 Bruna Kulmann-Leal
Bruna Kulmann-Leal Joel Henrique Ellwanger
Joel Henrique Ellwanger José Artur Bogo Chies
José Artur Bogo Chies- Laboratório de Imunobiologia e Imunogenética, Programa de Pós-Graduação em Genética e Biologia Molecular (PPGBM), Departamento de Genética, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, Brazil
The genetic background of Brazilians encompasses Amerindian, African, and European components as a result of the colonization of an already Amerindian inhabited region by Europeans, associated to a massive influx of Africans. Other migratory flows introduced into the Brazilian population genetic components from Asia and the Middle East. Currently, Brazil has a highly admixed population and, therefore, the study of genetic factors in the context of health or disease in Brazil is a challenging and remarkably interesting subject. This phenomenon is exemplified by the genetic variant CCR5Δ32, a 32 base-pair deletion in the CCR5 gene. CCR5Δ32 originated in Europe, but the time of origin as well as the selective pressures that allowed the maintenance of this variant and the establishment of its current frequencies in the different human populations is still a field of debates. Due to its origin, the CCR5Δ32 allele frequency is high in European-derived populations (~10%) and low in Asian and African native human populations. In Brazil, the CCR5Δ32 allele frequency is intermediate (4-6%) and varies on the Brazilian States, depending on the migratory history of each region. CCR5 is a protein that regulates the activity of several immune cells, also acting as the main HIV-1 co-receptor. The CCR5 expression is influenced by CCR5Δ32 genotypes. No CCR5 expression is observed in CCR5Δ32 homozygous individuals. Thus, the CCR5Δ32 has particular effects on different diseases. At the population level, the effect that CCR5Δ32 has on European populations may be different than that observed in highly admixed populations. Besides less evident due to its low frequency in admixed groups, the effect of the CCR5Δ32 variant may be affected by other genetic traits. Understanding the effects of CCR5Δ32 on Brazilians is essential to predict the potential use of pharmacological CCR5 modulators in Brazil. Therefore, this study reviews the impacts of the CCR5Δ32 on the Brazilian population, considering infectious diseases, inflammatory conditions, and cancer. Finally, this article provides a general discussion concerning the impacts of a European-derived variant, the CCR5Δ32, on a highly admixed population.
Introduction
Genetic Aspects of the Brazilian Population
Until the year 1500 CE, Brazil was inhabited only by Native Americans belonging to different linguistic groups, distributed along the coast and hinterland of the country. This scenario changed dramatically after the arrival of the Portuguese explorers in the Brazilian territory that year, affecting many cultural and biological aspects of the native populations. The European colonization of Brazil and the associated influx of Africans had a strong influence on the genetic makeup of the Brazilian population. In Brazil, as well as in other countries colonized by the Europeans, the Native American population deeply declined after colonization (contracted around 90% in the Americas) (1–3). The remaining native population underwent a strong process of genetic miscegenation. However, the processes of population change continued throughout Brazilian history, even in more recent times. Over the past 200 years, Brazil has received a large influx of European immigrants from various countries, also described as the last migration pulse, which added another layer to the genetic makeup of the Brazilian population (1–4).
In general terms, the genetic background of current Brazilians has Amerindian, African, and European components in different proportions (2, 3, 5–7), depending on the Brazilian region under investigation (North, Northeast, Center-West, Southeast, or South). For example, the genetic makeup of Brazilians in the southern region of Brazil was strongly influenced by migratory flows from Europe in the 19th and 20th centuries; although in the Northeast of the country, the African genetic component is high (1, 2, 8). Of note, the European component is preponderant in different Brazilian regions when the Amerindian, African, and European components are compared, but even observing some regional peculiarities as those mentioned above, the genetic composition of the Brazilian population is rather uniform in its miscegenation in different regions of the country (1).
Throughout history, Brazil also received migrants from other countries beyond those from Europe and Africa, including countries from Asia and Middle East (7, 9). The intense migration within the national territory (10) allowed the exchange of genetic information between Brazilians from different regions, ethnic and genetic groups. As a result of the interactions of these different groups, the Brazilian population is currently highly miscegenated, a characteristic evident in the rich genetic and phenotypic diversity observed among the Brazilian population (2, 6, 11, 12). Considering the scenario mentioned above, the Brazilian population can be considered genetically heterogeneous and admixed, in addition to being relatively uniform throughout the country (1). Interestingly, admixed Brazilian populations are probable “reservoirs” of the diverse Native American genetic component (3), currently the least prevalent genetic component in the population (1, 8).
Y-chromosome haplogroup analysis corroborates the high genetic miscegenation observed in the Brazilian population. Abe-Sandes et al. (13) investigated the frequency of different haplogroups in Brazilian individuals from different ethnicities. A significant frequency of typical European haplotypes in Afro-Brazilians was found, for example, in the Quilombola community of São Gonçalo, Bahia state, northeastern Brazil. Abe-Sandes et al. (13) also found the E-SRY4064 haplotype, usually observed in populations from Sub-Saharian Africa and almost absent in populations from Europe and Asia, in white Brazilians, in a notable frequency (13). Marrero et al. (14) also reported evidence of admixture in Native American populations, showing the presence of non-Amerindian haplotypes in Kaingang and Guarani peoples (14). Finally, numerous studies analyzing Y-chromosome haplogroups reinforce the miscegenation addressed in this article, pointing to European, Amerindian, African and Asian haplogroups in different ethnicities and population groups from different Brazilian regions (13–28).
In the same direction, evaluation of mitochondrial DNA in different populations of Brazil showed the presence of diverse haplogroups characteristic of African, European, Native American and Asian populations, again evidencing the high level of miscegenation in the Brazilian population (14, 29–37). Of note, Cardena et al. (38) assessed a population from São Paulo, southeastern state of Brazil, specifically evaluating mtDNA haplogroups and comparing such data with self-declared ethnicity. Interestingly, a significant parcel of the individuals classified as whites showed a high percentage of African mtDNA (37.6%), with less participation of Amerindian (31.6%) and European (30.8%) origins. When analyzing other genomic loci of the same individuals, a higher European contribution was noticed (63.3%), evidencing a considerable African participation of maternal origin in individuals simultaneously presenting high non mtDNA European ancestry (38, 39).
Pivotal Information Regarding the CCR5Δ32 Variant
The CCR5Δ32 polymorphism (reference SNP ID number: rs333) is a genetic variant that originated in the European population (40), and therefore can be used as an ancestry-informative marker in studies involving population genetics and genome ancestry (41, 42). This variant represents a 32-base pair deletion in the CCR5 gene (chromosome 3; 3p.21.31), a fundamental component of the immune system responsible for encoding the CCR5 protein, which acts mainly in the regulation of inflammatory cell migration. It is unclear what selective pressures (considering positive selection) were responsible for fixing CCR5Δ32 in the human genome. Smallpox, bubonic plague, and other infectious diseases have already been suggested, but there is no consensus on this aspect (40). Neutral evolution is also a possibility (43). What is somehow certain is that the variant probably originated in the European population at 700-5,000 years ago (43, 44), potentially even earlier than 5,000 years (45, 46), and later spread heterogeneously across the world.
The CCR5Δ32 allele presents a higher frequency in northern Europe (greater than 15% in Norway, Latvia, and Estonia), being less frequent in countries located in the south of the European continent. For example, the frequency of the CCR5Δ32 allele is 8.1% in Spain, 6.9% in Portugal, 6.2% in Italy, and 5.1% in Greece. The allele frequency is very low or even absent in most Asian and African countries: for example, 0.4% in China, 2.2% in Korea, 0.7% in Cameroon, 0.26% in Eritrea, and 2.9% in Egypt (47). A recent study reports the absence of the CCR5Δ32 allele in the Nepalese population (48). Similarly, CCR5Δ32 is rare in Native American groups, showing an overall CCR5Δ32 allele frequency of 0.2%, mostly probably due to miscegenation (42). In the contemporary Brazilian population, the overall frequency of the CCR5Δ32 allele usually ranges from 4 to 6% but showing significant variations between different Brazilian regions and ethnic groups (42, 49), as will be discussed in the next sections of this article.
The main function of the CCR5 is coordinating leukocyte migration during inflammatory reactions through interaction with different chemokines, especially CCL3, CCL4, and CCL5 (40). Of note, these chemokines were historically called “MIP-1α”, “MIP-1β” and “RANTES”, respectively, but that denomination has fallen into disuse (50, 51). The CCR5 protein is expressed on the cell surface and has seven transmembrane domains connected by three extracellular loops and three intracellular loops. Leukocytes are the main cells that express the CCR5 (40), although the protein is also detected in other cell types, such as human embryonic neurons (52), adipocytes (53), and several types of cancer cells and tissues (54–58), indicating that CCR5 performs immune functions that go beyond coordinating the migration of inflammatory cells.
Carriers of the wild-type CCR5 gene have CCR5 expression constitutively, with some variation between individuals. CCR5Δ32 causes important phenotypic effects, affecting the interaction of the CCR5 with chemokines. Due to the induction of a change in the CCR5 gene reading frame, the CCR5Δ32 produces a truncated protein that is not expressed on the cell surface, presenting a gene-dosage effect. In brief, the presence of the CCR5Δ32 allele in heterozygous causes a reduction in the expression of CCR5 at the membrane. The presence of the CCR5Δ32 allele in homozygosis culminate in virtually no expression of CCR5 molecules on the cell surface (59–63). The CCR5Δ32-derived molecules are not phosphorylated and remain retained in the endoplasmic reticulum (64). Interestingly, it was suggested that in addition to the gene-dosage effect associated to CCR5Δ32, the CCR5Δ32-derived truncated protein could promote the sequestration of the CCR5 and CXCR4 proteins, both HIV-1 co-receptors, from the cell surface (65, 66).
These changes in the expression of CCR5 associated to CCR5Δ32 culminate in a disrupted CCR5-mediated immune response, which can be beneficial in some situations or harmful in others (67) since the ‘chemokine system’ is not completely redundant. The absence of CCR5 can impact the cell signaling coordinated by CCL3, CCL4 and CCL5, thus perturbing the proper CCR5-mediated immune responses (68). Disruptions in the chemokine system can significantly alter the susceptibility and progression of different diseases. For instance, COVID-19 severe cases are associated with uncontrolled receptor-ligand interactions and consequent inflammatory dysregulation, which characterizes the cytokine storm frequently observed in such severe disease cases (69, 70). Recently, CCR5Δ32 deletion was identified as a protective factor in Czech First-Wave COVID-19 subjects (71). Different CCR5-editing techniques are currently available and can be used to test in vitro the impacts of the CCR5 absence in different conditions, simulating the consequences of CCR5Δ32 on the immune system and disease conditions (72, 73). However, it is essential to emphasize that the CCR5-editing in human embryos raises many ethical concerns and may have deleterious consequences (67, 74).
Looking at the desirable effects, CCR5Δ32 protects against HIV infection, since the homozygous state of the variant impairs the proper expression of CCR5, preventing the interaction of CCR5 (the main HIV co-receptor) with the virus on the cell surface, thus avoiding infection of the host (75, 76). As mentioned above, CCR5Δ32-derived molecules (CCR5 truncated proteins) can also have an important protective effect against HIV by sequestrating CCR5 and CXCR4 from cell surface (65, 66). The discovery of this effect was truly relevant because it gives support to the use of CCR5 blockers for the clinical control of HIV infection. The best example of this case is maraviroc, a noncompetitive CCR5 antagonist that prevents the proper interaction between the HIV envelope glycoprotein and the CCR5. Currently, other CCR5 blockers (e.g., cenicriviroc, leronlimab) are being tested to treat HIV infection and other inflammatory conditions, and maraviroc emerges as a potential drug to treat other diseases involving CCR5, especially some types of cancer (77). In Brazil, CCR5 blockers represent a good choice for HIV treatment, since most of the circulating viral strains show CCR5 tropism (78–80). Based on the scenario presented above, Figure 1 shown an alluvial diagram representing the classic outcomes associated with the CCR5Δ32, including “desirable” and “undesirable” effects.
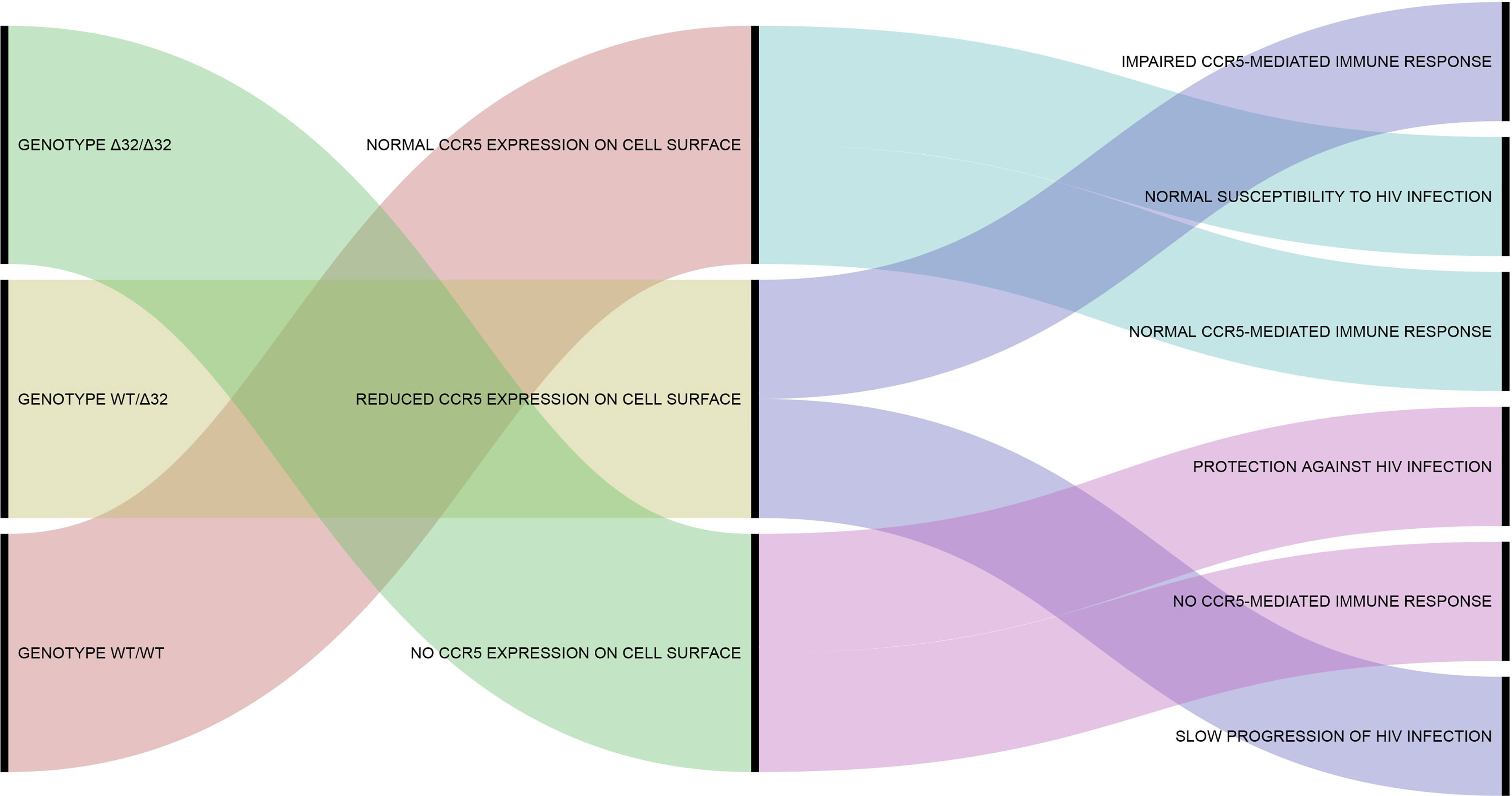
Figure 1 Alluvial diagram representing the classic outcomes associated with the CCR5Δ32. The CCR5Δ32 genotypes are shown in the left part of the diagram. The phenotypic effects of each genotype are shown in the center. The more classical consequences associated with each phenotype are shown in the right part of the diagram. Additional information concerning the phenotypic effects of the CCR5Δ32 on human cells and immune system can be found in previous studies of our group (40, 68, 81). This figure was created using RAWGraphs (https://rawgraphs.io/) (82).
Another major achievement involving CCR5Δ32, and HIV infection was the sustained remission of the infection in the ‘Berlin Patient’, reported in 2009 (83) and confirmed in 2011 (84), and in the ‘London Patient’, reported in 2019 (85) and confirmed in 2020 (86). Both individuals were HIV positive and developed hematological malignant diseases (acute myeloid leukemia and Hodgkin’s lymphoma, respectively), requiring allogeneic hematopoietic stem-cell transplantations. After receiving cell transplantations from CCR5Δ32 homozygous donors, both showed sustained remission of HIV infection. Other cases like Berlin and London patients are being followed up, such as the ‘Düsseldorf patient’ (87). The success of this strategy, although involving few cases, shows that sustained remission of HIV is possible to be achieved and subsequently maintained free of antiretroviral therapy. The Berlin patient, Timothy Ray Brown, passed away on September 29, 2020, due to the recurrence of acute myeloid leukemia, not HIV infection (88, 89). In addition to having collaborated enormously to advance research involving HIV, T. R. Brown created the Timothy Ray Brown Foundation and contributed significantly to the field of HIV/AIDS research, with a big and admirable impact on global society as an HIV activist (89–91).
Currently, it is known that the influence of CCR5 and CCR5Δ32 goes beyond protection against HIV infection and is much broader than previously believed, influencing the susceptibility and outcome of different conditions, such as other different viral, bacterial, and parasitic diseases (40, 92), as well as non-infectious inflammatory conditions (93–96). This occurs because the lack of CCR5 expression, in humans naturally due to CCR5Δ32, interferes with multiple aspects of inflammatory responses, including expression of immune system genes, levels of inflammatory markers, and activity of immune cells (97–103). On the other hand, now looking at the undesirable aspects of CCR5Δ32, this genetic variant increases the risk of serious complications caused by the West Nile virus and Tick-borne encephalitis virus (104–109).
Although Brazilians form a population of more than 210 million individuals, genetic studies in this population are still limited, with most genetic studies focusing on populations with European ancestry (6, 9). The Brazilian population can serve as a study case to understand the impact of genetic admixture on the frequency of genetic variants, such as CCR5Δ32, and its impacts on different conditions and pharmacogenomics (7). Understanding the extent to which the CCR5Δ32 variant influences the health of different populations is critical since it indicates which individuals and ethnic groups are more likely to benefit from therapies focused on modulating CCR5 in the context of cancer, infections, and inflammatory diseases. Focusing on HIV, knowing the frequency of CCR5Δ32 in different human populations is the initial step to guide potential new attempts at sustained remission of HIV infection through stem cell transplantation with CCR5Δ32 homozygous genotype. Moreover, it is also essential to understand how CCR5Δ32 impacts the health of the Brazilian population.
Considering that (I) the frequency of CCR5Δ32 is quite varied among Brazilians from different country’s regions and that (II) the role of CCR5Δ32 in various pathological conditions is an emerging topic with several knowledge gaps, the primary objective of this article is to review the effects of the genetic variant CCR5Δ32 on the Brazilian population, considering several diseases and clinical conditions. The secondary objective of this article is to discuss the impacts of a European-derived variant, the CCR5Δ32, on a highly mixed population.
Methods
For the initial selection of articles, the terms “CCR5”, “CCR5 delta 32”, “CCR5Δ32” and “rs333”, used in combination with “Brazil” or “Brazilian”, were searched on PubMed (https://pubmed.ncbi.nlm.nih.gov/). Subsequently, the same search strategy was used on Scientific Electronic Library Online - SciELO (https://scielo.org/). The articles were initially selected based on the title and abstract. Only articles addressing CCR5Δ32 in Brazilian populations were included in this review. Articles published in English and Portuguese were considered in the evaluation, without restriction concerning the date of publication. On some specific occasions, the reference list of selected articles was also used as an additional source of published works involving CCR5Δ32 in the Brazilian population. Additional unstructured searches were performed on PubMed to select the articles cited in the introduction section and additional points of the review.
CCR5Δ32 Frequency in Brazil
A study published in 2016 by Silva-Carvalho and collaborators (49) presented a very complete meta-analysis regarding the CCR5Δ32 frequency in Brazil. In addition to original data from those authors, the meta-analysis included 29 articles reporting the CCR5Δ32 frequency in Brazil, encompassing populations from ten Brazilian States. The study found an overall allelic frequency of 4% in the country (49). The frequencies of the CCR5Δ32 allele in the Brazilian States, including data compiled by Silva-Carvalho et al. (49), are summarized in Figure 2. Henceforward, we expand the information concerning the CCR5Δ32 frequency in Brazil, highlighting studies not included in the meta-analysis by Silva-Carvalho et al. (49), and including data obtained from studies with indigenous populations and quilombola communities, as discussed below.
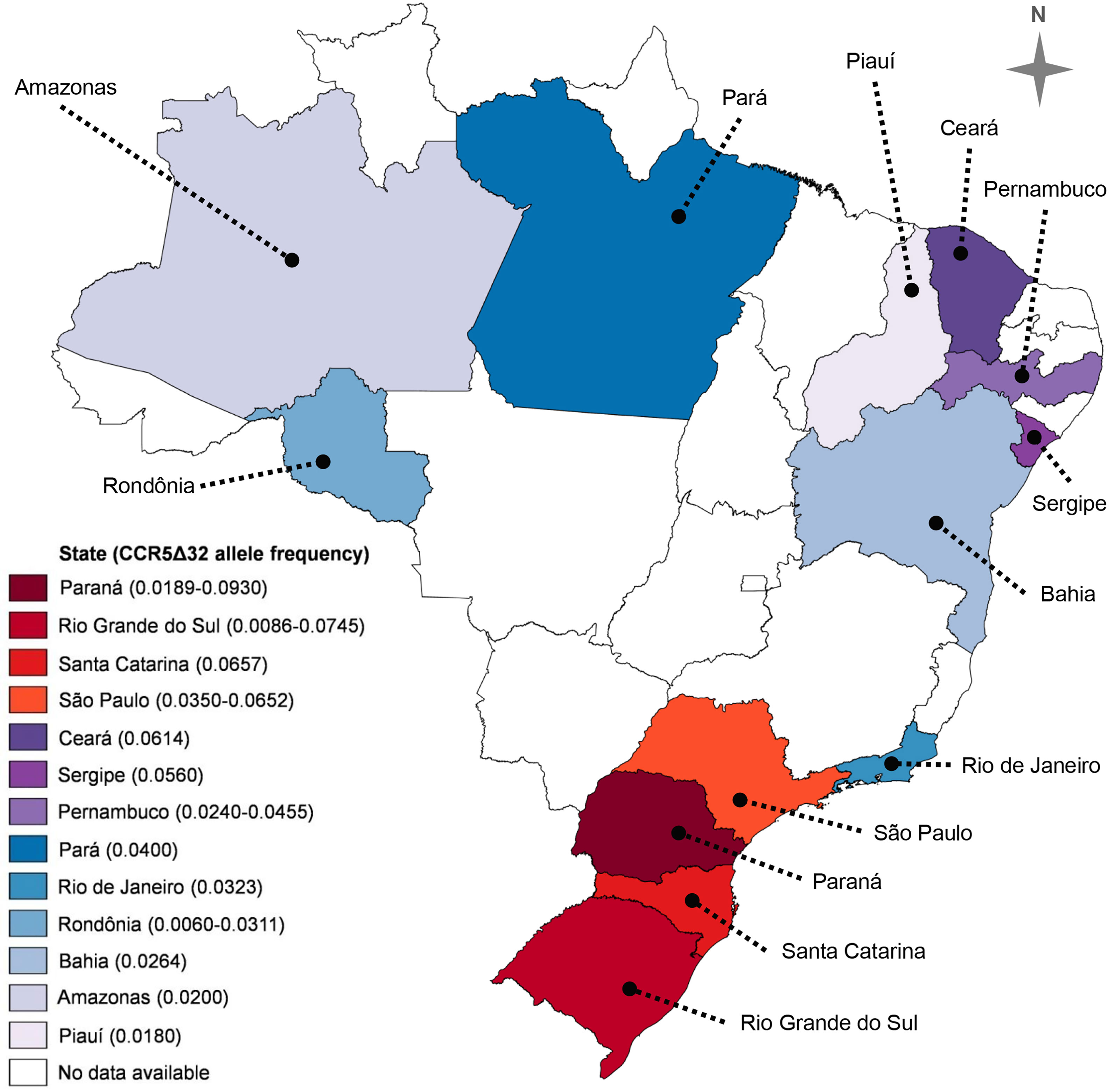
Figure 2 CCR5Δ32 allele frequency in thirteen Brazilian states. Two values in parentheses represent the lowest and the highest frequency observed in a given state. Data from Silva-Carvalho et al. (49), Hüneimeier et al. (110) (Mura population; Amazonas State), Carvalho et al. (41) (Mocambo community; Sergipe State), and Ferreira-Fernandes et al. (111) (Piauí State). The map was created with the help of MapChart (https://mapchart.net/), licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Leboute et al. (112) reported the absence of the CCR5Δ32 allele in a sample of 300 Amerindians from four indigenous populations of the Brazilian Amazon region, namely: Tikuna (n = 191), Baniwa (n = 46), Kashinawa (n = 29), and Kanamari (n = 34). Based on such data, we can argue that, at least until the date of publication of that work, the studied Amazonian tribes probably did not have a significant degree of miscegenation at a level sufficient for the introduction of the CCR5Δ32 allele into those indigenous groups. Alternatively, the allele could already be circulating in the groups, but it may not have been detected due to the small sample size (112).
Carvalhaes et al. (113) also described the frequency of the CCR5Δ32 allele in different ethnic groups of the Brazilian Amazon region, specifically from Pará State. The sample groups investigated were composed of 394 individuals from Belém (capital of Pará), 67 Afro-Brazilian individuals, 89 Amerindian individuals, and 111 Japanese immigrants. The CCR5Δ32 allele was not observed in Amerindian individuals and Japanese immigrants. In the sample of Afro-Brazilian individuals, only one individual carrying the allele in heterozygous was found, with the allele frequency, in this case, being 0.75%. In the sample of random individuals from Belém, one homozygous individual for the gene deletion and 22 heterozygous individuals were found, resulting in a CCR5Δ32 allele frequency of 3.04% (113).
Hünemeier et al. (110) evaluated the frequency of the CCR5Δ32 allele in Native American populations in Brazil and Paraguay: five Amazonian groups (Tiriyo, Mura, Cinta Larga, Gavião, and Zoró); a group from the Paraguayan Gran Chaco (Lengua); one from the Paraguayan forest (Aché); and one from southern Brazil (Kaingang). The CCR5Δ32 allele was found only in two groups: Mura (2%) and Kaingang (3%). The presence of the CCR5Δ32 allele in the samples of these two groups may be due to gene flow, which is explained by previous data showing that both populations have a degree of miscegenation. Thus, the CCR5Δ32 allele may have been introduced in American-native populations due to European miscegenation (110).
Vargas et al. (42) investigated the distribution of the CCR5Δ32 allele in individuals from Alegrete, a city in the western region of Rio Grande do Sul State. The population of Alegrete is highly admixed, with the genetic participation of Spanish, Portuguese, African, and Amerindian peoples. In the study, 103 healthy and unrelated individuals were analyzed, being divided into ‘white’ (n=59), ‘brown’ (n=31), and ‘black’ (n=13). No CCR5Δ32 homozygous individuals were found, and the frequency of heterozygotes was 14% in whites, 13% in browns, and 8% in blacks. Allele frequencies were 6.8%, 6.4%, and 3.8%, respectively (42). In Brazil, the classification of ethnicity performed by the government agency Instituto Brasileiro de Geografia e Estatística (Brazilian Institute of Geography and Statistics) is based on skin color, and for this reason many Brazilian studies classify individuals using this criterion. Alternatively, ‘white’ individuals can be classified as Caucasians, and ‘brown’ and ‘black’ can be classified as non-Caucasians.
Ferreira-Fernandes et al. (111) analyzed the CCR5Δ32 frequency in a sample of the population of the Piauí State. The sample consisted of 223 elderly individuals from the Network of Research on Frailty in Elderly Brazilians. The CCR5Δ32 allele was found only in heterozygous in the sample, with an allele frequency of 1.8%. In order to have a more robust investigation, the sample was also stratified according to sex and age (dividing the groups into individuals below or above 73 years old), but the frequencies were not statistically different between groups, ranging from 1.5% to 2.3%. The general CCR5Δ32 frequency observed is in accordance with other data presented by groups also from northeastern Brazil (111).
Carvalho et al. (41) evaluated the CCR5Δ32 frequency in three quilombola communities in the states of Sergipe (Mocambo community) and Bahia (Rio das Rãs and São Gonçalo communities). The groups were founded about 150 years ago by individuals from Sub-Saharan Africa and/or their descendants. The study evaluated individuals born in quilombola communities and recent immigrants, with a total of 100 inhabitants from Rio das Rãs, 71 from Mocambo, and 53 from São Gonçalo. In these communities, 28 were recent immigrants from Rio das Rãs, 18 from Mocambo, and 15 from São Gonçalo. Thus, the total sample size was 224 individuals: 163 born in the quilombos and 61 recent immigrants. In most cases, the oldest person in each family was chosen to participate in the study. The CCR5Δ32 allele was found in the three communities evaluated, but only in heterozygosis, with allele frequencies of 5.6% in Mocambo, 1% in Rio das Rãs, and 0.9% in São Gonçalo. According to the authors, the differences in allele frequencies can be due to several factors, including different proportions of parental populations in the founder’s individuals, a founder-effect, and different patterns of inter-ethnic contact (41).
Finally, we summarized in Figure 2 the frequencies of CCR5Δ32 allele in thirteen Brazilian States, according to data of ten states compiled by Silva-Carvalho et al. (49), and the frequencies observed by Hüneimeier et al. (110) in the Mura population (Amazonas State), by Carvalho et al. (41) in individuals from Mocambo community (Sergipe State), and Ferreira-Fernandes et al. (111) in individuals from Piauí. To the best of our knowledge, there are no data available in the literature on CCR5Δ32 in the other Brazilian States.
CCR5Δ32 in Infectious Diseases
CCR5 plays a critical role in the regulation of the immune response against infectious agents, controlling the traffic of immune cells [e.g., Natural Killer (NK) and T-regulatory (Treg) cells] towards inflammation sites. For instance, a recent study with mice showed that CCR5 has a pivotal role in the recruitment of NK cells to the kidney allowing an adequate neutrophil activity during systemic Candida albicans infection, acting as a fundamental molecule for a proper immune response. The absence of CCR5 expression resulted in uncontrolled inflammation and increased renal damage in face of C. albicans infection (114). Also, Treg cells play a fundamental role in resolving inflammatory conditions, providing an immunosuppressive activity. During infection by different pathogens (e.g., Schistosoma spp.), the poor recruitment of Treg cells to the inflammation sites due to CCR5 absence causes uncontrolled inflammation and related tissue damage (40, 115). On the other hand, during Rocio virus infection, the CCR5 absence was associated with reduced brain inflammation and better prognosis in animals (116). Taking together, imbalances in the CCR5-mediated immune responses due to CCR5Δ32 can cause both reduced and exacerbated inflammation, depending on the type of pathogen responsible for the infection (e.g., fungus, bacteria, virus), the infection site, or the immune cell type affected by the lack or reduction of CCR5 expression (40). In this context, studies addressing CCR5Δ32 and viruses in the Brazilian population will be discussed here, including HIV, Human T-lymphotropic virus (HTLV), Dengue, Influenza A, Hepatitis C virus (HCV), Hepatitis B virus (HBV), and Human papillomavirus (HPV).
As explained in the introduction section, CCR5Δ32 exerts its protective effect against HIV infection through two mechanisms: reduced expression of the CCR5 gene (gene-dosage effect; probably the most important mechanism) (60, 63) and sequestration of CCR5 and CXCR4 from the cell surface (65, 66). Many studies that evaluated CCR5Δ32 in the Brazilian population corroborated the protective effect of the variant on susceptibility or clinical aspects of HIV infection (e.g., 117–120), although other studies have not evidenced these effects, in some cases probably due to the small sample size (e.g., 121, 122). The main results of the studies involving CCR5Δ32 and HIV infection in Brazil are detailed in Table 1.
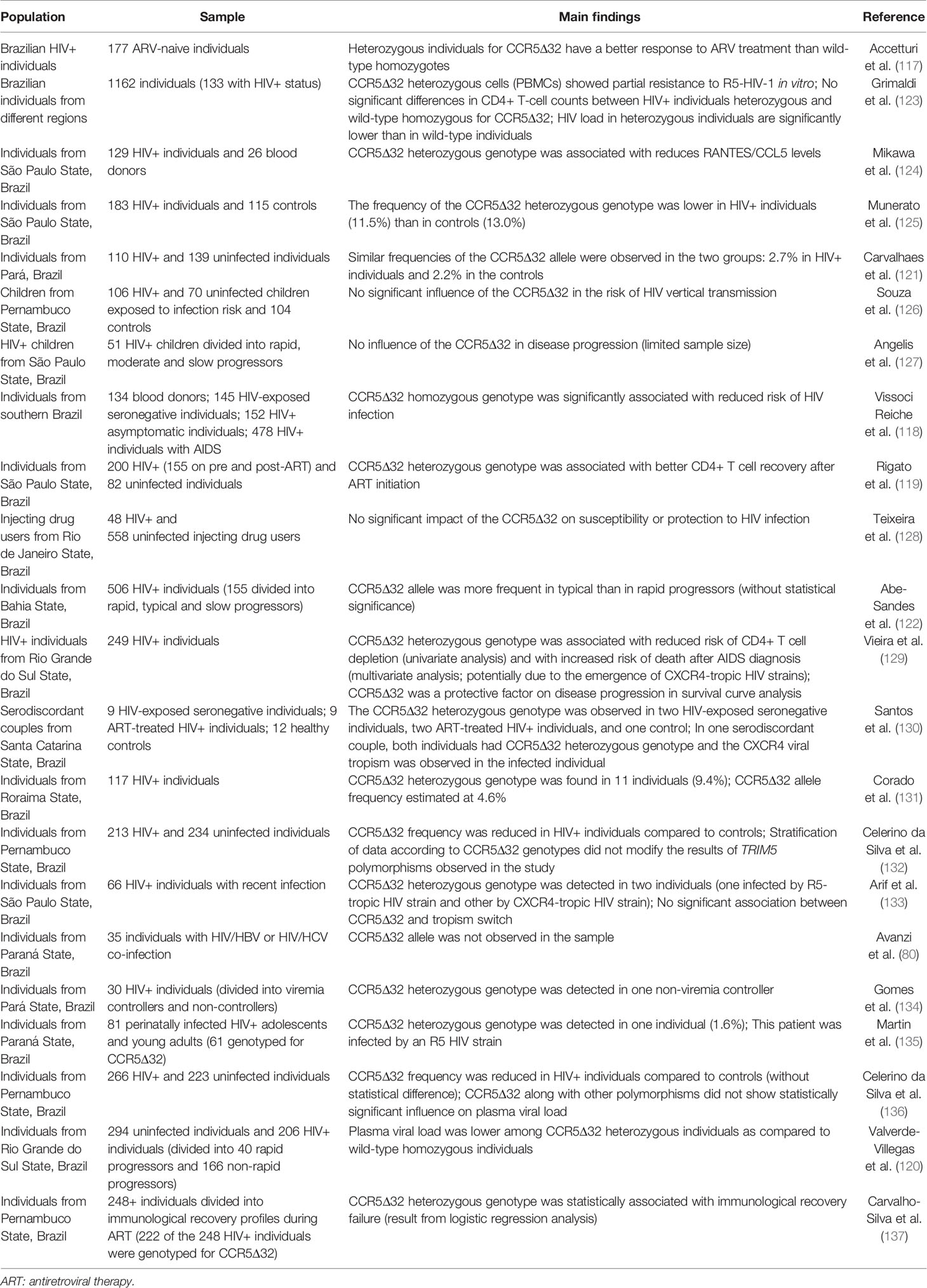
Table 1 Impacts of the CCR5Δ32 on HIV infection.
Experimental evidence indicated that the course of HTLV (type 1 and 2) infection and HIV/HTLV co-infection may be affected by CCR5 expression patterns, which can be modulated by such viruses (138, 139). The CCR5 and its ligands can also influence the course of Dengue infection (140, 141). CCR5Δ32 was associated with an increased risk of fatal Influenza virus infection in Spanish individuals (142). However, CCR5Δ32 has a limited impact on these infections in the Brazilian population. Studying HTLV-1 infection, no statistically significant association was found between CCR5Δ32 and susceptibility or presence/absence of a symptomatic infection (143). Only one study was found regarding this evaluation in a non-Brazilian population. Hisada et al. (144) investigated the CCR5Δ32 frequency in Jamaican HTLV-1-infected individuals and healthy controls. However, the frequency found was too low to further conclusions. That said, no study found an association between the variant and HTLV-1 infection (144). Also, no statistically significant association was observed when the frequencies of CCR5Δ32 were compared between severe Dengue cases and controls (145). A similar study carried out in an Australian population also found no association between the CCR5Δ32 allele and DENV infection (146). The CCR5Δ32 was not associated with hospitalization in individuals infected by Influenza A virus (2009 pandemic H1N1 strain) (147). Subsequently, a study addressing the same virus also reported no significant effect of CCR5Δ32 on H1N1 infection severity (148). A study conducted in a Spanish population identified an association between the CCR5Δ32 allele and fatality due to Influenza A (H1N1) infection (142). Also, an association of the variant with disease severity was observed in a Canadian population (149). Therefore, further studies evaluating the role of this polymorphism in Influenza virus infection are needed.
HCV and HBV are associated with the development of hepatocarcinoma and other liver diseases (150). CCR5 could affect both susceptibility to these viruses and associated diseases due to its regulatory role in inflammatory reactions. Our group evaluated the influence of CCR5Δ32 on susceptibility to HCV infection and HCV/HIV co-infection. In the same study, we also accessed the potential impact of the CCR5Δ32 on HCV-related fibrosis, cirrhosis, and hepatocarcinoma. In total, 1352 individuals were included in the study. No statistically significant associations of CCR5Δ32 with the evaluated criteria were observed (151). Looking at data reported in other populations [see discussion in reference (151)], we highlight that the association between the CCR5Δ32 variant and HCV infection can show important biases in some populations, and other studies corroborate our results showing a lack of association between the variant and HCV infection. Importantly, our work had the largest sample evaluated in the context of HCV infection (151).
More recently, we evaluated the influence of CCR5Δ32 on susceptibility to HBV infection and HBV/HIV co-infection in a study involving 1113 individuals. We found no significant effect of CCR5Δ32 on susceptibility to HBV mono-infection. On the other hand, the CCR5Δ32 allele exerted a protective influence on HBV/HIV co-infection. Of note, this result was potentially due to the known protective effect of CCR5Δ32 on HIV infection (92). In a study in the Indian population, the heterozygous genotype (WT/Δ32) was associated with a higher susceptibility to HBV infection, whereas in a study in the Iranian population, the variant was a protective factor against the infection (152, 153). Other studies carried out in different populations reported a lack of association between HBV infection and the CCR5Δ32 variant (154–156), which is in agreement with the major finding observed in our previous study (92).
HPV is strongly associated with the development of cervical cancer (157) and it was suggested that CCR5 could play a role in the context of HPV infection and related diseases. Nevertheless, Mangieri et al. (158) observed no significant effect of CCR5Δ32 on susceptibility to the infection or cervical lesions (158). Also, the CCR5Δ32 was not associated with infection by a particular HPV genotype (159). In contrast, in a Swedish population, the homozygous genotype for the variant was associated with an increased risk of HPV infection (160). Given the limited amount of data and the contradictory results concerning the involvement of CCR5 in HPV infection, further evaluation concerning the potential role of the CCR5Δ32 variant in the context of HPV infection and related diseases in Brazilian and other populations are needed.
The influence of CCR5Δ32 on parasitic diseases was also investigated in the Brazilian population, including Chagas disease, leishmaniasis, and toxoplasmosis. CCR5 can have two opposite effects on Chagas disease, a disease caused by Trypanosoma cruzi infection. CCR5 mediates the control of acute infection, assuming a favorable role for the host. In opposition, the increased expression of CCR5 during Chagas disease is associated with exacerbated inflammation and related cardiac complications (161). Thus, the levels of CCR5 expression are critical in the outcome of Chagas disease. However, two other studies found no association between the CCR5Δ32 variant and cardiac or digestive manifestations on chronic Chagas disease (162, 163). In a Peruvian population, the frequency of the Δ32 allele was not high enough to allow an analysis of association with T. cruzi infection, and a study with individuals from Venezuela did not find an association of the variant with the presence of disease symptoms (164, 165). Therefore, the potential CCR5Δ32 allele role in Chagas disease is still under discussion.
Brajão de Oliveira et al. (166) and Ribas et al. (167) reported no statistically significant difference between Leishmania-infected individuals and controls concerning CCR5Δ32 frequencies (166, 167). In the study performed by Brajão de Oliveira et al. (166), the CCR5Δ32 allele carriers showed a less severe spectrum of clinical manifestations, but without statistical significance (166). Ribas et al. (167) observed a higher frequency of the CCR5Δ32 polymorphism among a subgroup of patients with recurrent lesion, but this specific result was based on an exceedingly small cohort (167). Also, a study performed in a Pakistani population showed no association between the CCR5Δ32 variant and cutaneous leishmaniasis (168).
The CCR5Δ32 wild-type genotype in association with AA or AG genotypes (from the CCR5 rs1799987 polymorphism, an intron A/G SNP) was associated with increased risk of ocular toxoplasmosis, potentially due to the persistent CCR5-mediated inflammation in individuals with normal CCR5 expression (169). Also evaluating Brazilians, Vallochi et al. (170) found no association between the CCR5Δ32 and ocular toxoplasmosis (based on a brief description; detailed data not described by such authors) (170). No other studies evaluating the role of this variant in the context of ocular toxoplasmosis in non-Brazilian populations were found.
Based on the studies discussed above, apart from the protective effect of CCR5Δ32 on HIV infection, the impacts of CCR5Δ32 on viral and parasitic infections in Brazilian populations seem quite limited (details of each study are presented in Table 1 and Table 2). However, considering the recognized role of CCR5 in the regulation of inflammation, it is possible that potential influences of CCR5Δ32 on non-HIV infections have not been detected due to the small number of studies carried out in Brazil on these topics, many of them involving a small sample size.
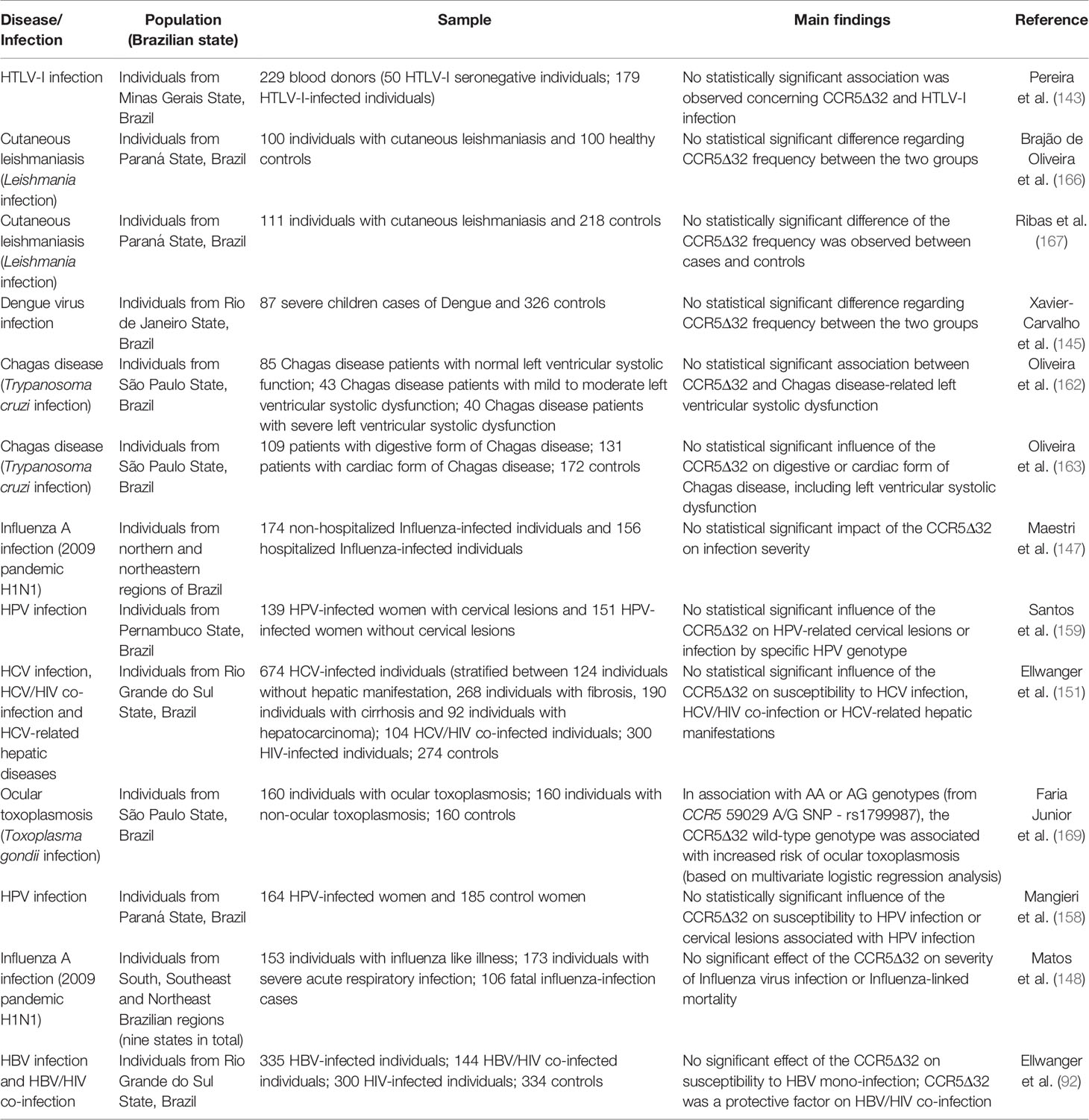
Table 2 Impacts of the CCR5Δ32 on infectious diseases.
Finally, the impact of the CCR5Δ32 on fungal infections is unknown in Brazilian populations and quite sparse in other human populations, and therefore research in this field is needed. Of note, Brazil is affected by several endemic mycoses, such as Dermatophytosis, Paracoccidioidomycosis, Histoplasmosis, and Cryptococcosis, among others (171). Understanding whether and how the CCR5Δ32 influences the susceptibility or clinical progression of these diseases can provide insights into the potential use of CCR5-based therapies for these diseases.
CCR5Δ32 in Inflammatory Conditions
Considering the critical role of CCR5 in the regulation of the inflammatory response, several authors have been investigating the effect of CCR5Δ32 on conditions that have their susceptibility or clinical course affected by different types (e.g., systemic, local) and intensity of inflammation. In this topic, we review the role of CCR5Δ32 on the following inflammatory diseases or inflammation-related clinical conditions: multiple sclerosis, systemic lupus erythematosus, preeclampsia, rheumatoid arthritis, juvenile idiopathic arthritis, periodontitis, osteomyelitis, transplant rejection, and sickle cell disease. Details of each study are described in Table 3 and discussed below.
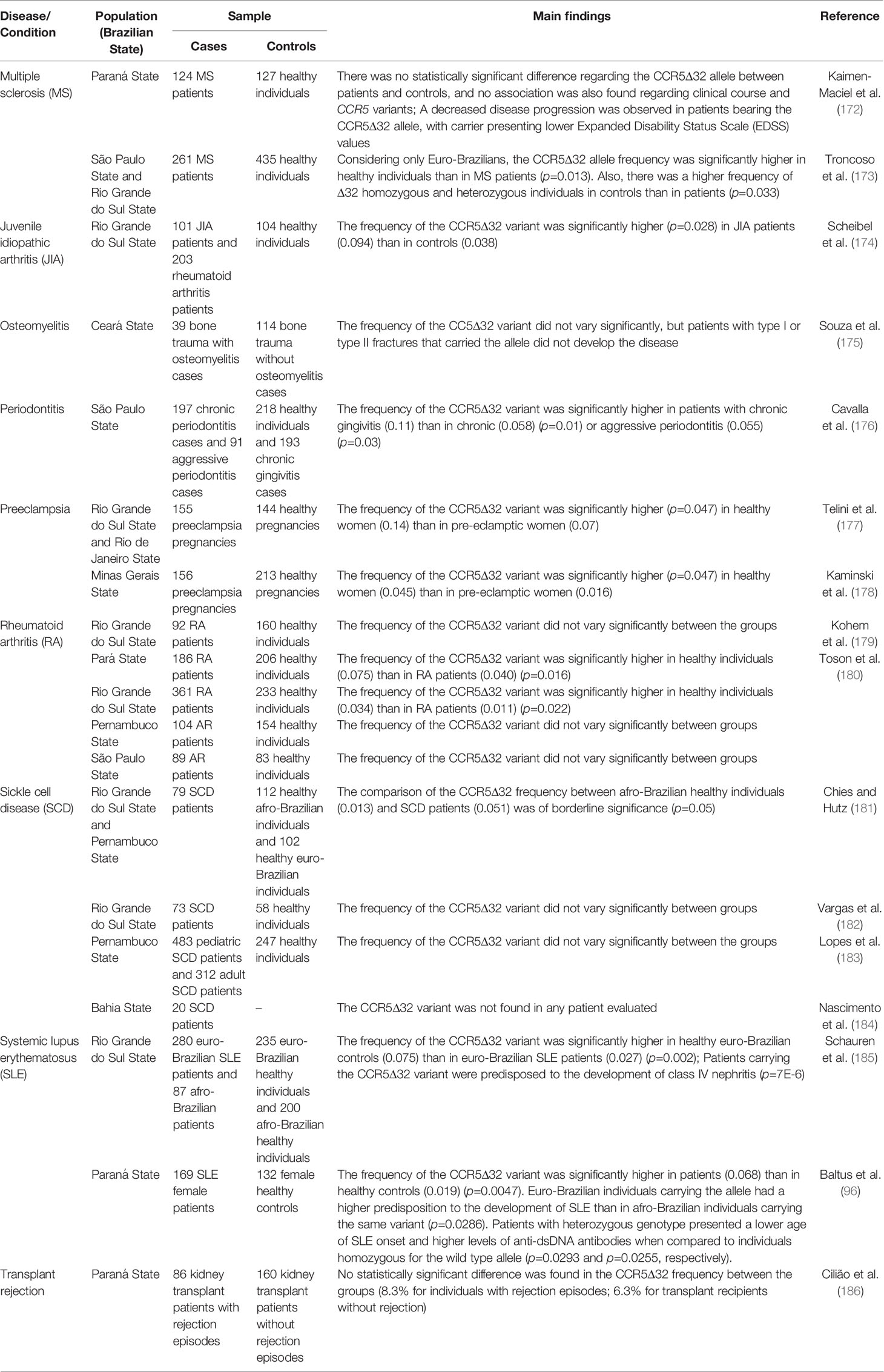
Table 3 Impacts of the CCR5Δ32 on inflammatory conditions.
Multiple sclerosis is an autoimmune, chronic, and inflammatory disease showing heterogeneity in clinical findings. Chemokines and chemokine receptors are molecules involved in the pathogenesis of multiple sclerosis (172, 194), and the CCR5Δ32 can influence different aspects of this disease, as shown in studies with non-Brazilian individuals (195–197). A meta-analysis carried out in 2014 evaluated the role of this variant in multiple sclerosis in different populations, and concluded that the CCR5Δ32 is not associated with susceptibility to the development of multiple sclerosis in Europeans, calling attention to the need for further studies involving other populations (198). In Australian individuals, this variant also did not show a protective role to multiple sclerosis (199). However, other studies have shown an association of the Δ32 allele with treatment response, disease severity, and susceptibility to multiple sclerosis (196, 200–202). In Brazil, only two papers explored the possible impact of the CCR5Δ32 on multiple sclerosis. Based on magnetic resonance imaging, Kaimen-Maciel et al. (172) observed a decreased disease progression in patients bearing the CCR5Δ32 allele (172). Subsequently, Troncoso et al. (173) described a statistically significant higher CCR5Δ32 allele frequency in Euro-Brazilian controls (7.4%) compared to Euro-Brazilian patients (3.3%), suggesting a protective role of the variant on the development of multiple sclerosis. Besides, the frequency of the CCR5Δ32 was higher in Euro-Brazilian patients with progressive multiple sclerosis than Euro-Brazilian patients with relapse remitting multiple sclerosis (173). Both studies carried out in Brazil show that the CCR5Δ32 variant can influence both the susceptibility and the clinical outcome of multiple sclerosis.
Systemic lupus erythematosus is a chronic inflammatory autoimmune disease characterized by the large production of autoantibodies, triggering generalized tissue damage. This disease has different clinical manifestations and a complex genetic influence, and chemokines and their receptors, such as CCR5, are implicated in the pathogenesis of lupus (96, 185, 203, 204). The CCR5Δ32 variant has already been studied in this context, being previously associated to protection against lupus development and, albeit in a contradictory manner, this polymorphism was also associated to susceptibility to nephritis in lupus patients (203, 204). In Brazil, two studies evaluated the CCR5Δ32 variant in lupus.
Schauren et al. (185) investigated the role of the CCR5Δ32 in healthy patients and controls of Rio Grande do Sul State (185). A lower frequency of the CCR5Δ32 allele was found in Euro-Brazilian patients (2.7%) compared to Euro-Brazilian controls (7.5%), suggesting a protective role of the variant against the development of systemic lupus erythematosus. However, in the same study, patients with the CCR5Δ32 allele had a greater predisposition to the development of class IV nephritis than patients without the allele, which suggests a more severe clinical outcome associated with the genetic variant (185).
Baltus et al. (96) evaluated the frequencies of the CCR5Δ32 in patients and controls in the Paraná State, also southern Brazil. Unlike the first study, the frequency of the CCR5Δ32 allele was statistically higher in patients (6.8%) than in controls (1.9%), suggesting the variant as a risk factor for systemic lupus erythematosus. Also, by stratifying the sample according to ethnicity, the researchers identified that Euro-Brazilian individuals carrying the CCR5Δ32 were more likely to develop systemic lupus erythematosus than Afro-Brazilian patients carrying the variant. In another analysis of the study, CCR5Δ32 carriers had a lower age of systemic lupus erythematosus onset and higher levels of anti-dsDNA antibodies. Thus, the CCR5Δ32 allele was associated with increased susceptibility to the development of systemic lupus erythematosus and severity in clinical outcomes (96). Studies performed in different populations have found no association between the variant and the development of systemic lupus erythematosus (205–208). Such divergence involving the results mentioned above deserves attention and, therefore, more studies in other populations are required.
Preeclampsia is a hypertensive gestational complication and an important cause of maternal-fetal mortality in Brazil. Relevant clinical findings of the disease, such as edema and proteinuria after the 20th week of pregnancy, are intricate with an excessive inflammatory process and endothelial dysfunction. In preeclampsia, increased systemic production of pro-inflammatory chemokines was observed, highlighting the role of the chemokine-ligand system in this condition (177, 178, 209). Two studies evaluating the CCR5Δ32 variant in preeclampsia were carried out in Brazil, both published by our group, but evaluating samples from different Brazilian regions. Firstly, Telini et al. (177) evaluated the frequency of the CCR5Δ32 in Brazilian women who developed preeclampsia and women who did not develop this condition during their pregnancies. The group of healthy women had a higher frequency of the CCR5Δ32 allele (14%) when compared to the group of women who developed preeclampsia (7%). The analysis revealed a protective role of the variant on preeclampsia development (177). More recently, Kaminski et al. (178) also investigated the role of CCR5Δ32 in women who developed preeclampsia and in women with healthy pregnancies (178). In accordance with the results of Telini et al. (177), healthy pregnant women also showed an increased CCR5Δ32 allele frequency (4.5%) compared to the group of pregnant women with preeclampsia (1.6%). Thus, the study corroborated the protective role of the CCR5Δ32 variant on preeclampsia development, endorsing the hypothesis that a reduced inflammatory millieu may contribute to a lower risk of developing preeclampsia (177, 178). A study conducted in a Turkish population found similar results, strengthening the conclusion here presented (210).
Rheumatoid arthritis is a systemic autoimmune disease characterized by progressive damage to the joints caused by chronic inflammation in the synovial fluid. Given the intense migration of immune cells to the inflammation sites, the role of CCR5 in rheumatoid arthritis appears to be of great importance (179, 180). In Brazil, two studies investigating the role of the CCR5Δ32 variant in rheumatoid arthritis were published. Kohem et al. (179) evaluated the frequency of the allele in healthy patients and controls from the Rio Grande do Sul State, and no statistically significant difference was found between the groups. Of note, the sample group was relatively small, with 92 patients and 160 healthy controls (179). Toson et al. (180) performed a similar study but evaluating the frequency of the CCR5Δ32 variant in different Brazilian populations, considering four different regions (south, southeast, northeast, and north). Two of the four sample groups, from southern and northern regions, showed a statistically significant difference between rheumatoid arthritis patients and healthy controls (4% vs. 7.5%; 1.1% vs. 3.4%, respectively), being precisely the groups with the largest sample sizes. The difference concerning the northeast region sample was not statistically significant but followed a similar trend to the groups in southern and northern. Only the southeastern sample deviated from the trend, with the small sample size possibly being the reason for the lack of statistical association. In sum, the study suggests a protective role for the CCR5Δ32 variant against the development of rheumatoid arthritis (180). A meta-analysis carried out in 2012 concluded that the variant may play a role in protection to rheumatoid arthritis in European populations, corroborating the data found in Brazil (211).
Juvenile idiopathic arthritis is a chronic inflammatory condition characterized in the synovial joints of young people up to 16 years of age (174, 212). Scheibel et al. (174) investigated the potential association of the CCR5Δ32 variant with juvenile idiopathic arthritis subtypes in a sample from Porto Alegre, southern Brazil. A statistically significant difference was found between patients (9.4%) and healthy controls (3.8%), especially considering the group of patients of the systemic juvenile idiopathic arthritis subtype (25%). The researchers conclude that the CCR5Δ32 variant, although not a risk factor for the development of juvenile idiopathic arthritis, contributes to the progression and clinical status of patients (174). Interestingly, the meta-analysis previously mentioned (211) also explored the role of the Δ32 allele in juvenile idiopathic arthritis, and concluded that the variant was a protective factor for this condition as well (211). A further study comprising children from different populations found an association between the heterozygous genotype and mild disease course, but no influence on susceptibility to disease development (213). That said, these controversial results evidence the importance of novel studies investigating the CCR5Δ32 variant in juvenile idiopathic arthritis.
Periodontitis is an oral disease characterized by a chronic infection accompanied by inflammatory processes, causing irreversible and progressive destruction of dental support structures. The CCR5-mediated immune responses affect multiple aspects of periodontitis. For instance, not only CCR5 and its ligands are important in the context of disease protection, but also influence periodontal destruction and bone resorption (176, 214–217). Cavalla et al. (176) investigated the CCR5Δ32 variant and its possible influence on periodontitis development. The CCR5Δ32 allele was significantly more frequent in individuals classified in the group of chronic gingivitis (11.1%) than in individuals with chronic periodontal disease (5.8%) or aggressive periodontal disease (5.5%). This result suggests a protective role of the variant concerning periodontitis (176). Other studies carried out in Taiwan and Germany found no association between the variant and periodontitis (218, 219). Considering the conflicting results, it is interesting to carry out further studies in other populations to better understand the role of CCR5 in the development of periodontitis.
Osteomyelitis is an infectious-inflammatory condition that can occur after bone trauma often following Staphylococcus aureus infection (175, 220). Souza et al. (175) evaluated the CCR5Δ32 frequency in patients who were admitted to a hospital in Fortaleza, northeastern Brazil, with bone trauma. The patients were prospectively studied to assess a possible development of osteomyelitis. There was no statistically significant difference between individuals who developed and those who did not develop the disease, but all patients with closed fractures (type I or type II) and who carried the CCR5Δ32 variant did not developed the condition. The researchers conclude that the lack of statistical significance observed in their study was probably due to the low sample size (175). No other studies regarding the potential role of the CCR5Δ32 in osteomyelitis were found in the literature.
The immune response and inflammatory processes that occur after an organ transplant are critical in the process of tissue rejection. Genetic variants related to the immune system can therefore influence the response to transplantation (186, 221–223). Studies carried out in non-Brazilian populations observed no association between the CCR5Δ32 allele and kidney transplant rejection (224–228). A study in a multicentric sample from Europe showed a higher survival rate after kidney transplantation in individuals with the CCR5Δ32 homozygous genotype (222). In Brazil, Cilião et al. (186) evaluated the CCR5Δ32 frequency in transplanted individuals who had episodes of rejection comparing to individuals who did not have such episodes. A sample of 246 patients was collected in a referral hospital in Londrina, Paraná State. However, the frequency of the CCR5Δ32 variant did not vary significantly between the groups (186).
Sickle cell disease is an inherited disorder caused by a single nucleotide substitution in the beta-globin gene. This mutation originated in Africa and is, therefore, more common in African populations and Afro-descendants. Sickle cell disease can be understood as a chronic inflammatory condition, which may be the cause of associated secondary complications. In this sense, high levels of inflammation in sickle cell disease patients are related to disease morbidity (181–184). A study in a population from Egypt found no association between the variant and sickle cell disease (229). In Brazil, four studies investigated the influence of the CCR5Δ32 variant in sickle cell disease, all detailed below.
Chies and Hutz (181) assessed the potential role of the CCR5Δ32 in severe and recurrent infections that could contribute to differentiated survival of sickle cell anemia patients. The study involved individuals from different ethnic groups and the frequencies of the CCR5Δ32 allele found were 4.4% in Euro-Brazilian controls, 1.3% in Afro-Brazilian controls, and 5.1% in sickle cell anemia patients. When comparing these frequencies between the different groups, no statistically significant difference was found. However, it is important to note that, considering the same ethnic background of the groups of patients and Afro-Brazilian controls, a difference in the allele frequency was evidenced, being the CCR5Δ32 allele three times more present in the group of sickle cell anemia patients. Given the low frequency of the allele in the sample of Afro-Brazilian controls, a 3-fold increase in the group of patients is quite important. The researchers suggested that the CCR5Δ32 allele was more frequent in the group of patients for conferring some advantages concerning the clinical course of the disease (181). As mentioned previously, sickle cell anemia can be considered a chronic inflammatory disease (93), and patients with the CCR5Δ32 allele would benefit from developing inflammatory responses at low levels. According to this hypothesis, the CCR5Δ32 allele was associated with an improvement in the general health status of the patients (93, 181).
Subsequently, Vargas et al. (182) evaluated CCR5Δ32 in sickle cell anemia patients from Porto Alegre, Rio Grande do Sul State. No statistically significant difference was observed in the study but, interestingly, the CCR5Δ32 allele was present only in the group of patients with a severe clinical course (when the pain rate was considered). Such data may indicate a trend towards the development of a severe clinical course associated with the CCR5Δ32 allele in sickle cell anemia patients (182). Lopes et al. (183) compared the CCR5Δ32 frequencies of two groups of patients (pediatric and adult) and between sick adults and healthy controls from Pernambuco, northeastern Brazil. There were no statistically significant differences in any of the comparisons made in the study (183). Finally, Nascimento et al. (184) evaluated the CCR5Δ32 frequency in sickle cell anemia patients from Bahia State. However, the CCR5Δ32 allele was not found in the study (184).
CCR5Δ32 in Cancer
Chemokines and chemokine receptors have fundamental participation in both antitumor response and pathogenesis of cancer. The migration of regulatory immune cells to tumor sites can create an immunosuppressor environment proper for cancer development. Also, cancer cells can subvert the anti-tumor action of chemokine-ligand interactions (187–191). Of note, CD4+ T cells are important modulators of the immune response, acting as drivers for the action of effector cells. Some CD4+ regulatory T cells express the CCR5 molecule, being this a key receptor of the cellular response against tumor development. The presence of the CCR5Δ32 variant can impair the action of CCR5+/CD4+ T cells, influencing the risk of cancer development. In brief, chemokine receptors can assume multiple roles in different tumoral processes, and more investigation is needed to unravel the connections between CCR5 and cancer (101, 192). Two meta-analyses published in 2014 evaluated the possible role of the Δ32 allele in cancer. Ying et al. (230) found no association of the variant with risk of tumorigenesis, while Lee et al. (205) found an association of the allele with susceptibility to cancer in Indians, specifically concerning breast cancer (205, 230). Further studies found associations of the CCR5Δ32 variant with improved metastasis-free survival in breast cancer patients and, contradictorily, also with an increased risk for developing breast cancer (231–233). In Brazil, the possible role of the CCR5Δ32 variant in cancer has been addressed (Table 4) and the available data will be presented below.
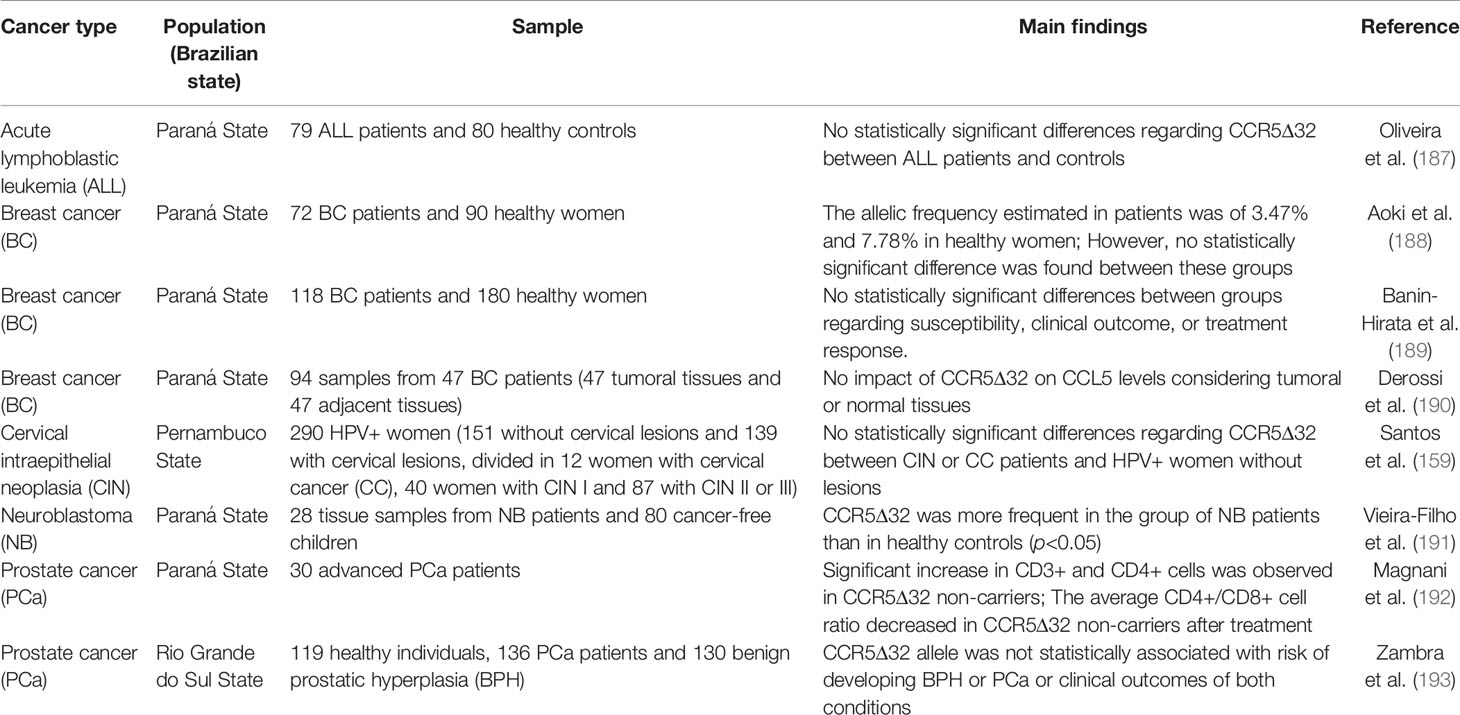
Table 4 Impacts of the CCR5Δ32 on cancer.
The action of CD8+ cytotoxic T cells is important in the antitumor immune response. The use of immunomodulators in antitumor treatment is increasingly common, with carboxymethyl-glucan (CM-G) being one of the best-described immunostimulators (192, 234). Magnani et al. (192) evaluated the CD3+, CD4+ and CD8+ cell populations of patients with advanced prostate cancer and compared this data with the CCR5 genotype, associating it with the administration of oral CM-G for 28 days. The CCR5Δ32 variant was found only in a heterozygous genotype, in six patients, at an allelic frequency of 10%. Five patients reported a family history of prostate cancer, two of whom had affected first-degree relatives. Both patients carried the CCR5Δ32 allele. In general, CCR5Δ32 non-carriers had higher counts on CD3+ and CD4+ cells when comparing respectively after and before treatment with CM-G, as well as higher counts of CD8+ cells when comparing to CCR5Δ32 carriers only after treatment with CM-G. In addition, the average CD4+/CD8+ cell ratio showed a worsened antitumor response after treatment in CCR5Δ32 allele carriers (192). Zambra et al. (193) also evaluated the CCR5Δ32 frequency in Brazilian prostate cancer patients, comparing to individuals affected by benign prostatic hyperplasia and healthy subjects. No association was found considering the variant and risk to both conditions, nor with clinical outcomes (193).
Aoki et al. (188) assessed the CCR5Δ32 frequency in individuals with breast cancer and healthy women. However, no significant difference was observed between groups. The impact of p53 genotypes, a known tumor suppressor gene, together with the CCR5Δ32 genotypes, was also evaluated revealing a higher frequency of individuals with the p53 Arg homozygous genotype and the CCR5Δ32 wild-type genotype amongst controls as compared to patients (188). Banin-Hirata et al. (189) also evaluated whether the CCR5Δ32 variant was associated with susceptibility, response to treatment, and clinical course of breast cancer. No association was found between CCR5Δ32 and the features analyzed (189). In accordance, Derossi et al. (190) did not found an association between the CCR5Δ32 and CCL5 levels in breast cancer (190).
HPV infection is the main cause of cervical cancer. However, factors other than HPV infection, including genetic, immune, and environmental factors, also affect tumorigenesis (159, 235, 236). In this context, Santos et al. (159) evaluated the CCR5Δ32 frequency in HPV+ women with and without cervical neoplastic lesions. No association was found between the variant and the presence of cancer or lesions severity (159).
In addition to the multiple roles of CCR5 in tumorigenesis and antitumor response, this molecule is also an important modulator of neuroinflammation (237–239), potentially affecting the development of brain-related diseases. In this sense, Vieira-Filho et al. (191) found an association between the presence of the CCR5Δ32 allele and susceptibility to neuroblastoma (191). Lastly, Oliveira et al. (187) investigated the role of the CCR5Δ32 variant in acute lymphoblastic leukemia, but no association was found between the variant and the disease development (187). In conclusion, the CCR5 has varied influences in different types of cancer.
Impacts of CCR5Δ32 on a Highly Admixed Population – A Critical Look
At a population level, the effects of CCR5Δ32 on European populations may be different than those potentially observed in highly admixed populations. However, the population-specific effects of CCR5Δ32 are not only due to its frequency, but also due to its interaction with different alleles. There are nine widely known CCR5 haplotypes, which are formed by combinations of eight CCR5 polymorphisms (including CCR5Δ32) and one polymorphism located in the CCR2 gene (40, 70). The impact of the CCR5 haplotypes on HIV disease progression differs between African Americans and Caucasians since the effects of the CCR5Δ32 can be modulated by other alleles heterogeneously distributed among the populations (240). In a broader perspective, this information indicates that the effect of the CCR5Δ32 observed in Europeans (or other non-Brazilian populations) may be modified by further genetic traits circulating in Brazilians, which may also vary in different regions of the country. In fact, the detection of the real effect of CCR5Δ32 on different health and disease conditions in the Brazilian population is not a simple task. Of note, gene-disease association studies performed with admixed populations can be difficult due to differential linkage disequilibrium patterns (241).
Pharmacogenomic approaches, including the use of CCR5 modulators based on the CCR5Δ32 genotyping, must be considered at an individual level, especially in highly admixed populations, where the frequency of polymorphisms may be quite different from those observed in populations with greater genetic homogeneity (7). The CCR5Δ32 genotyping could be considered in pharmacological treatments involving CCR5 blockade in the context of inflammatory diseases or types of cancer. The use of CCR5 modulators in individuals with the CCR5Δ32 genotype probably has a limited effect due to the natural absence of CCR5 expression on the cell surface. Although the number of individuals with this genotype is exceptionally low in an admixed population such as the Brazilian population, the cost-benefit of this strategy must be considered on a case-by-case basis. Despite the limitations, the area of pharmacogenomics involving CCR5Δ32 genotyping is expected to progress in the next years, especially considering the increasing use of CCR5 modulators to treat other diseases not associated with HIV infection. Some important advances have already been made. For instance, the CCR5Δ32 genotyping can help clinicians to predict the progression of human enteroviral cardiomyopathy, also helping the decision making concerning the early use of antiviral interferon-β therapy in such condition (242).
Conclusions
The CCR5Δ32 allele frequency is quite variable in Brazil, being extremely low in some regions (e.g., 0.6% in Rondônia), but high in others (e.g., up to 9.3% in Paraná and 7.4% in Rio Grande do Sul). In Native American populations, the allele is absent or occurs at low frequencies. In Brazil, CCR5Δ32 is not uncommon in non-Caucasian populations, because of the miscegenation that has occurred in the country.
Many studies corroborated the protective effect of the CCR5Δ32 on susceptibility or clinical aspects of HIV infection in the Brazilian population. On the other hand, there is no evidence pointing to a relevant role for CCR5Δ32 on Cutaneous leishmaniasis, Chagas disease, HTLV-1, Dengue virus, Influenza A, HPV, HBV and HCV infections, or HCV-HIV co-infection in Brazilians. Limited evidence indicates a potential involvement of CCR5Δ32 wild-type genotype in ocular toxoplasmosis and a protective effect of the variant on HBV/HIV co-infection.
Considering inflammatory conditions, the CCR5Δ32 can influence both the susceptibility and the clinical outcome of multiple sclerosis. Of note, CCR5Δ32 reduces the risk of preeclampsia and periodontitis development, potentially due to the CCR5Δ32-associated reduced inflammation. Moreover, CCR5Δ32 can reduce the risk of rheumatoid arthritis, but contributes to the progression and clinical status of juvenile idiopathic arthritis patients. CCR5Δ32 can also influence sickle cell anemia-related immune conditions. However, the impact of CCR5Δ32 on systemic lupus erythematosus is controversial. Concerning tumoral development, the CCR5Δ32 has varying influences on the development of different types of cancer, including prostate cancer and breast cancer. It is not possible to generalize the impact of the variant on cancer development, especially in the Brazilian population.
Understanding the real impact of the CCR5Δ32 variant in different conditions is essential to indicate in which diseases the use of CCR5 modulators may be relevant. This knowledge is fundamental for the advancement of CCR5-based therapies, especially in populations with a complex genetic structure. Finally, CCR5Δ32 influences should be assessed within the context of each population, since genetic admixture and interactions with other alleles may alter the expected phenotypic effects attributed to CCR5Δ32.
Author Contributions
BK-L and JE wrote the first version of the manuscript. JC revised and edited the text. All authors contributed to the article and approved the submitted version.
Funding
BK-L receives a fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil). JE receives a postdoctoral fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Programa Nacional de Pós-Doutorado – PNPD/CAPES, Brazil). JC receives a research fellowship from CNPq (Bolsa de Produtividade em Pesquisa - Nível 1A, Brazil) and has research projects funded by Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS, Brazil) and CAPES (Brazil).
Conflict of Interest
JE is Guest Associate Editor of Frontiers in Immunology but he was not involved in editing this article.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Pena SDJ, Di Pietro G, Fuchshuber-Moraes M, Genro JP, Hutz MH, de Souza Gomez Kehdy F, et al. The Genomic Ancestry of Individuals From Different Geographical Regions of Brazil is More Uniform Than Expected. PloS One (2011) 6:e17063. doi: 10.1371/journal.pone.0017063
2. Adhikari K, Chacón-Duque JC, Mendoza-Revilla J, Fuentes-Guajardo M, Ruiz-Linares A. The Genetic Diversity of the Americas. Annu Rev Genom Hum Genet (2017) 18:277–96. doi: 10.1146/annurev-genom-083115-022331
3. Mas-Sandoval A, Arauna LR, Gouveia MH, Barreto ML, Horta BL, Lima-Costa MF, et al. Reconstructed Lost Native American Populations From Eastern Brazil Are Shaped by Differential Jê/Tupi Ancestry. Genome Biol Evol (2019) 11:2593–604. doi: 10.1093/gbe/evz161
4. Castro e Silva MA, Nunes K, Lemes RB, Mas-Sandoval À, Amorim CEG, Krieger JE, et al. Genomic Insight Into the Origins and Dispersal of the Brazilian Coastal Natives. PNAS (2020) 117:2372–7. doi: 10.1073/pnas.1909075117
5. Lins TC, Vieira RG, Abreu BS, Grattapaglia D, Pereira RW. Genetic Composition of Brazilian Population Samples Based on a Set of Twenty-Eight Ancestry Informative SNPs. Am J Hum Biol (2010) 22:187–92. doi: 10.1002/ajhb.20976
6. Giolo SR, Soler JMP, Greenway SC, Almeida MAA, de Andrade M, Seidman JG, et al. Brazilian Urban Population Genetic Structure Reveals a High Degree of Admixture. Eur J Hum Genet (2012) 20:111–6. doi: 10.1038/ejhg.2011.144
7. Suarez-Kurtz G, Paula DP, Struchiner CJ. Pharmacogenomic Implications of Population Admixture: Brazil as a Model Case. Pharmacogenomics (2014) 15:209–19. doi: 10.2217/pgs.13.238
8. Callegari-Jacques SM, Grattapaglia D, Salzano FM, Salamoni SP, Crossetti SG, Ferreira ME, et al. Historical Genetics: Spatiotemporal Analysis of the Formation of the Brazilian Population. Am J Hum Biol (2003) 15:824–34. doi: 10.1002/ajhb.10217
9. Halagan M, Oliveira DC, Maiers M, Fabreti-Oliveira RA, Moraes MEH, Visentainer JEL, et al. The Distribution of HLA Haplotypes in the Ethnic Groups That Make Up the Brazilian Bone Marrow Volunteer Donor Registry (REDOME). Immunogenetics (2018) 70:511–22. doi: 10.1007/s00251-018-1059-1
10. Baptista EA, Abel GJ, Campos J. Internal Migration in Brazil Using Circular Visualization. Reg Stud Reg Sci (2018) 5:361–4. doi: 10.1080/21681376.2018.1526649
11. Ruiz-Linares A, Adhikari K, Acuña-Alonzo V, Quinto-Sanchez M, Jaramillo C, Arias W, et al. Admixture in Latin America: Geographic Structure, Phenotypic Diversity and Self-Perception of Ancestry Based on 7,342 Individuals. PloS Genet (2014) 10:e1004572. doi: 10.1371/journal.pgen.1004572
12. Chacón-Duque J-C, Adhikari K, Fuentes-Guajardo M, Mendoza-Revilla J, Acuña-Alonzo V, Barquera R, et al. Latin Americans Show Wide-Spread Converso Ancestry and Imprint of Local Native Ancestry on Physical Appearance. Nat Commun (2018) 9:5388. doi: 10.1038/s41467-018-07748-z
13. Abe-Sandes K, Silva WA, Zago MA. Heterogeneity of the Y Chromosome in Afro-Brazilian Populations. Hum Biol (2004) 76:77–86. doi: 10.1353/hub.2004.0014
14. Marrero AR, Silva-Junior WA, Bravi CM, Hutz MH, Petzl-Erler ML, Ruiz-Linares A, et al. Demographic and Evolutionary Trajectories of the Guarani and Kaingang Natives of Brazil. Am J Phys Anthropol (2007) 132:301–10. doi: 10.1002/ajpa.20515
15. de Azevedo DA, da Silva LAF, Gusmão L, de Carvalho EF. Analysis of Y Chromosome SNPs in Alagoas, Northeastern Brazil. Forensic Sci Int: Genet Suppl Ser (2009) 2:421–2. doi: 10.1016/j.fsigss.2009.08.166
16. Barcelos R, Ribeiro G, Silva W, Abe-Sandes K, Godinho N, Marinho-Neto F, et al. Male Contribution in the Constitution of the Brazilian Centro-Oeste Population Estimated by Y-Chromosome Binary Markers. Int Congr Ser (2006) 1288:228–30. doi: 10.1016/j.ics.2005.11.109
17. Carvalho-Silva DR, Santos FR, Rocha J, Pena SDJ. The Phylogeography of Brazilian Y-Chromosome Lineages. Am J Hum Genet (2001) 68:281–6. doi: 10.1086/316931
18. Carvalho-Silva DR, Tarazona-Santos E, Rocha J, Pena SDJ, Santos FR. Y Chromosome Diversity in Brazilians: Switching Perspectives From Slow to Fast Evolving Markers. Genetica (2006) 126:251–60. doi: 10.1007/s10709-005-1454-z
19. Grattapaglia D, Kalupniek S, Guimarães CS, Ribeiro MA, Diener PS, Soares CN. Y-Chromosome STR Haplotype Diversity in Brazilian Populations. Forensic Sci Int (2005) 149:99–107. doi: 10.1016/j.forsciint.2004.06.003
20. Hünemeier T, Carvalho C, Marrero AR, Salzano FM, Junho Pena SD, Bortolini MC. Niger-Congo Speaking Populations and the Formation of the Brazilian Gene Pool: mtDNA and Y-Chromosome Data. Am J Phys Anthropol (2007) 133:854–67. doi: 10.1002/ajpa.20604
21. Leite FPN, Callegari-Jacques SM, Carvalho BA, Kommers T, Matte CHF, Raimann PE, et al. Y-STR Analysis in Brazilian and South Amerindian Populations. Am J Hum Biol (2008) 20:359–63. doi: 10.1002/ajhb.20702
22. Mendes-Junior CT, Ferreira LB, Canas M do CT, Pimentel AL, Muniz YCN, Wiezel CEV, et al. Y-Chromosome STR Haplotypes in a Sample From São Paulo State, Southeastern Brazil. J Forensic Sci (2007) 52:495–7. doi: 10.1111/j.1556-4029.2006.00372.x
23. Nunes ACS, Silva DA, Teixeira MAD, Nunes DD, Lopes CMS, Tucunduva Netto OR, et al. Y Chromosome Comparative Analysis of Rondônia With Other Brazilian Populations. Legal Med (2011) 13:161–3. doi: 10.1016/j.legalmed.2010.12.007
24. Palha T, Ribeiro-Rodrigues E, Ribeiro-dos-Santos Â, Santos S. Fourteen Short Tandem Repeat Loci Y Chromosome Haplotypes: Genetic Analysis in Populations From Northern Brazil. Forensic Sci Int: Genet (2012) 6:413–8. doi: 10.1016/j.fsigen.2011.08.003
25. Ribeiro GGBL, De Lima RR, Wiezel CEV, Ferreira LB, Sousa SMB, Rocha DMS, et al. Afro-Derived Brazilian Populations: Male Genetic Constitution Estimated by Y-Chromosomes STRs and AluYAP Element Polymorphisms. Am J Hum Biol (2009) 21:354–6. doi: 10.1002/ajhb.20875
26. São-Bento M, Carvalho M, Bento AM, Andrade L, Lopes V, Serra A, et al. Y-Chromosome SNP Analysis in the Brazilian Population of São Paulo State (Ribeirão Preto). Forensic Sci Int: Genet Suppl Ser (2009) 2:427–8. doi: 10.1016/j.fsigss.2009.08.117
27. Silva DA, Carvalho E, Costa G, Tavares L, Amorim A, Gusmão L. Y-Chromosome Genetic Variation in Rio De Janeiro Population. Am J Hum Biol (2006) 18:829–37. doi: 10.1002/ajhb.20567
28. Vallinoto ACR, Cayres-Vallinoto IMV, Ribeiro Dos Santos ÂKC, Zago MA, Santos SEB, Guerreiro JF. Heterogeneity of Y Chromosome Markers Among Brazilian Amerindians. Am J Hum Biol (1999) 11:481–7. doi: 10.1002/(SICI)1520-6300(1999)11:4<481::AID-AJHB7>3.0.CO;2-2
29. Ambrosio IB, Braganholi DF, Polverari FS, Cicarelli RMB. Mitochondrial DNA 30-SNP Data Confirm High Prevalence of African Lineages in the Population of Espirito Santo, Brazil. Forensic Sci Int: Genet Suppl Ser (2015) 5:e346–7. doi: 10.1016/j.fsigss.2015.09.137
30. Barbosa ABG, da Silva LAF, Azevedo DA, Balbino VQ, Mauricio-da-Silva L. Mitochondrial DNA Control Region Polymorphism in the Population of Alagoas State, North-Eastern Brazil. J Forensic Sci (2008) 53:142–6. doi: 10.1111/j.1556-4029.2007.00619.x
31. Bortolini MC, Zago MA, Salzano FM, Silva-Júnior WA, Bonatto SL, Silva MCBOD, et al. Evolutionary and Anthropological Implications of Mitochondrial DNA Variation in African Brazilian Populations. Hum Biol (1997) 69:141–59.
32. Freitas JM, Fassio LH, Braganholi DF, Chemale G. Mitochondrial DNA Control Region Haplotypes and Haplogroup Diversity in a Sample From Brasília, Federal District, Brazil. Forensic Sci Int: Genet (2019) 40:e228–30. doi: 10.1016/j.fsigen.2019.02.006
33. Marrero AR, Pereira Das Neves Leite F, De Almeida Carvalho B, Martins Peres L, Kommers TC, Mânica Da Cruz I, et al. Heterogeneity of the Genome Ancestry of Individuals Classified as White in the State of Rio Grande do Sul, Brazil. Am J Hum Biol (2005) 17:496–506. doi: 10.1002/ajhb.20404
34. Palencia L, Valverde L, Álvarez A, Cainé LM, Cardoso S, Alfonso-Sánchez MA, et al. Mitochondrial DNA Diversity in a Population From Santa Catarina (Brazil): Predominance of the European Input. Int J Legal Med (2010) 124:331–6. doi: 10.1007/s00414-010-0464-2
35. Poletto MM, Malaghini M, Silva JS, Bicalho MG, Braun-Prado K. Mitochondrial DNA Control Region Diversity in a Population From Parana State—Increasing the Brazilian Forensic Database. Int J Legal Med (2019) 133:347–51. doi: 10.1007/s00414-018-1886-5
36. Sanches NM, Paneto GG, Figueiredo RF, de Mello AO, Cicarelli RMB. Mitochondrial DNA Control Region Diversity in a Population From Espirito Santo State, Brazil. Mol Biol Rep (2014) 41:6645–8. doi: 10.1007/s11033-014-3547-1
37. Santos MV, Mendes C, Carvalho M, Vide MC, Corte-Real F, Vieira DN. Mitochondrial Variation in the Bahia–Brazil Population. Int Congr Ser (2004) 1261:404–6. doi: 10.1016/S0531-5131(03)01785-0
38. Cardena MMSG, Ribeiro-dos-Santos Â, Santos S, Mansur AJ, Pereira AC, Fridman C. Assessment of the Relationship Between Self-Declared Ethnicity, Mitochondrial Haplogroups and Genomic Ancestry in Brazilian Individuals. PloS One (2013) 8:e62005. doi: 10.1371/journal.pone.0062005
39. Hermida RMSM, Domingues PM, Silva DA, Carvalho EF. Ancestry Evaluation of Sub Saharan Male Descendant Population in Rio De Janeiro Inferred by Analysis of Mitochondrial DNA. Forensic Sci Int: Genet Suppl Ser (2013) 4:e214–5. doi: 10.1016/j.fsigss.2013.10.110
40. Ellwanger JH, Kaminski V de L, Rodrigues AG, Kulmann-Leal B, Chies JAB. CCR5 and CCR5Δ32 in Bacterial and Parasitic Infections: Thinking Chemokine Receptors Outside the HIV Box. Int J Immunogenet (2020) 47:261–85. doi: 10.1111/iji.12485
41. Carvalho MWP, Leboute APM, Oliveira SF, Sousa SMB, Klautau-Guimarães M de N, Simões AL. CCR5D32 Mutation in Three Brazilian Populations of Predominantly Sub-Saharan African Ancestry. Genet Mol Biol (2004) 27:321–5. doi: 10.1590/S1415-47572004000300002
42. Vargas AE, Marrero AR, Salzano FM, Bortolini MC, Chies JAB. Frequency of CCR5delta32 in Brazilian Populations. Braz J Med Biol Res (2006) 39:321–5. doi: 10.1590/s0100-879x2006000300002
43. Sabeti PC, Walsh E, Schaffner SF, Varilly P, Fry B, Hutcheson HB, et al. The Case for Selection at CCR5-Delta32. PloS Biol (2005) 3:e378. doi: 10.1371/journal.pbio.0030378
44. Stephens JC, Reich DE, Goldstein DB, Shin HD, Smith MW, Carrington M, et al. Dating the Origin of the CCR5-Delta32 AIDS-Resistance Allele by the Coalescence of Haplotypes. Am J Hum Genet (1998) 62:1507–15. doi: 10.1086/301867
45. Lidén K, Linderholm A, Götherström A. Pushing it Back. Dating the CCR5–32 Bp Deletion to the Mesolithic in Sweden and its Implications for the Meso\Neo Transition. Doc Praehistorica (2006) 33:29–37. doi: 10.4312/dp.33.5
46. Faure E, Royer-Carenzi M. Is the European Spatial Distribution of the HIV-1-Resistant CCR5-Delta32 Allele Formed by a Breakdown of the Pathocenosis Due to the Historical Roman Expansion? Infect Genet Evol (2008) 8:864–74. doi: 10.1016/j.meegid.2008.08.007
47. Solloch UV, Lang K, Lange V, Böhme I, Schmidt AH, Sauter J. Frequencies of Gene Variant CCR5-Δ32 in 87 Countries Based on Next-Generation Sequencing of 1.3 Million Individuals Sampled From 3 National DKMS Donor Centers. Hum Immunol (2017) 78:710–7. doi: 10.1016/j.humimm.2017.10.001
48. Shrestha D, Shrestha R, Sherchand SP, Sherchan S, Hendriksen RS, Bhatta-Sharma L, et al. Identification of CCR5 Δ32 Allele in Different Ethnic Groups of Nepal. Nepal Med Coll J (2020) 22:153–7. doi: 10.3126/nmcj.v22i3.32643
49. Silva-Carvalho WHV, de Moura RR, Coelho AVC, Crovella S, Guimarães RL. Frequency of the CCR5-Delta32 Allele in Brazilian Populations: A Systematic Literature Review and Meta-Analysis. Infect Genet Evol (2016) 43:101–7. doi: 10.1016/j.meegid.2016.05.024
50. Zlotnik A, Yoshie O. Chemokines: A New Classification System and Their Role in Immunity. Immunity (2000) 12:121–7. doi: 10.1016/s1074-7613(00)80165-x
51. Bacon K, Baggiolini M, Broxmeyer H, Horuk R, Lindley I, Mantovani A, et al. IUIS/WHO Subcommittee on Chemokine Nomenclature. Chemokine/Chemokine Receptor Nomenclature. Cytokine (2003) 21:48–9. doi: 10.1016/s1043-4666(02)00493-3
52. Boutet A, Salim H, Leclerc P, Tardieu M. Cellular Expression of Functional Chemokine Receptor CCR5 and CXCR4 in Human Embryonic Neurons. Neurosci Lett (2001) 311:105–8. doi: 10.1016/S0304-3940(01)02149-8
53. Hazan U, Romero IA, Cancello R, Valente S, Perrin V, Mariot V, et al. Human Adipose Cells Express CD4, CXCR4, and CCR5 Receptors: A New Target Cell Type for the Immunodeficiency Virus-1? FASEB J (2002) 16:1254–6. doi: 10.1096/fj.01-0947fje
54. Vaday GG, Peehl DM, Kadam PA, Lawrence DM. Expression of CCL5 (RANTES) and CCR5 in Prostate Cancer. Prostate (2006) 66:124–34. doi: 10.1002/pros.20306
55. Sales KJ, Adefuye A, Nicholson L, Katz AA. CCR5 Expression is Elevated in Cervical Cancer Cells and is Up-Regulated by Seminal Plasma. Mol Hum Reprod (2014) 20:1144–57. doi: 10.1093/molehr/gau063
56. Kranjc MK, Novak M, Pestell RG, Lah TT. Cytokine CCL5 and Receptor CCR5 Axis in Glioblastoma Multiforme. Radiol Oncol (2019) 53:397–406. doi: 10.2478/raon-2019-0057
57. Liu J, Wang C, Ma X, Tian Y, Wang C, Fu Y, et al. High Expression of CCR5 in Melanoma Enhances Epithelial-Mesenchymal Transition and Metastasis via Tgfβ1. J Pathol (2019) 247:481–93. doi: 10.1002/path.5207
58. Suarez-Carmona M, Chaorentong P, Kather JN, Rothenheber R, Ahmed A, Berthel A, et al. CCR5 Status and Metastatic Progression in Colorectal Cancer. Oncoimmunology (2019) 8:e1626193. doi: 10.1080/2162402X.2019.1626193
59. Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, Horuk R, et al. Homozygous Defect in HIV-1 Coreceptor Accounts for Resistance of Some Multiply-Exposed Individuals to HIV-1 Infection. Cell (1996) 86:367–77. doi: 10.1016/s0092-8674(00)80110-5
60. Wu L, Paxton WA, Kassam N, Ruffing N, Rottman JB, Sullivan N, et al. CCR5 Levels and Expression Pattern Correlate With Infectability by Macrophage-Tropic HIV-1, In Vitro. J Exp Med (1997) 185:1681–91. doi: 10.1084/jem.185.9.1681
61. Roda Husman AM, Blaak H, Brouwer M, Schuitemaker H. CC Chemokine Receptor 5 Cell-Surface Expression in Relation to CC Chemokine Receptor 5 Genotype and the Clinical Course of HIV-1 Infection. J Immunol (1999) 163:4597–603.
62. Segerer S, MacK M, Regele H, Kerjaschki D, Schlöndorff D. Expression of the C-C Chemokine Receptor 5 in Human Kidney Diseases. Kidney Int (1999) 56:52–64. doi: 10.1046/j.1523-1755.1999.00544.x
63. Venkatesan S, Petrovic A, Van Ryk DI, Locati M, Weissman D, Murphy PM. Reduced Cell Surface Expression of CCR5 in CCR5Delta 32 Heterozygotes is Mediated by Gene Dosage, Rather Than by Receptor Sequestration. J Biol Chem (2002) 277:2287–301. doi: 10.1074/jbc.M108321200
64. Benkirane M, Jin D-Y, Chun RF, Koup RA, Jeang K-T. Mechanism of Transdominant Inhibition of CCR5-Mediated HIV-1 Infection by Ccr5Δ32*. J Biol Chem (1997) 272:30603–6. doi: 10.1074/jbc.272.49.30603
65. Agrawal L, Lu X, Qingwen J, VanHorn-Ali Z, Nicolescu IV, McDermott DH, et al. Role for CCR5Δ32 Protein in Resistance to R5, R5X4, and X4 Human Immunodeficiency Virus Type 1 in Primary CD4+ Cells. J Virol (2004) 78:2277–87. doi: 10.1128/JVI.78.5.2277-2287.2004
66. Agrawal L, Jin Q, Altenburg J, Meyer L, Tubiana R, Theodorou I, et al. Ccr5Δ32 Protein Expression and Stability Are Critical for Resistance to Human Immunodeficiency Virus Type 1 In Vivo. J Virol (2007) 81:8041–9. doi: 10.1128/JVI.00068-07
67. Ellwanger JH, Kaminski VL, Chies JAB. CCR5 Gene Editing - Revisiting Pros and Cons of CCR5 Absence. Infect Genet Evol (2019) 68:218–20. doi: 10.1016/j.meegid.2018.12.027
68. Ellwanger JH, Kulmann-Leal B, Kaminski VL, Rodrigues AG, Bragatte MAS, Chies JAB. Beyond HIV Infection: Neglected and Varied Impacts of CCR5 and CCR5Δ32 on Viral Diseases. Virus Res (2020) 286:198040. doi: 10.1016/j.virusres.2020.198040
69. Coperchini F, Chiovato L, Croce L, Magri F, Rotondi M. The Cytokine Storm in COVID-19: An Overview of the Involvement of the Chemokine/Chemokine-Receptor System. Cytokine Growth Factor Rev (2020) 53:25–32. doi: 10.1016/j.cytogfr.2020.05.003
70. Mehlotra RK. Chemokine Receptor Gene Polymorphisms and COVID-19: Could Knowledge Gained From HIV/AIDS be Important? Infect Genet Evol (2020) 85:104512. doi: 10.1016/j.meegid.2020.104512
71. Hubacek JA, Dusek L, Majek O, Adamek V, Cervinkova T, Dlouha D, et al. Ccr5Δ32 Deletion as a Protective Factor in Czech First-Wave COVID-19 Subjects. Physiol Res (2021) 70:111–5. doi: 10.33549/physiolres.934647
72. Badia R, Riveira-Muñoz E, Clotet B, Esté JA, Ballana E. Gene Editing Using a Zinc-Finger Nuclease Mimicking the CCR5Δ32 Mutation Induces Resistance to CCR5-Using HIV-1. J Antimicrob Chemother (2014) 69:1755–9. doi: 10.1093/jac/dku072
73. Liu Z, Liang J, Chen S, Wang K, Liu X, Liu B, et al. Genome Editing of CCR5 by AsCpf1 Renders CD4+T Cells Resistance to HIV-1 Infection. Cell Biosci (2020) 10:85. doi: 10.1186/s13578-020-00444-w
74. Niemiec E, Howard HC. Ethical Issues Related to Research on Genome Editing in Human Embryos. Comput Struct Biotechnol J (2020) 18:887–96. doi: 10.1016/j.csbj.2020.03.014
75. Huang Y, Paxton WA, Wolinsky SM, Neumann AU, Zhang L, He T, et al. The Role of a Mutant CCR5 Allele in HIV-1 Transmission and Disease Progression. Nat Med (1996) 2:1240–3. doi: 10.1038/nm1196-1240
76. Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, et al. Resistance to HIV-1 Infection in Caucasian Individuals Bearing Mutant Alleles of the CCR-5 Chemokine Receptor Gene. Nature (1996) 382:722–5. doi: 10.1038/382722a0
77. Miao M, De Clercq E, Li G. Clinical Significance of Chemokine Receptor Antagonists. Expert Opin Drug Metab Toxicol (2020) 16:11–30. doi: 10.1080/17425255.2020.1711884
78. Arruda LB, de Araújo ML, Martinez ML, Gonsalez CR, da Silva Duarte AJ, Coakley E, et al. Determination of Viral Tropism by Genotyping and Phenotyping Assays in Brazilian HIV-1-Infected Patients. Rev do Inst Med Trop São Paulo (2014) 56:287–90. doi: 10.1590/S0036-46652014000400003
79. Pessôa R, Sabino EC, Sanabani SS. Frequency of Coreceptor Tropism in PBMC Samples From HIV-1 Recently Infected Blood Donors by Massively Parallel Sequencing: The REDS II Study. Virol J (2015) 12:74. doi: 10.1186/s12985-015-0307-3
80. Avanzi VM, Vicente BA, Beloto NCP, Gomes-da-Silva MM, Ribeiro CEL, Tuon FF, et al. Profile of HIV Subtypes in HIV/HBV- and HIV/HCV-Coinfected Patients in Southern Brazil. Rev da Sociedade Bras Med Trop (2017) 50:470–7. doi: 10.1590/0037-8682-0450-2016
81. Ellwanger JH, Kaminski VL, Chies JAB. What We Say and What We Mean When We Say Redundancy and Robustness of the Chemokine System - How CCR5 Challenges These Concepts. Immunol Cell Biol (2020) 98:22–7. doi: 10.1111/imcb.12291
82. Mauri M, Elli T, Caviglia G, Uboldi G, Azzi M. (2017). RAWGraphs: A Visualisation Platform to Create Open Outputs, in: Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter, CHItaly ‘17, . pp. 1–5. New York, NY, USA: Association for Computing Machinery. doi: 10.1145/3125571.3125585
83. Hütter G, Nowak D, Mossner M, Ganepola S, Müssig A, Allers K, et al. Long-Term Control of HIV by CCR5 Delta32/Delta32 Stem-Cell Transplantation. N Engl J Med (2009) 360:692–8. doi: 10.1056/NEJMoa0802905
84. Allers K, Hütter G, Hofmann J, Loddenkemper C, Rieger K, Thiel E, et al. Evidence for the Cure of HIV Infection by CCR5Δ32/Δ32 Stem Cell Transplantation. Blood (2011) 117:2791–9. doi: 10.1182/blood-2010-09-309591
85. Gupta RK, Abdul-Jawad S, McCoy LE, Mok HP, Peppa D, Salgado M, et al. HIV-1 Remission Following CCR5Δ32/Δ32 Haematopoietic Stem-Cell Transplantation. Nature (2019) 568:244–8. doi: 10.1038/s41586-019-1027-4
86. Gupta RK, Peppa D, Hill AL, Gálvez C, Salgado M, Pace M, et al. Evidence for HIV-1 Cure After CCR5Δ32/Δ32 Allogeneic Haemopoietic Stem-Cell Transplantation 30 Months Post Analytical Treatment Interruption: A Case Report. Lancet HIV (2020) 7:e340–7. doi: 10.1016/S2352-3018(20)30069-2
87. Kalidasan V, Theva Das K. Lessons Learned From Failures and Success Stories of HIV Breakthroughs: Are We Getting Closer to an HIV Cure? Front Microbiol (2020) 11:46. doi: 10.3389/fmicb.2020.00046
88. UNAIDS Brasil. UNAIDS Lastima a Morte De Timothy Brown, O ´Paciente De Berlim. Brasília-DF, Brazil: UNAIDS Brasil (2020). Available at: https://unaids.org.br/2020/09/unaids-lastima-a-morte-de-timothy-brown-o-paciente-de-berlim.
90. UNAIDS Brasil. O “Paciente De Berlim”: A Constante Superação do Estigma. Brasil: Programa Conjunto das Nações Unidas sobre HIV/AIDS (2019). Available at: https://unaids.org.br/2019/08/o-paciente-de-berlim-a-constante-superacao-do-estigma.
91. Brown TR. The London Patient. AIDS Res Hum Retroviruses (2020) 36:251–2. doi: 10.1089/aid.2020.0058
92. Ellwanger JH, Kulmann-Leal B, Wolf JM, Michita RT, Simon D, Lunge VR, et al. Role of the Genetic Variant CCR5Δ32 in HBV Infection and HBV/HIV Co-Infection. Virus Res (2020) 277:197838. doi: 10.1016/j.virusres.2019.197838
93. Chies JAB, Nardi NB. Sickle Cell Disease: A Chronic Inflammatory Condition. Med Hypotheses (2001) 57:46–50. doi: 10.1054/mehy.2000.1310
94. Eri R, Jonsson JR, Pandeya N, Purdie DM, Clouston AD, Martin N, et al. CCR5-Delta32 Mutation is Strongly Associated With Primary Sclerosing Cholangitis. Genes Immun (2004) 5:444–50. doi: 10.1038/sj.gene.6364113
95. Boiardi L, Vaglio A, Nicoli D, Farnetti E, Palmisano A, Pipitone N, et al. CC Chemokine Receptor 5 Polymorphism in Chronic Periaortitis. Rheumatology (2011) 50:1025–32. doi: 10.1093/rheumatology/keq416
96. Baltus THL, Kallaur AP, Lozovoy MAB, Morimoto HK, Delongui F, Alfieri DF, et al. Ccr5Δ32 (Rs333) Polymorphism is Associated With the Susceptibility to Systemic Lupus Erythematosus in Female Brazilian Patients. Rheumatol Int (2016) 36:7–15. doi: 10.1007/s00296-015-3308-z
97. Afzal AR, Kiechl S, Daryani YP, Weerasinghe A, Zhang Y, Reindl M, et al. Common CCR5-Del32 Frameshift Mutation Associated With Serum Levels of Inflammatory Markers and Cardiovascular Disease Risk in the Bruneck Population. Stroke (2008) 39:1972–8. doi: 10.1161/STROKEAHA.107.504381
98. Hütter G, Neumann M, Nowak D, Klein S, Klüter H, Hofmann W-K. The Effect of the CCR5-Delta32 Deletion on Global Gene Expression Considering Immune Response and Inflammation. J Inflammation (Lond) (2011) 8:29. doi: 10.1186/1476-9255-8-29
99. Muntinghe FLH, Abdulahad WH, Huitema MG, Damman J, Seelen MA, Lems SPM, et al. Ccr5Δ32 Genotype Leads to a Th2 Type Directed Immune Response in ESRD Patients. PloS One (2012) 7:e31257. doi: 10.1371/journal.pone.0031257
100. Joo S, Suwanto A, Sato A, Nakahashi-Ouchida R, Mori H, Uchida Y, et al. A Role for the CCR5-CCL5 Interaction in the Preferential Migration of HSV-2-Specific Effector Cells to the Vaginal Mucosa Upon Nasal Immunization. Mucosal Immunol (2019) 12:1391–403. doi: 10.1038/s41385-019-0203-z
101. Kulmann-Leal B, Ellwanger JH, Chies JAB. A Functional Interaction Between the CCR5 and CD34 Molecules Expressed in Hematopoietic Cells can Support (or Even Promote) the Development of Cancer. Hematol Transfus Cell Ther (2020) 42:70–6. doi: 10.1016/j.htct.2019.10.001
102. Martín-Leal A, Blanco R, Casas J, Sáez ME, Rodríguez-Bovolenta E, de Rojas I, et al. CCR5 Deficiency Impairs CD4+ T-Cell Memory Responses and Antigenic Sensitivity Through Increased Ceramide Synthesis. EMBO J (2020) 39:e104749. doi: 10.15252/embj.2020104749
103. Matti C, Legler DF. CCR5 Deficiency/CCR5Δ32: Resistant to HIV Infection at the Cost of Curtailed CD4+ T Cell Memory Responses. EMBO J (2020) 39:e105854. doi: 10.15252/embj.2020105854
104. Glass WG, McDermott DH, Lim JK, Lekhong S, Yu SF, Frank WA, et al. CCR5 Deficiency Increases Risk of Symptomatic West Nile Virus Infection. J Exp Med (2006) 203:35–40. doi: 10.1084/jem.20051970
105. Lim JK, Louie CY, Glaser C, Jean C, Johnson B, Johnson H, et al. Genetic Deficiency of Chemokine Receptor CCR5 is a Strong Risk Factor for Symptomatic West Nile Virus Infection: A Meta-Analysis of 4 Cohorts in the US Epidemic. J Infect Dis (2008) 197:262–5. doi: 10.1086/524691
106. Kindberg E, Mickiene A, Ax C, Akerlind B, Vene S, Lindquist L, et al. A Deletion in the Chemokine Receptor 5 (CCR5) Gene is Associated With Tickborne Encephalitis. J Infect Dis (2008) 197:266–9. doi: 10.1086/524709
107. Lim JK, McDermott DH, Lisco A, Foster GA, Krysztof D, Follmann D, et al. CCR5 Deficiency is a Risk Factor for Early Clinical Manifestations of West Nile Virus Infection But Not for Viral Transmission. J Infect Dis (2010) 201:178–85. doi: 10.1086/649426
108. Mickienė A, Pakalnienė J, Nordgren J, Carlsson B, Hagbom M, Svensson L, et al. Polymorphisms in Chemokine Receptor 5 and Toll-Like Receptor 3 Genes are Risk Factors for Clinical Tick-Borne Encephalitis in the Lithuanian Population. PloS One (2014) 9:e106798. doi: 10.1371/journal.pone.0106798
109. Ellwanger JH, Chies JAB. Host Immunogenetics in Tick-Borne Encephalitis Virus Infection-The CCR5 Crossroad. Ticks Tick Borne Dis (2019) 10:729–41. doi: 10.1016/j.ttbdis.2019.03.005
110. Hünemeier T, Neves AG, Nornberg I, Hill K, Hurtado AM, Carnese FR, et al. T-Cell and Chemokine Receptor Variation in South Amerindian Populations. Am J Hum Biol (2005) 17:515–8. doi: 10.1002/ajhb.20407
111. Ferreira-Fernandes H, Santos ACC, Motta FJN, Canalle R, Yoshioka FKN, Burbano RR, et al. Prevalence of CCR5-Δ32 and CCR2-V64I Polymorphisms in a Mixed Population From Northeastern Brazil. Genet Mol Res (2015) 14:11710–8. doi: 10.4238/2015.October.2.4
112. Leboute AP, de Carvalho MW, Simões AL. Absence of the Deltaccr5 Mutation in Indigenous Populations of the Brazilian Amazon. Hum Genet (1999) 105:442–3. doi: 10.1007/s004390051128
113. Carvalhaes FA de PL, Cardoso GL, Hamoy IG, Liu YT, Guerreiro JF. Distribution of CCR5-[Delta]32, CCR2-64I, and SDF1-3’a Mutations in Populations From the Brazilian Amazon Region. Hum Biol (2004) 76:643–6. doi: 10.1353/hub.2004.0052
114. Nguyen NZN, Tran VG, Lee S, Kim M, Kang SW, Kim J, et al. CCR5-Mediated Recruitment of NK Cells to the Kidney Is a Critical Step for Host Defense to Systemic Candida Albicans Infection. Immune Netw (2020) 20:e49. doi: 10.4110/in.2020.20.e49
115. Souza ALS, Souza PRS, Pereira CA, Fernandes A, Guabiraba R, Russo RC, et al. Experimental Infection With Schistosoma Mansoni in CCR5-Deficient Mice is Associated With Increased Disease Severity, as CCR5 Plays a Role in Controlling Granulomatous Inflammation. Infect Immun (2011) 79:1741–9. doi: 10.1128/IAI.00502-10
116. Chávez JH, França RFO, Oliveira CJF, de Aquino MTP, Farias KJS, Machado PRL, et al. Influence of the CCR-5/MIP-1 α Axis in the Pathogenesis of Rocio Virus Encephalitis in a Mouse Model. Am J Trop Med Hygiene (2013) 89:1013–8. doi: 10.4269/ajtmh.12-0591
117. Accetturi CA, Pardini R, Pinto GHN, Turcato GJ, Lewi DS, Diaz RS. Effects of CCR5 Genetic Polymorphism and HIV-1 Subtype in Antiretroviral Response in Brazilian HIV-1-Infected Patients. JAIDS J Acquired Immune Deficiency Syndromes (2000) 24:399–400. doi: 10.1097/00126334-200008010-00016
118. Vissoci Reiche EM, Ehara Watanabe MA, Bonametti AM, Morimoto HK, Akira Morimoto A, Wiechmann SL, et al. Frequency of CCR5-Δ32 Deletion in Human Immunodeficiency Virus Type 1 (HIV-1) in Healthy Blood Donors, HIV-1-Exposed Seronegative and HIV-1-Seropositive Individuals of Southern Brazilian Population. Int J Mol Med (2008) 22:669–75. doi: 10.3892/ijmm_00000071
119. Rigato PO, Hong MA, Casseb J, Ueda M, de Castro I, Benard G, et al. Better CD4+ T Cell Recovery in Brazilian HIV-Infected Individuals Under HAART Due to Cumulative Carriage of SDF-1-3’a, CCR2-V64I, CCR5-D32 and CCR5-Promoter 59029A/G Polymorphisms. Curr HIV Res (2008) 6:466–73. doi: 10.2174/157016208785861131
120. Valverde-Villegas JM, de Medeiros RM, de Andrade KP, Jacovas VC, Dos Santos BR, Simon D, et al. Novel Genetic Associations and Gene-Gene Interactions of Chemokine Receptor and Chemokine Genetic Polymorphisms in HIV/AIDS. AIDS (2017) 31:1235–43. doi: 10.1097/QAD.0000000000001491
121. Carvalhaes FA de PL, Cardoso GL, Vallinoto ACR, Machado LF, Ishak M de OG, Ishak R, et al. Frequencies of CCR5-Δ32, CCR2-64I and SDF1-3’A Mutations in Human Immunodeficiency Virus (HIV) Seropositive Subjects and Seronegative Individuals From the State of Pará in Brazilian Amazonia. Genet Mol Biol (2005) 28:665–9. doi: 10.1590/S1415-47572005000500004
122. Abe-Sandes K, Bomfim TF, Machado TMB, Abe-Sandes C, Acosta AX, Alves CRB, et al. Ancestralidade Genômica, Nível Socioeconômico E Vulnerabilidade Ao HIV/aids Na Bahia, Brasil. Saúde e Sociedade (2010) 19:75–84. doi: 10.1590/S0104-12902010000600008
123. Grimaldi R, Shindo N, Acosta AX, Dourado I, Brites C, de Melo Carvalho O, et al. Prevalence of the CCR5Delta32 Mutation in Brazilian Populations and Cell Susceptibility to HIV-1 Infection. Hum Genet (2002) 111:102–4. doi: 10.1007/s00439-002-0747-x
124. Mikawa AY, Tagliavini SA, Costa PI. CCR5 Genotype and Plasma Beta-Chemokine Concentration of Brazilian HIV-Infected Individuals. Braz J Med Biol Res (2002) 35:1333–7. doi: 10.1590/s0100-879x2002001100011
125. Munerato P, Azevedo ML, Sucupira MCA, Pardini R, Pinto GHN, Catroxo M, et al. Frequency of Polymorphisms of Genes Coding for HIV-1 Co-Receptors CCR5 and CCR2 in a Brazilian Population. Braz J Infect Dis (2003) 7:236–40. doi: 10.1590/s1413-86702003000400002
126. Souza PR, Arraes LC, de Lima Filho JL, Bruneska D, Milanese M, Crovella S. CCR5 Promoter Polymorphisms and HIV-1 Perinatal Transmission in Brazilian Children. J Reprod Immunol (2006) 69:77–84. doi: 10.1016/j.jri.2005.09.001
127. de Angelis DSA, Freire WS, Pannuti CS, de Menezes Succi RC, Machado DM. CCR5 Genotypes and Progression to HIV Disease in Perinatally Infected Children. Braz J Infect Dis (2007) 11:196–8. doi: 10.1590/S1413-86702007000200004
128. Teixeira SLM, Bastos FI, Hacker MA, Morgado MG. Distribution of CCR5 Genotypes and HLA Class I B Alleles in HIV-1 Infected and Uninfected Injecting Drug Users From Rio De Janeiro, Brazil. Infect Genet Evol (2009) 9:638–42. doi: 10.1016/j.meegid.2009.03.007
129. Vieira VC, Barral MFM, Mendoza-Sassi RA, Silveira JM, Soares MA, de Martínez AMB. The Effect of Combined Polymorphisms in Chemokines and Chemokine Receptors on the Clinical Course of HIV-1 Infection in a Brazilian Population. Mem Inst Oswaldo Cruz (2011) 106:408–15. doi: 10.1590/s0074-02762011000400005
130. Santos ÍM, da Rosa EA, Gräf T, Ferreira LGE, Petry A, Cavalheiro F, et al. Analysis of Immunological, Viral, Genetic, and Environmental Factors That Might Be Associated With Decreased Susceptibility to HIV Infection in Serodiscordant Couples in Florianópolis, Southern Brazil. AIDS Res Hum Retroviruses (2015) 31:1116–25. doi: 10.1089/aid.2015.0168
131. Corado A de LG, da Silva GAV, Leão RAC, Granja F, Naveca FG. Frequency of CCR5 Genotypes in HIV-Infected Patients in Roraima, Brazil. Braz J Infect Dis (2016) 20:314–5. doi: 10.1016/j.bjid.2016.01.001
132. Celerino da Silva R, Coelho AVC, Arraes LC, Brandão LAC, Crovella S, Guimarães RL. TRIM5 Gene Polymorphisms in HIV-1-Infected Patients and Healthy Controls From Northeastern Brazil. Immunol Res (2016) 64:1237–42. doi: 10.1007/s12026-016-8810-1
133. Arif MS, Hunter J, Léda AR, Zukurov JPL, Samer S, Camargo M, et al. Pace of Coreceptor Tropism Switch in HIV-1-Infected Individuals After Recent Infection. J Virol (2017) 91:e00793-17. doi: 10.1128/JVI.00793-17
134. Gomes STM, Gomes ÉR, Dos Santos MB, Lima SS, Queiroz MAF, Machado LFA, et al. Immunological and Virological Characterization of HIV-1 Viremia Controllers in the North Region of Brazil. BMC Infect Dis (2017) 17:381. doi: 10.1186/s12879-017-2491-9
135. Martin BM, Santos JS, Scapinello FV, Ribeiro CE, Gomes-da-Silva MM, Raboni SM. Young Adults HIV-1 Infected by Vertical Transmission in Southern Brazil - Clinical, Demographic, and Virological Features. Int J STD AIDS (2017) 28:1419–25. doi: 10.1177/0956462417712880
136. Celerino da Silva R, Moura RR, Victor Campos Coelho A, Arraes LC, Brandão LAC, Crovella S, et al. HLA-C Single Nucleotide Polymorphism Associated With Increased Viral Load Level in HIV-1 Infected Individuals From Northeast Brazil. Curr HIV Res (2017) 15:266–72. doi: 10.2174/1570162X15666170511141741
137. Carvalho-Silva WHV, Andrade-Santos JL, Guedes MC dos S, Crovella S, Guimarães RL. CCR5 Genotype and Pre-Treatment CD4+ T-Cell Count Influence Immunological Recovery of HIV-Positive Patients During Antiretroviral Therapy. Gene (2020) 741:144568. doi: 10.1016/j.gene.2020.144568
138. Barrios CS, Abuerreish M, Lairmore MD, Castillo L, Giam C-Z, Beilke MA. Recombinant Human T-Cell Leukemia Virus Types 1 and 2 Tax Proteins Induce High Levels of CC-Chemokines and Downregulate CCR5 in Human Peripheral Blood Mononuclear Cells. Viral Immunol (2011) 24:429–39. doi: 10.1089/vim.2011.0037
139. Oo Z, Barrios CS, Castillo L, Beilke MA. High Levels of CC-Chemokine Expression and Downregulated Levels of CCR5 During HIV-1/HTLV-1 and HIV-1/HTLV-2 Coinfections. J Med Virol (2015) 87:790–7. doi: 10.1002/jmv.24070
140. Sierra B, Perez AB, Garcia G, Aguirre E, Alvarez M, Gonzalez D, et al. Role of CC Chemokine Receptor 1 and Two of its Ligands in Human Dengue Infection. Three Approaches Under the Cuban Situation. Microbes Infect (2014) 16:40–50. doi: 10.1016/j.micinf.2013.10.011
141. Marques RE, Guabiraba R, Del Sarto JL, Rocha RF, Queiroz AL, Cisalpino D, et al. Dengue Virus Requires the CC-Chemokine Receptor CCR5 for Replication and Infection Development. Immunology (2015) 145:583–96. doi: 10.1111/imm.12476
142. Falcon A, Cuevas MT, Rodriguez-Frandsen A, Reyes N, Pozo F, Moreno S, et al. CCR5 Deficiency Predisposes to Fatal Outcome in Influenza Virus Infection. J Gen Virol (2015) 96:2074–8. doi: 10.1099/vir.0.000165
143. Pereira RW, Pires ER, Duarte APM, de Moura RP, Monteiro E, Torloni H, et al. Frequency of the CCRdelta32 Allele in Brazilians: A Study in Colorectal Cancer and in HTLV-I Infection. Genet Mol Biol (2000) 23:523–6. doi: 10.1590/S1415-47572000000300003
144. Hisada M, Lal RB, Masciotra S, Rudolph DL, Martin MP, Carrington M, et al. Chemokine Receptor Gene Polymorphisms and Risk of Human T Lymphotropic Virus Type I Infection in Jamaica. J Infect Dis (2002) 185:1351–4. doi: 10.1086/340129
145. Xavier-Carvalho C, Gibson G, Brasil P, Ferreira RX, de Souza Santos R, Gonçalves Cruz O, et al. Single Nucleotide Polymorphisms in Candidate Genes and Dengue Severity in Children: A Case-Control, Functional and Meta-Analysis Study. Infect Genet Evol (2013) 20:197–205. doi: 10.1016/j.meegid.2013.08.017
146. Brestovac B, Halicki LA, Harris RP, Sampson I, Speers DJ, Mamotte C, et al. Primary Acute Dengue and the Deletion in Chemokine Receptor 5 (CCR5Δ32). Microbes Infect (2014) 16:518–21. doi: 10.1016/j.micinf.2014.02.007
147. Maestri A, dos Santos MC, Ribeiro-Rodrigues EM, de Mello WA, Sousa RCM, dos Santos SE, et al. The Ccr5Δ32 (Rs333) Polymorphism is Not a Predisposing Factor for Severe Pandemic Influenza in the Brazilian Admixed Population. BMC Res Notes (2015) 8:326. doi: 10.1186/s13104-015-1299-1
148. Matos AR, Martins JSCC, Oliveira MdeLA, Garcia CC, Siqueira MM. Human Ccr5Δ32 (Rs333) Polymorphism has No Influence on Severity and Mortality of Influenza A(H1N1)pdm09 Infection in Brazilian Patients From the Post Pandemic Period. Infect Genet Evol (2019) 67:55–9. doi: 10.1016/j.meegid.2018.10.024
149. Keynan Y, Juno J, Meyers A, Ball TB, Kumar A, Rubinstein E, et al. Chemokine Receptor 5 Δ32 Allele in Patients With Severe Pandemic (H1N1) 2009. Emerging Infect Dis (2010) 16:1621–2. doi: 10.3201/eid1610.100108
150. Perz JF, Armstrong GL, Farrington LA, Hutin YJF, Bell BP. The Contributions of Hepatitis B Virus and Hepatitis C Virus Infections to Cirrhosis and Primary Liver Cancer Worldwide. J Hepatol (2006) 45:529–38. doi: 10.1016/j.jhep.2006.05.013
151. Ellwanger JH, Leal BK, Valverde-Villegas JM, Simon D, Marangon CG, Mattevi VS, et al. Ccr5Δ32 in HCV Infection, HCV/HIV Co-Infection, and HCV-Related Diseases. Infect Genet Evol (2018) 59:163–6. doi: 10.1016/j.meegid.2018.02.002
152. Abdolmohammadi R, Azar SS, Khosravi A, Shahbazi M. CCR5 Polymorphism as a Protective Factor for Hepatocellular Carcinoma in Hepatitis B Virus-Infected Iranian Patients. Asian Pac J Cancer Prev (2016) 17:4643–6. doi: 10.22034/APJCP.2016.17.10.4643
153. Suneetha PV, Sarin SK, Goyal A, Kumar GT, Hissar S. Host Genetic Factors and HBV Induced Liver Disease: Association of Vitamin D Receptor, CCR5, TNF-A and TNF-B Gene Polymorphisms With Viral Load and Disease Severity. J Hepatol (2006) 44:S15. doi: 10.1016/S0168-8278(06)80033-4
154. Goel V, Bose PD, Sarma MP, Hazam RK, Das BC, Gondal R, et al. Chemokine Receptor 5 (CCR5) Polymorphism in Chronic Hepatitis B Patients Treated With Three Different Nucleos(T)Ide Analogues. Indian J Med Res (2013) 137:1208–9.
155. Khorramdelazad H, Hakimizadeh E, Hassanshahi G, Rezayati M, Sendi H, Arababadi MK. CCR5 Δ 32 Mutation is Not Prevalent in Iranians With Chronic HBV Infection. J Med Virol (2013) 85:964–8. doi: 10.1002/jmv.23510
156. Rebbani K, Ezzikouri S, Marchio A, Ababou M, Kitab B, Dejean A, et al. Common Polymorphic Effectors of Immunity Against Hepatitis B and C Modulate Susceptibility to Infection and Spontaneous Clearance in a Moroccan Population. Infect Genet Evol (2014) 26:1–7. doi: 10.1016/j.meegid.2014.04.019
157. Oyervides-Muñoz MA, Pérez-Maya AA, Rodríguez-Gutiérrez HF, Gómez-Macias GS, Fajardo-Ramírez OR, Treviño V, et al. Understanding the HPV Integration and its Progression to Cervical Cancer. Infect Genet Evol (2018) 61:134–44. doi: 10.1016/j.meegid.2018.03.003
158. Mangieri LFL, Sena MM, Cezar-Dos-Santos F, Trugilo KP, Okuyama NCM, Pereira ÉR, et al. CCR5 Genetic Variants and Epidemiological Determinants for HPV Infection and Cervical Premalignant Lesions. Int J Immunogenet (2019) 46:331–8. doi: 10.1111/iji.12444
159. dos Santos EUD, de Lima GDC, de Oliveira ML, de Andrade Heráclio SA, da Silva HDA, Crovella S, et al. CCR2 and CCR5 Genes Polymorphisms in Women With Cervical Lesions From Pernambuco, Northeast Region of Brazil: A Case-Control Study. Mem Inst Oswaldo Cruz (2016) 111:174–80. doi: 10.1590/0074-02760150367
160. Zheng B, Wiklund F, Gharizadeh B, Sadat M, Gambelunghe G, Hallmans G, et al. Genetic Polymorphism of Chemokine Receptors CCR2 and CCR5 in Swedish Cervical Cancer Patients. Anticancer Res (2006) 26:3669–74.
161. Oliveira AP, Ayo CM, Bestetti RB, Brandão de Mattos CC, Cavasini CE, de Mattos LC. The Role of CCR5 in Chagas Disease - a Systematic Review. Infect Genet Evol (2016) 45:132–7. doi: 10.1016/j.meegid.2016.08.012
162. Oliveira AP, Bernardo CR, Camargo AVS, Villafanha DF, Cavasini CE, de Mattos CCB, et al. CCR5 Chemokine Receptor Gene Variants in Chronic Chagas’ Disease. Int J Cardiol (2014) 176:520–2. doi: 10.1016/j.ijcard.2014.07.043
163. Oliveira AP, Bernardo CR, Camargo AV da S, Ronchi LS, Borim AA, de Mattos CCB, et al. Genetic Susceptibility to Cardiac and Digestive Clinical Forms of Chronic Chagas Disease: Involvement of the CCR5 59029 a/G Polymorphism. PloS One (2015) 10:e0141847. doi: 10.1371/journal.pone.0141847
164. Calzada JE, Nieto A, Beraún Y, Martín J. Chemokine Receptor CCR5 Polymorphisms and Chagas’ Disease Cardiomyopathy. Tissue Antigens (2001) 58:154–8. doi: 10.1034/j.1399-0039.2001.580302.x
165. Fernández-Mestre MT, Montagnani S, Layrisse Z. Is the CCR5-59029-G/G Genotype a Protective Factor for Cardiomyopathy in Chagas Disease? Hum Immunol (2004) 65:725–8. doi: 10.1016/j.humimm.2004.05.002
166. Brajão de Oliveira K, Reiche EMV, Kaminami Morimoto H, Pelegrinelli Fungaro MH, Estevão D, Pontello R, et al. Analysis of the CC Chemokine Receptor 5 Delta32 Polymorphism in a Brazilian Population With Cutaneous Leishmaniasis. J Cutan Pathol (2007) 34:27–32. doi: 10.1111/j.1600-0560.2006.00573.x
167. Ribas AD, Ribas RC, Da Silva Júnior WV, Aristides SMA, Lonardoni MVC, Watanabe MAE, et al. Effect of the Chemokine Receptor CCR5 in the Development of American Cutaneous Leishmaniasis in a Southern Brazilian Population. Mol Med Rep (2013) 8:189–94. doi: 10.3892/mmr.2013.1452
168. Sophie M, Hameed A, Muneer A, Samdani AJ, Saleem S, Azhar A. CC Chemokine Receptor 5 Δ32 Polymorphism: Association Analysis and Allele Distribution Among Cutaneous Leishmaniasis Patients From Pakistan. J Cutaneous Pathol (2016) 43:564–70. doi: 10.1111/cup.12712
169. Faria Junior GM, Ayo CM, de Oliveira AP, Lopes AG, Frederico FB, Silveira-Carvalho AP, et al. CCR5 Chemokine Receptor Gene Polymorphisms in Ocular Toxoplasmosis. Acta Trop (2018) 178:276–80. doi: 10.1016/j.actatropica.2017.12.012
170. Vallochi AL, Goldberg AC, Falcai A, Ramasawmy R, Kalil J, Silveira C, et al. Molecular Markers of Susceptibility to Ocular Toxoplasmosis, Host and Guest Behaving Badly. Clin Ophthalmol (2008) 2:837–48. doi: 10.2147/opth.s1629
171. Costa MC, Pereira de Sá N, Johann S, Santos DA. Social, Environmental and Microbiologic Aspects of Endemic Mycoses in Brazil. New Microbes New Infect (2019) 29:100496. doi: 10.1016/j.nmni.2018.11.004
172. Kaimen-Maciel DR, Vissoci Reiche EM, Brum Souza DG, Frota Comini ER, Bobroff F, Morimoto HK, et al. CCR5-Δ32 Genetic Polymorphism Associated With Benign Clinical Course and Magnetic Resonance Imaging Findings in Brazilian Patients With Multiple Sclerosis. Int J Mol Med (2007) 20:337–44. doi: 10.3892/ijmm.20.3.337
173. Troncoso LL, Pontillo A, de Oliveira EML, Finkelszteijn A, Schneider S, Chies JAB. Ccr5Δ32 - A Piece of Protection in the Inflammatory Puzzle of Multiple Sclerosis Susceptibility. Hum Immunol (2018) 79:621–6. doi: 10.1016/j.humimm.2018.04.015
174. Scheibel I, Veit T, Neves AG, Souza L, Prezzi S, Machado S, et al. Differential CCR5Delta32 Allelic Frequencies in Juvenile Idiopathic Arthritis Subtypes: Evidence for Different Regulatory Roles of CCR5 in Rheumatological Diseases. Scand J Rheumatol (2008) 37:13–7. doi: 10.1080/03009740701631935
175. Souza M do SQA, de Souza CA, Cunha LMP, de Souza AQA, de Morais MS, Rabenhorst SHB. A New Look at Osteomyelitis Development–Focus on CCR5delta32. Study in Patients From Northeast Brazil. Infect Genet Evol (2015) 31:61–3. doi: 10.1016/j.meegid.2015.01.012
176. Cavalla F, Biguetti CC, Dionisio TJ, Azevedo MCS, Martins W, Santos CF, et al. Ccr5Δ32 (Rs333) Polymorphism is Associated With Decreased Risk of Chronic and Aggressive Periodontitis: A Case-Control Analysis Based in Disease Resistance and Susceptibility Phenotypes. Cytokine (2018) 103:142–9. doi: 10.1016/j.cyto.2017.09.022
177. Telini B, Veit TD, Chies JAB, Vianna P. The Ccr5Δ32 Polymorphism as a Pre-Eclampsia Susceptibility Marker: An Evaluation in Brazilian Women. Arch Gynecol Obstet (2014) 290:1–3. doi: 10.1007/s00404-014-3246-0
178. Kaminski VL, Ellwanger JH, Sandrim V, Pontillo A, Chies JAB. Influence of NKG2C Gene Deletion and CCR5Δ32 in Pre-Eclampsia-Approaching the Effect of Innate Immune Gene Variants in Pregnancy. Int J Immunogenet (2019) 46:82–7. doi: 10.1111/iji.12416
179. Kohem CL, Brenol JCT, Xavier RM, Bredemeier M, Brenol CV, Dedavid e Silva TL, et al. The Chemokine Receptor CCR5 Genetic Polymorphism and Expression in Rheumatoid Arthritis Patients. Scand J Rheumatol (2007) 36:359–64. doi: 10.1080/03009740701393999
180. Toson B, Dos Santos EJ, Adelino JE, Sandrin-Garcia P, Crovella S, Louzada-Júnior P, et al. Ccr5Δ32 and the Genetic Susceptibility to Rheumatoid Arthritis in Admixed Populations: A Multicentre Study. Rheumatol (Oxford) (2017) 56:495–7. doi: 10.1093/rheumatology/kew398
181. Chies JAB, Hutz MH. High Frequency of the CCR5delta32 Variant Among Individuals From an Admixed Brazilian Population With Sickle Cell Anemia. Braz J Med Biol Res (2003) 36:71–5. doi: 10.1590/S0100-879X2003000100010
182. Vargas AE, da Silva MAL, Silla L, Chies JAB. Polymorphisms of Chemokine Receptors and eNOS in Brazilian Patients With Sickle Cell Disease. Tissue Antigens (2005) 66:683–90. doi: 10.1111/j.1399-0039.2005.00506.x
183. Lopes MP, Santos MNN, Faber EW, Bezerra MAC, Hatzlhofer BLD, Albuquerque DM, et al. The Ccr5Δ32 Polymorphism in Brazilian Patients With Sickle Cell Disease. Dis Markers (2014) 2014:678246. doi: 10.1155/2014/678246
184. Nascimento AF, Oliveira JS, Silva Junior JC, Barbosa AAL. Genomic Ancestry Evaluated by Ancestry-Informative Markers in Patients With Sickle Cell Disease. Genet Mol Res (2016) 15:gmr7604. doi: 10.4238/gmr.15017604
185. Schauren JS, Marasca JA, Veit TD, Monticielo OA, Xavier RM, Brenol JCT, et al. CCR5delta32 in Systemic Lupus Erythematosus: Implications for Disease Susceptibility and Outcome in a Brazilian Population. Lupus (2013) 22:802–9. doi: 10.1177/0961203313491848
186. Cilião HL, Camargo-Godoy RBO, de Souza MF, Dos Reis MB, Iastrenski L, Alvares Delfino VD, et al. Association of UGT2B7, UGT1A9, ABCG2, and IL23R Polymorphisms With Rejection Risk in Kidney Transplant Patients. J Toxicol Environ Health A (2017) 80:661–71. doi: 10.1080/15287394.2017.1286922
187. Oliveira CEC, Amarante MK, Perim A de L, Ozawa PMM, Hiroki C, Freire Vitiello GA, et al. Absence of Association Between CCR5 Rs333 Polymorphism and Childhood Acute Lymphoblastic Leukemia. Adv Hematol (2014) 2014:924030. doi: 10.1155/2014/924030
188. Aoki MN, da silva do amaral Herrera AC, Amarante MK, do Val Carneiro JL, Fungaro MHP, Watanabe MAE. CCR5 and P53 Codon 72 Gene Polymorphisms: Implications in Breast Cancer Development. Int J Mol Med (2009) 23:429–35. doi: 10.3892/ijmm_00000148
189. Banin-Hirata BK, Losi-Guembarovski R, Oda JMM, de Oliveira CEC, Campos CZ, Mazzuco TL, et al. CCR2-V64I Genetic Polymorphism: A Possible Involvement in HER2+ Breast Cancer. Clin Exp Med (2016) 16:139–45. doi: 10.1007/s10238-015-0342-9
190. Derossi DR, Amarante MK, Guembarovski RL, de Oliveira CEC, Suzuki KM, Watanabe MAE, et al. CCL5 Protein Level: Influence on Breast Cancer Staging and Lymph Nodes Commitment. Mol Biol Rep (2019) 46:6165–70. doi: 10.1007/s11033-019-05051-8
191. Vieira-Filho DRM, Amarante MK, Ishibashi CM, Ariza CB, Vitiello GAF, de Oliveira KB, et al. CCR5 and CXCL12 Allelic Variants: Possible Association With Childhood Neuroblastoma Susceptibility? J Neuroimmunol (2020) 342:577193. doi: 10.1016/j.jneuroim.2020.577193
192. Magnani M, Castro-Gomez RH, Aoki MN, Gregório EP, Libos F, Morimoto HK, et al. Analysis of Peripheral T Cells and the CC Chemokine Receptor (CCR5) Delta32 Polymorphism in Prostate Cancer Patients Treated With Carboxymethyl-Glucan (CM-G). Nat Prod Res (2012) 26:945–51. doi: 10.1080/14786419.2010.535159
193. Zambra FMB, Biolchi V, Brum IS, Chies JAB. CCR2 and CCR5 Genes Polymorphisms in Benign Prostatic Hyperplasia and Prostate Cancer. Hum Immunol (2013) 74:1003–8. doi: 10.1016/j.humimm.2013.04.031
194. Allanore Y, Simms R, Distler O, Trojanowska M, Pope J, Denton CP, et al. Systemic Sclerosis. Nat Rev Dis Primers (2015) 1:1–21. doi: 10.1038/nrdp.2015.2
195. Sellebjerg F, Madsen HO, Jensen CV, Jensen J, Garred P. CCR5 Delta32, Matrix Metalloproteinase-9 and Disease Activity in Multiple Sclerosis. J Neuroimmunol (2000) 102:98–106. doi: 10.1016/s0165-5728(99)00166-6
196. Otaegui D, Ruíz-Martínez J, Olaskoaga J, Emparanza JI, López de Munain A. Influence of CCR5-Delta32 Genotype in Spanish Population With Multiple Sclerosis. Neurogenetics (2007) 8:201–5. doi: 10.1007/s10048-007-0085-1
197. van Veen T, Nielsen J, Berkhof J, Barkhof F, Kamphorst W, Bö L, et al. CCL5 and CCR5 Genotypes Modify Clinical, Radiological and Pathological Features of Multiple Sclerosis. J Neuroimmunol (2007) 190:157–64. doi: 10.1016/j.jneuroim.2007.08.005
198. Song GG, Lee YH. A Meta-Analysis of the Relation Between Chemokine Receptor 5 Delta32 Polymorphism and Multiple Sclerosis Susceptibility. Immunol Invest (2014) 43:299–311. doi: 10.3109/08820139.2013.845204
199. Bennetts BH, Teutsch SM, Buhler MM, Heard RNS, Stewart GJ. The CCR5 Deletion Mutation Fails to Protect Against Multiple Sclerosis. Hum Immunol (1997) 58:52–9. doi: 10.1016/S0198-8859(97)00207-3
200. D’Angelo R, Crisafulli C, Rinaldi C, Ruggeri A, Amato A, Sidoti A. Ccr5Δ32 Polymorphism Associated With a Slower Rate Disease Progression in a Cohort of RR-MS Sicilian Patients. Multiple Sclerosis Int (2011) 2011:e153282. doi: 10.1155/2011/153282
201. Gade-Andavolu R, Comings DE, MacMurray J, Rostamkhani M, Cheng LS-C, Tourtellotte WW, et al. Association of CCR5 Δ32 Deletion With Early Death in Multiple Sclerosis. Genet Med (2004) 6:126–31. doi: 10.1097/01.GIM.0000127274.45301.54
202. Karam RA, Rezk NA, Amer MM, Fathy HA. Immune Response Genes Receptors Expression and Polymorphisms in Relation to Multiple Sclerosis Susceptibility and Response to INF-β Therapy. IUBMB Life (2016) 68:727–34. doi: 10.1002/iub.1530
203. Carvalho C, Calvisi SL, Leal B, Bettencourt A, Marinho A, Almeida I, et al. CCR5-Delta32: Implications in SLE Development. Int J Immunogenet (2014) 41:236–41. doi: 10.1111/iji.12094
204. Cheng F-J, Zhou X-J, Zhao Y-F, Zhao M-H, Zhang H. Chemokine Receptor 5 (CCR5) Delta 32 Polymorphism in Lupus Nephritis: A Large Case-Control Study and Meta-Analysis. Autoimmunity (2014) 47:383–8. doi: 10.3109/08916934.2014.906581
205. Lee YH, Kim J-H, Song GG. Chemokine Receptor 5 Δ32 Polymorphism and Systemic Lupus Erythematosus, Vasculitis, and Primary Sjogren’s Syndrome. Z Rheumatol (2014) 73:848–55. doi: 10.1007/s00393-014-1356-5
206. Martens HA, Gross S, van der Steege G, Brouwer E, Berden JHM, de Sevaux R, et al. Lack of Association of C-C Chemokine Receptor 5 Δ32 Deletion Status With Rheumatoid Arthritis, Systemic Lupus Erythematosus, Lupus Nephritis, and Disease Severity. J Rheumatol (2010) 37:2226–31. doi: 10.3899/jrheum.091468
207. Heydarifard Z, Tabarraei A, Abdollahi N, Moradi A, Khanjari Y. Evaluation of CCR5Δ32 Polymorphism in Patients With Systemic Lupus Erythematosus and Healthy Individuals. Med Lab J (2018) 12:38–43. doi: 10.29252/mlj.12.2.38
208. Rzeszotarska E, Sowinska A, Stypinska B, Walczuk E, Wajda A, Lutkowska A, et al. The Role of MECP2 and CCR5 Polymorphisms on the Development and Course of Systemic Lupus Erythematosus. Biomolecules (2020) 10:494. doi: 10.3390/biom10030494
209. Michita RT, Kaminski VL, Chies JAB. Genetic Variants in Preeclampsia: Lessons From Studies in Latin-American Populations. Front Physiol (2018) 9:1771. doi: 10.3389/fphys.2018.01771
210. Gurdol F, Yurdum LM, Ozturk U, Isbilen E, Cakmakoglu B. Association of the CC Chemokine Receptor 5 (CCR5) Polymorphisms With Preeclampsia in Turkish Women. Arch Gynecol Obstet (2012) 286:51–4. doi: 10.1007/s00404-012-2244-3
211. Lee YH, Bae S-C, Song GG. Association Between the Chemokine Receptor 5 Delta32 Polymorphism and Rheumatoid Arthritis: A Meta-Analysis. Modern Rheumatol (2013) 23:304–10. doi: 10.3109/s10165-012-0665-2
212. Veit TD, Scheibel I, Chies JA. Considerations About the Role of the CCR5 Gene in Juvenile Idiopathic Arthritis - Look at the Whole or Put All Parts Together? Open Rheumatol J (2011) 5:78–80. doi: 10.2174/1874312901105010078
213. Chasnyk V, Fedorova E, Egorov A, Ammosova T, Nekhai S, Kostik M, et al. A130: Is the CCR5-Delta32 Mutation Protective Against Systemic-Onset Juvenile Idiopathic Arthritis? Arthritis Rheumatol (2014) 66:S171–1. doi: 10.1002/art.38551
214. Gamonal J, Acevedo A, Bascones A, Jorge O, Silva A. Characterization of Cellular Infiltrate, Detection of Chemokine Receptor CCR5 and Interleukin-8 and RANTES Chemokines in Adult Periodontitis. J Periodontal Res (2001) 36:194–203. doi: 10.1034/j.1600-0765.2001.360309.x
215. Garlet GP, Martins W, Ferreira BR, Milanezi CM, Silva JS. Patterns of Chemokines and Chemokine Receptors Expression in Different Forms of Human Periodontal Disease. J Periodontal Res (2003) 38:210–7. doi: 10.1034/j.1600-0765.2003.02012.x
216. Ferreira SB, Repeke CE, Raimundo FM, Nunes IS, Avila-Campos MJ, Ferreira BR, et al. CCR5 Mediates Pro-Osteoclastic and Osteoclastogenic Leukocyte Chemoattraction. J Dent Res (2011) 90:632–7. doi: 10.1177/0022034510395021
217. Rossi AD, Alves SYF, Nelson-Filho P, Silva RAB, Silva LAB. Involvement of CCR5 in Bone Resorption Associated to Apical Periodontitis. J Immunol (2019) 202. 181.12-181.12.
218. Folwaczny M, Glas J, Török H-P, Fricke K, Folwaczny C. Prevalence of the Chemokine Receptor CCR5-Delta32 Gene Mutation in Periodontal Disease. Clin Immunol (2003) 109:325–9. doi: 10.1016/j.clim.2003.08.001
219. Shih Y-S, Fu E, Fu MM, Lin F-G, Chiu H-C, Shen E-C, et al. Association of CCL5 and CCR5 Gene Polymorphisms With Periodontitis in Taiwanese. J Periodontol (2014) 85:1596–602. doi: 10.1902/jop.2014.130651
220. Olson ME, Horswill AR. Staphylococcus Aureus Osteomyelitis: Bad to the Bone. Cell Host Microbe (2013) 13:629–31. doi: 10.1016/j.chom.2013.05.015
221. Akalin E, Murphy B. Gene Polymorphisms and Transplantation. Curr Opin Immunol (2001) 13:572–6. doi: 10.1016/S0952-7915(00)00261-2
222. Fischereder M, Luckow B, Hocher B, Wüthrich RP, Rothenpieler U, Schneeberger H, et al. CC Chemokine Receptor 5 and Renal-Transplant Survival. Lancet (2001) 357:1758–61. doi: 10.1016/s0140-6736(00)04898-4
223. Moench C, Uhrig A, Lohse AW, Otto G. CC Chemokine Receptor 5delta32 Polymorphism-a Risk Factor for Ischemic-Type Biliary Lesions Following Orthotopic Liver Transplantation. Liver Transpl (2004) 10:434–9. doi: 10.1002/lt.20095
224. Abdi R, Huong TTB, Sahagun-Ruiz A, Murphy PM, Brenner BM, Milford EL, et al. Chemokine Receptor Polymorphism and Risk of Acute Rejection in Human Renal Transplantation. JASN (2002) 13:754–8. doi: 10.1681/ASN.V133754
225. Azmandian J, Mandegary A, Saber A, Torshabi M, Etminan A, Ebadzadeh M-R, et al. Chemokine Receptor 2-V64I and Chemokine Receptor 5-Delta32 Polymorphisms and Clinical Risk Factors of Delayed Graft Function and Acute Rejection in Kidney Transplantation. Iran J Kidney Dis (2012) 6:56–62.
226. Firasat S, Raza A, Abid A, Aziz T, Mubarak M, Naqvi SAA, et al. The Effect of Chemokine Receptor Gene Polymorphisms (CCR2V64I, CCR5-59029g>A and CCR5Δ32) on Renal Allograft Survival in Pakistani Transplant Patients. Gene (2012) 511:314–9. doi: 10.1016/j.gene.2012.09.099
227. Singh R, Kapoor R, Srivastava A, Mittal RD. Impact of Chemokine Receptor CCR2 and CCR5 Gene Polymorphism on Allograft Outcome in North Indian Renal Transplant Recipients. Scand J Immunol (2009) 69:51–6. doi: 10.1111/j.1365-3083.2008.02192.x
228. Yigit B, Bozkurt N, Berber I, Titiz I, İsbir T. Analysis of CC Chemokine Receptor 5 and 2 Polymorphisms and Renal Transplant Survival. Cell Biochem Funct (2007) 25:423–6. doi: 10.1002/cbf.1322
229. El Sissy MH, Hafez AA, Moneim SEA, Eldemerdash DM. Association of the CCR5Δ32 Mutant Genotype With Sickle Cell Disease in Egyptian Patients. Hemoglobin (2019) 43:258–63. doi: 10.1080/03630269.2019.1680381
230. Ying H, Wang J, Gao X. CCL5-403, CCR5-59029, and Delta32 Polymorphisms and Cancer Risk: A Meta-Analysis Based on 20,625 Subjects. Tumor Biol (2014) 35:5895–904. doi: 10.1007/s13277-014-1780-9
231. Fatima F, Saleem S, Hameed A, Haider G, Ali Zaidi SA, Kanwal M, et al. Association Analysis and Allelic Distribution of Deletion in CC Chemokine Receptor 5 Gene (CCR5Δ32) Among Breast Cancer Patients of Pakistan. Mol Biol Rep (2019) 46:2387–94. doi: 10.1007/s11033-019-04699-6
232. Span PN, Pollakis G, Paxton WA, Sweep FCGJ, Foekens JA, Martens JWM, et al. Improved Metastasis-Free Survival in Nonadjuvantly Treated Postmenopausal Breast Cancer Patients With Chemokine Receptor 5 Del32 Frameshift Mutations. Int J Cancer (2015) 136:91–7. doi: 10.1002/ijc.28962
233. Tajbakhsh A, Farjami Z, Nesaei-Bajestani A, Afzaljavan F, Rivandi M, Moezzi A, et al. Evaluating the Association Between CCR5delta32 Polymorphism (Rs333) and the Risk of Breast Cancer in a Cohort of Iranian Population. Iran J Public Health (2021) 50:583–91. doi: 10.18502/ijph.v50i3.5604
234. Farhood B, Najafi M, Mortezaee K. CD8+ Cytotoxic T Lymphocytes in Cancer Immunotherapy: A Review. J Cell Physiol (2019) 234:8509–21. doi: 10.1002/jcp.27782
235. Yang X, Cheng Y, Li C. The Role of TLRs in Cervical Cancer With HPV Infection: A Review. Signal Transduct Target Ther (2017) 2:17055. doi: 10.1038/sigtrans.2017.55
236. Ellwanger JH, Zambra FMB, Guimarães RL, Chies JAB. MicroRNA-Related Polymorphisms in Infectious Diseases-Tiny Changes With a Huge Impact on Viral Infections and Potential Clinical Applications. Front Immunol (2018) 9:1316. doi: 10.3389/fimmu.2018.01316
237. Ubogu EE, Callahan MK, Tucky BH, Ransohoff RM. CCR5 Expression on Monocytes and T Cells: Modulation by Transmigration Across the Blood-Brain Barrier In Vitro. Cell Immunol (2006) 243:19–29. doi: 10.1016/j.cellimm.2006.12.001
238. Sorce S, Myburgh R, Krause K-H. The Chemokine Receptor CCR5 in the Central Nervous System. Prog Neurobiol (2011) 93:297–311. doi: 10.1016/j.pneurobio.2010.12.003
239. Martin-Blondel G, Brassat D, Bauer J, Lassmann H, Liblau RS. CCR5 Blockade for Neuroinflammatory Diseases–Beyond Control of HIV. Nat Rev Neurol (2016) 12:95–105. doi: 10.1038/nrneurol.2015.248
240. Gonzalez E, Bamshad M, Sato N, Mummidi S, Dhanda R, Catano G, et al. Race-Specific HIV-1 Disease-Modifying Effects Associated With CCR5 Haplotypes. Proc Natl Acad Sci U.S.A. (1999) 96:12004–9. doi: 10.1073/pnas.96.21.12004
241. Liu J, Lewinger JP, Gilliland FD, Gauderman WJ, Conti DV. Confounding and Heterogeneity in Genetic Association Studies With Admixed Populations. Am J Epidemiol (2013) 177:351–60. doi: 10.1093/aje/kws234
Keywords: CCR5, CCR5delta32, Brazil, cancer, inflammation, infectious disease, pathogen, population genetics
Citation: Kulmann-Leal B, Ellwanger JH and Chies JAB (2021) CCR5Δ32 in Brazil: Impacts of a European Genetic Variant on a Highly Admixed Population. Front. Immunol. 12:758358. doi: 10.3389/fimmu.2021.758358
Received: 13 August 2021; Accepted: 23 November 2021;
Published: 10 December 2021.
Edited by:
Masakazu Kamata, University of Alabama at Birmingham, United StatesReviewed by:
Harish Padh, Gujarat National Law University, IndiaFeng-Liang Liu, Kunming Institute of Zoology, China
Copyright © 2021 Kulmann-Leal, Ellwanger and Chies. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Artur Bogo Chies, jabchies@terra.com.br
†These authors have contributed equally to this work