 Paloma A. Harcha
Paloma A. Harcha Tamara López-López
Tamara López-López Adrián G. Palacios
Adrián G. Palacios Pablo J. Sáez
Pablo J. Sáez- 1Centro Interdisciplinario de Neurociencia de Valparaíso, Instituto de Neurociencia, Facultad de Ciencias, Universidad de Valparaíso, Valparaíso, Chile
- 2Cell Communication and Migration Laboratory, Institute of Biochemistry and Molecular Cell Biology, Center for Experimental Medicine, University Medical Center Hamburg-Eppendorf, Hamburg, Germany
The role of Pannexin (PANX) channels during collective and single cell migration is increasingly recognized. Amongst many functions that are relevant to cell migration, here we focus on the role of PANX-mediated adenine nucleotide release and associated autocrine and paracrine signaling. We also summarize the contribution of PANXs with the cytoskeleton, which is also key regulator of cell migration. PANXs, as mechanosensitive ATP releasing channels, provide a unique link between cell migration and purinergic communication. The functional association with several purinergic receptors, together with a plethora of signals that modulate their opening, allows PANX channels to integrate physical and chemical cues during inflammation. Ubiquitously expressed in almost all immune cells, PANX1 opening has been reported in different immunological contexts. Immune activation is the epitome coordination between cell communication and migration, as leukocytes (i.e., T cells, dendritic cells) exchange information while migrating towards the injury site. In the current review, we summarized the contribution of PANX channels during immune cell migration and recruitment; although we also compile the available evidence for non-immune cells (including fibroblasts, keratinocytes, astrocytes, and cancer cells). Finally, we discuss the current evidence of PANX1 and PANX3 channels as a both positive and/or negative regulator in different inflammatory conditions, proposing a general mechanism of these channels contribution during cell migration.
1 Introduction
Cell communication and cell migration are key phenomena for development, tissue repair, and immune response; thus coordination of these responses are key for sustaining life (1–4). Indeed, a fine coordination of leukocyte communication is required for migration to clear an infection, or recruit other migrating cells towards an injury site. Interestingly, immune cells use different migratory strategies associated with their immune function and location (5–7). For example, under resting conditions immune cells would randomly patrol the tissue, but upon activation undergo directional migration to reach the secondary lymphoid organs (8). Despite presenting unique features, immune cell migration follows the general rules of cell migration, and depends on cytoskeletal dynamics (actin, non-muscular myosin II [MyoII]), and microtubules, as described in detail in the following reviews (6, 9–11). The study of immune cell migration is directly linked to development of new techniques to monitor the behavior of these cells in their native microenvironment (5, 12, 13), although this is still very challenging. However, researchers have developed ex vivo (i.e., tissue slices), and in vitro systems that mimic some tissue properties (i.e., confinement, properties of the extracellular matrix, etc). Thus, motility has been studied in models with different levels of microenvironment complexity (i.e., 1D, 2D, and 3D), topographies (that do or do not impose cellular deformation), or that mimic their transmigration through tissue layers (6, 13, 14).
1.1 Danger Signals and the Role of Purinergic Signaling
Immune cell migration is also controlled by microenvironmental chemical cues, such as chemokines and danger signals, affecting cell positioning along the tissue (15–17). Danger signals are molecules that trigger the immune system, and are classified due to their origin: damage-associated molecular patterns (DAMPs) [such as extracellular adenosine-5′-triphosphate (ATP)], and pathogen-associated molecular patterns (PAMPs) [such as lipopolysaccharide (LPS) (16). Both DAMPs and PAMPs trigger immune cell maturation, and expression of receptors (i.e. chemokine receptors) promoting directional migration towards the injury site. Interestingly, extracellular ATP acts both as a DAMP when released from damaged cells, or as signaling molecule when released from healthy cells. In both cases, ATP activates purinergic receptors (P2) triggering subsequent downstream signaling that depends on the activated receptor.
P2 receptors are classified into ionotropic (P2X) receptors that allow calcium ions (Ca2+) influx, and metabotropic (P2Y) receptors that trigger Ca2+ release from intracellular stores (18). Both families of P2 are widely expressed on immune cells, controlling a plethora of functions (18), including cell communication. The activation of specific P2X and P2Y receptors family members depends on the exposure time and agonist concentration, which allows the spatiotemporal regulation of the signaling (19). In addition to the differential activation of P2 receptors the concentration of adenine nucleotides, such as ATP, is integrated by immune cells and decoded as low or high inflammatory state (18). Immune cells use different cell communication mechanisms dependent or independent on cell contacts, which amplify signals according to that inflammatory state (2, 6, 7, 20). The release of small molecules via plasma membrane channels, such as connexins (CXs) and pannexins (PANXs), and its coupling to purinergic signaling represents a widely used mechanism for cell communication, which plays a role in paracrine (between cells) and autocrine (cell autonomous) signaling (2, 18, 21).
1.2 Cell Polarity and PANX-Dependent Signaling
CXs and PANXs are membrane proteins that allow the exchange of small molecules (i.e., glucose, ATP) and ions between the cytoplasm and the extracellular milieu (22). Upon docking, CX channels of adjacent cells form intercellular channels that connect their cytoplasm, namely gap junction channels, although PANX channels until now are shown to form only hemichannels at the plasma membrane. The latter puts PANX channels and purinergic signaling in the center stage of both cell-autonomous signaling and contact-independent cell communication (23), which are required for efficient motility. Migrating cells use cellular polarity, the asymmetric organization of intracellular components, as the navigation system that determines the direction of migration (6, 10, 11). Cell polarity is dynamically set by changing the position of organelles, cytoskeleton, and signaling proteins (10, 24). Thus, polarization of the actin cytoskeleton allows the establishment of a front-rear migration axis, which subsequently polarizes other proteins (10). Interestingly, F-actin and its regulator Arp3 directly interact with PANX1 (25–27), suggesting that F-actin polarization and nucleation of new microfilaments might directly control PANX1 localization. Indeed, actin flow and polarization permits the concomitant polarization of PANX1 to the leading edge in migrating immune cells (28–30). Similarly, PANX3 stability at the plasma membrane requires intact actin cytoskeleton (25), suggesting that a similar mechanism might take place during cell migration.
Altogether, the co-polarization of PANXs with the actin cytoskeleton, and their indirect functional impact on microtubules, via protein-protein interaction with a microtubule stabilizer (31), could imply a polarization of the PANX-dependent signaling. Therefore, ATP and other molecules that permeate through PANX-channels will be released in a polarized manner that might sustain the cell polarity and direction of migration (28, 30, 32).
In this review, we summarize the contribution of PANX1 channels during cell migration. The first part of this review is focused on different immune cells, as example of PANX1 channel contribution to leukocyte migration and recruitment. Then, we describe PANX1 and PANX3 contribution to migration of non-immune cells, such as astrocytes, skin cells, and cancer cells, as well as the few available evidence for the PANX2 and its putative role during cell migration. Afterwards, we dedicate a final section of PANX1 contribution to cell migration during neuroinflammatory conditions, and aging.
2 Immune Cell Migration
Most leukocytes use amoeboid migration to move within tissues. This migration mode is characterized by limited adhesion to the extracellular matrix with little (or non-) proteolytic activity, preventing extracellular matrix modification (5). Therefore, in order to undergo fast migration after damage, immune cells must deform their cellular body while facing microenvironment obstacles (6, 9). Leukocytes highly rely on acto-myosin cytoskeleton contractility, and mechanosensitive channels, including PANXs channels (6, 9, 30). Interestingly, PANX1 is required for homing of bone-marrow derived immune cell precursors (33), suggesting that these channels are required from early stages of development. Moreover, since leukocytes reside in different tissues and are exposed to different mechanical and chemical signals, these cells exhibit different migration strategies. Despite this cell-specific migratory behavior, the contribution of PANX1 channels to purinergic and Ca2+ signaling, and to cytoskeleton regulation is well established. In general PANX1 channels are positive regulators of immune cell migration as summarized in this section (Table 1).
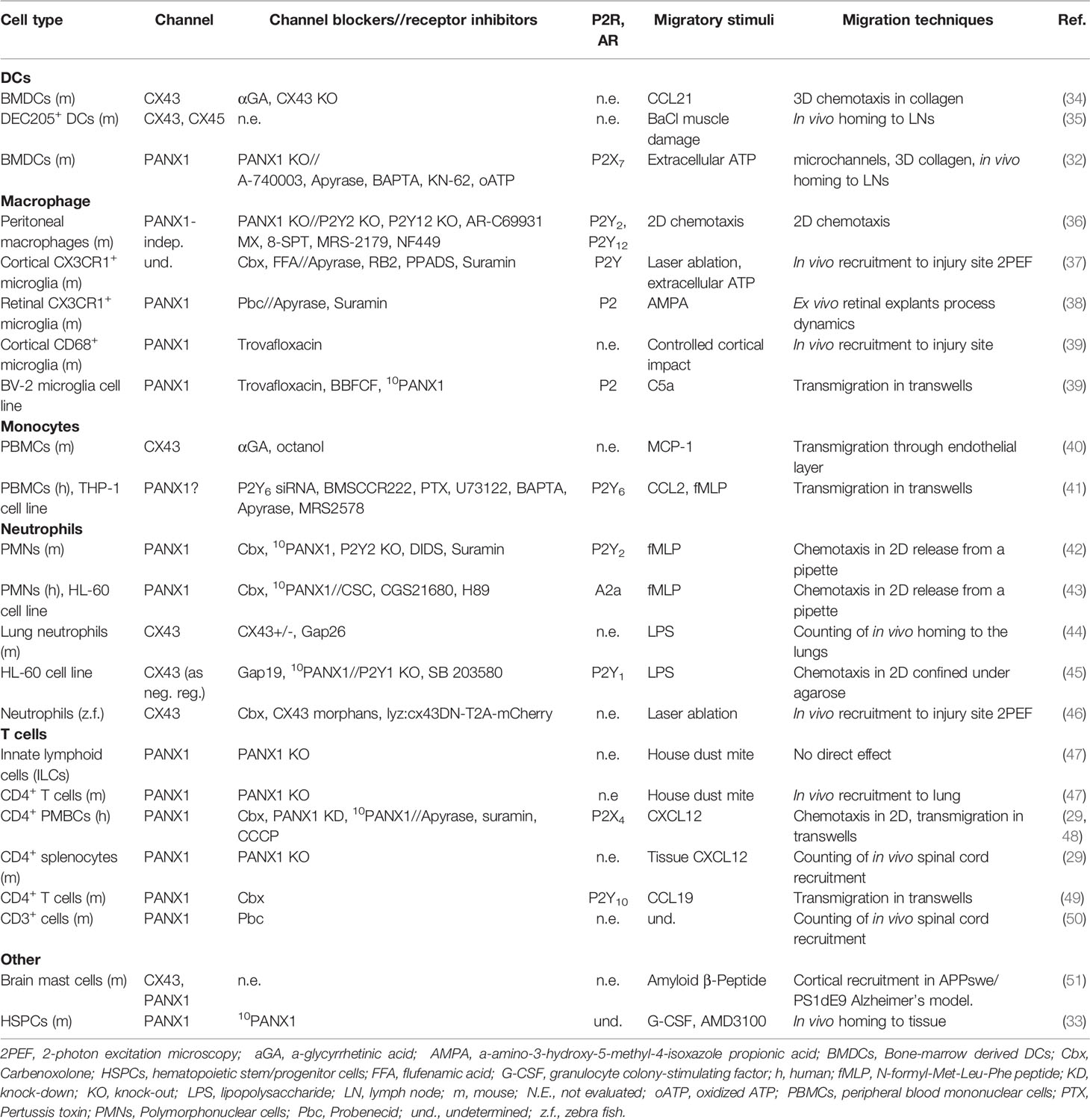
Table 1 Summary of CXs and PANX channels contribution to immune cell migration.
2.1 Neutrophils
Neutrophils, key components of the innate immune system, are polymorphonuclear phagocytic cells found in the bloodstream. Upon danger signal (i.e. chemokine/cytokine detection after infection or injury) neutrophils leave the bloodstream to invade vascular tissues (52). In the tissue, neutrophils have phagocytic activity, producing reactive oxygen species, forming neutrophil extracellular traps (NETs), and releasing cytokines, chemokines and bactericidal peptides (17, 53). PANX1 channels are highly expressed in neutrophils and contribute to their activation (42, 43, 54, 55). For instance, after exposure to non-esterified fatty acids (i.e., oleic and linoleic acid), NET formation requires activation of P2X1 receptors by extracellular ATP, released via PANX1 channels (55).
Indeed, adenine nucleotide release and purinergic signaling cascade activation are key for neutrophil migration, particularly when the release of ATP is used as a navigational cue (43, 54). Accordingly, several purinergic receptors regulate neutrophil migration, including P2X1 (45), P2Y2 (42, 54, 56), P2Y6 (57), P2Y11 (58), P2Y14 (59), A1 (60), and A2a (61). Interestingly, components of the purinergic signaling that contribute to migratory response are polarized (Figure 1), and therefore provide a spatio-temporal dimension to this phenomenon (43, 54).
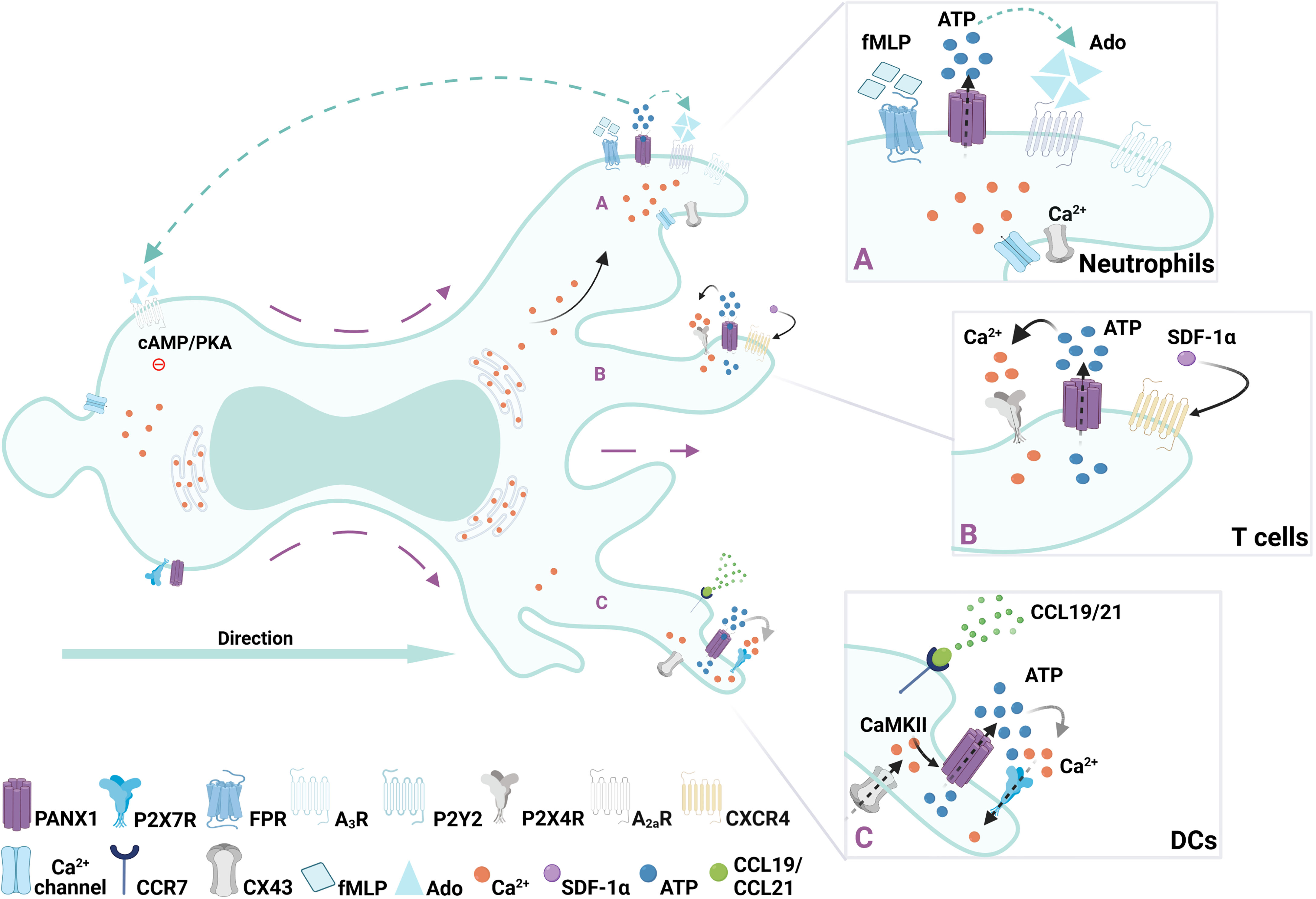
Figure 1 Regulation of immune cell migration by PANX1 channels. Redistribution of surface PANX1channels have been described during migration of immune cells. (A) In neutrophils after FMLP gradient sensing, opening of PANX1 channels polarize towards the leading edge (right side of the cell) allowing ATP release, subsequently activating local purinergic P2Y2 and -upon ATP degradation- adenosine A3 receptors. Then, at the rear of the migrating cell, activation of the adenosine A2a receptors by adenosine (Ado) promotes inhibitory cascades mediated by cAMP/PKA, leading to an orchestrated cytoskeleton rearrangement required for migration. (B) In T cells activation of CXCR4 receptor with SDF-1 triggers controlled burst of ATP after PANX1 channels opening, which is accompanied by mitochondria and P2X4 translocation to the leading edge where Ca2+ influx occurs. (C) In DCs, ATP-induced fast migration requires an autocrine feedback loop between PANX1 channels and P2X7 receptor, which triggers Ca2+ influx and subsequent activation of CaMKII, which maintains PANX1 channels opened.
Neutrophils migration towards a N-formyl-Met-Leu-Phe (FMLP) gradient, which mimics a bacterial-induced chemotactic response, also depends on extracellular ATP sensing. During FMLP-induced migration ATP is released in a polarized manner, as PANX1 is polarized to the leading edge together with F-actin (42). This polarization towards the leading edge relies on the direct interaction between F-actin and the C-terminus of PANX1 previously described (25, 27). ATP released via PANX1 leads to the subsequent activation of P2Y2 and A3 receptors, which are also localized at the leading edge generating an autocrine feedback loop required to maintain the polarization of the cell and to amplify the gradient sensing (42, 54). Interestingly, PANX1 channels also contribute to the inhibitory signals at the rear of the cell (43). The continuous degradation of ATP by ectonucleotidases produces adenosine, which activates adenosine A2a receptors at the cell rear, leading to intracellular cAMP/PKA signaling, and inhibiting excitatory signals from the leading edge (43). Since adenosine activates both A3 and A2a, PANX1 indirectly contributes to excitatory and inhibitory signals required for cell migration, as shown in Figure 1. So far, polarization of positive and negative signals regulated by PANX1 has only been reported on neutrophils, although is tempting to assume it as a general mechanism for immune cell migration.
Moreover, neutrophils present additional pathways to release ATP, such as CXs and tweety family member 3 (TTYH3) maxi anion channels (2, 42), but it seems that different channels and purinergic receptors are recruited depending on the stimuli of the immune response. For example, in a mouse model CX43 hemichannels contribute to LPS-induced neutrophil recruitment in the lungs (44). Similarly, CX43 hemichannels provide a path for ATP release that promotes neutrophil swarming during laser wound tissue damage in zebrafish (46). However, when CX43-mediated ATP release is coupled to activation of P2X1 receptors, this reduces the migration of human neutrophils and HL-60 neutrophil-like in an under agarose assay, and then acts as a stop signal (45).
Overall, PANX1 contributes to neutrophil cell migration by establishing a polarity axis, which is supported by re-localization of the actin cytoskeleton, and the purinergic signaling-related proteins involved in the migratory response. However, the putative role of PANX2 and PANX3, and in pathological conditions remains largely unexplored.
2.2 T Cells
T cell migration is a key step during the adaptive immune response, and its pattern varies with the activation state and the microenvironmental context. Before antigen exposure, and during antigen-presenting cells (APC) search in the lymph nodes, T cells have a diffusive and random migration (Brownian type), whereas less diffusive chemotactic movements (Lévy type) is exhibited by recently activated T cell to migrate into secondary lymphoid organs for priming; or highly directional migration (ballistic) induced by haptotaxis cues and chemotaxis gradient caused by cognate APC at the peripheral tissues (8). Adaptive immune response progression requires purinergic signaling to modulate T cell functions (28). For example, P2X7 receptor modulates the balance between the number of differentiated Th17 and Treg lymphocytes (62). Accordingly, PANX1 and purinergic receptors are essential for T cell activation and cell death (63–65). In both cases, ATP is released through PANX1 channels activating purinergic receptors, triggering intracellular cascades, and inducing the corresponding T cell response.
Acute chemokine stromal-derived factor 1α (SDF-1α, also known as CXCL12) stimulation induces PANX1 channel opening, cell polarization, and migration of CD4+ immature T cells (29, 48), as shown in Figure 1. Particularly SDF-1a recognition leads to a well-controlled PANX1 channel opening by a G protein-coupled receptor mechanism, leading to a rapid burst of ATP release, and subsequent focal adhesion kinase (FAK) phosphorylation (29). In this context, SDF-1α promotes translocation of P2X4 and mitochondria to the leading edge, increasing Ca2+ influx and pseudopod protrusion needed for cell polarization and migration (48). Polarized at the back of the cells, P2Y11 receptors are also activated, triggering inhibitory signaling via cAMP/PKA activation (Figure 1), and preventing mitochondria activity at the cell rear (66). Thus, polarized migratory T cells might present mitochondria at both leading edge (48), and/or at the cell rear (67), suggesting that specific chemical (i.e. chemokine treatments) and physical (i.e. adhesion of the surface) cues of the microenvironment differentially shape intracellular organelle location and migratory function. In fact, supporting the role of PANX-mediated signaling in T cell polarity, protein-protein interaction of PANX1 and collapsin response mediator protein 2 (CRMP2), indirectly controls microtubule stability (31). Interestingly, CRMP2 polarization towards the cell rear is required for T cell chemotaxis (68).
In freshly isolated naïve T cells subsets, CD4+ and CD8+ cells, PANX1 is abundantly present at the plasma membrane, whereas PANX2 abundance is much lower (65). Therefore, it is conceivable that upon stimulation the abundance of PANX2, and/or PANX3, could increase under specific conditions. Accordingly, in vivo data in murine model of experimental autoimmune encephalomyelitis disease suggests that stimulated T cells require functional PANX channels to transmigrate into the spinal cord (29, 50). PANX contribution during cell migration of specific T cell subsets remains largely unknown, but there is evidence that regulatory T cells required PANX1 to downregulate the response of effector T cells in vivo in a model of allergic airway inflammation (47). The latter data is very provocative, because it suggests that when PANX1 acts as a cell communication effector protein, it contributes to limit immune cell recruitment (47). Alternatively, PANX1 could promote T cell infiltration when is acting as a migration effector protein, likely by sustaining local signaling, which subsequently permits the polarization of the actin cytoskeleton or microtubule stability. Whether this migratory versus cell communication role of PANX1 is affected by the expression of other PANXs, or whether this is a mechanism present in other cell types is yet to be explored.
2.3 Dendritic Cells
Immature dendritic cells (DCs) normally reside in peripheral tissues where they scan the microenvironment in search of danger signals (69, 70). After antigen uptake, DCs migrate to secondary lymphoid organs, to initiate adaptive immune responses (69, 70). Resident DCs constantly internalize extracellular material, by phagocytosis or micropinocytosis, but upon danger signal detection these processes are downregulated and the migratory strategy changes from slow/random to fast/persistent (6, 30). Purinergic signaling is key for DCs response, as several P2X and P2Y receptors are expressed. In particular, P2X7 receptor together with PANX1, are required to sustain fast migratory phases (30, 32, 71). Transient exposure to high extracellular ATP concentrations activates P2X7 receptor, opening PANX1 channels to release ATP establishing an autocrine loop (32). This autocrine loop triggers Ca2+ influx via P2X7, calmodulin kinase II (CaMKII) activation and F-actin cytoskeleton to the cell rear (Figure 1), and is necessary to sustain DC fast migration (32). Interestingly, CaMKII directly controls the opening of PANX1 channels providing a direct link between purinergic and Ca2+ signaling that might be responsible for maintaining DCs migration (72). PANX1 appears to be equally distributed in migrating DCs, suggesting that this protein does not need to polarized to control DC speed. Since CaMKII regulates actin dynamics (73), its activity directly impacts on acto-myosin contractility, which is required for DC migration (30, 74). Thus, PANX1 contributes to DC migration by sustaining Ca2+ signaling via P2X7, which activates CaMKII maintaining actin polarization at the cell rear and subsequent contractility.
Activating signals trigger a maturation program in which several phenotypic changes, including the decrease of macropinocytosis, are required for the fast migration of DCs (6, 69). Interestingly, in other cells (i.e. neuroblastoma) ATP induces the internalization of PANX1 into macropinosomes (75), but it is not known whether the exact mechanism takes place during DC maturation and migration. Upon maturation, DCs increase the expression of cell surface molecules related to antigen presentation and directional migration, such as CCR7 chemokine receptor (6, 69). Directional migration in DCs largely depends on CCL19/CCL21 activation of CCR7 with the concomitant activation of CX43 channels (34). In the same line, migratory DCs increase the expression of CX43 and CX45 during homing to lymph nodes (35). Conversely, PANX1 channels were dispensable for this response as shown by the lack of effect observed with PANX blockers (34).
These data suggest that PANX1 channels might contribute to the early stages of DC migration upon danger signal detection, and that later stages including CCR7-dependent chemotaxis relies in the activity of CXs. This putative functional distinction between CXs and PANXs during DC migration might rely in different protein turnover at the plasma membrane, and/or the non-channel signaling function, as is shown for CX43 (76).
2.4 Monocytes and Macrophages
Although monocytes and macrophage originate from a common myeloid precursor and share several markers, they differ in their location, as monocytes are generally found in the bloodstream, while macrophages are tissue-resident cells. However, circulating monocytes differentiate into monocyte-derived macrophages (MMs) or monocyte-derived DCs (Mo-DCs) upon sensing of danger signals at the injury site (77). During CCL2-induced chemotaxis, monocytes release ATP, thus generating an autocrine loop with subsequent activation of P2Y6 receptors that is required for efficient migration (41), similar to other cells (see section Dendritic Cells, DCs). However, the molecular mechanism for ATP release was not elucidated. Monocytes, and macrophages, express both CXs and PANXs, but only CXs role has been shown during extravasation and migration. Particularly, CXs contribute by forming gap junction channels between monocytes and the endothelium (2, 40, 78). However, the functional role of PANXs expressed in monocytes (22), remains unaddressed.
Resident macrophages are named according to the different tissues in which they reside, although in general these cells share their primary functions: cellular detritus clearance, phagocytosis of pathogen particles, and in a lesser extent, antigen presentation (79). Resident brain macrophages, named microglia, quickly reacting to danger signals to prevent neuronal damage (80). ATP released during tissue damage acts as a chemoattractant of microglia in vivo, which extend their processes towards the injury site, a response that might require PANX1 channel activation (37). However, this need to be confirmed as only general blockers were used. Similarly, retinal microglia process extension induced by α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptor activation is prevented with Probenecid, a general PANX channel blocker (38). In addition, trovafloxacin another proposed PANX1 blocker (81), reduces microglia recruitment after traumatic brain injury (39). In a similar model PANX1 is required to induced the recruitment of microglia and other myeloid cells, and the lack of PANX1 improved the posttraumatic recovery of the mice (82). In addition, C5a-induced transmigration of microglia depends on PANX1-dependent ATP release, likely via an autocrine loop (39), such as the one shown in DCs (see section Dendritic Cells, DCs). However, this response might be specific different macrophage subsets, as PANX1 channels have only a mild contribution to C5a-induced chemotaxis in peritoneal macrophages, which was dependent on P2Y2 and P2Y12 receptors (36). Conversely, CX43 contributes to LPS-induced migration of peritoneal macrophages (83), suggesting that different channels might be required under other conditions.
Depending on the chemical cues of the microenvironment (i.e. cytokines), macrophages polarize to M1 or M2 macrophages, which exhibited different migratory properties (84). Interestingly, macrophage M1 polarization reduces PANX1 expression, whereas M2 polarization induces its upregulation (85). However, whether PANX channels play a role during the migration of these cells is still unknown.
3 Role of Tissue PANXs on Leukocyte Recruitment
Immune cell migration is not only induced by activation of PANX expressed in the migrating cell, but also can be indirectly promoted by PANX channels activated in the tissue. Initial observations by Chekeni et al, revealed that PANX1 channels were required for the release of “find me” signals (i.e., ATP and UTP) during T cell apoptosis, which triggered monocyte recruitment (86).
In liver, recruitment of monocytes is a hallmark of hepatic inflammation, involving apoptosis of hepatocytes induced by saturated free fatty acids (lipoapoptosis). Exposure to lipoapoptotic supernants elicits monocyte recruitment in an ATP-dependent chemokine-independent manner (87). PANX1 channels release ATP during lipoaptosis leading to c-Jun NH2-terminal kinase (JNK) activation in liver cells, revealing that hepatocytic PANX1 is key regulator of immune recruitment during nonalcoholic steatohepatitis (NASH) progression. In the same line, during obesity progression there is also accumulation of unsaturated fatty acids, which induce skeletal muscle inflammation and recruitment of immune cells. Thus, in vitro experiments with a muscle cell line that form myotubes show that treatment with palmitate induces ATP release from myotubes, which triggers monocyte recruitment (88). Interestingly, the release of ATP from the myotubes was independent of PANX1, but dependent on PANX3 channels (88). Consequently, myotubes that lack of PANX3 are unable to release ATP upon palmitate treatment, and do not trigger monocyte migration (88). A similar mechanism could occur during wound healing in a dorsal skin mouse model (89). PANX3 KO mice presented a delayed healing and inflammation resolution at the injury site. Indeed, the number of CD4+ T cells, neutrophils and macrophages was reduced in PANX3 KO mice, suggesting that tissue PANX3 was required for immune cell recruitment (89).
In the central nervous system the choroid plexus, located in the brain ventricle, is a key immune barrier between the cerebrospinal fluid and the blood. Epiplexus cells, resident innate immune cells of the choroid plexus, share markers and function with macrophages, DCs and other phagocytic cells (90). Under resting conditions epiplexus cells are sessile, but upon detection of extracellular ATP these cells increase their motility (91). This response depends on the ATP release via PANX1 channels from the epithelium of the choroid plexus, although epiplexus do not express PANX1 (91). Whether if during chronic inflammation or infection PANX expression is induced remains unknown.
Moreover, adipocyte-derived ATP release during adrenergic stimulation triggers macrophages recruitment (92). In addition, PANX1 opening is required for insulin-stimulated glucose uptake in adipocytes (93). Since insulin activates PANX1 channels causing the release of ATP, which in turn results in a signaling cascade indirectly allowing the transport of glucose into adipocytes, PANX1 might play a role in sustaining the inflammation observed during insulin resistance.
4 Role of PANXs on Migration of Non-Immune Cells
Unlike most immune cells that alter and deform little/transiently the extracellular matrix, mesenchymal cells require proteolytic enzymes to modify the microenvironment to undergo migration (5). Moreover, mesenchymal migratory cells use their actin cytoskeleton, form focal adhesion, and align with the extracellular matrix, with which and form focal adhesion (5, 10). However, despite these differences with amoeboid cell migration, mesenchymal cell migration also depends on acto-myosin contractility to move, although the motility of these cells is significantly slower (10). Another key protein for migration is the extracellular-signal-regulated protein kinase (ERK), a mitogen-activated protein kinase (MAPK), is a serine/threonine kinase, which modulates migration through phosphorylation of myosin light chain kinase (MLCK), calpain protease, paxillin, and focal adhesion kinase (FAK) (94). ERK activation can be triggered by cell matrix proteins (fibronectin, vitronectin, and collagen), growth factors (VEGF, FGF, EGF, insulin) and also indirectly by mechanical stress (94, 95). Indeed, mechanical stretch activates ERK through EGF receptor activation, triggering cell contraction (95). PANX1 channel opening is also affected by mechanical stress, although some evidence suggests that this occurs in an indirect fashion (96). However, regardless of the pathways activated PANX channel activity is affected by mechanotransduction and therefore these channels might contribute during cell migration and deformation of extracellular matrix, as it occurs during mesenchymal migration.
During the past years, increasing interest have grown on study PANX1 role during mesenchymal migration, particularly in the context of cancer progression (Table 2). For this reason, we decided to include in the following section, the latest publications associating PANX1 with non-immune cell migration.
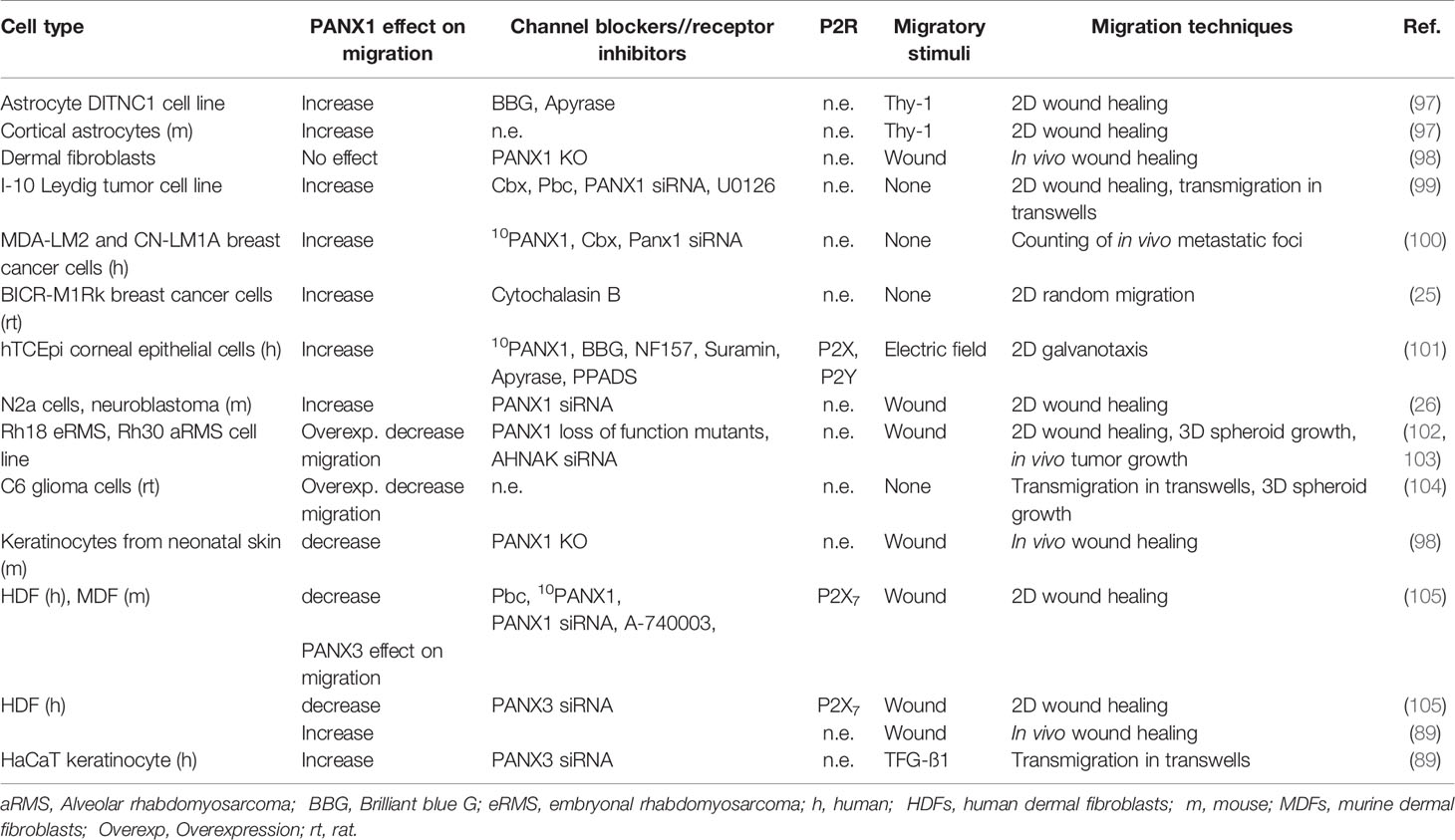
Table 2 Summary of PANX1 contribution to non-immune cell migration.
4.1 Fibroblasts and Keratinocytes, Role of PANXs During Skin Cell Migration
The skin forms an active barrier and provides the first layer of defense by preventing the entry of foreign agents. This organ is a complex multilayer organization of cells with different but complementary functions, such as keratinocytes, melanocytes, fibroblasts, and immune cells (106, 107). During tissue damage skin resident cells communicate by using contact-dependent and independent mechanisms to quickly heal the wound (108). Consequently, ATP release and associated purinergic signaling play a key role during skin inflammation and wound healing (108). In particular, PANX1 is expressed in keratinocytes and fibroblasts from human and mice skin, as discussed below (98, 105, 109–111).
PANX1 levels decrease during adulthood, but increase after tissue damage (98). PANX1 is required for skin wound healing, as shown in a murine model of skin punch biopsies with a lack of PANX1 reduced tissue repair, but simultaneously increased fibrosis (98). Surprisingly, in vitro experiments revealed that the lack of PANX1 in keratinocytes increases their migratory potential. In contrast isolated skin fibroblasts from PANX1 knockout mice were more proliferative but showed decreased contractile properties in comparison to control conditions (98), suggesting that PANX1 expression is cell type specific, and that tissue interaction controls the overall migratory response in a cell-type specific and tissue specific manner. In the same line, PANX1 negatively regulates human dermal fibroblast migration when kept in monocultures. Indeed, fibroblasts lacking PANX1 or those treated with PANX1 channel blockers increase their speed of collective migration in wound healing assays, a similar but smaller response occurs when cells lack PANX3 (105). The decrease in cell migration during wound healing in fibroblasts is linked to decreased ATP release and activation of purinergic receptors. Consequently, P2X7 receptors blockade increases the speed of migration in human dermal fibroblasts (105). However, this last data should be considered carefully depending on the working model to be compared with, as the authors also report no effect over migration with P2X7 blockade in murine dermal fibroblasts (105). Additionally, in vivo experiments in a mice model reveal that PANX3 was required for proper wound healing (89). This suggests that different models of study might require different purinergic receptors. In any case, PANX1- and PANX3-dependent ATP release was consistently associated with a decrease in the collective and single migration of dermal fibroblasts, a response that relies in reorganization of the actin cytoskeleton (105).
These data reveal that PANX1 channels contribute to cell migration as a positive or negative regulators depending on the cell type and components of the microenvironment, including the available adenine nucleotides (Figure 3). However, the analysis of specific downstream signaling, and how the chemical cues of the microenvironment affect the role of PANX1 during migration remains largely unexplored.
4.2 PANXs Role During Astrocytic Migration Under Inflammation
Astrocytes are the more numerous glial cell in the brain, where these cells protect and feed the neurons (112). Astrocytes are crucial for tissue repairing during brain injury, and avoid spreading of neuronal damage by glial scar formation (112). During inflammation reactive astrocytes exhibit functional and morphological changes, as well as an increase in the expression of DAMPs receptors (112). Neuronal interaction with astrocytes controls cell migration via direct interaction of membrane proteins, such asThy-1 (CD90) (113, 114). Thy-1 is a membrane glycophosphatidylinositol (GPI) anchored protein that binds to αVβ3-containing integrin and syndecan-4 to stimulate FAK and actin reorganization (i.e. stress fibers formation), leading to morphological changes and migration in DITNC1 cell line. This astrocyte like cell line express high levels of αVβ3 Integrin and Syndecan-4, which resemble those observed in reactive astrocytes after tissue damage (115, 116). In DITNC1 cells Thy-1 stimulation triggers activation of intracellular signaling (PI3K and PLCγ) leading to Ca2+ release from intracellular stores opening CX43 and PANX1 channels. The ATP released via these channels activates P2X7 receptors and subsequent Ca2+ influx (117, 118), revealing that Thy-1 induction of DITNC1 cell migration depends on PANX1 channels.
Under resting conditions primary astrocytes express very low levels of αVβ3 Integrin and Syndecan-4, but their expression is induced during neuroinflammation (97). TNF is a cytokine associated with neuroinflammation and accordingly triggers expression of αVβ3 Integrin and Syndecan-4 in astrocytes, accompanied by PANX1, CX43, P2X7 receptors allowing the establishment of molecular toolkit required for Thy-1 signaling (97). Indeed, TNF-stimulated astrocytes respond to Thy-1, which leads to astrocyte cell migration by triggering PANX1 and CX43 ATP release and subsequent P2X7 receptor activation (Figure 2) (97), supporting the data obtained in DITNC1 cells.
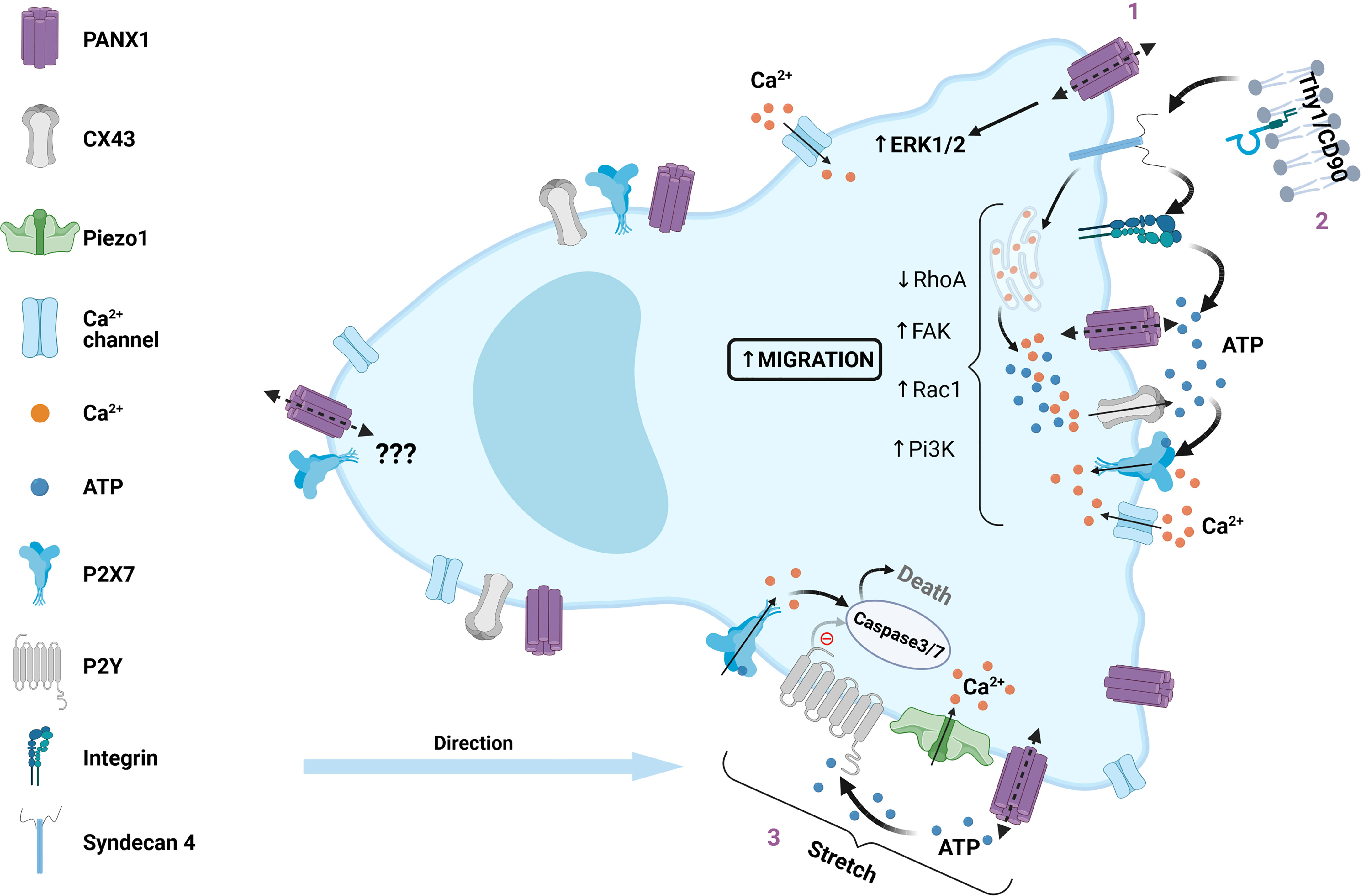
Figure 2 Role of PANX1 channels in mesenchymal cell migration. Intracellular regulation of PANX1 channels during migration of mesenchymal non-immune cells is described as a positive or negative regulation, here we summarize the positive regulation. (1) Increased cell migration is correlated with PANX1 overexpression via activation of ERK1/2 pathway, which could also play a major role during collective cell migration (not included in this figure). (2) Thy1/CD90, a surface protein expressed in normal cells, but overexpressed in certain pathologies, interacts with αVβ3-containing integrin and Syndecan 4, which promotes cell migration. The downstream signaling involves focal adhesion kinase (FAK) activation, PI3K, PLCγ and IP3 production, leading to Ca2+ release from intracellular stores, and opening of both CX43 and PANX1 channels. The latter promotes extracellular ATP release and subsequent P2X7 activation, Ca2+ influx and PKCα activation leading to Rac1 activation and RhoA inhibition. (3) Mechanical deformation during transepithelial migration lead to PANX1 channel activation, releasing ATP and activation of P2X7 receptors, leading to cell death by caspase 3/7 activation. Alternatively, PANX1 mutations that change its function lead to P2Y activation, preventing caspase activation and cell death.
In another pro-inflammatory context, reactive astrocytes derived from Amyotrophic Lateral Sclerosis (ALS) model hSOD1G93A transgenic mice present an increased abundance of several migration-related molecules, including αvβ3 Integrin, syndecan-4 proteoglycan, P2X7 receptor, PANX1, and CX43 (97). Thy-1 recognition induced both adhesion and migration of hSOD1G93A astrocytes (97). Intriguingly, TNF stimulation, and in ALS models triggers Thy-1 associated signaling expression, which pre-set a migratory phenotype in astrocytes, to which it is possible to speculate that, in general, pro-inflammatory conditions will induce a similar response preparing reactive astrocytes to migrate if needed, but whether PANX channels play a role during astrocyte migration in all pro-inflammatory conditions will require further studies.
4.3 Cancer Cells, Differential Contribution of PANXs to Tumor Progression
Tumor progression involves a series of sequential steps, which lead to tumor growth and metastasis. Cancer cell migration is the key step that allows invasion and colonization of new tissues. The stimulation of this response occurs during the epithelial to mesenchymal transition (EMT) (119, 120). Interestingly, dichotomic contribution of PANX1 on cancer cell migration has been reported (summarized in Table 2). Particularly, PANX1 can acts as a negative regulator for C6 cells motility, which are derived from rat glia. Indeed, overexpression of PANX1 reduces C6 glioma cell migration in different levels of complexity models of study (i.e. 2D, 3D spheroids, and in vivo) (104). In the same line a similar response is observed in rhabdomyosarcoma cells, in which inducible expression of PANX1 prevents cell migration (102). However, in this model PANX1 seems to play a role independent of its channel function, because it requires PANX1 physical interaction with AHNAK, a large scaffold protein (102, 103). Thus, at least in this rhabdomyosarcoma model it seems that PANX1 contribution might be related either to cytoskeleton re-organization and signaling, as shown for CX43 C-terminus (76). Interestingly, PANX1 expression induces gene and protein level upregulation of CX43, which has a tumor suppressive role in rhabdomyosarcoma (121). Altogether, these data supports the notion that PANX1 is a negative regulator tumor suppressor factor in cancer cells. However, in other cancer cell lines, PANX1 acts as a pro-migration factor as we discuss below (Figure 3).
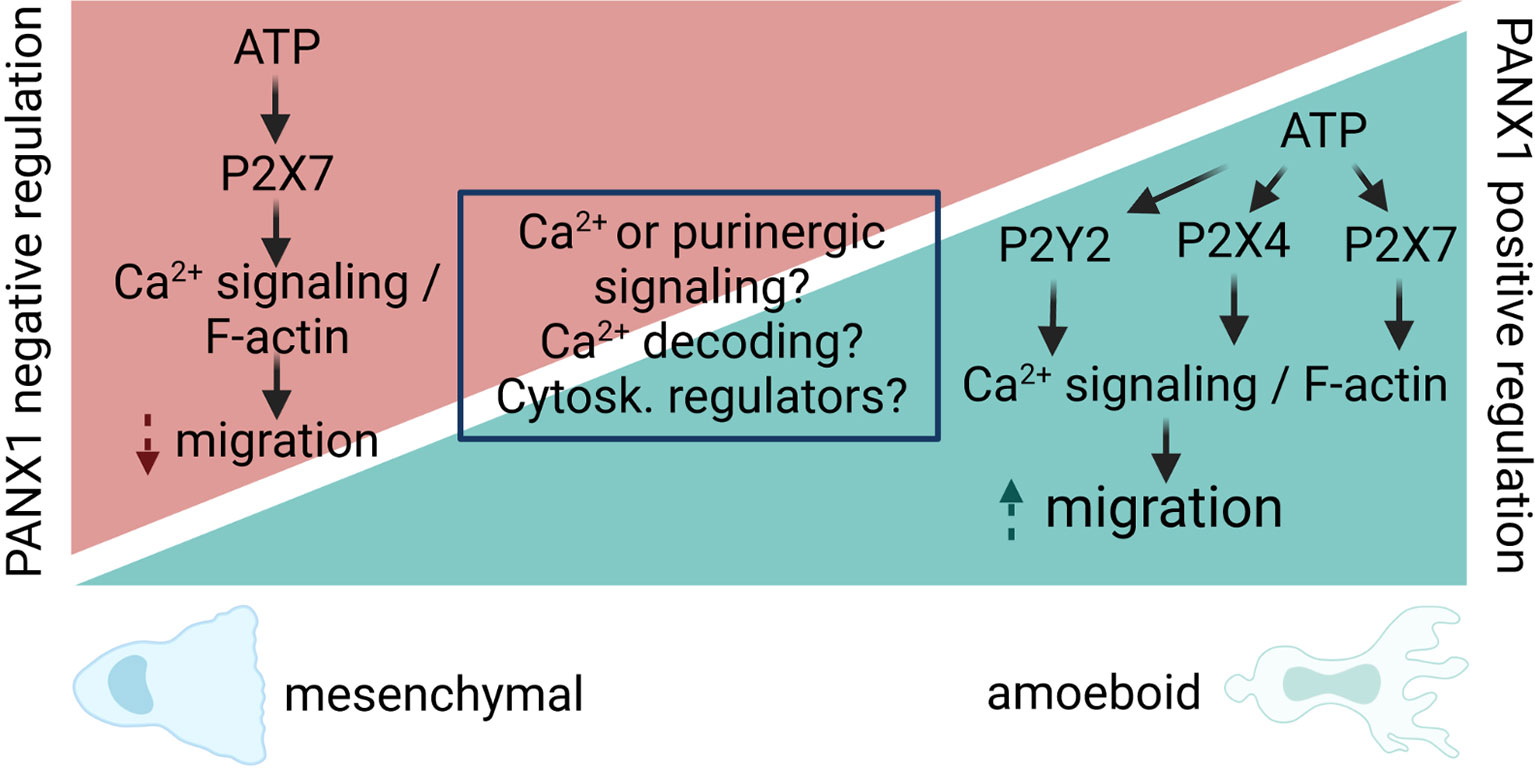
Figure 3 Proposed model for PANX1 channel regulation of cell migration. PANX1 channels act as positive or negative regulators of cell migration, despite the fact that some pathways and proteins are shared by those opposite responses. We propose that different outcomes are likely to occur due difference in Ca2+ and/or purinergic signaling (i.e. agonists, concentrations, etc). In particular, for Ca2+ signaling, the local (microdomain, nanodomain) regulation of it might activate different signaling cascades leading to increase or decrease of cell migration. In addition, the decoding of Ca2+ signals by Ca2+-sensitive enzymes (i.e. CaMKII) could directly modify the opening of PANX1, leading to changes in cytoskeleton dynamics directly or indirectly (i.e. cytoskeleton -Cytosk.- regulators) will result in signaling cascades that promote or inhibit cell migration. Therefore, the main contribution of PANX1 channels would be to amplify the initial response, and this would be downstream of the intrinsic cell-type specific Ca2+ response that ultimately determines the migratory outcome.
Pioneer studies that revealed PANX1 interaction with the actin cytoskeleton, suggested its pro-migratory phenotype in breast cancer (25). Indeed, PANX1 promotes motility, transmigration, and in vivo invasion in melanoma, breast and testicular cancer cells: B16-BL6, B16-F10; CN34, CN-LM1A, MDA-MB-231 MDA-MB-468, MDA-LM2, and I-10 cells, respectively (99, 100, 122, 123). In some cases, such as testicular cancer PANX1 activity was required for ERK1/2 activity, E-cadherin and metalloproteinase 9 (MMP-9) revealing that these channels contribute with different aspects of cell migration (99).
Like normal fibroblasts (see section above), MDA-MB-231 cells are sensitive to Thy-1 that increases cell migration by releasing ATP via PANX1 channels with subsequent activation of P2X7 receptors in a positive feedback loop (123). Consequently, a mutation that increases the permeability of PANX1 channels leading to an exacerbated ATP release, also promotes cell motility, but not trans-endothelial migration (100), suggesting that PANX1 contributes to specific steps during tumor progression.
Which are the signals that lead to a positive or negative modulation of PANX1 on migration? As reviewed in this section, we hypothesize that it might depend on the cell type (Table 2), but the exact molecular mechanisms and signaling pathways that determine the outcome of the response remain unknown (Figure 3). The involvement of the other membrane channels that share functional and structural similarities with PANXs, such as the Leucine-rich repeat-containing 8 (LRRC8) proteins (124), is largely unexplored. Indeed, LRRC8A acts as a positive regulator of cancer cell migration (125), but how the activity of these channels affect the opening of PANXs, and how LRRC8 protein expression changes during cancer progression is not yet shown.
4.4 PANXs and Cell Migration During Aging, Senescence, and Neurodegeneration
Cellular changes progressively alter immune function during chronic inflammation and natural ageing, increasing susceptibility to infections and tumors (126). Associated with a chronic low-grade inflammatory state, is well accepted that cell motility decreases over time, which on immune cells might be due to accumulation of DNA damage after nuclear breakage (127). However, studies of cell migration in aging models are still scattered. Impaired phagocytosis and migration of DCs have been reported in aged humans (128). Not only cell decline (129), but also naive CD8+ and CD4+ T cell from old mice present lower migration and microtubules gene expression (130). Migration of aged marginal zone B cells at the spleen has also been reported to be impaired, consequently affecting immunoglobulin production (131). Human monocytes form elder volunteers showed altered gene profile of cellular motility (132), while bone marrow mesenchymal stem cells from aged human donors also present lower proliferation and migration abilities (133).
In the central nervous system, with smaller branches and slower motility process, microglia from aged mice exhibit reduced protrusion activity and cell migration after acute injury (134). During chronic neuroinflammation, aged mice of an Alzheimer’s disease model present increased mast cell infiltration in the brain parenchyma (51). Conversely, aged neutrophils migrate faster to the injury (135, 136), despite having impaired phagocytic and degranulation activity (137, 138). Similarly, myoblast have augmented migratory features (speed and directionality) during wound healing (139). Aging is likely to exert a cell-specific effect, and therefore it is hard to anticipate whether PANX1, and other PANXs, will contribute as a positive or a negative regulator as this will depend on the spatiotemporal regulation and accumulation of different ligands in the microenvironment.
5 Concluding Remarks
In the present review, we summarize the contribution of PANX channels during cell migration, emphasizing PANX1, that has been more widely studied. We have focused on immune cells as the integration of cell communication and cell migration is key for their function, despite mainly migrating as single cells. An interesting aspect of PANX-dependent purinergic signaling is the possibility that single migrating cells would have an impact on their neighbors. Using a mathematical model, Agliari et al. explored the hypothesis that ATP release and autocrine signaling during immune cell migration might impact neighboring cells while migrating as a group of single cells (140). The statistical inference approach revealed that migrating DCs have no instantaneous cell communication via release of small soluble molecules, such as ATP (140). However, the model only predicts immediate interactions and the release of adenine nucleotides or other small molecules will act with a delay considering the diffusion time and other parameters. The latter shows the need for the simultaneous study of cell communication and cell migration in a coordinated manner by using computational methods and theoretical frameworks, as it has been done for chemotaxis, or collective cell migration (141–143).
Another key aspect to link PANX1 and cell migration is the direct association between PANX channels and cytoskeletal components (actin, MyoII, microtubules), which are master regulators of cell migration. There is evidence of direct and indirect modulation of PANX channels by the cytoskeleton, which modifies membrane dynamics (144). In addition, PANX1 channels are somehow mechanosensitive, although in a lesser extent in comparison to other channels such as Piezo1 and Piezo2 (145). Still PANX1 could quickly react to changes in the membrane tension (96, 100, 146), providing a fast feedback mechanism in which the opening of the channel can be controlled by the mechanical cues of the microenvironment that surrounds the migrating cell. This could be sustained in time by the activation of enzymes that directly modify the opening of the channel, such as Ca2+-sensitive CaMKII (72), or by a positive feedback with P2 receptors (147, 148) (Figures 2, 3). In addition, some evidence suggests a non-channel contribution of PANX1 during migration of rhabdomyosarcoma (102, 103), which has been observed in other channels that can act as signaling proteins such as CX43 (76), or as enzymes and therefore receive the name of ‘chanzymes’, such as TRPM7 (149).
PANX channels interplay with purinergic signaling and indirectly with Ca2+ signaling is well established (2, 30), but its direct contribution with Ca2+ influx seems to be cell specific (32, 51, 65, 150). Then, local Ca2+ signaling regulation could lead to different migratory outcome (Figure 3). Moreover, whether other ions, such as K+, could be relevant for the migratory is not yet demonstrated, although PANX1 channels directly contribute to migration induced by changes in the electric field, process named galvanotaxis (101). On the other hand, whether the opening of PANX channels contribute to ion flux and membrane voltage of the migrating cell is yet unexplored.
It is tempting to speculate about the necessity of PANX polarization during migration of leukocytes, but this should be carefully verified for each cell and stimuli. In neutrophils there is PANX1 polarization during migration (43, 86), but the same is not clear in T cells (29), and polarization seems to be not required for fast DC migration (32) (Figure 2). Therefore, the use of recently developed techniques, such as super resolution, optogenetics, and the development of new tools for live cell imaging monitoring of PANXs, will greatly improve our understanding of their role during cell migration.
Lastly, most of the studies have focused on PANX1, which seems to be a leading player during cell migration, but it is unclear the role of the other PANXs. Do PANXs have redundant functions? Are these cell-type specific? For example glioma cell migration is unaffected when changing the expression of PANX2, although cell growth was directly impacted (151). These data suggests that PANX2 and PANX3 might have other roles unrelated to cell migration and might be associated to cell growth and volume as recently reviewed (152). In the case of PANX3, which seems to act as a negative regulator of collective cell migration, it will be interesting to explore whether also prevent single cell migration. Finally, the role of PANXs has not yet been elucidated during chronic inflammation (i.e. obesity) or during aging (23, 152), conditions that change the responsiveness of immune cells (153). We propose that both conditions, chronic inflammation and aging, might induce the expression of different PANX isoforms in immune cells, leading to an increased migratory potential. However, how different PANXs isoforms orchestrate single and collective cell migration is still an unexplored field.
Author Contributions
PH wrote large parts of the manuscript and provided advice for the figures. TL-L wrote some sections of the manuscript, and drafted the figures. AP contributed to drafting the manuscript. PS conceived the review, wrote and edited the manuscript, and revised the text and the figures. All authors contributed to the article and approved the submitted version.
Funding
This project has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 953489, ITN EndoConnect (PS), MINECO Spain PID2020-116086RB-I00 (PS), Forschungszentrum Medizintechnik Hamburg (FMTHH) 04fmthh2021 (PS), ANID Postdoctoral Project N° 3200342 (PH), MILENIO ICM-ANID P09-022-F (AP and PH).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to acknowledge David McGrath and Nabil Al Dam for reading the review and providing feedback. The figures of this review were created with BioRender.com, to which BioRender’s Academic License Terms apply.
References
1. Mayor R, Carmona-Fontaine C. Keeping in Touch With Contact Inhibition of Locomotion. Trends Cell Biol (2010) 20:319–28. doi: 10.1016/j.tcb.2010.03.005
2. Sáez PJ, Shoji KF, Aguirre A, Sáez JC. Regulation of Hemichannels and Gap Junction Channels by Cytokines in Antigen-Presenting Cells. Mediators Inflamm (2014) 2014:742734. doi: 10.1155/2014/742734
3. Ma L, Li Y, Peng J, Wu D, Zhao X, Cui Y, et al. Discovery of the Migrasome, an Organelle Mediating Release of Cytoplasmic Contents During Cell Migration. Cell Res (2015) 25:24–38. doi: 10.1038/cr.2014.135
4. Kriebel PW, Majumdar R, Jenkins LM, Senoo H, Wang W, Ammu S, et al. Extracellular Vesicles Direct Migration by Synthesizing and Releasing Chemotactic Signals. J Cell Biol (2018) 217:2891–910. doi: 10.1083/jcb.201710170
5. Yamada KM, Sixt M. Mechanisms of 3D Cell Migration. Nat Rev Mol Cell Biol (2019) 20:738–52. doi: 10.1038/s41580-019-0172-9
6. Kameritsch P, Renkawitz J. Principles of Leukocyte Migration Strategies. Trends Cell Biol (2020) 30:818–32. doi: 10.1016/j.tcb.2020.06.007
7. Miskolci V, Klemm LC, Huttenlocher A. Cell Migration Guided by Cell-Cell Contacts in Innate Immunity. Trends Cell Biol (2021) 31:86–94. doi: 10.1016/j.tcb.2020.11.002
8. Krummel MF, Bartumeus F, Gerard A. T Cell Migration, Search Strategies and Mechanisms. Nat Rev Immunol (2016) 16:193–201. doi: 10.1038/nri.2015.16
9. Vargas P, Barbier L, Saez PJ, Piel M. Mechanisms for Fast Cell Migration in Complex Environments. Curr Opin Cell Biol (2017) 48:72–8. doi: 10.1016/j.ceb.2017.04.007
10. Seetharaman S, Etienne-Manneville S. Cytoskeletal Crosstalk in Cell Migration. Trends Cell Biol (2020) 30:720–35. doi: 10.1016/j.tcb.2020.06.004
11. Kopf A, Kiermaier E. Dynamic Microtubule Arrays in Leukocytes and Their Role in Cell Migration and Immune Synapse Formation. Front Cell Dev Biol (2021) 9:635511. doi: 10.3389/fcell.2021.635511
12. Germain RN, Robey EA, Cahalan MD. A Decade of Imaging Cellular Motility and Interaction Dynamics in the Immune System. Science (2012) 336:1676–81. doi: 10.1126/science.1221063
13. Garcia-Arcos JM, Chabrier R, Deygas M, Nader G, Barbier L, Saez PJ, et al. Reconstitution of Cell Migration at a Glance. J Cell Sci (2019) 132. doi: 10.1242/jcs.225565
14. Sáez PJ, Barbier L, Attia R, Thiam HR, Piel M, Vargas P. Leukocyte Migration and Deformation in Collagen Gels and Microfabricated Constrictions. Methods Mol Biol (2018) 1749:361–73. doi: 10.1007/978-1-4939-7701-7_26
15. Matzinger P. Tolerance, Danger, and the Extended Family. Annu Rev Immunol (1994) 12:991–1045. doi: 10.1146/annurev.iy.12.040194.005015
16. Kono H, Rock KL. How Dying Cells Alert the Immune System to Danger. Nat Rev Immunol (2008) 8:279–89. doi: 10.1038/nri2215
17. David BA, Kubes P. Exploring the Complex Role of Chemokines and Chemoattractants In Vivo on Leukocyte Dynamics. Immunol Rev (2019) 289:9–30. doi: 10.1111/imr.12757
18. Di Virgilio F, Sarti AC, Coutinho-Silva R. Purinergic Signaling, DAMPs, and Inflammation. Am J Physiol Cell Physiol (2020) 318:C832–5. doi: 10.1152/ajpcell.00053.2020
19. Bours MJ, Swennen EL, Di Virgilio F, Cronstein BN, Dagnelie PC. Adenosine 5’-Triphosphate and Adenosine as Endogenous Signaling Molecules in Immunity and Inflammation. Pharmacol Ther (2006) 112:358–404. doi: 10.1016/j.pharmthera.2005.04.013
20. Kienle K, Lammermann T. Neutrophil Swarming: An Essential Process of the Neutrophil Tissue Response. Immunol Rev (2016) 273:76–93. doi: 10.1111/imr.12458
21. Kepp O, Bezu L, Yamazaki T, Di Virgilio F, Smyth MJ, Kroemer G, et al. ATP and Cancer Immunosurveillance. EMBO J (2021) 40:e108130. doi: 10.15252/embj.2021108130
22. Seo JH, Dalal MS, Contreras JE. Pannexin-1 Channels as Mediators of Neuroinflammation. Int J Mol Sci (2021) 22. doi: 10.3390/ijms22105189
23. Giaume C, Naus CC, Saez JC, Leybaert L. Glial Connexins and Pannexins in the Healthy and Diseased Brain. Physiol Rev (2021) 101:93–145. doi: 10.1152/physrev.00043.2018
24. Vaidziulyte K, Coppey M, Schauer K. Intracellular Organization in Cell Polarity - Placing Organelles Into the Polarity Loop. J Cell Sci (2019) 132. doi: 10.1242/jcs.230995
25. Bhalla-Gehi R, Penuela S, Churko JM, Shao Q, Laird DW. Pannexin1 and Pannexin3 Delivery, Cell Surface Dynamics, and Cytoskeletal Interactions. J Biol Chem (2010) 285:9147–60. doi: 10.1074/jbc.M109.082008
26. Wicki-Stordeur LE, Swayne LA. Panx1 Regulates Neural Stem and Progenitor Cell Behaviours Associated With Cytoskeletal Dynamics and Interacts With Multiple Cytoskeletal Elements. Cell Commun Signal (2013) 11:62. doi: 10.1186/1478-811X-11-62
27. Boyce AK, Wicki-Stordeur LE, Swayne LA. Powerful Partnership: Crosstalk Between Pannexin 1 and the Cytoskeleton. Front Physiol (2014) 5:27. doi: 10.3389/fphys.2014.00027
28. Junger WG. Immune Cell Regulation by Autocrine Purinergic Signalling. Nat Rev Immunol (2011) 11:201–12. doi: 10.1038/nri2938
29. Velasquez S, Malik S, Lutz SE, Scemes E, Eugenin EA. Pannexin1 Channels Are Required for Chemokine-Mediated Migration of CD4+ T Lymphocytes: Role in Inflammation and Experimental Autoimmune Encephalomyelitis. J Immunol (2016) 196:4338–47. doi: 10.4049/jimmunol.1502440
30. Sáez PJ, Sáez JC, Lennon-Duménil AM, Vargas P. Role of Calcium Permeable Channels in Dendritic Cell Migration. Curr Opin Immunol (2018) 52:74–80. doi: 10.1016/j.coi.2018.04.005
31. Xu X, Wicki-Stordeur LE, Sanchez-Arias JC, Liu M, Weaver MS, Choi CSW, et al. Probenecid Disrupts a Novel Pannexin 1-Collapsin Response Mediator Protein 2 Interaction and Increases Microtubule Stability. Front Cell Neurosci (2018) 12:124. doi: 10.3389/fncel.2018.00124
32. Sáez PJ, Vargas P, Shoji KF, Harcha PA, Lennon-Duménil AM, Sáez JC. ATP Promotes the Fast Migration of Dendritic Cells Through the Activity of Pannexin 1 Channels and P2X7 Receptors. Sci Signal (2017) 10. doi: 10.1126/scisignal.aah7107
33. Cymer M, Brzezniakiewicz-Janus K, Bujko K, Thapa A, Ratajczak J, Anusz K, et al. Pannexin-1 Channel "Fuels" by Releasing ATP From Bone Marrow Cells a State of Sterile Inflammation Required for Optimal Mobilization and Homing of Hematopoietic Stem Cells. Purinergic Signal (2020) 16:313–25. doi: 10.1007/s11302-020-09706-1
34. Ruez R, Dubrot J, Zoso A, Bacchetta M, Molica F, Hugues S, et al. Dendritic Cell Migration Toward CCL21 Gradient Requires Functional Cx43. Front Physiol (2018) 9:288. doi: 10.3389/fphys.2018.00288
35. Corvalán LA, Araya R, Branes MC, Sáez PJ, Kalergis AM, Tobar JA, et al. Injury of Skeletal Muscle and Specific Cytokines Induce the Expression of Gap Junction Channels in Mouse Dendritic Cells. J Cell Physiol (2007) 211:649–60. doi: 10.1002/jcp.20971
36. Kronlage M, Song J, Sorokin L, Isfort K, Schwerdtle T, Leipziger J, et al. Autocrine Purinergic Receptor Signaling is Essential for Macrophage Chemotaxis. Sci Signal (2010) 3:ra55. doi: 10.1126/scisignal.2000588
37. Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, et al. ATP Mediates Rapid Microglial Response to Local Brain Injury In Vivo. Nat Neurosci (2005) 8:752–8. doi: 10.1038/nn1472
38. Fontainhas AM, Wang M, Liang KJ, Chen S, Mettu P, Damani M, et al. Microglial Morphology and Dynamic Behavior is Regulated by Ionotropic Glutamatergic and GABAergic Neurotransmission. PloS One (2011) 6:e15973. doi: 10.1371/journal.pone.0015973
39. Garg C, Seo JH, Ramachandran J, Loh JM, Calderon F, Contreras JE. Trovafloxacin Attenuates Neuroinflammation and Improves Outcome After Traumatic Brain Injury in Mice. J Neuroinflamm (2018) 15:42. doi: 10.1186/s12974-018-1069-9
40. Eugenin EA, Branes MC, Berman JW, Saez JC. TNF-Alpha Plus IFN-Gamma Induce Connexin43 Expression and Formation of Gap Junctions Between Human Monocytes/Macrophages That Enhance Physiological Responses. J Immunol (2003) 170:1320–8. doi: 10.4049/jimmunol.170.3.1320
41. Campwala H, Sexton DW, Crossman DC, Fountain SJ. P2Y(6) Receptor Inhibition Perturbs CCL2-Evoked Signalling in Human Monocytic and Peripheral Blood Mononuclear Cells. J Cell Sci (2014) 127:4964–73. doi: 10.1242/jcs.159012
42. Chen Y, Yao Y, Sumi Y, Li A, To UK, Elkhal A, et al. Purinergic Signaling: A Fundamental Mechanism in Neutrophil Activation. Sci Signal (2010) 3:ra45. doi: 10.1126/scisignal.2000549
43. Bao Y, Chen Y, Ledderose C, Li L, Junger WG. Pannexin 1 Channels Link Chemoattractant Receptor Signaling to Local Excitation and Global Inhibition Responses at the Front and Back of Polarized Neutrophils. J Biol Chem (2013) 288:22650–7. doi: 10.1074/jbc.M113.476283
44. Sarieddine MZ, Scheckenbach KE, Foglia B, Maass K, Garcia I, Kwak BR, et al. Connexin43 Modulates Neutrophil Recruitment to the Lung. J Cell Mol Med (2009) 13:4560–70. doi: 10.1111/j.1582-4934.2008.00654.x
45. Wang X, Qin W, Xu X, Xiong Y, Zhang Y, Zhang H, et al. Endotoxin-Induced Autocrine ATP Signaling Inhibits Neutrophil Chemotaxis Through Enhancing Myosin Light Chain Phosphorylation. Proc Natl Acad Sci USA (2017) 114:4483–8. doi: 10.1073/pnas.1616752114
46. Poplimont H, Georgantzoglou A, Boulch M, Walker HA, Coombs C, Papaleonidopoulou F, et al. Neutrophil Swarming in Damaged Tissue Is Orchestrated by Connexins and Cooperative Calcium Alarm Signals. Curr Biol (2020) 30:2761–76.e2767. doi: 10.1016/j.cub.2020.05.030
47. Medina CB, Chiu YH, Stremska ME, Lucas CD, Poon I, Tung KS, et al. Pannexin 1 Channels Facilitate Communication Between T Cells to Restrict the Severity of Airway Inflammation. Immunity (2021) 54:1715–27.e1717. doi: 10.1016/j.immuni.2021.06.014
48. Ledderose C, Liu K, Kondo Y, Slubowski CJ, Dertnig T, Denicolo S, et al. Purinergic P2X4 Receptors and Mitochondrial ATP Production Regulate T Cell Migration. J Clin Invest (2018) 128:3583–94. doi: 10.1172/JCI120972
49. Gurusamy M, Tischner D, Shao J, Klatt S, Zukunft S, Bonnavion R, et al. G-Protein-Coupled Receptor P2Y10 Facilitates Chemokine-Induced CD4 T Cell Migration Through Autocrine/Paracrine Mediators. Nat Commun (2021). doi: 10.1038/s41467-021-26882-9
50. Hainz N, Wolf S, Beck A, Wagenpfeil S, Tschernig T, Meier C. Probenecid Arrests the Progression of Pronounced Clinical Symptoms in a Mouse Model of Multiple Sclerosis. Sci Rep (2017) 7:17214. doi: 10.1038/s41598-017-17517-5
51. Harcha PA, Vargas A, Yi C, Koulakoff AA, Giaume C, Saez JC. Hemichannels Are Required for Amyloid Beta-Peptide-Induced Degranulation and Are Activated in Brain Mast Cells of APPswe/PS1dE9 Mice. J Neurosci (2015) 35:9526–38. doi: 10.1523/JNEUROSCI.3686-14.2015
52. Berzat A, Hall A. Cellular Responses to Extracellular Guidance Cues. EMBO J (2010) 29:2734–45. doi: 10.1038/emboj.2010.170
53. Thiam HR, Wong SL, Qiu R, Kittisopikul M, Vahabikashi A, Goldman AE, et al. NETosis Proceeds by Cytoskeleton and Endomembrane Disassembly and PAD4-Mediated Chromatin Decondensation and Nuclear Envelope Rupture. Proc Natl Acad Sci USA (2020) 117:7326–37. doi: 10.1073/pnas.1909546117
54. Chen Y, Corriden R, Inoue Y, Yip L, Hashiguchi N, Zinkernagel A, et al. ATP Release Guides Neutrophil Chemotaxis via P2Y2 and A3 Receptors. Science (2006) 314:1792–5. doi: 10.1126/science.1132559
55. Alarcon P, Manosalva C, Quiroga J, Belmar I, Alvarez K, Diaz G, et al. Oleic and Linoleic Acids Induce the Release of Neutrophil Extracellular Traps via Pannexin 1-Dependent ATP Release and P2X1 Receptor Activation. Front Vet Sci (2020) 7:260. doi: 10.3389/fvets.2020.00260
56. Bao Y, Ledderose C, Graf AF, Brix B, Birsak T, Lee A, et al. mTOR and Differential Activation of Mitochondria Orchestrate Neutrophil Chemotaxis. J Cell Biol (2015) 210:1153–64. doi: 10.1083/jcb.201503066
57. Sil P, Hayes CP, Reaves BJ, Breen P, Quinn S, Sokolove J, et al. P2Y6 Receptor Antagonist MRS2578 Inhibits Neutrophil Activation and Aggregated Neutrophil Extracellular Trap Formation Induced by Gout-Associated Monosodium Urate Crystals. J Immunol (2017) 198:428–42. doi: 10.4049/jimmunol.1600766
58. Alkayed F, Kashimata M, Koyama N, Hayashi T, Tamura Y, Azuma Y. P2Y11 Purinoceptor Mediates the ATP-Enhanced Chemotactic Response of Rat Neutrophils. J Pharmacol Sci (2012) 120:288–95. doi: 10.1254/jphs.12173fp
59. Barrett MO, Sesma JI, Ball CB, Jayasekara PS, Jacobson KA, Lazarowski ER, et al. A Selective High-Affinity Antagonist of the P2Y14 Receptor Inhibits UDP-Glucose-Stimulated Chemotaxis of Human Neutrophils. Mol Pharmacol (2013) 84:41–9. doi: 10.1124/mol.113.085654
60. Xu X, Zheng S, Xiong Y, Wang X, Qin W, Zhang H, et al. Adenosine Effectively Restores Endotoxin-Induced Inhibition of Human Neutrophil Chemotaxis via A1 Receptor-P38 Pathway. Inflamm Res (2017) 66:353–64. doi: 10.1007/s00011-016-1021-3
61. Liu L, Das S, Losert W, Parent CA. Mtorc2 Regulates Neutrophil Chemotaxis in a cAMP- and RhoA-Dependent Fashion. Dev Cell (2010) 19:845–57. doi: 10.1016/j.devcel.2010.11.004
62. Rivas-Yanez E, Barrera-Avalos C, Parra-Tello B, Briceno P, Rosemblatt MV, Saavedra-Almarza J, et al. P2X7 Receptor at the Crossroads of T Cell Fate. Int J Mol Sci (2020) 21. doi: 10.3390/ijms21144937
63. Schenk U, Westendorf AM, Radaelli E, Casati A, Ferro M, Fumagalli M, et al. Purinergic Control of T Cell Activation by ATP Released Through Pannexin-1 Hemichannels. Sci Signal (2008) 1:ra6. doi: 10.1126/scisignal.1160583
64. Woehrle T, Yip L, Elkhal A, Sumi Y, Chen Y, Yao Y, et al. Pannexin-1 Hemichannel-Mediated ATP Release Together With P2X1 and P2X4 Receptors Regulate T-Cell Activation at the Immune Synapse. Blood (2010) 116:3475–84. doi: 10.1182/blood-2010-04-277707
65. Shoji KF, Saez PJ, Harcha PA, Aguila HL, Saez JC. Pannexin1 Channels Act Downstream of P2X 7 Receptors in ATP-Induced Murine T-Cell Death. Channels (Austin) (2014) 8:142–56. doi: 10.4161/chan.28122
66. Ledderose C, Bromberger S, Slubowski CJ, Sueyoshi K, Aytan D, Shen Y, et al. The Purinergic Receptor P2Y11 Choreographs the Polarization, Mitochondrial Metabolism, and Migration of T Lymphocytes. Sci Signal (2020) 13. doi: 10.1126/scisignal.aba3300
67. Campello S, Lacalle RA, Bettella M, Manes S, Scorrano L, Viola A. Orchestration of Lymphocyte Chemotaxis by Mitochondrial Dynamics. J Exp Med (2006) 203:2879–86. doi: 10.1084/jem.20061877
68. Vincent P, Collette Y, Marignier R, Vuaillat C, Rogemond V, Davoust N, et al. A Role for the Neuronal Protein Collapsin Response Mediator Protein 2 in T Lymphocyte Polarization and Migration. J Immunol (2005) 175:7650–60. doi: 10.4049/jimmunol.175.11.7650
69. De Winde CM, Munday C, Acton ,SE. Molecular Mechanisms of Dendritic Cell Migration in Immunity and Cancer. Med Microbiol Immunol (2020) 209:515–29. doi: 10.1007/s00430-020-00680-4
70. Banchereau J, Steinman RM. Dendritic Cells and the Control of Immunity. Nature (1998) 392:245–52. doi: 10.1038/32588
71. Cekic C, Linden J. Purinergic Regulation of the Immune System. Nat Rev Immunol (2016) 16:177–92. doi: 10.1038/nri.2016.4
72. López X, Palacios-Prado N, Guiza J, Escamilla R, Fernandez P, Vega JL, et al. A Physiologic Rise in Cytoplasmic Calcium Ion Signal Increases Pannexin1 Channel Activity via a C-Terminus Phosphorylation by CaMKII. Proc Natl Acad Sci USA (2021). doi: 10.1073/pnas.2108967118
73. Khan S, Downing KH, Molloy JE. Architectural Dynamics of CaMKII-Actin Networks. Biophys J (2019) 116:104–19. doi: 10.1016/j.bpj.2018.11.006
74. Bretou M, Sáez PJ, Sanseau D, Maurin M, Lankar D, Chabaud M, et al. Lysosome Signaling Controls the Migration of Dendritic Cells. Sci Immunol (2017) 2. doi: 10.1126/sciimmunol.aak9573
75. Boyce AKJ, van der Slagt E, Sanchez-Arias JC, Swayen LA. ATP Triggers Macropinocytosis That Internalizes and is Regulated by PANX1. bioRxiv (2020) doi. doi: 10.1101/2020.11.19.389072
76. Cina C, Maass K, Theis M, Willecke K, Bechberger JF, Naus CC. Involvement of the Cytoplasmic C-Terminal Domain of Connexin43 in Neuronal Migration. J Neurosci (2009) 29:2009–21. doi: 10.1523/JNEUROSCI.5025-08.2009
77. Jakubzick CV, Randolph GJ, Henson PM. Monocyte Differentiation and Antigen-Presenting Functions. Nat Rev Immunol (2017) 17:349–62. doi: 10.1038/nri.2017.28
78. Rodjakovic D, Salm L, Beldi G. Function of Connexin-43 in Macrophages. Int J Mol Sci (2021) 22. doi: 10.3390/ijms22031412
79. Geissmann F, Gordon S, Hume DA, Mowat AM, Randolph GJ. Unravelling Mononuclear Phagocyte Heterogeneity. Nat Rev Immunol (2010) 10:453–60. doi: 10.1038/nri2784
80. Yu C, Roubeix C, Sennlaub F, Saban DR. Microglia Versus Monocytes: Distinct Roles in Degenerative Diseases of the Retina. Trends Neurosci (2020) 43:433–49. doi: 10.1016/j.tins.2020.03.012
81. Poon IK, Chiu YH, Armstrong AJ, Kinchen JM, Juncadella IJ, Bayliss DA, et al. Unexpected Link Between an Antibiotic, Pannexin Channels and Apoptosis. Nature (2014) 507:329–34. doi: 10.1038/nature13147
82. Seo JH, Dalal MS, Calderon F, Contreras JE. Myeloid Pannexin-1 Mediates Acute Leukocyte Infiltration and Leads to Worse Outcomes After Brain Trauma. J Neuroinflamm (2020) 17:245. doi: 10.1186/s12974-020-01917-y
83. Shen C, Chen JH, Lee Y, Hassan MM, Kim SJ, Choi EY, et al. mTOR- and SGK-Mediated Connexin 43 Expression Participates in Lipopolysaccharide-Stimulated Macrophage Migration Through the iNOS/Src/FAK Axis. J Immunol (2018) 201:2986–97. doi: 10.4049/jimmunol.1700954
84. Cui K, Ardell CL, Podolnikova NP, Yakubenko VP. Distinct Migratory Properties of M1, M2, and Resident Macrophages Are Regulated by Alphadbeta2 and Alphambeta2 Integrin-Mediated Adhesion. Front Immunol (2018) 9:2650. doi: 10.3389/fimmu.2018.02650
85. Lopez-Castejon G, Baroja-Mazo A, Pelegrin P. Novel Macrophage Polarization Model: From Gene Expression to Identification of New Anti-Inflammatory Molecules. Cell Mol Life Sci (2011) 68:3095–107. doi: 10.1007/s00018-010-0609-y
86. Chekeni FB, Elliott MR, Sandilos JK, Walk SF, Kinchen JM, Lazarowski ER, et al. Pannexin 1 Channels Mediate ’Find-Me’ Signal Release and Membrane Permeability During Apoptosis. Nature (2010) 467:863–7. doi: 10.1038/nature09413
87. Xiao F, Waldrop SL, Bronk SF, Gores GJ, Davis LS, Kilic G. Lipoapoptosis Induced by Saturated Free Fatty Acids Stimulates Monocyte Migration: A Novel Role for Pannexin1 in Liver Cells. Purinergic Signal (2015) 11:347–59. doi: 10.1007/s11302-015-9456-5
88. Pillon NJ, Li YE, Fink LN, Brozinick JT, Nikolayev A, Kuo MS, et al. Nucleotides Released From Palmitate-Challenged Muscle Cells Through Pannexin-3 Attract Monocytes. Diabetes (2014) 63:3815–26. doi: 10.2337/db14-0150
89. Zhang P, Ishikawa M, Rhodes C, Doyle A, Ikeuchi T, Nakamura K, et al. Pannexin-3 Deficiency Delays Skin Wound Healing in Mice Due to Defects in Channel Functionality. J Invest Dermatol (2019) 139:909–18. doi: 10.1016/j.jid.2018.08.033
90. Ling EA, Kaur C, Lu J. Origin, Nature, and Some Functional Considerations of Intraventricular Macrophages, With Special Reference to the Epiplexus Cells. Microsc Res Tech (1998) 41:43–56. doi: 10.1002/(SICI)1097-0029(19980401)41:1<43::AID-JEMT5>3.0.CO;2-V
91. Maslieieva V, Thompson RJ. A Critical Role for Pannexin-1 in Activation of Innate Immune Cells of the Choroid Plexus. Channels (Austin) (2014) 8:131–41. doi: 10.4161/chan.27653
92. Tozzi M, Hansen JB, Novak I. Pannexin-1 Mediated ATP Release in Adipocytes is Sensitive to Glucose and Insulin and Modulates Lipolysis and Macrophage Migration. Acta Physiol (Oxf) (2020) 228:e13360. doi: 10.1111/apha.13360
93. Adamson SE, Meher AK, Chiu YH, Sandilos JK, Oberholtzer NP, Walker NN, et al. Pannexin 1 is Required for Full Activation of Insulin-Stimulated Glucose Uptake in Adipocytes. Mol Metab (2015) 4:610–8. doi: 10.1016/j.molmet.2015.06.009
94. Huang C, Jacobson K, Schaller MD. MAP Kinases and Cell Migration. J Cell Sci (2004) 117:4619–28. doi: 10.1242/jcs.01481
95. Hino N, Rossetti L, Marin-Llaurado A, Aoki K, Trepat X, Matsuda M, et al. ERK-Mediated Mechanochemical Waves Direct Collective Cell Polarization. Dev Cell (2020) 53:646–60.e648. doi: 10.1016/j.devcel.2020.05.011
96. López X, Escamilla R, Fernandez P, Duarte Y, Gonzalez-Nilo F, Palacios-Prado N, et al. Stretch-Induced Activation of Pannexin 1 Channels Can Be Prevented by PKA-Dependent Phosphorylation. Int J Mol Sci (2020) 21. doi: 10.3390/ijms21239180
97. Lagos-Cabre R, Alvarez A, Kong M, Burgos-Bravo F, Cardenas A, Rojas-Mancilla E, et al. Alphavbeta3 Integrin Regulates Astrocyte Reactivity. J Neuroinflamm (2017) 14:194. doi: 10.1186/s12974-017-0968-5
98. Peñuela S, Kelly JJ, Churko JM, Barr KJ, Berger AC, Laird DW. Panx1 Regulates Cellular Properties of Keratinocytes and Dermal Fibroblasts in Skin Development and Wound Healing. J Invest Dermatol (2014) 134:2026–35. doi: 10.1038/jid.2014.86
99. Liu H, Yuan M, Yao Y, Wu D, Dong S, Tong X. In Vitro Effect of Pannexin 1 Channel on the Invasion and Migration of I-10 Testicular Cancer Cells via ERK1/2 Signaling Pathway. BioMed Pharmacother (2019) 117:109090. doi: 10.1016/j.biopha.2019.109090
100. Furlow PW, Zhang S, Soong TD, Halberg N, Goodarzi H, Mangrum C, et al. Mechanosensitive Pannexin-1 Channels Mediate Microvascular Metastatic Cell Survival. Nat Cell Biol (2015) 17:943–52. doi: 10.1038/ncb3194
101. Nakajima KI, Tatsumi M, Zhao M. An Essential and Synergistic Role of Purinergic Signaling in Guided Migration of Corneal Epithelial Cells in Physiological Electric Fields. Cell Physiol Biochem (2019) 52:198–211. doi: 10.33594/000000014
102. Xiang X, Langlois S, St-Pierre ME, Barre JF, Grynspan D, Purgina B, et al. Pannexin 1 Inhibits Rhabdomyosarcoma Progression Through a Mechanism Independent of its Canonical Channel Function. Oncogenesis (2018) 7:89. doi: 10.1038/s41389-018-0100-4
103. Xiang X, Langlois S, St-Pierre ME, Blinder A, Charron P, Graber TE, et al. Identification of Pannexin 1-Regulated Genes, Interactome, and Pathways in Rhabdomyosarcoma and its Tumor Inhibitory Interaction With AHNAK. Oncogene (2021) 40:1868–83. doi: 10.1038/s41388-020-01623-2
104. Lai CP, Bechberger JF, Thompson RJ, Macvicar BA, Bruzzone R, Naus CC. Tumor-Suppressive Effects of Pannexin 1 in C6 Glioma Cells. Cancer Res (2007) 67:1545–54. doi: 10.1158/0008-5472.CAN-06-1396
105. Flores-Munoz C, Maripillan J, Vasquez-Navarrete J, Novoa-Molina J, Ceriani R, Sanchez HA, et al. Restraint of Human Skin Fibroblast Motility, Migration, and Cell Surface Actin Dynamics, by Pannexin 1 and P2X7 Receptor Signaling. Int J Mol Sci (2021) 22. doi: 10.3390/ijms22031069
106. Werner S, Krieg T, Smola H. Keratinocyte-Fibroblast Interactions in Wound Healing. J Invest Dermatol (2007) 127:998–1008. doi: 10.1038/sj.jid.5700786
107. Russo B, Brembilla NC, Chizzolini C. Interplay Between Keratinocytes and Fibroblasts: A Systematic Review Providing a New Angle for Understanding Skin Fibrotic Disorders. Front Immunol (2020) 11:648. doi: 10.3389/fimmu.2020.00648
108. Burnstock G, Knight GE, Greig AV. Purinergic Signaling in Healthy and Diseased Skin. J Invest Dermatol (2012) 132:526–46. doi: 10.1038/jid.2011.344
109. Peñuela S, Bhalla R, Gong XQ, Cowan KN, Celetti SJ, Cowan BJ, et al. Pannexin 1 and Pannexin 3 are Glycoproteins That Exhibit Many Distinct Characteristics From the Connexin Family of Gap Junction Proteins. J Cell Sci (2007) 120:3772–83. doi: 10.1242/jcs.009514
110. Celetti SJ, Cowan KN, Penuela S, Shao Q, Churko J, Laird DW. Implications of Pannexin 1 and Pannexin 3 for Keratinocyte Differentiation. J Cell Sci (2010) 123:1363–72. doi: 10.1242/jcs.056093
111. Pinheiro AR, Paramos-De-Carvalho D, Certal M, Costa MA, Costa C, Magalhaes-Cardoso MT, et al. Histamine Induces ATP Release From Human Subcutaneous Fibroblasts, via Pannexin-1 Hemichannels, Leading to Ca2+ Mobilization and Cell Proliferation. J Biol Chem (2013) 288:27571–83. doi: 10.1074/jbc.M113.460865
112. Lagos-Cabre R, Burgos-Bravo F, Avalos AM, Leyton ,L. Connexins in Astrocyte Migration. Front Pharmacol (2019) 10:1546. doi: 10.3389/fphar.2019.01546
113. Dreyer EB, Leifer D, Heng JE, Mcconnell JE, Gorla M, Levin LA, et al. An Astrocytic Binding Site for Neuronal Thy-1 and its Effect on Neurite Outgrowth. Proc Natl Acad Sci USA (1995) 92:11195–9. doi: 10.1073/pnas.92.24.11195
114. Leyton L, Schneider P, Labra CV, Ruegg C, Hetz CA, Quest AF, et al. Thy-1 Binds to Integrin Beta(3) on Astrocytes and Triggers Formation of Focal Contact Sites. Curr Biol (2001) 11:1028–38. doi: 10.1016/s0960-9822(01)00262-7
115. Avalos AM, Arthur WT, Schneider P, Quest AF, Burridge K, Leyton L. Aggregation of Integrins and RhoA Activation are Required for Thy-1-Induced Morphological Changes in Astrocytes. J Biol Chem (2004) 279:39139–45. doi: 10.1074/jbc.M403439200
116. Kong M, Munoz N, Valdivia A, Alvarez A, Herrera-Molina R, Cardenas A, et al. Thy-1-Mediated Cell-Cell Contact Induces Astrocyte Migration Through the Engagement of Alphavbeta3 Integrin and Syndecan-4. Biochim Biophys Acta (2013) 1833:1409–20. doi: 10.1016/j.bbamcr.2013.02.013
117. Henriquez M, Herrera-Molina R, Valdivia A, Alvarez A, Kong M, Munoz N, et al. ATP Release Due to Thy-1-Integrin Binding Induces P2X7-Mediated Calcium Entry Required for Focal Adhesion Formation. J Cell Sci (2011) 124:1581–8. doi: 10.1242/jcs.073171
118. Alvarez A, Lagos-Cabre R, Kong M, Cardenas A, Burgos-Bravo F, Schneider P, et al. Integrin-Mediated Transactivation of P2X7R via Hemichannel-Dependent ATP Release Stimulates Astrocyte Migration. Biochim Biophys Acta (2016) 1863:2175–88. doi: 10.1016/j.bbamcr.2016.05.018
119. Clark AG, Vignjevic DM. Modes of Cancer Cell Invasion and the Role of the Microenvironment. Curr Opin Cell Biol (2015) 36:13–22. doi: 10.1016/j.ceb.2015.06.004
120. Vilchez Mercedes SA, Bocci F, Levine H, Onuchic JN, Jolly MK, Wong PK. Decoding Leader Cells in Collective Cancer Invasion. Nat Rev Cancer (2021) doi: 10.1038/s41568-021-00376-8
121. Proulx AA, Lin ZX, Naus CC. Transfection of Rhabdomyosarcoma Cells With Connexin43 Induces Myogenic Differentiation. Cell Growth Differ (1997) 8:533–40.
122. Peñuela S, Gyenis L, Ablack A, Churko JM, Berger AC, Litchfield DW, et al. Loss of Pannexin 1 Attenuates Melanoma Progression by Reversion to a Melanocytic Phenotype. J Biol Chem (2012) 287:29184–93. doi: 10.1074/jbc.M112.377176
123. Brenet M, Martinez S, Perez-Nunez R, Perez LA, Contreras P, Diaz J, et al. Thy-1 (CD90)-Induced Metastatic Cancer Cell Migration and Invasion Are Beta3 Integrin-Dependent and Involve a Ca(2+)/P2X7 Receptor Signaling Axis. Front Cell Dev Biol (2020) 8:592442. doi: 10.3389/fcell.2020.592442
124. Abascal F, Zardoya R. LRRC8 Proteins Share a Common Ancestor With Pannexins, and may Form Hexameric Channels Involved in Cell-Cell Communication. Bioessays (2012) 34:551–60. doi: 10.1002/bies.201100173
125. Friard J, Corinus A, Cougnon M, Tauc M, Pisani DF, Duranton C, et al. LRRC8/VRAC Channels Exhibit a Noncanonical Permeability to Glutathione, Which Modulates Epithelial-to-Mesenchymal Transition (EMT). Cell Death Dis (2019) 10:925. doi: 10.1038/s41419-019-2167-z
126. Weyand CM, Goronzy JJ. Aging of the Immune System. Mechanisms and Therapeutic Targets. Ann Am Thorac Soc (2016) 13(Suppl 5):S422–8. doi: 10.1513/AnnalsATS.201602-095AW
127. Raab M, Gentili M, De Belly H, Thiam HR, Vargas P, Jimenez AJ, et al. ESCRT III Repairs Nuclear Envelope Ruptures During Cell Migration to Limit DNA Damage and Cell Death. Science (2016) 352:359–62. doi: 10.1126/science.aad7611
128. Agrawal A, Agrawal S, Cao JN, Su H, Osann K, Gupta S. Altered Innate Immune Functioning of Dendritic Cells in Elderly Humans: A Role of Phosphoinositide 3-Kinase-Signaling Pathway. J Immunol (2007) 178:6912–22. doi: 10.4049/jimmunol.178.11.6912
129. Nikolich-Zugich J. Ageing and Life-Long Maintenance of T-Cell Subsets in the Face of Latent Persistent Infections. Nat Rev Immunol (2008) 8:512–22. doi: 10.1038/nri2318
130. Chen G, Lustig A, Weng NP. T Cell Aging: A Review of the Transcriptional Changes Determined From Genome-Wide Analysis. Front Immunol (2013) 4:121. doi: 10.3389/fimmu.2013.00121
131. Turner VM, Mabbott NA. Ageing Adversely Affects the Migration and Function of Marginal Zone B Cells. Immunology (2017) 151:349–62. doi: 10.1111/imm.12737
132. Saare M, Tserel L, Haljasmagi L, Taalberg E, Peet N, Eimre M, et al. Monocytes Present Age-Related Changes in Phospholipid Concentration and Decreased Energy Metabolism. Aging Cell (2020) 19:e13127. doi: 10.1111/acel.13127
133. Zhang Y, Zhu W, He H, Fan B, Deng R, Hong Y, et al. Macrophage Migration Inhibitory Factor Rejuvenates Aged Human Mesenchymal Stem Cells and Improves Myocardial Repair. Aging (Albany NY) (2019) 11:12641–60. doi: 10.18632/aging.102592
134. Damani MR, Zhao L, Fontainhas AM, Amaral J, Fariss RN, Wong WT. Age-Related Alterations in the Dynamic Behavior of Microglia. Aging Cell (2011) 10:263–76. doi: 10.1111/j.1474-9726.2010.00660.x
135. Uhl B, Vadlau Y, Zuchtriegel G, Nekolla K, Sharaf K, Gaertner F, et al. Aged Neutrophils Contribute to the First Line of Defense in the Acute Inflammatory Response. Blood (2016) 128:2327–37. doi: 10.1182/blood-2016-05-718999
136. Barkaway A, Rolas L, Joulia R, Bodkin J, Lenn T, Owen-Woods C, et al. Age-Related Changes in the Local Milieu of Inflamed Tissues Cause Aberrant Neutrophil Trafficking and Subsequent Remote Organ Damage. Immunity (2021) 54:1494–510.e1497. doi: 10.1016/j.immuni.2021.04.025
137. Wenisch C, Patruta S, Daxbock F, Krause R, Horl W. Effect of Age on Human Neutrophil Function. J Leukoc Biol (2000) 67:40–5. doi: 10.1002/jlb.67.1.40
138. Butcher SK, Chahal H, Nayak L, Sinclair A, Henriquez NV, Sapey E, et al. Senescence in Innate Immune Responses: Reduced Neutrophil Phagocytic Capacity and CD16 Expression in Elderly Humans. J Leukoc Biol (2001) 70:881–6. doi: 10.1189/jlb.70.6.881
139. Brown AD, Close GL, Sharples AP, Stewart CE. Murine Myoblast Migration: Influence of Replicative Ageing and Nutrition. Biogerontology (2017) 18:947–64. doi: 10.1007/s10522-017-9735-3
140. Agliari E, Saez PJ, Barra A, Piel M, Vargas P, Castellana M. A Statistical Inference Approach to Reconstruct Intercellular Interactions in Cell Migration Experiments. Sci Adv (2020) 6:eaay2103. doi: 10.1126/sciadv.aay2103
141. Buttenschon A, Edelstein-Keshet L. Bridging From Single to Collective Cell Migration: A Review of Models and Links to Experiments. PloS Comput Biol (2020) 16:e1008411. doi: 10.1371/journal.pcbi.1008411
142. Tweedy L, Thomason PA, Paschke PI, Martin K, Machesky LM, Zagnoni M, et al. Seeing Around Corners: Cells Solve Mazes and Respond at a Distance Using Attractant Breakdown. Science (2020) 369. doi: 10.1126/science.aay9792
143. Jin S, Guerrero-Juarez CF, Zhang L, Chang I, Ramos R, Kuan CH, et al. Inference and Analysis of Cell-Cell Communication Using CellChat. Nat Commun (2021) 12:1088. doi: 10.1038/s41467-021-21246-9
144. Sanchez-Arias JC, Candlish RC, van der Slagt E, Swayne LA. Pannexin 1 Regulates Dendritic Protrusion Dynamics in Immature Cortical Neurons. eNeuro (2020) 7. doi: 10.1523/ENEURO.0079-20.2020
145. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 and Piezo2 are Essential Components of Distinct Mechanically Activated Cation Channels. Science (2010) 330:55–60. doi: 10.1126/science.1193270
146. Zmurchok C, Collette J, Rajagopal V, Holmes WR. Membrane Tension Can Enhance Adaptation to Maintain Polarity of Migrating Cells. Biophys J (2020) 119:1617–29. doi: 10.1016/j.bpj.2020.08.035
147. Adamson SE, Leitinger N. The Role of Pannexin1 in the Induction and Resolution of Inflammation. FEBS Lett (2014) 588:1416–22. doi: 10.1016/j.febslet.2014.03.009
148. Giuliani AL, Sarti AC, Di Virgilio F. Extracellular Nucleotides and Nucleosides as Signalling Molecules. Immunol Lett (2019) 205:16–24. doi: 10.1016/j.imlet.2018.11.006
149. Montell C. The TRP Superfamily of Cation Channels. Sci STKE (2005) 2005:re3. doi: 10.1126/stke.2722005re3
150. Harcha PA, Lopez X, Saez PJ, Fernandez P, Barria I, Martinez AD, et al. Pannexin-1 Channels Are Essential for Mast Cell Degranulation Triggered During Type I Hypersensitivity Reactions. Front Immunol (2019) 10:2703. doi: 10.3389/fimmu.2019.02703
151. Lai CP, Bechberger JF, Naus ,CC. Pannexin2 as a Novel Growth Regulator in C6 Glioma Cells. Oncogene (2009) 28:4402–8. doi: 10.1038/onc.2009.283
152. O’donnell BL, Penuela S. Pannexin 3 Channels in Health and Disease. Purinergic Signal (2021). doi: 10.1007/s11302-021-09805-7
Keywords: cell communication, leukocytes, cancer, inflammation, Ca2+ signaling, amoeboid migration, mesenchymal migration, mechanotransduction
Citation: Harcha PA, López-López T, Palacios AG and Sáez PJ (2021) Pannexin Channel Regulation of Cell Migration: Focus on Immune Cells. Front. Immunol. 12:750480. doi: 10.3389/fimmu.2021.750480
Received: 30 July 2021; Accepted: 15 November 2021;
Published: 16 December 2021.
Edited by:
Emmanuel Donnadieu, Institut National de la Santé et de la Recherche Médicale (INSERM), FranceReviewed by:
Eliseo A. Eugenin, University of Texas Medical Branch at Galveston, United StatesLeigh Anne Swayne, University of Victoria, Canada
Copyright © 2021 Harcha, López-López, Palacios and Sáez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pablo J. Sáez, p.saez@uke.de