
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 19 April 2021
Sec. T Cell Biology
Volume 12 - 2021 | https://doi.org/10.3389/fimmu.2021.661990
This article is part of the Research TopicCD4 T cells in HIV: A Friend or Foe?View all 16 articles
 Lisa Van de Wijer1*†
Lisa Van de Wijer1*† Wouter A. van der Heijden1†
Wouter A. van der Heijden1† Rob ter Horst1
Rob ter Horst1 Martin Jaeger1
Martin Jaeger1 Wim Trypsteen2
Wim Trypsteen2 Sofie Rutsaert2
Sofie Rutsaert2 Bram van Cranenbroek3Esther van Rijssen3
Bram van Cranenbroek3Esther van Rijssen3 Irma Joosten3
Irma Joosten3 Leo Joosten1
Leo Joosten1 Linos Vandekerckhove2Till Schoofs4Jan van Lunzen4
Linos Vandekerckhove2Till Schoofs4Jan van Lunzen4 Mihai G. Netea1,5
Mihai G. Netea1,5 Hans J.P.M. Koenen3
Hans J.P.M. Koenen3 André J.A.M. van der Ven1
André J.A.M. van der Ven1 Quirijn de Mast1
Quirijn de Mast1Long-term changes in the immune system of successfully treated people living with HIV (PLHIV) remain incompletely understood. In this study, we assessed 108 white blood cell (WBC) populations in a cohort of 211 PLHIV on stable antiretroviral therapy and in 56 HIV-uninfected controls using flow cytometry. We show that marked differences exist in T cell maturation and differentiation between PLHIV and HIV-uninfected controls: PLHIV had reduced percentages of CD4+ T cells and naïve T cells and increased percentages of CD8+ T cells, effector T cells, and T helper 17 (Th17) cells, together with increased Th17/regulatory T cell (Treg) ratios. PLHIV also exhibited altered B cell maturation with reduced percentages of memory B cells and increased numbers of plasmablasts. Determinants of the T and B cell composition in PLHIV included host factors (age, sex, and smoking), markers of the HIV reservoir, and CMV serostatus. Moreover, higher circulating Th17 percentages were associated with higher plasma concentrations of interleukin (IL) 6, soluble CD14, the gut homing chemokine CCL20, and intestinal fatty acid binding protein (IFABP). The changes in circulating lymphocytes translated into functional changes with reduced interferon (IFN)- γ responses of peripheral blood mononuclear cells to stimulation with Candida albicans and Mycobacterium tuberculosis. In conclusion, this comprehensive analysis confirms the importance of persistent abnormalities in the number and function of circulating immune cells in PLHIV on stable treatment.
Combination antiretroviral therapy (cART) has drastically curtailed morbidity and mortality in people living with HIV (PLHIV) (1). Still, PLHIV remain at an increased risk for pneumococcal infections, Mycobacterium tuberculosis (M. tuberculosis) reactivation and impaired vaccine responses (2–6). Moreover, HIV infection predisposes to non-infectious comorbidities, such as cardiovascular disease and non-AIDS-related cancer, which share an underlying pathophysiological pathway characterized by a persisting and inappropriate activation of innate and adaptive immune cells (7, 8). Together, these observations point towards a disbalance in the homeostasis of the immune system, characterized by immunodeficiency on the one hand, and chronic inflammation on the other hand.
HIV-1 preferentially infects and kills activated CD4+ T cells, leading to rapid and severe CD4+ T cell depletion in the gut and increased microbial translocation (9–12). A small proportion of these cells remains latently infected with replication competent virus and defective proviruses, called the HIV-1 reservoir (13). The HIV-1 reservoir and increased microbial translocation, together with lifestyle factors and co-infections such as cytomegalovirus (CMV) may all contribute to the disrupted immune function in PLHIV (10, 13–17). However, published data have shown inconsistent or even contradictory findings. Heterogeneity in study populations, limited sample sizes of study populations and differences in lifestyle factors, including use of tobacco and recreational drugs, may underlie these inconsistencies and emphasize the need for an integrative approach in evaluating the immune system in PLHIV on stable cART.
The Human Functional Genomics Project (HFGP) aims to disentangle variation in the immune system in different cohorts of healthy individuals and individuals with underlying conditions. It combines multiple levels of analyses and data integration, including demographic and lifestyle data, data from ‘omics technologies’, and functional immune data (18). As part of this project, we previously identified relevant environmental and host factors for circulating white blood cell (WBC) populations in healthy individuals (19). Embedded within the HFGP, we established a cohort of 211 virally suppressed PLHIV (200HIV) and showed that these individuals exhibited a sustained pro-inflammatory immune phenotype with priming of the interleukin (IL)-1β pathway (20). In the present study, we used the same cohort to comprehensively assess the peripheral WBC composition during treated HIV infection with a special focus on the adaptive immune system. We compared their WBC populations with those of healthy individuals and assessed associations with demographic and lifestyle factors, different HIV-specific factors, and ex vivo cytokine responses of peripheral blood mononuclear cells (PBMCs) to stimulation with different bacterial, fungal and viral antigens.
This study is part of the HFGP (www.humanfunctionalgenomics.org) (18). Between 14 December 2015 and 6 February 2017, individuals living with HIV were recruited from the HIV clinic of Radboud university medical center. Inclusion criteria were Caucasian ethnicity, age ≥ 18 years, receiving cART > 6 months, and latest HIV-RNA levels ≤200 copies/ml. Exclusion criteria were: signs of acute or opportunistic infections, antibiotic use <1 month prior to study visit, and active hepatitis B/C. The control population consisted of 56 healthy adult individuals (56P cohort), who did not suffer from any acute or chronic conditions and who were longitudinally sampled four times in three-month intervals. Inclusion, sampling and sample processing of both cohorts were conducted simultaneously and by the same personnel. The 56P participants were selected as a subset of a larger cohort of 534 healthy individuals (500FG) which was phenotypically assessed two years earlier according to the same methods (19, 21). Differences in cell-cell associations were compared between 200 HIV and 500FG cohort. The reason is that the larger sample size of this control cohort (n=534 vs n=56) improved statistical power, whilst batch effects between PLHIV and 500FG were deemed of less significance when comparing within-group correlations between cohorts. General information from all participants was recorded in the Castor Electronic Data Capture program (Castor EDC, CIWIT B.V., Amsterdam, The Netherlands), using questionnaires on socio-demographic information, lifestyle and life-events. Clinical data were extracted from medical files and the ‘Stichting HIV Monitoring’ registry (Amsterdam, The Netherlands).
The study protocols were approved by the Medical Ethical Review Committee region Arnhem-Nijmegen (ref. 42561.091.122) and conducted in accordance with the principles of the Declaration of Helsinki. All study participants provided written informed consent.
Venous blood was collected in sterile 10 ml EDTA BD Vacutainer® tubes between 8 and 11 am and processed within 1-4 hours. Cell counts were determined using a Sysmex XN-450 automated hematology analyzer (Sysmex Corporation, Kobe, Japan). Erythrocyte-lysed whole blood samples were obtained after 10 minutes incubation of 1.5 ml EDTA-anticoagulated blood with lysis buffer containing 3.0 M NH4Cl, 0.2 M KHCO3 and 2 mM Na4EDTA. The remaining WBC were washed twice, by adding 25 ml phosphate-buffered saline 1x (PBS, Braun, Melsungen, Germany) and centrifuging at 452 x g for 5 min at room temperature. Before staining, cells were resuspended in 300 μl of PBS enriched with 0.2% bovine serum albumin (BSA, Sigma-Aldrich, Zwijndrecht, Netherlands). Isolation of PBMCs was performed by density centrifugation of EDTA-anticoagulated blood diluted 1:1 in pyrogen-free PBS over Ficoll-Paque (VWR, Amsterdam, The Netherlands) as described previously (22). Cells were adjusted to 5.0 x 106 cells/ml.
Supplementary Table 1 summarizes the antibody clones and the fluorochrome conjugates used for the different panel fluorescent staining mixes. Staining was performed on 100 μl/well erythrocyte-lysed blood (panel 1-3,5) or 0.5 x 106 cells/well freshly isolated PBMCs (panel 4). Cells were stained according to previously described procedures (see also Supplementary Methods) (19).
Samples were measured on a 10-color Navios flow cytometer (Beckman Coulter, Fullerton, CA, USA), equipped with three solid-state lasers (488 nm, 638 nm, and 405 nm) (19). Gating was conducted manually and verified by two independent specialists to prevent gating errors. Samples were analyzed within 4-5 hr after blood collection, using five distinct and complementary 10-color antibody panels: 1. general; 2. T cell; 3. B cell; 4. intracellular T cell/Treg; 5. chemokine receptors (CCR). Staining and gating strategies can be found in Supplementary Figure 1. Flow cytometry data were analyzed using Kaluza software version 1.3. In our analyses, we focused on a set of 108 manually annotated WBC subsets based on the original 500FG study (panel 1-4) (19), with the addition of a fifth panel in which we classified monocytes, CD4+ memory and regulatory T cells, and CD8+ cells according to their expression of the CXCR3, CCR4, and CCR6 (panel 5).
Freshly isolated PBMCs were incubated with different stimuli (Supplementary Table 2) including bacterial (Staphylococcus aureus, M. tuberculosis, Streptococcus pneumoniae [S. pneumoniae]), fungal (Cryptococcus gattii, Candida albicans [C. albicans] hyphae and yeast) and other relevant antigens (Imiquimod, TLR7 ligand), in round-bottom 96-well plates (Greiner Bio-One, Frickenhausen, Germany) with 0.5 x 106 cells/well at 37°C and 5% CO2 in the presence of 10% human pooled serum for seven days. Supernatants were stored at -20°C. Levels of the lymphocyte-derived cytokines IL-17, IL-22, and interferon (IFN)-γ were measured in the supernatants (PeliKine Compact or Duoset ELISA, R&D Systems).
Serum levels of immunoglobulin (Ig)M and IgG were measured by immunonephelometry using a Beckman Coulter Imager and Beckman Coulter reagents. Measurements were standardized using certified European reference material 470 (ERM®-DA470). CMV IgG antibodies were measured in serum using ELISA (Genway Biotech Inc., San Diego, CA, USA) according to the manufacturer’s instructions. Markers of systemic inflammation, high-sensitive C-reactive protein (hsCRP) and soluble CD14 (sCD14), and microbial translocation, intestinal fatty acid binding protein (IFABP), were measured by ELISA (Quantikine, R&D Systems) according to the manufacturer’s instructions. IL-6 was measured using a SimplePlex Cartridge (Protein Simple, San Jose, CA, USA). Circulating plasma CCL20, IL-17A, and IFN-γ were measured using the commercially available Olink Proteomics AB Inflammation Panel as described previously (23, 24).
The HIV reservoir was assessed by analyzing CD4+ cell-associated HIV-1 DNA (CA-DNA) and CA-RNA. In virally suppressed patients, the CA-DNA roughly equals the integrated HIV-1 DNA, being replication competent or not (25), while CA-RNA is associated with recent HIV-1 transcriptional activity and serves as a proxy for the active proviral reservoir (26). CA-DNA and CA-RNA were measured in triplicate by droplet digital PCR (ddPCR – Bio-Rad) in CD4+ T cells isolated using EasySep Human CD4+ T Cell Isolation Kit (Stemcell technologies, Vancouver, Canada) as described previously (27). CA-DNA was extracted by the DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol with the addition of 75µl elution buffer on the column heated at 56°C for 10 minutes. CA-RNA was extracted using the Innuprep RNA kit (Westburg, Leusden, The Netherlands) with 30µl elution buffer. Total RNA was reversely transcribed to cDNA by qScript cDNA SuperMix according to manufacturer’s protocol (Quantabio, Beverly, MA, USA). CA-DNA was normalized by measuring the reference gene RPP30 (Supplementary Table 3 and Supplementary Methods) in duplicate by ddPCR and expressed per million CD4+ T cells. CA-RNA was normalized using three reference genes per patient, (B2M, ACTB and GADPH), which were measured with LightCycler 480 SYBR Green I Master mix. HIV-1 RNA copies were divided by the geometric mean of the reference genes and expressed per million PBMCs. ddPCR data analysis was performed using the ddpcRquant tool with standard settings for thresholding and absolute quantification (28).
A detailed description of the statistical methods can be found in the Supplementary Methods. Given the impact of cohort differences in the absolute cell numbers of main WBC types (e.g. CD4+) on the numbers of their subsets (e.g. CD4+ regulatory T cells [Tregs]), results were primarily reported as WBC percentages, unless stated otherwise. WBC percentages were calculated by dividing the cell count of each subpopulation by its respective parent (one level up) or, where relevant, grandparent (two levels up; Supplementary Table 4). Data curation of cytokine, soluble marker, and immunoglobulin data was done according to previous methods (21). For comparisons between cohorts, we used all four longitudinally collected data points from the 56P cohort as independent measurements. Because of the known seasonality effects on WBC, this approach was considered preferable over selecting one of the data collection points or summarizing the data points. Data were normalized using an inverse rank transformation algorithm.
Comparisons in baseline characteristics between groups were made using Student’s t-test (or Mann-Whitney U test) for continuous variables, and Pearson’s Chi-square test (or Fisher’s exact) for categorical variables. Linear regression was used to compare WBC between cohorts, and to calculate associations between WBC and clinical and virological factors. All analyses were corrected for age, sex, time since January 2015 and seasonal effects. Cytokine analyses between cohorts were also corrected for CD4+ and CD8+ T cell percentages. Spearman correlations were used as the distance metric for unsupervised hierarchical WBC count clustering. Two-sided FDR-corrected p-values < 0.05 were considered statistically significant (29). Effect sizes are reported as Spearman’s Rho or standardized beta coefficients (β). Data were analyzed using the statistical programming language R (version 3.4.3, R Core Team, 2012).
Data from 211 PLHIV and 56 controls were analyzed in this study (Table 1). PLHIV were older (median [IQR] 52.5 [46.2 – 59.4] vs 30.0 [25.1 – 52.2] years, p<0.0001) and more often male (193/211 [91.5%] vs 34/56 [60.7%], p<0.0001) than controls. PLHIV had received cART for a median of 6.6 (4.2–11.9) years and 205/210 (98%) had plasma HIV-1 RNA <50 copies/mL at time of study visit. Analyzing the HIV reservoir, we found detectable levels of total CA-DNA in 207/208 (99.5%) and CA-RNA in 210/210 (100%); CA-DNA and CA-RNA levels were highly correlated (Supplementary Figure 2A; R=0.68, p<2.2·10-16). In general, PLHIV with higher CA-DNA and CA-RNA levels were older, had been living with and treated for HIV for a longer period, and had lower nadir CD4+ T cell counts (Supplementary Figure 2A). We observed no significant differences in CA-RNA and CA-DNA between PLHIV with plasma HIV-1 RNA <50 copies/mL (205/210 [98%]) and those with plasma HIV-1 RNA 50-200 copies/mL (5/210 [2%]; Supplementary Figure 2B). PLHIV with at least once a viral load of >50 copies/mL during year prior to study visit (23/210 [11.0%]) had higher levels of CA-RNA (p=0.0077), but no differences in CA-DNA (p=0.076; Supplementary Figure 2B).
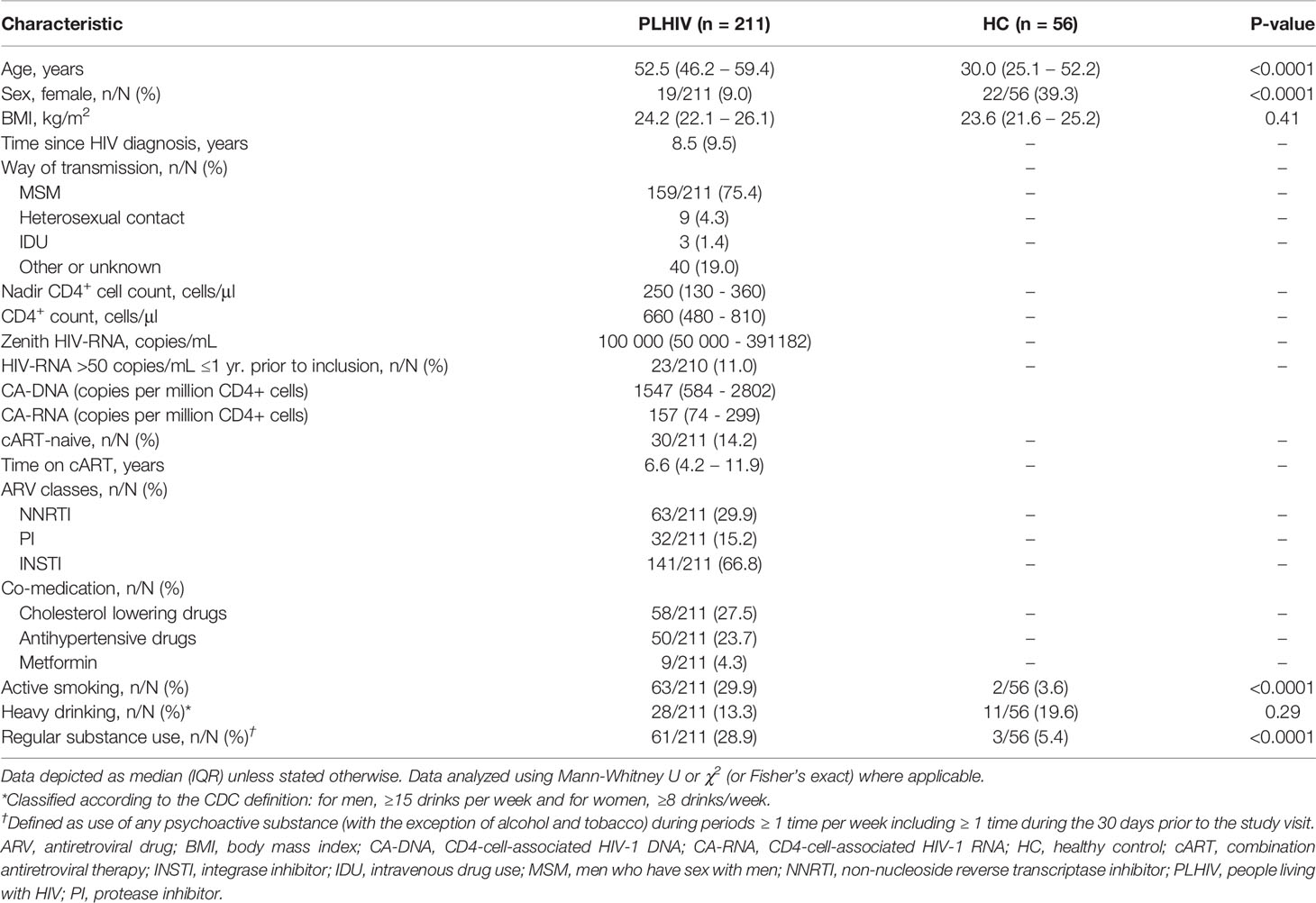
Table 1 General characteristics.
We analyzed 108 WBC populations, including 93/107 (87%) B and T cell subsets and 14/107 (13%) innate cell subsets (neutrophils, monocytes, natural killer [NK] and natural killer T cells [NKT]). Figure 1 and Supplementary Figure 3 show the differences in WBC composition between PLHIV and controls. An overview of median (IQR) WBC percentages and numbers in PLHIV and controls can be found in Supplementary Table 5.
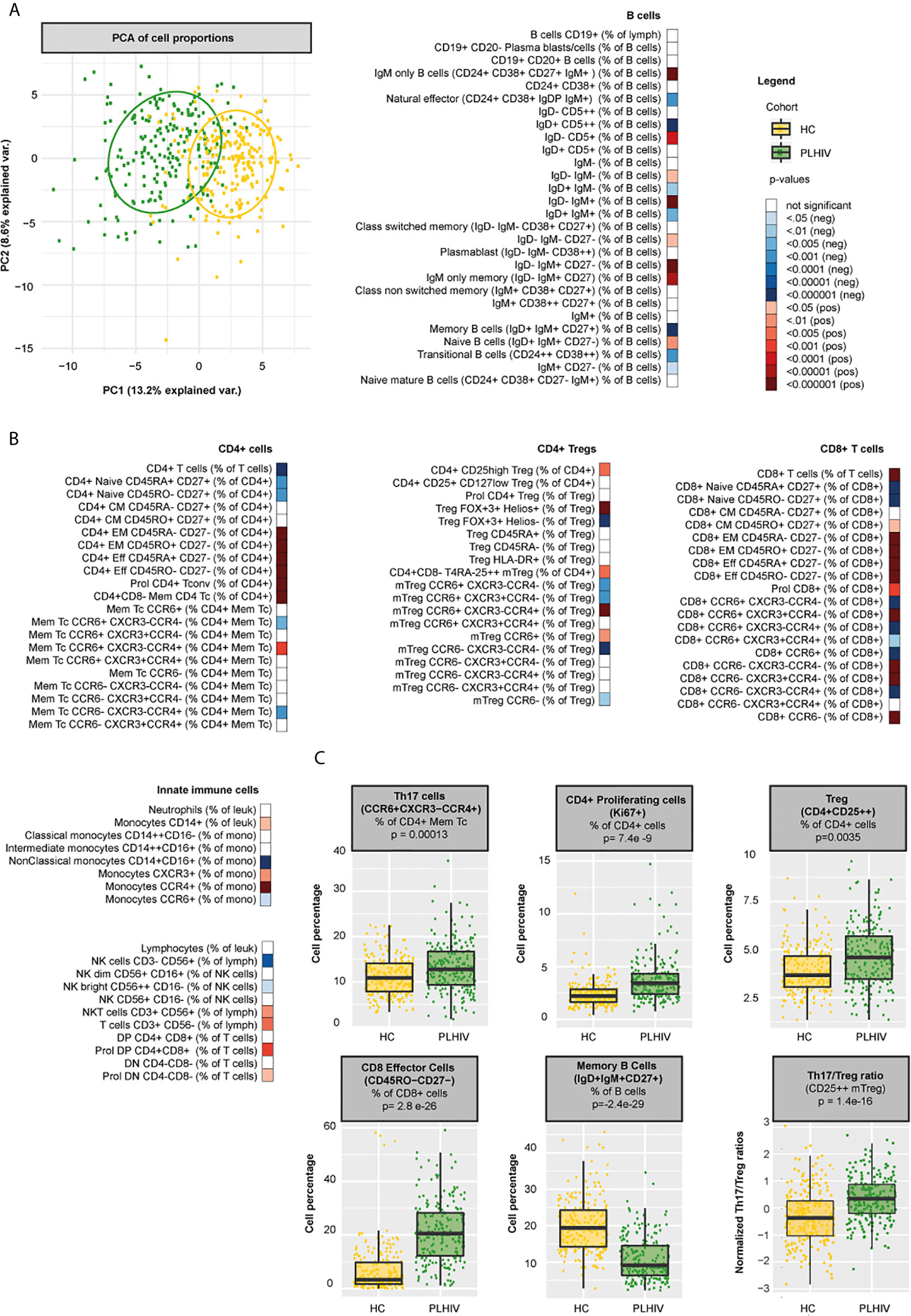
Figure 1 Differences in WBC percentages between PLHIV and healthy individuals (A) Principal component biplot showing standardized variance for the two principal component of all WBC subsets. The ovals represent the different cohorts. (B) Differences in WBC percentages (n=108 WBC subsets) between PLHIV (n=211) and healthy controls (n=56) (C) Boxplots showing examples of cohort differences in cell percentages and Th17/Treg ratios (Treg identification marker CD25++). Inverse-rank transformed data were analyzed using multiple linear regression and adjusted for age, sex, sampling time, and season. For color coding of the FDR-adjusted p-values see legend.
Principal component analysis (PCA) of all WBC populations only explained a limited amount of the total variance. Still, the PCA plot showed clear differences in clustering between PLHIV and controls (Figure 1A). As expected, PLHIV exhibited an expansion of CD8+ and a reduction of CD4+ T cell numbers and percentages (Figure 1B and Supplementary Table 5). Functionally, CD4+ T cells comprise a diverse population of cells. CD4+ T helper (Th) cells fulfill essential roles for viral (Th1, CCR6-CXCR3+CCR4-), parasitic (Th2, CCR6-CXCR3-CCR4+), and mucosal (Th17, CCR6+CXCR3-CCR4+) immunity (30). In addition, CD4+ Tregs are essential for controlling inflammation (31). Despite reduced CD4+ T cell counts, numbers of all Th cell subsets (CD4+ CD45RA- CD25-) were markedly increased in PLHIV compared to controls (Figure 1B and Supplementary Table 5). Within the Th pool, Th2 percentages were reduced in PLHIV compared to controls, whereas Th1 percentages did not differ. Remarkably, Th17 percentages and numbers were increased in PLHIV (Figures 1B, C and Supplementary Table 5). While Treg percentages (of CD4+ cells), including highly suppressive Tregs co-expressing the transcription factors FoxP3 and Helios, were also increased in PLHIV, absolute Treg numbers were reduced (Figures 1B, C and Supplementary Table 5). This relative increase of Tregs may result from a loss of other CD4+ subsets (32). Among Tregs, we found no differences in the percentage of activated (HLA-DR+) and effector Tregs (CD45RA-). The relationship between pro-inflammatory Th17 cells and Treg must remain balanced to preserve functional immunity. Altered ratios have been described in untreated HIV (lower Th17/Treg), autoimmune disease and cancer (higher Th17/Treg) (9, 33, 34). Here, we found increased Th17/Treg ratios among virally suppressed PLHIV, irrespective of the Treg identification marker used (Figure 1C and Supplementary Figure 4). Furthermore, out of the Treg population, the percentage of Th17-like CCR6+ Tregs was increased in PLHIV, further contributing to a pro-inflammatory state. Developmentally, T cells evolve from naive T cells to antigen experienced central memory (CM), effector memory (EM), and effector cells (35, 36). HIV not only differentially affects functional subpopulations, but also disrupts these developmental stages. While naïve CD4+ and CD8+ T cells were reduced, memory and effector cells were expanded in PLHIV (Figure 1B and Supplementary Table 5). Likewise, we found higher percentages of proliferating (Ki67+) CD4+ and CD8+ T cells in PLHIV, indicating increased cell turnover. These results suggest a shift from naive cells towards terminally differentiated cells in HIV, even if viral replication is under control (37), which cannot be explained by age, sex, or season as we corrected our models for these factors.
Changes in the B cell compartment have also been described in PLHIV, including loss of CD27+ memory B cells (38, 39). Furthermore, viremia and low CD4+ T cell counts have been associated with the expansion of terminally differentiated B cells and immature B cells respectively (38, 39). In our study, we observed clear differences in B cell development in PLHIV illustrated by increased percentages of naïve B cells (IgD+IgM+CD27-) and reduced percentages of memory B cells (IgD+IgM+CD27+), transitional B cells (CD24++CD38++) and natural effector B cells (CD24+CD38+IgD+IgM+). In addition, the number of plasmablasts (IgD- IgM- CD38++) was increased in PLHIV, (Figures 1B, C and Supplementary Table 5). Adequate B cell maturation further requires optimal communication between B and T cells. We therefore sought to identify differences in WBC co-regulation between PLHIV and HIV-uninfected individuals by comparing cell-cell associations between PLHIV and participants from the 500FG cohort. Details of this 500FG cohort have been reported previously (500FG, n=534) (19). Using the same immunophenotyping techniques, we observed weaker correlations between naïve, CM and EM CD4+ T cells and several B cell subpopulations (including class-switch memory B cells) within PLHIV than in participants of the 500FG cohort, suggesting altered B/T cell interactions (Figure 2). These differences were not attributable to the influence of age, sex, or season as these factors were regressed out of the analysis.
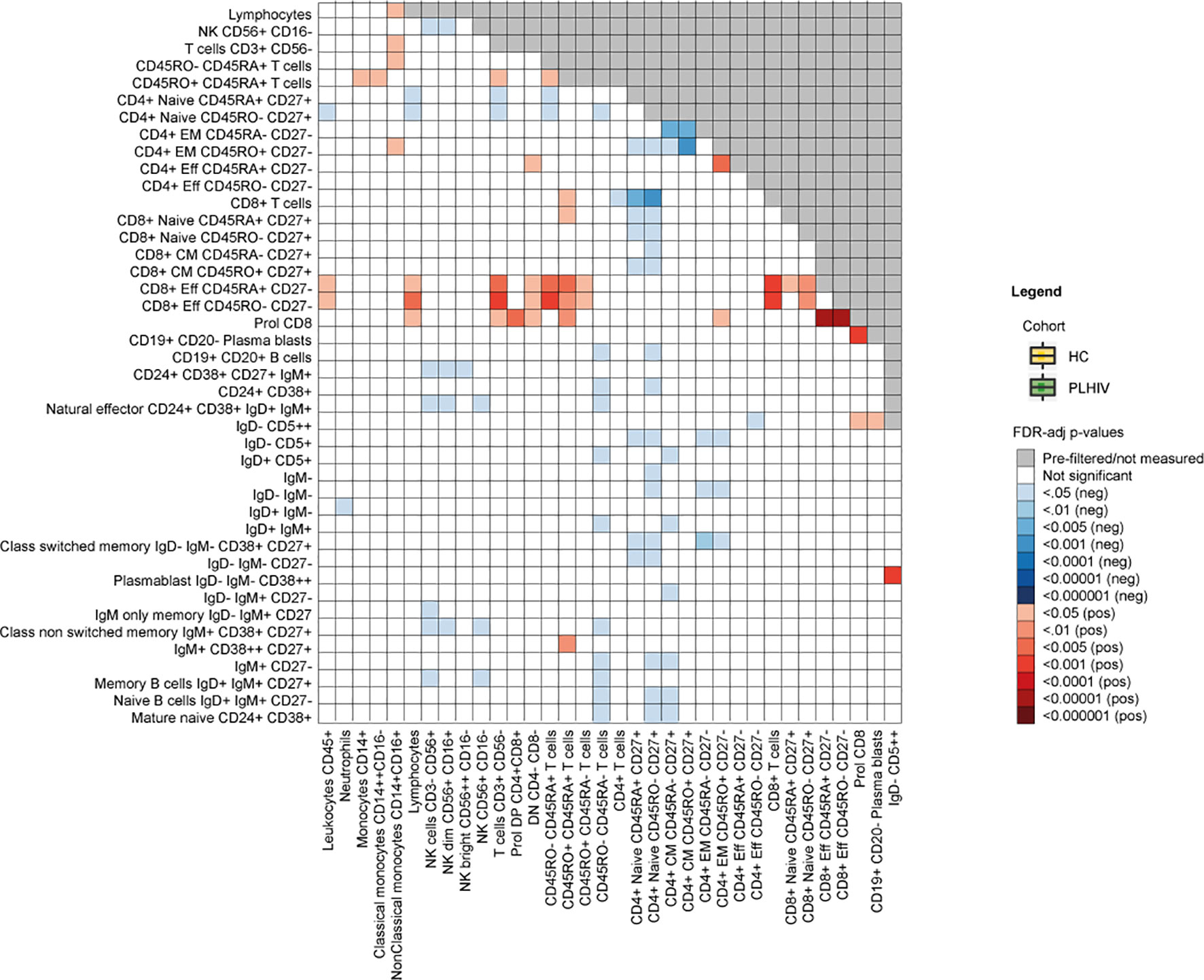
Figure 2 Differences in cell-cell associations between PLHIV and healthy individuals. Exploratory analysis depicting cell-cell associations between a total of 77 available WBC subsets that were significantly stronger (red) or weaker (blue) in PLHIV compared to healthy controls (n=534). FDR-adjusted p-values are obtained after 10 000 permutations and adjusted for age, sex, sampling time and season. HC, healthy control; PLHIV, people living with HIV; WBC, white blood cells.
Apart from changes in the adaptive cell compartment, we observed clear changes in the innate WBC compartment in PLHIV. Monocyte and NKT cell numbers and percentages were increased, whereas NK cell numbers and percentages were reduced, specifically the cytokine-producing NK bright cells (Figure 1B and Supplementary Table 5) Together, these data show a widespread functional and developmental dysregulation of the immune system in virally suppressed PLHIV. This dysregulation encompasses both the innate and the adaptive compartment and results in a pro-inflammatory immune environment with an expansion of monocytes, pro-inflammatory Th and effector T cells, and dysregulated B cell memory responses.
Apart from the effects of HIV, demographic and lifestyle-related factors may influence the composition of the circulating WBC populations. Indeed, we found that older age was associated with an increase of innate immune cells and differential effects on B and T cell populations, for example with lower percentages of naïve T cells (CD4+ T cells β=-0.29, p=0.00020; CD8+ T cells β=-0.29, p=0.00020) and B memory cells (β=-0.39, p=1.5·10-7) and higher percentages of memory T cells (e.g. CD8+ CM β=0.22, p=0.00064) and mature naïve B cells in PLHIV (β=0.25, p=0.00087, Figure 3 and Supplementary Table 6). Sex-dependent influences of the WBC composition, reflected by more effector and EM cells and fewer naïve (B and T) cells in males in HIV infection, resembled those observed previously observed in healthy individuals (21, 40).
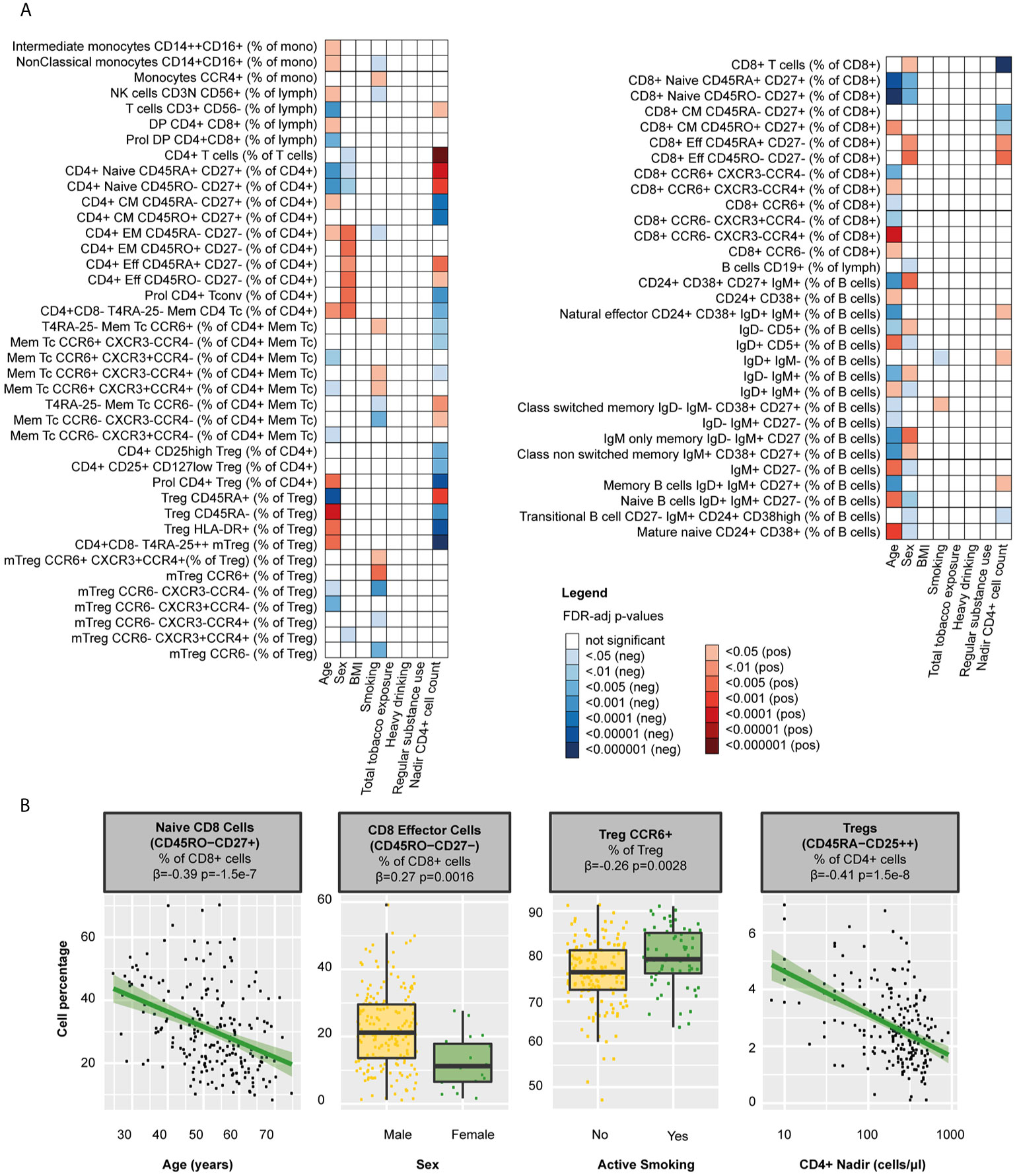
Figure 3 Clinical determinants of WBC percentages in PLHIV (A) Heatmap of WBC percentages (n=108 WBC subsets) that were significantly associated with any of the clinical determinants tested in 211 PLHIV; WBC subsets that showed no significant correlations with any of the parameters (n=37) have been removed from the figure. (B) Examples of WBC subsets that were significantly associated with age, sex, smoking, or CD4 nadir. Inverse-rank transformed data were analyzed using linear regression and adjusted for age, sex, sampling time, and season. For color coding of the FDR-adjusted p-values see legend. PLHIV, people living with HIV; WBC, white blood cells.
Lifestyle risk behaviors such as smoking [63/211 (29.9%)], heavy drinking [28/211 (13.3%)], and regular drug use [61/211 (28.9%)] were highly prevalent in PLHIV (Table 1). We found that packyears (reflecting total tobacco exposure) were associated with higher frequencies of neutrophils (β=0.22, p=0.033), Treg (β=0.22, p=0.033), CD8+ subsets (e.g. CCR6-CXCR3-CCR4+ β=0.24, p=0.025), and class switched memory B cells (β=0.20, p=0.042, Supplementary Table 6). Active smoking correlated with higher percentages of Th17 (β=0.21, p=0.031) and CCR6+ Tregs (β=0.26, p=0.0028, Figure 3 and Supplementary Table 6). Neither heavy drinking, nor regular drug use affected the WBC composition.
We further assessed the effects of relevant clinical factors on the WBC composition in PLHIV, by exploring associations with the history of immune suppression and treatment-related factors. We found that nadir CD4+ cell counts were closely associated with both B and T cell percentages in PLHIV (Figure 3 and Supplementary Table 6). For example, higher counts were associated with higher percentages of naïve CD4+ T cells (β=0.45, p=1.2·10-9) and memory B cells (β=0.18, p=0.034). In contrast, we observed no effects of the duration of HIV infection or cART (regimen) on the WBC composition (Supplementary Tables 6).
First, we explored the relationship with markers of the HIV reservoir. CA-DNA was negatively associated with CD4+ T cell percentages (β=-0.33, p=9.8·10-5) and positively with CD8+ T cell percentages (β=0.34, p=7.0·10-5). Within the CD4+ T cell pool, CA-DNA correlated with higher percentages of CM and Th17 cells (β=0.21, p=0.026 and β=0.24, p=0.0081 respectively; Figure 4 and Supplementary Table 7) Higher CA-DNA was also associated with more CD4+ T cell proliferation (Ki67+, β=0.35, p=3.3·10-5) and Treg activation (HLA-DR+, β=0.23, p=0.0081). We found similar associations between T cells and CA-RNA, whereas we found no relation between T cells and the relative HIV transcription level (CA-RNA/CA-DNA ratios; Supplementary Table 7). Lastly, we observed several associations between CA-DNA and B cells: higher CA-DNA levels correlated with reduced percentages of IgD+ only memory B cells (β=-0.27, p=0.00068) and, albeit non-significantly, with reduced percentages of natural effector B cells (CD24+CD38+IgD+IgM+, β=-0.18, p=0.056), and class unswitched memory B cells (IgD+IgM+CD27+, β=-0.18, p=0.053). Exclusion of PLHIV with HIV-RNA 50-200 copies/mL at time of study visit (5/210 [2%]) did not change the main conclusions of our paper (Supplementary Tables 8, 9).
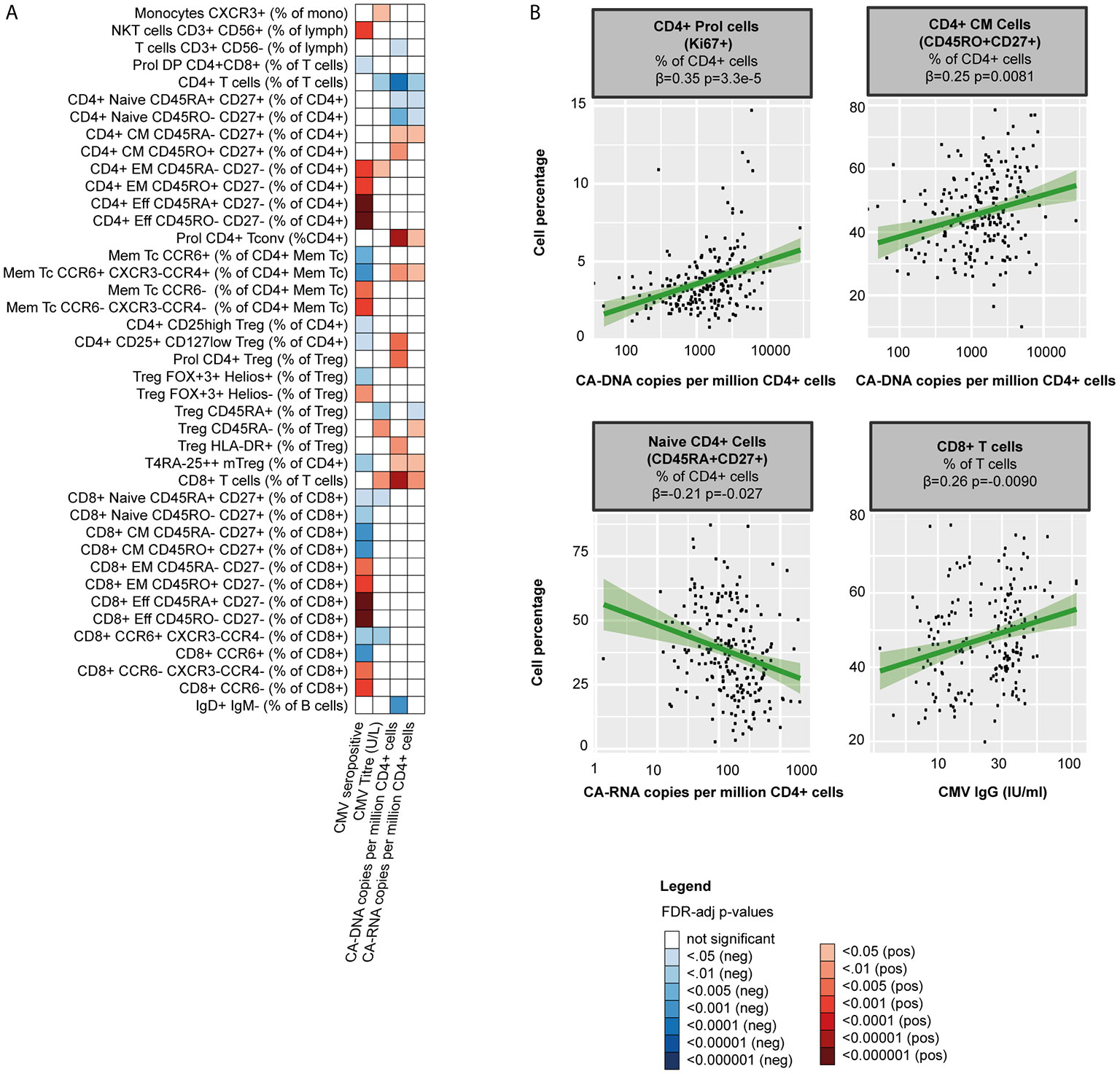
Figure 4 The HIV-1 reservoir, CMV serostatus and WBC percentages in PLHIV (A) Heatmap of significant associations between WBC percentages (total n=108), CMV co-infection (serostatus and IgG titers in 198 seropositive PLHIV) and markers of the HIV-1 reservoir (CA -DNA and CA- RNA) in 211 PLHIV; WBC subsets that showed no significant correlations with any of the parameters (n=41) have been removed from the figure. (B) Examples of WBC percentages that were significantly associated with the viral reservoir or CMV IgG titers. Inverse-rank transformed data were analyzed using linear regression and adjusted for age, sex, sampling time, and season. For color coding of the FDR-adjusted p-values see legend. CA-DNA, CD4-cell-associated HIV-1 DNA; CA-RNA, CD4-cell-associated HIV-1 RNA; CMV, cytomegalovirus; PLHIV, people living with HIV; WBC, white blood cells.
Second, CMV co-infection may contribute to immune dysregulation in both treated and untreated PLHIV. As CMV is known to affect WBC populations in healthy controls (41), we explored the association of CMV serostatus with WBC composition. 198 of the PLHIV (93.8%) were seropositive for CMV. In line with findings in healthy individuals (41), CMV seropositivity correlated with higher percentages of effector, EM T cells (e.g. with CD8+ effector cells β=0.43, p=1.7·10-9 and CD4+ EM cells β=0.30, p=0.00011; Figure 4 and Supplementary Table 7) and NKT cells (β=0.29, p=0.00029), yet with lower percentages of Th17 cells (β=-0.26, p=0.00065). We observed no associations between B cell subsets and CMV.
Our main findings include a loss of naïve T cells and an expansion of Th17 in PLHIV compared to healthy controls, which related with lower nadir CD4+ T cells counts and higher levels of CA-DNA. To further assess the potential underlying mechanisms, we explored the relationship between these WBC subsets, markers of chronic inflammation, and markers of microbial translocation. As discussed above, naïve T cells are able to differentiate into different Th subsets depending on the cytokine environment (42). We previously showed that PLHIV in our cohort exhibited a pro-inflammatory profile with increased levels of hsCRP (p=0.00022) and sCD14 (a marker of monocyte activation, p=0.0025; Figure 5A) as well as markedly elevated monocyte-derived cytokine responses, particularly IL-1β (20). Such a pro-inflammatory cytokine environment may push the differentiation of naive CD4+ cells into Th17 cells (42). Indeed, we observed positive associations between Th17 percentages and circulating IL-6 (β=0.21, p=0.014) and sCD14 (β =0.17, p=0.035; Figures 5B, C). Notably, Th17 cells fulfill an essential role in mucosal defense and gut Th17 cells are known to be severely depleted during acute HIV (9). To test whether gut integrity might have been compromised in our cohort, we measured levels of the microbial translocation marker IFABP and found increased levels in PLHIV compared to healthy controls (p=7.6·10-5; Figure 5D), suggesting ongoing microbial translocation in chronic treated HIV, especially in those with higher CA-RNA and CA-DNA (CA-RNA β =0.18, p=0.0091 and CA-DNA β=0.21, p=0.0025, Figure 5E). Higher levels of IFABP were associated with an expansion of peripheral blood Th17 (β=0.17, p0.043; Figure 5B). Homing of Th17 to the gut (and other tissues, such as the skin) is directed by the chemokine CCL20, which is produced by tissue and immune cells (neutrophils and monocytes) and binds uniquely to the CCR6+ receptor (43). Correspondingly, we found strong associations between CCL20 and Th17 percentages (β=0.22, p=0.0037), CCL20 and Th17/Treg (β=0.26, p=0.00081 for Th17/FoxP3+ Treg, β=0.27, p=0.00081 for Th17/CD127low Treg, and β=0.19, p=0.013 for Th17/CD25++ Treg), CCL20 and circulating IL-17A levels (β=0.47, p=1.2·10-11) and CCL20 and IFABP (β=0.24, p=0.0011; Figure 5F). Together, these results suggest that the pro-inflammatory environment in chronic HIV may promote the differentiation of circulating naïve CD4+ T cells into Th17 cells and that these changes may be associated with changes in gut permeability and gut homing.
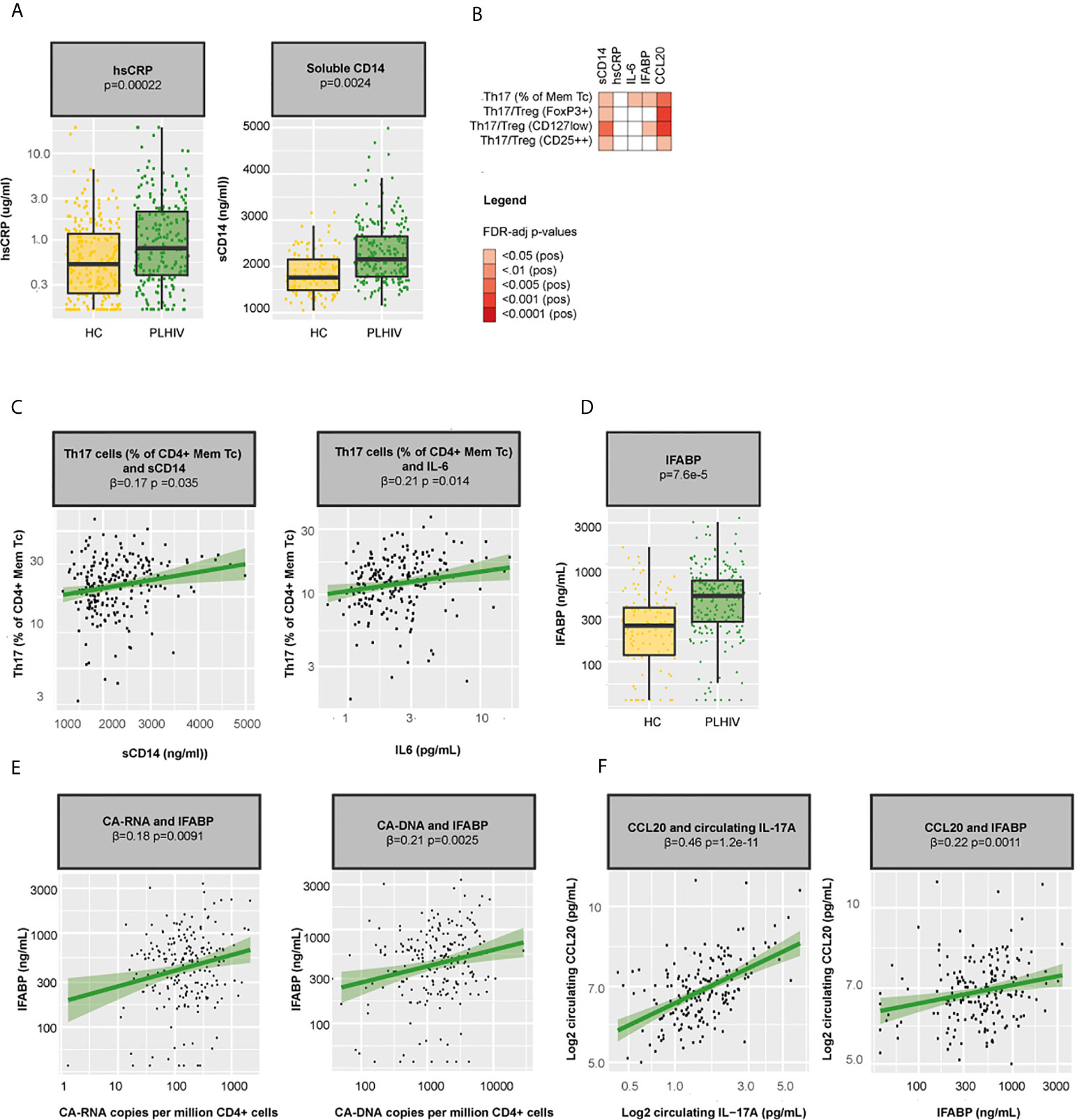
Figure 5 Inflammation, microbial translocation, and Th17 cells in PLHIV (A) Boxplots showing differences in hsCRP and sCD14 between PLHIV (n=211) and healthy controls (n=56). (B) Heatmap showing associations between Th17 percentages, Th17/Treg ratios and sCD14, hsCRP, IFABP, and CCL20. (C) Associations between Th17 percentages and sCD14 and circulating IL-6 in PLHIV. (D) Levels of the microbial translocation marker IFABP in PLHIV and healthy controls. (E) Associations between IFABP and CA-RNA and CA-DNA in PLHIV. (F) Associations between CCL20 and circulating IL-17A and IFABP. Inverse-rank transformed data were analyzed using linear regression analyses and corrected for sampling time. CA-DNA, CD4-cell-associated HIV-1 DNA; CA-RNA, CD4-cell-associated HIV-1 RNA; CCL20, Chemokine (C-C motif) ligand 20; HC, healthy control; hsCRP, high-sensitivity CRP; IFABP, intestinal fatty-acid binding protein; IL-6, interleukin 6; IL-17A, interleukin 17A; Mem Tc, CD4+ memory T cell; PLHIV, people living with HIV; sCD14, soluble CD14; Th17 percentages, T-helper 17 cell percentages (Mem Tc CCR6+ CXCR3-CCR4+ as percentage of CD4+ Mem Tc); Treg, regulatory T cells; WBC, white blood cells.
As our results indicated significant changes in circulating immune cell populations in PLHIV, we next analyzed the possible functional consequences. First, we measured ex vivo cytokine responses of PBMCs after stimulation with different stimuli. We found strong correlations between NK and T-cell percentages and ex vivo IFN-γ, IL-17, and IL-22 responses (Figure 6A and Supplementary Table 10): IFN-γ responses correlated with percentages of NK dim, CD4+ and CD8+ EM cells, whereas IL-22 responses correlated with CCR6+ CM CD4+ cell percentages. As expected, Th17 percentages were associated with increased circulating IL-17A (β=0.16, p=0.029) and ex vivo IL-17 responses to C. albicans (β=0.19, p=0.047) and with reduced IFN-γ responses (Figures 6A, E and Supplementary Table 10). Despite the expansion of Th17 cells among PLHIV compared to controls, ex vivo responses of IL-17 or IL-22 did not differ, suggesting that the functional capacity of these cells may be compromised. In contrast, IFN-γ production upon stimulation with C. albicans hyphae (p=0.012) and M. tuberculosis (p=0.0025) was reduced in PLHIV (Figure 6B). Among PLHIV, lower ex vivo IFN-γ production to stimulation with Imiquimod (β =-0.21, p=0.0020) and S. pneumoniae (β=-0.22, p=0.0011) and lower circulating IFN-γ concentrations (β=-0.18, p=0.014) were associated with higher CA-DNA levels (Figures 6C, D). No associations were found between ex vivo cytokine production and CMV seropositivity.
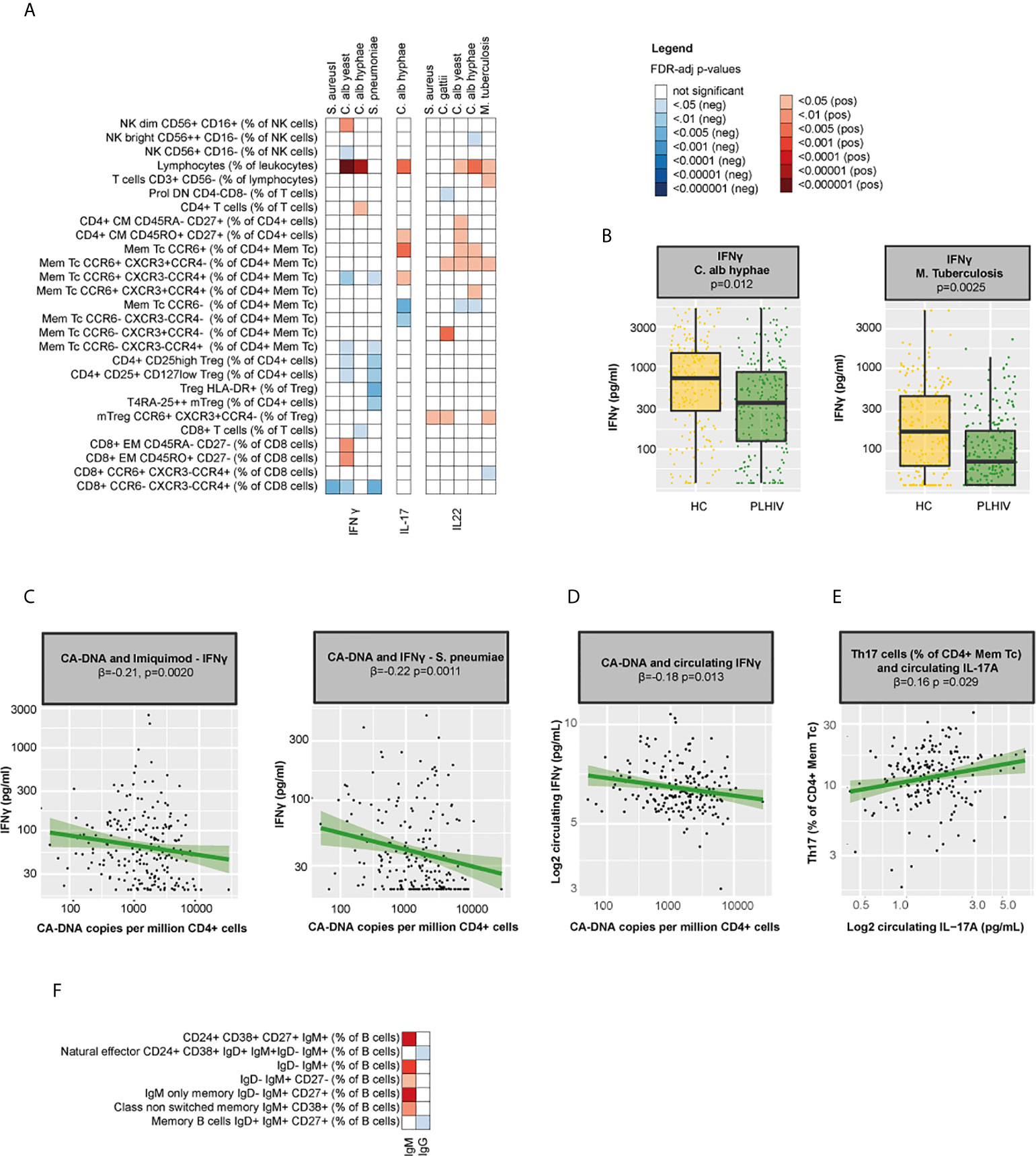
Figure 6 Functional consequences of WBC alterations in PLHIV (A) Heatmap of NK(T) and T cell percentages (total n=71) that were significantly associated with the ex vivo production of IFN-γ, IL-17, and IL-22 upon seven days ex vivo stimulation of PBMCs with different stimuli (n=7) in 211 PLHIV; WBC subsets that showed no significant correlations with any of the parameters (n=44) have been removed from the figure. (B) Boxplot showing differences in ex vivo IFN-γ responses upon stimulation with C. albicans and M. tuberculosis between PLHIV (n=211) and healthy controls (n=56). (C) Association between CA-DNA and ex vivo IFN-γ responses to stimulation with Imiquimod and S. pneumoniae in PLHIV. (D) Association between CA-DNA and circulating IFN-γ in PLHIV. (E) Association between Th17 percentages and circulating IL-17A in PLHIV. (F) Heatmap of B cell percentages (total n=28) that were significantly correlated with IgM or IgG levels in PLHIV; WBC subsets that showed no significant correlations with any of the parameters (n=21) have been removed from the figure. Inverse-rank transformed data were analyzed using linear regression analyses and corrected for sampling time. For cohort comparisons, data were corrected for age, sex, sampling time, season and the CD4+ and CD8+ cell percentages. For color coding of the FDR-adjusted p-values see legend. CA-DNA, CD4-cell-associated HIV-1 DNA; HC, healthy control; IFN-γ, interferon gamma; Ig, immunoglobulin; IL-17(A), interleukin 17(A); IL-22, interleukin 22; Mem Tc, CD4+ memory T cell; NK(T) cells, natural killer (T) cells; PBMC, peripheral blood mononuclear cell; PLHIV, people living with HIV; sCD14, soluble CD14; Th17 percentages, T-helper 17 cell percentages (Mem Tc CCR6+ CXCR3-CCR4+ as percentage of CD4+ Mem Tc); WBC, white blood cell.
Second, we measured serum immunoglobulins and found significant correlations between serum IgM and IgM+ B cell populations (Figure 6F), but no cohort differences in IgM levels [median (IQR) 0.76 (0.56 – 1.05) g/L in PLHIV versus 0.82 (0.65 – 1.21) g/L in controls, p= 0.22] or IgG levels [10.03 (8.34 - 11.59) g/L in PLHIV versus 9.10 (7.59 – 11.38) g/L in controls, p= 0.12]. These results indicate that while some of the alterations in adaptive immune function may be reversed by long-term cART (44, 45), others, such as impaired IFN-γ responses, remain.
In this study we show that, despite suppressive cART, the circulating innate and adaptive immune cell composition in PLHIV differs from that of HIV-uninfected individuals. We confirm that PLHIV exhibit a WBC profile characterized by proliferating memory and effector CD4+ and CD8+ T cells. While untreated HIV infection has been associated with a loss of circulating Th17 cells (9), we observed an expansion of circulating Th17 cells and increased Th17/Treg ratios during stable suppressive treatment, which was associated with plasma concentrations of IL-6, CCL20 and the microbial translocation marker IFABP. Furthermore, PLHIV showed clear changes in B cell maturation with reduced memory B cell percentages and increased plasmablast numbers. In the innate compartment, we observed an expansion of monocytes together with a loss of NK cells, specifically NK bright cells. In addition to age, sex, smoking, and CMV, we found strong associations between WBC populations and markers of the HIV-1 reservoir (CA-DNA and CA-RNA). Functionally, Th17 responses seemed to be preserved, whereas IFN-γ responses to C. albicans and M. tuberculosis were compromised, especially in those with higher CA-DNA. The compromised IFN-γ responses may affect host defense against some important bacterial pathogens (including M. tuberculosis) and the HIV-1 reservoir.
Prior studies have shown that untreated HIV infection results in a massive depletion of Th17 cells from the peripheral blood and the mucosa (9), a process that may partially be reversed by cART (46–48). We recently showed that PLHIV from the same cohort exhibited a pro-inflammatory profile with increased monocyte-derived cytokines, particularly IL-1β (20). IL-1β and IL-6 are among the critical cytokines driving differentiation of Th17 (42), and we postulate that these cytokines may have contributed to the higher circulating Th17 numbers. Th17 cells in the peripheral blood poorly reflect mucosal Th17 numbers (49) and it is possible that mucosal Th17 depletion with increased microbial translocation and altered Th17 recruitment occurs in the participants of our study. The gut-inflammatory marker generating islet-derived protein 3α (REG3α) would be of interest for future studies on the interplay between Th17 and epithelial gut damage in PLHIV (50).Concurrently to increased Th17 cells and pro-inflammatory Th17-like (CCR6+) mTregs, PLHIV showed increased peripheral blood Th17/Treg ratios. Increased Th17/Treg ratios have been linked to cardiovascular disease and atherosclerosis (33), cancer (34), and major depressive disorder (51), which are all highly prevalent in long-term treated PLHIV. Th17-mediated auto-immune diseases like psoriasis are also common among PLHIV, although they mostly occur during periods of severe immunosuppression and resolve upon cART initiation (52). Despite these changes in Th17, IL-17 and IL-22 cytokine responses did not differ between PLHIV and healthy controls. In contrast, we observed reduced ex vivo IFN-γ responses to C. albicans and M. tuberculosis in PLHIV. IFN-γ is predominantly produced by NK(T) cells, Th1, and CD8+ cells (44). Given that CD8+ cells were increased in PLHIV and Th1 cells did not differ between PLHIV and controls, we postulate that the reduced IFN-γ responses may have resulted from the loss of NK cells, which has been reported previously in both untreated and treated PLHIV (53, 54). In line with prior data, we observed an inverse relationship between IFN-γ responses and CA-DNA, suggesting that the failure to restore the NK cell compartment after cART initiation may be important for the containment of the HIV-1 reservoir (55).Moreover, IFN-γ is a key cytokine in the immune response against M. tuberculosis which remains an important pathogen in treated PLHIV (56). Improving IFN-γ responses, may therefore be relevant in the context of M. tuberculosis and HIV cure.
Different factors may contribute to the variation in T-cells repertoire in PLHIV. First, the effects of demographic factors such as age, sex, and smoking resembled those observed previously in healthy individuals (19, 41). Second, PLHIV in our study were almost universally coinfected with CMV and, in line with earlier data in HIV-infected and uninfected individuals, CMV seropositivity was associated with the expansion of effector and EM CD4+ and CD8+ T cells (14, 41, 57, 58). Of note, high CMV IgG levels may reflect frequent CMV reactivations or result from a stronger immune response (including adequate B/T cell interactions and B cell responses) and, consequently, fewer activations (59). Importantly, higher CMV IgG titers have been linked to microbial translocation and the development of non-AIDS-defining events such as cardiovascular disease (14, 60). Third, we found substantial associations between WBC subsets and CD4+ nadir and markers of the HIV-1 reservoir. Overall, CA-DNA showed more and stronger correlations with WBC subsets than did CA-RNA, which may be explained by the fact that CA-RNA levels, reflecting transcriptional activity, are low during viral suppression and subtle effects may be missed (13, 61).
Next to T cell dysfunction, HIV is characterized by aberrant B cell responses and B cell dysfunction. Using a different set of B cell markers than earlier studies in PLHIV (62, 63), we confirm their observations that percentages of naïve B cells are increased and memory B cells are reduced in PLHIV. Moreover, adequate B cell maturation requires optimal communication between T and B cells, which, according to our data, might be compromised in chronic stable PLHIV. B/T cell interactions take place in the germinal centers in lymph nodes and are orchestrated by follicular Th cells (Tfh) (64). As these cells are known to be highly permissive to HIV infection and serve as reservoirs during chronic infection, they could potentially explain these disrupted B/T cell interactions (64–66). Clinically, compromised B/T cells interactions may contribute to impaired immune responses to vaccination as well as increased risks for infections, such as invasive pneumococcal disease (5, 67). Finally, improvement of B/T cell communication is crucial for the development of broadly neutralizing antibodies and thus functional cure in PLHIV (68).
Our findings support the relevance of new immune modulating strategies in ART-treated PLHIV. Examples of interventions with potent anti-inflammatory properties in PLHIV include the IL-1β-inhibiting agent canakinumab (69) and the epigenetic modifier panobinostat (HDACi) (70, 71). Moreover, checkpoint inhibitors (e.g. those targeting PD1, PD-L1, and CTLA4) have been shown to, transiently, reverse latency of the viral reservoir, to restore cytotoxic T cell functions (72–75), and to enhance B-cell germinal responses to HIV-1 envelope vaccines (76). Further research is required to establish whether these agents are safe and effective long-term options to mitigate inflammation and to reverse immune exhaustion and HIV latency in stable HIV infection.
Several limitations should be considered when interpreting our data. First, the observational study design limits causal inferences. Second, generalizability of our findings to women, children and non-European populations requires further studying. Third, participants from the HIV cohort were older and more often male and age and sex both have known effects on the immune system (19, 21) To adjust for these differences, we used multivariable regression models which allowed us to take into account the effects of these independent predictors (age and sex) on our outcome of interest (HIV status). Fourth, we used a predefined set of markers and flow cytometry panels. While this standardized approach enhances validity and reproducibility and enabled us to compare our results with those of a large cohort of healthy controls, some interesting WBC subsets and markers for HIV have not been included (e.g. Tfh cells, CD38 HLA-DR co-expression, and PD-1/CD57 expression in the context of B/T cell interactions, T-cell activation, and immune cell exhaustion respectively). Furthermore, data on intracellular production of IL-17 and IFN-γ in maximally stimulated sorted cell populations (e.g. Th17 or NK cells) may provide a better understanding of the changes in cytokine production capacity per WBC subset. Finally, our WBC data are limited to the peripheral blood and we have no data on tissue-specific WBC composition (e.g. lymph nodes or the gut).
In summary, we show that the circulating innate and adaptive immune cell composition is altered in a large group of PLHIV receiving cART for more than six months. Furthermore, our findings suggest that some of the adaptive immune responses (Th17) are preserved while IFN-γ responses are compromised. Our comprehensive approach provides new insight into the changes in the immune cell architecture and functional immunity in treated HIV and highlights associations with the HIV-reservoir, underlining the need for early cART initiation. Our results are currently validated and extended in a multi-omics study including 2000 virally suppressed PLHIV (clinicaltrials.gov identifier: NCT03994835).
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving human participants were reviewed and approved by Medical Ethical Review Committee region Arnhem-Nijmegen (ref. 42561.091.122). The patients/participants provided their written informed consent to participate in this study.
WH, QM, MN, HK, IJ, and AV designed the study. WH, LW and MJ recruited and included the participants. WH, LW, MJ, WT, SR, BC, and ER performed the laboratory experiments. RH, LW and WH analyzed the data and interpreted the data together with QM, AV, MN, LK, HK, IJ and JL. LW, WH, AV and QM wrote the manuscript. All authors contributed to the article and approved the submitted version.
QM, AV, and MN receive research support from ViiV Healthcare. The funders were not involved in the study design, data interpretation or the submission.
This study was supported by an AIDS-fonds (#P-29001) Netherlands and an ERC Advanced Grant (#833247). LV was supported by FWO (grant 1.8.020.09.N.00) and Collen-Francqui Research Professor Mandate. SR received a strategic basic research fund of the Research Foundation – Flanders (FWO, 1S32916N). TS and JL were employed by ViiV Healthcare.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all volunteers from the 200HIV and 56P cohorts of the HFGP for participation in the study. We thank the clinicians and nurse practitioners of the HIV clinic of the Radboud university medical center for their help with participant recruitment and Anouk Janssen, Sanne Smeekens and Marije Oosting for their help with participant inclusion.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.661990/full#supplementary-material
1. Smit M, Brinkman K, Geerlings S, Smit C, Thyagarajan K, Sighem A, et al. Future challenges for clinical care of an ageing population infected with HIV: a modelling study. Lancet Infect Dis (2015) 15(7):810–8. doi: 10.1016/S1473-3099(15)00056-0
2. Harboe ZB, Larsen MV, Ladelund S, Kronborg G, Konradsen HB, Gerstoft J, et al. Incidence and Risk Factors for Invasive Pneumococcal Disease in HIV-Infected and Non-HIV-Infected Individuals Before and After the Introduction of Combination Antiretroviral Therapy: Persistent High Risk Among HIV-Infected Injecting Drug Users. Clin Infect Dis (2014) 59(8):1168–76. doi: 10.1093/cid/ciu558
3. Kerneis S, Launay O, Turbelin C, Batteux F, Hanslik T, Boelle PY. Long-term Immune Responses to Vaccination in HIV-Infected Patients: A Systematic Review and Meta-Analysis. Clin Infect Dis (2014) 58(8):1130–9. doi: 10.1093/cid/cit937
4. O’Connor J, Vjecha MJ, Phillips AN, Angus B, Cooper D, Grinsztejn B, et al. Effect of immediate initiation of antiretroviral therapy on risk of severe bacterial infections in HIV-positive people with CD4 cell counts of more than 500 cells per muL: secondary outcome results from a randomised controlled trial. Lancet HIV (2017) 4(3):E105–E12. doi: 10.1016/S2352-3018(16)30216-8
5. Garcia Garrido HM, Mak AMR, Wit F, Wong GWM, Knol MJ, Vollaard A, et al. Incidence and Risk Factors for Invasive Pneumococcal Disease and Community-acquired Pneumonia in Human Immunodeficiency Virus-Infected Individuals in a High-income Setting. Clin Infect Dis (2020) 71(1):41–50. doi: 10.1093/cid/ciz728
6. Ganatra SR, Bucsan AN, Alvarez X, Kumar S, Chatterjee A, Quezada M, et al. Antiretroviral therapy does not reduce tuberculosis reactivation in a tuberculosis-HIV coinfection model. J Clin Invest (2020) 130(10):5171–9. doi: 10.1172/JCI136502
7. Titanji B, Gavegnano C, Hsue P, Schinazi R, Marconi VC. Targeting Inflammation to Reduce Atherosclerotic Cardiovascular Risk in People With HIV Infection. J Am Heart Assoc (2020) 9(3):e014873. doi: 10.1161/JAHA.119.014873
8. Deeks SG. HIV infection, inflammation, immunosenescence, and aging. Annu Rev Med (2011) 62:141–55. doi: 10.1146/annurev-med-042909-093756
9. Chevalier MF, Petitjean G, Dunyach-Remy C, Didier C, Girard PM, Manea ME, et al. The Th17/Treg Ratio, IL-1RA and sCD14 Levels in Primary HIV Infection Predict the T-cell Activation Set Point in the Absence of Systemic Microbial Translocation. PloS Pathog (2013) 9(6). doi: 10.1371/journal.ppat.1003453
10. Brenchley JM. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Retrovirology (2006) 3. doi: 10.1186/1742-4690-3-S1-S98
11. Perkins, Wolinsky D. The Interplay Between Host Genetic Variation, Viral Replication, and Microbial Translocation in Untreated HIV-Infected Individuals (vol 212, pg 578, 2015). J Infect Dis (2015) 212(10):1686–6. doi: 10.1093/infdis/jiv089
12. Siedner MJ, Zanni M, Tracy RP, Kwon DS, Tsai AC, Kakuhire B, et al. Increased Systemic Inflammation and Gut Permeability Among Women With Treated HIV Infection in Rural Uganda. J Infect Dis (2018) 218(6):922–6. doi: 10.1093/infdis/jiy244
13. Cohn LB, Chomont N, Deeks SG. The Biology of the HIV-1 Latent Reservoir and Implications for Cure Strategies. Cell Host Microbe (2020) 27(4):519–30. doi: 10.1016/j.chom.2020.03.014
14. Ramendra R, Isnard S, Lin J, Fombuena B, Ouyang J, Mehraj V, et al. Cytomegalovirus Seropositivity Is Associated With Increased Microbial Translocation in People Living With Human Immunodeficiency Virus and Uninfected Controls. Clin Infect Dis (2020) 71(6):1438–46. doi: 10.1093/cid/ciz1001
15. Isnard S, Ramendra R, Dupuy FP, Mehraj V, Lin J, Kokinov N, et al. Relevance of Reg3 alpha and I-FABP on microbial translocation, inflammation and reservoir size in people living with HIV. J Int AIDS Soc (2019) 22:67–8. doi: 10.1016/S2055-6640(20)31030-X
16. Mehraj V, Ramendra R, Isnard S, Dupuy FP, Ponte R, Chen J, et al. Circulating (1–>3)-beta-D-glucan Is Associated With Immune Activation During Human Immunodeficiency Virus Infection. Clin Infect Dis (2020) 70(2):232–41. doi: 10.1093/cid/ciz212
17. Olson A, Coote C, Snyder-Cappione JE, Lin N, Sagar M. HIV-1 transcription but not intact provirus levels are associated with systemic inflammation. J Infect Dis (2020). doi: 10.1093/infdis/jiaa657
18. Netea MG, Joosten LA, Li Y, Kumar V, Oosting M, Smeekens S, et al. Understanding human immune function using the resources from the Human Functional Genomics Project. Nat Med (2016) 22(8):831–3. doi: 10.1038/nm.4140
19. Aguirre-Gamboa R, Joosten I, Urbano PC, van der Molen RG, van Rijssen E, van Cranenbroek B, et al. Differential Effects of Environmental and Genetic Factors on T and B Cell Immune Traits. Cell Rep (2016) 17(9):2474–87. doi: 10.1016/j.celrep.2016.10.053
20. van der Heijden WA, van de Wijer L, Keramati F, Trypsteen W, Rutsaert S, Ter Horst R, et al. Chronic HIV infection induces transcriptional and functional reprogramming of innate immune cells. JCI Insight (2021) 6(7):e145928. doi: 10.1172/jci.insight.145928
21. Ter Horst R, Jaeger M, Smeekens SP, Oosting M, Swertz MA, Li Y, et al. Host and Environmental Factors Influencing Individual Human Cytokine Responses. Cell (2016) 167(4):1111–24.e13. doi: 10.1016/j.cell.2016.10.018
22. Oosting M, Buffen K, Cheng SC, Verschueren IC, Koentgen F, van de Veerdonk FL, et al. Borrelia-induced cytokine production is mediated by spleen tyrosine kinase (Syk) but is Dectin-1 and Dectin-2 independent. Cytokine (2015) 76(2):465–72. doi: 10.1016/j.cyto.2015.08.005
23. Assarsson E, Lundberg M, Holmquist G, Bjorkesten J, Thorsen SB, Ekman D, et al. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PloS One (2014) 9(4):e95192. doi: 10.1371/journal.pone.0095192
24. Koeken VACM, de Bree LCJ, Mourits VP, Moorlag SJCFM, Walk J, Cirovic B, et al. BCG vaccination in humans inhibits systemic inflammation in a sex-dependent manner. J Clin Invest (2020) 130(10):5591–602. doi: 10.1172/JCI133935
25. Chomont N, El-Far M, Ancuta P, Trautmann L, Procopio FA, Yassine-Diab B, et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med (2009) 15(8):893–900. doi: 10.1038/nm.1972
26. Pasternak AO, Lukashov VV, Berkhout B. Cell-associated HIV RNA: a dynamic biomarker of viral persistence. Retrovirology (2013) 10:41. doi: 10.1186/1742-4690-10-41
27. Rutsaert S, De Spiegelaere W, De Clercq L, Vandekerckhove L. Evaluation of HIV-1 reservoir levels as possible markers for virological failure during boosted darunavir monotherapy. J Antimicrobial Chemotherapy (2019) 74(10):3030–4. doi: 10.1093/jac/dkz269
28. Trypsteen W, Vynck M, De Neve J, Bonczkowski P, Kiselinova M, Malatinkova E, et al. ddpcRquant: threshold determination for single channel droplet digital PCR experiments. Anal Bioanal Chem (2015) 407(19):5827–34. doi: 10.1007/s00216-015-8773-4
29. Benjamini Y, Hochberg Y. Controlling the False Discovery Rate - a Practical and Powerful Approach to Multiple Testing. J R Stat Soc Ser B-Statistical Method (1995) 57(1):289–300. doi: 10.1111/j.2517-6161.1995.tb02031.x
30. Becattini S, Hochberg Y. T cell immunity. Functional heterogeneity of human memory CD4(+) T cell clones primed by pathogens or vaccines. Science (2015) 347(6220):400–6. doi: 10.1126/science.1260668
31. Li ZY, Latorre D, Mele F, Foglierini M, De Gregorio C, Cassotta A, et al. FOXP3(+) regulatory T cells and their functional regulation. Cell Mol Immunol (2015) 12(5):558–65. doi: 10.1038/cmi.2015.10
32. Schulze Zur Wiesch J, Li D, Tsun A, Li B. Comprehensive analysis of frequency and phenotype of T regulatory cells in HIV infection: CD39 expression of FoxP3+ T regulatory cells correlates with progressive disease. J Virol (2011) 85(3):1287–97. doi: 10.1128/JVI.01758-10
33. Saigusa R, Winkels H, Ley K. T cell subsets and functions in atherosclerosis. Nat Rev Cardiol (2020) 17(7):387–401. doi: 10.1038/s41569-020-0352-5
34. Knochelmann HM, Dwyer CJ, Bailey SR, Amaya SM, Elston DM, Mazza-McCrann JM, et al. When worlds collide: Th17 and Treg cells in cancer and autoimmunity. Cell Mol Immunol (2018) 15(5):458–69. doi: 10.1038/s41423-018-0004-4
35. Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol (2004) 22:745–63. doi: 10.1146/annurev.immunol.22.012703.104702
36. Mahnke YD, Brodie TM, Sallusto F, Roederer M, Lugli E. The who’s who of T-cell differentiation: human memory T-cell subsets. Eur J Immunol (2013) 43(11):2797–809. doi: 10.1002/eji.201343751
37. Sachsenberg N, Perelson AS, Yerly S, Schockmel GA, Leduc D, Hirschel B, et al. Turnover of CD4(+) and CD8(+) T lymphocytes in HIV-1 infection as measured by Ki-67 antigen. J Exp Med (1998) 187(8):1295–303. doi: 10.1084/jem.187.8.1295
38. Moir S, Fauci AS. Insights into B cells and HIV-specific B-cell responses in HIV-infected individuals. Immunol Rev (2013) 254(1):207–24. doi: 10.1111/imr.12067
39. Moir S, Buckner CM, Ho J, Wang W, Chen J, Waldner AJ, et al. B cells in early and chronic HIV infection: evidence for preservation of immune function associated with early initiation of antiretroviral therapy. Blood (2010) 116(25):5571–9. doi: 10.1182/blood-2010-05-285528
40. Arts RJ, Novakovic B, Ter Horst R, Carvalho A, Bekkering S, Lachmandas E, et al. Glutaminolysis and Fumarate Accumulation Integrate Immunometabolic and Epigenetic Programs in Trained Immunity. Cell Metab (2016) 24(6):807–19. doi: 10.1016/j.cmet.2016.10.008
41. Patin E, Hasan M, Bergstedt J, Rouilly V, Libri V, Urrutia A, et al. Natural variation in the parameters of innate immune cells is preferentially driven by genetic factors. Nat Immunol (2018) 19(3):302–14 doi: 10.1038/s41590-018-0049-7.
42. Acosta-Rodriguez EV, Napolitani G, Lanzavecchia A, Sallusto F. Interleukins 1 beta and 6 but not transforming growth factor-beta are essential for the differentiation of interleukin 17-producing human T helper cells. Nat Immunol (2007) 8(9):942–9. doi: 10.1038/ni1496
43. Schutyser E, Struyf S, Van Damme J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor Rev (2003) 14(5):409–26. doi: 10.1016/S1359-6101(03)00049-2
44. Roff SR, Noon-Song EN, Yamamoto JK. The Significance of Interferon-gamma in HIV-1 Pathogenesis, Therapy, and Prophylaxis. Front Immunol (2014) 4:498. doi: 10.3389/fimmu.2013.00498
45. Kim CJ, Nazli A, Rojas OL, Chege D, Alidina Z, Huibner S, et al. A role for mucosal IL-22 production and Th22 cells in HIV-associated mucosal immunopathogenesis. Mucosal Immunol (2012) 5(6):670–80. doi: 10.1038/mi.2012.72
46. Gosselin A, Monteiro P, Chomont N, Diaz-Griffero F, Said EA, Fonseca S, et al. Peripheral Blood CCR4(+)CCR6(+) and CXCR3(+)CCR6(+) CD4(+) T Cells Are Highly Permissive to HIV-1 Infection. J Immunol (2010) 184(3):1604–16. doi: 10.4049/jimmunol.0903058
47. He Y, Li J, Zheng YH, Luo Y, Zhou HY, Yao YH, et al. A Randomized Case-Control Study of Dynamic Changes in Peripheral Blood Th17/Treg Cell Balance and Interleukin-17 Levels in Highly Active Antiretroviral-Treated HIV Type 1/AIDS Patients. AIDS Res Hum Retroviruses (2012) 28(4):339–45. doi: 10.1089/aid.2011.0140
48. Kim CJ, McKinnon LR, Kovacs C, Kandel G, Huibner S, Chege D, et al. Mucosal Th17 cell function is altered during HIV infection and is an independent predictor of systemic immune activation. J Immunol (2013) 191(5):2164–73. doi: 10.4049/jimmunol.1300829
49. Nayrac M, Requena M, Loiseau C, Cazabat M, Suc B, Carrere N, et al. Th22 cells are efficiently recruited in the gut by CCL28 as an alternative to CCL20 but do not compensate for the loss of Th17 cells in treated HIV-1-infected individuals. Mucosal Immunol (2021) 14(1):219–28. doi: 10.1038/s41385-020-0286-6
50. Isnard S, Ramendra R, Dupuy FP, Lin J, Fombuena B, Kokinov N, et al. Plasma Levels of C-Type Lectin REG3alpha and Gut Damage in People With Human Immunodeficiency Virus. J Infect Dis (2020) 221(1):110–21. doi: 10.1093/infdis/jiz423
51. Ghosh R, Kumar PK, Mitra P, Purohit P, Nebhinani N, Sharma P. Circulating T helper 17 and IFN-gamma positive Th17 cells in Major Depressive Disorder. Behav Brain Res (2020) 394:112811. doi: 10.1016/j.bbr.2020.112811
52. Morar N, Willis-Owen SA, Maurer T, Bunker CB. HIV-associated psoriasis: pathogenesis, clinical features, and management. Lancet Infect Dis (2010) 10(7):470–8. doi: 10.1016/S1473-3099(10)70101-8
53. Dillon SM, Lee EJ, Bramante JM, Barker E, Wilson CC. The Natural Killer Cell Interferon-Gamma Response to Bacteria Is Diminished in Untreated HIV-1 Infection and Defects Persist Despite Viral Suppression. Jaids-Journal Acquired Immune Deficiency Syndromes (2014) 65(3):259–67. doi: 10.1097/01.qai.0000435603.50598.2b
54. Azzoni L, Papasavvas E, Chehimi J, Kostman JR, Mounzer K, Ondercin J, et al. Sustained impairment of IFN-gamma secretion in suppressed HIV-infected patients despite mature NK cell recovery: evidence for a defective reconstitution of innate immunity. J Immunol (2002) 168(11):5764–70. doi: 10.4049/jimmunol.168.11.5764
55. Marras F, Casabianca A, Bozzano F, Ascierto ML, Orlandi C, Di Biagio A, et al. Control of the HIV-1 DNA Reservoir Is Associated In Vivo and In Vitro with NKp46/NKp30 (CD335 CD337) Inducibility and Interferon Gamma Production by Transcriptionally Unique NK Cells. J Virol (2017) 91(23). doi: 10.1128/JVI.00647-17
56. Flynn JL, Chan J. Tuberculosis: latency and reactivation. Infect Immun (2001) 69(7):4195–201. doi: 10.1128/IAI.69.7.4195-4201.2001
57. Tu WJ, Rao S. Mechanisms Underlying T Cell Immunosenescence: Aging and Cytomegalovirus Infection. Front Microbiol (2016) 7:2111. doi: 10.3389/fmicb.2016.02111
58. Freeman ML, Mudd JC, Shive CL, Younes SA, Panigrahi S, Sieg SF, et al. CD8 T-Cell Expansion and Inflammation Linked to CMV Coinfection in ART-treated HIV Infection. Clin Infect Dis (2016) 62(3):392–6. doi: 10.1093/cid/civ840
59. Gianella S, Morris SR, Tatro E, Vargas MV, Haubrich RH, Daar ES, et al. Virologic Correlates of Anti-CMV IgG Levels in HIV-1-Infected Men. J Infect Dis (2014) 209(3):452–6. doi: 10.1093/infdis/jit434
60. Lichtner M, Cicconi P, Vita S, Cozzi-Lepri A, Galli M, Lo Caputo S, et al. Cytomegalovirus Coinfection Is Associated With an Increased Risk of Severe Non-AIDS-Defining Events in a Large Cohort of HIV-Infected Patients. J Infect Dis (2015) 211(2):178–86. doi: 10.1093/infdis/jiu417
61. Pasternak AO, Berkhout B. What do we measure when we measure cell-associated HIV RNA. Retrovirology (2018) 15(1):13. doi: 10.1186/s12977-018-0397-2
62. Pensieroso S, Galli L, Nozza S, Ruffin N, Castagna A, Tambussi G, et al. B-cell subset alterations and correlated factors in HIV-1 infection. AIDS (2013) 27(8):1209–17. doi: 10.1097/QAD.0b013e32835edc47
63. Buckner CM, Kardava L, Zhang X, Gittens K, Justement JS, Kovacs C, et al. Maintenance of HIV-Specific Memory B-Cell Responses in Elite Controllers Despite Low Viral Burdens. J Infect Dis (2016) 214(3):390–8. doi: 10.1093/infdis/jiw163
64. Vinuesa CG. HIV and T follicular helper cells: a dangerous relationship. J Clin Invest (2012) 122(9):3059–62. doi: 10.1172/JCI65175
65. Lindqvist M, van Lunzen J, Soghoian DZ, Kuhl BD, Ranasinghe S, Kranias G, et al. Expansion of HIV-specific T follicular helper cells in chronic HIV infection. J Clin Invest (2012) 122(9):3271–80. doi: 10.1172/JCI64314
66. Planchais C, Hocqueloux L, Ibanez C, Gallien S, Copie C, Surenaud M, et al. Early Antiretroviral Therapy Preserves Functional Follicular Helper T and HIV-Specific B Cells in the Gut Mucosa of HIV-1-Infected Individuals. J Immunol (2018) 200(10):3519–29. doi: 10.4049/jimmunol.1701615
67. Abzug MJ, Warshaw M, Rosenblatt HM, Levin MJ, Nachman SA, Pelton SI, et al. Immunogenicity and Immunologic Memory after Hepatitis B Virus Booster Vaccination in HIV-Infected Children Receiving Highly Active Antiretroviral Therapy. J Infect Dis (2009) 200(6):935–46. doi: 10.1086/605448
68. Moir S, Fauci AS. B-cell responses to HIV infection. Immunological Rev (2017) 275(1):33–48. doi: 10.1111/imr.12502
69. Hsue PY, Li D, Ma Y, Ishai A, Manion M, Nahrendorf M, et al. IL-1beta Inhibition Reduces Atherosclerotic Inflammation in HIV Infection. J Am Col Cardiol (2018) 72:2809–11. doi: 10.1016/j.jacc.2018.09.038
70. Brinkmann CR, Hojen JF, Rasmussen TA, Kjaer AS, Olesen R, Denton PW, et al. Treatment of HIV-Infected Individuals with the Histone Deacetylase Inhibitor Panobinostat Results in Increased Numbers of Regulatory T Cells and Limits Ex Vivo Lipopolysaccharide-Induced Inflammatory Responses. mSphere (2018) 3(1):e00616–17. doi: 10.1128/mSphere.00616-17
71. Hogh Kolbaek Kjaer AS, Brinkmann CR, Dinarello CA, Olesen R, Ostergaard L, Sogaard OS, et al. The histone deacetylase inhibitor panobinostat lowers biomarkers of cardiovascular risk and inflammation in HIV patients. AIDS (2015) 29:1195–200. doi: 10.1097/QAD.0000000000000678
72. Chen H, Moussa M, Catalfamo M. The Role of Immunomodulatory Receptors in the Pathogenesis of HIV Infection: A Therapeutic Opportunity for HIV Cure? Front Immunol (2020) 11(1):1223. doi: 10.3389/fimmu.2020.01223
73. Evans VA, van der Sluis RM, Solomon A, Dantanarayana A, McNeil C, Garsia R, et al. Programmed cell death-1 contributes to the establishment and maintenance of HIV-1 latency. AIDS (2018) 32(1):1491–7. doi: 10.1097/QAD.0000000000001849
74. Le Garff G, Samri A, Lambert-Niclot S, Even S, Lavole A, Cadranel J, et al. Transient HIV-specific T cells increase and inflammation in an HIV-infected patient treated with nivolumab. AIDS (2017) 31:1048–51. doi: 10.1097/QAD.0000000000001429
75. Gay CL, Bosch RJ, Ritz J, Hataye JM, Aga E, Tressler RL, et al. Clinical Trial of the Anti-PD-L1 Antibody BMS-936559 in HIV-1 Infected Participants on Suppressive Antiretroviral Therapy. J Infect Dis (2017) 215:1725–33. doi: 10.1093/infdis/jix191
Keywords: HIV, Th17 & Tregs cells, CD4+/CD8+ lymphocytes, B cell, HIV reservoir, CMV, Interferon gama (IFN-γ), Natural killer cell (NK cells)
Citation: Van de Wijer L, van der Heijden WA, Horst Rt, Jaeger M, Trypsteen W, Rutsaert S, van Cranenbroek B, van Rijssen E, Joosten I, Joosten L, Vandekerckhove L, Schoofs T, van Lunzen J, Netea MG, Koenen HJPM, van der Ven AJAM and de Mast Q (2021) The Architecture of Circulating Immune Cells Is Dysregulated in People Living With HIV on Long Term Antiretroviral Treatment and Relates With Markers of the HIV-1 Reservoir, Cytomegalovirus, and Microbial Translocation. Front. Immunol. 12:661990. doi: 10.3389/fimmu.2021.661990
Received: 31 January 2021; Accepted: 29 March 2021;
Published: 19 April 2021.
Edited by:
Constantinos Petrovas, Centre Hospitalier Universitaire Vaudois (CHUV), SwitzerlandReviewed by:
Stéphane Isnard, McGill University Health Centre, CanadaCopyright © 2021 Van de Wijer, van der Heijden, Horst, Jaeger, Trypsteen, Rutsaert, van Cranenbroek, van Rijssen, Joosten, Joosten, Vandekerckhove, Schoofs, van Lunzen, Netea, Koenen, van der Ven and de Mast. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lisa Van de Wijer, TC52YW5kZXdpamVyQHJhZGJvdWR1bWMubmw=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.