 Petros P. Sfikakis1,2*
Petros P. Sfikakis1,2* Kleio-Maria Verrou1,2
Kleio-Maria Verrou1,2 Giannis Ampatziadis-Michailidis1
Giannis Ampatziadis-Michailidis1 Ourania Tsitsilonis3
Ourania Tsitsilonis3 Dimitrios Paraskevis4Efstathios Kastritis5
Dimitrios Paraskevis4Efstathios Kastritis5 Evi Lianidou6Paraskevi Moutsatsou7
Evi Lianidou6Paraskevi Moutsatsou7 Evangelos Terpos5
Evangelos Terpos5 Ioannis Trougakos8Vasiliki Chini1Menelaos Manoloukos1
Ioannis Trougakos8Vasiliki Chini1Menelaos Manoloukos1 Panagiotis Moulos1,9
Panagiotis Moulos1,9 Georgios A. Pavlopoulos1,9George Kollias1,2,10
Georgios A. Pavlopoulos1,9George Kollias1,2,10 Pantelis Hatzis1,9
Pantelis Hatzis1,9 Meletios A. Dimopoulos1,5
Meletios A. Dimopoulos1,5- 1Center of New Biotechnologies & Precision Medicine, National and Kapodistrian University of Athens Medical School, Athens, Greece
- 2Joint Rheumatology Program, National and Kapodistrian University of Athens Medical School, Athens, Greece
- 3Department of Biology, National and Kapodistrian University of Athens (NKUA), Athens, Greece
- 4Department of Hygiene, Epidemiology and Medical Statistics, Medical School, National and Kapodistrian University of Athens, Athens, Greece
- 5Department of Clinical Therapeutics, School of Medicine, National and Kapodistrian University of Athens, Athens, Greece
- 6Department of Chemistry, National and Kapodistrian University of Athens (NKUA), Athens, Greece
- 7Department of Clinical Biochemistry, School of Medicine, University General Hospital Attikon, NKUA, Haidari, Greece
- 8Department of Cell Biology and Biophysics, Faculty of Biology, National and Kapodistrian University of Athens, Athens, Greece
- 9Institute for Fundamental Biomedical Research, Biomedical Sciences Research Center (BSRC) Alexander Fleming, Vari, Greece
- 10Institute for Bioinnovation, Biomedical Sciences Research Center (BSRC) Alexander Fleming, Vari, Greece
The reasons behind the clinical variability of SARS-CoV-2 infection, ranging from asymptomatic infection to lethal disease, are still unclear. We performed genome-wide transcriptional whole-blood RNA sequencing, bioinformatics analysis and PCR validation to test the hypothesis that immune response-related gene signatures reflecting baseline may differ between healthy individuals, with an equally robust antibody response, who experienced an entirely asymptomatic (n=17) versus clinical SARS-CoV-2 infection (n=15) in the past months (mean of 14 weeks). Among 12.789 protein-coding genes analysed, we identified six and nine genes with significantly decreased or increased expression, respectively, in those with prior asymptomatic infection relatively to those with clinical infection. All six genes with decreased expression (IFIT3, IFI44L, RSAD2, FOLR3, PI3, ALOX15), are involved in innate immune response while the first two are interferon-induced proteins. Among genes with increased expression six are involved in immune response (GZMH, CLEC1B, CLEC12A), viral mRNA translation (GCAT), energy metabolism (CACNA2D2) and oxidative stress response (ENC1). Notably, 8/15 differentially expressed genes are regulated by interferons. Our results suggest that subtle differences at baseline expression of innate immunity-related genes may be associated with an asymptomatic disease course in SARS-CoV-2 infection. Whether a certain gene signature predicts, or not, those who will develop a more efficient immune response upon exposure to SARS-CoV-2, with implications for prioritization for vaccination, warrant further study.
Introduction
Since December 2019 the SARS-CoV-2 has spread throughout the world infecting dozens of millions of people and resulting in over 2.8 million deaths, as of April 2021. Although the case fatality rate in hospitalized patients may exceed 10% (1, 2), 35-50% of infected adults do not develop, perceive and report any clinical symptom (3, 4). Asymptomatic infected persons may be responsible for viral transmission for more days than aware self-isolated cases, which may also explain, at least partially, the exponential increase in the number of infections globally (5–7). Notably, we only know in retrospect who was indeed asymptomatic, since individuals without symptoms at the time of a positive molecular test should be followed for 14 days to determine the clinical picture, being “pre-symptomatic” if they develop symptoms later.
The proportion of asymptomatic individuals varies widely among different viral infections, whereas relevant biomarkers do not currently exist due to our limited knowledge of the molecular host-pathogen interactions and immune response to particular infections (8). For example, a significant fraction of cytomegalovirus infections, similarly to SARS-CoV-2, are asymptomatic and unsuspected (9). In contrast, measles infected individuals are very rarely asymptomatic (10). The reasons why certain individuals, including even people living with HIV (11) or other immunodeficiencies (12), do not develop clinical symptoms during SARS-CoV-2 infection are essentially unknown (13, 14). So far, studies assessing the immune response during asymptomatic infection are few. In an elegant study, Long et al. showed that asymptomatic individuals presented with significantly longer duration of viral shedding compared to symptomatic patients, lower levels of IgG antibodies to SARS-CoV-2, and lower serum levels of 18/48 cytokines, including interferon-gamma levels, suggesting that asymptomatic individuals indeed displayed a weaker anti-virus-reactive immune response to SARS-CoV-2 (15). More recently, Chan et al. also showed in a whole blood transcriptomic analyses that asymptomatic patients display a less robust response to type-I interferon than symptomatic patients, whereas differences between asymptomatic and symptomatic patients may be present at the cellular, innate, and adaptive immune response levels (16)
While the role of genetics in determining immune and clinical response to the SARS-CoV-2 virus is currently under investigation, it is well established that individual human immune systems are highly variable (17). Most of this inter-individual immune variation is explained by environmental exposures early in life (18) but genetic factors are clearly also involved. For example, a gene expression signature dominated by interferon-inducible genes in the blood is prominent in systemic lupus erythematosus (19), whereas interferon-α is increased not only in the serum of these patients but also in their healthy first-degree relatives (20) pointing to genetic influences on the interferon-mediated immune interactions.
Clearly, the most successful immune response against SARS-CoV-2 occurs in those individuals who, while remaining asymptomatic, develop a robust adaptive response. This is not always the case since antibodies are not detected in a proportion of asymptomatic infections (21). We have recently examined the humoral immune response to SARS-CoV-2 in members of the National and Kapodistrian University of Athens, Greece (22). Overall, among 4.996 people the unweighted seroprevalence of SARS-CoV-2 antibodies was 1.58%, whereas 49% of the seropositive individuals denied having had any clinical symptom compatible with previous SARS-CoV-2 infection, which was also unsuspected for 33% of them. Interestingly, in our study, the mean levels of antibodies to both the nucleocapsid (N) protein and the receptor-binding-domain (RBD) of the spike (S) protein were comparable between asymptomatic and clinical infection cases and not associated with age or sex (4). Others have also reported that IgG antibodies are commonly observed in both asymptomatic and clinical infections (85% versus 94% of patients, respectively) (23), whereas durable B cell-mediated immunity against SARS-CoV-2 after mild or severe disease occurs in most individuals (24).
To further study COVID pathogenesis, herein we aimed to identify mRNA expression patterns that could serve as baseline correlates for development, or not,of clinical symptoms following contact with SARS-CoV-2. Since variations in the strength and/or extent of the immune response may be critical for the clinical picture and progress after infection with SARS-CoV-2, existing inter-individual differences at the transcriptome level may be observed even later, after convalescence. Indeed, gene expression pattern in blood samples collected 21 days after influenza infection are indistinguishable from baseline (25). Therefore, we performed 3’ mRNA next generation sequencing-based genome-wide transcriptional whole blood profiling to test the hypothesis that the baseline mRNA expression of theimmune response-related genes are differentially expressed between healthy individuals who developed an equally robust antibody response following either an entirely asymptomatic or clinical SARS-CoV-2 infection.
Materials and Methods
Blood Collection and Anti-SARS-CoV-2 Antibody Testing
Blood samples were collected from members of the NKUA, Athens, Greece in June–November 2020. The protocol was approved by the Ethics and Bioethics Committee of the School of Medicine, NKUA (protocol #312/02-06-2020) and study participants provided written informed consent. All plasma samples were analyzed as previously described (4) using, a) the CE-IVD Roche Elecsys® Anti-SARS-CoV-2 test, an electrochemiluminescence immunoassay (ECLIA) for the detection of total antibodies (IgG, IgM, and IgA; pan-Ig) to SARS-CoV-2 N-protein (Roche Diagnostics GmbH, Mannheim, Germany), and b) the CE-IVD Roche Elecsys® Anti-SARS-CoV-2 S, an ECLIA for the quantitative determination of antibodies (including IgGs) to the SARS-CoV-2 S-protein RBD (Roche Diagnostics).
3’ mRNA Sequencing, Mapping, Quality Control, and Quantifications
Total RNA was isolated from whole blood, stored in paxgene, using the ExtractionMonarch® Total RNA Miniprep Kit (NEB #T2010). Upon blood isolation, Monarch DNA/RNA Protection Reagent (supplied as a 2x concentrate) was added undiluted to an equal volume of blood. Addition of the protection reagent and the following RNA isolation was performed as described in the kit’s manual for Total RNA Purification from Mammalian Whole Blood Samples.
After quantification on a NanoDrop ND-1000 (Thermofisher) and Bioanalyzer RNA 6000 Nano assay (Agilent), 140-300ng of total RNA from samples passing quality control were processed using the QuantSeq 3’ mRNA-Seq Library Prep Kit FWD (Lexogen, 015.96) for library preparation. Libraries were assessed for molarity and median library size using Bioanalyzer High Sensitivity DNA Analysis (Agilent, 5067-4626). After multiplexing and addition of 13% PhiX Control v3 (Illumina, FC-110-3001) as spike in, the NGS was performed on a NextSeq550 with NextSeq 500/550 High Output Kit v2.5 - 75 cycles (Illumina, 20024906).
The quality of FASTQ files was assessed using FastQC (version 0.11.9) (26). The reads were mapped to the GRCh38 reference human genome using STAR, as part of a pipeline provided by Lexogen and BlueBee. After quality control, we obtained quantifications for 16.737 (12.789 protein coding) genes with more than five reads in more than 25% of the 17 asymptomatic and 15 clinical disease samples. Raw bam files, one for each sample, were summarized to a 3’UTR read counts table, using the Bioconductor package GenomicRanges (27), through metaseqR2 (28). The gene counts table was normalized for inherent systematic or experimental biases (e.g., sequencing depth, gene length, GC content bias) using the Bioconductor package EDASeq (29). For the downstream analysis, 12 hemoglobin (HBQ1, HBG2, HBZ, HBA2, HBA1, HBM, HBZP1, HBE1, HBG1, HBD, HBBP1, HBB) genes were removed from all samples.
Blood Immune Cell Subsets Deconvolution
CIBERSORTx (30) was used to estimate the proportion of blood immune cell subsets for each individual. As a signature matrix, the LM22 signature matrix for 22 subsets obtained at the single cell level was used. The Mann-Whitney U test was applied in order to calculate the significance of the difference in distributions between the asymptomatic and clinical groups. Statistical significance calculation and plotting were applied with R.
Differential Gene Expression
The resulting gene counts table was subjected to differential expression analysis (DEA) to compare individuals with a history of asymptomatic versus clinical (“symptomatic”) infection using the Bioconductor packages DESeq (31), edgeR (32), NOISeq (33), limma (34), NBPSeq (35), baySeq (36). In order to combine the statistical significance from multiple algorithms and perform meta-analysis, the PANDORA weighted P-value across results method was applied through metaseqR2. The weighted meta p-value was used as metric for the statistical significance for the differentially expressed genes. Multidimensional scaling was also applied through metaseqR2. DAVID analysis (37) was performed for the increased and decreased genes, both for enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways and for biological processes [Gene Ontology (GO)]. For the prediction of enriched regulons in asymptomatic disease we used the TRRUST (v2) reference transcription factor (TF)–target interaction database (38) and enrichR (39) focusing on the ChEA prediction with the increased genes in asymptomatic disease as input. For the identification of interferon-regulated genes the interferome database (v2) (40) was used.
Real Time PCR
Validation of the gene expression signatures was performed not only on the 32 samples for which the RNAseq propocol was followed, but also for 9 additional available samples (5 and 4 from prior asymptomatic or clinical infection, respectively). Briefly, 1000 ng starting material (or 500 ng for the samples with limited available RNA) was reverse transcribed to cDNA using a Takara PrimeScript RT Reagent Kit (Takara RR037A), following the manufacturer’s protocol. Real-time quantitative PCR (qPCR) was performed using the SYBR™ Select Master Mix, Applied Biosystems, ThermoFisher Scientific on the LightCycler® 96, Roche. Primers specific for each gene were designed with Primer Blast, while GAPDH served as the housekeeping gene (Supplementary Table S1.). All samples were measured in duplicates. Relative expression of each sample was defined as 2^-DCt, where DCt = Ct (gene target) – Ct (housekeeping gene).
Results
Whole Blood Transcriptional Profiling and Determination of Immune Cell Subsets in Healthy Seropositive Individuals With Prior Asymptomatic Versus Clinical Infection Time PCR
As shown in Table 1, the two groups under study comprised 15 seropositive individuals (9 men, mean age 34 years) who experienced clinical infection 7 to 25 weeks before sampling (mean 13; SD5.3)and 17 seropositive individuals (11 men, mean age 37 years)with entirely asymptomatic infection. Of those with clinical infection, 12/15 experienced fever and only one was in need of hospitalization which was uneventful. Individuals were considered asymptomatic in the absence of any symptoms since the onset of the pandemic, according to a detailed history obtained by a physician (absence of fever of any grade, fatigue, conjunctivitis/sweating coughs, headaches, respiratory distress/dyspnea, smell or taste loss, diarrhea). These individuals reported putative exposure to SARS-CoV-2 5 to 21 weeks before sampling (mean 14; SD 6.8).

Table 1 Demographics and antibody measurements.
Whole blood-derived, 3’ mRNA next generation sequencing-based, genome-wide transcriptional profiling was performed and, overall, more than 386 million reads were generated. Genes with fewer than five counts in fewer than 25% of the samples were filtered out, resulting to 16.737 profiled genes, of which 12.789 were protein coding. Twelve hemoglobin genes (HBQ1, HBG2, HBZ, HBA2, HBA1, HBM, HBZP1, HBE1, HBG1, HBD, HBBP1, HBB) were removed. A multidimensional scaling (MDS) plot generated using all 16.737 expressed genes, in order to avoid gene-type biases, revealed no clear separation of the two sample groups (Figure 1A).
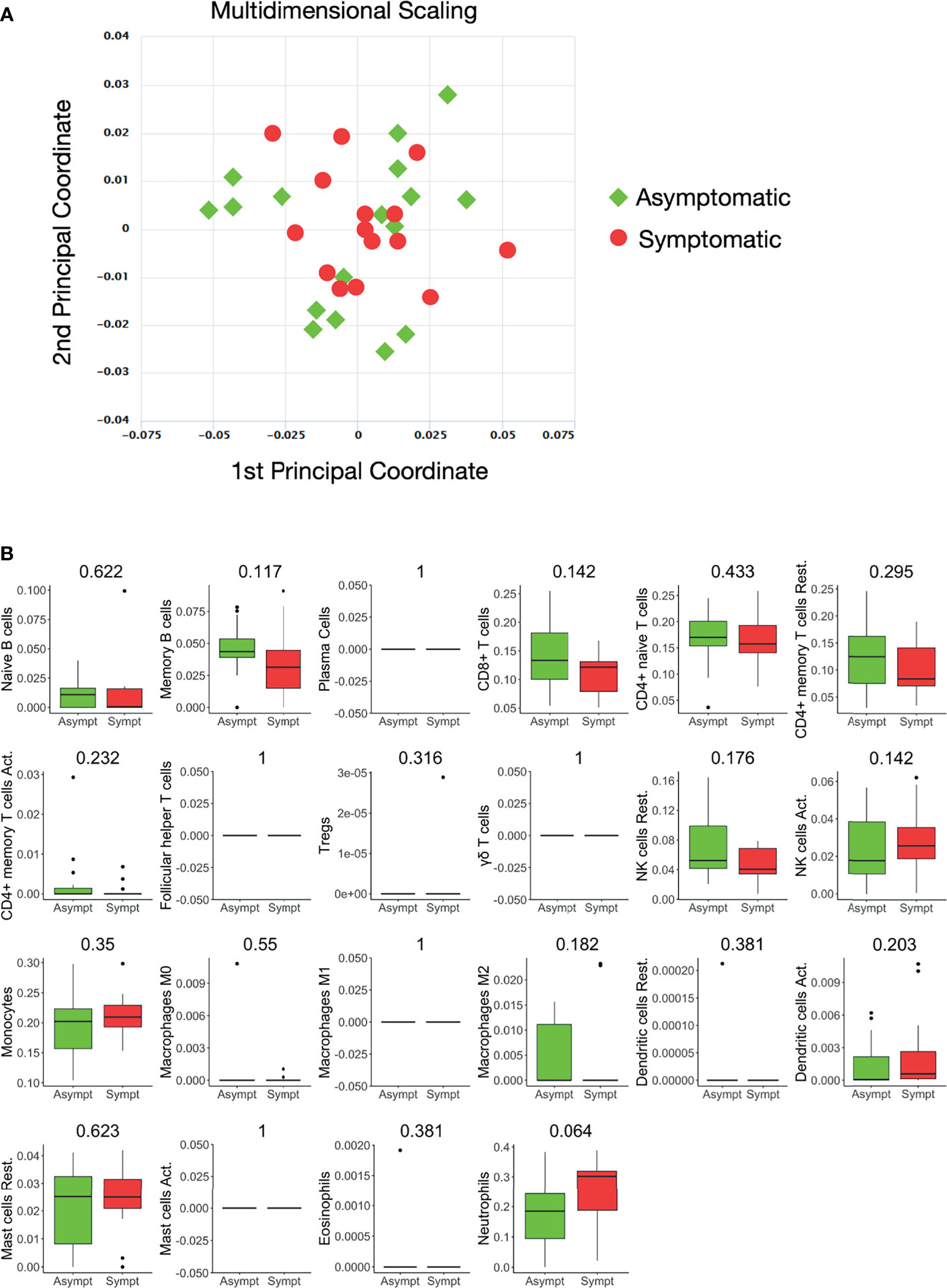
Figure 1 Whole blood transcriptional profiles and immune cell subsets in seropositive healthy individuals with prior asymptomatic or clinical SARS-CoV-2 infection. (A) Dimensionality reduction of all samples: Multidimensional scaling of all samples from individuals with prior clinical (n=15) and prior asymptomatic (n=17) infection. Each dot corresponds to the sample of one individual. All expressed elements were used (16.737, out of which 12.789 were non-zero protein-coding genes), in order to avoid gene type biases. The smaller the distance between each sample pair, the greater the similarity of the gene expression profile of the samples. No separation of the two sample groups is revealed, reflecting their similarity. (B) Blood transcriptome deconvolution with CYBERSORTx in prior asymptomatic (AS) and prior clinical (CL) disease groups. For every cell type, the Mann-Whitney U test p-value comparing the two groups is displayed on top. No statistically significan differences (meta p-value< 0.05) were detected between the two groups.
The proportions of immune cell populations, namely, naive B cells, memory B cells, plasma cells, CD8+ T cells, naive CD4+ T cells, resting memory CD4+ T cells, activated memory CD4+ T cells, follicular helper T cells, regulatory T cells, gamma delta T cells, resting NK cells, activated NK cells, monocytes, M0 macrophages, M1 macrophages, M2 macrophages, resting dendritic cells, activated dendritic cells, resting mast cells, activated mast cells, eosinophils and neutrophils in the peripheral blood estimated by CIBERSORTx were also comparable between the two groups (Figure 1B).
Differentially Expressed Genes Are Associated With Innate Immunity and Interferon Activity
Although the differential expression analysis of 12.777 protein coding genes did not reveal a distinct transcriptional profile between the two groups of healthy individuals, 24 genes were returned as differentially expressed (logFC=|1|, meta p-value<0.05) in a primary analysis (Supplementary Figure S1). Because the number of these genes was small, we repetitively applied the DEA pipeline, removing samples that were possible outliers in terms of expression of each differentially expressed gene. Therefore, genes that were repeatedly returned as significantly differentially expressed in those with prior asymptomatic infection relatively to those with clinical SARS-CoV-2 infection were characterized as differently expressed (Figure 2). Brief description of the function of six and nine genes that were found significantly decreased (Supplementary Table S2) and increased (Supplementary Table S3), respectively, in prior asymptomatic versus clinical SARS-CoV-2 infection is shown in Supplementary Tables.
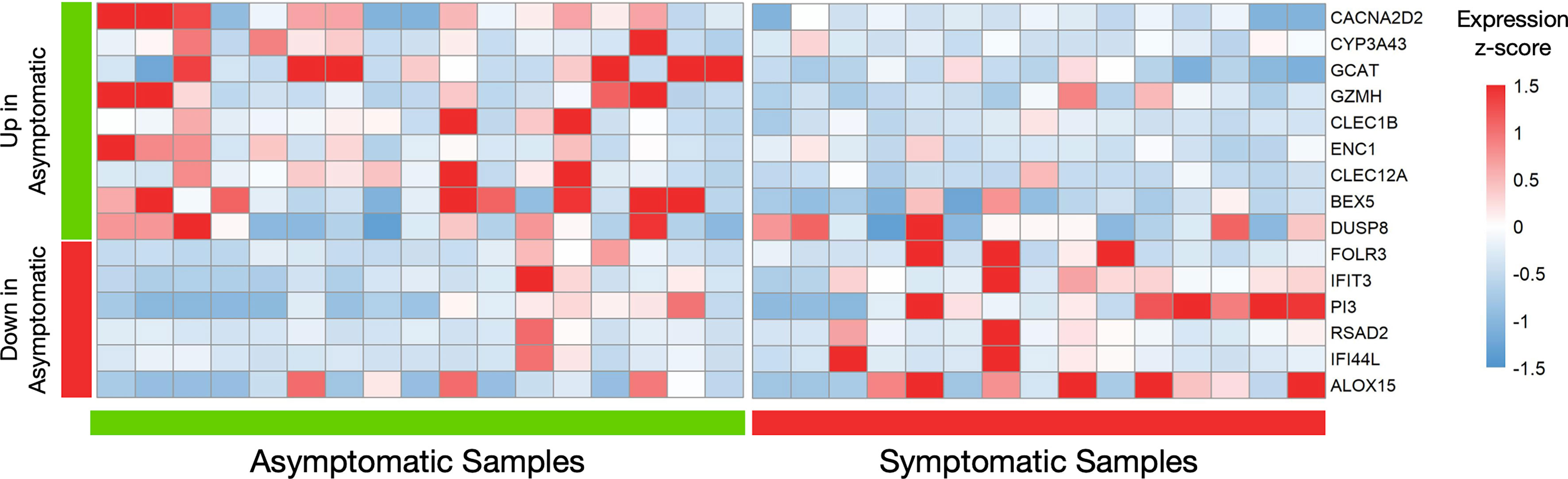
Figure 2 Differential gene expression analysis in seropositive healthy individuals with prior asymptomatic or clinical SARS-CoV-2 infection. Heatmap of robustly differentially expressed genes (genes that were differentially expressed and highly expressed in three or more samples, logFC>|1|, meta p-value<0.05) in individuals with prior asymptomatic infection relatively to those with clinical (“symptomatic”) SARS-CoV-2 infection, with raw expression values being scaled. The values for all samples (17 asymptomatic on the left and 15 clinical on the right) is plotted. The first nine genes are increased in the Asymptomatic group, while the next six are decreased.
In order to validate our DGEA results we performed SYBR Green-based qPCR to quantify the mRNA expression of 5 randomly selected differentially expressed genes Indeed, all 5 genes showed the same pattern of expression as in the RNA-sequencing experiment, reaching statistical significance in 4 genes, despite the small sample size (Supplementary Figure S1).The statistical significance remained excluding the 9 additional samples that were not included in the RNAseq run from the PCR validation (data not shown).
Notably, all six decreased genes in asymptomatic SARS-CoV-2 infection (IFIT3, IFI44L, FOLR3, RSAD2, PI3, ALOX15), are involved in innate immune responses (41–45) while the first two are interferon-inducible genes. Similarly, three increased genes (GZMH, CLEC1B, CLEC12A) are involved in innate immunity mechanisms (43, 46, 47), one (GCAT) in viral mRNA translation (48), one (CACNA2D2) in the integration of energy metabolism (49) and one (ENC1) in oxidative stress responses (50). The expression patterns of these 15 genes across all samples are depicted in Figure 2. Enrichment analysis returned no statistically significant enriched KEGG or GO terms. Similarly, there were no common upstream transcriptional regulators revealed by transcription factor (TF)–target interaction databases for these genes.
Furthermore, the interferome database which hosts genomic and transcriptomic data generated from cells or tissues treated with interferons was used for the 15 genes that were found to be differently expressed in asymptomatic versus clinical SARS-CoV-2 infections. Collectively, 8 out of 15 genes are regulated by interferons (ENC1, FOLR3, IFIT3, PI3, RSAD2, IFI44L, CLEC12A, ALOX15). Specifically, six genes are regulated by both type I and type II Interferons (ENC1, FOLR3, IFIT3, PI3, RSAD2, IFI44L), whereas the remaining two are targets of interferon type II only (CLEC12A, ALOX15) (40) (Figure 3).
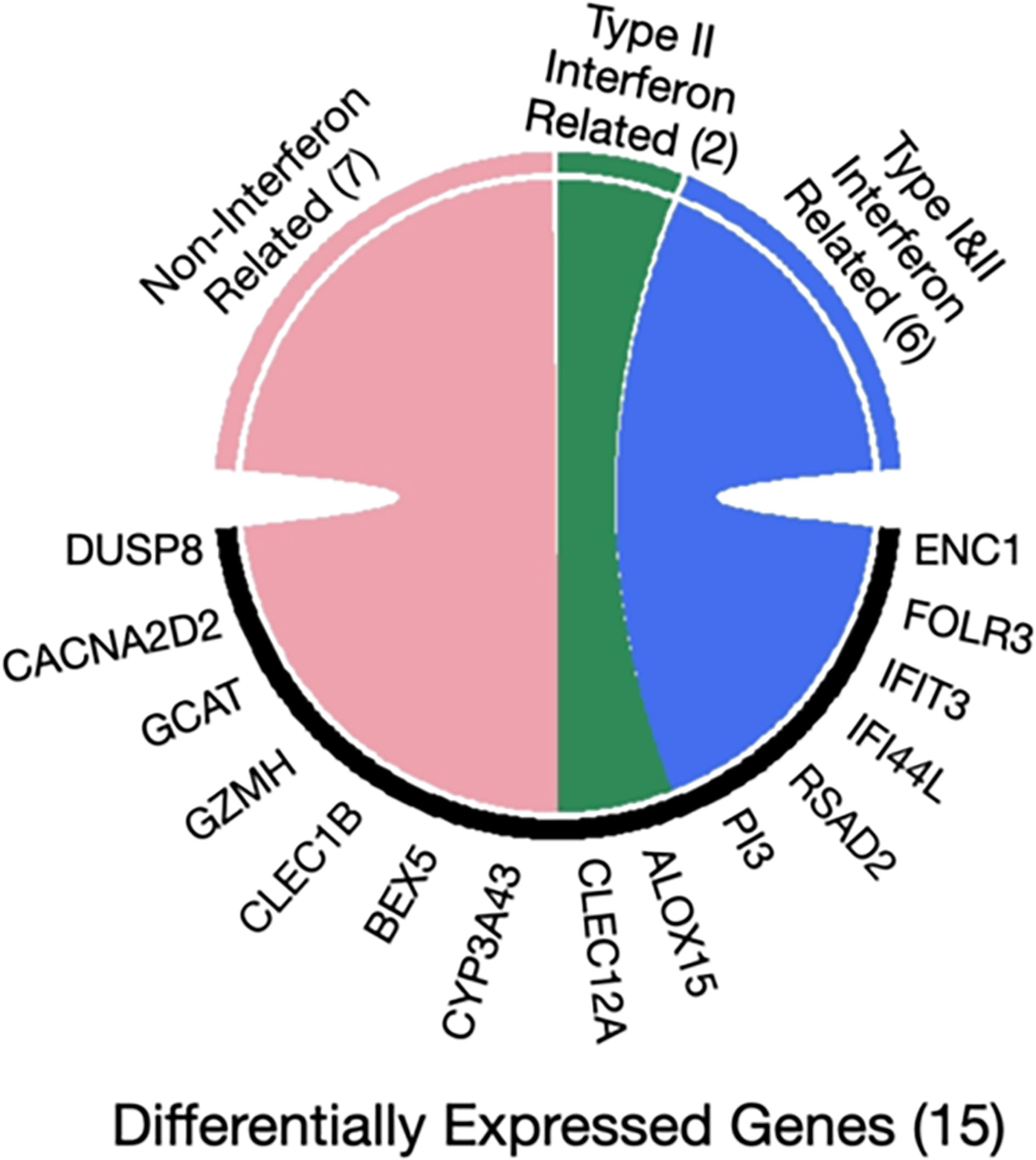
Figure 3 Chord diagram of differentially expressed genes in seropositive healthy individuals with prior asymptomatic or clinical SARS-CoV-2 infection with respect to interferon activity. The 15 genes characterized as differentially expressed in those with prior asymptomatic relatively to those with clinical infection were queried in the Interferome database and 8/15 were found to be associated with interferon activity. Of those, 6 are regulated by both interferon type I and II (blue), while 2 genes only by type II (green) and none by type III; the 7 remaining genes are not regulated by interferons (pink).
Discussion
Genome-wide transcriptome analyses studies using next generation sequencing technology in patients infected with SARS-CoV-2 provide evidence that transcriptome-wide changes may serve as predictors of morbidity and possibly of response to specific therapies (51). In addition, transcriptomic analyses may provide mechanistic insights into certain complications associated with SARS-CoV-2 infection (52). To our knowledge, this is the first whole blood genome-wide transcriptomic comparative analysis in seropositive healthy individuals who either experienced a clinical SARS-CoV-2 infection or an entirely asymptomatic infection around 3 months before sampling and developed an equally robust antibody response. In one previous study published so far in asymptomatic seropositive individuals infected in a super spreading event, the transcriptome in peripheral blood mononuclear cells was similar to that of seronegative highly exposed individuals from the same community. The putative time of infection of seropositive asymptomatic individuals was 4–6 weeks prior to sample collection, suggesting that the development of antibody response following viral exposure in asymptomatic cases is not necessarily associated with sustained alterations in the immune transcriptome (53).
Variations in innate immune system responses and cytokine networks could explain, at least in part, the wide heterogeneity in clinical presentation of SARS-CoV-2 infection (54). The symptom that best reflects the potency of the immune response, namely fever, has been repeatedly shown to be a poor diagnostic marker in severe disease (55, 56). Along these lines, it has been speculated that asymptomatic infection could be partly explained by the examples of altered innate immunity mechanisms operating in bats and pangolins. Despite carrying an enormous load of viral species, these animals display an apparent genetic resistance to coronavirus pathology (57). For example, decomposition of many type I interferon genes (58) and partial loss of function in stimulator of interferon genes (STING) is observed in bats (59). Regarding pangolins, recent findings suggest that these animals have lost interferon-ϵ (60) as well as interferon-induced with helicase C domain 1 (IFIH1), also known as IFIH1/MDA5 (61).
Our results provide evidence that among 12.777 genes, there were only 15 with significantly different expression when comparing healthy, relatively young individuals after convalescence from a previous entirely asymptomatic SARS-CoV-2 infection to those with a clinical infection history. While there were no apparent differences in cellular components and no specific immune deficiencies or co-morbidities to explain the different clinical presentation, the small, only, number of differentially expressed genes is expected since the cohort comprised healthy individuals at the time of sampling, who experienced in the past a SARS-CoV-2 infection. The small number of differentially expressed genes was also the reason why further bioinformatics analysis, i.e. enrichment and/or functionality analysis could not be applied. It should be highlighted that the transcriptome analysis was performed several weeks after the time of active infection; thus, certain potential differential responses may have been blunted during assessment after infection. This could also explain the limitation of the absence of differentially expressed genes with >2-fold change in our primary analysis. However, such differential responses should be more robust at the time of infection and more genes and immune networks may be differentially expressed.
Among the six genes that were found with significantly decreased expression in previously asymptomatic cases relatively to clinical cases, and in line with our research hypothesis, all are involved in innate immune responses (Supplementary Table S2) and two of these genes (IFIT3, IFI44L) belong to the interferon-induced family of genes. Overall, 8 of the 15 differentially expressed genes in those with prior asymptomatic infection relatively to those with clinical SARS-CoV-2 infection can be found in datasets that include genes which have been implicated in interferon related signaling pathways in vitro (38). A detailed explanation according the functionality of those genes and the pathways though which they act requires further studies. However, on the basis of these findings some assumptions can be made, since, indeed, the first 6 genes with higher expression in individuals with clinical infection compared to asymptomatic individuals, namely IFIT3, IFI44L, FOLR3, RSAD3, PI3, and ALOX15, share some common characteristics that can be relevant. For example, IFIT3, IFI44L and RSAD3 expression can be induced by viruses which in turn enhances the progress of the viral infection (62–64), whereas higher intrinsic expression of IFIT3, FOLR3, PI3 and ALOX-15 have all been associated with immune-mediated chronic diseases (65–68) In contrast, a protective effect of GZMH, CLEC1B, CLEC12A, that have a higher expression in asymptomatic individuals, may be associated with the effectiveness of GZMH in viral eradication (69) and the ability of CLEC1B and CLEC12A to enhance neutrophil extracellular trap formation, thus presenting an antiviral effect that helps to control systemic virus levels (70). Despite the fact that our findings have to be validated in a larger independent cohort of prior SARS-CoV-2 infected individuals, taken together with those of the literature support the hypothesis that there are differences in the innate immune responses between clinical and asymptomatic individuals during SARS-CoV-2 infections (15, 16).
As happens in all viral infections, type I interferon response plays a major protective role for the host because not only promotes viral clearance but also triggers a prolonged adaptive immune response (71). Insights into the innate and adaptive immune responses to SARS-CoV-2 have been gained by many research efforts over the past year (52). The innate immune responses that protect against disease and particularly the role of type I and III interferons have been addressed in numerous studies, mainly in patients with severe disease at the time of sampling. Important findings by Casanova and collaborators have shown that either neutralizing autoantibodies to type I interferons (72) or deleterious mutations in components involved in interferon induction or signaling (17) predispose patients to life-threatening infections. Along these lines, a highly impaired type I interferon response has been reported in patients with severe disease (73). However, in contrast to these findings, increased levels of interferons and interferon-stimulated genes have been observed in severe and life-threatening infections in many other studies (74–76). Indeed, increased interferon-alpha levels are a biomarker of mortality (77).
Moreover, the SARS-CoV-2 receptor ACE2, which is expressed in specific cell subsets across tissues is an interferon-stimulated gene in human airway epithelial cells (78), suggesting that a weaker individual interferon response may be protective. The latter may explain the low infection levels and morbidity in children (55, 56) who, relative to adults, display, in general lower interferon responses (79) and lower ACE2 expression (80). Taken together, in individuals infected with SARS-CoV-2, interferon-mediated responses may be protective or detrimental depending on the timing and the stage of infection, in addition to other factors, including viral load, age, and co-morbidities (71, 81, 82).
To conclude, our results suggest that subtle differences in the expression levels of innate immunity-related genes, including lower expression of genes involved in interferon signaling, may be beneficial for the host upon SARS-CoV-2 infection. The current study attempts to fill the existing gap regarding the potential implication of certain pathways in the clinical phenotype of SARS-CoV-2 infection. The described association of a ‘weaker’ immune response to SARS-CoV-2 with a lack of clinical symptoms needs further investigation, which hopefully will be performed in the near future. Whether a certain innate immunity signature predicts, or not, those who will develop a more successful immune response upon contact with SARS-CoV-2, with possible implications for prioritization of vaccination, warrant further study.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/, GSE173317.
Ethics Statement
The protocol was approved by the Ethics and Bioethics Committee of the School of Medicine, NKUA (protocol #312/02-06-2020). The patients/participants provided their written informed consent to participate in this study.
Author Contributions
Conceptualization, PS, and MD. Methodology, PS and KMV. Formal analysis and investigation, PS and KMV. Writing - original draft preparation, PS. Writing - review and editing, PS, KMV, GA-M, OT, DP, EK, EL, ParM, ET, IT, VC, MM, PanM, GP, GK, PH, and MD. Funding acquisition, PS and MD. Resources, OT, EL, ParM, ET, and IT. Supervision: PS and MD. All authors contributed to the article and approved the submitted version.
Funding
We acknowledge financial support of this work by the project pMedGR (MIS 5002802), funded by the Operational Programme “Competitiveness, Entrepreneurship and Innovation’ (NSRF 2014-2020) and co-financed by Greece and the European Union (European Regional Development Fund)”. Furthermore, we acknowledge financial support of “SAFEA” (SCIENTIFIC ASSOCIATION FOR THE SUPPORT OF THE FDPM EDUCATIONAL & RESEARCH ACTIVITIES), National Kapodistrian University of Athens.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Professor Argyrios Theofilopoulos for critical review of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.746203/full#supplementary-material
References
1. Macedo A, Gonçalves N, Febra C. COVID-19 Fatality Rates in Hospitalized Patients: Systematic Review and Meta-Analysis. Ann Epidemiol (2021) 57:14–21. doi: 10.1016/j.annepidem.2021.02.012
2. Lim ZJ, Subramaniam A, Reddy MP, Blecher G, Kadam U, Afroz A, et al. Case Fatality Rates for Patients With COVID-19 Requiring Invasive Mechanical Ventilation. A Meta-anal Am J Respir Crit Care Med (2021) 203(1):54. doi: 10.1164/rccm.202006-2405OC
3. Oran DP, Topol EJ. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Ann Internal Med (2020) 173(5):362–7. doi: 10.7326/M20-6976
4. Tsitsilonis OE, Paraskevis D, Lianidou E, Terpos E, Akalestos A, Pierros V, et al. SARS-CoV-2 Infection Is Asymptomatic in Nearly Half of Adults With Robust Anti-Spike Protein Receptor-Binding Domain Antibody Response. Vaccines (2021) 9(3):207. doi: 10.3390/vaccines9030207
5. Oran DP, Topol EJ. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Ann Internal Med (2020) 173(5):362–7. doi: 10.7326/M20-3012
6. Li R, Pei S, Chen B, Song Y, Zhang T, Yang W, et al. Substantial Undocumented Infection Facilitates the Rapid Dissemination of Novel Coronavirus (SARS-CoV-2). Science (2020) 368(6490):489–93. doi: 10.1126/science.abb3221
7. Buitrago-Garcia D, Egli-Gany D, Counotte MJ, Hossmann S, Imeri H, Ipekci AM, et al. Occurrence and Transmission Potential of Asymptomatic and Presymptomatic SARS-CoV-2 Infections: A Living Systematic Review and Meta-Analysis. PloS Med (2020) 17(9):e1003346. doi: 10.1371/journal.pmed.1003346
8. Morzunov S, Deyde V, Abrahamyan L. New Biomarkers of Innate and Adaptive Immunity in Infectious Diseases. J Immunol Res (2017) 2017:1–3. doi: 10.1155/2017/7047405
9. Gupta M, Mohanta SS, Rao A, Parameswaran GG, Agarwal M, Arora M, et al. Transmission Dynamics of the COVID-19 Epidemic in India and Modeling Optimal Lockdown Exit Strategies. Int J Infect Dis (2020 103:579–89. doi: 10.1101/2020.05.13.20096826
10. Misin A, Antonello RM, Bella SD, Campisciano G, Zanotta N, Giacobbe DR, et al. Measles: An Overview of a Re-Emerging Disease in Children and Immunocompromised Patients. Microorganisms (2020) 8(2):276. doi: 10.3390/microorganisms8020276
11. Martins MS, Pimentel F, Costa W, Fernandes C. COVID-19 in a Patient Newly Diagnosed With AIDS: Asymptomatic Disease. Eur J Case Rep Internal Med (2021) 8(1):002204. doi: 10.12890/2021_002204.
12. Marcus N, Frizinsky S, Hagin D, Ovadia A, Hanna S, Farkash M, et al. Minor Clinical Impact of COVID-19 Pandemic on Patients With Primary Immunodeficiency in Israel. Front Immunol (2021) 11:3505. doi: 10.3389/fimmu.2020.614086
13. Goodnow CC. COVID-19, Varying Genetic Resistance to Viral Disease and Immune Tolerance Checkpoints. Immunol Cell Biol (2021) 99(2):177–91. doi: 10.1111/imcb.12419
14. García LF. Immune Response, Inflammation, and the Clinical Spectrum of COVID-19. Front Immunol (2020) 11:1441. doi: 10.3389/fimmu.2020.01441
15. Long QX, Tang XJ, Shi QL, Li Q, Deng HJ, Yuan J, et al. Clinical and Immunological Assessment of Asymptomatic SARS-CoV-2 Infections. Nat Med (2020) 26(8):1200–4. doi: 10.1038/s41591-020-0965-6
16. Chan YH, Fong SW, Poh CM, Carissimo G, Yeo NK, Amrun SN, et al. Asymptomatic COVID-19: Disease Tolerance With Efficient Anti-Viral Immunity Against SARS-CoV-2. EMBO Mol Med (2021) 13(6):e14045. doi: 10.15252/emmm.202114045
17. Casanova JL, Su HC, Abel L, Aiuti A, Almuhsen S, Arias AA, et al. A Global Effort to Define the Human Genetics of Protective Immunity to SARS-CoV-2 Infection. Cell (2020) 181(6):1194–9. doi: 10.1016/j.cell.2020.05.016
18. Brodin P, Davis MM. Human Immune System Variation. Nat Rev Immunol (2017) 17(1):21–9. doi: 10.1038/nri.2016.125
19. Brodin P, Jojic V, Gao T, Bhattacharya S, Angel CJ, Furman D, et al. Variation in the Human Immune System Is Largely Driven by Non-Heritable Influences. Cell (2015) 160(1-2):37–47. doi: 10.1016/j.cell.2014.12.020
20. Kono DH, Baccala R, Theofilopoulos AN. Inhibition of Lupus by Genetic Alteration of the Interferon-α/β Receptor. Autoimmunity (2003) 36(8):503–10. doi: 10.1080/08916930310001624665
21. Ramos I, Goforth C, Soares-Schanoski A, Weir DL, Samuels EC, Phogat S, et al. Antibody Responses to SARS-CoV-2 Following an Outbreak Among Marine Recruits With Asymptomatic or Mild Infection. Front Immunol (2021) 12:681586. doi: 10.3389/fimmu.2021.681586
22. Tsitsilonis OE, Paraskevis D, Lianidou E, Pierros V, Akalestos A, Kastritis E, et al. Seroprevalence of Antibodies Against SARS-CoV-2 Among the Personnel and Students of the National and Kapodistrian University of Athens, Greece: A Preliminary Report. Life (2020) 10(9):214. doi: 10.3390/life10090214
23. Jiang C, Wang Y, Hu M, Wen L, Wen C, Wang Y, et al. Antibody Seroconversion in Asymptomatic and Symptomatic Patients Infected With Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin Trans Immunol (2020) 9(9):e1182. doi: 10.1002/cti2.1182
24. Ogega CO, Skinner NE, Blair PW, Park HS, Littlefield K, Ganesan A, et al. Durable SARS-CoV-2 B Cell Immunity After Mild or Severe Disease. J Clin Invest (2021) 131(7):e145516. doi: 10.1172/JCI145516
25. Zhai Y, Franco LM, Atmar RL, Quarles JM, Arden N, Bucasas KL, et al. Host Transcriptional Response to Influenza and Other Acute Respiratory Viral Infections–a Prospective Cohort Study. PloS Pathog (2015) 11(6):e1004869. doi: 10.1371/journal.ppat.1004869
26. Fastqc. Available at: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
27. Lawrence M, Huber W, Pagès H, Aboyoun P, Carlson M, Gentleman R, et al. Software for Computing and Annotating Genomic Ranges. PloS Comput Biol (2013) 9. doi: 10.1371/journal.pcbi.1003118
28. Fanidis D, Moulos P. Integrative, Normalization-Insusceptible Statistical Analysis of RNA-Seq Data, With Improved Differential Expression and Unbiased Downstream Functional Analysis. Briefings Bioinf (2021) 22(3):bbaa156. doi: 10.1093/bib/bbaa156
29. Risso D, Schwartz K, Sherlock G, Dudoit S. GC-Content Normalization for RNA-Seq Data. BMC Bioinformatics (2011) 12:480. doi: 10.1186/1471-2105-12-480
30. Newman AM, Steen CB, Liu CL, Gentles AJ, Chaudhuri AA, Scherer F, et al. Determining Cell Type Abundance and Expression From Bulk Tissues With Digital Cytometry. Nat Biotechnol (2019) 37(7):773–82. doi: 10.1038/s41587-019-0114-2
31. Anders S, Huber W. Differential Expression Analysis for Sequence Count . Nat Prec (2010). doi: 10.1038/npre.2010.4282.1
32. Robinson MD, McCarthy DJ, Smyth GK. Edger: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics (2010) 26:139–40. doi: 10.1093/bioinformatics/btp616
33. Tarazona S, Garcia-Alcalde F, Dopazo J, Ferrer A, Conesa A. Differential Expression in RNA-Seq: A Matter of Depth. Genome Res (2011) 21:2213–23. doi: 10.1101/gr.124321.111
34. Smyth GK. limma: Linear Models for Microarray Data. In: Gentleman R, Carey VJ, Huber W, Irizarry RA, Dudoit S, editors. Bioinformatics and Computational Biology Solutions Using R and Bioconductor. Statistics for Biology and Health. New York, NY: Springer. (2005) doi: 10.1007/0-387-29362-0_23.
35. Di Y, Schafer D, Cumbie JS, Chang JH. The NBP Negative Binomial Model for Assessing Differential Gene Expression From RNA-Seq. Stat Appl Genet Mol Biol (2011) 10(1):1–28. doi: 10.2202/1544-6115.1637
36. Hardcastle TJ, Kelly KA. Bayseq: Empirical Bayesian Methods for Identifying Differential Expression in Sequence Count Data. BMC Bioinf (2010) 11:422. doi: 10.1186/1471-2105-11-422
37. Huang DW, Sherman BT, Lempicki RA. Bioinformatics Enrichment Tools: Paths Toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res (2009) 37(1):1–13. doi: 10.1093/nar/gkn923
38. Han H, Cho J-W, Lee S, Yun A, Kim H, Bae D, et al. TRRUST V2: An Expanded Reference Database of Human and Mouse Transcriptional Regulatory Interactions. Nucleic Acids Res (2018) 46:D380–6. doi: 10.1093/nar/gkx1013
39. Kuleshov MV, Jones MR, Rouillard AD, Fernandez NF, Duan Q, Wang Z, et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res (2016) 44:W90–97. doi: 10.1093/nar/gkw377
40. Rusinova I, Forster S, Yu S, Kannan A, Masse M, Cumming H, et al. INTERFEROME V2. 0: An Updated Database of Annotated Interferon-Regulated Genes. Nucleic Acids Res (2013) 41(database issue):D1040–6. doi: 10.1093/nar/gks1215
41. Schmeisser H, Mejido J, Balinsky CA, Morrow AN, Clark CR, Zhao T, et al. Identification of Alpha Interferon-Induced Genes Associated With Antiviral Activity in Daudi Cells and Characterization of IFIT3 as a Novel Antiviral Gene. J Virol (2010) 84(20):10671–80. doi: 10.1128/JVI.00818-10
42. Gaudet P, Livstone MS, Lewis SE, Thomas PD. Phylogenetic-Based Propagation of Functional Annotations Within the Gene Ontology Consortium. Briefings Bioinf (2011) 12(5):449–62. doi: 10.1093/bib/bbr042
43. Pruitt KD, Brown GR, Hiatt SM, Thibaud-Nissen F, Astashyn A, Ermolaeva O, et al. RefSeq: An Update on Mammalian Reference Sequences. Nucleic Acids Res (2014) 42(D1):D756–63. doi: 10.1093/nar/gkt1114
44. Chin KC, Cresswell P. Viperin (Cig5), an IFN-Inducible Antiviral Protein Directly Induced by Human Cytomegalovirus. Proc Natl Acad Sci (2001) 98(26):15125–30. doi: 10.1073/pnas.011593298
45. Zhao J, O'Donnell VB, Balzar S, Croix CM, Trudeau JB, Wenzel SE. 15-Lipoxygenase 1 Interacts With Phosphatidylethanolamine-Binding Protein to Regulate MAPK Signaling in Human Airway Epithelial Cells. Proc Natl Acad Sci (2011) 108(34):14246–51. doi: 10.1073/pnas.1018075108
46. Kerrigan AM, Dennehy KM, Mourão-Sá D, Faro-Trindade I, Willment JA, Taylor PR, et al. CLEC-2 Is a Phagocytic Activation Receptor Expressed on Murine Peripheral Blood Neutrophils. J Immunol (2009) 182(7):4150–7. doi: 10.4049/jimmunol.0802808
47. Pyż E, Huysamen C, Marshall AS, Gordon S, Taylor PR, Brown GD. Characterisation of Murine MICL (CLEC12A) and Evidence for an Endogenous Ligand. Eur J Immunol (2008) 38(4):1157–63. doi: 10.1002/eji.200738057
48. Guo Y, Parker JS. The Paradoxes of Viral mRNA Translation During Mammalian Orthoreovirus Infection. Viruses (2021) 13(2):275. doi: 10.3390/v13020275
49. Carboni GL, Gao B, Nishizaki M, Xu K, Minna JD, Roth JA, et al. CACNA2D2-Mediated Apoptosis in NSCLC Cells Is Associated With Alterations of the Intracellular Calcium Signaling and Disruption of Mitochondria Membrane Integrity. Oncogene (2003) 22(4):615–26. doi: 10.1038/sj.onc.1206134
50. Wang XJ, Zhang DD. Ectodermal-Neural Cortex 1 Down-Regulates Nrf2 at the Translational Level. PloS One (2009) 4(5):e5492. doi: 10.1371/journal.pone.0005492
51. Vabret N, Britton GJ, Gruber C, Hegde S, Kim J, Kuksin M, et al. Immunology of COVID-19: Current State of the Science. Immunity (2020) 52(6):910–41. doi: 10.1016/j.immuni.2020.05.002
52. Carvalho T, Krammer F, Iwasaki A. The First 12 Months of COVID-19: A Timeline of Immunological Insights. Nat Rev Immunol (2021) 15:1–2. doi: 10.1038/s41577-021-00522-1
53. Lee HK, Knabl L, Pipperger L, Volland A, Furth PA, Kang K, et al. Immune Transcriptomes of Highly Exposed SARS-CoV-2 Asymptomatic Seropositive Versus Seronegative Individuals From the Ischgl Community. Sci Rep (2021) 11(1):1–0. doi: 10.1038/s41598-021-83110-6
54. Birra D, Benucci M, Landolfi L, Merchionda A, Loi G, Amato P, et al. COVID 19: A Clue From Innate Immunity. Immunol Res (2020) 68(3):161–8. doi: 10.1007/s12026-020-09137-5
55. Guan Wj, Ni ZY, Hu Y, Liang WH, Ou CQ, He JX, et al. Clinical Characteristics of Coronovirus Disease 2019 in China. N Engl J Med (2020) 382:1708–20. doi: 10.1056/NEJMoa2002032
56. Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical Features of Patients Infected With 2019 Novel Coronovirus in Wuhan, China. lancet (2020) 395(10223):497–506. doi: 10.1016/S0140-6736(20)30183-5
57. Shankar EM, Che KF, Yong YK, Girija AS, Velu V, Ansari AW, et al. Asymptomatic SARS-CoV-2 Infection: Is it All About Being Refractile to Innate Immune Sensing of Viral Spare-Parts?—Clues From Exotic Animal Reservoirs. Pathog Dis (2021) 79(1):ftaa076. doi: 10.1093/femspd/ftaa076
58. Zhou P, Tachedjian M, Wynne JW, Boyd V, Cui J, Smith I, et al. Contraction of the Type I IFN Locus and Unusual Constitutive Expression of IFN-α in Bats. Proc Natl Acad Sci (2016) 113(10):2696–701. doi: 10.1073/pnas.1518240113
59. Xie J, Li Y, Shen X, Goh G, Zhu Y, Cui J, et al. Dampened STING-Dependent Interferon Activation in Bats. Cell Host Microbe (2018) 23(3):297–301. doi: 10.1016/j.chom.2018.01.006
60. Choo SW, Rayko M, Tan TK, Hari R, Komissarov A, Wee WY, et al. Pangolin Genomes and the Evolution of Mammalian Scales and Immunity. Genome Res (2016) 26(10):1312–22. doi: 10.1101/gr.203521.115
61. Du Z, Sharma SK, Spellman S, Reed EF, Rajalingam R. KIR2DL5 Alleles Mark Certain Combination of Activating KIR Genes. Genes Immunity (2008) 9(5):470–80. doi: 10.1038/gene.2008.39
62. Xu F, Song H, An B, Xiao Q, Cheng G, Tan G. NF-κb-Dependent IFIT3 Induction by HBx Promotes Hepatitis B Virus Replication. Front Microbiol (2019) 10:2382. doi: 10.3389/fmicb.2019.02382
63. Li Y, Zhang J, Wang C, Qiao W, Li Y, Tan J. IFI44L Expression Is Regulated by IRF-1 and HIV-1. FEBS Open Bio (2021) 11(1):105–13. doi: 10.1002/2211-5463.13030
64. Seo JY, Yaneva R, Hinson ER, Cresswell P. Human Cytomegalovirus Directly Induces the Antiviral Protein Viperin to Enhance Infectivity. Science (2011) 332(6033):1093–7. doi: 10.1126/science.1202007
65. He T, Xia Y, Yang J. Systemic Inflammation and Chronic Kidney Disease in a Patient Due to the RNASEH2B Defect. Pediatr Rheumatol (2021) 19(1):1–9. 45. doi: 10.1186/s12969-021-00497-2
66. Tseng CC, Lin YZ, Lin CH, Li RN, Yen CY, Chan HC, et al. Next-Generation Sequencing Profiles of the Methylome and Transcriptome in Peripheral Blood Mononuclear Cells of Rheumatoid Arthritis. J Clin Med (2019) 8(9):1284. doi: 10.3390/jcm8091284
67. Rohde G, Message SD, Haas JJ, Kebadze T, Parker H, Laza-Stanca V, et al. Johnston SL: CXC-Chemokines and Antimicrobial Peptides in Rhinovirus-Induced Experimental Asthma Exacerbations. Clin Exp Allergy (2014) 44:930–9. doi: 10.1111/cea.12313
68. Singh NK, Rao GN. Emerging Role of 12/15-Lipoxygenase (ALOX15) in Human Pathologies. Prog Inlipid Res (2019) 73:28–. doi: 10.1016/j.plipres.2018.11.001
69. Tang H, Li C, Wang L, Zhang H, Fan Z. Granzyme H of Cytotoxic Lymphocytes Is Required for Clearance of the Hepatitis B Virus Through Cleavage of the Hepatitis B Virus X Protein. J Immunol (2012) 188(2):824–31. doi: 10.4049/jimmunol.1102205
70. Hiroki CH, Toller-Kawahisa JE, Fumagalli MJ, Colon DF, Figueiredo L, Fonseca BA, et al. Neutrophil Extracellular Traps Effectively Control Acute Chikungunya Virus Infection. Front Immunol (2020) 10:3108. doi: 10.3389/fimmu.2019.03108
71. Sa Ribero M, Jouvenet N, Dreux M, Nisole S. Interplay Between SARS-CoV-2 and the Type I Interferon Response. PloS Pathog (2020) 16(7):e1008737. doi: 10.1371/journal.ppat.1008737
72. Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, et al. Autoantibodies Against Type I IFNs in Patients With Life-Threatening COVID-19. Science (2020) 370(6515):eabd4585.
73. Zhang SY, Boisson-Dupuis S, Chapgier A, Yang K, Bustamante J, Puel A, et al. Inborn Errors of Interferon (IFN)-Mediated Immunity in Humans: Insights Into the Respective Roles of IFN-α/β, IFN-γ, and IFN-λ in Host Defense. Immunol Rev (2008) 226(1):29–40. doi: 10.1111/j.1600-065X.2008.00698.x
74. Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, Smith N, et al. Impaired Type I Interferon Activity and Inflammatory Responses in Severe COVID-19 Patients. Science (2020) 369(6504):718–24. doi: 10.1126/science.abc6027
75. Broggi A, Granucci F, Zanoni I. Type III Interferons: Balancing Tissue Tolerance and Resistance to Pathogen Invasion. J Exp Med (2020) 217(1):e20190295. doi: 10.1084/jem.20190295
76. Zhou Z, Ren L, Zhang L, Zhong J, Xiao Y, Jia Z, et al. Heightened Innate Immune Responses in the Respiratory Tract of COVID-19 Patients. Cell Host Microbe (2020) 27(6):883–90. doi: 10.1016/j.chom.2020.04.017
77. Galani IE, Rovina N, Lampropoulou V, Triantafyllia V, Manioudaki M, Pavlos E, et al. Untuned Antiviral Immunity in COVID-19 Revealed by Temporal Type I/III Interferon Patterns and Flu Comparison. Nat Immunol (2021) 22(1):32–40. doi: 10.1038/s41590-020-00840-x
78. Lucas C, Wong P, Klein J, Castro TB, Silva J, Sundaram M, et al. Longitudinal Analyses Reveal Immunological Misfiring in Severe COVID-19. Nature (2020) 584(7821):463–9. doi: 10.1038/s41586-020-2588-y
79. Ziegler CG, Miao VN, Owings AH, Navia AW, Tang Y, Bromley JD, et al. Impaired Local Intrinsic Immunity to SARS-CoV-2 Infection in Severe COVID-19. bioRxiv (2021). doi: 10.1101/2021.02.20.431155
80. Bunyavanich S, Do A, Vicencio A. Nasal Gene Expression of Angiotensin-Converting Enzyme 2 in Children and Adults. JAMA (2020) 323(23):2427–9. doi: 10.1001/jama.2020.8707
81. Park A, Iwasaki A. Type I and Type III Interferons–Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host Microbe (2020) 27(6):870–8. doi: 10.1016/j.chom.2020.05.008
82. Triggle CR, Bansal D, Ding H, Islam MM, Farag EA, Hadi HA, et al. A Comprehensive Review of Viral Characteristics, Transmission, Pathophysiology, Immune Response, and Management of SARS-CoV-2 and COVID-19 as a Basis for Controlling the Pandemic. Front Immunol (2021) 12:338. doi: 10.3389/fimmu.2021.631139
Keywords: innate immunity, anti-SARS-CoV2 antibody, asympomatic, RNAseq, whole-blood
Citation: Sfikakis PP, Verrou KM, Ampatziadis-Michailidis G, Tsitsilonis O, Paraskevis D, Kastritis E, Lianidou E, Moutsatsou P, Terpos E, Trougakos I, Chini V, Manoloukos M, Moulos P, Pavlopoulos GA, Kollias G, Hatzis P and Dimopoulos MA (2021) Blood Transcriptomes of Anti-SARS-CoV-2 Antibody-Positive Healthy Individuals Who Experienced Asymptomatic Versus Clinical Infection. Front. Immunol. 12:746203. doi: 10.3389/fimmu.2021.746203
Received: 23 July 2021; Accepted: 15 September 2021;
Published: 05 October 2021.
Edited by:
Levon Abrahamyan, Université de Montréal, CanadaReviewed by:
Hye Kyung Lee, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health (NIH), United StatesMatthew Wood, Seattle Children’s Research Institute, United States
Copyright © 2021 Sfikakis, Verrou, Ampatziadis-Michailidis, Tsitsilonis, Paraskevis, Kastritis, Lianidou, Moutsatsou, Terpos, Trougakos, Chini, Manoloukos, Moulos, Pavlopoulos, Kollias, Hatzis and Dimopoulos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Petros P. Sfikakis, cHNmaWtha2lzQG1lZC51b2EuZ3I=