 Zhitao Qi
Zhitao Qi Dong Yan
Dong Yan Lu Cao
Lu Cao Yang Xu
Yang Xu Mingxian Chang
Mingxian Chang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 31 August 2021
Sec. Comparative Immunology
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.707426
Bid (BH3-interacting domain death agonist), a member of the Bcl-2 family, plays a crucial role in the initiation of apoptosis. Independent of its apoptotic function, Bid is also involved in the regulation of inflammation and innate immunity. However, the role of Bid during bacterial pathogen infection remains unclear. In the present study, Bid of zebrafish (Dario rerio) was cloned and its functions during Edwardsiella ictaluri infection were investigated. Zebrafish Bid enhances the apoptosis rate of Epithelioma papulosum cyprini (EPC) cells following E. ictaluri infection. Importantly, in vitro and in vivo bacterial invasion assays showed that overexpressed Bid could significantly inhibit the invasion and proliferation of E. ictaluri. Real-time qPCR analysis revealed that p53 gene expression was downregulated in embryos microinjected with Bid-FLAG. Further, in vitro and in vivo bacterial invasion assays showed that overexpressed p53 increased the invasion and proliferation of E. ictaluri. Moreover, the invasion and proliferation of E. ictaluri were inhibited when co-overexpressing Bid and p53 in vivo and in vitro. Further, the numbers of E. ictaluri in larvae treated with Z-IETD-FMK (caspase-8 inhibitor) were higher than those of larvae without Z-IETD-FMK treatment, while the number of E. ictaluri in larvae microinjected with bid-Flag decreased significantly, even if the larvae were treated in advance with Z-IETD-FMK. Collectively, our study demonstrated a novel antibacterial activity of fish Bid, providing evidence for understanding the function of apoptosis associated gene in pathogen infection.
Apoptosis has crucial roles in biological processes ranging from embryogenesis to aging, and from normal tissue homeostasis to many human diseases (1, 2). The Bcl-2 (B cell lymphoma-2) family is a class of proteins involved in apoptosis. According to their structures and functions, the members of the Bcl-2 family can be divided into three categories, including the multi-domain anti-apoptotic proteins, the multi-domain pro-apoptotic proteins, and the pro-apoptotic BH-3-only proteins (3). The multi-domain anti-apoptotic proteins, including Bcl-2, Bcl-xL, Mcl-1, A1, Bcl-B, and Bcl-w, contain four conserved Bcl-2 homology (BH) domains (BH1-4) and are important for cell survival. The multi-domain pro-apoptotic proteins, Bax, Bak, and Bok, contain BH1-4 domains. The pro-apoptotic BH-3 only proteins include Bid, Bad, Bmf, Bik, Puma, Noxa, Hrk, and Bnip3. By interacting with each other through molecular motifs such as the BH domains, in addition to apoptosis, the Bcl-2 family members are also involved in immune regulation and host immune response to pathogens (4).
Bid (BH3-interacting domain death agonist), a pro-apoptotic protein of the Bcl-2 family, plays a crucial role in the initiation of apoptosis (5). In most cases, Bid is found in the cytoplasm in an inactive state. Under apoptotic stimuli, Bid can be a substrate for a series of proteases, including caspase-8, Granzyme B, calpains, and cathepsins, which generate a truncated Bid (tBid). The tBid then migrates to mitochondria and induces the Bax/Bak-dependent permeabilization of the outer mitochondrial membrane (OM), leading to apoptosis initiation (5, 6). In addition to its role in apoptosis, full-length Bid was found to play crucial roles in inflammation and innate immunity. Full-length Bid could interact with nucleotide-binding and oligomerization (NOD) 1, NOD2, and the IκB kinase (IKK) complex. The cytokine production in colonocytes depleted of Bid or macrophages from Bid-/- mice were markedly defective in response to NOD activation (7).
Teleost fish are important animal models for scientific research (8), and Bid is conserved in fish species (9). However, the understanding on the functions of fish Bid is limited. Zebrafish Bid was found to induce apoptosis in embryos (1). Rare minnow (Gobiocypris rarus) Bid is involved in grass carp reovirus (GCRV)-triggered apoptosis (10). These results indicated that Bid had conserved pro-apoptotic activity in fish. Interestingly, channel catfish Bid was found to be significantly upregulated at 48 h following Flavobacterium columnare infection (11), giving evidence that fish Bid may play some function during bacterial invasion. Further research is needed to elucidate the function and mechanism of fish Bid during bacterial infection.
Edwardsiella ictaluri, a Gram-negative bacteria, is a deadly pathogen in zebrafish (Danio rerio) (12, 13). In the present study, we found that zebrafish Bid could induce apoptosis in piscine cells, similar to previous observations in mammals (1, 14). Furthermore, zebrafish Bid exhibited antibacterial activity, which could significantly inhibit the invasion and proliferation of E. ictaluri in vivo and in vitro. More importantly, we found that the antibacterial activity of zebrafish Bid works in a caspase-8 independent fashion. This is the first report about the antibacterial activity of fish Bid. Our study enriches the knowledge about the function of fish Bid, which could be used as a potential treatment choice for bacterial infection in fish.
Epithelioma papulosum cyprinid (EPC) cells were cultured in an M199 (HyClone, USA) culture medium with 10% fetal bovine serum (FBS) (Lonsa, USA). Wild-type E. ictaluri (PPD130/91 strain) were cultured in tryptic soy broth (TSB, BD Biosciences) at 28°C.
The specific primers (bid-F1: CCGGAATTCATGGACTTCAACAGGAAC and bid-R1: were designed according to the zebrafish Bid EST sequence (GenBank accession No: XM_005159109.4) and used for RT-PCR to clone the full open reading frame (ORF) of zebrafish Bid with the cDNA of the wild type of zebrafish as templates. The PCR products and p3xFLAG-CMV™-14 were digested using EcoR I and Xba I restriction enzymes and ligated using T4 DNA ligase (TaKaRa, USA). All plasmids were confirmed by sequencing.
To investigate the expression changes of zebrafish Bid during early development, wild-type zebrafish embryos or larvae were collected at 6 h, 12 h post-fertilization (hpf), 1 day (d), 3 days, 5 days, and 10 days post-fertilization (dpf). For each time point, 30–50 embryos or larvae were collected and used for RNA extraction.
To study the inducible expression pattern of Bid, zebrafish larvae at 4 dpf were infected with 2 × 108 CFU/ml E. ictaluri in a total volume of 5 ml. After immersion in the bacterial suspension for 6 h, zebrafish larvae were maintained in 60-mm sterile disposable Petri dishes with 25 ml water. Larvae were collected at 12, 24, and 48 h post-infection (hpi). All samples were snap-frozen in liquid nitrogen and stored at −80°C.
To determine the role of Bid in apoptosis, EPC cells seeded overnight in six-well plates at 1 × 106 cells per well were transiently transfected using Lipofectamine 2000 (Invitrogen, USA) with 2 μg p3xFLAG or Bid-FLAG plasmid. At 24 h post-transfection, the cells were infected with various concentrations of E. ictaluri with an MOI of 1°C, 5°C, and 10 at 28°C. After infection for 1 h, the medium was removed and replaced with MEM medium containing gentamicin (50 µg/ml) to kill the extracellular bacteria. At 6 hpi, the culture medium was discarded and trypsin without EDTA was added for 2 min. The cell suspension was collected, centrifuged at 4°C and 500 g, and washed twice with precooled PBS solution. Apoptosis was detected using an Annexin V-FITC/PI Apoptosis Detection Kit (Vazyme, China) on a CytoFLEX LX flow cytometer (Beckman, USA).
For in vitro bacterial invasion assays, EPC cells seeded overnight in 24-well plates at 3 × 106 cells per well were transiently transfected using Lipofectamine 2000 (Invitrogen, USA) with 500 ng p3×FLAG or Bid-FLAG plasmid. Twenty-four hours post-transfection, the cells were infected with E. ictaluri at MOI 10. After infection for 1.5 h, the medium was removed and replaced with MEM medium containing gentamicin (50 µg/ml) to kill the extracellular bacteria. At 3 and 6 hpi, the cells were washed and lysed in PBS containing 1% (v/v) Triton X-100 at room temperature for 20 min. The lysates were diluted with TBS and then plated onto TSA to calculate the intracellular bacterial colony-forming units (CFU). The invasion assays were performed in triplicate.
For in vivo bacterial invasion assays, the p3×FLAG empty plasmid or Bid-FLAG plasmid was diluted to the desired concentration of 200 ng/μl and microinjected into fertilized eggs at the one-cell or two-cell stage with a injected volume of 2 nl. The injected embryos were raised to 4 dpf at 28°C and then were divided into two groups (three replicates per group, with 10 larvae per replicate) used for E. ictaluri infection. Briefly, the larvae were infected with E. ictaluri at an MOI of 10 for 6 h, then 20 ml aquaculture water was added. At 48 hpi, 10 larvae per group were collected, rinsed with 1 ml PBS, and ground via a glass homogenizer to obtain homogenates. Serial dilutions of the homogenates were plated onto TSB agar, and CFU were enumerated after 24 h of incubation at 28°C.
The p3×FLAG empty plasmid or Bid-FLAG plasmid was diluted to the desired concentration of 200 ng/μl and microinjected into fertilized eggs at the one-cell or two-cell stage with an injected volume of 2 nl. The injected embryos were raised to 4 dpf at 28°C, and the apoptosis-related genes, including birc5, epidermal growth factor receptor (egfr), caspase 8, caspase 3a, caspase 3b, caspase 9, programmed cell death protein 8 (PDCD8), bax, and bid, were examined using quantitative real-time PCR (qPCR). The expression of these genes at 12 h post-E. ictaluri infection was also examined using qPCR. The bacterial infection was completed as mentioned above. All primers used for gene expressions were listed in Table S1.
To further analyze the roles of Bid and p53 for fish survival and antibacterial activity, the fertilized eggs at the one-cell or two-cell stage were randomly divided into three groups (three replicates per group, with 30 eggs per replicate): the control group, which was microinjected with p3×FLAG empty plasmid; the p53 group, which was injected with p53-FLAG; and the p53-Bid group, which was co-injected with Bid-FLAG and p53-FLAG. The injected embryos were raised to 4 dpf at 28°C and then were infected with E. ictaluri at an MOI of 10 for 6 days. The numbers of surviving larvae were counted daily, and the survival curves were generated by GraphPad Prism 7. The numbers of E. ictaluri in larvae were examined at 24 hpi as the in vivo bacterial invasion assays.
The fertilized eggs at the one-cell or two-cell stage were pretreated with 50 μM Z-IETD-FMK (caspase-8 inhibitor) (15) for 8 h, then infected with 2 × 108 CFU/ml E. ictaluri. The survival rates of larvae were counted for 6 days. The larvae were also collected at 6, 24, and 48 hpi, and the number of E. ictaluri in larvae was examined as an in vivo bacterial invasion assay. In addition, the 4-dpf larvae, developed from the one-cell or two-cell stage fertilized eggs that were microinjected with p3×FLAG empty plasmid or Bid-FLAG plasmid, were treated with 50 μM Z-IETD-FMK for 8 h and then infected with 2 × 108 CFU/ml E. ictaluri. The numbers of E. ictaluri in larvae at 24 and 48 hpi were calculated as mentioned above.
Total RNA was extracted from 30–50 zebrafish embryos or larvae using TRIzol reagent (Invitrogen, USA). The cDNA was reverse transcribed using a RevertAid™ First Strand cDNA Synthesis Kit (Thermo Fisher, USA). The qPCR was performed on a Thermo Fisher Scientific QuantStudio™ 3 Real-Time PCR Instrument (96-well 0.2 ml Block) under the following conditions: 3 min at 95°C, followed by 50 cycles of 15 s at 94°C, 15 s at 52°C–60°C, and 30 s at 72°C. All reactions were performed in triplicate in a 96-well plate, and the mean values were recorded. The specificity of primers was confirmed by sequencing and melting curves. The relative expression of target genes was normalized to the expression of housekeeping genes gapdh and EF-1α and expressed as fold changes relative to the corresponding control group. The primers used for qRT-PCR are listed in Table S1.
Statistical analysis and graphs were performed and produced using GraphPad Prism 7.0 software. Expression data of qPCR are presented as means and standard error of the mean (SEM). Two-tailed Student’s t-test or ANOVA was used to compare means and SEM between groups. All data are representative of two or three biologic replications. The level of significance is shown as follows: *p < 0.05, **p < 0.01. Significance testing in the cumulative survival analysis used the log-rank test.
The ORF length of zebrafish bid is 564 bp (GenBank accession No. XM_005159109), encoding 187 amino acids. Zebrafish Bid contains a conserved BH3 domain and several conserved cleavage sites (9), including the caspase-8 cleavage site (LETD), the granzyme B cleavage site (HELQ), and the calpain cleave site (Figure 1). Zebrafish Bid shares 70.4%, 66.5%, 34.5%, 16.3%, and 19.7% overall sequence identities with the Bid of common carp, grass carp, rainbow trout, mouse, and human, respectively (Figure 1). To further understand the evolutionary history of vertebrates’ Bid, a phylogenetic tree was constructed using Mega X software with the neighbor-joining (N-J) method (16). The tree is divided into four main clades (clades A to D), supported by high bootstrap values (≥ 99%) (Figure 2). Clade A is composed of Bid of fish. In clade A, zebrafish Bid clustered with the Bid of Cyprinidae. Clade B is composed of Bid from amphibians, and clade C is composed of Bid from birds. Clade D includes the Bid of mammals.

Figure 1 Multiple-sequence alignment of vertebrate Bid. The Bid from each species were aligned using the Clustal O program and decorated with BoxShade software. The conserved BH3 domains were boxed. The caspase-8 cleavage site (LETD), the granzyme B cleavage site (HELQ), and the calpain cleavage site was marked by red arrow, blue arrow, and green arrow, respectively.
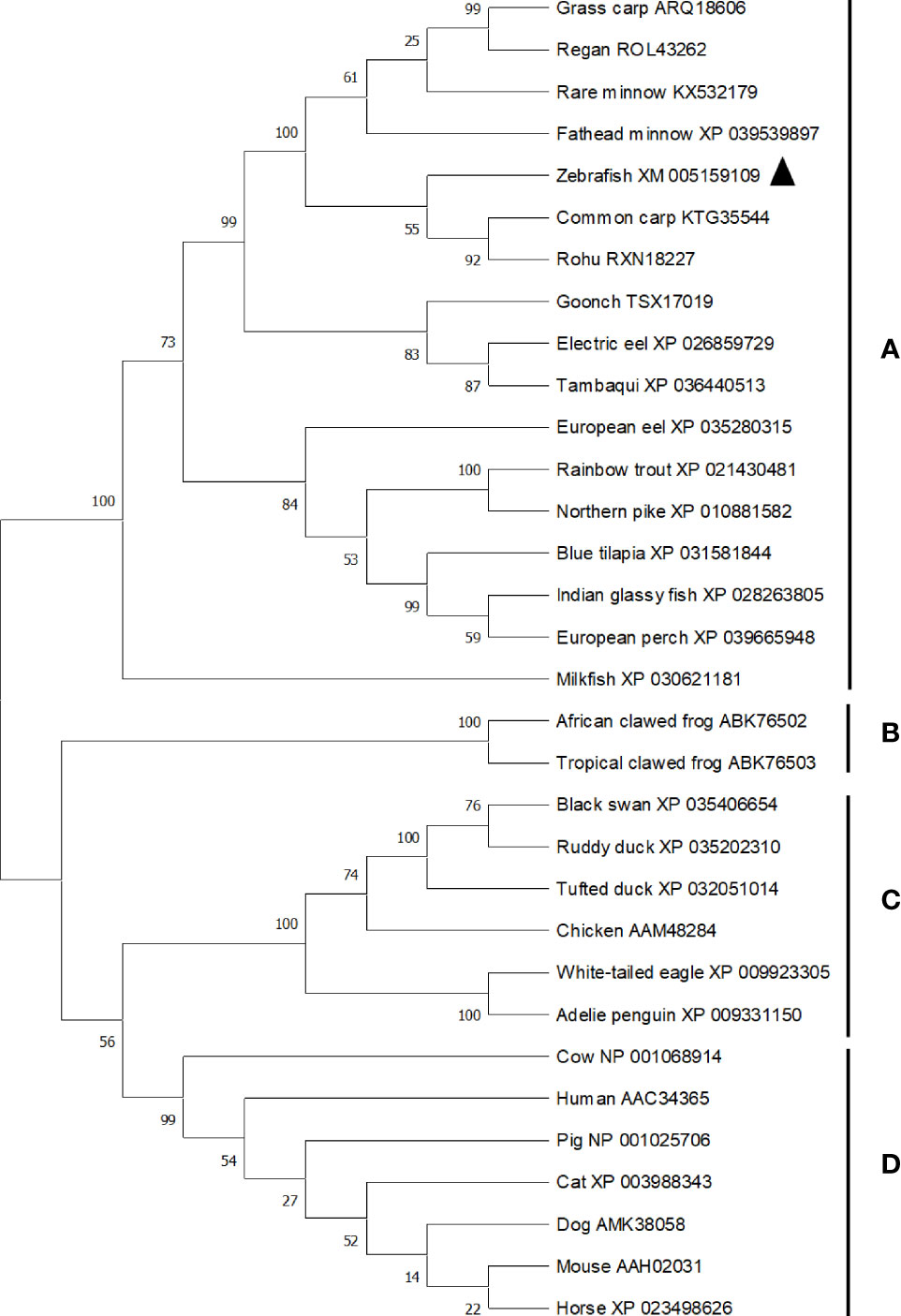
Figure 2 Phylogenetic tree analysis of vertebrates Bid. The phylogenetic tree was constructed using Mega X software using N-J method. Bootstrap was calculated with 10,000 repeats. The tree is divided into four main clades (clade A–D), with clade (A) composed of fish Bid, clade (B) composed of amphibian Bid, clade (C) composed of birds Bid and clade (D) composed of mammalian Bid. Accession numbers for sequences were listed following the common names of species. Zebrafish Bid was marked by triangle in the tree.
First, we analyzed the expression of zebrafish bid by qPCR. Results showed that zebrafish bid was expressed ubiquitously across all early developmental stages examined, showing a tendency first to decrease and then increase slowly (Figure 3A). We also assessed the expression changes of zebrafish bid in larvae infected with E. ictaluri. Results showed that zebrafish Bid was significantly upregulated at 48 hpi following E. ictaluri infection (Figure 3B).
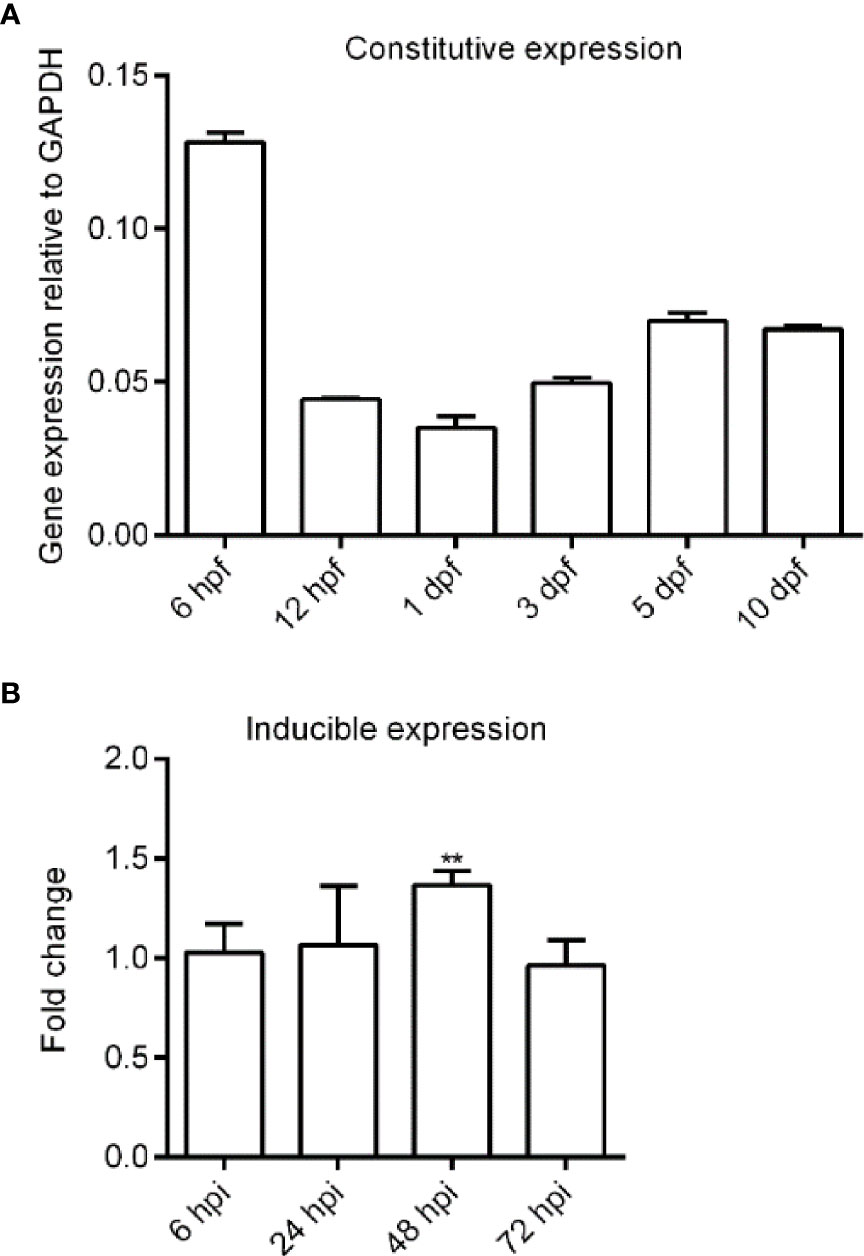
Figure 3 Expression analysis of zebrafish bid in normal embryos and embryos infected with E. ictaluri by using qPCR. (A) Expression of bid in normal embryos. Thirty to 50 embryos were collected for each time point. Expression of bid was normalized to the housekeeping gene gapdh. (B) Expression of bid in embryos infected with E. ictaluri. The zebrafish larvae at 4 dpf were infected with 2 × 108 CFU/ml E. ictaluri and were collected at 12, 24, and 48 h postinfection (hpi). The fold changes of bid in embryos infected with E. ictaluri at each time point were analyzed using the 2-ΔΔCT method relative to the corresponding control group. Asterisks indicated that the expression level of bid in embryos infected with E. ictaluri was significantly higher than that of the control. **p < 0.01.
To test whether Bid can induce apoptosis, we first transfected the EPC cells with Bid-FLAG or p3×FLAG empty plasmid. Then, the cells were infected with E. ictaluri, and the apoptosis rates were examined. At the early apoptosis stage, the apoptosis rates for cells transfected with Bid-FLAG were significantly higher than that transfected with empty plasmid when the MOIs of E. ictaluri were 5 and 10 (p < 0.05) (Figure 4A). At the late apoptosis stage, higher apoptosis rates for cells transfected with Bid-FLAG were also observed when the MOI of E. ictaluri was 10, compared with those for cells transfected with empty plasmid (Figure 4B).
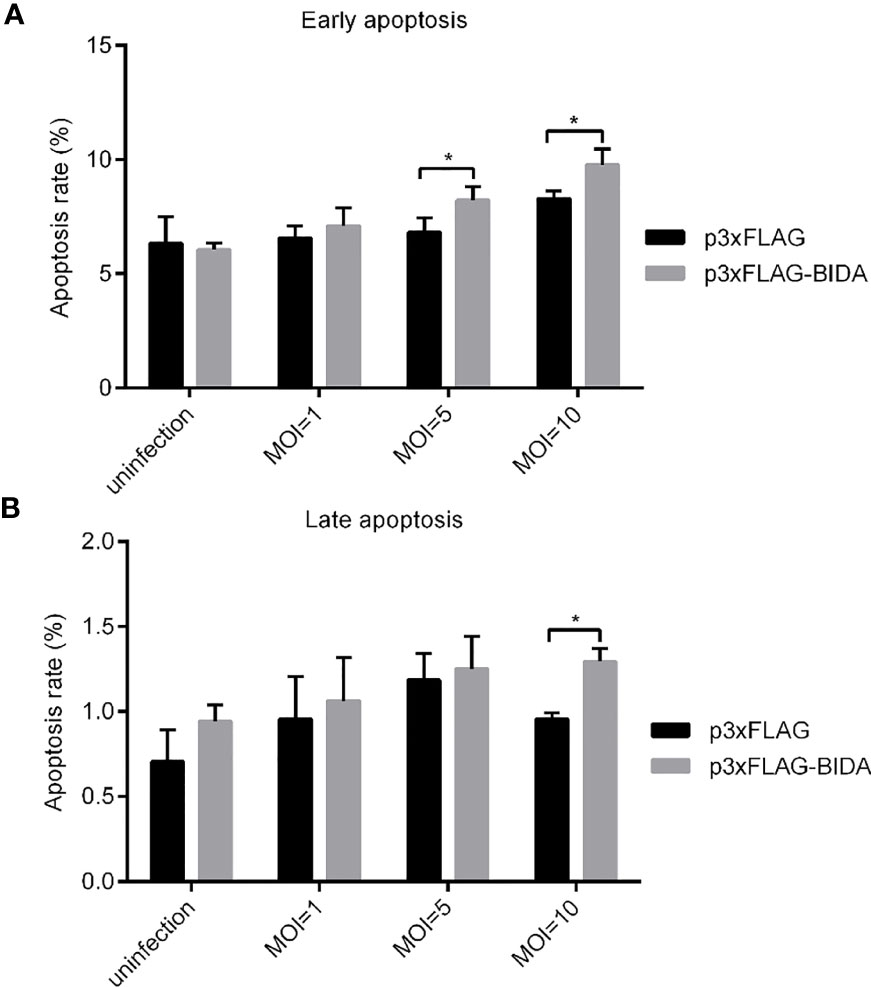
Figure 4 Apoptotic activity of zebrafish Bid at the early apoptosis stage (A) and the later apoptosis stage (B). EPC cells were transiently transfected with a 2 μg p3xFLAG or Bid-FLAG plasmid. At 24 hours post-transfection, the cells were infected with E. ictaluri with an MOI of 1, 5, and 10 at 28°C. After infection for 1 h, the extracellular bacteria were killed using gentamicin (50 µg/mL). Apoptosis at 6 hpi was detected using an Annexin V-FITC/PI Apoptosis Detection Kit on the CytoFLEX LX flow cytometer. *p< 0.05.
The antibacterial activities of zebrafish Bid were assessed using bacterial invasion assays in vitro and in vivo. Results showed that overexpression of zebrafish Bid could significantly inhibit the proliferation of E. ictaluri in EPC cells at 3 and 6 hpi (Figure 5A). In vivo bacterial invasion assays showed that the numbers of E. ictaluri in fish microinjected with Bid-FLAG were significantly lower than those in fish microinjected with empty plasmid at 48 hpi (Figure 5B).
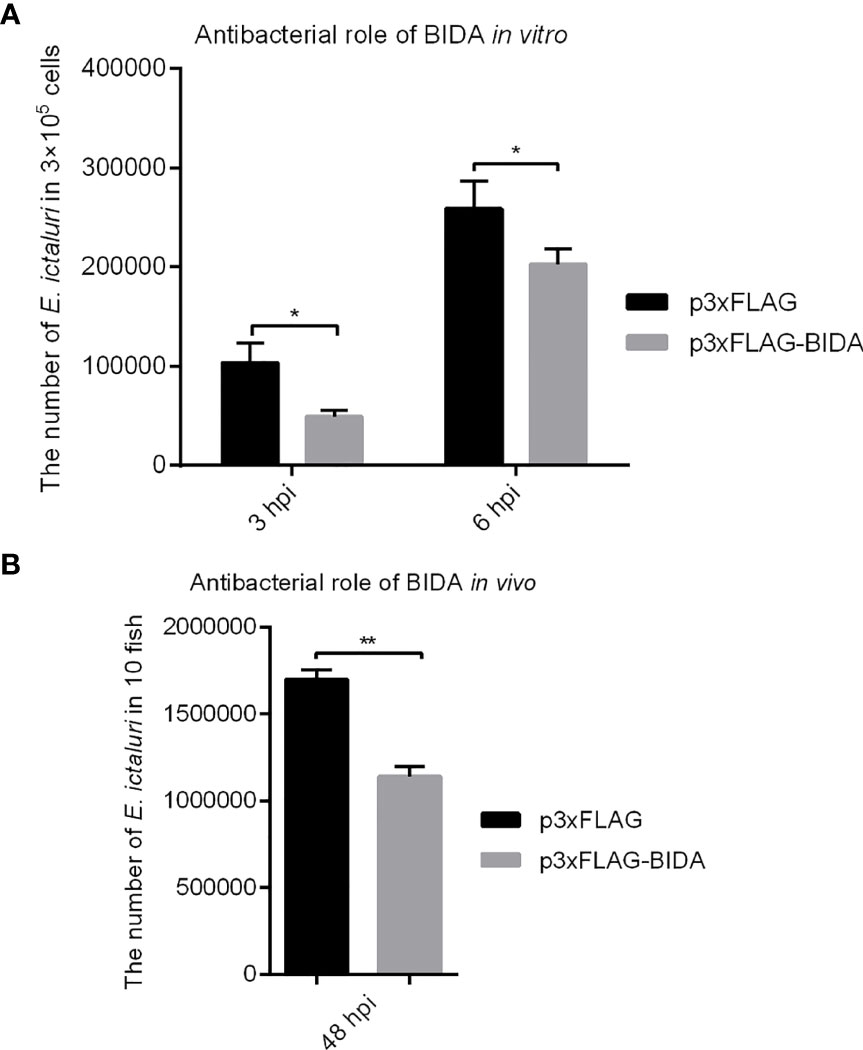
Figure 5 Antibacterial activity of zebrafish Bid in vitro (A) and in vivo (B). For in vitro bacterial invasion assays (A), the EPC cells were transiently transfected with 500 ng p3×FLAG or Bid-FLAG plasmid. At 24 h post-transfection, the cells were infected with E. ictaluri at an MOI of 10. At 1.5 h post-infection, the extracellular bacteria were killed using gentamicin (50 µg/ml). At 3 and 6 hpi, the cells were lysed, and the intracellular bacterial colony-forming units (CFU) were calculated. For in vivo bacterial invasion assays (B), the fertilized eggs were microinjected with 200 ng/μl p3×FLAG or Bid-FLAG. At 4 dpf, the larvae were infected with E. ictaluri at an MOI of 10. At 48 hpi, the fish were ground, and the CFU was calculated. *p < 0.05, **p < 0.01.
To further understand the antibacterial mechanism of zebrafish Bid, we first examined the expression of bicr5, egfr, p53, caspase8, caspase3a, caspase3b, caspase9, pdcd8, and bax in larvae microinjected with bid-FLAG. Results showed that without E. ictaluri infection, the expression of p53 was significantly inhibited in larvae microinjected with Bid. No significant changes were observed for other examined genes (Figures 6A–C). The larvae microinjected with Bid-FLAG were then infected with E. ictaluri, and the expressions of the above genes were examined at 12 hpi. Results showed that the expression of p53 was significantly downregulated and that birc5 and egfr were significantly upregulated (Figures 6D–F), when compared with those in larvae microinjected with the p3×FLAG empty plasmid.
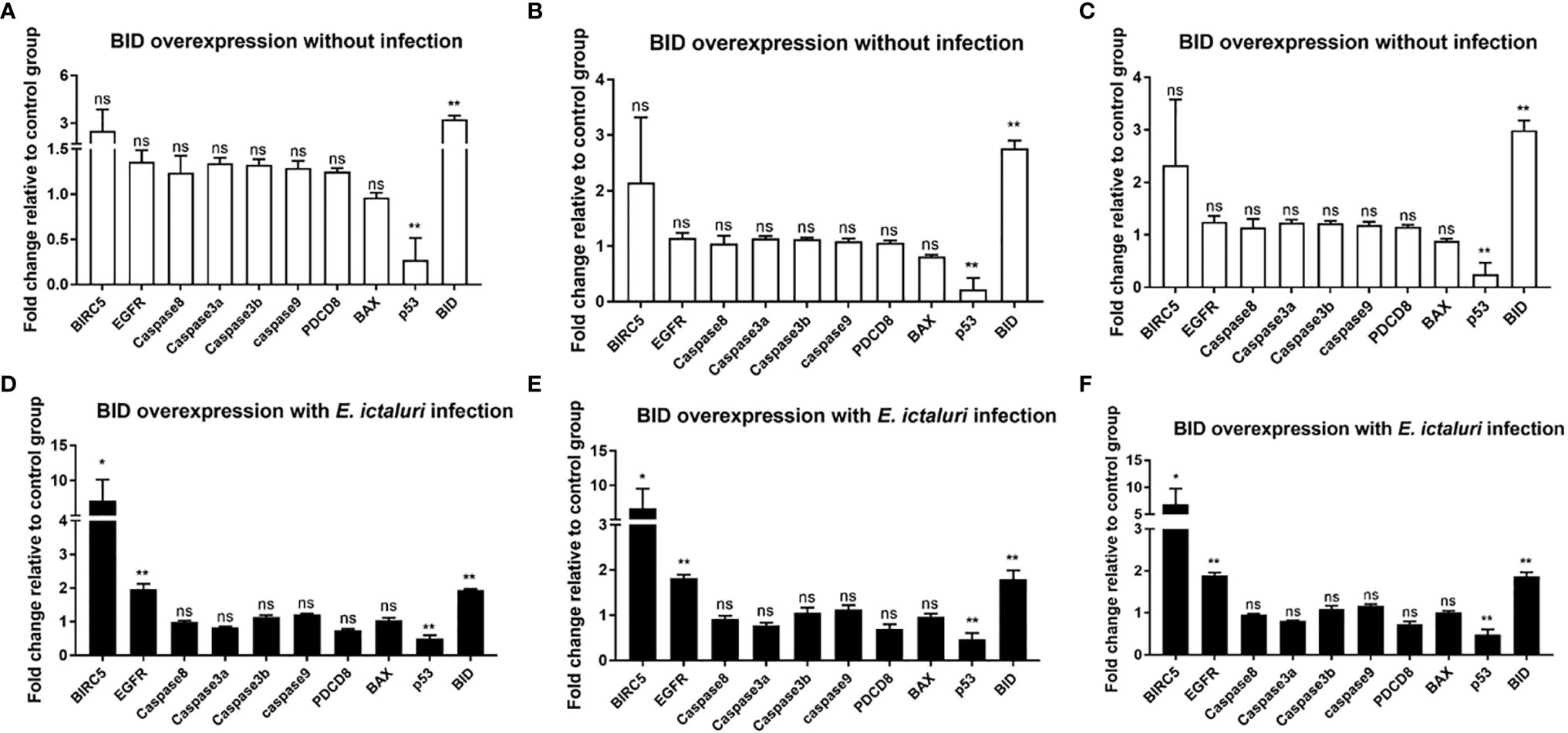
Figure 6 Expression analysis of apoptosis-related genes in bid-overexpressed embryos without (A–C) or with (D–F) E. ictaluri infection. The fertilized eggs were microinjected with 200 ng/μl p3×FLAG empty plasmid or Bid-FLAG plasmid. The microinjected eggs were raised to 4 dpf, and the expressions of apoptosis-related genes were examined using qPCR. The 4-dpf microinjected larvae were infected with E. ictaluri, and the expressions of apoptosis-related genes were examined using qPCR at 12 hpi. The expression levels of target genes were analyzed using the 2-ΔΔCT method by respectively calculating with the Ct values of gapdh (A, D), EF-1α (B, E), and the average Ct of gapdh and EF-1α (C, F). The average Ct values of the two reference genes are listed in Table S2. *p < 0.05, **p < 0.01. ns, no significance.
The finding that overexpression of Bid could significantly inhibit the transcript of p53 with or without E. ictaluri infection (Figure 6) indicated that Bid exerts its antibacterial activity by regulating p53. Thus, we first analyzed the effects of p53 on the survival rates of fish infected with E. ictaluri. The survival rate of larvae microinjected with p53-Flag was significantly lower than that of fish microinjected with the p3×FLAG empty plasmid (p = 0.0002) (Figure 7A). We also found that the survival rates of larvae co-microinjected with Bid-FLAG and p53-FLAG were significantly increased (p < 0.01) compared with those of larvae microinjected with p53-Flag (Figure 7A). Furthermore, the in vivo bacterial invasion assays showed that the number of bacteria in larvae transfected with p53 was significantly higher than that of cells transfected with p3×FLAG empty plasmid (p < 0.01) and cells co-transfected with Bid-Flag and p53-FLAG (p < 0.01) (Figure 7B). Further, the number of bacteria in cells co-transfected with Bid-FLAG and p53-FLAG was significantly lower than that of cells transfected with p3×FLAG empty plasmid (p < 0.01) (Figure 7B).
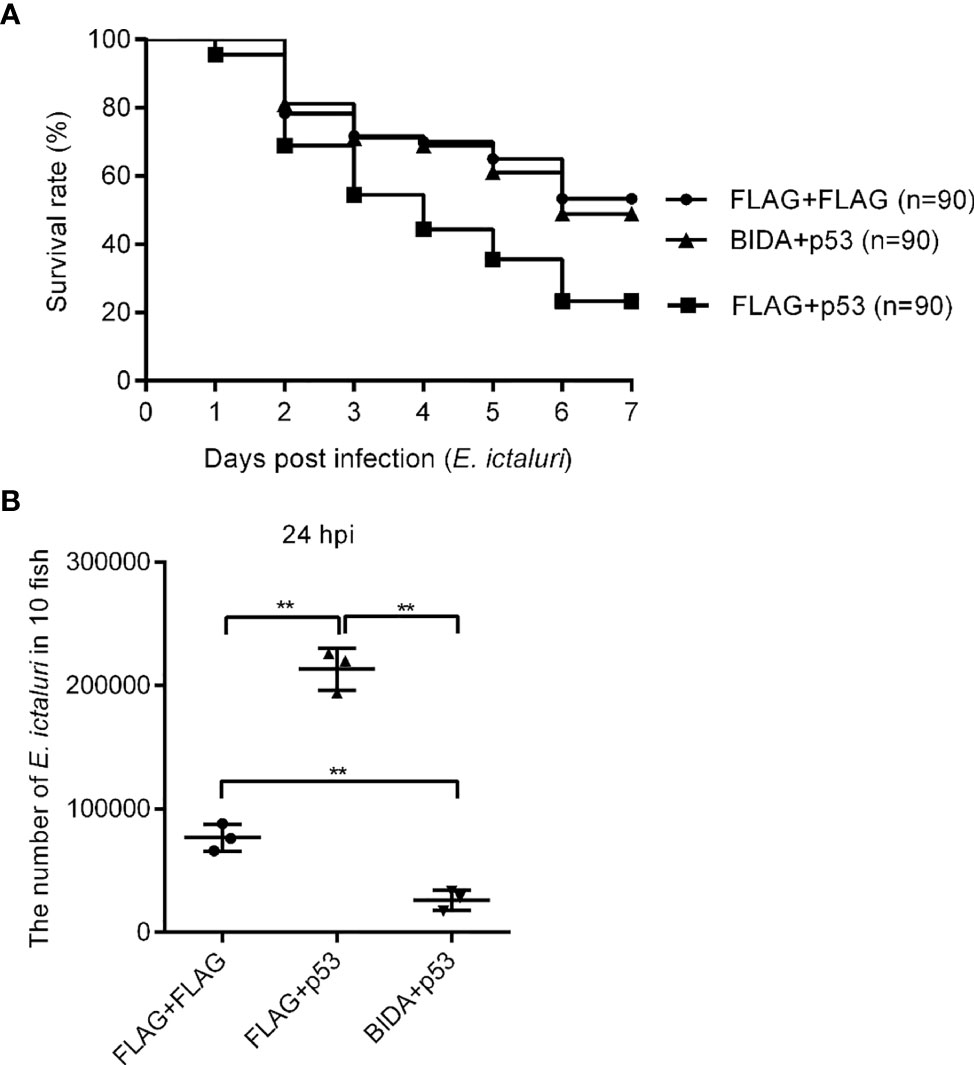
Figure 7 The effects of Bid and p53 on fish survival (A) and the number of E. ictaluri in larvae (B). For fish survival analysis, the fertilized eggs were microinjected with p3×FLAG empty plasmid, p53-Flag, or co-microinjected p53-Flag and bid-Flag and raised to 4 dpf, and then infected with E. ictaluri for 6 days. The numbers of surviving larvae were counted daily, and survival curves were generated. The number of E. ictaluri in larvae was calculated as the in vivo bacterial invasion assays. **p < 0.01.
Z-IETD-FMK is the inhibitor of the caspase-8 inhibitor (15). We found that the larval survival at 6 dpi was decreased significantly when the larvae were pretreated with Z-IETD-FMK (Figure 8A). The in vivo bacterial invasion assays revealed that the numbers of E. ictaluri in larvae treated with Z-IETD-FMK at 24 and 48 hpi were higher than those of larvae without Z-IETD-FMK treatment (Figure 8B). Furthermore, we found that the number of E. ictaluri in larvae microinjected with bid-Flag decreased significantly, even if the larvae were treated in advance with Z-IETD-FMK (Figure 8C).
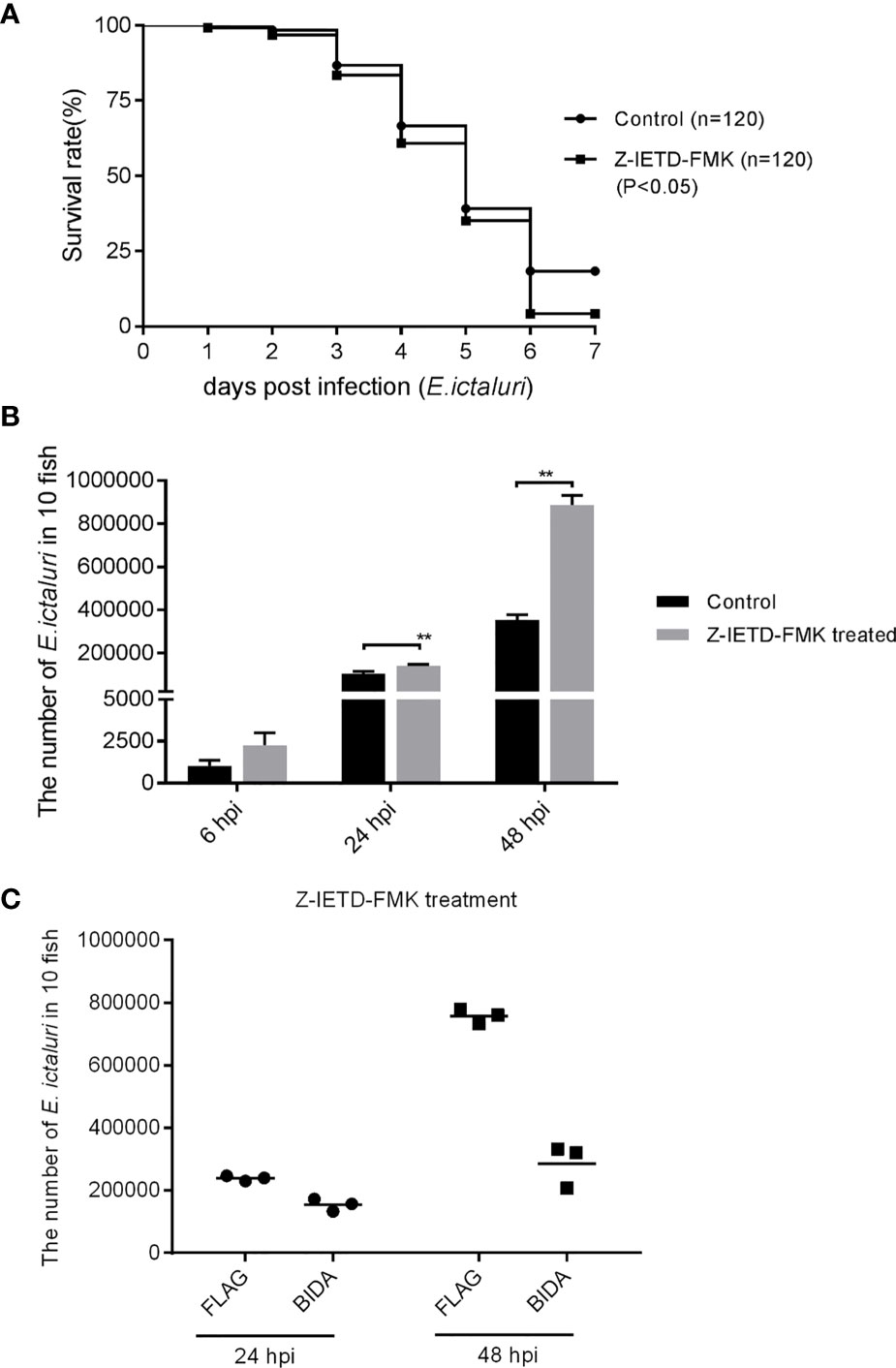
Figure 8 Antibacterial activity of Bid is caspase-8 independent. (A) Survival rates for larvae treated with Z-IETD-FMK. (B) Number of E. ictaluri in larvae treated with Z-IETD-FMK. (C) Number of E. ictaluri in larvae microinjected with Bid-Flag and treated with Z-IETD-FMK. **p < 0.01.
Bid, a member of the Bcl-2 family, plays a crucial role in mammalian apoptotic signaling (17). However, studies about its function in fish are scarce. In the present study, the bid of zebrafish was cloned, and its antibacterial function was investigated. Our results demonstrated that in addition to its pro-apoptotic function, zebrafish Bid has a strong antibacterial role against E. ictaluri, an important fish pathogen affecting cultured and wild fish worldwide (12, 18).
Zebrafish Bid encodes 187 amino acids, which share a high sequence identity with Bid of other fish species, ranging from 34.5% to 70.4% (Figure 1). Zebrafish Bid contains a conserved BH3 region that is required for Bid to bind to Bcl-2-like pro-survival proteins and initiate apoptosis (17). Phylogenetic tree analysis revealed that zebrafish Bid was well clustered with fish Bid (Figure 2), confirming that zebrafish Bid is a member of the Bid family. Phylogenetic tree analysis also showed that Bid forms from the different animal classes were separated, suggesting that Bid underwent divergent evolutionary processes.
Further analysis revealed some differences at the potential cleavage sites between zebrafish Bid and mammalian Bid (Figure 1). The caspase-8 cleavage site for mammalian Bid is “LQTD” (19), while the same site in zebrafish Bid is “IETD.” Also, mammalian Bid could be cleaved by granzyme B, which plays an important role in the target cell killing by cytotoxic T cells (20). The granzyme B cleavage site in mammalian Bid is “IEPD,” while that in zebrafish is “HELQ.” Mammalian Bid can also be cleaved by calpain, and the cleavage site is between Gly70 and Arg71 (21); a similar site in zebrafish is Leu67–Leu68. The poor conservation of these cleavage sites indicates that these enzymes might not be critical for the functions of zebrafish Bid.
E. ictaluri causes Edwardsiellosis in zebrafish (12). We found that following E. ictaluri infection, zebrafish bid was upregulated. Furthermore, zebrafish Bid could enhance the apoptosis rate of EPC cells following E. ictaluri infection (Figure 3). Our results are in line with the literature (1). A bid of Chinese giant salamander played a positive role in Chinese giant salamander iridovirus (GSIV)-induced apoptosis (22). Bid-deficient rare minnow delayed the grass carp reovirus (GCRV) replication and attenuated GCRV-induced apoptosis (10). Previous studies indicated that Edwardsiella could induce apoptosis in fish cells. For example, E. tarda (now renamed as E. piscicida) could induce apoptosis in EPC cells (14). These results indicated that Bid forms in lower vertebrates possess pro-apoptotic activity, similar to that of mammalian Bid.
We observed that the full length of zebrafish Bid has antibacterial activity against E. ictaluri, which could significantly inhibit the proliferation of E. ictaluri, as confirmed by the in vitro and in vivo bacterial invasion assays (Figure 5). To further understand the potential mechanism for Bid antibacterial activity, we checked the expression of potential genes related to Bid in larvae. As expected, we found that the overexpression of the full-length Bid could significantly upregulate the expressions of birc5 and egfr in larvae infected with E. ictaluri. Birc5, also known as survivin, is an anti-apoptotic protein in vertebrates that can inhibit cell death (23). Egfr, a cell surface receptor, involves regulating cell growth in both normal and malignant tissues (24). The upregulation of these two genes indicated that they are involved in the cell apoptosis induced by Bid during E. ictaluri infection. Importantly, we observed that overexpression of the full-length Bid downregulated the expression of p53 both in uninfected larvae and in larvae infected with E. ictaluri (Figure 6). The p53 gene, an important transcription factor initially described as an oncogene, and later studies demonstrated it is part of the immune system (25). Downregulation of p53 suggested that Bid might regulate p53 to exert antibacterial activity. Furthermore, we found that overexpression of p53 could significantly decrease the survival rates of embryos following E. ictaluri infection. Further, the numbers of E. ictaluri in larvae were greatly increased, indicating that p53 plays a negative role during E. ictaluri infection. We also found that the survival rates of larvae co-microinjected with Bid-FLAG and p53-FLAG were significantly increased, and the numbers of E. ictaluri in EPC cells co-transfected with Bid-FLAG and p53-FLAG were decreased. Collectively, these results indicated that Bid could inhibit the negative role of p53 during E. ictaluri infection.
Mammalian Bid needs to be cleaved by caspase-8 to exert most of its functions. However, some studies also found that functional Bid does not need to be cleaved by caspase-8. For example, a mutant Bid that lacks the caspase-8 cleavage site could still kill cells (26). Moreover, endogenous Bid could be translocated to the mitochondria without cleavage in an anoikis model (27). Here, we found that pretreatment of larvae with Z-IETD-FMK, an inhibitor of caspase-8, could significantly decrease the survival rate of larvae and increase the number of E. ictaluri in larvae, indicating that when caspase-8 was inhibited, the antibacterial properties were also inhibited. However, we found that despite the pretreatment of larvae with Z-IETD-FMK, the overexpression of Bid still had perfect antibacterial activity against E. ictaluri, indicating that the antibacterial activity of Bid does not rely on the cleavage of caspase-8. These results confirmed that the full-length Bid inhibit the p53 role to exert its antibacterial activity, which do not rely on the caspase-8-mediated Bid cleavage (Figure 9).
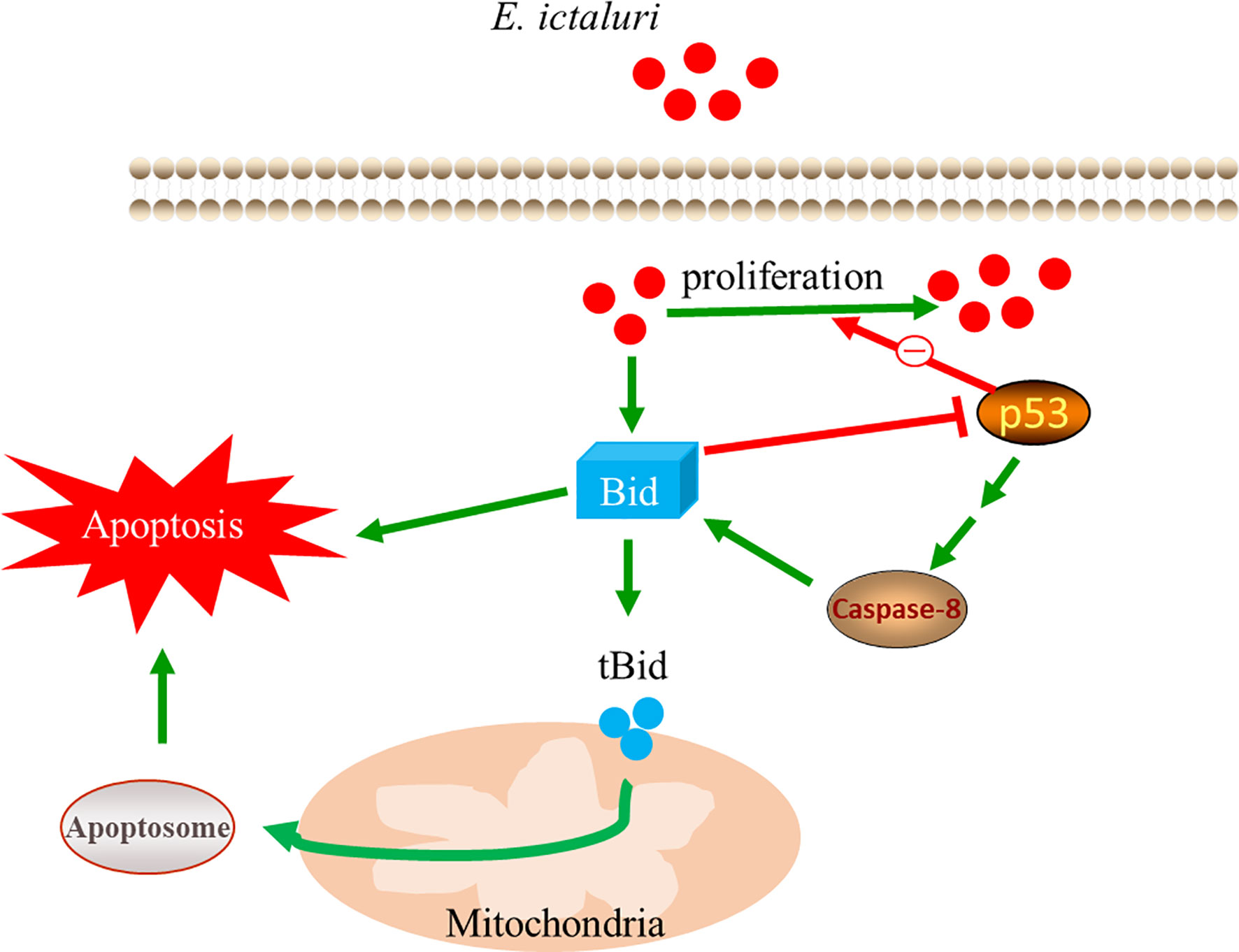
Figure 9 Mechanism of the antibacterial activity of full-length Bid.
In summary, zebrafish Bid possess pro-apoptotic activity, similar to that of mammalian Bid. More importantly, zebrafish Bid had antibacterial activity, which could significantly inhibit the invasion and proliferation of E. ictaluri in vivo and in vitro. The antibacterial activity of Bid was achieved by inhibiting the negative role of p53 during E. ictaluri infection but does not rely on caspase-8-mediated Bid cleavage.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The animal study was reviewed and approved by the Animal Care and Use Committee of the Institute of Hydrobiology, CAS.
MC and ZQ designed the experiments and wrote the manuscript. DY performed the experiments. LC and YX helped in preparing the manuscript. All authors contributed to the article and approved the submitted version.
This work was funded by the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant XDA24010308) and the National Natural Science Foundation of China (Grant 31873046 and 31302221). ZQ was supported financially by the projects for “six talents” of Jiangsu Province (Grant No. NY-115).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank LetPub (www.letpub.com) for its linguistic assistance and scientific consultation during the preparation of this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.707426/full#supplementary-material
1. Kratz E, Eimon PM, Mukhyala K, Stern H, Zha J, Strasser A, et al. Functional Characterization of the Bcl-2 Gene Family in the Zebrafish. Cell Death Differ (2006) 13:1631–40. doi: 10.1038/sj.cdd.4402016
2. Bruckheimer EM, Cho SH, Sarkiss M, Herrmann J, McDonnell TJ. The Bcl-2 Gene Family and Apoptosis. Adv Biochem Eng Biotechnol (1998) 62:75–105. doi: 10.1007/BFb0102306
3. Choudhury S. A Comparative Analysis of Bcl-2 Family. Bioinformation (2019) 15(4):299–306. doi: 10.6026/97320630015299
4. Droin NM, Green DR. Role of Bcl-2 Family Members in Immunity and Disease. Biochim Biophys Acta (2004) 1644:179–88. doi: 10.1016/j.bbamcr.2003.10.011
5. Yin XM. Bid, a BH3-Only Multi-Functional Molecule, Is at the Cross Road of Life and Death. Gene (2006) 369:7–19. doi: 10.1016/j.gene.2005.10.038
6. Billen LP, Shamas-Din A, Andrews DW. Bid: A Bax-Like BH3 Protein. Oncogene (2009) 27:S93–S104. doi: 10.1038/onc.2009.47
7. Yeretssian G, Correa RG, Doiron K, Fitzgerald P, Dillon CP, Green DR, et al. Non-Apoptotic Role of BID in Inflammation and Innate Immunity. Nature (2011) 474:96–9. doi: 10.1038/nature09982
8. Toni M, Manciocco A, Angiulli E, Alleva E, Cioni C. Malavasi. Review: Assessing Fish Welfare in Research and Aquaculture, With a Focus on European Directives. Animal (2019) 13:161–70. doi: 10.1017/S1751731118000940
9. Coultas L, Huang DCS, Adams JM, Strasser A. Pro-Apoptotic BH3-Only Bcl-2 Family Members in Vertebrate Model Organisms Suitable for Genetic Experimentation. Cell Death Differ (2002) 9:1163–6. doi: 10.1038/sj.cdd.4401096
10. He L, Wang H, Luo L, Li Y, Huang R, Liao L, et al. Bid-Deficient Fish Delay Grass Carp Reovirus (GCRV) Replication and Attenuate GCRV-Triggered Apoptosis. Oncotarget (2017) 8(44):76408–22. doi: 10.18632/oncotarget.19460
11. Yuan ZH, Liu SK, Yao J, Zeng QF, Tan SX, Liu ZJ. Expression of Bcl-2 Genes in Channel Catfish After Bacterial Infection and Hypoxia Stress. Dev Comp Immunol (2016) 65:79–90. doi: 10.1016/j.dci.2016.06.018
12. Hawke JP, Kent M, Rogge M, Baumgartner W, Wiles J, Shelley J, et al. Edwardsiellosis Caused by Edwardsiella Ictaluri in Laboratory Populations of Zebrafish Danio Rerio. J Aquat Anim Health (2013) 25:171–83. doi: 10.1080/08997659.2013.782226
13. Kent ML, Sanders JL, Spagnoli S, Al-Samarrie CE, Murray KN. Review of Diseases and Health Management in Zebrafish Danio Rerio (Hamilton 1822) in Research Facilities. J Fish Dis (2020) 43:637–50. doi: 10.1111/jfd.13165
14. Wang B, Yu T, Dong X, Zhang ZH, Song L, Xu Y, et al. Edwardsiella Tarda Invasion of Fish Cell Lines and the Activation of Divergent Cell Death Pathways. Vet Microbiol (2013) 163:282–9. doi: 10.1016/j.vetmic.2012.12.027
15. Cowburn AS, White JF, Deighton J, Walmsley SR, Chilvers ER. Z-VAD-Fmk Augmentation of TNF Alpha-Stimulated Neutrophil Apoptosis Is Compound Specific and Does Not Involve the Generation of Reactive Oxygen Species. Blood (2005) 105(7):2970–2. doi: 10.1182/blood-2004-07-2870
16. Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol Biol Evol (2018) 35:1547–9. doi: 10.1093/molbev/msy096
17. Puthalakath H, Strasser A. Keeping Killers on a Tight Leash: Transcriptional and Posttranslational Control of the Pro-Apoptotic Activity of BH3-Only Proteins. Cell Death Differ (2002) 9:505–12. doi: 10.1038/sj.cdd.4400998
18. Tekedar HC, Blom J, Kalindamar S, Nho S, Karsi A, Lawrence ML. Comparative Genomics of the Fish Pathogens Edwardsiella Ictaluri 93-146 and Edwardsiella Piscicida C07-087. Microb Genom (2020) 6:e000322. doi: 10.1099/mgen.0.000322
19. Schendel SL, Azimov R, Pawlowski K, Godzik A, Kagan BL, Reed JC. Ion Channel Activity of the BH3 Only Bcl-2 Family Member, BID. J Biol Chem (1999) 274(31):21932–6. doi: 10.1074/jbc.274.31.21932
20. Sutton VR, Davis JE, Cancilla M, Johnstone RW, Ruefli AA, Sedelies K, et al. Initiation of Apoptosis by Granzyme B Requires Direct Cleavage of Bid, But Not Direct Granzyme B-Mediated Caspase Activation. J Exp Med (2000) 192(10):1403–14. doi: 10.1084/jem.192.10.1403
21. Mandic A, Viktorsson K, Strandberg L, Heiden T, Hansson J, Linder S. Etal.Calpain-Mediated Bid Cleavage and Calpain-Independent Bak Modulation: Two Separate Pathways in Cisplatin-Induced Apoptosis. Mol Cell Biol (2002) 22(9):3003–13. doi: 10.1128/MCB.22.9.3003-3013.2002
22. Li YQ, Liu YN, Zhou Y, Liu WZ, Fan YD, Jiang N, et al. Bid is Involved in Apoptosis Induced by Chinese Giant Salamander Iridovirus and Contributes to the Viral Replication in an Amphibian Cell Line. Dev Comp Immunol (2021) 116:103935. doi: 10.1016/j.dci.2020.103935
23. Wheatley SP, Altieri D. Survivin at a Glance. J Cell Sci (2019) 132:jcs223826. doi: 10.1242/jcs.223826
24. Jackson NM, Ceresa BP. EGFR-Mediated Apoptosis via STAT3. Exp Cell Res (2017) 356:93–103. doi: 10.1016/j.yexcr.2017.04.016
25. Levine A. P53 and the Immune Response: 40 Years of Exploration-a Plan for the Future. Int J Mol Sci (2020) 21:541. doi: 10.3390/ijms21020541
26. Sarig R, Zaltsman Y, Marcellus RC, Flavell R, Mak TW. Gross A.BID-D59A is a Potent Inducer of Apoptosis in Primary Embryonic Fibroblasts. J Biol Chem (2003) 278:10707–15. doi: 10.1074/jbc.M210296200
Keywords: BID, apoptosis, antibacterial activity, p53, Edwardsiella ictaluri
Citation: Qi Z, Yan D, Cao L, Xu Y and Chang M (2021) Zebrafish BID Exerts an Antibacterial Role by Negatively Regulating p53, but in a Caspase-8-Independent Manner. Front. Immunol. 12:707426. doi: 10.3389/fimmu.2021.707426
Received: 10 May 2021; Accepted: 10 August 2021;
Published: 31 August 2021.
Edited by:
Humberto Lanz-Mendoza, National Institute of Public Health, MexicoReviewed by:
Javier Santander, Memorial University of Newfoundland, CanadaCopyright © 2021 Qi, Yan, Cao, Xu and Chang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mingxian Chang, bWluZ3hpYW5jaGFuZ0BpaGIuYWMuY24=; Zhitao Qi, cWl6aGl0YW9AeWNpdC5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.