 Bin Yuan
Bin Yuan Xiao-jie Lu1,2*
Xiao-jie Lu1,2*
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 24 December 2021
Sec. Microbial Immunology
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.800796
This article is part of the Research Topic Current Trends in Exploiting Molecular Signaling in Bacteria-Host Crosstalk View all 30 articles
Acute central nervous system (CNS) injuries, including stroke, traumatic brain injury (TBI), and spinal cord injury (SCI), are the common causes of death or lifelong disabilities. Research into the role of the gut microbiota in modulating CNS function has been rapidly increasing in the past few decades, particularly in animal models. Growing preclinical and clinical evidence suggests that gut microbiota is involved in the modulation of multiple cellular and molecular mechanisms fundamental to the progression of acute CNS injury-induced pathophysiological processes. The altered composition of gut microbiota after acute CNS injury damages the equilibrium of the bidirectional gut-brain axis, aggravating secondary brain injury, cognitive impairments, and motor dysfunctions, which leads to poor prognosis by triggering pro-inflammatory responses in both peripheral circulation and CNS. This review summarizes the studies concerning gut microbiota and acute CNS injuries. Experimental models identify a bidirectional communication between the gut and CNS in post-injury gut dysbiosis, intestinal lymphatic tissue-mediated neuroinflammation, and bacterial-metabolite-associated neurotransmission. Additionally, fecal microbiota transplantation, probiotics, and prebiotics manipulating the gut microbiota can be used as effective therapeutic agents to alleviate secondary brain injury and facilitate functional outcomes. The role of gut microbiota in acute CNS injury would be an exciting frontier in clinical and experimental medicine.
Acute injuries to the central nervous system (CNS), such as stroke, traumatic brain injury (TBI), spinal cord injury (SCI), are critical global health problems that result in lifelong disabilities or death, leading to catastrophic changes to the injured individuals, alongside their family and even the entire community (1, 2). The processes secondary to acute CNS injuries involve a sequence of complex pathophysiological mechanisms, including excitotoxicity, electrolyte imbalance, oxidative stress, inflammation, apoptosis, pyroptosis, ferroptosis, autophagy, and cerebral edema (3). These cellular and molecular damages exacerbate neuronal cell death. Although some preclinical researchers have made many efforts to develop efficacious treatment strategies, patients with severe acute CNS injuries still have a poor prognosis. Given the prevalence of acute CNS injury-induced disabilities or death, exploring a novel and effective therapeutic regimen is imperative. In addition, a growing body of studies has shown that gut microbiota plays a pivotal role in health and disease in the host, particularly in the CNS (4–7).
Gut microbiota refers to the assemblage of bacteria, archaea, viruses, and eukaryotic microbes that colonize in the digestive tract (8). The gut microbiota contains trillions of microorganisms, over 1000 different species of known bacteria, and approximately 100 ~ 150-fold more genes than the human genome (9). At the phylum level, the gut microbiota primarily consists of Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, and Verrucomicrobia (10). Thereinto, Firmicutes, and Bacteroidetes comprise about 90% of all the bacteria (11). Additionally, the composition of gut microbiota among individuals was influenced by diet, age, gender, environment, and genes (12). Although the microbiome is spatially restricted to the gut, it has been shown to regulate the functions of distant organs (13). Notably, advances in the sequencing of gut microbiota have revealed the close correlation between the complex ecosystem and CNS (14). Previous studies focused on exploring the bidirectional communication pathways between gut microbiota and CNS, termed the “microbiota-gut-brain” axis (MGBA) (15). The bidirectional communication pathways between the CNS and gut microbiota involve immunological, endocrine, metabolic, and neural pathways (16, 17). Recent findings have implicated that MGBA partakes in the pathogenesis of many neurological disorders such as neurodegenerative diseases(e.g., Alzheimer’s disease and Parkinson’s disease), neurodevelopmental and neuropsychiatric diseases (e.g., anxiety, depression, autism, and schizophrenia), autoimmune disease (e.g., multiple sclerosis), and acute CNS injuries (e.g., stroke, TBI and SCI) (7, 15, 18–20). In this review, we provide an update on the link between gut microbiota and acute CNS injuries.
In top-down signaling, the pathways involve the autonomic nervous system, enteric nervous system, the hypothalamic-pituitary-adrenal (HPA) axis, and immunological pathway (Figure 1). The autonomic nervous system regulates intestinal homeostasis, and the neurotransmitters, released by the activated sympathetic and parasympathetic nerve fibers, modulate gut motility, gut barrier permeability, fluid maintenance, bile secretion, resident immune cell activation, and gut microbiota makeup (21). The enteric nervous system is also responsible for gut functions, such as gut motility and fluid maintenance, a neuronal connection between the microbiota and the host. Moreover, the HPA axis is one of the vital nonneuronal transmission pathways within the MGBA, releasing cortisol to influence gut homeostasis in response to various stimuli. With the discovery of meningeal lymphatic vessels, the brain is no longer an immune-privileged site. Functional meningeal lymphatic vessels lined mainly in the dorsal part of the skull are responsible for the clearance of cerebrospinal fluid and drainage of immune cells and the periphery, communicating with the host organs (22).
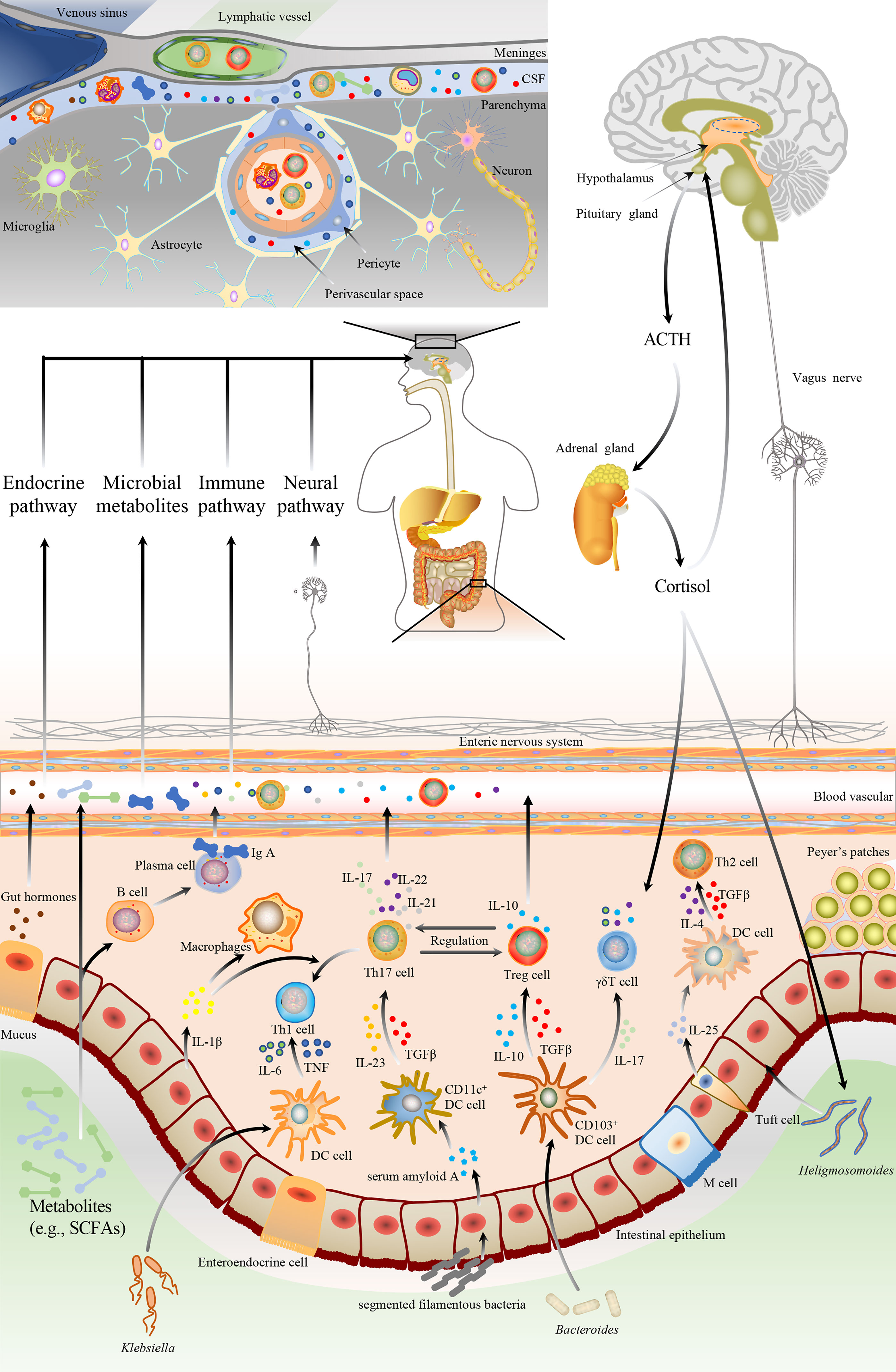
Figure 1 The bidirectional communication pathways between the gut microbiota and brain. The gut microbiota could bi-directionally communicate with the brain through multiple pathways, including neuronal and non-neuronal. The brain regulates the gut microbiota via neuronal pathways (e.g., autonomic nervous system and enteric nervous system), hypothalamic-pituitary-adrenal axis, etc. Neuronal pathways release neurotransmitters to modulate gut motility, gut barrier permeability, fluid maintenance, resident immune cell activation, and gut microbiota composition. HPA also releases cortisol to regulate gut homeostasis. Additionally, gut microbiota affects the development and pathophysiology of the brain by immunological, endocrine, metabolic, and neural pathways. Microbiomes and their metabolites could modulate the brain and behavior by affecting intestinal epithelial cells to alter gut barrier function, enteroendocrine cells to secret hormones, as well as dendritic cells and macrophage, to regulate immune and microglia activation. Gut microbiota can modulate the CD4+ T cells differentiation through epithelial cells or DC cells-mediated signals. ① Ectopic colonizing microbes, such as Klebsiella, can invade intestinal epithelium and stimulate DC cells to secrete proinflammatory cytokines, including IL-6 and TNF, which drive the polarization of Th1cells. ② SFB promotes Th17 polarization via epithelial cell-mediated CD11c+ DC cells activation. Epithelial cells release serum amyloid A to activate CD11c+ DC cells, leading to the TGF-β, IL-12, and IL-23 secretion. ③ Resident microbes, such as Bacteroides, modulate Treg cells generation by TGF-β and IL-10, which are secreted by CD103+ DC cells. CD103+ DC cells also can release IL-17 to promote γδT cell polarization. ④ Microbiota-associated Th2 cell polarization is correlated with parasite colonization such as Heligmosomoides, mediated by tuft cells secreting IL-25 to DC cells. Then activated DC cells release IL-4 and TGF-β to drive Th2 polarization. DC, dendritic cell; IL, interleukin; TNF, tumor necrosis factor; Th, T helper; TGF-β, transforming growth factor-β; SFB, segmented filamentous bacteria; SCFA, short-chain fatty acid; Treg, regulatory T cell.
Two different mechanisms involved in the bottom-up signaling are neuronal and nonneuronal pathways (Figure 1). The vagus nerve, composed of both afferent and efferent fibers (80% vs. 20%), plays a pivotal role in bidirectionally transmitting vital information between the gut and brain. The afferent fibers stimulated by microbial metabolites and enteroendocrine neuropeptides convey hypothalamic neurons that promote pituitary secretions. In addition, the interaction between the gut and brain primarily relies on the nonneuronal pathway. Singh et al. reported that the gut microbiota-mediated neuroprotection was absent in lymphocyte-deficient mice after an experimental stroke of permanent distal middle cerebral artery occlusion (MCAO), indicating that the gut communicate with the brain by immunological pathway (23). In a transient MCAO model, intestinal CD45+ and CD11c+ cells significantly migrated from the gut to the brain and meninges at 3 days post-stroke (24). A study of a thoracic level 9 contusion SCI also showed that commercial probiotics (VSL#3) feeding triggered a protective immune response in gut-associated lymphatic tissues (GALTs) and conferred neuroprotection with the improvement of locomotor recovery after SCI (25). TBI-induced leaky gut released lipopolysaccharide (LPS), a toxic bacterial component, into the circulation that exacerbated neuroinflammation by activating microglia (26).
The MGBA is critical to the development of the human central nervous system. A prospective longitudinal study conducted by Carlson et al. investigated the correlation between gut microbial composition and cognitive ability in 89 infants. It revealed that 2-year-old cognitive function assessed with the Early Learning Composite of Mullen Scale was significantly correlated with the gut microbiota composition at one year (27). Infants with a relatively high abundance of Bacteroides showed higher performance, while those with a relatively high quantity of Faecalibacterium had a lower performance. In another clinical study of 39 infants, the α-diversity of gut microbiota was associated with functional connectivity between the amygdala and thalamus, between the anterior cingulate cortex and anterior insula, and between the supplementary motor area and the inferior parietal lobule (28). Additionally, the functional connectivity was also related to 2-year-old cognitive outcomes. These studies have demonstrated that the gut microbiota significantly affects neurodevelopment in the early stage through the MGBA.
Microbial components and metabolites such as lipopolysaccharide (LPS), long-chain fatty acids (LCFAs), short-chain fatty acids (SCFAs), trimethylamine-N-oxide (TAMO), tryptophan, and polysaccharide A (PSA) are considered to induce neuroinflammation and modulate the function of CNS either directly or by activating migration of peripheral immune cells to the brain. Although this regulation of immune cells by the microbiota occurs in the gut, peripheral immune cells could also migrate to the brain meninges and modulate the brain function (29).
Previous studies have shown that the gut microbiota-host interaction contributes to the maturation and modulation of the host immune system. Through constant contact with the gut microbiota, immune cells and epithelial cells located in the gut have achieved a homeostatic state and promote tolerogenic responses to the host commensal microbes. Under physiological conditions, all types of immune cells such as T lymphocytes, B lymphocytes, macrophages, dendritic cells (DCs), etc., counterbalance each other to preserve the host homeostasis. T helper (Th) cells and regulatory T (Treg) cells are a requisite component of the host immune system, especially in the gut-associated immune responses. Compared with germ-free (GF) mice, conventional specific-pathogen-free (SPF) mice had a more significant proportion of CD4+ T cells (30). Th1 cells were polarized by DCs-secreted pro-inflammatory cytokines such as interleukin(IL)-6 and IL-12 stimulated by Klebsiella (31). Parasites, such as Heligmosomoides, could activate Th2 cells through DCs-derived transforming growth factor-β(TGF-β) and IL-4 (32). Besides, segmented filamentous bacteria drive Th17 polarization via activation of CD11c+ DCs (33). The activated Th17 cells secrete high-affinity IL-17 and promote IgA transportation, memory CD4+ T cell differentiation, and mucin production (33). Bacteroides fragilis stimulate regulatory CD4+ T cells to make themselves colonize the intestinal epithelium and induce immunosuppression, mediated by CD103+ DCs (34). γδ T cells, another innate immune cell population in the gut epithelium, are vital for gut homeostasis regulation and pathological reaction. CD103+ DCs activated by gut microbiota also communicate with IL-17+ γδ T cells via cell-to-cell contact or different cytokines (35). γδ T cells protect the host from intestinal barrier damage and pathogenic bacterial invasion and exert pro-inflammatory or anti-inflammatory effects depending on the milieu (36). Additionally, the development of intestinal mucosa B cells is regulated by commensal microbes, which promote antibody production and control the expression of a differentiation-related gene through enhancement of fatty acid synthesis, glycolysis, and oxidative phosphorylation (37). Commensal microbiota-induced secretion of IL-1β by mucosal macrophages is closely correlated with the development of steady-state Th17 cells (38)(Figure 1).
Gut microbiota-derived small molecules are inextricably linked with the crosstalk between gut microbiota and host. Moreover, there is now a considerable body of experimental evidence that some metabolites of the intestinal microbiota can participate in the modulation of inflammatory cytokine production and immune cell differentiation. SCFAs produced by gut microbiota such as Faecalibacterium prausnitzii, Roseburia intestinalis, and Anaerostipes butyraticus regulate the activation and differentiation of immune cells (e.g., neutrophils, macrophages, DCs, and T cells), mediated by inhibiting histone deacetylases (HDACs) as well as activating G-protein-coupled receptors (GPCRs) (e.g., GPR41, GPR43, and GPR109A) and olfactory receptor-78 (Olfr-78) (39–41). TMAO is another gut microbiota metabolite oxidized from trimethylamine (TMA) generated by hepatic flavin-containing monooxygenases (FMOs) from dietary nutrients such as choline and L-carnitine. TAMO regulates pro-inflammatory responses via activating the NOD-like receptor family, pyrin domain-containing protein 3 (NLRP3) inflammasome, mitogen-activated protein kinase (MAPK), and nuclear factor-κB (NF-κB) signaling pathway (42). Additionally, tryptophan has also been demonstrated to be involved in the functional modulation of intestinal intraepithelial lymphocytes and innate lymphocytes via activating the aryl hydrocarbon receptor (AHR) (43). PSA, produced by Bacteroides fragilis, activates toll-like receptor 2(TLR2) expressed on DCs and Treg cells, triggering anti-inflammatory cytokine IL-10 (43). PSA also regulates the differentiation of naive CD4+ T cells into Th1 cells, skewing Th1/Th2 ratio towards Th1 cells (44). Interestingly, a study found that CD39 expression is required for Treg cells to migrate to the CNS, depending on PSA-driven effects on the Treg cells (45).
Stroke is the second leading cause of death worldwide. Morbidity and mortality of stroke grow in many countries, contributing to financial burden and loss of life quality, and thus diminishing national happiness index. Approximately 15 million people around the world are victims of a stroke every year (46). There are two types of strokes: ischemic stroke and hemorrhagic stroke. Recent evidence shows that shifts in the gut microbial composition occur rapidly after stroke in clinical and experimental studies. Therefore, alteration of the gut microbiota directly regulates the reactions secondary to brain injury through central and peripheral immunity and indirectly determines the brain’s sequel to these types of catastrophic events.
Reportedly, ischemic stroke accounts for ~80% of all strokes (47), and the gut microbiota plays an essential role in the pathogenesis and prognosis of ischemic stroke. Multiple studies have shown that ischemic stroke significantly changes the gut microbiota composition (21, 23, 48–66). These studies have been summarized in Table 1. Although dysbiosis of the gut microbiota has been proved in previous studies, controversy still exists on the specific microbiota difference. Compelling evidence has been identified that confounding factors such as age, diet, behavior, antibiotic use, prolonged stress, environment, and genetics compose the gut microbiota, which may be influenced by the contradictory results of the above studies. Thus, more studies are needed to clarify the role of gut microbiota dysbiosis in the pathogenesis and prognosis of ischemic stroke. Recently, a preclinical study also suggested that the alteration in the gut microbiota was associated with hemorrhagic transformation (HT) (71). The relative abundance of Proteobacteria and Actinobacteria was significantly increased in HT rats after experimental stroke, indicating that the gut microbiota is involved in the progression of ischemic stroke.
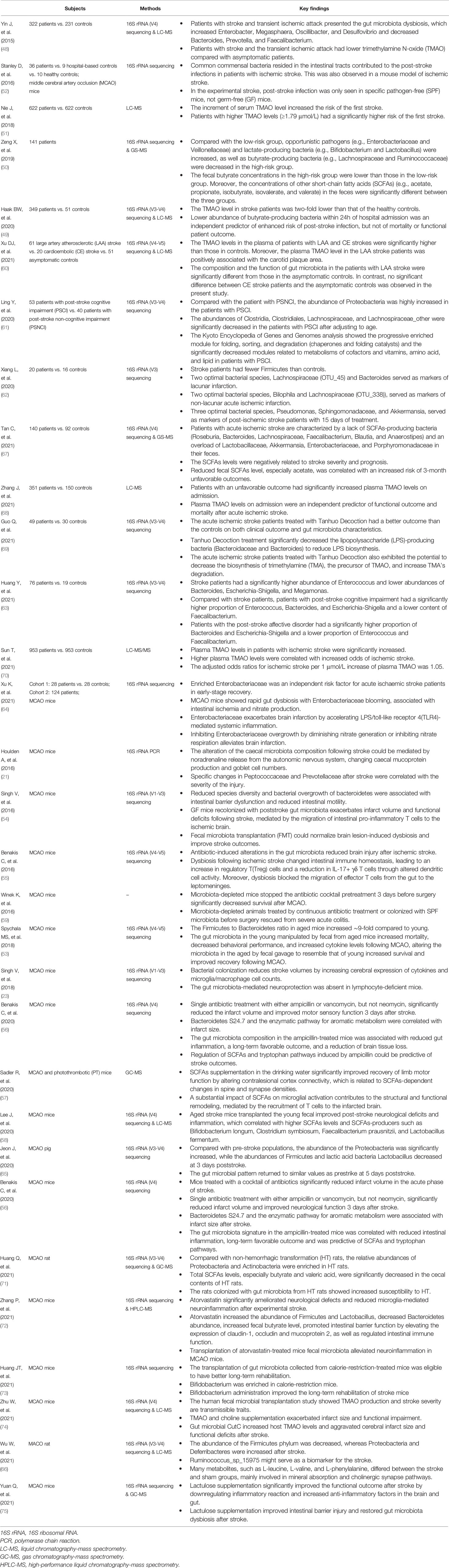
Table 1 A summary of preclinical and human studies on the gut microbiota and ischemic stroke.
Mechanistically, the gut microbiota-mediated neuro-protection greatly depended on the microglia and lymphocyte responses, significantly increasing Th cells, polarized Treg cells, and Th17 cell counts in the intestinal Peyer’s patches (23). Pro-inflammatory Th1, Th17, and γδT cells often increase inflammatory damage, while Treg cells suppress post-stroke inflammation by secreting the anti‐inflammatory cytokine IL‐10. Alteration of gut microbiota following a stroke in the bacterial population triggers pro-inflammatory T cells responses, migrates intestinal immune cells to the meninges involved in secondary brain injury, and worsens stroke outcome. In GF MCAO animal models, mice transplanted with post-stroke fecal content presented increased infarct volume and functional deficits by inducing pro-inflammatory T cell polarization. Moreover, restoration of gut microbiota homeostasis with fecal microbiota transplantation (FMT) reduced infarct volume, improved stroke outcome and promoted the migration of intestinal Treg cells to the ischemic area in the brain (54). Also, intestinal dysbiosis following ischemic stroke was found to regulate immune homeostasis in the small intestine with increased Treg cells and decreased IL-17+ γδT cells, mediated by DCs. The neuroprotective effect of IL-10 was identified as a regulator of Treg cell-mediated IL-17+ γδT cell suppression (55).
Microbial-derived metabolites also correlate with the progression and prognosis of ischemic stroke. Ischemic stroke patients had significant gut microbiota dysbiosis with an increased abundance of SCFAs-producing bacteria such as Odoribacter, Akkermansia, which closely correlated with the stroke outcome (76). However, Zeng et al. reported that people with a high risk of stroke had lower levels of butyrate-producing bacteria (e.g., Lachnospiraceae and Ruminococcaceae) and fecal butyrate (50). Tan et al. also reported a lack of SCFAs-producing bacteria and a low fecal SCFAs level in acute ischemic stroke patients (67). Moreover, the reduced fecal SCFAs were correlated with an increased risk of 3-month unfavorable outcomes (67). The differences in the results of these clinical studies may be due to the small cohorts of the studies. These findings need to be further validated by higher-quality clinical studies with large cohorts. In experimental stroke, reduced plasma SCFAs level correlated with a worse stroke outcome in mice, and SCFAs supplementation improved behavioral recovery with modified poststroke cortical connectivity and synaptic plasticity by recruiting T lymphocytes on modulation of microglial activation, as reflected by the increase in Treg cells (57). Oral gavage of SCFAs-producing bacteria such as Bifidobacterium longum, Clostridium symbiosum, Faecalibacterium prausnitzii, and Lactobacillus fermentum alleviated post-stroke neurological deficits and inflammation by increasing populations of Treg cells and reducing the percentage of IL-17+ γδT cells (58). Pretreatment with Clostridium butyricum improved neurological deficits and decreased hippocampal apoptosis by increasing butyrate and reducing brain oxidative stress in experimental stroke (77). Furthermore, Zhou et al. reported that butyrate alleviated neuronal apoptosis following stroke via GPR41/Gβγ/PI3K/Akt pathway (78). SCFAs could also improve outcomes by protecting gut epithelial cells against stroke-induced gut leakiness by enhancing tight junction proteins (79). Furthermore, sodium butyrate reduced infarct volume and improved neurological function Recently, Huang et al. found that the significant decrease of SCFAs in cecal contents, especially butyrate and valeric acid, was closely related hemorrhagic transformation after ischemic stroke (71).
It has been demonstrated that there is a significant association between TMAO level and various diseases, including stroke (80). Although several clinical studies have identified a correlation between TMAO level and stroke, the results remain controversial. Most studies show that the plasma TMAO concentrations in stroke patients are significantly higher than those in control patients, and its high level is positively related to the severity of the stroke (51, 68). A large-scale case-control study with 953 sex- and age-matched pairs performed by Sun et al. suggested that the plasma TMAO concentrations in patients with first acute ischemic stroke were significantly elevated (70). Furthermore, further analysis revealed that the multivariable-adjusted odds ratios for ischemic stroke per 1 μmol/L increase of plasma TMAO level were 1.05. Tan et al. reported that TMAO concentrations decreased with time after stroke, and elevated TMAO levels at an early stage predicted poor stroke outcomes (81). A meta-analysis also showed that compared with non-stroke controls, TMAO increased the stroke risks by 68% and accreted 2.201 umol/L on the average level of TMAO in stroke patients (82). Zhu et al.’s study also suggested gut microbiota can impact stroke severity via the gut microbial CutC-mediated TMAO pathway, which exacerbated cerebral infarct size and functional deficits (74).
The activation of the kynurenine pathway for tryptophan degradation correlates with stroke-induced inflammatory responses and unfavorable outcomes53. Tryptophan catabolites regulate intestinal immune cell function by activating AHR. Pharmacological and genetic inhibition of neural cell-specific AHR activation improved stroke outcomes in the MCAO mice model (83). Furthermore, tryptophan catabolism positively correlated with the severity of stroke outcome and might be associated with stroke-induced inflammatory response (51). Besides, xenobiotic/aromatic compound metabolism was a predictive marker of the size of the ischemic lesion (56).
Multiple studies have demonstrated that antibiotic-induced dysbiosis promotes the proliferation and differentiation of T cells in the gut to either improve or worsen outcomes in experimental stroke. Mice pretreated with ampicillin or vancomycin significantly improved the outcome of stroke, whose neuroprotection is related to a shift with increased Proteobacteria and Firmicutes and reduction of Bacteroidetes caused by antibiotics (56). Particularly, Bacteroidetes S24.7 was closely associated with infarct size. However, Winek et al. reported stroke mice pretreated with quintuple broad-spectrum antibiotics presented with the damaged gut epithelium and worsened outcome (59). This controversial result still needs further study to clarify. FMT is a novel and potent treatment strategy in patients with gut microbiota dysbiosis obtained from fecal microbiota in healthy individuals. FMT exerts a neuroprotective effect by altering gut microbial metabolites production and reducing pro-inflammatory gut bacteria, alleviating inflammatory response and oxidative stress in the brain. Restoring gut microbiota homeostasis with FMT from healthy donors reduced lesion size by increasing Treg cells (54). MCAO mice receiving FMT from anti-inflammatory donors reduced the infarct volume by 54% (55). Additionally, FMT from young microbiota was also beneficial to stroke recovery (53). Oral gavage of SCFAs-producing bacteria or SCFAs supplementation also alleviated neurological deficits and improved poststroke recovery by reducing IL-17+ γδT cells in the ischemic brain (57, 58). Recently, Zhang et al. found that atorvastatin significantly alleviated the defects in sensorimotor behaviors and reduced microglia-mediated neuroinflammation by increasing the abundance of Firmicutes and Lactobacillus, decreasing the abundance of Bacteroidetes abundance, increasing fecal butyrate level, promoting intestinal barrier function, as well as regulating intestinal immune function (reduced monocyte chemotactic protein 1(MCP-1), tumor necrosis factor-α(TNF-α) and increased IL-10) in the mice with permanent MCAO (72). Calorie restriction also can promote ischemic stroke rehabilitation via enriching the abundance of Bifidobacterium (73). Tanhuo decoction also promoted poststroke recovery by decreasing the biosynthesis of TMA, the precursor of TMAO, and increasing the expression of trimethylamine-corrinoid protein Co-methyltransferase (mttB), which catabolizes TMA to methane (69). Additionally, Lactulose supplementation was shown to significantly improve the functional outcome of stroke, which is possibly mediated by repairing intestinal barrier injury and improving gut microbiota dysbiosis following stroke (75).
Hemorrhagic stroke includes intracranial and subarachnoid hemorrhage. Intracranial hemorrhage accounts for 80% of hemorrhagic stroke and 10-15% of all strokes, which is primarily caused by hypertension-induced small vessel rupture, while subarachnoid bleeding is mainly due to intracranial aneurysms rupture (84). Hemorrhagic stroke is characterized by high mortality and morbidity, which burdens society and families. However, there are, to date, few studies focused on the exploration of the correlation between hemorrhagic stroke and gut microbiota. A few studies reported that gut microbiota dysbiosis contributes to hypertension and intracranial aneurysms. However, the direct relationship between gut microbiota and hemorrhagic stroke has not been studied. Both clinical and animal studies are warranted in the future.
A case-control metagenome-wide association study showed that the structural heterogeneity of intestinal microbiota in patients with intracranial aneurysm (IA) was significantly decreased compared to healthy controls, which had an increased abundance of Bacteroides, Parabacteroides, Ruminococcus, and Blautia in IA patients and an enrichment of Faecalibacterium, Eubacterium, Collinsella, and Lactobacillus (85). Recently, another multicenter, prospective case-control study reported that the abundance of the genus Campylobacter and Campylobacter ureolyticus was significantly higher in patients with ruptured IA than that in patients with unruptured IA, which may be associated with the rupture of IA (86). Further analysis suggested that gut microbiota promoted the pathogenesis of IA by regulating plasma amino acids (e.g., taurine, hypotaurine, L-histidine, and L-citrulline) and fatty acid levels (85). Compared to mice transplanted with healthy control feces, the incidence of IA in mice transplanted with the feces of IA patients was significantly increased (85). Mechanistically, supplementation with taurine or H.hathewayi reduces the formation and rupture of IA by blunting cerebrovascular inflammatory processes, reducing extracellular matrix remodeling, and maintaining the structural integrity of cerebral blood vessels. Similarly, Fumiaki Shikata et al. also reported that the gut microbiota contributes to the development of IA by modulating inflammation in the experimental IA model (87). Gut microbiota depletion by an oral antibiotic cocktail of ampicillin, metronidazole, neomycin, and vancomycin (AMNV) significantly reduced the incidence of IA via decreasing macrophage infiltration and the expression of pro-inflammatory cytokines such as IL-1β and IL-6 in vascular wells (87). These results suggest that gut microbiota is closely correlated with the development of IA. Additionally, human studies are needed to determine the exact contribution of the gut microbiota to the pathophysiology of IA and aneurysmal subarachnoid hemorrhage in humans.
Traumatic brain injury (TBI) is one of the most common neurological diseases, with an estimated incidence of approximately 50 million people worldwide annually, leading to thousands of deaths and disabilities (88). TBI induces various secondary progressive brain damage contributing to varied functional outcomes. TBI also influences the gut barrier integrity, gut function, and gut microbiota composition (89). In turn, gut microbiota alterations may regulate a pro-inflammatory response following TBI and aggravate secondary brain injury. However, the information on TBI-induced gut microbiota dysbiosis is scarce for now. The relevant studies are summarized in Table 2.
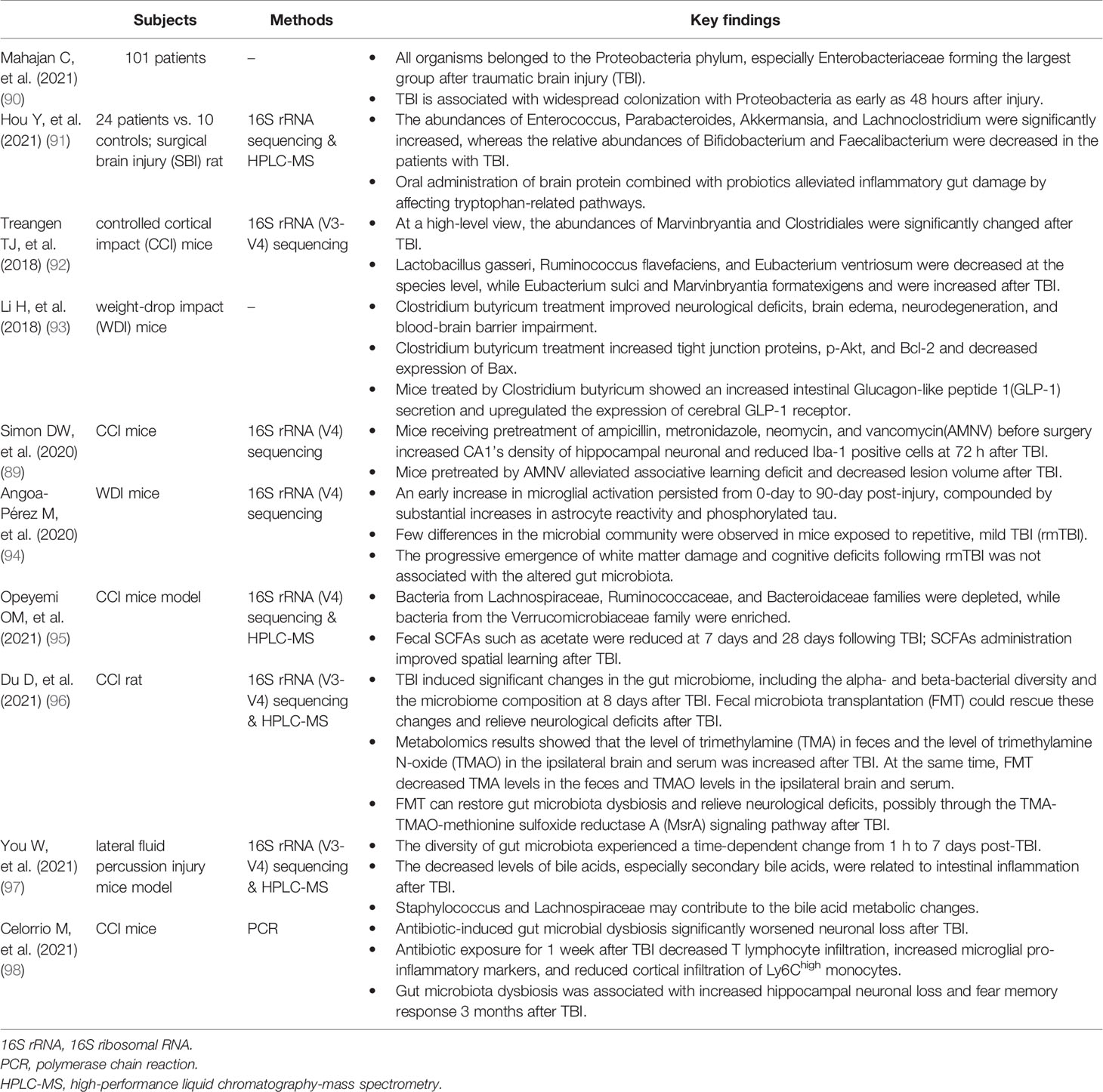
Table 2 A summary of preclinical and human studies on the gut microbiota and traumatic brain injury.
Recently, an observational study investigated the characteristics of gut microbiota in 101 TBI patients and found that organisms from rectal swabs obtained on days 0, 3, and 7 after admission belonged to the Proteobacteria phylum, with Enterobacteriaceae forming the largest group (90). Hou et al. also analyzed the gut microbiota composition in a small cohort (10 healthy control volunteers vs. 24 TBI patients) and reported that the abundance of Enterococcus, Parabacteroides, Akkermansia, and Lachnoclostridium were significantly increased, while the abundance of Bifidobacterium and Faecalibacterium were decreased in TBI patients (91).
In the controlled cortical impact (CCI) mouse model, the gut microbiota significantly decreased in Lactobacillus gasseri, Ruminococcus flavefaciens, and Eubacterium ventriosum and increased dramatically in Eubacterium sulci and Marvinbryantia formatexigens at 24h post-CCI (92). In an experimental weight-drop injury model, the severity of TBI is correlated with the alteration in Bacteroidetes, Porphyromonadaceae, Firmicutes, and α-Proteobacteria (21). Nicholson et al. found a reduced Firmicutes/Bacteroidetes ratio in gut microbiota composition occurring at 2h post-injury was significantly related to MRI-determined lesion volume and behavioral function defects (99). In the lateral fluid percussion injury mice model, You et al. also observed the alterations of gut microbiota and bile acid profile (97). Further analysis found that specific bacterial taxa such as Staphylococcus and Lachnospiraceae could be associated with the bile acid metabolic changes, resulting in intestinal inflammation. Interestingly, Angoa-Pérez et al. found that repetitive, mild TBI did not cause alterations in the gut microbiota composition (94). Although differences in gut microbiota composition have been observed after TBI in animal models, the exact regulatory mechanism remains elusive. A study considered that vagal afferent alterations, TBI-induced increase of cholecystokinin level, might be responsible for gut dysfunction through activation of the vago-vagal NTS-inhibitory pathway (100). Additionally, in another experimental TBI, the gut upregulated the expression of glycoproteins to recruit immune cells and activate inflammatory signals, resulting in altered mucosal integrity (101). The leaky gut allowed toxic bacterial components such as LPS to enter the circulation that mediates neuroinflammation by activating microglia following TBI (26). Furthermore, the permeability of the blood-brain barrier (BBB) can increase up to 4 times more than normal within 6h following TBI. The increased BBB permeability aggravates the gut dysbiosis-induced neuroinflammation by LPS exposure, γδT cell activation, and activated microglia differentiation into the M1 phenotype. A recent preclinical study performed by Celorrio et al. suggested that antibiotic-induced gut microbial dysbiosis established before TBI significantly worsened neuronal loss, reduced cortical infiltration of Ly6Chigh monocytes and T lymphocyte, increased microglial pro-inflammatory markers, and impaired neurogenesis after TBI (98).
CCI mice pretreated with AMNV 2 weeks before CCI presented increased neuronal density in the hippocampus at 72h post-injury, while mice treated with AMNV right after CCI showed reduced lesion volume and attenuated associative learning deficit at 22 days (89). TBI mice treated by Clostridium butyricum also improved neurological deficits, attenuates brain edema, ameliorated neurodegeneration, and alleviated BBB impairment via elevating intestinal Glucagon-like peptide 1(GLP-1) secretion (93). SCFAs supplementation also improved spatial learning following CCI-induced TBI, mediated by activating the neurotrophic tyrosine kinase receptor type 1 (TrkA) pathway (95, 102). Probiotic supplementation also significantly remedied the gut microbiota dysbiosis and decreased the intestinal permeability following experimental TBI by reducing the intestinal mucosa damage, alleviating brain injury (103). In human studies, probiotic treatment could relieve systemic inflammatory response, decrease nosocomial infection rate, and improve the recovery of patients with TBI (104, 105). Interestingly, vagal stimulation reduced gut barrier permeability after TBI, mediated by the suppression of TNF-α release (106). Recently, it has been demonstrated that FMT can restore gut microbiota dysbiosis following TBI and ameliorate neurological deficits, mediated by the TMA-TMAO-MsrA signaling pathway (96).
Traumatic spinal cord injury (SCI) is another acute CNS injury that affects millions worldwide every year (107). The studies involving SCI and the bidirectional effect on the gut microbiota have been carried out in recent years, which are summarized in Table 3. This section reviews the study published on the changes in the gut microbiota that occur following SCI.
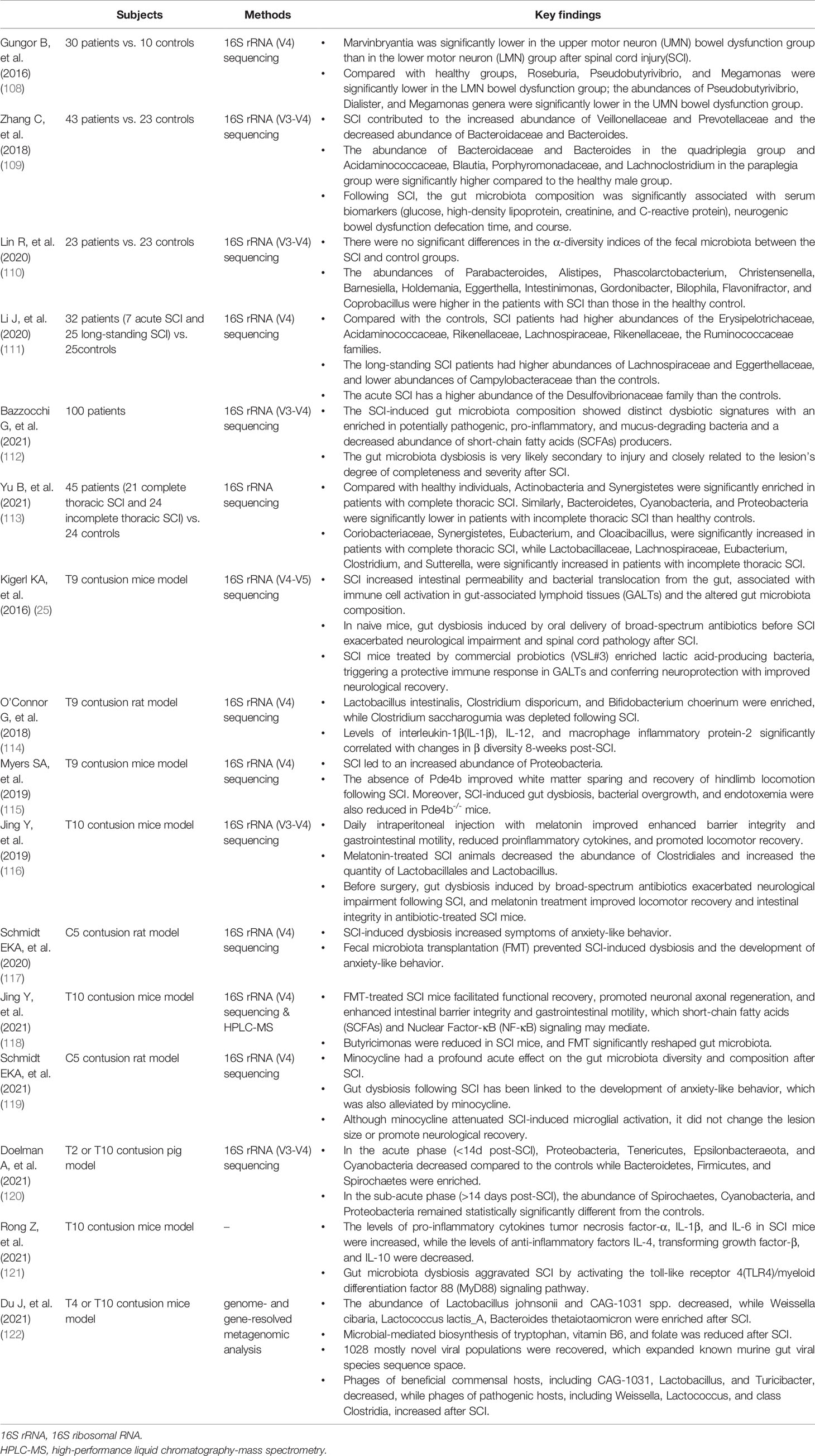
Table 3 A summary of preclinical and human studies on the gut microbiota and spinal cord injury.
In a Chinese cohort study, Zhang et al. observed an increase in Proteobacteria and Verrucomicrobia and reduced Bacteroidaceae and Bacteroides in patients with chronic traumatic complete SCI (109). Lin et al. also analyzed 46 Chinese subjects (23 SCI patients vs. 23 healthy controls) and reported that the abundances of Parabacteroides, Alistipes, Phascolarctobacterium, Christensenella, Barnesiella, Holdemania, Eggerthella, Intestinimonas, Gordonibacter, Bilophila, Flavonifractor, and Coprobacillus were higher in the patients with SCI than those in the health individuals (110). Another clinical study with 54 Turkish participants (41 SCI patients vs. 13 healthy controls) identified that butyrate-producing microbes of the Firmicutes phylum are significantly reduced in SCI patients than healthy controls (108). Recently, Bazzocchi et al. investigated a large Italian SCI population acute phase after injury and age- and gender-matched healthy Italians (112). Their study revealed that the abundance of SCI patients’ gut microbiota increased in potentially pathogenic, pro-inflammatory, and mucus-degrading bacteria and decreased in SCFAs producers. Moreover, gut microbiome dysbiosis is closely associated with the severity of the lesion after SCI. A case-control study carried by Yu et al. (45 SCI patients vs. 24 healthy individuals) showed that the abundance of Actinobacteria and Synergistetes in patients with complete thoracic SCI (CTSCI) was significantly higher than that in healthy individuals. At the same time, the Bacteroidetes, Cyanobacteria, and Proteobacteria were significantly decreased in patients with incomplete thoracic SCI (ITSCI) as compared to the healthy (113). Furthermore, they compared the gut microbiota composition between patients with CTSCI and ITSCI and found a significantly increased abundance of Coriobacteriaceae, Synergistetes, Eubacterium, and Cloacibacillus was observed in patients with CTSCI, while patients with ITSCI were abundant with Lactobacillaceae, Lachnospiraceae, Eubacterium, Clostridium, and Sutterella.
Similarly, a thoracic level 9 (T9) contusion SCI-induced gut microbiota dysbiosis in the experimental SCI mice was also characterized by an expansion of Bacteroidetes and a reduction of Firmicutes (115). However, a preclinical work in a T9 contusion SCI mouse model by Kigerl et al. reported that SCI mice presented a decrease in Bacteroidales and an increase in Clostridiales (25). In an SCI rat model, gut microbiota composition was significantly changed with an increased abundance of Lactobacillus intestinalis, Clostridium disporicum, and Bifidobacterium choerinum and a reduced level of Clostridium saccharogumia (114). The difference in the results may be caused by experimental deviation. Additionally, the above analyses of SCI-induced gut microbiota dysbiosis were assessed by 16S rRNA amplicon sequencing, which cannot profile microbiota function or identify viruses (123). Du et al. studied gut microbiota dysbiosis after experimental SCI at T4 or T10 using genome- and gene-resolved metagenomic analysis (122). The results suggested that the abundance of beneficial commensals (Lactobacillus johnsonii and CAG-1031 spp.) significantly decreased, while potentially pathogenic bacteria (Weissella cibaria, Lactococcus lactis_A, Bacteroides thetaiotaomicron) increased after SCI. Functionally, tryptophan, vitamin B6, and folate biosynthesis, encoded by microbial genes, were reduced in the feces after SCI. Interestingly, the study performed by Du et al. reported that phages of beneficial commensal hosts (CAG-1031, Lactobacillus, and Turicibacter) decreased. In contrast, phages of pathogenic hosts (Weissella, Lactococcus, and class Clostridia) increased after SCI (122). In a Yucatan minipig model with a contusion-compression SCI at T2 or T10, Doelman et al. presented a dynamic view of the microbiome changes following SCI and identified acute stage, 0-14 post-SCI, as a special time-frame that many of the bacterial fluctuations occur before returning to “baseline” levels (120).
SCI promotes intestinal leakiness and bacterial translocation associated with activation of immune cells in GALTs, by increasing the population of B cells, CD8+ T cells, DCs, and macrophages and decreasing CD4+ T cell counts (25). γδT cell-deficient mice improved functional recovery after SCI (124). Moreover, changes in gut microbiota composition following SCI could predict locomotor impairment (125). Additionally, gut microbiota dysbiosis can aggravate SCI by activating the TLR4/Myeloid differentiation factor 88 signaling pathway (121).
SCI mice fed with commercial probiotics (VSL#3) reduced neuropathology, improved locomotor recovery, and promoted an anti-inflammatory response by increasing the number of Treg cells in GALTs (25). Additionally, SCI mice daily treated with melatonin improved gut barrier integrity and functional recovery by reducing the abundance of Clostridiales and enhancing the quantity of Lactobacillales and Lactobacillus, which were related to a more favorable cytokine profile (116). Lactic acid supplementation was also proved to improve functional recovery following SCI (25). FMT prevented both SCI-induced dysbiosis, locomotor function, and the development of anxiety-like behavior (117). FMT could increase the amount of fecal SCFAs and downregulate IL-1β/NF-κB signaling in the spinal cord and NF-κB signaling in the gut following SCI (118). A recent study also reported that minocycline treatment attenuated SCI-induced anxiety-like behavior and systemic inflammatory response via altering the Firmicutes/Bacteroidetes ratio (119). Engineered liposomes targeting the MGBA may also be a potential treatment (126).
Gut microbiota is closely involved in the development and progression of acute CNS disease through multiple mechanisms, including immunological, endocrine, metabolic, and neural pathways. FMT and probiotics significantly improve brain injury by restoring the acute CNS injury-induced gut microbiota dysbiosis. Gut microbiota may be a potential target to assist in the treatment of acute CNS injury. However, several aspects are still needed to ponder despite a growing number of studies concerning the gut microbiota. Firstly, human gut microbiota composition is different from rodents. Although Firmicutes and Bacteroidetes are the most abundant microbiota both in mice and humans, more than 80% of the bacteria found in the mice intestine are not colonized in the human intestines based on genus level (127). Secondly, immunological features are also different between rodents and humans. A previous study suggests that the intestinal properties of humans are similar to those of mice. However, differences in intestinal immunity between mice and humans have already been found that γδT cells are found significantly less frequently in the intraepithelial compartment of humans than in mice (128). Thirdly, the effects of enteroviruses, fungi, and bacteriophages cannot be ignored. Bacteriophages have high host specificity that shapes the gut microbiota composition and regulates the host immune response by altering bacterial pathogen-associated molecular patterns and maintaining the host mucosal barrier (129). Although the effects of enteroviruses on health and disease are still unclear, phage-virus-fungi-bacterial-host interaction in the gut should also be considered. Moreover, their role in human acute brain injury or animal models has not been studied so far. Fourthly, developmental disturbances in GF mice should be considered. GF mice have hypoplastic immune structures and differ from SPF mice in the intestinal immune cell populations, such as IgA-producing plasma cells and lamina propria CD4+ T cells (130). Additionally, GF mice contain fewer serum immunoglobulins, particularly IgG (131). In the absence of gut microbiota, CNS is also altered with a “leaky” BBB and an abnormal microglia morphology and function (132, 133). Finally, criteria for identifying qualified, healthy donors in the FMT treatment have not yet been fully established. The safety and efficiency of FMT need to be extensively investigated.
BY wrote the manuscript. X-jL and QW revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the Scientific Research Project of Jiangsu Provincial Health commission under Grant No. H2018066.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sun P, Liu DZ, Jickling GC, Sharp FR, Yin KJ. Microrna-Based Therapeutics in Central Nervous System Injuries. J Cereb Blood Flow Metab (2018) 38(7):1125–48. doi: 10.1177/0271678x18773871
2. Wang Y, Tan H, Hui X. Biomaterial Scaffolds in Regenerative Therapy of the Central Nervous System. BioMed Res Int (2018) 2018(2):7848901–19. doi: 10.1155/2018/7848901
3. Sorby-Adams AJ, Marcoionni AM, Dempsey ER, Woenig JA, Turner RJ. The Role of Neurogenic Inflammation in Blood-Brain Barrier Disruption and Development of Cerebral Oedema Following Acute Central Nervous System (CNS) Injury. IJMS (2017) 18(8):1788. doi: 10.3390/ijms18081788
4. Zmora N, Suez J, Elinav E. You are What You Eat: Diet, Health and the Gut Microbiota. Nat Rev Gastroenterol Hepatol (2019) 16(1):35–56. doi: 10.1038/s41575-018-0061-2
5. O’Toole PW, Jeffery IB. Gut Microbiota and Aging. Sci (New York NY) (2015) 350(6265):1214–5. doi: 10.1126/science.aac8469
6. Sánchez B, Delgado S, Blanco-Míguez A, Lourenço A, Gueimonde M, Margolles A. Probiotics, Gut Microbiota, and Their Influence on Host Health and Disease. Mol Nutr Food Res (2017) 61(1):1600240. doi: 10.1002/mnfr.201600240
7. Järbrink-Sehgal E, Andreasson A. The Gut Microbiota and Mental Health in Adults. Curr Opin Neurobiol (2020) 62:102–14. doi: 10.1016/j.conb.2020.01.016
8. Sender R, Fuchs S, Milo R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PloS Biol (2016) 14(8):e1002533. doi: 10.1371/journal.pbio.1002533
9. Flowers SA, Ellingrod VL. The Microbiome in Mental Health: Potential Contribution of Gut Microbiota in Disease and Pharmacotherapy Management. Pharmacotherapy (2015) 35(10):910–6. doi: 10.1002/phar.1640
10. Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the Human Gut Microbiome. Nature (2011) 473(7346):174–80. doi: 10.1038/nature09944
11. Tyler Patterson T, Grandhi R. Gut Microbiota and Neurologic Diseases and Injuries. Adv Exp Med Biol (2020) 1238:73–91. doi: 10.1007/978-981-15-2385-4_6
12. Takagi T, Naito Y, Inoue R, Kashiwagi S, Uchiyama K, Mizushima K, et al. Differences in Gut Microbiota Associated With Age, Sex, and Stool Consistency in Healthy Japanese Subjects. J Gastroenterol (2019) 54(1):53–63. doi: 10.1007/s00535-018-1488-5
13. Delgado Jiménez R, Benakis C. The Gut Ecosystem: A Critical Player in Stroke. Neuromolecular Med. (2021) 23(2):236–241. doi: 10.1007/s12017-020-08633-z
14. Knight R, Vrbanac A, Taylor BC, Aksenov A, Callewaert C, Debelius J, et al. Best Practices for Analysing Microbiomes. Nat Rev Microbiol (2018) 16(7):410–22. doi: 10.1038/s41579-018-0029-9
15. Arya AK, Hu B. Brain-Gut Axis After Stroke. Brain Circ (2018) 4(4):165–73. doi: 10.4103/bc.bc_32_18
16. Russo R, Cristiano C, Avagliano C, De Caro C, La Rana G, Raso GM, et al. Gut-Brain Axis: Role of Lipids in the Regulation of Inflammation, Pain and CNS Diseases. Curr Med Chem (2018) 25(32):3930–52. doi: 10.2174/0929867324666170216113756
17. Fung TC, Olson CA, Hsiao EY. Interactions Between the Microbiota, Immune and Nervous Systems in Health and Disease. Nat Neurosci (2017) 20(2):145–55. doi: 10.1038/nn.4476
18. Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, et al. The Microbiota-Gut-Brain Axis. Physiol Rev (2019) 99(4):1877–2013. doi: 10.1152/physrev.00018.2018
19. Benakis C, Martin-Gallausiaux C, Trezzi JP, Melton P, Liesz A, Wilmes P. The Microbiome-Gut-Brain Axis in Acute and Chronic Brain Diseases. Curr Opin Neurobiol (2020) 61:1–9. doi: 10.1016/j.conb.2019.11.009
20. Cryan JF, O’Riordan KJ, Sandhu K, Peterson V, Dinan TG. The Gut Microbiome in Neurological Disorders. Lancet Neurol (2020) 19(2):179–94. doi: 10.1016/S1474-4422(19)30356-4
21. Houlden A, Goldrick M, Brough D, Vizi ES, Lénárt N, Martinecz B, et al. Brain Injury Induces Specific Changes in the Caecal Microbiota of Mice via Altered Autonomic Activity and Mucoprotein Production. Brain Behav Immun (2016) 57:10–20. doi: 10.1016/j.bbi.2016.04.003
22. Antila S, Karaman S, Nurmi H, Airavaara M, Voutilainen MH, Mathivet T, et al. Development and Plasticity of Meningeal Lymphatic Vessels. J Exp Med (2017) 214(12):3645–67. doi: 10.1084/jem.20170391
23. Singh V, Sadler R, Heindl S, Llovera G, Roth S, Benakis C, et al. The Gut Microbiome Primes a Cerebroprotective Immune Response After Stroke. J Cereb Blood Flow Metab (2018) 38(8):1293–8. doi: 10.1177/0271678X18780130
24. Brea D, Poon C, Benakis C, Lubitz G, Murphy M, Iadecola C, et al. Stroke Affects Intestinal Immune Cell Trafficking to the Central Nervous System. Brain Behav Immun (2021) 96:295–302. doi: 10.1016/j.bbi.2021.05.008
25. Kigerl KA, Hall JCE, Wang L, Mo X, Yu Z, Popovich PG. Gut Dysbiosis Impairs Recovery After Spinal Cord Injury. J Exp Med (2016) 213(12):2603–20. doi: 10.1084/jem.20151345
26. Wen L, You W, Wang H, Meng Y, Feng J, Yang X. Polarization of Microglia to the M2 Phenotype in a Peroxisome Proliferator-Activated Receptor Gamma-Dependent Manner Attenuates Axonal Injury Induced by Traumatic Brain Injury in Mice. J neurotrauma (2018) 35(19):2330–40. doi: 10.1089/neu.2017.5540
27. Carlson AL, Xia K, Azcarate-Peril MA, Goldman BD, Ahn M, Styner MA, et al. Infant Gut Microbiome Associated With Cognitive Development. Biol Psychiatry (2018) 83(2):148–59. doi: 10.1016/j.biopsych.2017.06.021
28. Gao W, Salzwedel AP, Carlson AL, Xia K, Azcarate-Peril MA, Styner MA, et al. Gut Microbiome and Brain Functional Connectivity in Infants-A Preliminary Study Focusing on the Amygdala. Psychopharmacology (2019) 236(5):1641–51. doi: 10.1007/s00213-018-5161-8
29. Harms AS, Thome AD, Yan Z, Schonhoff AM, Williams GP, Li X, et al. Peripheral Monocyte Entry Is Required for Alpha-Synuclein Induced Inflammation and Neurodegeneration in a Model of Parkinson Disease. Exp Neurol (2018) 300:179–87. doi: 10.1016/j.expneurol.2017.11.010
30. Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL. An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Cell (2005) 122(1):107–18. doi: 10.1016/j.cell.2005.05.007
31. Atarashi K, Suda W, Luo C, Kawaguchi T, Motoo I, Narushima S, et al. Ectopic Colonization of Oral Bacteria in the Intestine Drives TH1 Cell Induction and Inflammation. Science (2017) 358(6361):359–65. doi: 10.1126/science.aan4526
32. Fort MM, Cheung J, Yen D, Li J, Zurawski SM, Lo S, et al. IL-25 Induces IL-4, IL-5, and IL-13 and Th2-Associated Pathologies In Vivo. Immunity (2001) 15(6):985–95. doi: 10.1016/s1074-7613(01)00243-6
33. Wang Y, Yin Y, Chen X, Zhao Y, Wu Y, Li Y, et al. Induction of Intestinal Th17 Cells by Flagellins From Segmented Filamentous Bacteria. Front Immunol (2019) 10:2750. doi: 10.3389/fimmu.2019.02750
34. Round JL, Lee SM, Li J, Tran G, Jabri B, Chatila TA, et al. The Toll-Like Receptor 2 Pathway Establishes Colonization by a Commensal of the Human Microbiota. Sci (New York NY) (2011) 332(6032):974–7. doi: 10.1126/science.1206095
35. Fleming C, Cai Y, Sun X, Jala VR, Xue F, Morrissey S, et al. Microbiota-Activated CD103(+) Dcs Stemming From Microbiota Adaptation Specifically Drive Gammadeltat17 Proliferation and Activation. Microbiome (2017) 5(1):46. doi: 10.1186/s40168-017-0263-9
36. Yang Y, Xu C, Wu D, Wang Z, Wu P, Li L, et al. Γδ T Cells: Crosstalk Between Microbiota, Chronic Inflammation, and Colorectal Cancer. Front Immunol (2018) 9:1483. doi: 10.3389/fimmu.2018.01483
37. Wesemann DR, Portuguese AJ, Meyers RM, Gallagher MP, Cluff-Jones K, Magee JM, et al. Microbial Colonization Influences Early B-Lineage Development in the Gut Lamina Propria. Nature (2013) 501(7465):112–5. doi: 10.1038/nature12496
38. Shaw MH, Kamada N, Kim Y-G, Núñez G. Microbiota-Induced IL-1β, But Not IL-6, Is Critical for the Development of Steady-State TH17 Cells in the Intestine. J Exp Med (2012) 209(2):251–8. doi: 10.1084/jem.20111703
39. Chang PV, Hao L, Offermanns S, Medzhitov R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc Natl Acad Sci United States America (2014) 111(6):2247–52. doi: 10.1073/pnas.1322269111
40. Singh N, Gurav A, Sivaprakasam S, Brady E, Padia R, Shi H, et al. Activation of Gpr109a, Receptor for Niacin and the Commensal Metabolite Butyrate, Suppresses Colonic Inflammation and Carcinogenesis. Immunity (2014) 40(1):128–39. doi: 10.1016/j.immuni.2013.12.007
41. Parada Venegas D, de la Fuente MK, Landskron G, Gonzalez MJ, Quera R, Dijkstra G, et al. Short Chain Fatty Acids (Scfas)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol (2019) 10:277. doi: 10.3389/fimmu.2019.00277
42. Sun X, Jiao X, Ma Y, Liu Y, Zhang L, He Y, et al. Trimethylamine N-Oxide Induces Inflammation and Endothelial Dysfunction in Human Umbilical Vein Endothelial Cells via Activating ROS-TXNIP-NLRP3 Inflammasome. Biochem Biophys Res Commun (2016) 481(1-2):63–70. doi: 10.1016/j.bbrc.2016.11.017
43. Dasgupta S, Erturk-Hasdemir D, Ochoa-Reparaz J, Reinecker H-C, Kasper DL. Plasmacytoid Dendritic Cells Mediate Anti-Inflammatory Responses to a Gut Commensal Molecule via Both Innate and Adaptive Mechanisms. Cell Host Microbe (2014) 15(4):413–23. doi: 10.1016/j.chom.2014.03.006
44. Johnson JL, Jones MB, Cobb BA. Polysaccharide a From the Capsule of Bacteroides Fragilis Induces Clonal CD4+ T Cell Expansion. J Biol Chem (2015) 290(8):5007–14. doi: 10.1074/jbc.M114.621771
45. Wang Y, Begum-Haque S, Telesford KM, Ochoa-Reparaz J, Christy M, Kasper EJ, et al. A Commensal Bacterial Product Elicits and Modulates Migratory Capacity of CD39(+) CD4 T Regulatory Subsets in the Suppression of Neuroinflammation. Gut Microbes (2014) 5(4):552–61. doi: 10.4161/gmic.29797
46. Feigin VL, Norrving B, Mensah GA. Global Burden of Stroke. Circ Res (2017) 120(3):439–48. doi: 10.1161/CIRCRESAHA.116.308413
47. Ojaghihaghighi S, Vahdati SS, Mikaeilpour A, Ramouz A. Comparison of Neurological Clinical Manifestation in Patients With Hemorrhagic and Ischemic Stroke. World J Emerg Med (2017) 8(1):34–8. doi: 10.5847/wjem.j.1920-8642.2017.01.006
48. Yin J, Liao S-X, He Y, Wang S, Xia G-H, Liu F-T, et al. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients With Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. J Am Heart Assoc (2015) 4(11):e002699. doi: 10.1161/JAHA.115.002699
49. Haak BW, Westendorp WF, van Engelen TSR, Brands X, Brouwer MC, Vermeij J-D, et al. Disruptions of Anaerobic Gut Bacteria Are Associated With Stroke and Post-Stroke Infection: A Prospective Case-Control Study. Trans Stroke Res (2020) 11:110–12. doi: 10.1007/s12975-020-00863-4
50. Zeng X, Gao X, Peng Y, Wu Q, Zhu J, Tan C, et al. Higher Risk of Stroke is Correlated With Increased Opportunistic Pathogen Load and Reduced Levels of Butyrate-Producing Bacteria in the Gut. Front Cell Infect Microbiol (2019) 9:4. doi: 10.3389/fcimb.2019.00004
51. Nie J, Xie L, Zhao B-X, Li Y, Qiu B, Zhu F, et al. Serum Trimethylamine N-Oxide Concentration Is Positively Associated With First Stroke in Hypertensive Patients. Stroke (2018) 49(9):2021–8. doi: 10.1161/STROKEAHA.118.021997
52. Stanley D, Mason LJ, Mackin KE, Srikhanta YN, Lyras D, Prakash MD, et al. Translocation and Dissemination of Commensal Bacteria in Post-Stroke Infection. Nat Med (2016) 22(11):1277–84. doi: 10.1038/nm.4194
53. Spychala MS, Venna VR, Jandzinski M, Doran SJ, Durgan DJ, Ganesh BP, et al. Age-Related Changes in the Gut Microbiota Influence Systemic Inflammation and Stroke Outcome. Ann Neurol (2018) 84(1):23–36. doi: 10.1002/ana.25250
54. Singh V, Roth S, Llovera G, Sadler R, Garzetti D, Stecher B, et al. Microbiota Dysbiosis Controls the Neuroinflammatory Response After Stroke. J Neurosci (2016) 36(28):7428–40. doi: 10.1523/JNEUROSCI.1114-16.2016
55. Benakis C, Brea D, Caballero S, Faraco G, Moore J, Murphy M, et al. Commensal Microbiota Affects Ischemic Stroke Outcome by Regulating Intestinal Γδ T Cells. Nat Med (2016) 22(5):516–23. doi: 10.1038/nm.4068
56. Benakis C, Poon C, Lane D, Brea D, Sita G, Moore J, et al. Distinct Commensal Bacterial Signature in the Gut Is Associated With Acute and Long-Term Protection From Ischemic Stroke. Stroke (2020) 51(6):1844–54. doi: 10.1161/STROKEAHA.120.029262
57. Sadler R, Cramer JV, Heindl S, Kostidis S, Betz D, Zuurbier KR, et al. Short-Chain Fatty Acids Improve Poststroke Recovery via Immunological Mechanisms. J Neurosci (2020) 40(5):1162–73. doi: 10.1523/JNEUROSCI.1359-19.2019
58. Lee J, d’Aigle J, Atadja L, Quaicoe V, Honarpisheh P, Ganesh BP, et al. Gut Microbiota-Derived Short-Chain Fatty Acids Promote Poststroke Recovery in Aged Mice. Circ Res (2020) 127(4):453–65. doi: 10.1161/CIRCRESAHA.119.316448
59. Winek K, Engel O, Koduah P, Heimesaat MM, Fischer A, Bereswill S, et al. Depletion of Cultivatable Gut Microbiota by Broad-Spectrum Antibiotic Pretreatment Worsens Outcome After Murine Stroke. Stroke (2016) 47(5):1354–63. doi: 10.1161/STROKEAHA.115.011800
60. Xu DJ, Wang KC, Yuan LB, Li HF, Xu YY, Wei LY, et al. Compositional and Functional Alterations of Gut Microbiota in Patients With Stroke. Nutr Metab Cardiovasc Dis (2021) 31(12):3434–48. doi: 10.1016/j.numecd.2021.08.045
61. Ling Y, Gong T, Zhang J, Gu Q, Gao X, Weng X, et al. Gut Microbiome Signatures Are Biomarkers for Cognitive Impairment in Patients With Ischemic Stroke. Front Aging Neurosci 12:511562. doi: 10.3389/fnagi.2020.511562
62. Xiang L, Lou Y, Liu L, Liu Y, Zhang W, Deng J, et al. Gut Microbiotic Features Aiding the Diagnosis of Acute Ischemic Stroke. Front Cell Infect Microbiol (2020) 10:587284. doi: 10.3389/fcimb.2020.587284
63. Huang Y, Shen Z, He W. Identification of Gut Microbiome Signatures in Patients With Post-Stroke Cognitive Impairment and Affective Disorder. Front Aging Neurosci (2021) 13:706765. doi: 10.3389/fnagi.2021.706765
64. Xu K, Gao X, Xia G, Chen M, Zeng N, Wang S, et al. Rapid Gut Dysbiosis Induced by Stroke Exacerbates Brain Infarction in Turn. Gut (2021) 70:1486–94. doi: 10.1136/gutjnl-2020-323263
65. Jeon J, Lourenco J, Kaiser EE, Waters ES, Scheulin KM, Fang X, et al. Dynamic Changes in the Gut Microbiome at the Acute Stage of Ischemic Stroke in a Pig Model. Front Neurosci (2020) 14:587986. doi: 10.3389/fnins.2020.587986
66. Wu W, Sun Y, Luo N, Cheng C, Jiang C, Yu Q, et al. Integrated 16S Rrna Gene Sequencing and LC-MS Analysis Revealed the Interplay Between Gut Microbiota and Plasma Metabolites in Rats With Ischemic Stroke. J Mol Neurosci (2021) 71(10):2095–106. doi: 10.1007/s12031-021-01828-4
67. Tan C, Wu Q, Wang H, Gao X, Xu R, Cui Z, et al. Dysbiosis of Gut Microbiota and Short-Chain Fatty Acids in Acute Ischemic Stroke and the Subsequent Risk for Poor Functional Outcomes. JPEN J Parenter Enteral Nutr (2021) 45(3):518–29. doi: 10.1002/jpen.1861
68. Zhang J, Wang L, Cai J, Lei A, Liu C, Lin R, et al. Gut Microbial Metabolite TMAO Portends Prognosis in Acute Ischemic Stroke. J Neuroimmunol (2021) 354:577526. doi: 10.1016/j.jneuroim.2021.577526
69. Guo Q, Jiang X, Ni C, Li L, Chen L, Wang Y, et al. Gut Microbiota-Related Effects of Tanhuo Decoction in Acute Ischemic Stroke. Oxid Med Cell Longev (2021) 2021:5596924. doi: 10.1155/2021/5596924
70. Sun T, Zhang Y, Yin J, Peng X, Zhou L, Huang S, et al. Association of Gut Microbiota-Dependent Metabolite Trimethylamine N-Oxide With First Ischemic Stroke. J Atheroscler Thromb (2021) 28(4):320–8. doi: 10.5551/jat.55962
71. Huang Q, Di L, Yu F, Feng X, Liu Z, Wei M, et al. Alterations in the Gut Microbiome With Hemorrhagic Transformation in Experimental Stroke. CNS Neurosci Ther (2022) 28(1):77–91. doi: 10.1111/cns.13736
72. Zhang P, Zhang X, Huang Y, Chen J, Shang W, Shi G, et al. Atorvastatin Alleviates Microglia-Mediated Neuroinflammation via Modulating the Microbial Composition and the Intestinal Barrier Function in Ischemic Stroke Mice. Free Radic Biol Med (2021) 162:104–17. doi: 10.1016/j.freeradbiomed.2020.11.032
73. Huang JT, Mao YQ, Han B, Zhang ZY, Chen HL, Li ZM, et al. Calorie Restriction Conferred Improvement Effect on Long-Term Rehabilitation of Ischemic Stroke via Gut Microbiota. Pharmacol Res (2021) 170:105726. doi: 10.1016/j.phrs.2021.105726
74. Zhu W, Romano KA, Li L, Buffa JA, Sangwan N, Prakash P, et al. Gut Microbes Impact Stroke Severity via the Trimethylamine N-Oxide Pathway. Cell Host Microbe (2021) 29(7):1199–208. doi: 10.1016/j.chom.2021.05.002
75. Yuan Q, Xin L, Han S, Su Y, Wu R, Liu X, et al. Lactulose Improves Neurological Outcomes by Repressing Harmful Bacteria and Regulating Inflammatory Reactions in Mice After Stroke. Front Cell Infect Microbiol (2021) 11:644448. doi: 10.3389/fcimb.2021.644448
76. Li N, Wang X, Sun C, Wu X, Lu M, Si Y, et al. Change of Intestinal Microbiota in Cerebral Ischemic Stroke Patients. BMC Microbiol (2019) 19(1):191–8. doi: 10.1186/s12866-019-1552-1
77. Sun J, Ling Z, Wang F, Chen W, Li H, Jin J, et al. Clostridium Butyricum Pretreatment Attenuates Cerebral Ischemia/Reperfusion Injury in Mice via Anti-Oxidation and Anti-Apoptosis. Neurosci Lett (2016) 613:30–5. doi: 10.1016/j.neulet.2015.12.047
78. Zhou Z, Xu N, Matei N, McBride DW, Ding Y, Liang H, et al. Sodium Butyrate Attenuated Neuronal Apoptosis via GPR41/Gbetagamma/PI3K/Akt Pathway After MCAO in Rats. J Cereb Blood Flow Metab (2021) 41(2):267–81. doi: 10.1177/0271678X20910533
79. Yan H, Ajuwon KM. Butyrate Modifies Intestinal Barrier Function in IPEC-J2 Cells Through a Selective Upregulation of Tight Junction Proteins and Activation of the Akt Signaling Pathway. PloS One (2017) 12(6):e0179586. doi: 10.1371/journal.pone.0179586
80. Wu C, Li C, Zhao W, Xie N, Yan F, Lian Y, et al. Elevated Trimethylamine N-Oxide Related to Ischemic Brain Lesions After Carotid Artery Stenting. Neurology (2018) 90(15):e1283–90. doi: 10.1212/WNL.0000000000005298
81. Tan C, Wang H, Gao X, Xu R, Zeng X, Cui Z, et al. Dynamic Changes and Prognostic Value of Gut Microbiota-Dependent Trimethylamine-N-Oxide in Acute Ischemic Stroke. Front Neurol (2020) 11:2020.00029. doi: 10.3389/fneur.2020.00029
82. Farhangi MA, Vajdi M, Asghari-Jafarabadi M. Gut Microbiota-Associated Metabolite Trimethylamine N-Oxide and the Risk of Stroke: A Systematic Review and Dose-Response Meta-Analysis. Nutr J (2020) 19(1):76. doi: 10.1186/s12937-020-00592-2
83. Chen W-C, Chang L-H, Huang S-S, Huang Y-J, Chih C-L, Kuo H-C, et al. Aryl Hydrocarbon Receptor Modulates Stroke-Induced Astrogliosis and Neurogenesis in the Adult Mouse Brain. J Neuroinflamm (2019) 16(1):187–13. doi: 10.1186/s12974-019-1572-7
84. Macdonald RL, Schweizer TA. Spontaneous Subarachnoid Haemorrhage. Lancet (2017) 389(10069):655–66. doi: 10.1016/S0140-6736(16)30668-7
85. Li H, Xu H, Li Y, Jiang Y, Hu Y, Liu T, et al. Alterations of Gut Microbiota Contribute to the Progression of Unruptured Intracranial Aneurysms. Nat Commun (2020) 11(1):3218. doi: 10.1038/s41467-020-16990-3
86. Kawabata S, Takagaki M, Nakamura H, Oki H, Motooka D, Nakamura S, et al. Dysbiosis of Gut Microbiome is Associated With Rupture of Cerebral Aneurysms. Stroke (2021), STROKEAHA121034792. doi: 10.1161/STROKEAHA.121.034792
87. Shikata F, Shimada K, Sato H, Ikedo T, Kuwabara A, Furukawa H, et al. Potential Influences of Gut Microbiota on the Formation of Intracranial Aneurysm. Hypertension (2019) 73(2):491–6. doi: 10.1161/HYPERTENSIONAHA.118.11804
88. Maas AIR, Menon DK, Adelson PD, Andelic N, Bell MJ, Belli A, et al. Traumatic Brain Injury: Integrated Approaches to Improve Prevention, Clinical Care, and Research. Lancet Neurol (2017) 16(12):987–1048. doi: 10.1016/S1474-4422(17)30371-X
89. Simon DW, Rogers MB, Gao Y, Vincent G, Firek BA, Janesko-Feldman K, et al. Depletion of Gut Microbiota Is Associated With Improved Neurologic Outcome Following Traumatic Brain Injury. Brain Res (2020) 1747:147056. doi: 10.1016/j.brainres.2020.147056
90. Mahajan C, Khurana S, Kapoor I, Sokhal S, Kumar S, Prabhakar H, et al. Characteristics of Gut Microbiome After Traumatic Brain Injury. J Neurosurg Anesthesiol (2021). doi: 10.1097/ANA.0000000000000789
91. Hou Y, Xu L, Song S, Fan W, Wu Q, Tong X, et al. Oral Administration of Brain Protein Combined With Probiotics Induces Immune Tolerance Through the Tryptophan Pathway. Front Mol Neurosci (2021) 14:634631. doi: 10.3389/fnmol.2021.634631
92. Treangen TJ, Wagner J, Burns MP, Villapol S. Traumatic Brain Injury in Mice Induces Acute Bacterial Dysbiosis Within the Fecal Microbiome. Front Immunol (2018) 9:2757. doi: 10.3389/fimmu.2018.02757
93. Li H, Sun J, Du J, Wang F, Fang R, Yu C, et al. Clostridium Butyricum Exerts a Neuroprotective Effect in a Mouse Model of Traumatic Brain Injury via the Gut-Brain Axis. Neurogastroenterol Motil (2018) 30(5):e13260. doi: 10.1111/nmo.13260
94. Angoa-Pérez M, Zagorac B, Anneken JH, Briggs DI, Winters AD, Greenberg JM, et al. Repetitive, Mild Traumatic Brain Injury Results in a Progressive White Matter Pathology, Cognitive Deterioration, and a Transient Gut Microbiota Dysbiosis. Sci Rep (2020) 10(1):8949. doi: 10.1038/s41598-020-65972-4
95. Opeyemi OM, Rogers MB, Firek BA, Janesko-Feldman K, Vagni V, Mullett SJ, et al. Sustained Dysbiosis and Decreased Fecal Short-Chain Fatty Acids After Traumatic Brain Injury and Impact on Neurologic Outcome. J Neurotrauma (2021) 38(18):2610–21. doi: 10.1089/neu.2020.7506
96. Du D, Tang W, Zhou C, Sun X, Wei Z, Zhong J, et al. Fecal Microbiota Transplantation Is a Promising Method to Restore Gut Microbiota Dysbiosis and Relieve Neurological Deficits After Traumatic Brain Injury. Oxid Med Cell Longevity (2021) 2021:5816837. doi: 10.1155/2021/5816837
97. You W, Zhu Y, Wei A, Du J, Wang Y, Zheng P, et al. Traumatic Brain Injury Induces Gastrointestinal Dysfunction and Dysbiosis of Gut Microbiota Accompanied by Alterations of Bile Acid Profile. J Neurotrauma (2021). doi: 10.1089/neu.2020.7526
98. Celorrio M, Abellanas MA, Rhodes J, Goodwin V, Moritz J, Vadivelu S, et al. Gut Microbial Dysbiosis After Traumatic Brain Injury Modulates the Immune Response and Impairs Neurogenesis. Acta Neuropathol Commun (2021) 9(1):40. doi: 10.1186/s40478-021-01137-2
99. Nicholson SE, Watts LT, Burmeister DM, Merrill D, Scroggins S, Zou Y, et al. Moderate Traumatic Brain Injury Alters the Gastrointestinal Microbiome in a Time-Dependent Manner. Shock (2019) 52(2):240–8. doi: 10.1097/SHK.0000000000001211
100. Blanke EN, Holmes GM, Besecker EM. Altered Physiology of Gastrointestinal Vagal Afferents Following Neurotrauma. Neural Regener Res (2021) 16(2):254–63. doi: 10.4103/1673-5374.290883
101. Hang C-H, Shi J-X, Li J-S, Li W-Q, Yin H-X. Up-Regulation of Intestinal Nuclear Factor Kappa B and Intercellular Adhesion Molecule-1 Following Traumatic Brain Injury in Rats. World J Gastroenterol (2005) 11(8):1149–54. doi: 10.3748/wjg.v11.i8.1149
102. Lu J, Frerich JM, Turtzo LC, Li S, Chiang J, Yang C, et al. Histone Deacetylase Inhibitors Are Neuroprotective and Preserve NGF-Mediated Cell Survival Following Traumatic Brain Injury. Proc Natl Acad Sci United States America (2013) 110(26):10747–52. doi: 10.1073/pnas.1308950110
103. Zhang X, Jiang X. Effects of Enteral Nutrition on the Barrier Function of the Intestinal Mucosa and Dopamine Receptor Expression in Rats With Traumatic Brain Injury. JPEN J Parenter Enteral Nutr (2015) 39(1):114–23. doi: 10.1177/0148607113501881
104. Wan G, Wang L, Zhang G, Zhang J, Lu Y, Li J, et al. Effects of Probiotics Combined With Early Enteral Nutrition on Endothelin-1 and C-Reactive Protein Levels and Prognosis in Patients With Severe Traumatic Brain Injury. J Int Med Res (2020) 48(3):300060519888112. doi: 10.1177/0300060519888112
105. Tan M, Zhu J-C, Du J, Zhang L-M, Yin H-H. Effects of Probiotics on Serum Levels of Th1/Th2 Cytokine and Clinical Outcomes in Severe Traumatic Brain-Injured Patients: A Prospective Randomized Pilot Study. Crit Care (London England) (2011) 15(6):R290–10. doi: 10.1186/cc10579
106. Bansal V, Costantini T, Ryu SY, Peterson C, Loomis W, Putnam J, et al. Stimulating the Central Nervous System to Prevent Intestinal Dysfunction After Traumatic Brain Injury. J Trauma (2010) 68(5):1059–64. doi: 10.1097/TA.0b013e3181d87373
107. Stothers L, Macnab AJ, Mukisa R, Mutabazi S, Bajunirwe F. Traumatic Spinal Cord Injury in Uganda: A Prevention Strategy and Mechanism to Improve Home Care. Int J Epidemiol (2017) 46(4):1086–90. doi: 10.1093/ije/dyx058
108. Gungor B, Adiguzel E, Gursel I, Yilmaz B, Gursel M. Intestinal Microbiota in Patients With Spinal Cord Injury. PloS One (2016) 11(1):e0145878. doi: 10.1371/journal.pone.0145878
109. Zhang C, Zhang W, Zhang J, Jing Y, Yang M, Du L, et al. Gut Microbiota Dysbiosis in Male Patients With Chronic Traumatic Complete Spinal Cord Injury. J Trans Med (2018) 16(1):353–16. doi: 10.1186/s12967-018-1735-9
110. Lin R, Xu J, Ma Q, Chen M, Wang L, Wen S, et al. Alterations in the Fecal Microbiota of Patients With Spinal Cord Injury. PloS One (2020) 15(8):e0236470. doi: 10.1371/journal.pone.0236470
111. Li J, van der Pol W, Eraslan M, McLain A, Cetin H, Cetin B, et al. Comparison of the Gut Microbiome Composition Among Individuals With Acute or Long-Standing Spinal Cord Injury vs. Able-Bodied Controls. J Spinal Cord Med (2020) 4:1–9. doi: 10.1080/10790268.2020.1769949
112. Bazzocchi G, Turroni S, Bulzamini MC, D’Amico F, Bava A, Castiglioni M, et al. Changes in Gut Microbiota in the Acute Phase After Spinal Cord Injury Correlate With Severity of the Lesion. Sci Rep (2021) 11(1):12743. doi: 10.1038/s41598-021-92027-z
113. Yu B, Qiu H, Cheng S, Ye F, Li J, Chen S, et al. Profile of Gut Microbiota in Patients With Traumatic Thoracic Spinal Cord Injury and Its Clinical Implications: A Case-Control Study in a Rehabilitation Setting. Bioengineered (2021) 12(1):4489–99. doi: 10.1080/21655979.2021.1955543
114. O’Connor G, Jeffrey E, Madorma D, Marcillo A, Abreu MT, Deo SK, et al. Investigation of Microbiota Alterations and Intestinal Inflammation Post-Spinal Cord Injury in Rat Model. J Neurotrauma (2018) 35(18):2159–66. doi: 10.1089/neu.2017.5349
115. Myers SA, Gobejishvili L, Saraswat Ohri S, Garrett Wilson C, Andres KR, Riegler AS, et al. Following Spinal Cord Injury, PDE4B Drives an Acute, Local Inflammatory Response and a Chronic, Systemic Response Exacerbated by Gut Dysbiosis and Endotoxemia. Neurobiol Dis (2019) 124:353–63. doi: 10.1016/j.nbd.2018.12.008
116. Jing Y, Yang D, Bai F, Zhang C, Qin C, Li D, et al. Melatonin Treatment Alleviates Spinal Cord Injury-Induced Gut Dysbiosis in Mice. J Neurotrauma (2019) 36(18):2646–64. doi: 10.1089/neu.2018.6012
117. Schmidt EKA, Torres-Espin A, Raposo PJF, Madsen KL, Kigerl KA, Popovich PG, et al. Fecal Transplant Prevents Gut Dysbiosis and Anxiety-Like Behaviour After Spinal Cord Injury in. PloS One (2020) 15(1):e0226128. doi: 10.1371/journal.pone.0226128
118. Jing Y, Yu Y, Bai F, Wang L, Yang D, Zhang C, et al. Effect of Fecal Microbiota Transplantation on Neurological Restoration in a Spinal Cord Injury Mouse Model: Involvement of Brain-Gut Axis. Microbiome (2021) 9(1):59–21. doi: 10.1186/s40168-021-01007-y
119. Schmidt EKA, Raposo PJF, Torres-Espin A, Fenrich KK, Fouad K. Beyond the Lesion Site: Minocycline Augments Inflammation and Anxiety-Like Behavior Following SCI in Rats Through Action on the Gut Microbiota. J Neuroinflammation (2021) 18(1):144. doi: 10.1186/s12974-021-02123-0
120. Doelman A, Tigchelaar S, McConeghy B, Sinha S, Keung MS, Manouchehri N, et al. Characterization of the Gut Microbiome in a Porcine Model of Thoracic Spinal Cord Injury. BMC Genomics (2021) 22(1):775. doi: 10.1186/s12864-021-07979-3
121. Rong Z, Huang Y, Cai H, Chen M, Wang H, Liu G, et al. Gut Microbiota Disorders Promote Inflammation and Aggravate Spinal Cord Injury Through the TLR4/Myd88 Signaling Pathway. Front Nutr (2021) 8:702659. doi: 10.3389/fnut.2021.702659
122. Du J, Zayed AA, Kigerl KA, Zane K, Sullivan MB, Popovich PG. Spinal Cord Injury Changes the Structure and Functional Potential of Gut Bacterial and Viral Communities. mSystems (2021) 6(3):e01356–20. doi: 10.1128/mSystems.01356-20
123. Kigerl KA, Zane K, Adams K, Sullivan MB, Popovich PG. The Spinal Cord-Gut-Immune Axis as a Master Regulator of Health and Neurological Function After Spinal Cord Injury. Exp Neurol (2020) 323:113085. doi: 10.1016/j.expneurol.2019.113085
124. Sun G, Yang S, Cao G, Wang Q, Hao J, Wen Q, et al. Γδ T Cells Provide the Early Source of IFN-Γ to Aggravate Lesions in Spinal Cord Injury. J Exp Med (2018) 215(2):521–35. doi: 10.1084/jem.20170686
125. Kigerl KA, Mostacada K, Popovich PG. Gut Microbiota are Disease-Modifying Factors After Traumatic Spinal Cord Injury. Neurotherapeutics (2018) 15(1):60–7. doi: 10.1007/s13311-017-0583-2
126. Wang X, Wu J, Liu X, Tang K, Cheng L, Li J, et al. Engineered Liposomes Targeting the Gut-CNS Axis for Comprehensive Therapy of Spinal Cord Injury. J Control Release (2021) 331:390–403. doi: 10.1016/j.jconrel.2021.01.032
127. Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. Obesity Alters Gut Microbial Ecology. Proc Natl Acad Sci USA (2005) 102(31):11070–5. doi: 10.1073/pnas.0504978102
128. Mann ER, Landy JD, Bernardo D, Peake STC, Hart AL, Al-Hassi HO, et al. Intestinal Dendritic Cells: Their Role in Intestinal Inflammation, Manipulation by the Gut Microbiota and Differences Between Mice and Men. Immunol Lett (2013) 150(1-2):30–40. doi: 10.1016/j.imlet.2013.01.007
129. De Paepe M, Leclerc M, Tinsley CR, Petit M-A. Bacteriophages: An Underestimated Role in Human and Animal Health? Front Cell Infect Microbiol (2014) 4:39. doi: 10.3389/fcimb.2014.00039
130. Sommer F, Bäckhed F. The Gut Microbiota–Masters of Host Development and Physiology. Nat Rev Microbiol (2013) 11(4):227–38. doi: 10.1038/nrmicro2974
131. Macpherson AJ, Harris NL. Interactions Between Commensal Intestinal Bacteria and the Immune System. Nat Rev Immunol (2004) 4(6):478–85. doi: 10.1038/nri1373
132. Erny D, Hrabě de Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat Neurosci (2015) 18(7):965–77. doi: 10.1038/nn.4030
Keywords: gut microbiota, stroke, traumatic brain injury, spinal cord injury, gut-brain axis
Citation: Yuan B, Lu X-j and Wu Q (2021) Gut Microbiota and Acute Central Nervous System Injury: A New Target for Therapeutic Intervention. Front. Immunol. 12:800796. doi: 10.3389/fimmu.2021.800796
Received: 24 October 2021; Accepted: 09 December 2021;
Published: 24 December 2021.
Edited by:
Alina Maria Holban, University of Bucharest, RomaniaReviewed by:
Stuart Friess, Washington University in St. Louis, United StatesCopyright © 2021 Yuan, Lu and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiao-jie Lu, eGlhb2ppZWx1d3V4aUAxNjMuY29t; Qi Wu, bnN3dXFpQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.