, Girisha SK, Akash N, Deepanjali A, et al. Recombinant aeromonas hydrophila outer membrane protein 48 (Omp48) induces a protective immune response against aeromonas hydrophila and edwardsiella tarda. Res Microbiol (2012) 163:286–91. doi: 10.1016/j.resmic.2012.03.001
177. Xu H, Xing J, Tang X, Sheng X, Zhan W. Intramuscular administration of a DNA vaccine encoding OmpK antigen induces humoral and cellular immune responses in flounder (Paralichthys olivaceus) and improves protection against vibrio anguillarum. Fish Shellfish Immunol (2019) 86:618–26. doi: 10.1016/j.fsi.2018.11.073
178. Xing J, Zhang Z, Luo K, Tang X, Sheng X, Zhan W. T And b lymphocytes immune responses in flounder (Paralichthys olivaceus) induced by two forms of outer membrane protein K from vibrio anguillarum: Subunit vaccine and DNA vaccine. Mol Immunol (2020) 118:40–51. doi: 10.1016/j.molimm.2019.12.002
179. Fu Y, Zhang YA, Shen J, Tu J. Immunogenicity study of OmpU subunit vaccine against vibrio mimicus in yellow catfish, pelteobagrus fulvidraco. Fish Shellfish Immunol (2021) 108:80–5. doi: 10.1016/j.fsi.2020.11.030
180. Nguyen HT, Nguyen TTT, Chen YC, Vu-Khac H, Wang PC, Chen SC. Enhanced immune responses and effectiveness of refined outer membrane protein vaccines against vibrio harveyi in orange-spotted grouper (Epinephelus coioides). J Fish Dis (2018) 41:1349–58. doi: 10.1111/jfd.12828
181. Tang X, Wang H, Liu F, Sheng X, Xing J, Zhan W, et al. Outer membrane protein a: An immunogenic protein induces highly protective efficacy against vibrio ichthyoenteri. Microb Pathog (2017) 113:152–9. doi: 10.1016/j.micpath.2017.10.043
182. Tang X, Wang H, Liu F, Sheng X, Xing J, Zhan W. Recombinant outer membrane protein T (OmpT) of vibrio ichthyoenteri, a potential vaccine candidate for flounder (Paralichthys olivaceus). Microb Pathog (2019) 126:185–92. doi: 10.1016/j.micpath.2018.11.001
183. Liu F, Tang X, Sheng X, Xing J, Zhan W. Comparative study of the vaccine potential of six outer membrane proteins of edwardsiella tarda and the immune responses of flounder (Paralichthys olivaceus) after vaccination. Vet Immunol Immunopathol (2017) 185:38–47. doi: 10.1016/j.vetimm.2017.01.008
184. Sharma M, Dixit A. Immune response characterization and vaccine potential of a recombinant chimera comprising b-cell epitope of aeromonas hydrophila outer membrane protein c and LTB. Vaccine (2016) 34:6259–66. doi: 10.1016/j.vaccine.2016.10.064
185. Gonzalez-Stegmaier R, Pena A, Villarroel-Espindola F, Aguila P, Oliver C, MacLeod-Carey D, et al. Full recombinant flagellin b from vibrio anguillarum (rFLA) and its recombinant D1 domain (rND1) promote a pro-inflammatory state and improve vaccination against p. salmonis in Atlantic salmon (S. salar). Dev Comp Immunol (2021) 117. doi: 10.1016/j.dci.2020.103988
186. Rout SS, de Grahl I, Yu X, Reumann S. Production of a viral surface protein in nannochloropsis oceanica for fish vaccination against infectious pancreatic necrosis virus. Appl Microbiol Biotechnol (2022) 106:6535–49. doi: 10.1007/s00253-022-12106-7
187. Makesh M, Rajendran KV. Viral vaccines for farmed finfish. Fish Immune Syst Vaccines (2022) (Singapore: Springer) 95. doi: 10.1007/978-981-19-1268-9_5
188. Ma H, Cheng C, Deng Y, Liu G, Fan S, Feng J, et al. Effective protection against ranavirus-type grouper iridovirus and vibrio harveyi by priming a bivalent GrouperVAC-Irido-R-Vh followed by boosting a monovalent V. harveyi Vaccine Aquaculture (2022) 560:738455. doi: 10.1016/j.aquaculture.2022.738455
189. Kole S, Shin SM, Kwak IS, Cho SH, Jung SJ. Efficacy of chitosan-PLGA encapsulated trivalent oral vaccine against viral haemorrhagic septicemia virus, streptococcus parauberis, and miamiensis avidus in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol (2022) 127:843–54. doi: 10.1016/j.fsi.2022.07.029
190. Aly SM, Eissa AE, ElBanna NI, Albutti A. Efficiency of monovalent and polyvalent vibrio alginolyticus and vibrio parahaemolyticus vaccines on the immune response and protection in gilthead sea bream, sparus aurata (L.) against vibriosis. Fish Shellfish Immunol (2021) 111:145–51. doi: 10.1016/j.fsi.2020.10.011
191. Mohammadi Y, Mesbah M, Dezfoulnejad MC, Mehrgan MS, Islami HR. Growth performance, blood biochemical parameters, immune response, and antioxidant defense of Asian seabass (Lates calcarifer) fingerlings exposed to monovalent and bivalent vaccines against streptococcus iniae and vibrio harveyi. Aquac Int (2021) 29:2751–67. doi: 10.1007/s10499-021-00776-5
192. Lan NGT, Salin KR, Longyant S, Senapin S, Dong HT. Systemic and mucosal antibody response of freshwater cultured Asian seabass (Lates calcarifer) to monovalent and bivalent vaccines against streptococcus agalactiae and streptococcus iniae. Fish Shellfish Immunol (2021) 108:7–13. doi: 10.1016/j.fsi.2020.11.014
193. Secombes CJ. Cytokines and immunity. In: Principles of fish immunology. (Springer, Cham) (2022) 301–53. doi: /10.1007/978-3-030-85420-1_10
194. Xu H, Xing J, Tang X, Sheng X, Zhan W. The effects of CCL3, CCL4, CCL19 and CCL21 as molecular adjuvants on the immune response to VAA DNA vaccine in flounder (Paralichthys olivaceus). Dev Comp Immunol (2020) 103:103492. doi: 10.1016/j.dci.2019.103492
195. Guo M, Tang X, Sheng X, Xing J, Zhan W. The immune adjuvant effects of flounder (Paralichthys olivaceus) interleukin-6 on e. tarda subunit vaccine OmpV. Int J Mol Sci (2017) 18:1445. doi: 10.3390/ijms18071445
196. Guo M, Tang X, Sheng X, Xing J, Zhan W. The effects of IL-1β, IL-8, G-CSF and TNF-α as molecular adjuvant on the immune response to an e. tarda subunit vaccine in flounder (Paralichthys olivaceus). Fish Shellfish Immunol (2018) 77:374–84. doi: 10.1016/j.fsi.2018.04.009
197. Rathore G. Vaccines against bacterial pathogens of fish. In: Fish immune system and vaccines. (Singapore: Springer) (2022) 65–94. doi: 10.1007/978-981-19-1268-9_4
198. van Eerde A, Várnai A, Jameson JK, Paruch L, Moen A, Anonsen JH, et al. In-depth characterization of trichoderma reesei cellobiohydrolase TrCel7A produced in nicotiana benthamiana reveals limitations of cellulase production in plants by host-specific post-translational modifications. Plant Biotechnol J (2020) 18:631–43. doi: 10.1111/pbi.13227
199. Siriwattananon K, Manopwisedjaroen S, Kanjanasirirat P, Purwono PB, Rattanapisit K, Shanmugaraj B, et al. Development of plant-produced recombinant ACE2-fc fusion protein as a potential therapeutic agent against SARS-CoV-2. Front Plant Sci (2021) 11:604663. doi: 10.3389/fpls.2020.604663
200. El-Turkey A, El-Attar AK, Aboulata AE, Othman B, El-dougdoug KA. Expression of recombinant gD2 protein in transgenic tomato plants for development of a plant-derived vaccine against human herpes virus 2. Egypt J Virol (2014) 11:1–13. Available at: https://www.researchgate.net/%20publication/307175698.
201. Su H, Su J. Cyprinid viral diseases and vaccine development. Fish Shellfish Immunol (2018) 83:84–95. doi: 10.1016/j.fsi.2018.09.003
202. Buyel JF. Plant molecular farming - integration and exploitation of side streams to achieve sustainable biomanufacturing. Front Plant Sci (2019) 9:1893. doi: 10.3389/fpls.2018.01893
203. Lamichhane A, Azegami T, Kiyono H. The mucosal immune system for vaccine development. Vaccine (2014) 32:6711–23. doi: 10.1016/j.vaccine.2014.08.089
204. Singhal D, Mishra R. Edible vaccine-an effective way for immunization. Endocrine Metab Immune Disord Drug Targets (2022) . 22:1871–5303. doi: 10.2174/1871530322666220621102818
205. Mičúchová A, Piačková V, Frébort I, Korytávr T. Molecular farming: Expanding the field of edible vaccines for sustainable fish aquaculture. Rev Aquac (2022) 14:1978–2001. doi: 10.1111/raq.12683
206. Shin YJ, Kwon TH, Seo JY, Kim TJ. Oral immunization of fish against iridovirus infection using recombinant antigen produced from rice callus. Vaccine (2013) 31:5210–5. doi: 10.1016/j.vaccine.2013.08.085
207. Vidal-Meireles A, Neupert J, Zsigmond L, Rosado-Souza L, Kovács L, Nagy V, et al. Regulation of ascorbate biosynthesis in green algae has evolved to enable rapid stress-induced response via the VTC2 gene encoding GDP-l-galactose phosphorylase. New Phytol (2017) 214:668–81. doi: 10.1111/nph.14425
208. Charoonnart P, Purton S, Saksmerprome V. Applications of microalgal biotechnology for disease control in aquaculture. Biology-Basel (2018) 7:14. doi: 10.3390/biology7020024
209. Tabatabaei I, Bosco CD, Bednarska M, Ruf S, Meurer J, Bock R. A highly efficient sulfadiazine selection system for the generation of transgenic plants and algae. Plant Biotechnol J (2019) 17:638–49. doi: 10.1111/pbi.13004
210. Feng S, Feng W, Zhao L, Gu H, Li Q, Shi K, et al. Preparation of transgenic dunaliella salina for immunization against white spot syndrome virus in crayfish. Arch Virol (2014) 159:519–25. doi: 10.1007/s00705-013-1856-7
211. Kiataramgul A, Maneenin S, Purton S, Areechon N, Hirono I, Brocklehurst TW, et al. An oral delivery system for controlling white spot syndrome virus infection in shrimp using transgenic microalgae. Aquaculture (2020) 521:521. doi: 10.1016/j.aquaculture.2020.735022
212. Somchai P, Jitrakorn S, Thitamadee S, Meetam M, Saksmerprome V. Use of microalgae chlamydomonas reinhardtii for production of double-stranded RNA against shrimp virus. Aquac Rep (2016) 3:178–83. doi: 10.1016/j.aqrep.2016.03.003
213. Marsian J, Hurdiss DL, Ranson NA, Ritala A, Paley R, Cano I, et al. Plant-made nervous necrosis virus-like particles protect fish against disease. Front Plant Sci (2019) 10:880. doi: 10.3389/fpls.2019.00880
214. Shahin K, Pirezan F, Rogge M, LaFrentz BR, Shrestha RP, Hildebrand M, et al. Development of IglC and GroEL recombinant vaccines for francisellosis in Nile tilapia, oreochromis niloticus. Fish Shellfish Immunol (2020) 105:341–9. doi: 10.1016/j.fsi.2020.07.045
215. Ma K, Bao Q, Wu Y, Chen S, Zhao S, Wu H, et al. Evaluation of microalgae as immunostimulants and recombinant vaccines for diseases prevention and control in aquaculture. Front Bioeng Biotechnol (2020) 8:590431. doi: 10.3389/fbioe.2020.590431
216. Laere E, Ling APK, Wong YP, Koh RY, Lila MAM, Hussein S. Plant-based vaccines: Production and challenges. J Bot (2016) 2016:4928637. doi: 10.1155/2016/4928637
217. Anderson ED, Mourich DV, Fahrenkrug SC, LaPatra S, Shepherd J, Leong JAC. Genetic immunization of rainbow trout (Oncorhynchus mykiss) against infectious hematopoietic necrosis virus. Mol Mar Biol Biotechnol (1996) 5:114–22.
218. Hølvold LB, Myhr AI, Dalmo RA. Strategies and hurdles using DNA vaccines to fish. Vet Res (2014) 45:1–11. doi: 10.1186/1297-9716-45-21
219. Dalmo RA. DNA Vaccines for fish: Review and perspectives on correlates of protection. J Fish Dis (2018) 41:1–9. doi: 10.1111/jfd.12727
220. Huo X, Fan C, Ai T, Su J. The combination of molecular adjuvant CCL35. 2 and DNA vaccine significantly enhances the immune protection of carassius auratus gibelio against CyHV-2 infection. Vaccines (2020) 8:567. doi: 10.3390/vaccines8040567
221. Xing J, Xu H, Tang X, Sheng X, Zhan W. A DNA vaccine encoding the VAA gene of vibrio anguillarum induces a protective immune response in flounder. Front Immunol (2019) 10:499. doi: 10.3389/fimmu.2019.00499
222. Xu H, Xing J, Tang X, Sheng X, Zhan W. The effects of CCL3, CCL4, CCL19 and CCL21 as molecular adjuvants on the immune response to VAA DNA vaccine in flounder (Paralichthys olivaceus). Dev Comp Immunol (2020) 103:103492. doi: 10.1016/j.dci.2019.103492
223. Xing J, Jiang X, Xu H, Sheng X, Tang X, Chi H, et al. Local immune responses to VAA DNA vaccine against listonella anguillarum in flounder (Paralichthys olivaceus). Mol Immunol (2021) 134:141–9. doi: 10.1016/j.molimm.2021.03.012
224. Liu F, Tang X, Sheng X, Xing J, Zhan W. DNA Vaccine encoding molecular chaperone GroEL of edwardsiella tarda confers protective efficacy against edwardsiellosis. Mol Immunol (2016) 79:55–65. doi: 10.1016/j.molimm.2016.09.024
225. Sun Y, Ding S, He M, Liu A, Long H, Guo W, et al. Construction and analysis of the immune effect of vibrio harveyi subunit vaccine and DNA vaccine encoding TssJ antigen. Fish Shellfish Immunol (2020) 98:45–51. doi: 10.1016/j.fsi.2019.12.079
226. Saksida SM. Infectious haematopoietic necrosis epidemic (2001 to 2003) in farmed Atlantic salmon salmo salar in British Columbia. Dis Aquat Organ (2006) 72:213–23. doi: 10.3354/dao072213
227. Standish IF, Millard EV, Brenden TO, Faisal M. A DNA vaccine encoding the viral hemorrhagic septicemia virus genotype IVb glycoprotein confers protection in muskellunge (Esox masquinongy), rainbow trout (Oncorhynchus mykiss), brown trout (Salmo trutta), and lake trout (Salvelinus namaycush). Virol J (2016) 13:1–12. doi: 10.1186/s12985-016-0662-8
228. Thorarinsson R, Wolf JC, Inami M, Phillips L, Jones G, Macdonald AM, et al. Effect of a novel DNA vaccine against pancreas disease caused by salmonid alphavirus subtype 3 in Atlantic salmon (Salmo salar). Fish Shellfish Immunol (2021) 108:116-126. doi: 10.1016/j.fsi.2020.12.002
229. Chang CJ. Development and evaluation of DNA vaccine against salmonid alphavirus. In: Thomas S (eds) Vaccine Design. Methods in molecular biology (Humana, New York, NY) (2022) 2411. doi: 10.1007/978-1-0716-1888-2_12
230. Josepriya TA, Chien KH, Lin HY, Huang HN, Wu CJ, Song YL. Immobilization antigen vaccine adjuvanted by parasitic heat shock protein 70C confers high protection in fish against cryptocaryonosis. Fish Shellfish Immunol (2015) 45:517–27. doi: 10.1016/j.fsi.2015.04.036
231. Xu DH, Zhang D, Shoemaker C, Beck B. Immune response of channel catfish (Ictalurus punctatus) against ichthyophthirius multifiliis post vaccination using DNA vaccines encoding immobilization antigens. Fish Shellfish Immunol (2019) 94:308–17. doi: 10.1016/j.fsi.2019.08.071
232. Xu DH, Zhang D, Shoemaker C, Beck B. Dose effects of a DNA vaccine encoding immobilization antigen on immune response of channel catfish against ichthyophthirius multifiliis. Fish Shellfish Immunol (2020) 106:1031–41. doi: 10.1016/j.fsi.2020.07.063
233. Parenrengi A, Tenriulo A, Suryati E, Rosmiati R, Lante S, Azis AA, et al. Application of dsRNA VP15-WSSV by oral vaccination to increase survival rate and response immunes of tiger shrimp penaeus monodon. Indian J Anim Res (2022) 56:893–8. doi: 10.18805/IJAR.BF-1460
234. Jorge S, Dellagostin OA. The development of veterinary vaccines: a review of traditional methods and modern biotechnology approaches. Biotechnol Res Innov (2017) 1:6–13. doi: 10.1016/j.biori.2017.10.001
235. Jiang H, Bian Q, Zeng W, Ren P, Sun H, Lin Z, et al. Oral delivery of bacillus subtilis spores expressing grass carp reovirus VP4 protein produces protection against grass carp reovirus infection. Fish Shellfish Immunol (2019) 84:768–80. doi: 10.1016/j.fsi.2018.10.008
236. Yao YY, Chen DD, Cui ZW, Zhang XY, Zhou YY, Guo X, et al. Oral vaccination of tilapia against streptococcus agalactiae using bacillus subtilis spores expressing sip. Fish Shellfish Immunol (2019) 86:999–1008. doi: 10.1016/j.fsi.2018.12.060
237. Naderi-Samani M, Soltani M, Dadar M, Taheri-Mirghaed A, Zargar A, Ahmadivand S, et al. Oral immunization of trout fry with recombinant lactococcus lactis NZ3900 expressing G gene of viral hemorrhagic septicaemia virus (VHSV). Fish Shellfish Immunol (2020) 105:62–70. doi: 10.1016/j.fsi.2020.07.007
238. Sun H, Shang M, Tang Z, Jiang H, Dong H, Zhou X, et al. Oral delivery of bacillus subtilis spores expressing clonorchis sinensis paramyosin protects grass carp from cercaria infection. Appl Microbiol Biotechnol (2020) 104:1633–46. doi: 10.1007/s00253-019-10316-0
239. Zhang DX, Kang YH, Chen L, Siddiqui SA, Wang CF, Qian AD, et al. Oral immunization with recombinant lactobacillus casei expressing OmpAI confers protection against aeromonas veronii challenge in common carp, cyprinus carpio. Fish Shellfish Immunol (2018) 72:552–63. doi: 10.1016/j.fsi.2017.10.043
240. Kong Y, Kang YH, Tian JX, Zhang DX, Zhang L, Tao LT, et al. Oral immunization with recombinant lactobacillus casei expressing flaB confers protection against aeromonas veronii challenge in common carp, cyprinus carpio. Fish Shellfish Immunol (2019) 87:627–37. doi: 10.1016/j.fsi.2019.01.032
241. Kato T, Sugioka S, Itagaki K, Park EY. Gene transduction in mammalian cells using bombyx mori nucleopolyhedrovirus assisted by glycoprotein 64 of autographa californica multiple nucleopolyhedrovirus. Sci Rep (2016) 6:1–9. doi: 10.1038/srep32283
242. Kwang TW, Zeng X, Wang S. Manufacturing of AcMNPV baculovirus vectors to enable gene therapy trials. Mol Therapy-Methods Clin Dev (2016) 3:15050. doi: 10.1038/mtm.2015.50
243. Zhao JZ, Liu M, Xu LM, Zhang ZY, Cao YS, Shao YZ, et al. A chimeric recombinant infectious hematopoietic necrosis virus induces protective immune responses against infectious hematopoietic necrosis and infectious pancreatic necrosis in rainbow trout. Mol Immunol (2019) 116:180–90. doi: 10.1016/j.molimm.2019.10.015
244. Li K, Yuan R, Zhang M, Zhang T, Gu Y, Zhou Y, et al. Recombinant baculovirus BacCarassius-D4ORFs has potential as a live vector vaccine against CyHV-2. Fish Shellfish Immunol (2019) 92:101–10. doi: 10.1016/j.fsi.2019.05.065
245. Vallejos-Vidal E, Reyes-López F, MacKenzie S. Immunostimulant diets and oral vaccination in fish. In: Diagnosis and control of diseases of fish and shellfish (2017) 147-184. doi: 10.1002/9781119152125.ch6
246. Zhang C, Zhao Z, Liu GY, Li J, Wang GX, Zhu B. Immune response and protective effect against spring viremia of carp virus induced by intramuscular vaccination with a SWCNTs-DNA vaccine encoding matrix protein. Fish Shellfish Immunol (2018) 79:256–64. doi: 10.1016/j.fsi.2018.05.029
247. Ramírez-Paredes JG, Mendoza-Roldan MA, Lopez-Jimena B, Shahin K, Metselaar M, Thompson KD, et al. Whole cell inactivated autogenous vaccine effectively protects red Nile tilapia (Oreochromis niloticus) against francisellosis via intraperitoneal injection. J Fish Dis (2019) 42:1191–200. doi: 10.1111/jfd.13041
248. Huising MO, Guichelaar T, Hoek C, Verburg-van Kemenade BML, Flik G, Savelkoul HFJ, et al. Increased efficacy of immersion vaccination in fish with hyperosmotic pretreatment. Vaccine (2003) 21:4178–93. doi: 10.1016/S0264-410X(03)00497-3
249. Chettri JK, Deshmukh S, Holten-Andersen L, Jafaar RM, Dalsgaard I, Buchmann K, et al. Comparative evaluation of administration methods for a vaccine protecting rainbow trout against yersinia ruckeri O1 biotype 2 infections. Vet Immunol Immunopathol (2013) 154:42–7. doi: 10.1016/j.vetimm.2013.04.001
250. Weiner HL, da Cunha AP, Quintana F, Wu H. Oral tolerance. Immunol Rev (2011) 241:241–59. doi: 10.1111/j.1600-065X.2011.01017.x
251. Tobar JA, Jerez S, Caruffo M, Bravo C, Contreras F, Bucarey SA, et al. Oral vaccination of Atlantic salmon (Salmo salar) against salmonid rickettsial septicaemia. Vaccine (2011) 29:2336–40. doi: 10.1016/j.vaccine.2010.12.107
252. Liang R, Li J, Ruan Y, Zhu S. The design and parameter optimization of automatic positioning mechanism for automatic injection system of fry vaccine. Am Soc Agric Biol Engineers Annu Int Meeting 2015 (2015) 1:1-18. doi: 10.13031/aim.20152188971
253. Corbeil S, Kurath G, Lapatra SE. Fish DNA vaccine against infectious hematopoietic necrosis virus: Efficacy of various routes of immunisation. Fish Shellfish Immunol (2000) 10:711–23. doi: 10.1006/fsim.2000.0286
254. Wolf A, Hodneland K, Frost P, Hoeijmakers M, Rimstad E. Salmonid alphavirus-based replicon vaccine against infectious salmon anemia (ISA): Impact of immunization route and interactions of the replicon vector. Fish Shellfish Immunol (2014) 36:383-392. doi: 10.1016/j.fsi.2013.12.018
255. Droshnev AE, Zavyalova EA, Bulina KY, Alontseva DA. Prevention of salmon yersiniosis in industrial cultivation. AIP Conf Proc (2022) 2390:030014-1–4. doi: 10.1063/5.0070600
256. Du Y, Tang X, Sheng X, Xing J, Zhan W. The influence of concentration of inactivated edwardsiella tarda bacterin and immersion time on antigen uptake and expression of immune-related genes in Japanese flounder (Paralichthys olivaceus). Microb Pathog (2017) 103:19–28. doi: 10.1016/j.micpath.2016.12.011
257. Song YL, Kou GH. The immuno-responses of eel (Anguilla japonica) against edwardsiella anguillimortifera as studied by the immersion method. Fish Pathol (1981) 15:249–55. doi: 10.3147/jsfp.15.249
258. Mikkelsen H, Lund V, Larsen R, Seppola M. Vibriosis vaccines based on various sero-subgroups of vibrio anguillarum O2 induce specific protection in Atlantic cod (Gadus morhua l. ) juveniles. Fish Shellfish Immunol (2011) 30:330–9. doi: 10.1016/j.fsi.2010.11.007
259. Liang H-J, Peng R-H, Chiou PP. Development of immersion subunit vaccine against grouper iridovirus (GIV). Fish Shellfish Immunol (2016) 53:104. doi: 10.1016/j.fsi.2016.04.062
260. Aonullah AA, Nuryati S, Alimuddin, Murtini S. Efficacy of koi herpesvirus DNA vaccine administration by immersion method on cyprinus carpio field scale culture. Aquac Res (2017) 48:2655–62. doi: 10.1111/are.13097
261. Skov J, Chettri JK, Jaafar RM, Kania PW, Dalsgaard I, Buchmann K. Effects of soluble immunostimulants on mucosal immune responses in rainbow trout immersion-vaccinated against yersinia ruckeri. Aquaculture (2018) 492:237–46. doi: 10.1016/j.aquaculture.2018.04.011
262. Rombout JHWM, Kiron V. Mucosal vaccination of fish. Fish vaccination (2014), 56–67. doi: 10.1002/9781118806913.ch6
263. Plant KP, LaPatra SE. Advances in fish vaccine delivery. Dev Comp Immunol (2011) 35:1256–62. doi: 10.1016/j.dci.2011.03.007
264. Du Y, Tang X, Sheng X, Xing J, Zhan W. Immune response of flounder (Paralichthys olivaceus) was associated with the concentration of inactivated edwardsiella tarda and immersion time. Vet Immunol Immunopathol (2015) 167:44–50. doi: 10.1016/j.vetimm.2015.07.002
265. Chettri JK, Jaafar RM, Skov J, Kania PW, Dalsgaard I, Buchmann K, et al. Booster immersion vaccination using diluted yersinia ruckeri bacterin confers protection against ERM in rainbow trout. Aquaculture (2015) 440:1–5. doi: 10.1016/j.aquaculture.2015.01.027
266. Gao YL, Tang XQ, Sheng XZ, Xing J, Zhan WB. Immune responses of flounder paralichthys olivaceus vaccinated by immersion of formalin-inactivated edwardsiella tarda following hyperosmotic treatment. Dis Aquat Organ (2015) 116:111–20. doi: 10.3354/dao02909
267. Vinay TN, Bedekar MK. Methods of vaccine Delivery[M]. In: Fish immune system and vaccines. Singapore: Springer (2022). p. 217–30.
268. Linh NV, Dien LT, Sangpo P, Senapin S, Thapinta A, Panphut W, et al. Pre-treatment of Nile tilapia (Oreochromis niloticus) with ozone nanobubbles improve efficacy of heat-killed streptococcus agalactiae immersion vaccine. Fish Shellfish Immunol (2022) 123:229–37. doi: 10.1016/j.fsi.2022.03.007
269. Bøgwald J, Dalmo RA. Review on immersion vaccines for fish: An update 2019. Microorganisms (2019) 7:627. doi: 10.3390/microorganisms7120627
270. Thangsunan P, Kitiyodom S, Srisapoome P, Pirarat N, Yata T, Thangsunan P, et al. Novel development of cationic surfactant-based mucoadhesive nanovaccine for direct immersion vaccination against francisella noatunensis subsp. orientalis in red tilapia (Oreochromis sp.). Fish Shellfish Immunol (2022) 127:1051–60. doi: 10.1016/j.fsi.2022.07.056
271. Li L, Zhang T, Zhang G, Zhou G, Yang F, Wang E, et al. High immune efficiency of bacterial nanocellulose loaded MSRV G protein vaccine for bath immunization. Aquaculture (2022) 560:738579. doi: 10.1016/j.aquaculture.2022.738579
272. Kitiyodom S, Kaewmalun S, Nittayasut N, Suktham K, Surassmo S, Namdee K, et al. The potential of mucoadhesive polymer in enhancing efficacy of direct immersion vaccination against flavobacterium columnare infection in tilapia. Fish Shellfish Immunol (2019) 86:635–40. doi: 10.1016/j.fsi.2018.12.005
273. Alawdi S, Solanki AB. Mucoadhesive drug delivery systems: A review of recent developments. J Sci Res Med Biol Sci (2021) 2:50–64. doi: 10.47631/jsrmbs.v2i1.213
274. Bunnoy A, Thangsunan P, Chokmangmeepisarn P, Yata T, Klongklaew N, Pirarat N, et al. Mucoadhesive cationic lipid-based flavobacterium oreochromis nanoencapsulation enhanced the efficacy of mucoadhesive immersion vaccination against columnaris disease and strengthened immunity in Asian sea bass (Lates calcarifer). Fish Shellfish Immunol (2022) 127:633–46. doi: 10.1016/j.fsi.2022.06.059
275. Embregts CWE, Forlenza M. Oral vaccination of fish: Lessons from humans and veterinary species. Dev Comp Immunol (2016) 64:118–37. doi: 10.1016/j.dci.2016.03.024
276. Jin P, Sun F, Liu Q, Wang Q, Zhang Y, Liu X. An oral vaccine based on chitosan/aluminum adjuvant induces both local and systemic immune responses in turbot (Scophthalmus maximus). Vaccine (2021) 39:7477–84. doi: 10.1016/j.vaccine.2021.10.063
277. Andresen AMS, Gjoen T. Chitosan nanoparticle formulation attenuates poly (I:C) induced innate immune responses against inactivated virus vaccine in Atlantic salmon (Salmo salar). Comp Biochem Physiol D-Genomics Proteomics (2021) 40:100915. doi: 10.1016/j.cbd.2021.100915
278. Sukkarun P, Kitiyodom S, Yostawornkul J, Chaiin P, Yata T, Rodkhum C, et al. Chitosan-polymer based nanovaccine as promising immersion vaccine against aeromonas veronii challenge in red tilapia (Oreochromis sp. ). Fish Shellfish Immunol (2022) 129:30–5. doi: 10.1016/j.fsi.2022.08.035
279. Angulo C, Sanchez V, Delgado K, Monreal-Escalante E, Hernandez-Adame L, Angulo M, et al. Oral organic nanovaccines against bacterial and viral diseases. Microb Pathog (2022) 169:105648. doi: 10.1016/j.micpath.2022.105648
280. Samms KA, Alkie TN, Jenik K, de Jong J, Klinger KM, DeWitte-Orr SJ. Oral delivery of a dsRNA-phytoglycogen nanoparticle complex enhances both local and systemic innate immune responses in rainbow trout. Fish Shellfish Immunol (2022) 121:215–22. doi: 10.1016/j.fsi.2021.12.038
281. Ballesteros NA, Alonso M, Saint-Jean SR, Perez-Prieto SI. An oral DNA vaccine against infectious haematopoietic necrosis virus (IHNV) encapsulated in alginate microspheres induces dose-dependent immune responses and significant protection in rainbow trout (Oncorrhynchus mykiss). Fish Shellfish Immunol (2015) 45:877–88. doi: 10.1016/j.fsi.2015.05.045
282. Caruffo M, Maturana C, Kambalapally S, Larenas J, Tobar JA. Protective oral vaccination against infectious salmon anaemia virus in salmo salar. Fish Shellfish Immunol (2016) 54:54–9. doi: 10.1016/j.fsi.2016.03.009
283. Kim Y, Kim H, Beuchat LR, Ryu JH. Development of non-pathogenic bacterial biofilms on the surface of stainless steel which are inhibitory to salmonella enterica. Food Microbiol (2018) 69:136–42. doi: 10.1016/j.fm.2017.08.003
284. Siriyappagouder P, Shankar KM, Kumar BTN, Patil R, Byadgi OV. Evaluation of biofilm of aeromonas hydrophila for oral vaccination of channa striatus. Fish Shellfish Immunol (2014) 41:581–5. doi: 10.1016/j.fsi.2014.09.021
285. Mohamad A, Mursidi F-A, Zamri-Saad M, Amal MNA, Annas S, Monir MS, et al. Laboratory and field assessments of oral vibrio vaccine indicate the potential for protection against vibriosis in cultured marine fishes. Aniamls (2022) 12:133. doi: 10.3390/ani12020133
286. Perrie Y, Crofts F, Devitt A, Griffiths HR, Kastner E, Nadella V. Designing liposomal adjuvants for the next generation of vaccines. Adv Drug Delivery Rev (2016) 99:85–96. doi: 10.1016/j.addr.2015.11.005
287. Embregts CWE, Rigaudeau D, Tacchi L, Pijlman GP, Kampers L, Veselý T, et al. Vaccination of carp against SVCV with an oral DNA vaccine or an insect cells-based subunit vaccine. Fish Shellfish Immunol (2019) 85:66–77. doi: 10.1016/j.fsi.2018.03.028
288. Kang SH, Hong SJ, Lee YK, Cho S. Oral vaccine delivery for intestinal immunity-biological basis, barriers, delivery system, and m cell targeting. Polymers (Basel) (2018) 10:948. doi: 10.3390/polym10090948
289. Klug B, Robertson JS, Condit RC, Seligman SJ, Laderoute MP, Sheets R, et al. Adventitious agents and live viral vectored vaccines: Considerations for archiving samples of biological materials for retrospective analysis. Vaccine (2016) 34:6617–25. doi: 10.1016/j.vaccine.2016.02.015
290. Yu YY, Kong WG, Xu HY, Huang ZY, Zhang XT, Ding LG, et al. Convergent evolution of mucosal immune responses at the buccal cavity of teleost fish. iScience (2019) 19:821–35. doi: 10.1016/j.isci.2019.08.034
291. Soleto I, Granja AG, Simón R, Morel E, Díaz-Rosales P, Tafalla C. Identification of CD8α+ dendritic cells in rainbow trout (Oncorhynchus mykiss) intestine. Fish Shellfish Immunol (2019) 89:309–18. doi: 10.1016/j.fsi.2019.04.001
292. Jan JHWMR, Abelli L, Picchietti S, Scapigliati G, Kiron V. Teleost intestinal immunology. Fish Shellfish Immunol (2011) 31:616–26. doi: 10.1016/j.fsi.2010.09.001
293. Sunyer JO, Boudinot P. B-cell responses and antibody repertoires in teleost fish: From Ag receptor diversity to immune memory and vaccine development. Princ Fish Immunol (2022), 253–78. doi: 10.1007/978-3-030-85420-1_8
294. Johansson MEV, Phillipson M, Petersson J, Velcich A, Holm L, Hansson GC. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc Natl Acad Sci USA (2008) 105:15064–9. doi: 10.1073/pnas.0803124105
295. Wong G, Kaattari SL, Christensen JM. Effectiveness of an oral enteric coated vibrio vaccine for use in salmonid fish. Immunol Invest (1992) 21:353–64. doi: 10.3109/08820139209069375
296. Salinas I. The mucosal immune system of teleost fish. Biol (Basel) (2015) 4:525–39. doi: 10.3390/biology4030525
297. Rességuier J, Delaune E, Coolen AL, Levraud JP, Boudinot P, Guellec D, et al. Specific and efficient uptake of surfactant-free poly(lactic acid) nanovaccine vehicles by mucosal dendritic cells in adult zebrafish after bath immersion. Front Immunol (2017) 8:190. doi: 10.3389/fimmu.2017.00190
298. Salinas I, Ding Y, Fernández-Montero Á, Sunyer JO. Mucosal immunity in fish. In: Principles of fish immunology. (Springer, Cham) (2022) 387–443. doi: 10.1007/978-3-030-85420-1_12
299. Miller NW, Sizemore RC, Clem LW. Phylogeny of lymphocyte heterogeneity: the cellular requirements for in vitro antibody responses of channel catfish leukocytes. J Immunol (1985) 134:2884–8.
300. Li J, Barreda DR, Zhang YA, Boshra H, Gelman AE, LaPatra S, et al. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Nat Immunol (2006) 7:1116–24. doi: 10.1038/ni1389
301. van der Wal YA, Jenberie S, Nordli H, Greiner-Tollersrud L, Kool J, Jensen I, et al. The importance of the Atlantic salmon peritoneal cavity b cell response: Local IgM secreting cells are predominant upon piscirickettsia salmonis infection. Dev Comp Immunol (2021) 123:104125. doi: 10.1016/j.dci.2021.104125
302. Pilström L, Lundqvist ML, Wermenstam NE. The immunoglobulin light chain in poikilothermic vertebrates. Immunol Rev (1998) 166:123–32. doi: 10.1111/j.1600-065X.1998.tb01257.x
303. Saha NR, Suetake H, Kikuchi K, Suzuki Y. Fugu immunoglobulin d: A highly unusual gene with unprecedented duplications in its constant region. Immunogenetics (2004) 56:438–47. doi: 10.1007/s00251-004-0693-y
304. Danilova N, Bussmann J, Jekosch K, Steiner LA. The immunoglobulin heavy-chain locus in zebrafish: Identification and expression of a previously unknown isotype, immunoglobulin z. Nat Immunol (2005) 6:295–302. doi: 10.1038/ni1166
305. Hansen JD, Landis ED, Phillips RB. Discovery of a unique ig heavy-chain (IgT) in rainbow trout: Implications for a distinctive b cell developmental pathway in teleost fish. Proc Natl Acad Sci USA (2005) 102:6919–24. doi: 10.1073/pnas.0500027102
306. Savan R, Aman A, Nakao M, Watanuki H, Sakai M. Discovery of a novel immunoglobulin heavy chain gene chimera from common carp (Cyprinus carpio l.). Immunogenetics (2005) 57:458–63. doi: 10.1007/s00251-005-0015-z
307. Sheng X, Qian X, Tang X, Xing J, Zhan W. Polymeric immunoglobulin receptor mediates immune excretion of mucosal IgM-antigen complexes across intestinal epithelium in flounder (Paralichthys olivaceus). Front Immunol (2018) 9:1562. doi: 10.3389/fimmu.2018.01562
308. Vervarcke S, Ollevier F, Kinget R, Michoel A. Mucosal response in African catfish after administration of vibrio anguillarum O2 antigens via different routes. Fish Shellfish Immunol (2005) 18:125–33. doi: 10.1016/j.fsi.2004.06.004
309. Zeng C, Tang X, Du Y, Sheng X, Xing J, Zhan W. Dynamic distribution of formalin-inactivated edwardsiella tarda in olive flounder (Paralichthys olivaceus) post intramuscular injection. Vet Immunol Immunopathol (2018) 199:53–60. doi: 10.1016/j.vetimm.2018.03.007
310. Silvaraj S, Yasin ISM, Karim MMA, Saad MZ. Transcriptome analysis of immune response in recombinant cell vaccine expressing OmpK vaccinated juvenile seabass (lates calcarifer) head kidney against vibrio harveyi infection. Aquac Rep (2021) 21:100799. doi: 10.1016/j.aqrep.2021.100799
311. Xue T, Liu Y, Cao M, Li J, Tian M, Zhang L, et al. Transcriptome analysis reveals deep insights into the early immune response of turbot (Scophthalmus maximus) induced by inactivated aeromonas salmonicida vaccine. Fish Shellfish Immunol (2021) 119:163–72. doi: 10.1016/j.fsi.2021.09.027
312. Tian H, Xing J, Tang X, Chi H, Sheng X, Zhan W. Cluster of differentiation antigens: essential roles in the identification of teleost fish T lymphocytes. Mar Life Sci Technol (2022) 4:303–16. doi: 10.1007/s42995-022-00136-z
313. Restifo NP, Ying H, Hwang L, Leitner WW. The promise of nucleic acid vaccines. Gene Ther (2000) 7:89–92. doi: 10.1038/sj.gt.3301117
314. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature (1998) 392:245–52. doi: 10.1038/32588
315. Bedekar MK, Kole S. DNA Vaccines for fish. In: Advances in fisheries biotechnology. (Singapore: Springer) (2021) 289–336. doi: 10.1007/978-981-16-3215-0
316. Jakob T, Walker PS, Krieg AM, Udey MC, Vogel JC. Activation of cutaneous dendritic cells by CpG-containing oligodeoxynucleotides: a role for dendritic cells in the augmentation of Th1 responses by immunostimulatory DNA. J Immunol (1998) 161:3042–9.
317. Tonheim TC, Bøgwald J, Dalmo RA. What happens to the DNA vaccine in fish? a review of current knowledge. Fish Shellfish Immunol (2008) 25:1–18. doi: 10.1016/j.fsi.2008.03.007
318. Latz E, Schoenemeyer A, Visintin A, Fitzgerald KA, Monks BG, Knetter CF, et al. TLR9 signals after translocating from the ER to CpG DNA in the lysosome. Nat Immunol (2004) 5:190–8. doi: 10.1038/ni1028
319. Chang CJ. Immune sensing of DNA and strategies for fish DNA vaccine development. Fish Shellfish Immunol (2020) 101:252–60. doi: 10.1016/j.fsi.2020.03.064
320. Xu DH, Klesius PH, Shelby RA. Cutaneous antibodies in excised skin from channel catfish, ictalurus punctatus rafinesque, immune to ichthyophthirius multifiliis. J Fish Dis (2002) 25:45–52. doi: 10.1046/j.1365-2761.2002.00339.x
321. Mulder IE, Wadsworth S, Secombes CJ. Cytokine expression in the intestine of rainbow trout (Oncorhynchus mykiss) during infection with aeromonas salmonicida. Fish Shellfish Immunol (2007) 23:747–59. doi: 10.1016/j.fsi.2007.02.002
322. Giri SS, Park SC. Application of carbon nanotubes in the advancement of fish vaccine. In: Biotechnological advances in aquaculture health management. (Singapore: Springer) (2021) 61–78. doi: 10.1007/978-981-16-5195-3_4
323. Esteve-Gassent MD, Fouz B, Amaro C. Efficacy of a bivalent vaccine against eel diseases caused by vibrio vulnificus after its administration by four different routes. Fish Shellfish Immunol (2004) 16:93–105. doi: 10.1016/S1050-4648(03)00036-6
324. Midtlyng PJ. Current use and need for new fish vaccines. In: Principles of fish immunology. (Springer, Cham) (2022) 599–608. doi: 10.1007/978-3-030-85420-1_19
325. Stosik M, Tokarz-Deptuła B, Deptuła W. Immunological memory in teleost fish. Fish Shellfish Immunol (2021) 115:95–103. doi: 10.1016/j.fsi.2021.05.022
326. Xu Z, Parra D, Gomez D, Salinas I, Zhang YA, von Gersdorff Jørgensen L, et al. Teleost skin, an ancient mucosal surface that elicits gut-like immune responses. Proc Natl Acad Sci USA (2013) 110:13097–102. doi: 10.1073/pnas.1304319110
327. Xu Z, Takizawa F, Parra D, Gómez D, Jørgensen LVG, Lapatra SE, et al. Mucosal immunoglobulins at respiratory surfaces mark an ancient association that predates the emergence of tetrapods. Nat Commun (2016) 7:1–14. doi: 10.1038/ncomms10728
328. Lumsden JS, Ostland VE, MacPhee DD, Ferguson HW. Production of gill-associated and serum antibody byrainbow trout (Oncorhynchus mykiss) following immersion immunization with acetone-killed flavobacterium branchiophilum and the relationship to protection from experimental challenge. Fish Shellfish Immunol (1995) 5:151–65. doi: 10.1016/S1050-4648(05)80024-5
329. Morrison RN, Cooper GA, Koop BF, Rise ML, Bridle AR, Adams MB, et al. Transcriptome profiling the gills of amoebic gill disease (AGD)-affected Atlantic salmon (Salmo salar l.): A role for tumor suppressor p53 in AGD pathogenesis? Physiol Genomics (2006) 26:15–34. doi: 10.1152/physiolgenomics.00320.2005
330. Muñoz-Atienza E, Díaz-Rosales P, Tafalla C. Systemic and mucosal b and T cell responses upon mucosal vaccination of teleost fish. Front Immunol (2021) 11:622377. doi: 10.3389/fimmu.2020.622377
331. Oliveira TF, Quieroz GA, Leibowitz MP, Leal CAG. Development of an inactivated whole cell vaccine through immersion for immunoprophylaxis of francisella orientalis infections in Nile tilapia (Oreochromis niloticus l.) fingerlings and juveniles. Fish Shellfish Immunol (2022) 127:405–11. doi: 10.1016/j.fsi.2022.06.040
332. Schep LJ, Tucker IG, Young G, Ledger R, Butt AG. Controlled release opportunities for oral peptide delivery in aquaculture. J Control Release (1999) 59:1–14. doi: 10.1016/S0168-3659(98)00175-8
333. Press C, Evensen Ø. The morphology of the immune system in teleost fishes. Fish Shellfish Immunol (1999) 9:309–18. doi: 10.1006/fsim.1998.0181
334. Monir MS, Yusoff MSM, Zamri-Saad M, Amal MNA, Mohamad A, Azzam-Sayuti M, et al. Effect of an oral bivalent vaccine on immune response and immune gene profiling in vaccinated red tilapia (Oreochromis spp.) during infections with streptococcus iniae and aeromonas hydrophila. Biology-Basel (2022) 11:1628. doi: 10.3390/biology11091268
335. Mutoloki S, Munang’andu HM, Evensen Ø. Oral vaccination of fish - antigen preparations, uptake, and immune induction. Front Immunol (2015) 6:519. doi: 10.3389/fimmu.2015.00519
336. Companjen AR, Florack DEA, Bastiaans JHMW, Matos CI, Bosch D, Rombout JHWM. Development of a cost-effective oral vaccination method against viral disease in fish. Developments Biologicals. (2005) 121:143–50.
337. Salinas I, Zhang YA, Sunyer JO. Mucosal immunoglobulins and b cells of teleost fish. Dev Comp Immunol (2011) 35:1346–65. doi: 10.1016/j.dci.2011.11.009
338. Sun J, Zhang M, Zhao D, Yang J, Shi Y, Xu B, et al. Immunological effects of recombinant lactobacillus casei expressing IHNV G protein and rainbow trout (Oncorhynchus mykiss) chemokine CK6 as an oral vaccine. Front Immunol (2022) 13:9274439. doi: 10.3389/fimmu.2022.9274439
339. Zhang YA, Salinas I, Li J, Parra D, Bjork S, Xu Z, et al. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat Immunol (2010) 11:827–35. doi: 10.1038/ni.1913
340. Ballesteros NA, Castro R, Abos B, Saint-Jean SSR, Pérez-Prieto SI, Tafalla C. The pyloric caeca area is a major site for IgM and IgT b cell recruitment in response to oral vaccination in rainbow trout. PloS One (2013) 8:e66118. doi: 10.1371/journal.pone.0066118
341. Tordesillas L, Berin MC. Mechanisms of oral tolerance. Clin Rev Allergy Immunol (2018) 55:107–17. doi: 10.1007/s12016-018-8680-5
342. Maurice S, Nussinovitch A, Jaffe N, Shoseyov O, Gertler A. Oral immunization of carassius auratus with modified recombinant a-layer proteins entrapped in alginate beads. Vaccine (2004) 23:450–9. doi: 10.1016/j.vaccine.2004.06.022
343. Joosten PHM, Engelsma MY, Van Der Zee MD, Rombout JHWM. Induction of oral tolerance in carp (Cyprinus carpio l.) after feeding protein antigens. Vet Immunol Immunopathol (1997) 60:187–96. doi: 10.1016/S0165-2427(97)00124-4
344. Yu Y, Wang Q, Huang Z, Ding L, Xu Z. Immunoglobulins, mucosal immunity and vaccination in teleost fish. Front Immunol (2020) 11:567941. doi: 10.3389/fimmu.2020.567941
345. Chen L, Klaric G, Wadsworth S, Jayasinghe S, Kuo TY, Evensen Ø, et al. Augmentation of the antibody response of Atlantic salmon by oral administration of alginate-encapsulated IPNV antigens. PloS One (2014) 9:e109337. doi: 10.1371/journal.pone.0109337
346. Hoare R, Leigh W, Limakom T, Wongwaradechkul R, Metselaar M, Shinn AP, et al. Oral vaccination of Nile tilapia (Oreochromis niloticus) against francisellosis elevates specific antibody titres in serum and mucus. Fish Shellfish Immunol (2021) 113:86–8. doi: 10.1016/j.fsi.2021.03.019
347. Guo M, Li C. An overview of cytokine used as adjuvants in fish: current state and future trends. Rev Aquaculture (2021) 13:996–1014. doi: 10.1111/raq.12509
348. Ghimire TR, Benson RA, Garside P, Brewer JM. Alum increases antigen uptake, reduces antigen degradation and sustains antigen presentation by DCs in vitro. Immunol Lett (2012) 147:55–62. doi: 10.1016/j.imlet.2012.06.002
349. Sughra F, Hafeez-Ur-Rahman M, Abbas F, Altaf I. Evaluation of three alum-precipitated aeromonas hydrophila vaccines administered to labeo rohita, cirrhinus mrigala and ctenopharyngodon idella: Immunokinetics, immersion challenge and histopathology. Braz J Biol (2023) 83:249913. doi: 10.1590/1519-6984.249913
350. Sun F, Zhou J, Zhang Y, Liu Q, Wang Q, Liu X. A compound ginseng stem leaf saponins and aluminium adjuvant enhances the potency of inactivated aeromonas salmonicida vaccine in turbot. Fish Shellfish Immunol (2022) 128:60–6. doi: 10.1016/j.fsi.2022.07.027
351. Oreskovic Z, Nechvatalova K, Krejci J, Kummer V, Faldyna M. Aspects of intradermal immunization with different adjuvants: The role of dendritic cells and Th1/Th2 response. PloS One (2019) 14:e0211896. doi: 10.1371/journal.pone.0211896
352. Raman RP, Kumar S. Adjuvants for fish vaccines. In: Fish immune system and vaccines. Singapore: Springer (2022). p. 231–44.
353. Mutoloki S, Alexandersen S, Evensen Ø. Sequential study of antigen persistence and concomitant inflammatory reactions relative to side-effects and growth of Atlantic salmon (Salmo salar l.) following intraperitoneal injection with oil-adjuvanted vaccines. Fish Shellfish Immunol (2004) 16:633–44. doi: 10.1016/j.fsi.2003.10.002
354. Apostólico JDS, Lunardelli VAS, Coirada FC, Boscardin SB, Rosa DS. Adjuvants: Classification, modus operandi, and licensing. J Immunol Res (2016) 2016:16. doi: 10.1155/2016/1459394
355. Hwang JY, Kwon MG, Kim YJ, Jung SH, Park MA, Son MH. Montanide IMS 1312 VG adjuvant enhances the efficacy of immersion vaccine of inactivated viral hemorrhagic septicemia virus (VHSV) in olive flounder, paralichthys olivaceus. Fish Shellfish Immunol (2017) 60:420–5. doi: 10.1016/j.fsi.2016.12.011
356. Fan Y, Moon JJ. Particulate delivery systems for vaccination against bioterrorism agents and emerging infectious pathogens. Wiley Interdiscip Rev Nanomedicine Nanobiotechnology (2017) 9:e1403. doi: 10.1002/wnan.1403
357. Nasr-Eldahan S, Nabil-Adam A, Shreadah MA, Maher AM, El-Sayed Ali T. A review article on nanotechnology in aquaculture sustainability as a novel tool in fish disease control. Aquac Int (2021) 29:1459–80. doi: 10.1007/s10499-021-00677-7
358. Sarkar B, Mahanty A, Gupta SK, Choudhury AR, Daware A, Bhattacharjee S. Nanotechnology: A next-generation tool for sustainable aquaculture. Aquaculture (2022) 546:737330. doi: 10.1016/j.aquaculture.2021.737330
359. Boroumand H, Badie F, Mazaheri S, Seyedi ZS, Nahand JS, Nejati M, et al. Chitosan-based nanoparticles against viral infections. Front Cell Infect Microbiol (2021) 11:643953. doi: 10.3389/fcimb.2021.643953
360. Pumchan A, Sae-Ueng U, Prasittichai C, Sirisuay S, Areechon N, Unajak S. A novel efficient piscine oral nano-vaccine delivery system: Modified halloysite nanotubes (HNTs) preventing streptococcosis disease in tilapia (Oreochromis sp.). Vaccines (2022) 10:1180. doi: 10.3390/vaccines10081180
361. Bhat IA. Nanotechnology in reproduction, breeding and conservation of fish biodiversity: Current status and future potential. Rev Aquac (2022) 2022:1–11. doi: 10.1111/raq.12736
362. Zhu S, Miao B, Zhang YZ, Zeng WW, Wang DS, Su SQ. In vitro neutralization of nervous necrosis virus by a nanobody binding to the protrusion domain of capsid protein. Aquaculture (2022) 548:737654. doi: 10.1016/j.aquaculture.2021.737654
363. Wang Z, Xu C, Zhang Y, Huo X, Su J. Dietary supplementation with nanoparticle CMCS-20a enhances the resistance to GCRV infection in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol (2022) 127:572–84. doi: 10.1016/j.fsi.2022.07.004
364. Huo X, Wang Z, Xiao X, Yang C, Su J. Oral administration of nanopeptide CMCS-20H conspicuously boosts immunity and precautionary effect against bacterial infection in fish. Front Immunol (2022) 12:811616. doi: 10.3389/fimmu.2021.811616
365. Peres C, Matos AI, Conniot J, Sainz V, Zupančič E, Silva JM, et al. Poly(lactic acid)-based particulate systems are promising tools for immune modulation. Acta Biomater (2017) 48:41–57. doi: 10.1016/j.actbio.2016.11.012
366. Behera T, Nanda PK, Mohanty C, Mohapatra D, Swain P, Das BK, et al. Parenteral immunization of fish, labeo rohita with poly d, l-lactide-co-glycolic acid (PLGA) encapsulated antigen microparticles promotes innate and adaptive immune responses. Fish Shellfish Immunol (2010) 28:320–5. doi: 10.1016/j.fsi.2009.11.009
367. Guy B. The perfect mix: Recent progress in adjuvant research. Nat Rev Microbiol (2007) 5:396–7. doi: 10.1038/nrmicro1681
368. Vinay TN, Park CS, Kim HY, Jung SJ. Toxicity and dose determination of quillaja saponin, aluminum hydroxide and squalene in olive flounder (Paralichthys olivaceus). Vet Immunol Immunopathol (2014) 158:73–85. doi: 10.1016/j.vetimm.2013.03.007
369. Wang P. Natural and synthetic saponins as vaccine adjuvants. Vaccines (2021) 9:222. doi: 10.3390/vaccines9030222
370. Wang Y, Wang X, Huang J, Li J. Adjuvant effect of quillaja saponaria saponin (QSS) on protective efficacy and IgM generation in turbot (Scophthalmus maximus) upon immersion vaccination. Int J Mol Sci (2016) 17:325. doi: 10.3390/ijms17030325
371. Lange MD, Abernathy J, Farmer BD, Beck BH. Use of an immersion adjuvant with a flavobacterium columnare recombinant protein vaccine in channel catfish. Fish Shellfish Immunol (2021) 117:136–9. doi: 10.1016/j.fsi.2021.07.025
372. Chang CJ, Sun B, Robertsen B. Adjuvant activity of fish type I interferon shown in a virus DNA vaccination model. Vaccine (2015) 33:2442–8. doi: 10.1016/j.vaccine.2015.03.093
373. Robertsen B, Chang CJ, Bratland L. IFN-adjuvanted DNA vaccine against infectious salmon anemia virus: Antibody kinetics and longevity of IFN expression. Fish Shellfish Immunol (2016) 54:328–32. doi: 10.1016/j.fsi.2016.04.027
374. Zhao Z, Li Y, Chen G, Zhang C, Wang GX, Zhu B. Protective immunity against infectious spleen and kidney necrosis virus induced by mannose modified subunit vaccine with carbon nanotubes in mandarin fish. Aquac Res (2022) 53:2175–84. doi: 10.1111/are.15736
375. Gong YM, Wei XF, Zhou GQ, Liu MZ, Li PF, Zhu B. Mannose functionalized biomimetic nanovaccine enhances immune responses against tilapia lake virus. Aquaculture (2022) 560:738535. doi: 10.1016/j.aquaculture.2022.738535
376. Bedekar MK, Kole S. Development of nano-conjugated DNA vaccine against edwardsiellosis disease in fish. Methods Mol Biol (2022) 2411:195–204. doi: 10.1007/978-1-0716-1888-2_11
377. Collado-Gonzalez M, Esteban MA. Chitosan-nanoparticles effects on mucosal immunity: A systematic review. Fish Shellfish Immunol (2022) 130:1–8. doi: 10.1016/j.fsi.2022.08.030
378. Lagos L, Tandberg JI, Becker MI, Winther-Larsen HC. Immunomodulatory properties of concholepas concholepas hemocyanin against francisellosis in a zebrafish model. Fish Shellfish Immunol (2017) 67:571–4. doi: 10.1016/j.fsi.2017.06.046
379. Wang Z, Mu D, Jiang Y, He J, Zhang Y, Yang D, et al. Dietary supplementation of propolis enhanced the innate immune response against edwardsiella piscicida challenge in turbot (Scophthalmus maximus). Fish Shellfish Immunol (2022) 124:273–9. doi: 10.1016/j.fsi.2022.03.016
380. Martinez-Lopez A, Garcia-Valtanen P, Ortega-Villaizan M, Chico V, Gomez-Casado E, Coll JM, et al. VHSV G glycoprotein major determinants implicated in triggering the host type I IFN antiviral response as DNA vaccine molecular adjuvants. Vaccine (2014) 32:6012–9. doi: 10.1016/j.vaccine.2014.07.111
381. Nguyen HT, Nguyen TTT, Wang YT, Wang PC, Chen SC. Effectiveness of formalin-killed vaccines containing CpG oligodeoxynucleotide 1668 adjuvants against vibrio harveyi in orange-spotted grouper. Fish Shellfish Immunol (2017) 68:124–31. doi: 10.1016/j.fsi.2017.07.018
382. Yan Y, Huo X, Ai T, Su J. β-glucan and anisodamine can enhance the immersion immune efficacy of inactivated cyprinid herpesvirus 2 vaccine in carassius auratus gibelio: Immersion immune adjuvants. Fish Shellfish Immunol (2020) 98:285-395. doi: 10.1016/j.fsi.2020.01.025
383. Alishahi M, Tollabi M, Ghorbanpoor M. Comparison of the adjuvant effect of propolis and freund on the efficacy of aeromonas hydrophila vaccine in common carp (Cyprinus carpio). Iran J Fish Sci (2019) 18:428–44. doi: 10.22092/ijfs.2019.118393
384. Ko EJ, Lee YT, Lee Y, Kim KH, Kang SM. Distinct effects of monophosphoryl lipid a, oligodeoxynucleotide CpG, and combination adjuvants on modulating innate and adaptive immune responses to influenza vaccination. Immune Netw (2017) 17:326–42. doi: 10.4110/in.2017.17.5.326
Keywords: aquaculture, fish disease, fish immunity, aquatic vaccines, inactivated vaccine, liveattenuated vaccine, genetic engineering vaccine
Citation: Du Y, Hu X, Miao L and Chen J (2022) Current status and development prospects of aquatic vaccines. Front. Immunol. 13:1040336. doi: 10.3389/fimmu.2022.1040336
Received: 09 September 2022; Accepted: 26 October 2022;
Published: 10 November 2022.
Edited by:
Erlong Wang, Northwest A & F University, ChinaReviewed by:
Maria José Ranzani-Paiva, University of Sao Paulo, BrazilXiaojun Zhang, Yangzhou University, China
Copyright © 2022 Du, Hu, Miao and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiong Chen, Y2hlbmppb25nQG5idS5lZHUuY24=
†These authors have contributed equally to this work