 Shruti Mishra
Shruti Mishra Saranya Srinivasan
Saranya Srinivasan Chaoyu Ma
Chaoyu Ma Nu Zhang
Nu Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 18 August 2021
Sec. Immunological Tolerance and Regulation
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.708874
Regulatory T cells (Treg) are essential to maintain immune homeostasis and prevent autoimmune disorders. While the function and molecular regulation of Foxp3+CD4+ Tregs are well established, much of CD8+ Treg biology remains to be revealed. Here, we will review the heterogenous subsets of CD8+ T cells have been named “CD8+ Treg” and mainly focus on CD122hiLy49+CD8+ Tregs present in naïve mice. CD122hiLy49+CD8+ Tregs, which depends on transcription factor Helios and homeostatic cytokine IL-15, have been established as a non-redundant regulator of germinal center (GC) reaction. Recently, we have demonstrated that TGF-β (Transforming growth factor-β) and transcription factor Eomes (Eomesodermin) are essential for the function and homeostasis of CD8+ Tregs. In addition, we will discuss several open questions regarding the differentiation, function and true identity of CD8+ Tregs as well as a brief comparison between two regulatory T cell subsets critical to control GC reaction, namely CD4+ TFR (follicular regulatory T cells) and CD8+ Tregs.
Apart from eliminating specific pathogens and the development of memory B and T cells, another remarkable function of acquired immune response is to differentiate between self and non-self-antigens. This is achieved by educating lymphocytes not to respond to self-antigens and removal of autoreactive cells. This process of unresponsiveness is referred to as self-immune tolerance. Immune tolerance mechanisms ensure that lymphocytes that can mediate a response to the host’s own body are eliminated without impacting the immunogenicity of cells to infection and other foreign antigens. However, despite strict vigilance, checkpoints, and selection process some autoreactive lymphocytes manage to escape and enter the periphery. Regulatory T cells (Treg) are essential regulator of immune tolerance and prevent autoimmunity. In contrast to well-established CD4+Foxp3+ Tregs (1), recent advances have started to reveal the biology of CD8+ Tregs.
Various CD8+ T cell subsets carrying certain regulatory properties have been discovered in different experimental systems and have been named “CD8+ Tregs” (Table 1). For example, CD8+Foxp3+ Tregs have been identified in tissue transplantation and alloantigen-induced immune responses (2–7). CD8+CD103+ cells are induced from naïve CD8+ T cells by short-term in vitro stimulation in the presence of TGF-β and exhibit suppressive functions when transferred in vivo (8, 9). Further, CD8+CD103+ cells have been reported in an aggressive tumor model and express Foxp3 (10). However, these cells are often undetectable or extremely rare in unmanipulated mice under steady states. CD8+CD28- T cells, carrying suppressive activities exhibit age-dependent accumulation in human (11–14). Similar to CD8+Foxp3+ cells, CD8+CD28- subset is often associated with chronic antigen exposure and does not represent a major population in naïve young adults. Moreover, a sub-population of CD8+CD122+CD49d+ cells express both PD-1 and IL-10, and suppress alloantigen-induced transplant rejection (15–18). To be noted, CD49d expression has been used to identify T cells with prior TCR stimulation (21). Thus, although all these CD8+ regulatory T cell populations bear great therapeutic potential, they do not represent a major Treg subset in the absence of overt immune challenge. Importantly, even though CD8+CD122+CD49d+ cells share CD122 expression with CD8+CD122hiLy49+ CD8 Treg (see discussion below), there is no evidence to support that CD8+ T cells can differentiate from one subset to the other. In the following sections, we will mainly focus on CD8+CD122hiLy49+ CD8 Treg, present in young naïve mice under homeostatic settings.
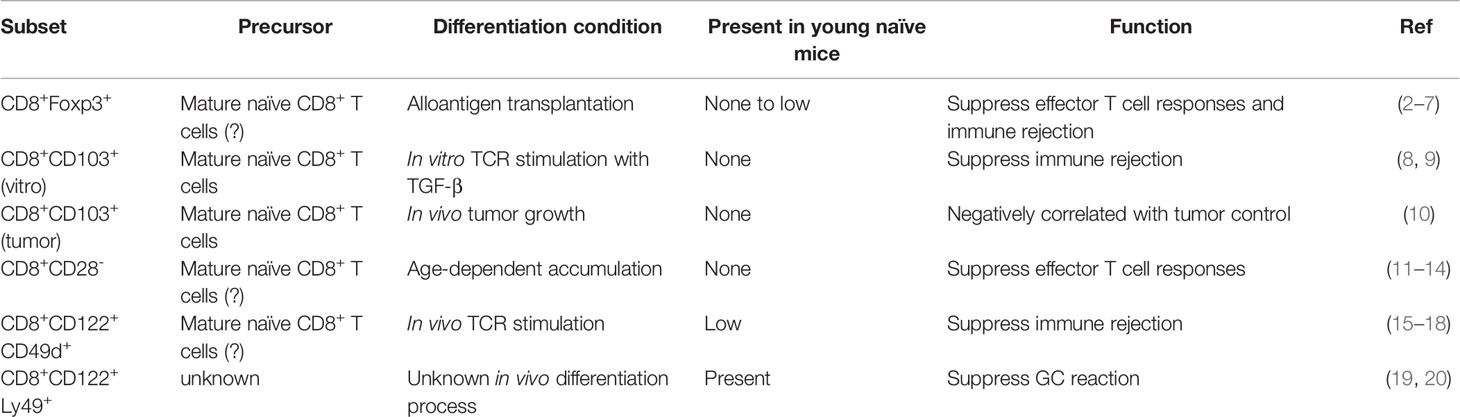
Table 1 Major “CD8+ Treg” subsets.
The discovery of these CD8+ Tregs is intertwined with the history of suppressor T cells (22). Initial work by Gershon and Cantor demonstrated Qa-1 dependent CD4 suppression by CD8 cells in vitro (23–25). This was further confirmed by others studying CD8+ T cells in experimental autoimmune encephalomyelitis (EAE) model (26, 27). Later, a series of investigations of non-classical MHC-I molecule Qa-1-restricted T cells has led to the discovery that the regulatory activity of CD8+ T cells is largely limited to a subset of memory-like CD8+ T cells co-expressing CD122hi and NK marker Ly49 in mice (19, 28–30). Importantly, these CD8+ Tregs do not express Foxp3, but instead depend on transcription factor Helios (encoded by Ikzf2) (31). Ly49 represents a collection of NK surface receptors (including inhibitory receptors Ly49A, Ly49C/I, Ly49F and Ly49G as well as activating receptor Ly49D and Ly49H). At functional level, but not at sequence level, mouse Ly49 gene cluster is homologous to human KIR (killer-cell immunoglobulin-like receptor) genes (32). On CD8+ Tregs, only inhibitory receptors Ly49F, Ly49A, Ly49G and Ly49C/I are detectable (29). All these discoveries together demonstrate that a Qa-1-restricted subset of memory-like CD8+ T cells exhibits potent regulatory activity, most likely via suppressing or eliminating activated CD4+ T cells (33).
Recently, it has been shown that under autoimmunity EAE setting, CD8+CD122hiLy49+ Tregs are not Qa-1-restricted and can be derived from classical MHC-I-restricted CD8+ T cells (34). This work also demonstrates that immune activation induced CD8+ Tregs may be a general player required for immune homeostasis. Importantly, these immune activation-induced CD8+ Tregs bear gene expression signature similar to Qa-1 restricted CD8+ Tregs (34).
In a separated line of research, it has been discovered that CD122hiCD8+ T cells suppress lymphopenia-induced T cell proliferation and autoimmunity in CD122 deficient mice (35). Putting all these elegant works together, it becomes clear that in naïve mice, a subset of both non-classical MHC-I molecule Qa-1-dependent and classical MHC-I-dependent CD8+CD122hiLy49+ T cells express Helios and suppress autoimmunity. Upon immune activation, these CD8+ Tregs further respond to activation-associated epitopes on CD4+ T cells to fine tune ongoing immune responses.
In addition to abovementioned EAE model (19, 36, 37), CD8+CD122hiLy49+ Tregs have documented functions in various settings, including colitis (38, 39), hepatitis (40), arthritis (41), diabetes (42, 43), viral infection (44), tumor immunity (45), atherogenesis (46) and organ transplantation (47). The best demonstrated role of CD8+ Tregs is to suppress GC reaction. Multiple different animal models with defective CD8+ Tregs exhibit enhanced GC reaction and autoantibody production (20, 28, 31, 48), suggesting a non-redundant role for CD8+ Tregs to mitigate GC response.
IL-10 was dispensable for CD8+ Treg regulatory action as anti-IL10 antibody treatment did not alter CD8+ Treg function (28). CD8+ T cells are considered cytotoxic. CD8+ Tregs perform their regulatory function by killing antigen activated CD4+ T cells and TFH cells. The main cytotoxic molecule shown to be important for CD8+ Treg function is perforin. CD8+ Treg from Prf1-/- mice were not able to suppress antigen-activated TFH cells in Rag2-/- mice (28). In another study done in EAE mice model, CD8+ Tregs from Prf1-/- mice were unable to suppress the proliferation of antigen-activated CD4+ T cells further confirming the importance of perforin in CD8+ Treg mediated suppression (34). However, more studies need to be performed to better understand the mechanism of perforin action. For example, our recent findings have demonstrated that WT CD8+ Tregs do not express granzyme A and granzyme B under steady states (20). It will be interesting to examine whether the expression of granzyme A and granzyme B is only induced upon detection of auto-reactive targeting cells or other granzyme(s) are involved in the function of CD8+ Tregs.
In addition, FasL has been demonstrated to be required for CD8+ Tregs to suppress activation and proliferation of both CD4+ and CD8+ T cells (49). Importantly, Fas and FasL signals are essential controllers of CD8+ Treg homeostasis as both lpr and gld mice carry an increased number of CD8+ Tregs. However, whether Fas/FasL directly affects CD8+ Tregs or indirectly impacts CD8+ Tregs via other immune cells remain uninvestigated. To be noted, similar to other effector T cells, TCR engagement is essential for CD8+ Tregs (34, 49). Together, accumulating evidence has demonstrated that upon cognate antigen encounter, CD8+CD122hiLy49+ Tregs target activated CD4+ T cells via a perforin and/or Fas/FasL dependent mechanism to suppress ongoing GC reaction.
Not surprisingly, IL-15 signal is essential for CD8+ Treg homeostasis as they carry high levels of IL-15 receptor CD122 (37). Regulatory T cell associated transcription factor Helios is essential for the function of CD8+ Treg, partially via controlling IL-15/STAT5 pathway (31). Cytoskeleton regulator moesin controls CD8+ Tregs via regulating IL-15 receptor internalization (48). Further, boosting IL-15 signal is sufficient to expand CD8+ Tregs and suppress autoimmune diabetes (42).
We recently discovered a lethal autoimmune phenotype in a T cell-specific conditional knockout mice line lacking both TGF-β receptor and transcription factor Eomes (20). In these double knockout (DKO) mice, greatly enhanced spontaneous GC reaction and autoantibody production are tightly associated with gradual decrease and eventually disappearance of CD8+ Treg population. Importantly, CD4+ Treg compartment is largely intact in DKO mice. In addition, we discover that TGF-β signal and Eomes play critical roles in three seemingly different aspects of CD8+ Treg biology. First, TGF-β signal is essential for the induction of Helios and suppression of effector molecules (e.g., granzyme A, granzyme B and KLRG-1). Second, Eomes controls a molecular program critical for the follicular location and migration of CD8+ Tregs, which includes the induction of CXCR5 and inhibition of integrins α1 and α4. Importantly, these functions of Eomes are not shared with the closely related transcription factor T-bet. Finally, TGF-β and Eomes coordinate to promote the expression of IL-15 receptor and pro-survival molecule Bcl-2. Further, the abovementioned functions of TGF-β and Eomes are CD8+ T cell-specific without impacting CD4+ T cells including CD4+ Tregs. Thus, our findings may establish DKO mice as a valuable CD8+ Treg deficient model with an intact CD4+ Treg compartment (50).
Strikingly, our results also reveal that CD8+ Tregs are extremely potent suppressors of autoimmunity (20). Adoptive transfer of a small number of WT CD8+ Treg into DKO mice is sufficient to fully suppress the spontaneous GC reaction without fully reconstituting the CD8+ Treg compartment. In both TGF-βR single knockout and Eomes single knockout mice, even though CD8+ Treg population is significantly reduced, no overt autoimmunity is detected. Thus, even a small portion of CD8+ Tregs is sufficient to mitigate autoimmunity at steady states, which imply significant translational potential for treating human autoimmune disorders.
As we have discussed that the best-established function of CD8+CD122hiLy49+ Tregs is to suppress GC reaction. In addition to these CD8+ Tregs, a long line of literature has suggested the suppressive function of CD4+Foxp3+TFR (Foxp3+ follicular regulatory T cell) in dampening GC reactions. An obvious question appears–why does GC need two distinct regulatory T cell subsets? In this section, we would like to briefly summarize the recent findings in TFR research and present a side-by-side comparison of CD8+CD122hiLy49+ Tregs with CD4+ TFRs (Table 2).

Table 2 TFR and CD8+CD122+Ly49+ comparison.
To specifically targeting CD4+ TFR without affecting other CD4+Foxp3+ Tregs, several elegant genetic models have been developed. First, a Bcl6f/fFoxp3-Cre mouse line has been demonstrated to lack TFR cells. Only subtle and late onset autoimmunity has been reported for this TFR-deficient mouse line (51). However, upon immunization there is decreased antigen-specific IgG and increased IgA response along with decreased antigen-specific antibody avidity (52). During influenza viral infection, TFR either has no impacts (51, 53) or promotes antigen-specific B cell responses (54) via an IL-10-dependent mechanism (55). Similarly, in a peanut allergy model, TFR cells help maintain GC and antigen-specific IgE response in an IL-10 dependent manner (56).
In a different model, where the expression of CXCR5 is deleted on Foxp3+ cells in an inducible manner, the reduction of TFR leads to no alteration in GC reaction or antibody production (57). All these data together suggest that TFR cells either have very subtle impacts or more of a helper function to promote antigen-specific GC response.
In an inducible TFR-deleter mouse line, it is shown that TFR selectively suppresses early GC response and spares later GC response after protein immunization (58). House dust mite (HDM) induced Th2 response is greatly enhanced in this TFR-deleter mice (58), which is consistent with another report using Bcl6f/fFxop3-cre mice and HDM challenge model (59). A recent publication using Bcl6f/fFxop3-cre mice identifies a key effector molecule neuritin used by TFR to directly inhibit GC B cells, especially under IgE skewing Th2 condition (60). Taken together, considerable discrepancy exists regarding the in vivo function of TFR. In addition to the intrinsic difference between various genetic models (e.g., Bcl6f/fFoxp3-Cre may have defects in other Foxp3+ cells and CXCR5 deletion may not eliminate all TFR-associated activity), the difference between immune challenge models may be critical. For example, while TFR only target early GC reaction (day 5-9) following protein immunization (58), the number of TFR significantly increases only late (>day 30) after influenza viral infection (53, 54). Th2 allergic response provides a distinct immune environment from viral infection. Food allergy is different from HDM allergy (e.g., targeting tissues). Different immune environment may create different niche for TFR, which may correlate with their various demonstrated functions.
In contrast to CD4+TFR, CD8+ Treg is generally considered as immune suppressor. To answer the question why GCs need two subsets of regulatory T cells, both population kinetics of CD4+ TFR vs CD8+ Tregs and various immune challenging conditions need to be taken into consideration. A significant number of CD8+ Tregs (around 0.3x106 cells/spleen) is present before immune challenge in secondary lymphoid organs. We have recently found that CD8+ Treg population is significantly reduced in response to an autoimmune stimulus (20). In contrast, TFR-deficient mice do not exhibit severe autoimmunity in the absence of challenge. Thus, CD8+ Treg may be the critical gate keeper of spontaneous GC reaction at steady states. Depending on the nature of immune stimuli, either CD8+ Treg or CD4+ Treg or both will be required to keep the fine balance between an effective immune response and excess tissue damage/autoimmunity.
In spite of commonly expressed Treg associated transcription factor Helios, unbiased transcriptional analysis suggests that CD8+ Tregs are closely resemble antigen-experienced CD8+ T cells, but distinct from CD4+ Tregs (20). While investigating the lineage relationship between CD122hiLy49+CD8+ Tregs and other CD8+ T cells, two prominent features of CD8+ Tregs are hard to ignore.
First, CD8+ Tregs bear similar markers of senescent T cells. Studies in human have identified age-dependent accumulation of CD28-CD8+ T cells carrying certain regulatory activities (11–14). Even though the relationship between CD28-CD8+ and CD122hiLy49+ CD8+ Tregs is not clear at present, it has been shown that CD27-CD28-CD8+ senescent T cells carries increased levels of NK-associated markers, including Ly49 in mice (61). Further, CD8+ Treg gene signature has been established as a reference of dysfunctional CD8+ T cells during chronic antigen exposure (62). A few more pieces of evidence include age-dependent accumulation of Ly49+CD8+ T cells in mice (61, 63, 64) and age-dependent reduction of regulatory activity of CD122hiCD8+ T cells (35). Together, all the evidence points to an interesting relationship between CD8+ Tregs and age-dependent accumulation of senescent T cells.
Secondly, virtual memory T cells (TVM) represent a subset of memory-like CD8+ T cells differentiated via homeostatic proliferation without previous cognate antigen encounter (65–67). TVM cells bear similar markers as CD8+ Tregs (e.g., CD122hi and CD49dlo). At molecular levels, IL-15 is essential for TVM homeostasis (68). TGF-β signal controls T cell homeostatic proliferation (69, 70), therefore most likely impacts TVM differentiation. Eomes is required for TVM differentiation (71). All of these molecular regulators control both TVM and CD8+ Treg. The only known surface marker to distinguish TVM vs CD8+ Treg is Ly49. Recently, using an Eomes-reporter mouse line, a thymic precursor for TVM has been identified (72). Whether a subset of these Eomes+ precursors will differentiate into CD8+ Tregs remains an open question. It remains to be established what are the differential signals control the differentiation of TVM vs CD8+ Treg.
Significant progress has been made in CD8+ Treg field in recent decades. Both Qa-1-restricted and classical MHC-I-restricted CD8+ Tregs bearing surface CD122 and Ly49 are present in young naïve mice. These CD8+ Tregs play a non-redundant function to mitigate spontaneous GC reaction and various immune responses via suppressing activated CD4+ T cells. Transcription factor Helios is required for the regulatory function of CD8+ Tregs while Eomes is essential for their follicular location. Cytokine IL-15 controls their homeostasis and TGF-β is required to maintain their regulatory identity. In spite of these advances, open questions regarding their true identity and differentiation/development path remain to be addressed in the future. Further, even though both our group and others have provided evidence that a similar human CD8+ Treg subset does exist (20, 43), due to the facts that Ly49 is a mouse-specific gene cluster without human homologs at sequence level, human CD8+ Treg remains to be firmly established. Future investigation into CD8+ Treg will not only provide an opportunity to design future therapeutic interventions for autoimmune disorders, but will also elucidate a mysterious path of CD8+ T cell differentiation.
SM and NZ researched and wrote the manuscript. SS and CM edited the manuscript. All authors contributed to the article and approved the submitted version.
This work is supported by NIH grant AI125701, American Cancer Society grant RGS-18-222-01-LIB, a W.M. Keck Foundation grant and by funds from the UT Health San Antonio to NZ.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Josefowicz SZ, Lu LF, Rudensky AY. Regulatory T Cells: Mechanisms of Differentiation and Function. Annu Rev Immunol (2012) 30:531–64. doi: 10.1146/annurev.immunol.25.022106.141623
2. Agle K, Vincent BG, Piper C, Belle L, Zhou V, Shlomchik W, et al. Bim Regulates the Survival and Suppressive Capability of CD8(+) FOXP3(+) Regulatory T Cells During Murine GVHD. Blood (2018) 132:435–47. doi: 10.1182/blood-2017-09-807156
3. Beres AJ, Haribhai D, Chadwick AC, Gonyo PJ, Williams CB, Drobyski WR. CD8+ Foxp3+ Regulatory T Cells Are Induced During Graft-Versus-Host Disease and Mitigate Disease Severity. J Immunol (2012) 189:464–74. doi: 10.4049/jimmunol.1200886
4. Robb RJ, Lineburg KE, Kuns RD, Wilson YA, Raffelt NC, Olver SD, et al. Identification and Expansion of Highly Suppressive CD8(+)Foxp3(+) Regulatory T Cells After Experimental Allogeneic Bone Marrow Transplantation. Blood (2012) 119:5898–908. doi: 10.1182/blood-2011-12-396119
5. Sawamukai N, Satake A, Schmidt AM, Lamborn IT, Ojha P, Tanaka Y, et al. Cell-Autonomous Role of Tgfbeta and IL-2 Receptors in CD4+ and CD8+ Inducible Regulatory T-Cell Generation During GVHD. Blood (2012) 119:5575–83. doi: 10.1182/blood-2011-07-367987
6. Lerret NM, Houlihan JL, Kheradmand T, Pothoven KL, Zhang ZJ, Luo X. Donor-Specific CD8+ Foxp3+ T Cells Protect Skin Allografts and Facilitate Induction of Conventional CD4+ Foxp3+ Regulatory T Cells. Am J Transplant (2012) 12:2335–47. doi: 10.1111/j.1600-6143.2012.04120.x
7. Churlaud G, Pitoiset F, Jebbawi F, Lorenzon R, Bellier B, Rosenzwajg M, et al. Human and Mouse CD8(+)CD25(+)FOXP3(+) Regulatory T Cells at Steady State and During Interleukin-2 Therapy. Front Immunol (2015) 6:171. doi: 10.3389/fimmu.2015.00171
8. Zhong H, Liu Y, Xu Z, Liang P, Yang H, Zhang X, et al. TGF-Beta-Induced CD8(+)CD103(+) Regulatory T Cells Show Potent Therapeutic Effect on Chronic Graft-Versus-Host Disease Lupus by Suppressing B Cells. Front Immunol (2018) 9:35. doi: 10.3389/fimmu.2018.00035
9. Liu Y, Lan Q, Lu L, Chen M, Xia Z, Ma J, et al. Phenotypic and Functional Characteristic of a Newly Identified CD8+ Foxp3- CD103+ Regulatory T Cells. J Mol Cell Biol (2014) 6:81–92. doi: 10.1093/jmcb/mjt026
10. Gabriely G, da Cunha AP, Rezende RM, Kenyon B, Madi A, Vandeventer T, et al. Targeting Latency-Associated Peptide Promotes Antitumor Immunity. Sci Immunol (2017) 2. doi: 10.1126/sciimmunol.aaj1738
11. Arosa FA, Esgalhado AJ, Padrao CA, Cardoso EM. Divide, Conquer, and Sense: CD8(+)CD28(-) T Cells in Perspective. Front Immunol (2016) 7:665. doi: 10.3389/fimmu.2016.00665
12. Liu Z, Tugulea S, Cortesini R, Suciu-Foca N. Specific Suppression of T Helper Alloreactivity by Allo-MHC Class I-Restricted CD8+CD28- T Cells. Int Immunol (1998) 10:775–83. doi: 10.1093/intimm/10.6.775
13. Strioga M, Pasukoniene V, Characiejus D. CD8+ CD28- and CD8+ CD57+ T Cells and Their Role in Health and Disease. Immunology (2011) 134:17–32. doi: 10.1111/j.1365-2567.2011.03470.x
14. Vuddamalay Y, van Meerwijk JP. CD28(-) and CD28(Low)CD8(+) Regulatory T Cells: Of Mice and Men. Front Immunol (2017) 8:31. doi: 10.3389/fimmu.2017.00031
15. Dai H, Wan N, Zhang S, Moore Y, Wan F, Dai Z. Cutting Edge: Programmed Death-1 Defines CD8+CD122+ T Cells as Regulatory Versus Memory T Cells. J Immunol (2010) 185:803–7. doi: 10.4049/jimmunol.1000661
16. Okuno Y, Murakoshi A, Negita M, Akane K, Kojima S, Suzuki H. CD8+ CD122+ Regulatory T Cells Contain Clonally Expanded Cells With Identical CDR3 Sequences of the T-Cell Receptor Beta-Chain. Immunology (2013) 139:309–17. doi: 10.1111/imm.12067
17. Dai Z, Zhang S, Xie Q, Wu S, Su J, Li S, et al. Natural CD8+CD122+ T Cells are More Potent in Suppression of Allograft Rejection Than CD4+CD25+ Regulatory T Cells. Am J Transplant (2014) 14:39–48. doi: 10.1111/ajt.12515
18. Liu H, Qiu F, Wang Y, Zeng Q, Liu C, Chen Y, et al. CD8+CD122+PD-1+ Tregs Synergize With Costimulatory Blockade of CD40/CD154, But Not B7/CD28, to Prolong Murine Allograft Survival. Front Immunol (2019) 10:306. doi: 10.3389/fimmu.2019.00306
19. Hu D, Ikizawa K, Lu L, Sanchirico ME, Shinohara ML, Cantor H. Analysis of Regulatory CD8 T Cells in Qa-1-Deficient Mice. Nat Immunol (2004) 5:516–23. doi: 10.1038/ni1063
20. Mishra S, Liao W, Liu Y, Yang M, Ma C, Wu H, et al. TGF-Beta and Eomes Control the Homeostasis of CD8+ Regulatory T Cells. J Exp Med (2021) 218. doi: 10.1084/jem.20200030
21. McDermott DS, Varga SM. Quantifying Antigen-Specific CD4 T Cells During a Viral Infection: CD4 T Cell Responses Are Larger Than We Think. J Immunol (2011) 187:5568–76. doi: 10.4049/jimmunol.1102104
22. Germain RN. Special Regulatory T-Cell Review: A Rose by Any Other Name: From Suppressor T Cells to Tregs, Approbation to Unbridled Enthusiasm. Immunology (2008) 123:20–7. doi: 10.1111/j.1365-2567.2007.02779.x
23. Cantor H, Hugenberger J, McVay-Boudreau L, Eardley DD, Kemp J, Shen FW, et al. Immunoregulatory Circuits Among T-Cell Sets. Identification of a Subpopulation of T-Helper Cells That Induces Feedback Inhibition. J Exp Med (1978) 148:871–7. doi: 10.1084/jem.148.4.871
24. Cantor H, McVay-Boudreau L, Hugenberger J, Naidorf K, Shen FW, Gershon RK. Immunoregulatory Circuits Among T-Cell Sets. II. Physiologic Role of Feedback Inhibition In Vivo: Absence in NZB Mice. J Exp Med (1978) 147:1116–25. doi: 10.1084/jem.147.4.1116
25. Eardley DD, Hugenberger J, McVay-Boudreau L, Shen FW, Gershon RK, Cantor H. Immunoregulatory Circuits Among T-Cell Sets. I. T-Helper Cells Induce Other T-Cell Sets to Exert Feedback Inhibition. J Exp Med (1978) 147:1106–15. doi: 10.1084/jem.147.4.1106
26. Jiang H, Zhang SI, Pernis B. Role of CD8+ T Cells in Murine Experimental Allergic Encephalomyelitis. Science (1992) 256:1213–5. doi: 10.1126/science.256.5060.1213
27. Koh DR, Fung-Leung WP, Ho A, Gray D, Acha-Orbea H, Mak TW. Less Mortality But More Relapses in Experimental Allergic Encephalomyelitis in CD8-/- Mice. Science (1992) 256:1210–3. doi: 10.1126/science.256.5060.1210
28. Kim HJ, Verbinnen B, Tang X, Lu L, Cantor H. Inhibition of Follicular T-Helper Cells by CD8(+) Regulatory T Cells is Essential for Self Tolerance. Nature (2010) 467:328–32. doi: 10.1038/nature09370
29. Kim HJ, Wang X, Radfar S, Sproule TJ, Roopenian DC, Cantor H. CD8+ T Regulatory Cells Express the Ly49 Class I MHC Receptor and are Defective in Autoimmune Prone B6-Yaa Mice. Proc Natl Acad Sci USA (2011) 108:2010–5. doi: 10.1073/pnas.1018974108
30. Lu L, Kim HJ, Werneck MB, Cantor H. Regulation of CD8+ Regulatory T Cells: Interruption of the NKG2A-Qa-1 Interaction Allows Robust Suppressive Activity and Resolution of Autoimmune Disease. Proc Natl Acad Sci USA (2008) 105:19420–5. doi: 10.1073/pnas.0810383105
31. Kim HJ, Barnitz RA, Kreslavsky T, Brown FD, Moffett H, Lemieux ME, et al. Stable Inhibitory Activity of Regulatory T Cells Requires the Transcription Factor Helios. Science (2015) 350:334–9. doi: 10.1126/science.aad0616
32. Rahim MM, Tu MM, Mahmoud AB, Wight A, Abou-Samra E, Lima PD, et al. Ly49 Receptors: Innate and Adaptive Immune Paradigms. Front Immunol (2014) 5:145. doi: 10.3389/fimmu.2014.00145
33. Nakagawa H, Wang L, Cantor H, Kim HJ. New Insights Into the Biology of CD8 Regulatory T Cells. Adv Immunol (2018) 140:1–20. doi: 10.1016/bs.ai.2018.09.001
34. Saligrama N, Zhao F, Sikora MJ, Serratelli WS, Fernandes RA, Louis DM, et al. Opposing T Cell Responses in Experimental Autoimmune Encephalomyelitis. Nature (2019) 572:481–7. doi: 10.1038/s41586-019-1467-x
35. Rifa’i M, Kawamoto Y, Nakashima I, Suzuki H. Essential Roles of CD8+CD122+ Regulatory T Cells in the Maintenance of T Cell Homeostasis. J Exp Med (2004) 200:1123–34. doi: 10.1084/jem.20040395
36. Lee YH, Ishida Y, Rifa’i M, Shi Z, Isobe K, Suzuki H. Essential Role of CD8+CD122+ Regulatory T Cells in the Recovery From Experimental Autoimmune Encephalomyelitis. J Immunol (2008) 180:825–32. doi: 10.4049/jimmunol.180.2.825
37. Yu P, Bamford RN, Waldmann TA. IL-15-Dependent CD8+ CD122+ T Cells Ameliorate Experimental Autoimmune Encephalomyelitis by Modulating IL-17 Production by CD4+ T Cells. Eur J Immunol (2014) 44:3330–41. doi: 10.1002/eji.201444675
38. Endharti AT, Okuno Y, Shi Z, Misawa N, Toyokuni S, Ito M, et al. CD8+CD122+ Regulatory T Cells (Tregs) and CD4+ Tregs Cooperatively Prevent and Cure CD4+ Cell-Induced Colitis. J Immunol (2011) 186:41–52. doi: 10.4049/jimmunol.1000800
39. Shimokawa C, Kato T, Takeuchi T, Ohshima N, Furuki T, Ohtsu Y, et al. CD8(+) Regulatory T Cells are Critical in Prevention of Autoimmune-Mediated Diabetes. Nat Commun (2020) 11:1922. doi: 10.1038/s41467-020-15857-x
40. Varthaman A, Khallou-Laschet J, Clement M, Fornasa G, Kim HJ, Gaston AT, et al. Control of T Cell Reactivation by Regulatory Qa-1-Restricted CD8+ T Cells. J Immunol (2010) 184:6585–91. doi: 10.4049/jimmunol.0903109
41. Leavenworth JW, Tang X, Kim HJ, Wang X, Cantor H. Amelioration of Arthritis Through Mobilization of Peptide-Specific CD8+ Regulatory T Cells. J Clin Invest (2013) 123:1382–9. doi: 10.1172/JCI66938
42. Stocks BT, Wilson CS, Marshall AF, Hoopes EM, Moore DJ. Regulation of Diabetogenic Immunity by IL-15-Activated Regulatory CD8 T Cells in Type 1 Diabetes. J Immunol (2019) 203:158–66. doi: 10.4049/jimmunol.1800976
43. Jiang H, Canfield SM, Gallagher MP, Jiang HH, Jiang Y, Zheng Z, et al. HLA-E-Restricted Regulatory CD8(+) T Cells are Involved in Development and Control of Human Autoimmune Type 1 Diabetes. J Clin Invest (2010) 120:3641–50. doi: 10.1172/JCI43522
44. Holderried TA, Lang PA, Kim HJ, Cantor H. Genetic Disruption of CD8+ Treg Activity Enhances the Immune Response to Viral Infection. Proc Natl Acad Sci USA (2013) 110:21089–94. doi: 10.1073/pnas.1320999110
45. Alvarez Arias DA, Kim HJ, Zhou P, Holderried TA, Wang X, Dranoff G, et al. Disruption of CD8+ Treg Activity Results in Expansion of T Follicular Helper Cells and Enhanced Antitumor Immunity. Cancer Immunol Res (2014) 2:207–16. doi: 10.1158/2326-6066.CIR-13-0121
46. Taghavie-Moghadam PL, Waseem TC, Hattler J, Glenn LM, Dobrian AD, Kaplan MH, et al. STAT4 Regulates the CD8(+) Regulatory T Cell/T Follicular Helper Cell Axis and Promotes Atherogenesis in Insulin-Resistant Ldlr(-/-) Mice. J Immunol (2017) 199:3453–65. doi: 10.4049/jimmunol.1601429
47. Choi JY, Eskandari SK, Cai S, Sulkaj I, Assaker JP, Allos H, et al. Regulatory CD8 T Cells That Recognize Qa-1 Expressed by CD4 T-Helper Cells Inhibit Rejection of Heart Allografts. Proc Natl Acad Sci USA (2020) 117:6042–6. doi: 10.1073/pnas.1918950117
48. Satooka H, Nagakubo D, Sato T, Hirata T. The ERM Protein Moesin Regulates CD8(+) Regulatory T Cell Homeostasis and Self-Tolerance. J Immunol (2017) 199:3418–26. doi: 10.4049/jimmunol.1700074
49. Akane K, Kojima S, Mak TW, Shiku H, Suzuki H. CD8+CD122+CD49dlow Regulatory T Cells Maintain T-Cell Homeostasis by Killing Activated T Cells via Fas/Fasl-Mediated Cytotoxicity. Proc Natl Acad Sci USA (2016) 113:2460–5. doi: 10.1073/pnas.1525098113
50. Cantor H, Kim HJ. A New Chapter in the CD8 T Reg Story. J Exp Med (2021) 218. doi: 10.1084/jem.20201746
51. Fu W, Liu X, Lin X, Feng H, Sun L, Li S, et al. Deficiency in T Follicular Regulatory Cells Promotes Autoimmunity. J Exp Med (2018) 215:815–25. doi: 10.1084/jem.20170901
52. Wu H, Chen Y, Liu H, Xu LL, Teuscher P, Wang S, et al. Follicular Regulatory T Cells Repress Cytokine Production by Follicular Helper T Cells and Optimize Igg Responses in Mice. Eur J Immunol (2016) 46:1152–61. doi: 10.1002/eji.201546094
53. Botta D, Fuller MJ, Marquez-Lago TT, Bachus H, Bradley JE, Weinmann AS, et al. Dynamic Regulation of T Follicular Regulatory Cell Responses by Interleukin 2 During Influenza Infection. Nat Immunol (2017) 18:1249–60. doi: 10.1038/ni.3837
54. Lu Y, Jiang R, Freyn AW, Wang J, Strohmeier S, Lederer K, et al. CD4+ Follicular Regulatory T Cells Optimize the Influenza Virus-Specific B Cell Response. J Exp Med (2021) 218. doi: 10.1084/jem.20200547
55. Laidlaw BJ, Lu Y, Amezquita RA, Weinstein JS, Vander Heiden JA, Gupta NT, et al. Interleukin-10 From CD4(+) Follicular Regulatory T Cells Promotes the Germinal Center Response. Sci Immunol (2017) 2. doi: 10.1126/sciimmunol.aan4767
56. Xie MM, Chen Q, Liu H, Yang K, Koh B, Wu H, et al. T Follicular Regulatory Cells and IL-10 Promote Food Antigen-Specific Ige. J Clin Invest (2020) 130:3820–32. doi: 10.1172/JCI132249
57. Vanderleyden I, Fra-Bido SC, Innocentin S, Stebegg M, Okkenhaug H, Evans-Bailey N, et al. Follicular Regulatory T Cells can Access the Germinal Center Independently of CXCR5. Cell Rep (2020) 30:611–9.e4. doi: 10.1016/j.celrep.2019.12.076
58. Clement RL, Daccache J, Mohammed MT, Diallo A, Blazar BR, Kuchroo VK, et al. Follicular Regulatory T Cells Control Humoral and Allergic Immunity by Restraining Early B Cell Responses. Nat Immunol (2019) 20:1360–71. doi: 10.1038/s41590-019-0472-4
59. Koh B, Ulrich BJ, Nelson AS, Panangipalli G, Kharwadkar R, Wu W, et al. Bcl6 and Blimp1 Reciprocally Regulate ST2(+) Treg-Cell Development in the Context of Allergic Airway Inflammation. J Allergy Clin Immunol (2020) 146:1121–36.e9. doi: 10.1016/j.jaci.2020.03.002
60. Gonzalez-Figueroa P, Roco JA, Papa I, Nunez Villacis L, Stanley M, Linterman MA, et al. Follicular Regulatory T Cells Produce Neuritin to Regulate B Cells. Cell (2021) 184(7):1775–89. doi: 10.1016/j.cell.2021.02.027
61. Pereira BI, De Maeyer RPH, Covre LP, Nehar-Belaid D, Lanna A, Ward S, et al. Sestrins Induce Natural Killer Function in Senescent-Like CD8(+) T Cells. Nat Immunol (2020) 21:684–94. doi: 10.1038/s41590-020-0643-3
62. Singer M, Wang C, Cong L, Marjanovic ND, Kowalczyk MS, Zhang H, et al. A Distinct Gene Module for Dysfunction Uncoupled From Activation in Tumor-Infiltrating T Cells. Cell (2016) 166:1500–11.e9. doi: 10.1016/j.cell.2016.08.052
63. Coles MC, McMahon CW, Takizawa H, Raulet DH. Memory CD8 T Lymphocytes Express Inhibitory MHC-Specific Ly49 Receptors. Eur J Immunol (2000) 30:236–44. doi: 10.1002/1521-4141(200001)30:1<236::AID-IMMU236>3.0.CO;2-X
64. Anfossi N, Robbins SH, Ugolini S, Georgel P, Hoebe K, Bouneaud C, et al. Expansion and Function of CD8+ T Cells Expressing Ly49 Inhibitory Receptors Specific for MHC Class I Molecules. J Immunol (2004) 173:3773–82. doi: 10.4049/jimmunol.173.6.3773
65. Haluszczak C, Akue AD, Hamilton SE, Johnson LD, Pujanauski L, Teodorovic L, et al. The Antigen-Specific CD8+ T Cell Repertoire in Unimmunized Mice Includes Memory Phenotype Cells Bearing Markers of Homeostatic Expansion. J Exp Med (2009) 206:435–48. doi: 10.1084/jem.20081829
66. Akue AD, Lee JY, Jameson SC. Derivation and Maintenance of Virtual Memory CD8 T Cells. J Immunol (2012) 188:2516–23. doi: 10.4049/jimmunol.1102213
67. Lee JY, Hamilton SE, Akue AD, Hogquist KA, Jameson SC. Virtual Memory CD8 T Cells Display Unique Functional Properties. Proc Natl Acad Sci USA (2013) 110:13498–503. doi: 10.1073/pnas.1307572110
68. Sosinowski T, White JT, Cross EW, Haluszczak C, Marrack P, Gapin L, et al. CD8alpha+ Dendritic Cell Trans Presentation of IL-15 to Naive CD8+ T Cells Produces Antigen-Inexperienced T Cells in the Periphery With Memory Phenotype and Function. J Immunol (2013) 190:1936–47. doi: 10.4049/jimmunol.1203149
69. Zhang N, Bevan MJ. TGF-Beta Signaling to T Cells Inhibits Autoimmunity During Lymphopenia-Driven Proliferation. Nat Immunol (2012) 13:667–73. doi: 10.1038/ni.2319
70. Johnson LD, Jameson SC. TGF-Beta Sensitivity Restrains CD8+ T Cell Homeostatic Proliferation by Enforcing Sensitivity to IL-7 and IL-15. PloS One (2012) 7:e42268. doi: 10.1371/journal.pone.0042268
71. White JT, Cross EW, Burchill MA, Danhorn T, McCarter MD, Rosen HR, et al. Virtual Memory T Cells Develop and Mediate Bystander Protective Immunity in an IL-15-Dependent Manner. Nat Commun (2016) 7:11291. doi: 10.1038/ncomms11291
Keywords: CD8+ Treg, transforming growth factor-b, eomes, germinal center, virtual memory T cells, senescence
Citation: Mishra S, Srinivasan S, Ma C and Zhang N (2021) CD8+ Regulatory T Cell – A Mystery to Be Revealed. Front. Immunol. 12:708874. doi: 10.3389/fimmu.2021.708874
Received: 12 May 2021; Accepted: 04 August 2021;
Published: 18 August 2021.
Edited by:
Shohei Hori, The University of Tokyo, JapanReviewed by:
Angela Thornton, National Institute of Allergy and Infectious Diseases (NIH), United StatesCopyright © 2021 Mishra, Srinivasan, Ma and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nu Zhang, emhhbmduM0B1dGhzY3NhLmVkdQ==
†Present address:Shruti Mishra, Department of Cancer Immunology and Virology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.