 Sylvie Bouvier1,2,3†
Sylvie Bouvier1,2,3† Elise Kaspi4†
Elise Kaspi4† Ahmad Joshkon5
Ahmad Joshkon5 Odile Paulmyer-Lacroix6Marie-Dominique Piercecchi-Marti7Akshita Sharma8Aurélie S. Leroyer5Alexandrine Bertaud5
Odile Paulmyer-Lacroix6Marie-Dominique Piercecchi-Marti7Akshita Sharma8Aurélie S. Leroyer5Alexandrine Bertaud5 Jean-Christophe Gris1,2,3,9Françoise Dignat-George10Marcel Blot-Chabaud5
Jean-Christophe Gris1,2,3,9Françoise Dignat-George10Marcel Blot-Chabaud5 Nathalie Bardin11*
Nathalie Bardin11*- 1Department of Hematology, Nîmes University Hospital, Nîmes, France
- 2Faculty of Pharmaceutical and Biological Sciences, University of Montpellier, Montpellier, France
- 3UA11 Institute Desbrest of Epidemiology and Public Health, INSERM, Univ Montpellier, Montpellier, France
- 4Aix Marseille Univ, APHM, INSERM, MMG, Hôpital la Timone, Service de Biologie Cellulaire, Marseille, France
- 5Aix-Marseille Univ, INSERM, INRAE, C2VN, Marseille, France
- 6Aix Marseille Univ, APHM, Hôpital la Conception, Laboratory of Histology-Embryology/Biology of Reproduction, Marseille, France
- 7Aix Marseille Univ, APHM, CNRS, EFS, ADES, Hôpital La Timone, Forensic Department, Marseille, France
- 8Department of Stem Cell and Regenerative Medicine, Centre for Interdisciplinary Research, DY Patil University, Kolhapur, India
- 9I.M. Sechenov First Moscow State Medical University, Moscow, Russia
- 10Aix Marseille Univ, APHM, INSERM, INRAE, C2VN, Hôpital la Conception, Laboratoire d’Hématologie, Marseille, France
- 11Aix Marseille Univ, APHM, INSERM, INRAE, C2VN, Hôpital la Conception, Laboratoire d’Immunologie, Marseille, France
CD146 is an adhesion molecule essentially located in the vascular system, which has been described to play an important role in angiogenesis. A soluble form of CD146, called sCD146, is detected in the bloodstream and is known as an angiogenic factor. During placental development, CD146 is selectively expressed in extravillous trophoblasts. A growing body of evidence shows that CD146 and, in particular, sCD146, regulate extravillous trophoblasts migration and invasion both in vitro and in vivo. Hereby, we review expression and functions of CD146/sCD146 in the obstetrical field, mainly in pregnancy and in embryo implantation. We emphasized the relevance of quantifying sCD146 in the plasma of pregnant women or in embryo supernatant in the case of in vitro fertilization (IVF) to predict pathological pregnancy such as preeclampsia or implantation defect. This review will also shed light on some major results that led us to define CD146/sCD146 as a biomarker of placental development and paves the way toward identification of new therapeutic targets during implantation and pregnancy.
Introduction
Successful embryo implantation, placentation, and subsequent gestation depend on complex coordinated interactions between maternal and fetal tissues. Invasion of trophoblasts into the deciduum and the myometrium to subsequently establish the uteroplacental vasculature is indispensable for an effective pregnancy (1). During the early phase of pregnancy, cytotrophoblasts differentiate into two major cell lineages, the syncytiotrophoblasts and the extravillous trophoblasts (EVTs) that form endovascular and interstitial invasive trophoblasts. The interstitial invasive trophoblasts invade uterine tissue and anchor the placenta to the uterus, while the endovascular invasive trophoblasts migrate to the maternal uterine spiral arteries transforming them into large diameter conduit vessels of low resistance to establish the uteroplacental circulation (2). Trophoblasts invasion is regulated by not only various angiogenic growth factors but also adhesion molecules and oxygen concentration (3). Thus, migrating EVTs secrete the angiogenic growth factors: vascular endothelial growth factor (VEGF) and the soluble form of CD146 (sCD146), which promote angiogenesis in the decidua.
CD146, often referred to as MUC18, melanoma cell adhesion molecule (MCAM, Mel-CAM), is an adhesion molecule belonging to the immunoglobulin superfamily. This transmembrane glycoprotein of 113 kDa is present in several isoforms, short and long membrane isoforms (4), and a soluble form (sCD146) generated by a membrane proteolysis via metalloproteinases (5). CD146 has a preferential localization at endothelial cell junctions (6) and is expressed not only on all types of human endothelial cells but also on other cell types such as TH17 lymphocytes (7, 8), EVTs (9), and cancers of various origins as melanoma cells or malignant mesothelioma cells (10–13). Soluble CD146 is detected in the supernatant of cultured cells and in human sera from healthy patients and patients with pathologies associated with vascular disorders (14).
The angiogenic function of CD146 and its soluble form under both physiological and pathological conditions including cancers is well-documented and has been recently reviewed (15). A growing body of evidence shows that CD146, and in particular its soluble form, regulates obstetrical angiogenesis (16–18). This review will summarize our current understanding of CD146/sCD146 contribution in obstetrics.
Expression and Localization of CD146 at the Human Materno-Fetal Interface
CD146 is only expressed in uteri of pregnant women and is totally absent in uteri of non-pregnant women (17). Its expression on the placental villi exhibits a pattern of spatial selectivity, progressively increasing in the zone of anchoring between the villi and the deciduum, allowing the attachment of the placenta to the uterine wall (19). Indeed, after implantation, the outermost cell layer of the blastocyst, the trophectoderm, gives rise to mononuclear cytotrophoblasts forming placental villi through branching morphogenesis. Then, syncytiotrophoblasts are generated by cell fusion of villous cytotrophoblasts. Whereas syncytiotrophoblasts of floating villi represent the transport units of the human placenta, anchoring villi of the placental basal plate form another differentiated trophoblast type, the so-called invasive extravillous trophoblast (20). Thus, CD146 is mainly expressed on intermediate, a subtype of trophoblasts morphologically and functionally between syncytiotrophoblasts and cytotrophoblasts (21), and extravillous trophoblasts that are characterized by their high migrative and invasive capabilities. CD146 expression on intermediate trophoblasts facilitates their binding to uterine smooth muscle cells, which limits the extension of the trophoblast migration zone to the site of implantation (22).
Besides, CD146 is highly expressed in the endometrium and cumulus–oocyte complex and in the trophectoderm and inner cell mass of the blastocyst (23). Moreover, Liu et al. showed that CD146 is potently upregulated both in receptive maternal uteri and invasive embryonic trophoblasts only during early stages of pregnancy, which progressively fades afterwards (17). These data reinforce the importance of CD146 at the human materno-fetal interface.
Figure 1 highlights CD146 expression on human embryos and EVTs from human placenta.
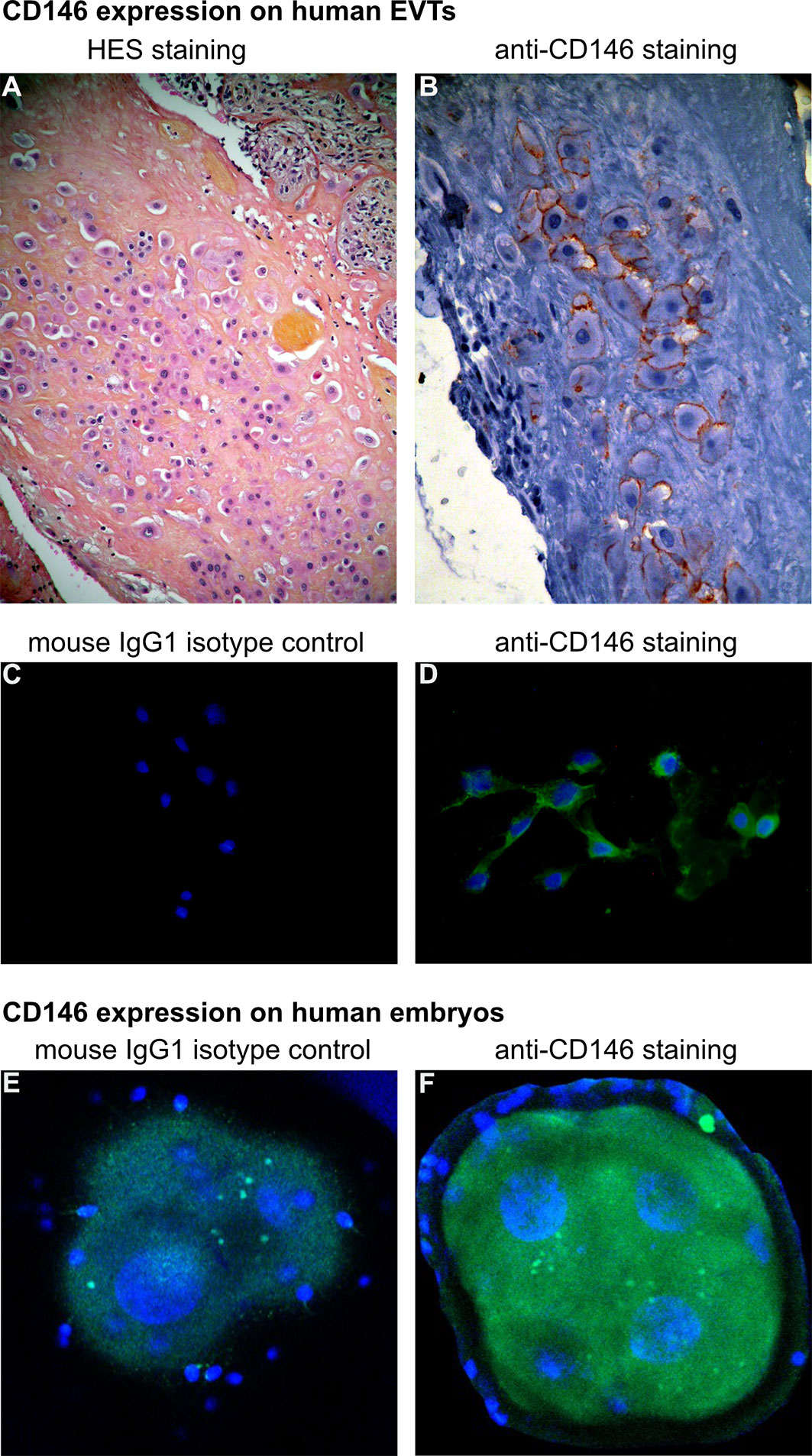
Figure 1 CD146 expression on human extravillous trophoblasts (EVTs) and embryos. (A, B) Characterization of EVTs in human placenta, from a normal term pregnancy. (A) EVTs are stained with hematoxylin and eosin (H&E) (objective 10×). (B) Immunohistochemistry with avidin–biotin–peroxidase complex, with CD146 antibody (monoclonal mouse antihuman, BioCytex S-Endo 1S-Endo1) (objective 20×). (C, D) Expression of CD146 on EVTs outgrowth from human placental villous explants culture (ex vivo model) [for model, see Kaspi et al., (18)]. (C) Cells were labeled with mouse IgG1 isotype control and (D) anti-CD146 antibody (Novocastra-Leica N1238). 4′,6-Diamidino-2-phenylindole (DAPI) was used to counterstain DNA (objective 20×). (E, F) Expression of CD146 on early embryo stage (day 2): immunostaining of (E) mouse IgG1 isotype control and (F) anti-CD146 (S-Endo1). DAPI was used to counterstain DNA. [Picture is from Bouvier et al, (24)] (objective 63×).
Strategic Roles of CD146 in Obstetrics
Extravillous trophoblasts play a crucial role in establishing the fetal–maternal circulation, essentially by invading the deciduum and remodeling the uterine spiral arteries. During early phases of pregnancy, CD146 appears to play an essential role in regulating intermediate and extravillous trophoblasts invasion and migration to ensure adequate embryo implantation and vascularization (16, 17). Indeed, in vitro and in vivo experiments showed that anti-CD146 blocking antibodies prevent embryo implantation by inhibiting MMP-2 and MMP-9 collagenase activity, impeding trophoblast proliferation and blocking deciduum angiogenesis and vascularization (17).
However, there is scarcity of knowledge concerning factors that regulate CD146 expression on extravillous trophoblasts. Data revealed that reduced expression of CD146 on the placenta is directly linked to preeclampsia, a pregnancy complication related to high blood pressure (16). Besides, it has been shown that during placentation, the deciduum cells promote CD146 expression on extravillous trophoblasts and enhance their differentiation by activating cAMP-dependent signaling pathway (25).
What About the Soluble Form of CD146 in Obstetrics?
Recent studies demonstrated the role of sCD146 as a regulator of trophoblast migration and potent stimulator of placental vascularization (18). In vitro experiments performed on EVTs cell line, HTR8/SVneo, showed that sCD146 inhibits their migration, invasion, and ability to form pseudo-capillary tubes in Matrigel. Likewise, ex vivo experiments on placental villous explants showed that sCD146 suppressed outgrowth and migration of EVTs. Accordingly, sCD146 was identified as a negative regulator of EVTs migration (18). Kaspi et al. (18) additionally investigated the in vivo functions of sCD146 in the placentation and fertility in a rat model. We showed that rats treated with recombinant sCD146 exhibit not only diminished number of pregnancies but also decreased number of embryos. Interestingly, histological studies performed on placenta evidenced reduced migration of Glycogen cells (GC), analogue to human extravillous cytotrophoblasts, in treated rats as compared to the controls (18). These results corroborate the inhibitory effect of sCD146 on extravillous trophoblasts migration. Therefore, it was proposed that sCD146 acts early in gestation, probably during implantation step and/or during placental development to regulate extravillous trophoblasts invasion.
How Does it Work?
Similarities between physiological blastocyst implantation and pathological neoplasm invasion are reported (26). Indeed, CD146 was first proposed as a marker of melanoma metastasis (27), but later, Liu et al. showed that CD146 promoted trophoblast invasion, implantation, and placentation (17). It is now well-recognized that the soluble form of multiple adhesion molecules implicated in pregnancy can modulate the function of the membrane protein, the best-known model being the effect of the soluble form of the vascular endothelial growth factor receptor-1 (VEGFR1), also called sFlt1 (28). In line with these data, it is assumed that sCD146 prevents the interaction between CD146 and its binding partners that act to induce trophoblasts invasion, differentiation, and placentation.
CD146 and galectin-1 (Gal1) are proangiogenic factors that are expressed in EVTs (29). To elucidate sCD146 mechanism of action on trophoblasts, HTR8/SVneo cells were treated with Gal1 and sCD146 (submitted manuscript). Results showed that these two molecules exerted an opposite effect on cell migration: sCD146 significantly decreasing HTR8/SVneo cells migration while Gal1 potentiating it. Of importance, sCD146 blocked Gal1-induced migratory effects on trophoblasts by inhibiting its secretion. This suggests that sCD146 acts as a ligand trap and antagonizes the effects and signaling mediated by membrane CD146. Besides, in vitro experiments using blocking anti-CD146 antibody or knocking down VEGFR2 inhibited Gal1-induced trophoblasts migration. Thus, it is proposed that the binding of Gal1 to CD146 on trophoblasts activates VEGFR2 signaling pathway.
In addition, the cognate interaction between VEGF and VEGFR2 is known to generate reactive oxygen species via NADPH-oxidase complex (NOX4) and Rac1 (protein of the Rho family) (30). As oxygen concentration is implicated in modifying trophoblasts invasion and differentiation (31), future work will validate if oxygen controls CD146 expression on trophoblasts or sCD146 generation.
Soluble CD146: A Potential Candidate Biomarker to Predict Pathological Pregnancies or Implantation Defects?
Under physiological conditions, the serum concentration of sCD146 progressively decreases throughout normal pregnancy, a result confirmed in two independent cohorts of patients (18, Bouvier et al., submitted manuscript). However, sCD146 concentration was found to be elevated in patients with at least one unexplained fetal loss as compared to women with at least one viable child (32). Bouvier et al. have quantified sCD146 plasma concentration in a cohort of women with placental-mediated pregnancy complications. They found sCD146 to be upregulated (21%) in women with preeclampsia as compared to women with normal pregnancy (submitted manuscript; clinicaltrials.gov identifier: NCT 01736826).
In view of these results, sCD146 may represent an attractive biomarker to assess abnormality in placental vascular development and constitute a potential predictive biomarker to discriminate between normal pregnancies and pathological ones.
In addition, since CD146 is detected only at early developmental stages of human embryos (day 2) and as in vitro fertilized eggs secreted sCD146 into their culture media, it has been proposed that sCD146 may act as a biomarker in reproductive medicine for evaluating embryos’ implantation potential. Data revealed that high concentration of sCD146 in embryo culture media is associated with lower implantation potentials (24). The sensitivity analysis performed on single embryo transfer showed that the optimal sCD146 concentration for a successful embryo implantation is just under 1,164 pg/ml. Beyond this value, the implantation rate decreased significantly [9% with sCD146 levels >1,164 pg/ml vs. 22% with sCD146 levels ≤1,164 pg/ml (24)]. Therefore, sCD146 was considered as an innovative biomarker for selecting the best embryos during in vitro fertilization (IVF).
Conclusion
In this review, we provide an overview of the diverse roles of CD146/sCD146 in human embryo implantation and pregnancy as summarized in Figure 2. CD146/sCD146 can be proposed as a biomarker of placental development. Taking into account the functional studies of CD146/sCD146 performed on EVTs and the sCD146 seric concentration in placental-mediated pregnancy complications, the description of the expression and functions of CD146/sCD146 also paves the way to the development of new therapeutic agents targeting CD146/sCD146 in obstetrics complications.
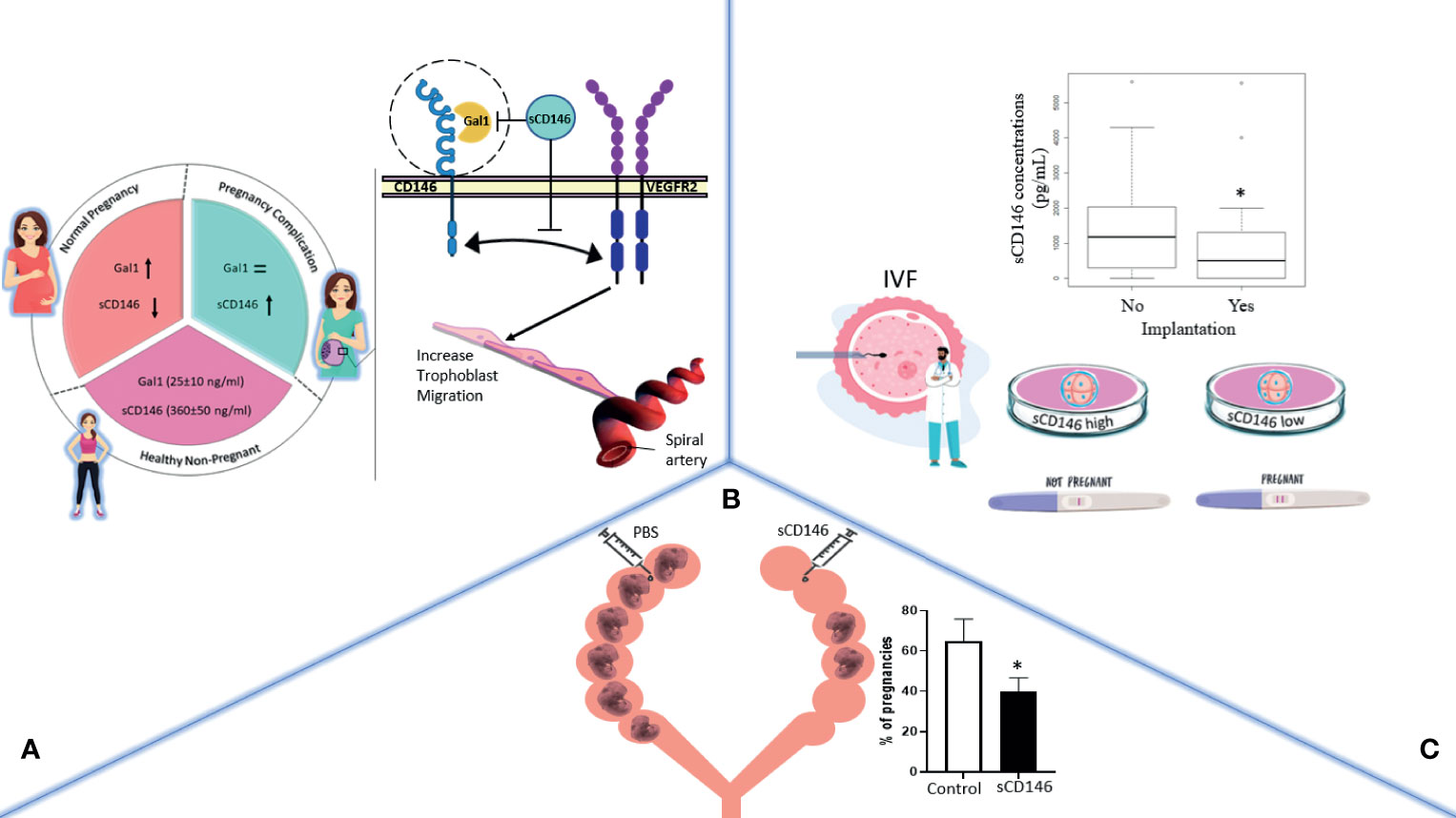
Figure 2 The general effects of CD146/sCD146 in obstetrics are summarized in three illustrations. (A) In pregnancy, sCD146 and galectin 1 (Gal1) plasma concentration varies throughout the different stages of pregnancy. In case of pregnancy complications, sCD146 concentration surges, which in turn hinders Gal1/membrane CD146 interaction, mutually known to activate trophoblasts migration toward the spiral arteries. (B) In implantation, sCD146 injection into female pregnant rats significantly decreases the number of embryos per litter. (C) In IVF, in vitro fertilized eggs producing high concentrations of sCD146 into their culture media fail implantation when transferred into the uterus, a boxplot of sCD146 concentrations between implanted (Yes, n = 37) and non-implanted embryos (No, n = 185) (p = 0.024) is shown. *p < 0.05.
Author Contributions
SB and EK contribute equally to this work and performed the majority of experiments. AJ made experiments and contribute to the redaction of the manuscript. OP-L and MP-M analyzed embryo and obstetrical data. AL and AB analyzed data concerning mechanism. J-CG and FD-G contribute to the reviewing of the manuscript. MB-C contribute to the writing. NB designed the studies and wrote the review. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the research team at the “Direction de la Recherche Clinique et de l’Innovation” at Nîmes University hospital: S. Clément, C. Meyzonnier, N. Best, A. Megzari, E. Dupeyron, S. Granier, B. Lafont, C. Masseguin, O. Albert, S. Doulay, M. Hnida, S.de Bouard, P. Rataboul, and M. P. Francheschi. We are also grateful to Teresa Sawyers, Medical Writer at the B.E.S.P.I.M, Nîmes University Hospital, for revising and editing this article. Our thanks also go to the Department of Gynecology and Obstetrics at Nîmes University Hospital, particularly Prof. Vincent Letouzey and Dr Eve Mousty. We are also most grateful to Nîmes University Hospital for its structural, human, and financial support.
References
1. Huppertz B, Peeters LLH. Vascular Biology in Implantation and Placentation. Angiogenesis (2005) 8(2):157–67. doi: 10.1007/s10456-005-9007-8
2. Fisher SJ, Damsky CH. Humancytotrophoblast Invasion. Semin Cell Biol (1993) 4(3):183–8. doi: 10.1006/scel.1993.1022
3. Knöfler M. Critical Growth Factors and Signalling Pathways Controlling Human Trophoblast Invasion. Int J Dev Biol (2010) 54:269–80. doi: 10.1387/ijdb.082769mk
4. Guezguez B, Vigneron P, Alais S, Jaffredo T, Gavard J, Mège RM, et al. A Dileucine Motif Targets MCAM-L Cell Adhesion Molecule to the Basolateral Membrane in MDCK Cells. FEBS Lett (2006) 580:3649–56. doi: 10.1016/j.febslet.2006.05.048
5. Boneberg EM, Illges H, Legler DF, Fürstenberger G. Soluble CD146 Is Generated by Ectodomain Shedding of Membrane CD146 in a Calcium-Induced, Matrix Metalloprotease-Dependent Process. Microvasc Res (2009) 78(3):325–31. doi: 10.1016/j.mvr.2009.06.012
6. Bardin N, Anfosso F, Massé JM, Cramer E, Sabatier F, Le Bivic A, et al. Identification of CD146 as a Component of the Endothelial Junction Involved in the Control of Cell-Cell Cohesion. Blood (2001) 98:3677–84. doi: 10.1182/blood.v98.13.3677
7. Dagur PK, Biancotto A, Wei L, Sen HN, Yao M, Strober W, et al. MCAM-Expressing CD4(+) T Cells in Peripheral Blood Secrete IL-17A and Are Significantly Elevated in Inflammatory Autoimmune Diseases. J Autoimmun (2001) 37:319–27. doi: 10.1016/j.jaut.2011.09.003
8. Kamiyama T, Watanabe H, Iijima M, Miyazaki A, Iwamoto S. Coexpression of CCR6 and CD146 (MCAM) Is a Marker of Effector Memory T-Helper 17 Cells. J Dermatol (2012) 39:838–42. doi: 10.1111/j.1346-8138.2012.01544
9. Shih IM, Kurman RJ. Expression of Melanoma Cell Adhesion Molecule in Intermediate Trophoblast. Lab Invest (1996) 75(3):377–88.
10. Sers C, Riethmüller G, Johnson JP. MUC18, a Melanoma-Progression Associated Molecule, and Its Potential Role in Tumor Vascularization and Hematogenous Spread. Cancer Res (1994) 54:5689–94.
11. Wu GJ, Wu MWH, Wang C, Liu Y. Enforced Expression of METCAM/MUC18 Increases Tumorigenesis of Human Prostate Cancer LNCaP Cells in Nude Mice. J Urol (2011) 185:1504–12. doi: 10.1016/j.juro.2010.11.052
12. Oka S, Uramoto H, Chikaishi Y, Tanaka F. The Expression of CD146 Predicts a Poor Overall Survival in Patients With Adenocarcinoma of the Lung. Anticancer Res (2012) 32:861–4.
13. Zabouo G, Imbert AM, Jacquemier J, Finetti P, Moreau T, Esterni B, et al. CD146 Expression Is Associated With a Poor Prognosis in Human Breast Tumors and With Enhanced Motility in Breast Cancer Cell Lines. Breast Cancer Res (2009) 11(1):R1. doi: 10.1186/bcr2215
14. Bardin N, Francès V, Combes V, Sampol J, Dignat-George F. CD146: Biosynthesis and Production of a Soluble Form in Human Cultured Endothelial Cells. FEBS Lett (1998) 421(1):12–4. doi: 10.1016/s0014-5793(97)01455-5
15. Joshkon A, Heim X, Dubrou C, Bachelier R, Traboulsi W, Stalin J, et al. Role of CD146 (MCAM) in Physiological and Pathological Angiogenesis-Contribution of New Antibodies for Therapy. Biomedicines (2020) 8(12):633. doi: 10.3390/biomedicines8120633
16. Liu Q, Yan X, Li Y, Zhang Y, Zhao X, Shen Y. Pre-Eclampsia Is Associated With the Failure of Melanoma Cell Adhesion Molecule (MCAM/CD146) Expression by Intermediate Trophoblast. Lab Invest (2004) 84(2):221–8. doi: 10.1038/labinvest.3700033
17. Liu Q, Zhang B, Zhao X, Zhang Y, Liu Y, Yan X. Blockade of Adhesion Molecule CD146 Causes Pregnancy Failure in Mice. J Cell Physiol (2008) 215:621–6. doi: 10.1002/jcp.21341
18. Kaspi E, Guillet B, Piercecchi-Marti MD, Alfaidy N, Bretelle F, Bertaud-Foucault A, et al. Identification of Soluble CD146 as a Regulator of Trophoblast Migration: Potential Role in Placental Vascular Development. Angiogenesis (2013) 16(2):329–42. doi: 10.1007/s10456-012-9317-6
19. Lee Y, Kim KR, McKeon F, Yang A, Boyd TK, Crum CP, et al. A Unifying Concept of Trophoblastic Differentiation and Malignancy Defined by Biomarker Expression. Hum Pathol (2007) 38(7):1003–13. doi: 10.1016/j.humpath.2006.12.012
20. Pollheimer J, Vondra S, Baltayeva J, Beristain AG, Knöfler M. Regulation of Placental Extravillous Trophoblasts by the Maternal Uterine Environment. Front Immunol (2018) 9:2597. doi: 10.3389/fimmu.2018.02597
21. Cierna Z, Varga I, Danihel L Jr, Kuracinova K, Janegova A, Danihel L. Intermediate Trophoblast–A Distinctive, Unique and Often Unrecognized Population of Trophoblastic Cells. Ann Anat (2016) 204:45–50. doi: 10.1016/j.aanat.2015.10.003
22. Shih T, Wang T, Wu R, Kurman J, Gearhart JD. Expression of Mel-CAM in Implantation Site Intermediate Trophoblastic Cell Line, IST-1, Limits Its Migration on Uterine Smooth Muscle Cells. J Cell Sci (1998) 111(Pt 17):2655–64. doi: 10.1242/jcs.111.17.2655
23. Wang H, Du X, Xu Y, Wang Z. Identification of CD146 Expression in Human and Mouse Preimplantation Embryo. J Reprod Contracept (2008) 19(1):7–26. doi: 10.1016/S1001-7844(08)60003-6
24. Bouvier S, Paulmyer-Lacroix O, Molinari N, Bertaud A, Paci M, Leroyer A, et al. Soluble CD146, an Innovative and Non-Invasive Biomarker of Embryo Selection for In Vitro Fertilization. PloS One (2017) 12(3):e0173724. doi: 10.1371/journal.pone.0173724
25. Higuchi T, Fujiwara H, Egawa H, Sato Y, Yoshioka S, Tatsumi K, et al. JEG3 and Human Chorionic Villous Explant Cultures. Mol Hum Reprod (2003) 9(6):359–66. doi: 10.1093/molehr/gag044
26. Dey SK, Lim H, Das SK, Reese J, Paria BC, Daikoku T, et al. Molecular Cues to Implantation. Endocr Rev (2004) 25(3):341–73. doi: 10.1210/er.2003-0020
27. Mills L, Tellez C, Huang S, Baker C, McCarty M, Green L, et al. Fully Human Antibodies to MCAM/MUC18 Inhibit Tumor Growth and Metastasis of Human Melanoma. Cancer Res (2002) 62(17):5106–14.
28. Maynard SE, Min JY, Merchan J, Lim KH, Jianyi Li J, Susanta Mondal S, et al. Excess Placental Soluble Fms-Like Tyrosine Kinase 1 (Sflt1) May Contribute to Endothelial Dysfunction, Hypertension, and Proteinuria in Preeclampsia. J Clin Invest (2003) 111:649–58. doi: 10.1172/JCI17189
29. Kolundžić N, Bojić-Trbojević Ž, Kovačević T, Stefanoska I, Kadoya T, Vićovac L. Galectin-1 Is Part of Human Trophoblast Invasion Machinery–A Functional Study In Vitro. PloS One (2011) 6(12):e28514. doi: 10.1371/journal.pone.0028514
30. Kim YM, Kim SJ, Tatsunami R, Yamamura H, TohruFukai T, Ushio-Fukai M. ROS-Induced ROS Release Orchestrated by Nox4, Nox2, and Mitochondria in VEGF Signaling and Angiogenesis. Am J Physiol Cell Physiol (2017) 312(6):C749–64. doi: 10.1152/ajpcell.00346.2016
31. Onogi A, Naruse K, Sado T, Tsunemi T, Shigetomi H, Noguchi T, et al. Hypoxia Inhibits Invasion of Extravillous Trophoblast Cells Through Reduction of Matrix Metalloproteinase (MMP)-2 Activation in the Early First Trimester of Human Pregnancy. Placenta (2011) 32(9):665–70. doi: 10.1016/j.placenta.2011.06.023
Keywords: biomarker, CD146/sCD146, fertility, implantation, pregnancy, preeclampsia
Citation: Bouvier S, Kaspi E, Joshkon A, Paulmyer-Lacroix O, Piercecchi-Marti M-D, Sharma A, Leroyer AS, Bertaud A, Gris J-C, Dignat-George F, Blot-Chabaud M and Bardin N (2021) The Role of the Adhesion Receptor CD146 and Its Soluble Form in Human Embryo Implantation and Pregnancy. Front. Immunol. 12:711394. doi: 10.3389/fimmu.2021.711394
Received: 18 May 2021; Accepted: 27 July 2021;
Published: 26 August 2021.
Edited by:
Surendra Sharma, Women & Infants Hospital of Rhode Island, United StatesReviewed by:
Nelson Fernandez, University of Essex, United KingdomElba Mónica Vermeulen, Academia Nacional de Medicina, Argentina
Copyright © 2021 Bouvier, Kaspi, Joshkon, Paulmyer-Lacroix, Piercecchi-Marti, Sharma, Leroyer, Bertaud, Gris, Dignat-George, Blot-Chabaud and Bardin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nathalie Bardin, bmF0aGFsaWUuYmFyZGluQHVuaXYtYW11LmZy
†These authors have contributed equally to this work and share first authorship